 Xavier Solanich1*†
Xavier Solanich1*† Gardenia Vargas-Parra2,3†
Gardenia Vargas-Parra2,3† Caspar I. van der Made4,5,6†
Caspar I. van der Made4,5,6† Annet Simons4Janneke Schuurs-Hoeijmakers4
Annet Simons4Janneke Schuurs-Hoeijmakers4 Arnau Antolí1Jesús del Valle2,3
Arnau Antolí1Jesús del Valle2,3 Gemma Rocamora-Blanch1Fernando Setién7Manel Esteller3,7,8,9Simon V. van Reijmersdal4Antoni Riera-Mestre1,10Joan Sabater-Riera11
Gemma Rocamora-Blanch1Fernando Setién7Manel Esteller3,7,8,9Simon V. van Reijmersdal4Antoni Riera-Mestre1,10Joan Sabater-Riera11 Gabriel Capellá2,3
Gabriel Capellá2,3 Frank L. van de Veerdonk5,6
Frank L. van de Veerdonk5,6 Ben van der Hoven12
Ben van der Hoven12 Xavier Corbella1,13
Xavier Corbella1,13 Alexander Hoischen4,5,6*Conxi Lázaro2,3*
Alexander Hoischen4,5,6*Conxi Lázaro2,3*- 1Department of Internal Medicine, Hospital Universitari de Bellvitge, Bellvitge Biomedical Research Institute (IDIBELL), L’Hospitalet de Llobregat, Barcelona, Spain
- 2Hereditary Cancer Program, Catalan Institute of Oncology, Program in Molecular Mechanisms and Experimental Therapy in Oncology (Oncobell), Bellvitge Biomedical Research Institute (IDIBELL), L’Hospitalet de Llobregat, Barcelona, Spain
- 3Centro de Investigación Biomédica en Red de Cáncer (CIBERONC), Madrid, Spain
- 4Department of Human Genetics, Radboud University Medical Center, Nijmegen, Netherlands
- 5Department of Internal Medicine and Radboud Center for Infectious Diseases (RCI), Radboud Institute for Molecular Life Sciences (RIMLS), Radboud University Medical Center, Nijmegen, Netherlands
- 6Radboud Expertise Center for Immunodeficiency and Autoinflammation and Radboud Center for Infectious Disease (RCI), Radboud University Medical Center, Nijmegen, Netherlands
- 7Josep Carreras Leukaemia Research Institute (IJC), Badalona, Spain
- 8Institucio Catalana de Recerca i Estudis Avançats (ICREA), Barcelona, Spain
- 9Physiological Sciences Department, School of Medicine and Health Sciences, University of Barcelona (UB), Barcelona, Spain
- 10Faculty of Medicine and Health Sciences, Universitat de Barcelona, Barcelona, Spain
- 11Department of Intensive Care, Hospital Universitari de Bellvitge, Bellvitge Biomedical Research Institute (IDIBELL), L’Hospitalet de Llobregat, Barcelona, Spain
- 12Department of Intensive Care, Erasmus MC, Rotterdam, Netherlands
- 13School of Medicine, Universitat Internacional de Catalunya, Barcelona, Spain
Introduction: Loss-of-function TLR7 variants have been recently reported in a small number of males to underlie strong predisposition to severe COVID-19. We aimed to determine the presence of these rare variants in young men with severe COVID-19.
Methods: We prospectively studied males between 18 and 50 years-old without predisposing comorbidities that required at least high-flow nasal oxygen to treat COVID-19. The coding region of TLR7 was sequenced to assess the presence of potentially deleterious variants.
Results: TLR7 missense variants were identified in two out of 14 patients (14.3%). Overall, the median age was 38 (IQR 30-45) years. Both variants were not previously reported in population control databases and were predicted to be damaging by in silico predictors. In a 30-year-old patient a maternally inherited variant [c.644A>G; p.(Asn215Ser)] was identified, co-segregating in his 27-year-old brother who also contracted severe COVID-19. A second variant [c.2797T>C; p.(Trp933Arg)] was found in a 28-year-old patient, co-segregating in his 24-year-old brother who developed mild COVID-19. Functional testing of this variant revealed decreased type I and II interferon responses in peripheral mononuclear blood cells upon stimulation with the TLR7 agonist imiquimod, confirming a loss-of-function effect.
Conclusions: This study supports a rationale for the genetic screening for TLR7 variants in young men with severe COVID-19 in the absence of other relevant risk factors. A diagnosis of TLR7 deficiency could not only inform on treatment options for the patient, but also enables pre-symptomatic testing of at-risk male relatives with the possibility of instituting early preventive and therapeutic interventions.
Introduction
A proportion of patients with COVID-19 develop fatal lung injury and multi-organ failure due to systemic host-immune inflammatory processes triggered by the viral infection (1). Advanced age, male sex and chronic disease such as diabetes and obesity are common in patients with more severe forms of COVID-19 (2–4). However, these risk factors cannot explain why critical disease also occurs in young (below 50 years of age) and apparently healthy individuals.
In the past months, several publications have identified loci and genes associated with COVID-19 susceptibility for severe COVID-19 by using comprehensive GWAS studies or genome, exome or candidate gene analyses (5). However, most of these susceptibility alleles showed risk values too low (odds ratio <2) to be regarded as predictive genomic markers. Some of the loci reported include ABO blood group (6, 7), ACE2 (8), TMPRSS2 (8), several HLA alleles (5, 9), APOE (10), and IFITM3 (11). On the other hand, rare variants in genes encoding for members of the type I/III interferon (IFN) pathway showed a higher estimated risk to severe COVID-19 (12, 13).
In July 2020 rare, deleterious germline variants in the X-chromosomal Toll-like receptor 7 (TLR7) gene were reported in young and, otherwise, healthy males with severe COVID-19. In these two brother pairs, rapid whole-exome sequencing identified both a maternally inherited 4-nucleotide deletion [c.2129_2132del; p.(Gln710Argfs*18)] and a missense variant [c.2383G>T; p.(Val795Phe)]. Both variants were associated with impaired type I and II IFN responses upon stimulation with the TLR7 agonist imiquimod, supporting the importance of intact TLR7 signaling in COVID-19 pathogenesis (13). Most recently these findings were replicated in an independent Italian cohort study of males <60 years of age with severe COVID-19 (79 severe cases versus 77 control cases), showing that 2.1% of severely affected males harbored deleterious TLR7 variants compared to none of the asymptomatic participants. These variants were demonstrated to decrease transcription of type I and II IFN-related genes in patient peripheral blood mononuclear cells (PBMC) treated with imiquimod, supporting a loss of function effect (14).
In follow-up of the recent discovery of TLR7 deficiency in patients with severe COVID-19, we aimed to prospectively determine the presence of TLR7 variants in young and previously healthy men with severe COVID-19.
Methods
This is a joint study performed at the Hospital Universitari de Bellvitge – IDIBELL, L’Hospitalet de Llobregat, Barcelona, Spain; and the Radboud University Medical Center, Nijmegen, The Netherlands and the Erasmus Medical Center, Rotterdam, The Netherlands.
The Barcelona Cases
From March to July 2020, researchers from Hospital Universitari de Bellvitge - IDIBELL prospectively collected biological samples from young patients without comorbidities related to severe COVID-19. Selection criteria were: 1) patients aged between 18 and 50 years old; 2) absence of known comorbidities associated with most severe forms of COVID-19 (BMI ≥ 30kg/m2, diabetes mellitus, hypertension, chronic heart, pulmonary or kidney disease, or an immunocompromised state); and 3) SARS-CoV-2 related lung injury requiring high flow oxygen devices or mechanical ventilation. Ten male patients fulfilled these selection criteria (Table 1). Eight patients (patients 1-8) were also evaluated by the COVID Human Genetic Effort, although no pathogenic variants were identified in any of the 13 type I IFN pathway genes studied (12).
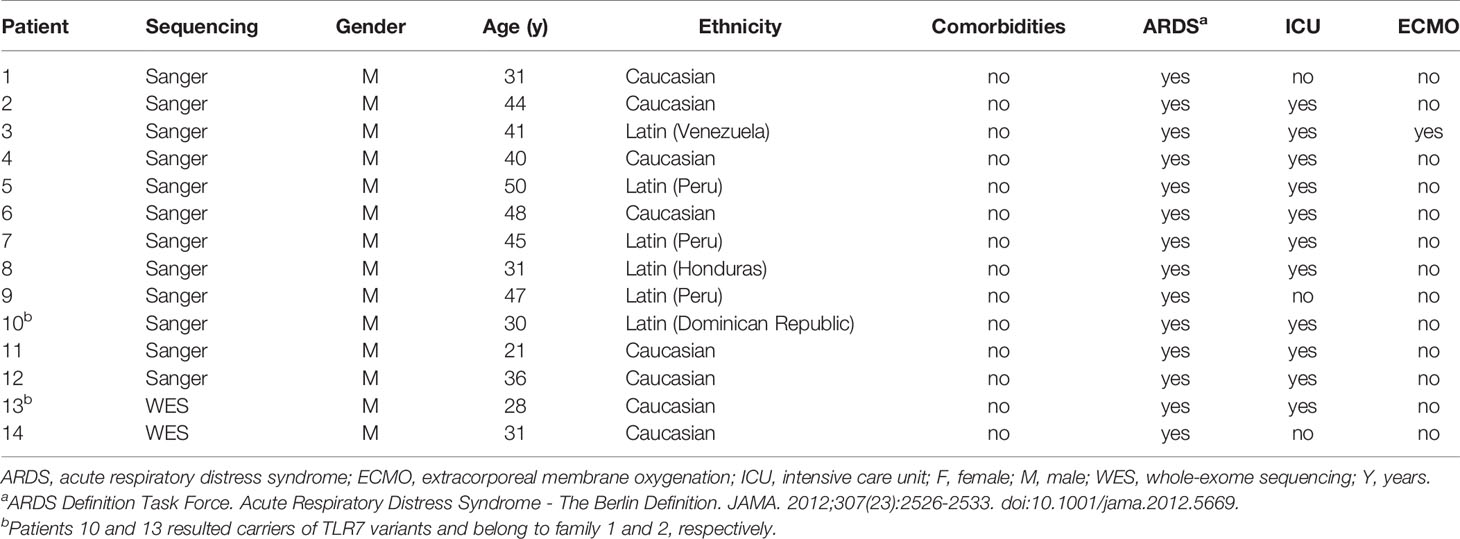
Table 1 Demographic and clinical findings of investigated patients.
Informed consent was obtained from all patients and relatives, and the IDIBELL Research Ethics Committee approved this study (PR152/20). Demographic, epidemiological, laboratory and clinical data were collected. Treatments specifically used to treat COVID-19 at any time during admission were also documented.
DNA was isolated from total blood either using a Maxwell instrument RSC (Promega, Madison, WI, USA) or QIAGEN Flexigene DNA kit (Qiagen, Germany). Nine PCR primer pairs (Sigma-Aldrich, MO, USA) were designed to cover the whole coding region of TLR7 (HGNC ID:15631). PCR was performed using DreamTaq MasterMix (ThermoFisher Scientific, Waltham, MA, USA), products were purified using EXO-SAP (New England Biolabs) and sequenced using the BigDye Terminator v.3.1 Sequencing Kit (Applied Biosystems, CA, USA) in an ABI Prism 3730 XL Genetic Analyzer (Applied Biosystems CA, USA). Primers and PCR conditions are available upon request. Mutation Surveyor software was used to detect variants and nomenclature was given according to HGVS guidelines. All variants identified were submitted to Alamut Software Suite v2.15.0 (Interactive Biosoftware) to retrieve population frequency and in silico prediction data.
The Dutch Cases
At the Radboud University Medical Center in Nijmegen and the Erasmus Medical Center in Rotterdam, the Netherlands, patients were screened prospectively in a clinical setting from December 2020 to February 2021 with the following criteria: 1) males between 18 and 40 years of age; 2) absence of comorbidities known to be associated with severe COVID-19 (BMI ≥ 30kg/m2, diabetes mellitus, hypertension, chronic heart, pulmonary or kidney disease, or an immunocompromised state); and 3) PCR-confirmed SARS-CoV-2 infection requiring high-flow oxygen therapy or ICU admission. A total of 4 patients (patients 11-14) fulfilled these inclusion criteria and underwent clinical Sanger sequencing (patients 11 and 12) or rapid whole-exome sequencing (patient 13 and 14) to specifically assess genetic variants in TLR7 (Table 1). Written informed consent was obtained from patient 13 whose clinical data has been included in this study. Rapid whole-exome sequencing was performed similar to previous reports (13). Sanger sequencing was done according to standard diagnostic procedures at the Department of Human Genetics, Radboud University Medical Center, protocols and primers sequences are available upon request.
For patient 13, in whom a rare TLR7 missense variant was identified, functional testing was performed to evaluate impaired type I and II IFN responses due to TLR7 loss-of-function, as described previously (13). In brief, venous blood was drawn and collected in EDTA tubes (Monject). Subsequently, peripheral mononuclear blood cells (PBMC) were isolated by density centrifugation of blood, diluted 1:1 in pyrogen-free saline over Ficoll-Paque (Pharmacia Biotech). Cells were washed twice in saline and resuspended in cell culture medium (Roswell Park Memorial Institute [RPMI] 1640, Gibco) supplemented with gentamicin, 10 mg/mL; L-glutamine, 10 mM; and pyruvate, 10 mM. PBMC were then stimulated at 5 × 105 cells/well in round-bottom 96-wells plates (Greiner) for either 4 hours (for transcription of type I IFN genes) and 7 days [for production of the type II IFN, (IFNγ)] in the presence of 10% human pool serum at 37°C and 5% carbon dioxide, in two separate experiments. Apart from a negative medium (RPMI) control, the TLR7 agonist imiquimod (imidazoquinoline compound, Invivogen) was used at a concentration of 5 μg/mL. After the 4 hour incubation period, the supernatants were discarded and the remaining cell pellets were resuspended in RLT buffer (Qiagen) and snap frozen to be stored at −80°C until processing for RNA isolation. In addition, after the 7 day incubation and a centrifugation step, supernatants were collected and stored at −20°C until measured using enzyme-linked immunosorbent assay in case of the 7 day timepoint.
Results
TLR7 Sequencing Results
A total of 14 patients were included, with a median age of 38 (IQR 30-45) years. Putative deleterious TLR7 missense variants were identified in two patients (patient 10 and 13). Both variants, c.644A>G; p.(Asn215Ser) in patient 10 and c.2797T>C; p.(Trp933Arg) in patient 13, were not previously reported in our in-house databases nor in the population database gnomAD (15). The p.(Asn215Ser) variant affects a highly conserved nucleotide and amino acid in the TLR7 leucine-rich region domain and is predicted damaging or possibly damaging by in silico software (Table S1). The p.(Trp933Arg) variant is also located at an evolutionarily highly conserved position within the TIR domain, important for downstream signaling via adapter proteins, and is considered deleterious by in silico effect predictors (Table S1). These TLR7 variants and other previously reported variants in COVID-19 patients are shown schematically in Figure 1.
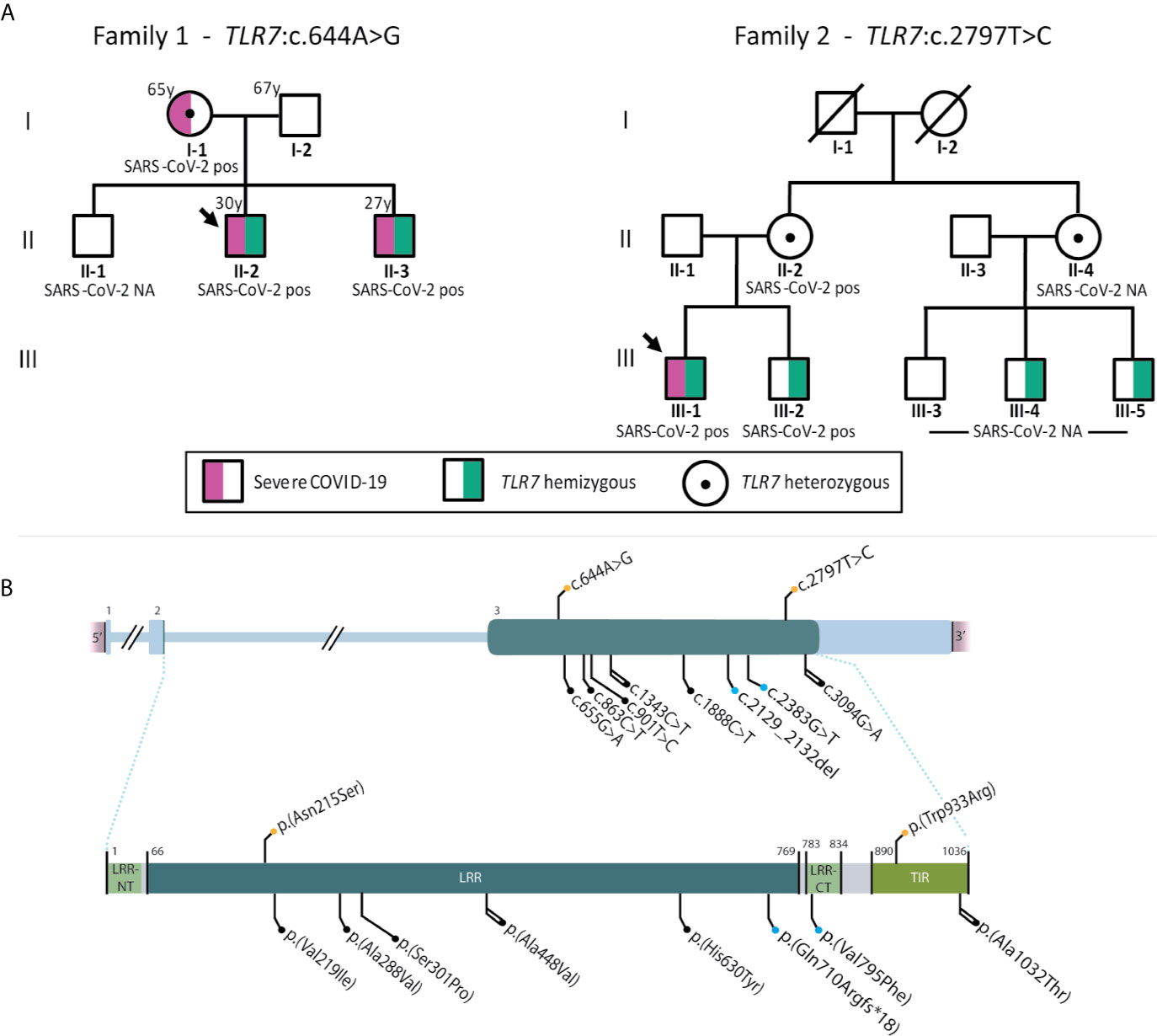
Figure 1 (A) displays pedigrees of families 1 (patient 10) and 2 (patient 13) with segregation analysis. (B) shows schematic representation of TLR7 (HGNC ID:15631) variants reported to date in severely affected COVID-19 cases. Variants in cDNA (top) and protein (bottom). Color code: orange, variants found in the present series; blue, previously reported in van der Made (13); black, previously reported in Fallerini (14). Shape code: circle, missense variants; square, frameshift variants. Line code: single, reported in one case; double, reported in 2 cases.
Patients’ Characteristics
Patient 10 (family 1, proband) was a 30-year-old man from Latin origin (Dominican Republic) with no general risk factors predisposing to severe COVID-19. He developed pneumonia with bilateral consolidations on a computed tomography (CT) scan and fulfilled the criteria of acute respiratory distress syndrome (ARDS) secondary to PCR-proven COVID-19 (Table 2). The patient received antiviral treatment with remdesivir and immunosuppressive therapy with dexamethasone. Due to respiratory insufficiency, the patient was intubated and admitted in ICU. The patient was successfully extubated after 4 days of mechanical ventilation and was discharged from ICU after 6 days. After the identification of the TLR7:p.(Asn215Ser) variant in the patient, segregation analysis confirmed co-segregation in both his brother and mother, in hemi- and heterozygous state, respectively (Figure 1). The eldest brother lives in the Dominican Republic and was therefore not available for testing. According to relatives, there is no evidence that he has been infected with SARS-CoV-2. The youngest brother had no previous medical history but also contracted severe COVID-19, requiring mechanical ventilation and ICU admission at another hospital (Table 2). These brothers therefore represent the third brother pair with severe COVID-19, following the initial report (13). The 65 year-old mother suffered from obesity, dyslipidemia, type 2 diabetes and hypertension, and was also admitted in ICU due to critical respiratory failure caused by COVID-19. She was discharged from the ICU 16 days after admission. Main demographic, clinical, laboratory, and radiological findings of the three relatives are summarized in Table 2.
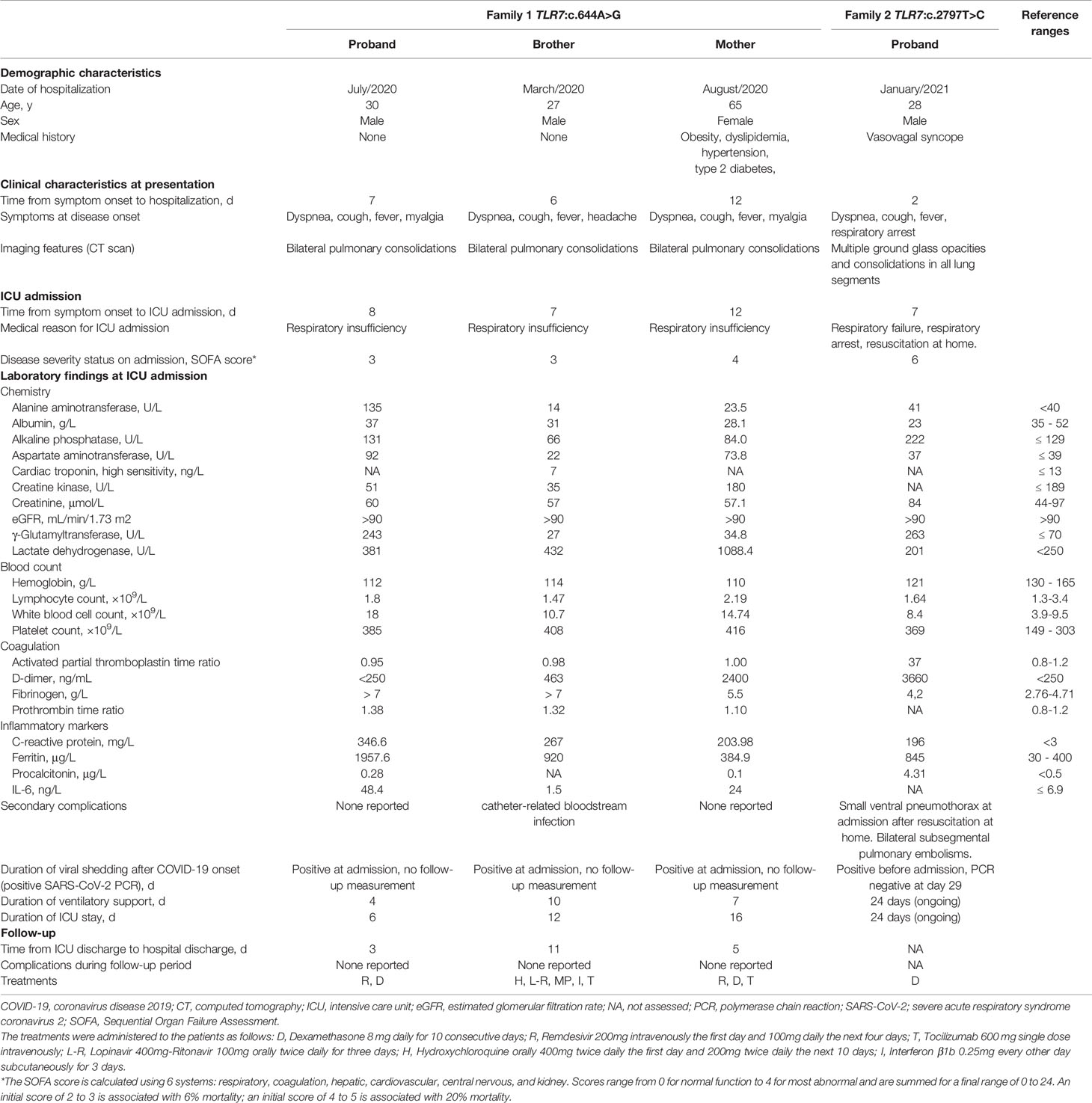
Table 2 Demographic, clinical, laboratory, and radiological findings of investigated patients.
Patient 13 (family 2, proband) was a 28-year-old male from Caucasian origin (The Netherlands) without previous medical history or comorbidities. The patient complained of progressive dyspnea and shortness of breath. Following rapid clinical deterioration and respiratory insufficiency, the patient was intubated and hospitalized in ICU at a peripheral hospital. A CT-scan showed multiple diffuse ground-glass opacities and consolidations in all lung segments, meeting the criteria for ARDS. Treatment consisted of mechanical ventilation with prone positioning, intravenous dexamethasone, and the antibiotics ceftriaxone and ciprofloxacin. Despite this therapeutic regiment, the patient’s condition further deteriorated and he was referred to the Erasmus Medical Center for possible ECMO treatment. However, with continuing prone positioning he gradually improved before ECMO was required. A repeated CT scan also showed subsegmental pulmonary embolisms for which intravenous heparin was started. In the following weeks, the patient gradually recovered and was successfully extubated and eventually discharged from the hospital. The patients’ first-degree family contracted COVID-19 at the time the patient developed symptoms, including his 24-year-old brother, who had only minor symptoms but was shown to be a carrier of the TLR7 variant (Figure 2B). Information on the exact SARS-CoV-2 viral titers during infections were unavailable. In addition, two male cousins, sons of a maternal aunt, were proven to be carriers of this variant. These individuals had not contracted (symptomatic) COVID-19. Based on the TLR7 variant carriership, these individuals were fast-tracked for early vaccination.
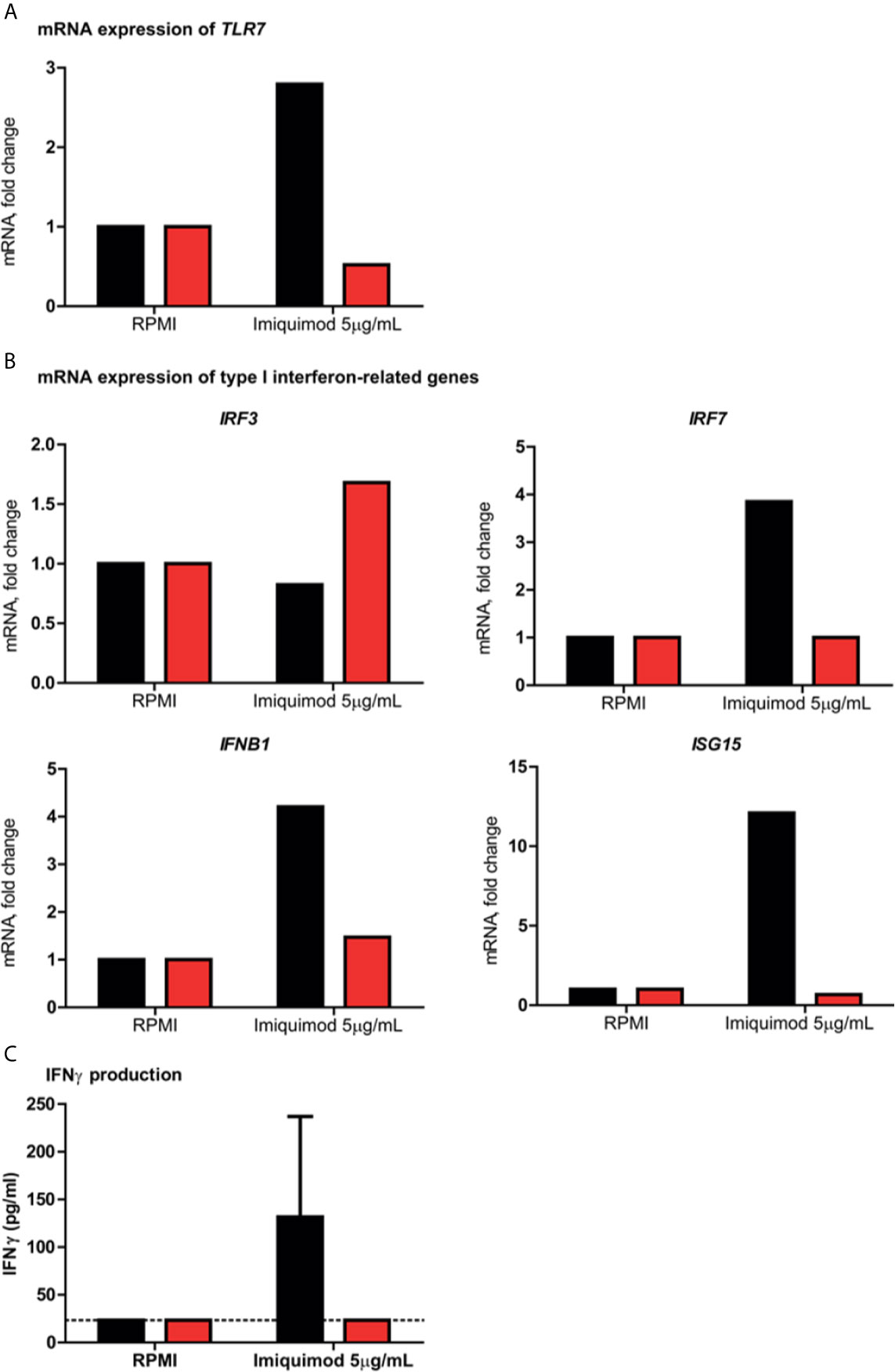
Figure 2 Assessment of ex vivo/in vitro type I and II interferon responses in peripheral blood mononuclear cells. In all panels, patient 13 is indicated in red and the healthy controls in black. (A) shows the fold change in TLR7 mRNA expression in patient 13 and a healthy male control after stimulation for 4 hours with the TLR7 agonist imiquimod, compared to the negative medium control RPMI. (B) displays the fold change in mRNA expression of type I IFN–related genes IRF3, IRF7, IFNB1 and ISG15 induced by imiquimod as compared to RPMI. (C) shows the production of IFN-γ production after imiquimod stimulation for 7 days as compared with unstimulated control cells (RPMI) from patient 13 and two healthy male controls. The dotted line indicates the assay’s detection limit. Abbreviations: IRF3, interferon regulatory factor 3; IRF7, interferon regulatory factor 7; ISG15, interferon-stimulated gene 15; and IFNB1, interferon beta 1.
Functional Testing in Primary Immune Cells
In PBMC isolated from patient 13, the type I and II IFN responses were evaluated upon stimulation with the TLR7 agonist imiquimod, in order to assess the functional impact of the p.(Trp933Arg) variant. It was shown that TLR7 stimulation in the patient resulted in a defective upregulation of TLR7 mRNA expression, as well as that of the type I IFN-related genes involved in the TLR7 pathway IRF7, IFNB1 and ISG15, as compared to a healthy male control (Figures 2A, B). Expression of IRF3, a transcription factor that is induced by TLR3 and not TLR7, showed a modest increase of almost 1.7 fold in the patient, but was not induced in the healthy control. Moreover, the production of IFNγ upon exposure with imiquimod was deficient in the patient, compared to two healthy male controls (Figure 2C).
Discussion
In this cohort of young males affected with severe COVID-19 rare TLR7 variants were prospectively identified in 2 out of 14 cases (14.3%). In 10 cases from Spain, one patient was shown to carry a new missense variant [c.644A>G; p.(Asn215Ser)]. The variant was predicted as damaging by in silico programs and segregated in a brother who also contracted severe COVID-19. In addition, four Dutch cases were studied, leading to the identification of another patient with a novel, unique missense variant [c.2797T>C; p.(Trp933Arg)] located in the highly conserved TIR domain. Functional analysis of the latter variant demonstrated defective type I and II IFN responses, similar to those documented in the previous reports (13, 14).
A role for the TLR7 receptor has been described in the host defense against single-stranded RNA viruses as well as in the pathogenesis of autoimmune disease such as SLE (16). However, no human mutations in TLR7 had been reported before the SARS-CoV-2 pandemic. It should be noted that complete TLR7 deficiency is estimated to be extremely rare, because endosomal TLRs (TLR3, TLR7, TLR8, and TLR9) play an essential, non-redundant biological role in host survival (17, 18). Therefore, these rare TLR7 variants are unlikely to be an explanation for severe COVID-19 in the general population. Distinct from the previously published studies that have identified (enrichment of) rare TLR7 variants in severe COVID-19 who were 1) males <35 years of age without comorbidities (13), 2) males <60 years of age with or without comorbidities (14) or 3) a group of unselected patients (19), in this study we have selected males <50 years of age without comorbidities predisposing to severe COVID-19. Despite the small cohort, the yield of TLR7 variants was unexpectedly high (14.3%) encouraging that screening for TLR7 rare variants in severely affected men may be fruitful. Elderly individuals also carry rare, loss of function TLR7 variants (14), but are more difficult to identify due to other predisposing general risk factors for severe disease in people of advanced age. Therefore, we suggest the following screening criteria: young men (<50 year of age) suffering from severe COVID-19 requiring at least high-flow oxygen therapy, and who were previously healthy. Affected young brother pairs, as well as pedigrees suggestive for X-linked segregation, should be further prioritized.
We furthermore hypothesize that some rare variants compromise essential functional domains in TLR7 and lead to full TLR7 deficiency (20), while other less rare variants might lead only to a partial TLR7 deficiency, and consequently impact a larger group of individuals, but exert a lower relative risk to develop severe COVID-19. Accordingly, these low effect size genetic variants in TLR7 have been proposed as a possible explanation of the male sex bias in COVID-19 severity because of its localization on the X chromosome and well-established function in innate immunity (21). Furthermore, it is possible, but only speculative, that the addition of factors that deteriorate TLR7 function [e.g. circulating 25-hydroxy vitamin D [25OHD] levels decline with age (22) and may be accompanied by a lower TLR7 expression and defective function (23)] in patients with these low effect variants may be a common cause of progression to the most severe stages of COVID-19 in male. The same speculative hypothesis could be applicable to other genes involved in immune response regulation after SARS-CoV-2 infection (4, 24).
Strong host genetic factors that confer an increased risk to develop severe COVID-19 might serve as genomic biomarkers, along with other factors, that could be used for early diagnosis and preventative measures, and could allow the identification of possible molecular targets for treatment. In this respect, there would be a strong argument to offer hemizygous TLR7 deficient males that have not had COVID-19 direct access to early vaccination as an effective preventative measure, similar to other patients with primary immunodeficiencies. This option has indeed been offered to the hemizygous carriers in the family of patient 13. Although no data is yet available to support specific management of TLR7 deficiency or the at-risk hemizygous carriers, early hospitalization and IFN-based therapies in inborn errors of IFN signaling form rational treatment options (18, 25).
This study has several limitations. First, no biological samples were available to functionally validate the pathogenicity of the p.(Asn215Ser) variant and therefore the functional impact of this variant remains unknown. Nevertheless, the absence of this variant in population databases, the high prediction scores for pathogenicity and the general scarcity of rare variants in TLR7 (13) suggest this finding is unlikely due to chance. Second, the TLR7 variant identified in patient 13 segregates in a brother who only experienced mild COVID-19. There are some explanations that may explain why a pathogenic variant in TLR7 does not become fully penetrant, since protection against SARS-CoV-2 depends on other genetic and environmental factors (e.g. exposure to a lower initial viral load, previous exposure to common cold coronaviruses) (26). Moreover, TLR7 function might be specifically influenced by epidemiological factors and certain comorbidities (e.g. 25OHD). Thirdly, the size of this study does not permit to draw any firm conclusions on the prevalence of loss of function TLR7 variants in males with severe COVID-19. Hence, larger cohort studies are required. Most recently, in pan-ancestry whole-exome sequencing data of 586,157 individuals, encompassing 20.592 patients who contracted COVID-19 and 1.266 of those who had severe disease, TLR7 was the only gene in which the burden of rare variants was significantly increased among patients with severe COVID-19, using a less conservative significance threshold [OR 4.53 (2.64, 7.77)] (19). This is one of the few occasions in which a Mendelian disease gene is replicated through the use of an association study, even without stratifying for male gender or younger age.
In summary, we have identified two novel germline variants in the X chromosomal TLR7 that likely lead to TLR7 deficiency, which is further corroborated by impaired type I and II IFN responses in the patient with the p.(Trp933Arg) variant. These findings reinforce the notion that TLR7 plays a critical role in the recognition of SARS-CoV-2 and the subsequent initiation of an early antiviral immune response that could prevent the development of severe COVID-19. More importantly, a better understanding of strong genetic factors predisposing to severe COVID-19 may help physicians to identify patients at risk. We therefore propose clinical screening criteria for the identification of TLR7 variants in male patients with severe COVID-19. This could not only enable targeted therapeutic management of the patient, but could also offer relatives the option of pre-symptomatic testing and by extension preventative measures such as early vaccination.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The studies involving human participants were reviewed and approved by IDIBELL Research Ethics Committee (approval number PR152/20). The patients/participants provided their written informed consent to participate in this study. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
Author Contributions
XS, GV-P, and CM contributed equally to this work. XS, AH, and CL devised the study. GC, CM, AR-M, FS, ME, and XC provided input on the study design. AA, BH, JS-H, FV, and G-RB assisted in patient management. GV-P, AS, JV, and SR designed and performed the sequence and functional analysis. XS, GV-P, CM, AH, and CL had full access to all data and take responsibility for the integrity and the accuracy of the data. XS, GV-P, CM, AH, and CL drafted the manuscript. All authors contributed to the article and approved the submitted version.
Funding
Contract grant sponsor: Supported by La Marató de TV3 foundation (202115-30-31), the Carlos III National Health Institute funded by FEDER funds – a way to build Europe – [PI19/00553 and CIBERONC]; the Government of Catalonia [2017SGR1282 and 2017SGR496]. AH is supported by the Solve-RD project. The Solve-RD project has received funding from the European Union’s Horizon 2020 research and innovation programme under grant agreement No. 779257. FV was supported by a ZonMW (The Netherlands Organization for Health Research and Development) Vidi grant (No. 91718351). This research was part of the Netherlands X-omics Initiative and partially funded by NWO (The Netherlands Organization for Scientific Research; project 184.034.019).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We sincerely thank the patients and their families for their participation. With the support of COVID-19 funding from the Departament de Salut de la Generalitat de Catalunya. We thank CERCA Programme/Generalitat de Catalunya for institutional support, and the unstoppable campaign of the Josep Carreras Leukaemia Foundation and the Cellex Foundation. We also thank the Radboud Genomics Technology Center for their technical support.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.719115/full#supplementary-material
References
1. Siddiqi HK, Mehra MR. COVID-19 Illness in Native and Immunosuppressed States: A Clinical-Therapeutic Staging Proposal. J Heart Lung Transplant (2020) 39(5):405–7. doi: 10.1016/j.healun.2020.03.012
2. Del Sole F, Farcomeni A, Loffredo L, Carnevale R, Menichelli D, Vicario T, et al. Features of Severe COVID-19: A Systematic Review and Meta-Analysis. Eur J Clin Invest (2020) 50(10):e13378. doi: 10.1111/eci.13378
3. Rubio-Rivas M, Corbella X, Mora-Luján JM, Loureiro-Amigo J, López Sampalo A, Yera Bergua C, et al. Predicting Clinical Outcome With Phenotypic Clusters in COVID-19 Pneumonia: An Analysis of 12,066 Hospitalized Patients From the Spanish Registry SEMI-COVID-19. J Clin Med (2020) 9(11):3488. doi: 10.3390/jcm9113488
4. Anastassopoulou C, Gkizarioti Z, Patrinos GP, Tsakris A. Human Genetic Factors Associated With Susceptibility to SARS-CoV-2 Infection and COVID-19 Disease Severity. Hum Genomics (2020) 14(1):40. doi: 10.1186/s40246-020-00290-4
5. Novelli G, Biancolella M, Mehrian-Shai R, Colona VL, Brito AF, Grubaugh ND, et al. COVID-19 One Year Into the Pandemic: From Genetics and Genomics to Therapy, Vaccination, and Policy. Hum Genomics (2021) 15(1):27. doi: 10.1186/s40246-021-00326-3
6. Ellinghaus D, Degenhardt F, Bujanda L, Buti M, Albillos A, Invernizzi P, et al. Genomewide Association Study of Severe Covid-19 With Respiratory Failure. N Engl J Med (2020) 383(16):1522–34. doi: 10.1056/NEJMoa2020283
7. Zhao J, Yang Y, Huang H, Li D, Gu D, Lu X, et al. Relationship Between the ABO Blood Group and the COVID-19 Susceptibility. Clin Infect Dis (2020) 4:ciaa1150. doi: 10.1093/cid/ciaa1150.
8. Hou Y, Zhao J, Martin W, Kallianpur A, Chung MK, Jehi L, et al. New Insights Into Genetic Susceptibility of COVID-19: An ACE2 and TMPRSS2 Polymorphism Analysis. BMC Med (2020) 18(1):216. doi: 10.1186/s12916-020-01673-z
9. Nguyen A, David JK, Maden SK, Wood MA, Weeder BR, Nellore A, et al. Human Leukocyte Antigen Susceptibility Map for Severe Acute Respiratory Syndrome Coronavirus 2. J Virol (2020) 94(13):e00510–20. doi: 10.1128/JVI.00510-20
10. Kuo CL, Pilling LC, Atkins JL, Masoli JAH, Delgado J, Kuchel GA, et al. APOE E4 Genotype Predicts Severe COVID-19 in the UK Biobank Community Cohort. J Gerontol A Biol Sci Med Sci (2020) Oct 1575(11):2231–2. doi: 10.1093/gerona/glaa131
11. Thevarajan I, Nguyen THO, Koutsakos M, Druce J, Caly L, van de Sandt CE, et al. Breadth of Concomitant Immune Responses Prior to Patient Recovery: A Case Report of non-Severe COVID-19. Nat Med (2020) 26(4):453–5. doi: 10.1038/s41591-020-0819-2
12. Zhang Q, Bastard P, Liu Z, Le Pen Z, Moncada-Velez Z, Chen Z, et al. Inborn Errors of Type I IFN Immunity in Patients With Life-Threatening COVID-19. Science (2020) 370(6515):eabd4570. doi: 10.1126/science.abd4570
13. van der Made CI, Simons A, Schuurs-Hoeijmakers J, van den Heuvel G, Mantere T, Kersten S, et al. Presence of Genetic Variants Among Young Men With Severe COVID-19. JAMA (2020) 324(7):663–73. doi: 10.1001/jama.2020.13719
14. Fallerini C, Daga S, Mantovani S, Benetti E, Picchiotti N, Francisci D, et al. Association of Toll-Like Receptor 7 Variants With Life-Threatening COVID-19 Disease in Males: Findings From a Nested Case-Control Study. Elife (2021) 10:e67569. doi: 10.7554/eLife.67569
15. Karczewski KJ, Francioli LC, Tiao G, Cummings BB, Alföldi J, Wang Q, et al. The Mutational Constraint Spectrum Quantified From Variation in 141,456 Humans. Nature (2020) 581(7809):434–43. doi: 10.1038/s41586-020-2308-7
16. Shen N, Fu Q, Deng Y, Qian X, Zhao J, Kaufman KM, et al. Sex-Specific Association of X-Linked Toll-Like Receptor 7 (TLR7) With Male Systemic Lupus Erythematosus. Proc Natl Acad Sci USA (2010) 107(36):15838–43. doi: 10.1073/pnas.1001337107
17. Casanova JL, Abel L, Quintana-Murci L. Human TLRs and IL-1Rs in Host Defense: Natural Insights From Evolutionary, Epidemiological, and Clinical Genetics. Annu Rev Immunol (2011) 29:447–91. doi: 10.1146/annurev-immunol-030409-101335
18. Plenge RM. Molecular Underpinnings of Severe Coronavirus Disease 2019. JAMA (2020). doi: 10.1001/jama.2020.14015
19. Kosmicki JA, Horowitz JE, Banerjee N, Lanche R, Marcketta A, Maxwell E, et al. Pan-Ancestry Exome-Wide Association Analyses of COVID-19 Outcomes in 586, 157 Individuals. Am J Hum Genet (2021) 108(7):1350–5. doi: 10.1016/j.ajhg.2021.05.017
20. van de Veerdonk FL, Netea MG. Rare Variants Increase the Risk of Severe COVID-19. Elife (2021) 10:e67860. doi: 10.7554/eLife.67860
21. Scully EP, Haverfield J, Ursin RL, Tannenbaum C, Klein SL. Considering How Biological Sex Impacts Immune Responses and COVID-19 Outcomes. Nat Rev Immunol (2020) 20(7):442–7. doi: 10.1038/s41577-020-0348-8
22. Pérez-López FR, Chedraui P, Fernández-Alonso AM. Vitamin D and Aging: Beyond Calcium and Bone Metabolism. Maturitas (2011) 69(1):27–36. doi: 10.1016/j.maturitas.2011.02.014
23. Alvarez-Rodriguez L, Lopez-Hoyos M, Garcia-Unzueta M, Amado JA, Cacho PM, Martinez-Taboada VM. Age and Low Levels of Circulating Vitamin D are Associated With Impaired Innate Immune Function. J Leukoc Biol (2012) 91(5):829–38. doi: 10.1189/jlb.1011523
24. Breidenbach JD, Dube P, Ghosh S, Abdullah BN, Modyanov NN, Malhotra D, et al. Impact of Comorbidities on SARS-CoV-2 Viral Entry-Related Genes. J Pers Med (2020) 10(4):E146. doi: 10.3390/jpm10040146
25. Lévy R, Bastard P, Lanternier F, Lecuit M, Zhang SY, Casanova JL. IFN-α2a Therapy in Two Patients With Inborn Errors of TLR3 and IRF3 Infected With SARS-CoV-2. J Clin Immunol (2021) 41(1):26–7. doi: 10.1007/s10875-020-00933-0
Keywords: COVID-19, SARS-CoV-2, host genetics, TLR7, immunodeficiency, genetic screening
Citation: Solanich X, Vargas-Parra G, van der Made CI, Simons A, Schuurs-Hoeijmakers J, Antolí A, del Valle J, Rocamora-Blanch G, Setién F, Esteller M, van Reijmersdal SV, Riera-Mestre A, Sabater-Riera J, Capellá G, van de Veerdonk FL, van der Hoven B, Corbella X, Hoischen A and Lázaro C (2021) Genetic Screening for TLR7 Variants in Young and Previously Healthy Men With Severe COVID-19. Front. Immunol. 12:719115. doi: 10.3389/fimmu.2021.719115
Received: 01 June 2021; Accepted: 08 July 2021;
Published: 23 July 2021.
Edited by:
Antonio Condino-Neto, University of São Paulo, BrazilReviewed by:
Alessandro Aiuti, Vita-Salute San Raffaele University, ItalyAlexandra Freeman, National Institutes of Health (NIH), United States
Giuseppe Novelli, University of Rome Tor Vergata, Italy
Copyright © 2021 Solanich, Vargas-Parra, van der Made, Simons, Schuurs-Hoeijmakers, Antolí, del Valle, Rocamora-Blanch, Setién, Esteller, van Reijmersdal, Riera-Mestre, Sabater-Riera, Capellá, van de Veerdonk, van der Hoven, Corbella, Hoischen and Lázaro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xavier Solanich, eHNvbGFuaWNoQGJlbGx2aXRnZWhvc3BpdGFsLmNhdA==; Alexander Hoischen, QWxleGFuZGVyLkhvaXNjaGVuQHJhZGJvdWR1bWMubmw=; Conxi Lázaro, Y2xhemFyb0BpY29uY29sb2dpYS5uZXQ=
†These authors have contributed equally to this work and share first authorship