 Laura Massarenti1
Laura Massarenti1 Christian Enevold1Dres Damgaard1
Christian Enevold1Dres Damgaard1 Niels Ødum2
Niels Ødum2 Peter Garred3
Peter Garred3 Morten Frisch4
Morten Frisch4 Miriam A. Shelef5,6
Miriam A. Shelef5,6 Søren Jacobsen7*†Claus Henrik Nielsen1,8†
Søren Jacobsen7*†Claus Henrik Nielsen1,8†- 1Institute for Inflammation Research, Center for Rheumatology and Spine Diseases, Section 7521, Copenhagen University Hospital Rigshospitalet, Copenhagen, Denmark
- 2LEO Foundation Skin Immunology Research Center, Department of Immunology and Microbiology, University of Copenhagen, Copenhagen, Denmark
- 3Laboratory of Molecular Medicine, Department of Clinical Immunology, Section 7631, Copenhagen University Hospital Rigshospitalet, Copenhagen, Denmark
- 4Department of Epidemiology Research, Statens Serum Institut, Copenhagen, Denmark
- 5Department of Medicine, University of Wisconsin–Madison, Madison, WI, United States
- 6William S. Middleton Memorial Veterans Hospital, Madison, WI, United States
- 7Copenhagen Lupus and Vasculitis Clinic, Center for Rheumatology and Spine Diseases, Section 4242, Copenhagen University Hospital Rigshospitalet, Copenhagen, Denmark
- 8Department of Odontology, Faculty of Health and Medical Sciences, University of Copenhagen, Copenhagen, Denmark
Peptidylarginine deiminases (PADs) catalyze citrullination, a post-translational modification playing a pathogenic role in anti-citrullinated protein antibody (ACPA)-positive rheumatoid arthritis (RA). The interplay between single nucleotide polymorphisms (SNPs) in the PADI genes and known risk factors for ACPA-positive RA, including smoking, HLA-DR4 and -1, and the PTPN22 R620W polymorphism, was investigated. We typed four PADI2 SNPs, four PADI4 SNPs, and the PTPN22 R620W SNP in 445 Danish RA patients and 533 age-matched healthy controls, as well as in 200 North American RA patients and 100 age- and sex-matched controls. The HLA-DRB1 locus was typed in the Danish cohort. Logistic regression analyses, adjusted for age, sex, smoking status, and PTPN22 R620W, revealed increased risk of anti-CCP-positive RA in carriers of rs11203367(T) (OR: 1.22, p=0.03) and reduced risk in carriers of rs2240335(A) in PADI4 (OR: 0.82, p=0.04). rs74058715(T) in PADI4 conferred reduced risk of anti-CCP-negative RA (OR: 0.38, p=0.003). In HLA-DRB1*04-positive individuals, specifically, the risk of anti-CCP-positive RA was increased by carriage of PADI4 rs1748033(T) (OR: 1.54, p=0.007) and decreased by carriage of PADI4 rs74058715(T) (OR: 0.44, p=0.01), and we observed an interaction between these SNPs and HLA-DRB1*04 (p=0.004 and p=0.008, respectively) Thus, PADI4 polymorphisms associate with ACPA-positive RA, particularly in HLA-DRB1*04-positive individuals, and with ACPA-negative RA independently of HLA-DRB1*04.
Introduction
Peptidylarginine deiminases (PADs) catalyze the post-translational conversion of peptidylarginine to peptidylcitrulline. Citrullination is involved in many physiological functions, including skin keratinization, neuron myelination, and formation of neutrophil extracellular traps (NETs) (1–3) but is also thought play a pathogenic role in anti-citrullinated protein-antibody (ACPA)-positive rheumatoid arthritis (RA) (4). ACPAs, routinely detected using the anti-cyclic citrullinated peptide (anti-CCP) test (5), are often present years before clinically overt disease (6), and anti-CCP-positive RA patients have a worse prognosis with a higher degree of erosive damage than anti-CPP-negative RA patients (7). ACPA-positive RA is strongly associated with HLA molecules containing the so-called shared epitope (SE), which are capable of binding certain citrullinated peptides (8–10). The shared epitope is present in most HLA-DRB1*04 subtypes (HLA-DRB1*0401, HLA-DRB1*0404, HLA-DRB1*0405, and HLA-DRB1*0408 but not in HLA-DR4B1*0402) and HLA-DRB1*01 subtypes (HLA-DRB1*0101 and HLA-DRB1*0102). However, HLA-DRB1*04, which is carried by 67–78% of anti-CCP-positive RA patients (10–12), correlates more strongly with anti-CCP-positive RA than HLA-DRB1*01 (reported in 22–51% of anti-CCP-positive RA patients) (10–12). In addition, the R620W variant of protein tyrosine phosphatase, non-receptor type 22 (PTPN22), confers increased risk of ACPA-positive RA (13, 14), as does smoking (9).
The five existing PAD isoforms in humans are encoded by the PADI1-4 and -6 genes (15). PAD2 and PAD4 are expressed by immune cells (16–18) and are present in the synovium of RA patients, where their expression levels correlate with those of inflammatory markers (18, 19). Some investigators have found that PAD4 generates autoantigens recognized by ACPAs more efficiently than PAD2 (20), while others have shown that the two PAD isoforms are approximately equally efficient (21).
Studies on Asian populations have shown associations between RA and single nucleotide polymorphisms (SNPs) in PADI2 (22, 23) as well as SNPs in PADI4 (22–27). However, the observed associations with PADI2 SNPs in Asian populations have not been replicated in studies on European or North American cohorts, while a few studies have confirmed the association of the PADI4 SNPs with RA in those populations (28–34). Notably, one study reported associations between one SNP in PADI4 (rs2240340) and RA in a North American cohort, alone or when combined with a Swedish cohort, but not in the Swedish cohort alone (31). A haplotype of PADI4 has been reported to confer susceptibility to RA in Asians (24), but not in a British study (30). The haplotype consists of the minor alleles of rs11203366, rs11203367, and rs874881 encoding the amino acid substitutions Gly55Ser, Val82Ala, and Gly112Ala, respectively, and rs1748033 which is a synonymous SNP (Figure 1). Besides, the minor allele (A) of the synonymous SNP rs2240035 has been shown to decrease the risk of RA in Asians and North Americans (26, 27, 35). Together with rs74058715, which is located in the 5’ untranslated region (UTR) of PADI4, rs2240035 has been shown to regulate PAD4 expression levels in neutrophils and monocytes (36).
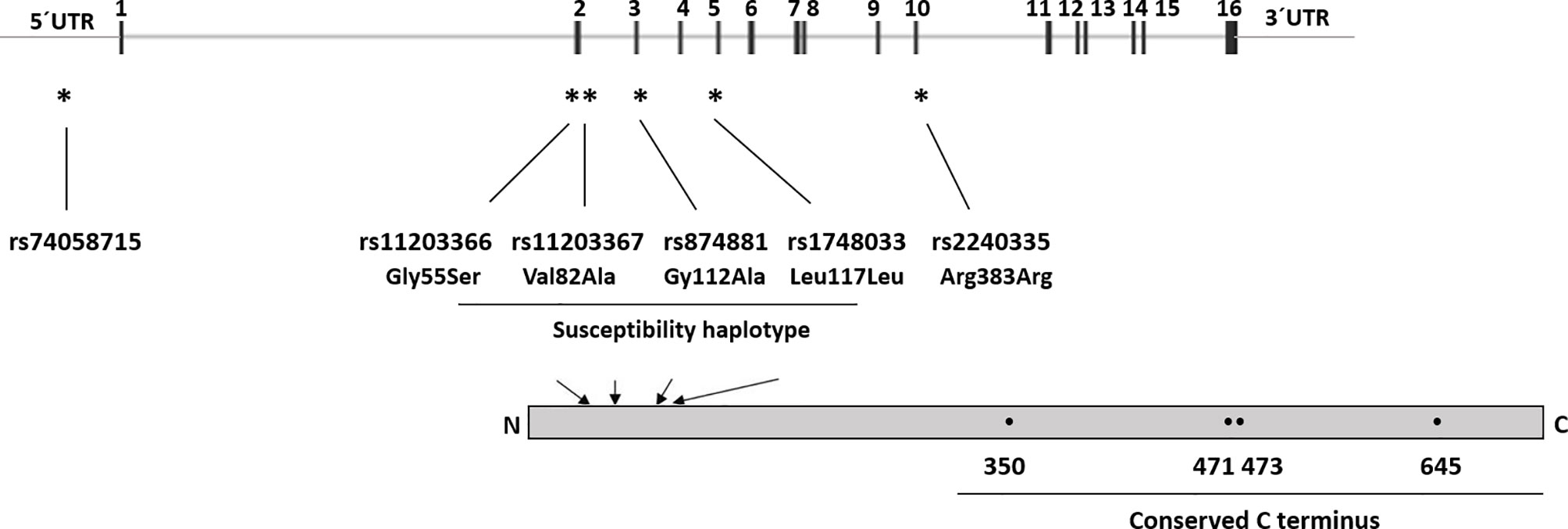
Figure 1 PADI4 gene structure and SNP location. UTR, Untranslated region, 1–16 indicates the exon number. * indicates SNPs location within the PADI4 gene, arrows indicate location of the SNP corresponding amino acids in the protein, and • indicates active site residues (Asp350, His471, Asp473, and Cys645).
Only few studies on PADI polymorphism in RA have stratified the patients according to HLA types (28, 32, 37, 38) and, generally, studies addressing the influence of HLA types on development of RA have not taken PADI polymorphism into account. We hypothesized that PADI polymorphisms exert their greatest influence in subjects carrying HLA types that bind citrullinated peptides most strongly, i.e., HLA-DRB1*04.
Our aim was to determine if anti-CCP-positive or anti-CCP-negative RA is associated with SNPs in PADI2 or PADI4 in a Danish and a North American cohort of RA patients and healthy controls. Moreover, we aimed at elucidating the interaction of PADI SNPs with HLA-DRB1*04 or HLA-DRB1*01 in predisposing to RA.
Materials and Methods
Patients and Controls
The Danish cohort included 445 RA patients clinically identified at 21 rheumatology departments across Denmark and 533 population controls matched for age (Table 1). The cohort has previously been reported on (39). The RA patients had less than 5 years of disease duration and fulfilled the American College of Rheumatology 1987 classification criteria for RA (40). Healthy controls, frequency-matched by birth year, were randomly selected from the Danish population, and blood samples were collected by general practitioners. Written informed consent was obtained from all study subjects, and the study was approved by the Scientific Ethical Committees for Copenhagen and Frederiksberg (KF 01-039/01), the Danish Data Protection Agency (2001–41–0658), and the Institutional Review Board at Statens Serum Institut (21–00050). The North American cohort included 200 RA patients and 100 age- and sex-matched controls without autoimmune disease selected from the University of Wisconsin (UW) Rheumatology Biorepository described in (35, 41) (Table 1). The RA patients were initially identified by having at least two outpatient visits with RA-associated ICD codes within 24 months (42), or one visit and a positive anti-CCP test. Diagnosis was confirmed by manual review of rheumatology notes. All subjects gave written informed consent, and the study was approved by the Institutional Review Board of the University of Wisconsin-Madison (#2015-0156).
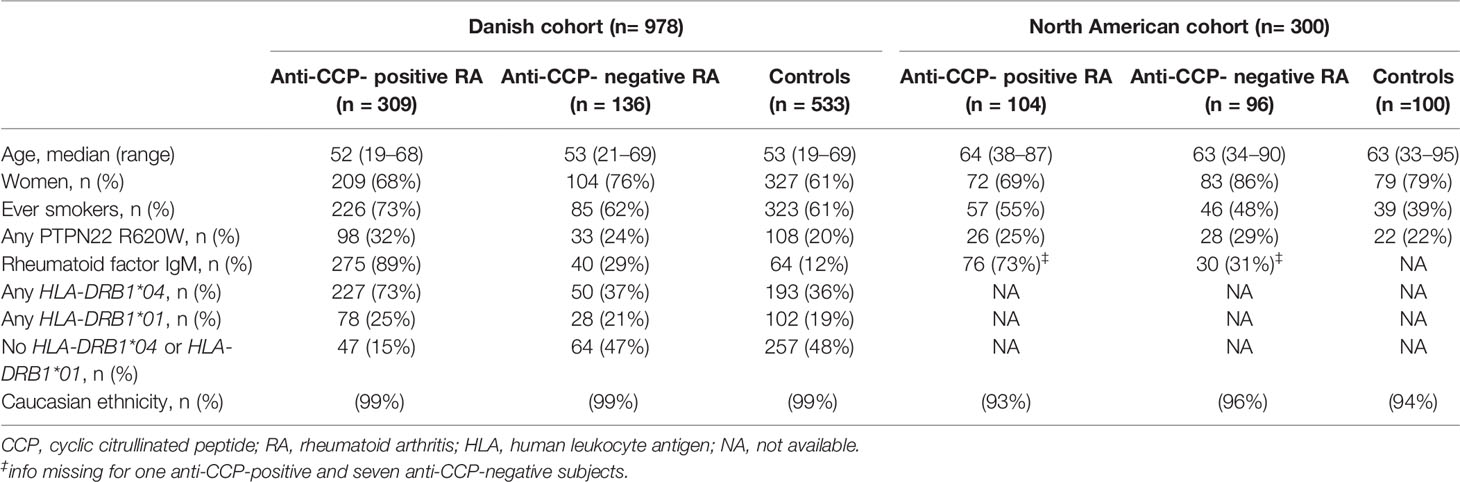
Table 1 Demographic, clinical, and genetic characteristics of anti-CCP-positive and anti-CCP-negative rheumatoid arthritis (RA) patients and controls.
SNP Selection
SNPs in PADI2 (rs2057094, rs2076616, rs2235912, and rs1005753) and PADI4 (rs74058715, rs11203367, rs1748033, and rs2240335) were selected based on literature search of previously reported associations with RA (22–27, 43). rs11203367 was chosen as a tag-SNP for the two other SNPs encoding amino acid substitutions in the RA susceptibility haplotype (rs11203366 and rs874881), given the strong linkage disequilibrium (LD) between these three SNPs (44). Additionally, as previously shown, rs11203367 is also in strong LD (r2 > 0.8), with the intronic SNPs rs2240340 and rs11203368 (44), two other SNPs associated with RA in previous studies (23, 31); therefore these intron located SNPs were excluded from typing. The PTPN22 missense SNP rs2476601 (R620W) was also included, given the reported association with RA and its known interactions with other risk factors like smoking and carriage of the SE (14). None of the SNPs investigated deviated significantly from Hardy-Weinberg equilibrium, in either cases or controls.
Genotyping
DNA was extracted from whole blood samples from the two cohorts as previously described (35, 39) and stored at −80°C until use. Samples were genotyped for PADI SNPs and the rs2476601 SNP in PTPN22, encoding R620W, by means of an in-house multiplex SNP assay protocol, as previously described (45). In brief, the method included a polymerase chain reaction (PCR) to amplify the selected SNP sites and an allele-specific primer extension (ASPE) reaction for labeling ASPE-oligonucleotides, followed by hybridization to MagPlex-TAG™ bead sets (Luminex Corporation, Austin, TX, USA) for analysis on the Luminex platform (Luminex Corporation, Austin, TX, USA). All assay runs included a panel of control samples with known genotypes (Coriell Cell Repository, Camden, NJ, USA) as well as no template negative controls. Low-resolution HLA-DRB1 typing of the Danish cohort had been performed prior to this study, according to the 11th Histocompatibility Workshop protocol (46).
Statistical Analysis
Hardy-Weinberg equilibrium was calculated with the “hwde”-package in R (R Foundation for Statistical Computing, Vienna, Austria). Associations between SNPs and disease were tested using multiple logistic regressions with adjustment for age, sex, smoking status, and carriage of PTPN22 R620W. P-values and odds ratios (ORs) with 95% confidence intervals (95% CI) were calculated for minor allele counts (trend test). Analyses of the Danish and North American cohorts were performed separately and, in order to increase statistical power, repeated after merging of the two cohorts. Post-hoc analyses after stratification for HLA-DRB1 alleles were performed in the Danish cohort, with further adjustment for the number of HLA-DRB1*04 or HLA-DRB1*01 alleles carried. Disease risk due to combined effects of PTPN22 R620W, HLA-DRB1 alleles, PADI4 alleles, and smoking was calculated by means of logistic-regression models, with adjustment for age and sex. Gene-gene interaction and gene-exposure interaction were calculated by logistic regression analyses including all single variables and interaction variables in the regression model. A full factorial set of interactions was presented to the model and only conditionally included if they significantly improved the predictive performance of the model. Analyses were performed in R Studio Version 1.0.153 (RStudio Inc., Boston, MA, USA) using R version 3.4.2 (R Foundation for Statistical Computing, Vienna, Austria) and in SPSS 25.0 (IBM, New York, USA). The level of statistical significance was set at p<0.05.
Results
The characteristics of the Danish and North American cohorts included in this study are shown in Table 1. The North American cohort had a median age of 63 years (range 33–95), which was higher than the 53 years (range 19–69) of the Danish cohort, and it contained more women than the Danish cohort, particularly within the anti-CCP-negative RA and healthy control groups. Anti-CCP-positive RA patients were more often smokers than anti-CCP-negative patients in both cohorts, but the Danish cohort contained more ever smokers than the North American cohort in all three study groups. All analyses were adjusted for age, sex, smoking status, and cohort origin.
Associations Between PADI Polymorphisms and RA
We studied associations of the PADI SNPs with anti-CCP-positive and anti-CCP-negative RA separately. Given that carriage of PTPN22 R620W is a known risk factor for anti-CCP-positive RA (14), we adjusted all analyses for this variable too. To increase statistical power, we merged the two cohorts in these analyses but adjusted the analyses for the origin of the cohort. We found that none of the PADI2 SNPs included in this study associated with anti-CCP-positive or anti-CCP-negative RA (Table 2).
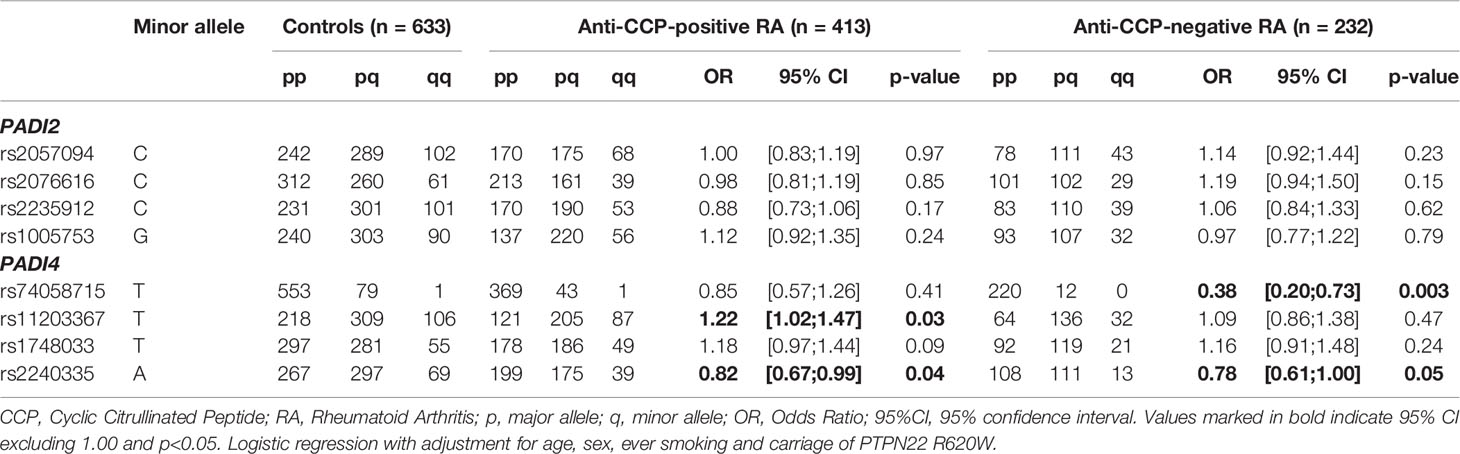
Table 2 Associations between selected polymorphisms in the PADI genes and anti-CCP-positive or anti-CCP-negative RA.
Among the PADI4 SNPs examined, the minor allele (T) of rs11203367 [the tag-SNP for the previously reported RA susceptibility haplotype (24, 30)], was associated with an increased risk of anti-CCP-positive RA (OR: 1.22, p=0.03) (Table 2). The minor allele (A) of rs2240335, on the other hand, appeared to protect against both anti-CCP-positive RA (OR: 0.82, p=0.04) and anti-CCP-negative RA (OR: 0.78, p=0.05). Moreover, the minor allele (T) of rs74058715 appeared to protect against anti-CCP-negative RA (OR: 0.38, p=0.003) (Table 2).
The majority of these reported associations was largely driven by the North American cohort, with exception of the association of rs74058715 and anti-CCP-negative RA (Supplementary Table 1).
Association of PADI Polymorphisms With RA by HLA-Type
The Danish cohort was HLA-typed, so we reanalyzed their data, taking this variable into account. We chose to examine the influence of HLA-DRB1*04 and HLA-DRB1*01 separately, since HLA-DRB1*04 is more strongly associated with anti-CCP-positive RA than HLA-DRB1*01 (12) (subjects were grouped as illustrated in Supplementary Figure 1; of note, subjects carrying both alleles were included in the analysis of HLA-DRB1*04).
Inclusion of HLA-DR type in the analysis did not influence the overall conclusions with respect to SNPs in PADI2 (data not shown). In HLA-DRB1*04-positive subjects, on the other hand, the minor allele (T) of rs74058715 was associated with a significantly reduced risk of anti-CCP-positive RA after adjustment for age, sex, smoking status, PTPN22 R620W, and number of HLA-DRB1*04 alleles (p=0.01; Table 3). This effect can be observed in Figure 2A, which shows the cumulative ORs associated with different combinations of four risk factors (HLA-DRB1*04 alleles, PADI4 alleles, PTPN22 R620W and smoking) compared to the reference group without any of these risk factors. Hence, the ORs for HLA-DRB1*04-positive subjects who carried the minor allele (light blue bars) were generally lower than the ORs for HLA-DRB1*04-positive subjects who did not carry this allele (dark blue bars). By contrast, the minor allele (T) of rs11203367 conferred a borderline significant increase in OR for anti-CCP-positive RA in HLA-DRB1*04-positive subjects (p=0.06, Table 3), as visualized in Figure 2B (light blue bars for carriers versus dark blue bars for non-carriers). Notably, rs1748033(T), which belongs to the same RA susceptibility haplotype as rs11203367(T), associated strongly with anti-CCP-positive RA in HLA-DRB1*04-positive individuals (p=0.007; Table 3), with higher cumulative ORs for carriers of this allele compared to non-carriers (Figure 2C, light blue bars vs dark blue bars).
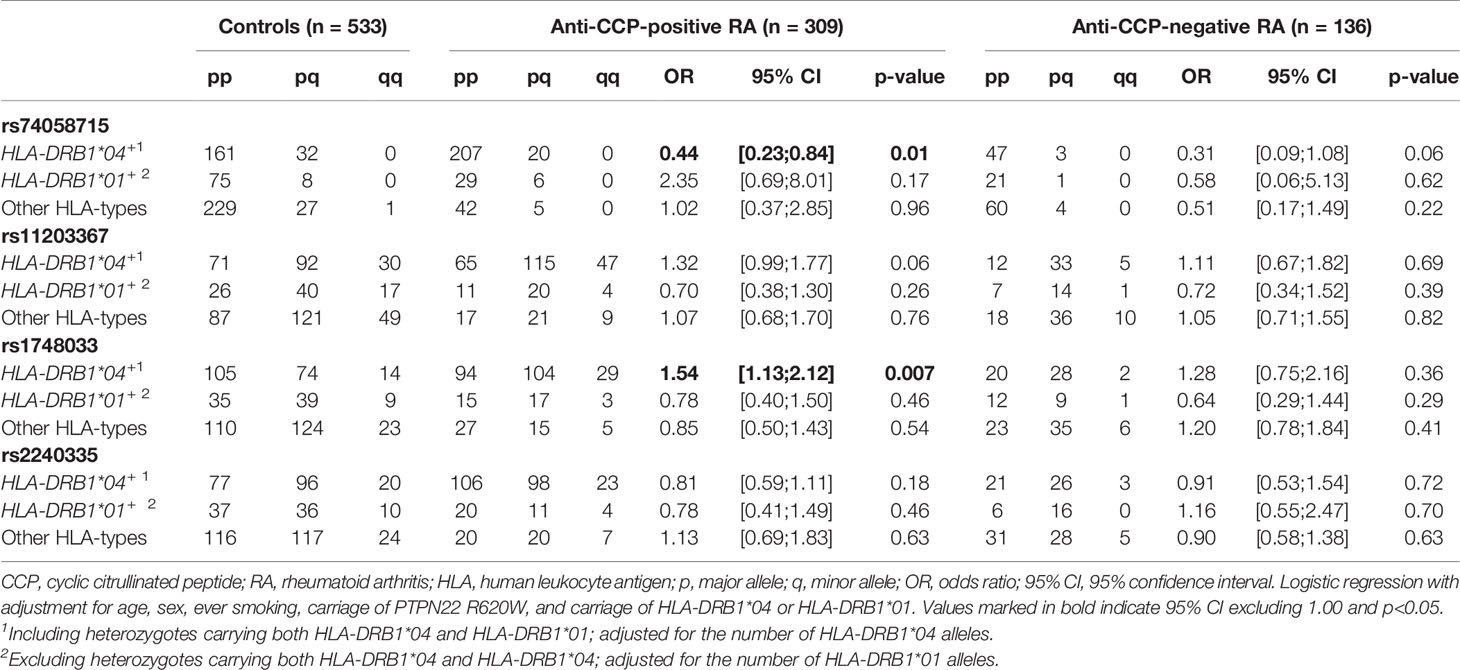
Table 3 Association of PADI4 SNPs with RA by HLA-type.
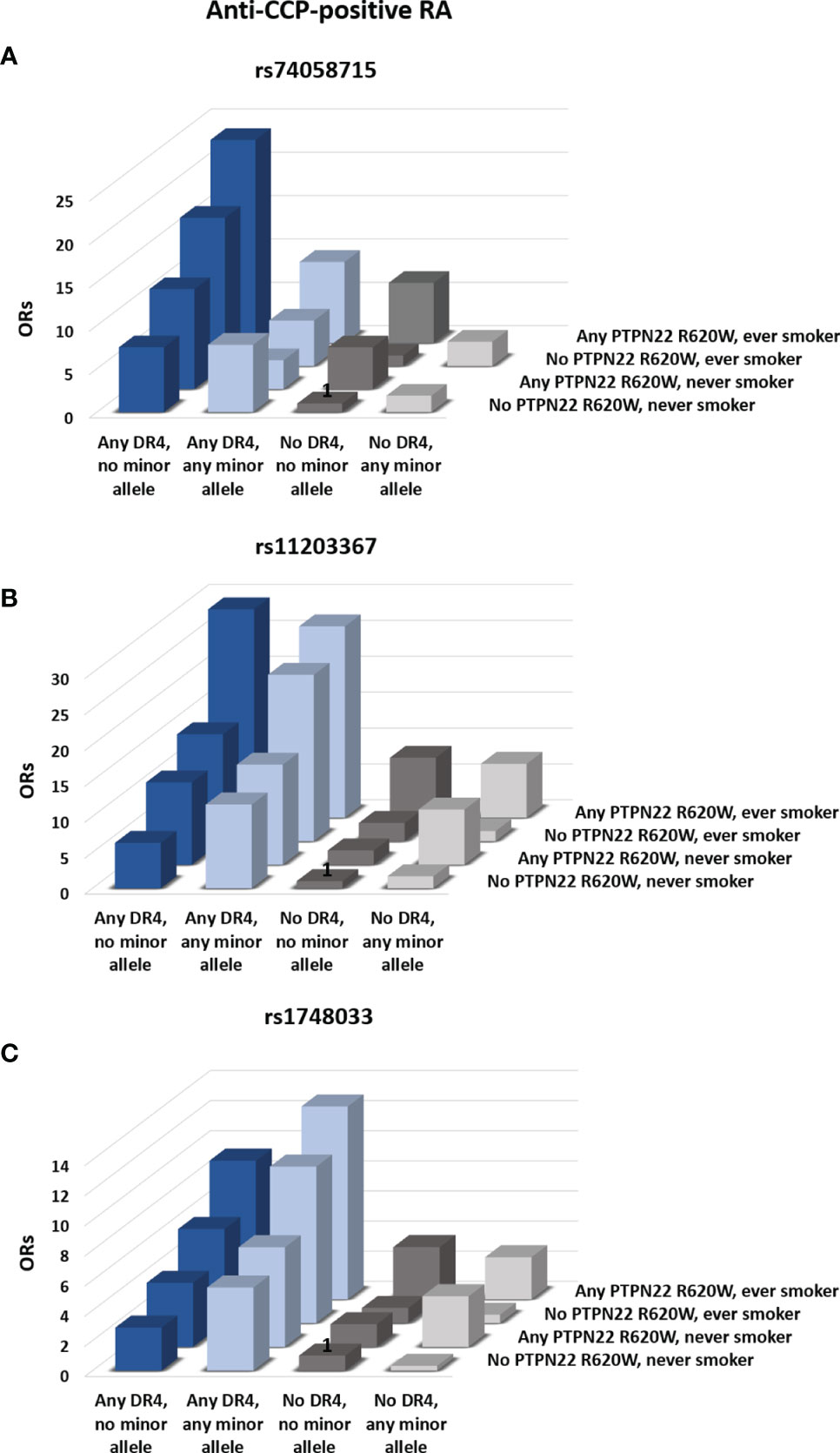
Figure 2 Influence of PADI4 SNPs on the risk of RA. The influence of the minor alleles of rs74058715 (A), rs11203367 (B), and rs1748033 (C) in PADI4 on the cumulative risk of anti-CCP–positive RA was assessed. Bars represent odds-ratios (ORs) for different combinations of absence or presence of any copy of the minor allele of the three PADI4 SNPs, any HLA-DRB1*04 allele, any PTPN22 R620W, and smoking. Subjects who carried HLA-DRB1*01 in combination with other HLA types than HLA-DRB1*04 were excluded from the analysis. Bars marked with “1” served as the reference condition. All ORs are adjusted for age and sex. ORs and p-values for all comparisons are reported in Supplementary Table 3.
No clear pattern was observed for HLA-DRB1*01-positive individuals or for subjects who carried neither HLA-DRB1*04 nor HLA-DRB1*01 (Table 3 and gray bars in Figure 2). Neither were any associations between SNPs in PADI and anti-CCP-negative RA revealed after adjustment for HLA type (Table 3).
Gene-Gene and Gene-Exposure Interactions
In view of the associations between SNPs in PADI4 and anti-CCP-positive RA and the known interaction between the SE and smoking as risk factors for anti-CCP-positive RA, we analyzed the interactions between PADI4 SNPs and other risk factors, i.e., HLA-DRB1*04, HLA-DRB1*01, PTPN22 R620W, and smoking on the risk of anti-CCP-positive or anti-CCP-negative RA.
We observed a strong interaction between the major allele (C) of rs74058715, HLA-DRB1*04 and ever-smoking, that associated with increased risk of anti-CCP-positive RA (p=0.008, Supplementary Table 2). Similarly, the minor allele (T) of rs1748033 and HLA-DRB1*04 interacted synergistically increasing the risk of anti-CCP-positive RA, irrespective of smoking status (p=0.004, Supplementary Table 2). Regarding the risk of anti-CCP-negative RA, we also observed an interaction between the minor allele (T) of rs1748033, HLA-DRB1*04 and ever-smoking (p=0.05, Supplementary Table 2).
Discussion
PAD2 and PAD4 are thought to be involved in the pathogenesis of ACPA-positive RA by catalyzing the formation of citrullinated autoantigens that are recognized by T cells and targeted by ACPAs (8). However, citrullination by PADs may have other potentially pathogenic roles, including regulation of T-cell responses (47, 48) and induction of NET formation (3, 49). Accordingly, SNPs in PADI2 and PADI4 have been shown to associate with RA, mainly in Asian populations (22–27).
This study did not show significant associations between PADI2 SNPs and RA in the two mainly Caucasian cohorts examined. We cannot rule out, however, that the minor allele (C) of rs2235912 confers resistance to anti-CCP-positive RA, as suggested by a weak tendency in that direction, which is in agreement with findings from a previous meta-analysis (43).
After merging the Danish and North American cohorts and adjusting the model for the origin of the cohorts, we found that the minor allele (T) of rs11203367 in PADI4 (tag-SNP for a haplotype containing Gly55Ser, Val82Ala, and Gly112Ala) associated with anti-CCP-positive RA with an OR of 1.22. The association was largely driven by the North American cohort, in accordance with a previous study showing association with rs2240340, which is in high LD with the SNP in our study (31). Furthermore, the minor allele (A) of the exon-synonymous SNP rs2240335 was associated with a decreased risk of anti-CCP-positive RA with an OR of 0.82, in agreement with previous results (35).
An especially strong association was observed in HLA-DRB1*04-positive subjects in the Danish cohort between the minor allele (T) of rs1748033 and anti-CCP-positive RA, and a similar tendency was observed for rs11203367, which is also part of RA the susceptibility haplotype. In individuals carrying HLA-DRB1*04, the minor (T) allele of rs74058715 was also associated with reduced risk of anti-CCP-positive RA with an OR of 0.44. This SNP is not part of the RA susceptibility haplotype but is associated with decreased expression of PAD4 in neutrophils and monocytes (36). These results suggest that SNPs in PADI4 alter the cumulative risk of disease conferred by other well-characterized risk factors (HLA-DRB1*04, PTPN22 R620W, and smoking). In accordance, we observed an interaction between the major allele (C) of rs74058715, HLA-DRB1*04, and ever-smoking, as well as between the minor allele (T) of rs1748033 and HLA-DRB1*04 in increasing the risk of anti-CCP-positive RA.
Somewhat unexpectedly, we observed that the minor (T) allele of rs74058715 was associated with reduced risk of anti-CCP-negative RA, and the minor allele (A) of rs2240335 showed a similar tendency. Furthermore, we observed an interaction between the minor allele (T) of rs1748033, HLA-DRB1*04, and ever-smoking in conferring risk of anti-CCP-negative RA. Generation of citrullinated autoantigens does not explain a pathogenic role for citrullination in ACPA-negative RA, but PADI polymorphisms may be associated with differences in citrullination patterns that may have an adverse influence on the generation of pro-inflammatory mediators (50), regulation of T-cell responses (47, 48), or NET formation (3, 49) potentially leading to tissue damage.
Our study is limited by the relatively low number of subjects included and the lack of HLA-typing of the North American cohort. Furthermore, only low-resolution HLA-typing was performed so the HLA-DRB1*04-positive group may have included individuals carrying HLA-DRB1*0402, which does not contain the SE and has been associated with a protective effect against RA (51).
It is important to note that although HLA-DRB1*04 and HLA-DRB1*01 are often grouped together as SE-containing serotypes, we chose to separate them in our analysis, since the association of HLA-DRB1*04 with RA is considerably stronger than that of HLA-DRB1*01 (12). We found no association between SNPs in PADI and RA in HLA-DRB1*01-positive individuals, and no interaction between PADI4 SNPs and HLA-DRB1*01, which may be due to the low power of the study. On the other hand, cohort stratifications and lack of correction due to the exploratory nature of the study may have led to type I errors. Thus, similar and more robust analyses in larger cohorts are warranted.
Our study suggests that certain SNPs in PADI4 are risk factors for ACPA-positive RA, especially in HLA-DRB1*04-positive individuals. Furthermore, it shows that PADI4 SNPs may also play a role in ACPA-negative RA.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics Statement
The studies involving human participants were reviewed and approved by the Scientific Ethical Committees for Copenhagen and Frederiksberg (KF 01-039/01), the Danish Data Protection Agency (2001–41–0658), the Institutional Review Board at Statens Serum Institut (21–00050), and the Institutional Review Board of the University of Wisconsin–Madison (#2015-0156). The patients/participants provided their written informed consent to participate in this study.
Author Contributions
LM, CN, DD, MS, MF, PG, SJ, and CE designed the study. LM and PG carried out the experiments. LM, CE, and SJ carried out the statistical analyses. LM, CN, and CE drafted the manuscript. SJ, MF, and MS provided the sample material and the clinical data for the Danish and North American cohorts, respectively. DD, NØ, and MS revised the manuscript critically. All authors contributed to the article and approved the submitted version.
Funding
This study was provided by the Independent Research Fund, Denmark (grant number DFF - 7016-00233) to CN as well as from the University of Wisconsin School of Medicine and Public Health from the Wisconsin Partnership Program (grant number 2951) and the Doris Duke Charitable Foundation (grant number 2015099) to MS. SJ was supported by grants from the Danish Rheumatism Association (grant number A3865) and the Novo Nordisk Foundation. MS was additionally supported by NIH/NIAMS (grant number K08AR065500) and U.S. Army Medical Research Acquisition Activity through the Peer Reviewed Medical Research Program (grant number W81XWH-18-1-0717). The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors thank MSc Pia Grothe Meinke for excellent technical assistance and BSc (Hons), PhD Robert Graham Quinton Leslie for editing the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.707690/full#supplementary-material
Abbreviations
ACPAs, anti-citrullinated protein antibodies; CCP, cyclic citrullinated peptide; HLA, human leukocyte antigen; NETs, neutrophil extracellular traps; PAD, peptidylarginine deiminase; RA, rheumatoid arthritis; SE, shared epitope; SNP, single nucleotide polymorphism.
References
1. Senshu T, Kan S, Ogawa H, Manabe M, Asaga H. Preferential Deimination of Keratin K1 and Filaggrin During the Terminal Differentiation of Human Epidermis. Biochem Biophys Res Commun (1996) 719:712–9. doi: 10.1006/bbrc.1996.1240
2. Falcão AM, Meijer M, Scaglione A, Rinwa P, Agirre E, Liang J, et al. PAD2-Mediated Citrullination Contributes to Efficient Oligodendrocyte Differentiation and Myelination. Cell Rep (2019) 27(4):1090–102.e10. doi: 10.1016/j.celrep.2019.03.108
3. Holmes CL, Shim D, Kernien J, Johnson CJ, Nett JE, Shelef MA. Insight Into Neutrophil Extracellular Traps Through Systematic Evaluation of Citrullination and Peptidylarginine Deiminases. J Immunol Res (2019) 2019. doi: 10.1155/2019/2160192
4. Schellekens GA, De Jong BAW, Van Den Hoogen FHJ, Van De Putte LBA, Van Venrooij WJ. Citrulline Is an Essential Constituent of Antigenic Determinants Recognized by Rheumatoid Arthritis-Specific Autoantibodies. J Clin Invest (1998) 101(1):273–81. doi: 10.1172/JCI1316
5. Schellekens GA, Visser H, De Jong BA, Van Den Hoogen FH, Hazes JM, Breedveld FC, et al. The Diagnostic Properties of Rheumatoid Arthritis Antibodies Recognizing a Cyclic Citrullinated Peptide. Arthritis Rheum (2000) 43(1):155–63. doi: 10.1002/1529-0131(200001)43:1<155::AID-ANR20>3.0.CO;2-3
6. Rantapää-Dahlqvist S, De Jong BAW, Berglin E, Hallmans G, Wadell G, Stenlund H, et al. Antibodies Against Cyclic Citrullinated Peptide and IgA Rheumatoid Factor Predict the Development of Rheumatoid Arthritis. Arthritis Rheum (2003) 48(10):2741–9. doi: 10.1002/art.11223
7. van der Helm-van Mil AHM, Verpoort KN, Breedveld FC, Toes REM, Huizinga TWJ. Antibodies to Citrullinated Proteins and Differences in Clinical Progression of Rheumatoid Arthritis. Arthritis Res Ther (2005) 7(5):R949–58. doi: 10.1186/ar1767
8. Hill JA, Southwood S, Sette A, Jevnikar AM, Bell DA, Cairns E. Cutting Edge: The Conversion of Arginine to Citrulline Allows for a High-Affinity Peptide Interaction With the Rheumatoid Arthritis-Associated HLA-DRB1*0401 MHC Class II Molecule. J Immunol (2003) 171(2):538–41. doi: 10.4049/jimmunol.171.2.538
9. Klareskog L, Stolt P, Lundberg K, Källberg H, Bengtsson C, Grunewald J, et al. A New Model for an Etiology of Rheumatoid Arthritis: Smoking may Trigger HLA-DR (Shared Epitope)-Restricted Immune Reactions to Autoantigens Modified by Citrullination. Arthritis Rheum (2006) 54(1):38–46. doi: 10.1002/art.21575
10. Ding B, Padyukov L, Lundström E, Seielstad M, Plenge RM, Oksenberg JR, et al. Different Patterns of Associations With Anti–Citrullinated Protein Antibody–Positive and Anti–Citrullinated Protein Antibody– Negative Rheumatoid Arthritis in the Extended Major Histocompatibility Complex Region. Arthritis Rheum (2009) 60(1):30–8. doi: 10.1002/art.24135
11. Pedersen M, Jacobsen S, Garred P, Madsen HO, Klarlund M, Svejgaard A, et al. Strong Combined Gene-Environment Effects in Anti-Cyclic Citrullinated Peptide-Positive Rheumatoid Arthritis: A Nationwide Case-Control Study in Denmark. Arthritis Rheum (2007) 56(5):1446–53. doi: 10.1002/art.22597
12. Lundstr̈om E, Källberg H, Alfredsson L, Klareskog L, Padyukov L. Gene-Environment Interaction Between the DRB1 Shared Epitope and Smoking in the Risk of Anti-Citrullinated Protein Antibody-Positive Rheumatoid Arthritis: All Alleles are Important. Arthritis Rheum (2009) 60(6):1597–603. doi: 10.1002/art.24572
13. Begovich AB, Carlton VEH, Honigberg LA, Schrodi SJ, Chokkalingam AP, Alexander HC, et al. Report A Missense Single-Nucleotide Polymorphism in a Gene Encoding a Protein Tyrosine Phosphatase (PTPN22) Is Associated With Rheumatoid Arthritis. Am J Hum Genet (2004) 75(2):330–7. doi: 10.1086/422827
14. Källberg H, Padyukov L, Plenge RM, Rönnelid J, Gregersen PK, van der Helm-van Mil AHM, et al. Gene-Gene and Gene-Environment Interactions Involving HLA-BRB1, PTPN22, and Smoking in Two Subsets of Rheumatoid Arthritis. Am J Hum Genet (2007) 80(5):867–75. doi: 10.1086/516736
15. Vossenaar ER, Zendman AJW, Van Venrooij WJ, Pruijn GJM. PAD, a Growing Family of Citrullinating Enzymes: Genes, Features and Involvement in Disease. BioEssays (2003) 25:1106–18. doi: 10.1002/bies.10357
16. Asaga H, Nakashima K, Senshu T, Ishigami A. Immunocytochemical Localization of Peptidylarginine Deiminase in Human Eosinophils and Neutrophils. J Leuk Biol (2001) 70(1):46–51.
17. Vossenaar ER, Radstake TRD, van der Heijden A, van Mansum MAM, Dieteren C, de Rooij D-J, et al. Expression and Activity of Citrullinating Peptidylarginine Deiminase Enzymes in Monocytes and Macrophages. Ann Rheum Dis (2004) 63(4):373–81. doi: 10.1136/ard.2003.012211
18. Foulquier C, Sebbag M, Clavel C, Chapuy-Regaud S, Al Badine R, Méchin MC, et al. Peptidyl Arginine Deiminase Type 2 (PAD-2) and PAD-4 But Not PAD-1, PAD-3, and PAD-6 are Expressed in Rheumatoid Arthritis Synovium in Close Association With Tissue Inflammation. Arthritis Rheum (2007) 56(11):3541–53. doi: 10.1002/art.22983
19. Damgaard D, Senolt L, Nielsen MF, Pruijn GJ, Nielsen CH. Demonstration of Extracellular Peptidylarginine Deiminase (PAD) Activity in Synovial Fluid of Patients With Rheumatoid Arthritis Using a Novel Assay for Citrullination of Fibrinogen. Arthritis Res Ther (2014) 16(6):498. doi: 10.1186/s13075-014-0498-9
20. Blachère NE, Parveen S, Frank MO, Dill BD, Molina H, Orange DE. High-Titer Rheumatoid Arthritis Antibodies Preferentially Bind Fibrinogen Citrullinated by Peptidylarginine Deiminase 4. Arthritis Rheumatol (2017) 69(5):986–95. doi: 10.1002/art.40035
21. Nakayama-Hamada M, Suzuki A, Kubota K, Takazawa T, Ohsaka M, Kawaida R, et al. Comparison of Enzymatic Properties Between Hpadi2 and Hpadi4. Biochem Biophys Res Commun (2005) 327(1):192–200. doi: 10.1016/j.bbrc.2004.11.152
22. Too CL, Murad S, Dhaliwal JS, Larsson P, Jiang X, Ding B, et al. Polymorphisms in Peptidylarginine Deiminase Associate With Rheumatoid Arthritis in Diverse Asian Populations: Evidence From MyEIRA Study and Meta-Analysis. Arthritis Res Ther (2012) 14(6):R250. doi: 10.1186/ar4093
23. Chang X, Xia Y, Pan J, Meng Q, Zhao Y, Yan X. PADI2 is Significantly Associated With Rheumatoid Arthritis. PloS One (2013) 8(12):e81259. doi: 10.1371/journal.pone.0081259
24. Suzuki A, Yamada R, Chang X, Tokuhiro S, Sawada T, Suzuki M, et al. Functional Haplotypes of PADI4, Encoding Citrullinating Enzyme Peptidylarginine Deiminase 4, are Associated With Rheumatoid Arthritis. Nat Genet (2003) 34(4):395–402. doi: 10.1038/ng1206
25. Lee YH, Rho YH, Choi SJ, Ji JD, Song GG. PADI4 Polymorphisms and Rheumatoid Arthritis Susceptibility: A Meta-Analysis. Rheumatol Int (2007) 27(9):827–33. doi: 10.1007/s00296-007-0320-y
26. Terao C, Yamada R, Ohmura K, Takahashi M, Kawaguchi T, Kochi Y, et al. The Human AIRE Gene at Chromosome 21q22 is a Genetic Determinant for the Predisposition to Rheumatoid Arthritis in Japanese Population. Hum Mol Genet (2011) 20(13):2680–5. doi: 10.1093/hmg/ddr161
27. Freudenberg J, Lee HS, Han BG, Shin HD, Kang YM, Sung YK, et al. Genome-Wide Association Study of Rheumatoid Arthritis in Koreans: Population-Specific Loci as Well as Overlap With European Susceptibility Loci. Arthritis Rheum (2011) 63(4):884–93. doi: 10.1002/art.30235
28. Gandjbakhch F, Fajardy I, Ferré B, Dubucquoi S, Flipo RM, Roger N, et al. A Functional Haplotype of PADI4 Gene in Rheumatoid Arthritis: Positive Correlation in a French Population. J Rheumatol (2009) 36(5):881–6. doi: 10.3899/jrheum.080398
29. Martinez A, Valdivia A, Pascual-Salcedo D, Lamas JR, Fernandez-Arquero M, Balsa A, et al. PADI4 Polymorphisms Are Not Associated With Rheumatoid Arthritis in the Spanish Population. Rheumatol (Oxford) (2005) 44(10):1263–6. doi: 10.1093/rheumatology/kei008
30. Barton A, Bowes J, Eyre S, Spreckley K, Hinks A, John S, et al. A Functional Haplotype of the PADI4 Gene Associated With Rheumatoid Arthritis in a Japanese Population Is Not Associated in a United Kingdom Population. Arthritis Rheum (2004) 50(4):1117–21. doi: 10.1002/art.20169
31. Plenge RM, Padyukov L, Remmers EF, Purcell S, Lee AT, Karlson EW, et al. Replication of Putative Candidate-Gene Associations With Rheumatoid Arthritis in >4,000 Samples From North America and Sweden: Association of Susceptibility With PTPN22, CTLA4, and PADI4. Am J Hum Genet (2005) 77(6):1044–60. doi: 10.1086/498651
32. Burr ML, Naseem H, Hinks A, Eyre S, Gibbons LJ, Bowes J, et al. PADI4 Genotype Is Not Associated With Rheumatoid Arthritis in a Large UK Caucasian Population. Ann Rheum Dis (2010) 69(4):666–70. doi: 10.1136/ard.2009.111294
33. Hoppe B, Häupl T, Gruber R, Kiesewetter H, Burmester GR, Salama A, et al. Detailed Analysis of the Variability of Peptidylarginine Deiminase Type 4 in German Patients With Rheumatoid Arthritis: A Case-Control Study. Arthritis Res Ther (2006) 8(2):R34. doi: 10.1186/ar1889
34. Farago B, Talian GC, Maasz A, Magyari L, Horvatovich K, Kovacs B, et al. Prevalence of Functional Haplotypes of the Peptidylarginine Deiminase Citrullinating Enzyme Gene in Patients With Rheumatoid Arthritis: No Influence of the Presence of Anti-Citrullinated Peptide Antibodies. Clin Exp Rheumatol (2007) 25(4):523–8.
35. Mergaert MA, Bawadekar M, Nguyen QT, Massarenti L, Holmes LC, Rebernick R, et al. Reduced Anti-Histone Antibodies and Increased Risk of Rheumatoid Arthritis Associated With a Single Nucleotide Polymorphism in PADI4 in North Americans. Vol. 20 Int J Mol Sci (2019) p:3093. doi: 10.3390/ijms20123093
36. Naranbhai V, Fairfax BP, Makino S, Humburg P, Wong D, Ng E, et al. Genomic Modulators of Gene Expression in Human Neutrophils. Nat Commun (2015) 6(May):1–13. doi: 10.1038/ncomms8545
37. Bang SY, Han TU, Choi CB, Sung YK, Bae SC, Kang C. Peptidyl Arginine Deiminase Type IV (PADI4) Haplotypes Interact With Shared Epitope Regardless of Anti-Cyclic Citrullinated Peptide Antibody or Erosive Joint Status in Rheumatoid Arthritis: A Case Control Study. Arthritis Res Ther (2010) 12(3):R115. doi: 10.1186/ar3051
38. Poór G, Nagy Z, Schmidt Z, Brózik M, Merétey K, Gergely P. Genetic Background of Anticyclic Citrullinated Peptide Autoantibody Production in Hungarian Patients With Rheumatoid Arthritis. Ann N Y Acad Sci (2007) 1110:23–32. doi: 10.1196/annals.1423.004
39. Pedersen M, Jacobsen S, Klarlund M, Pedersen BV, Wiik A, Wohlfahrt J, et al. Environmental Risk Factors Differ Between Rheumatoid Arthritis With and Without Auto-Antibodies Against Cyclic Citrullinated Peptides. Arthritis Res Ther (2006) 8(4):R133. doi: 10.1186/ar2022
40. Arnett FC, Edworthy SM, Bloch DA, Mcshane DJ, Fries JF, Cooper NS, et al. The American Rheumatism Association 1987 Revised Criteria for the Classification of Rheumatoid Arthritis. Arthritis Rheumatol (1988) 31(3):315–24. doi: 10.1002/art.1780310302
41. Rebernick R, Fahmy L, Glover C, Bawadekar M, Shim D, Holmes CL, et al. DNA Area and NETosis Analysis (DANA): A High-Throughput Method to Quantify Neutrophil Extracellular Traps in Fluorescent Microscope Images. Biol Proced Online (2018) 20(1):1–9. doi: 10.1186/s12575-018-0072-y
42. Katz JN, Barrett J, Liang MH, Bacon AM, Kaplan H, Kieval RI, et al. Sensitivity and Positive Predictive Value of Medicare Part B Physician Claims for Rheumatologic Diagnoses and Procedures. Arthritis Rheum (1997) 40(9):1594–600. doi: 10.1002/art.1780400908
43. Okada Y, Wu D, Trynka G, Raj T, Terao C, Ikari K, et al. Genetics of Rheumatoid Arthritis Contributes to Biology and Drug Discovery. Nature (2014) 506(7488):376–81. doi: 10.1038/nature12873
44. Massarenti L, Enevold C, Damgaard D, Ødum N, Nielsen CH, Jacobsen S. Peptidylarginine Deiminase-4 Gene Polymorphisms are Associated With Systemic Lupus Erythematosus and Lupus Nephritis. Scand J Rheumatol (2019) 48(2):133–40. doi: 10.1080/03009742.2018.1488273
45. Enevold C, Oturai AB, Sørensen PS, Ryder LP, Koch-Henriksen N, Bendtzen K. Multiple Sclerosis and Polymorphisms of Innate Pattern Recognition Receptors TLR1-10, NOD1-2, DDX58, and IFIH1. J Neuroimmunol (2009) 212(1–2):125–31. doi: 10.1016/j.jneuroim.2009.04.008
46. Kimura A, Sasazuki T. Eleventh International Histocompatibility Workshop Reference Protocol for the HLA DNA-Typing Technique, in: Tsuji K, Azaiwa M, Sazasuki T (eds): HLA 1991. Proceedings of the Eleventh International Histocompatibility Workshop and Conference, vol I, Oxford, Oxford Science Publications, (1992) pp. 397–419.
47. Kawalkowska J, Quirke AM, Ghari F, Davis S, Subramanian V, Thompson PR, et al. Abrogation of Collagen-Induced Arthritis by a Peptidyl Arginine Deiminase Inhibitor Is Associated With Modulation of T Cell-Mediated Immune Responses. Sci Rep (2016) 6(May):1–12. doi: 10.1038/srep26430
48. Sun B, Chang HH, Salinger A, Tomita B, Bawadekar M, Holmes CL, et al. Reciprocal Regulation of Th2 and Th17 Cells by PAD2-Mediated Citrullination. JCI Insight (2019) 4(22):e129687. doi: 10.1172/jci.insight.129687
49. Khandpur R, Carmona-rivera C, Vivekanandan-giri A, Yalavarthi S, Knight JS, Friday S, et al. NETs Are a Source of Citrullinated Autoantigens and Stimulate Inflammatory Responses in Rheumatoid Arthritis. Sci Transl Med (2013) 5(178):178ra40. doi: 10.1126/scitranslmed.3005580
50. Sun B, Dwivedi N, Bechtel TJ, Paulsen JL, Muth A, Bawadekar M, et al. Citrullination of NF-kB P65 Promotes Its Nuclear Localization and TLR-Induced Expression of IL-1β and TNF-α. Sci Immunol (2017) 2:1–13. doi: 10.1126/sciimmunol.aal3062
Keywords: rheumatoid arthritis, peptidylarginine deiminase (PAD), anti-citrullinated proteins antibodies (ACPA), HLA-DRB1*04, single nucleotide polymorphism (SNP)
Citation: Massarenti L, Enevold C, Damgaard D, Ødum N, Garred P, Frisch M, Shelef MA, Jacobsen S and Nielsen CH (2021) PADI4 Polymorphisms Confer Risk of Anti-CCP-Positive Rheumatoid Arthritis in Synergy With HLA-DRB1*04 and Smoking. Front. Immunol. 12:707690. doi: 10.3389/fimmu.2021.707690
Received: 10 May 2021; Accepted: 14 September 2021;
Published: 18 October 2021.
Edited by:
Dimitrios Petrou Bogdanos, University of Thessaly, GreeceReviewed by:
Aleksandra Antovic, Karolinska Institutet, SwedenMei-Ling Yang, Yale University, United States
Copyright © 2021 Massarenti, Enevold, Damgaard, Ødum, Garred, Frisch, Shelef, Jacobsen and Nielsen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Søren Jacobsen, c29lcmVuLmphY29ic2VuLjAxQHJlZ2lvbmguZGs=
†These authors share senior authorship