Abstract
Galectins play relevant roles in tumor development, progression and metastasis. Accordingly, galectins are certainly enticing targets for medical intervention in cancer. To date, however, clinical trials based on galectin inhibitors reported inconclusive results. This review summarizes the galectin inhibitors currently being evaluated and discusses some of the biological challenges that need to be addressed to improve these strategies for the benefit of cancer patients.
Introduction
Galectins are a family of proteins defined by their Carbohydrate Recognition Domain (CRD). Through that domain, galectins bind to galactosides, such as N-acetyllactosamine residues attached to biomolecules (1). Interestingly, the binding of glycans to galectins’ CRD is subject to allosteric regulations (2, 3). Even if carbohydrate binding is the classifying criteria for these proteins, it has long been known that galectins can also interact with other biological molecules in a carbohydrate-independent manner (4) [reviewed in (5, 6)]. Altogether, the list of galectin interactors reported so far has dramatically grown in the last years (extensive bibliography (7–12), cited as examples). Through this panoply of interactions, galectins regulate physiological cell properties such as differentiation; adhesion and migration; cell cycle and survival, immune patrolling, RNA splicing, and gene transcription (5, 6, 13).
Expression of galectins is strongly altered in cancer; comprehensive reviews address this point elsewhere (8, 14, 15). Albeit not oncogenic drivers, galectins exacerbate the malignant phenotype (16–18). Indeed, galectins regulate homotypic and heterotypic aggregation of cancer cells, cancer cell migration and invasion [reviewed in (17)], tumor angiogenesis [reviewed in (19, 20)] and immune escape [reviewed in (7, 8, 15)]. Consequently, increased galectin production in cancers generally predicts a poor clinical outcome for patients (21–24). Among the 16 galectins identified in mammals (12 in humans, as found in GenBank https://www.ncbi.nlm.nih.gov/genbank/ accessed on 20 November 2022), galectins-1, -3, -7, -8, and -9 have been extensively evaluated in cancer patient samples. Pre-clinical experimentation has demonstrated that galectin inhibitors are interesting anti-tumor tools, particularly when combined with irradiation (25–34), chemo- (34–42), anti-angiogenic- (43, 44), and immune-therapies (37, 45, 46). Interestingly, some of the described galectin inhibitors are currently being evaluated at the clinical level. This review aims to summarize galectins’ inhibitory strategies being tested, those that gave encouraging results in pre-clinical studies, and the challenges their effective use may entail.
Current galectin inhibitors
Current galectin inhibitors are listed in Table 1 (in vivo pre-clinical evaluations) and Table 2 (clinical trials). This topic was previously covered by (112–116). However, this manuscript aims to update on the current developments in the field, including some strategies not previously considered. It also assesses the challenges to scaling up the use of galectin inhibitors in the clinic. In this review, compounds are classified according to their mechanism of action (their influence over CRD -competitive vs. allosteric inhibitions-) or their glycan independence (Figure 1).
Table 1
| Inhibitor | Structure | Pre-clinical model | References |
|---|---|---|---|
| a) Carbohydrate compounds | |||
| β-D-lactosyl-steroid | 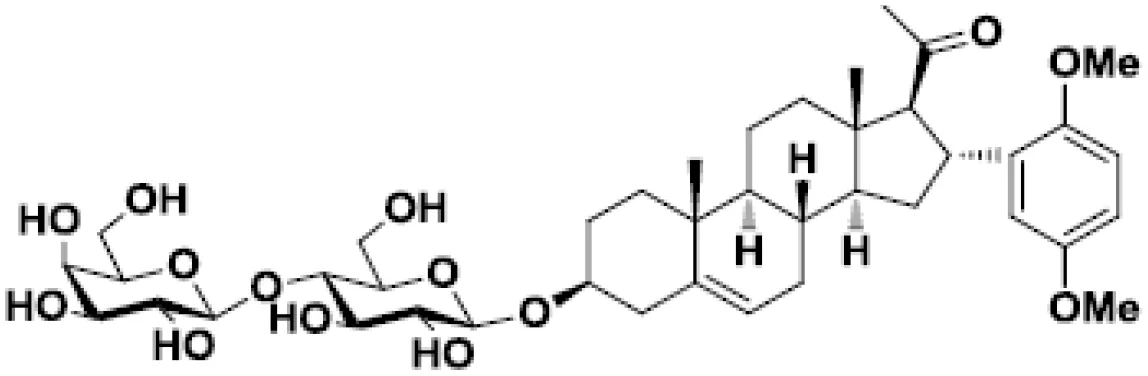 | Lymphoma and glioblastoma | (47) |
| Thiodigalactose (TDG) | 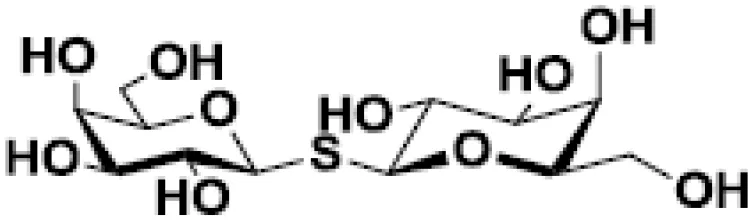 | Pulmonary metastasis in murine breast and colon cancer models | (48, 49) |
| Modified-thiodigalactose (TD139) | 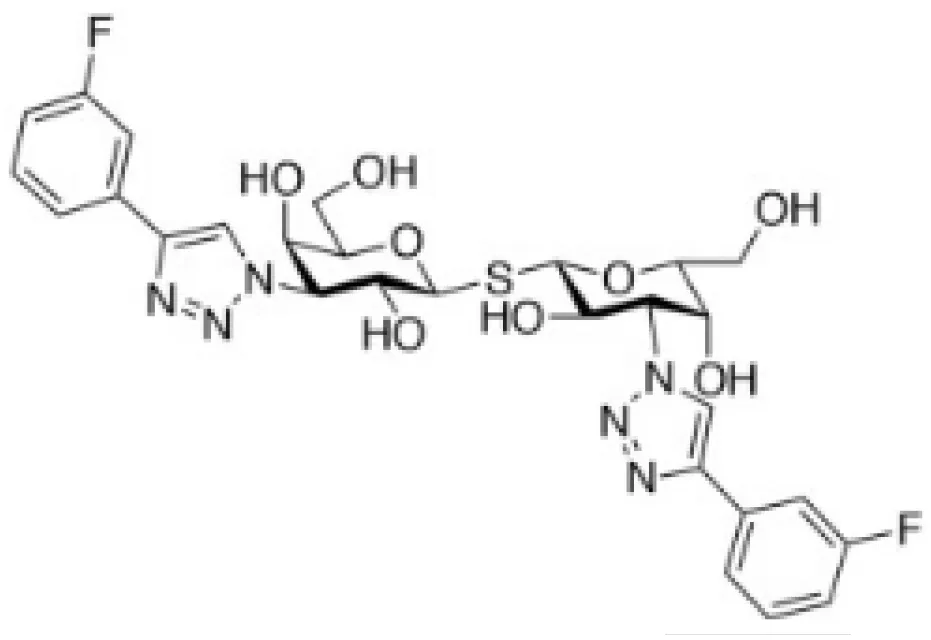 | Lung fibrosis | (50, 51) |
| GB1107 | 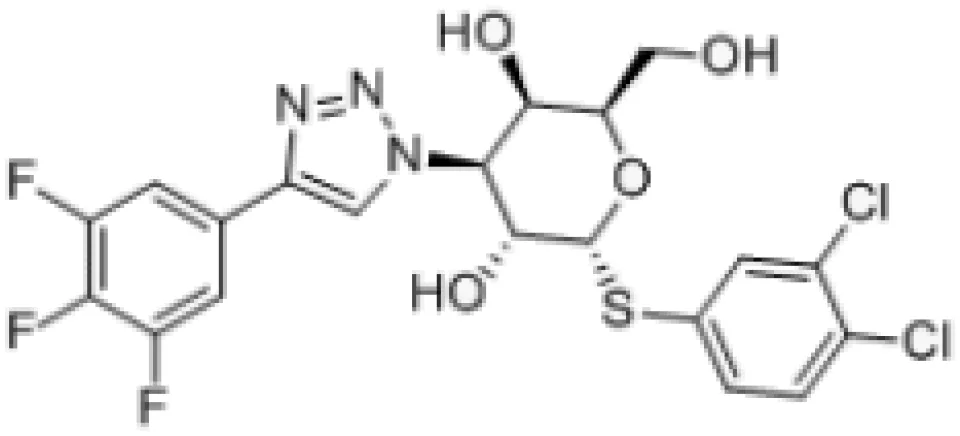 | Human and mouse lung adenocarcinoma in murine models Synergy with negative immune checkpoint. Oral squamous cell carcinoma; synergy with cetuximab | (52, 53) (54) |
| Lactulose-L-leucine | 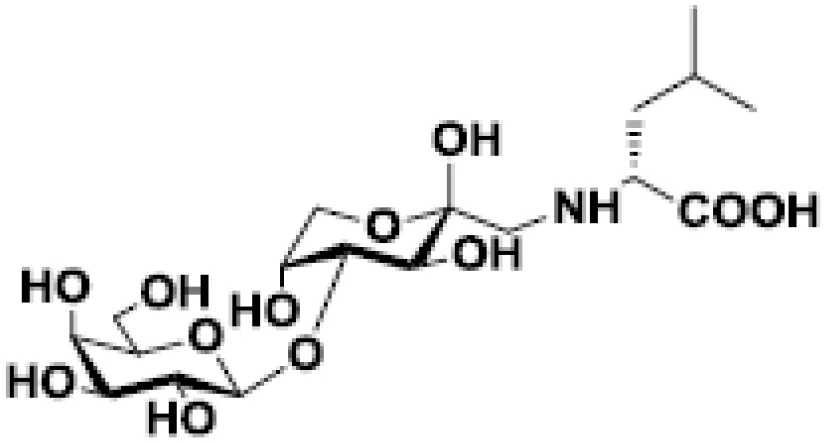 | Breast and prostate cancers in murine models | (55, 56) |
| Dendrimers : galactose- or lactose-conjugated porphyrin derivatives | 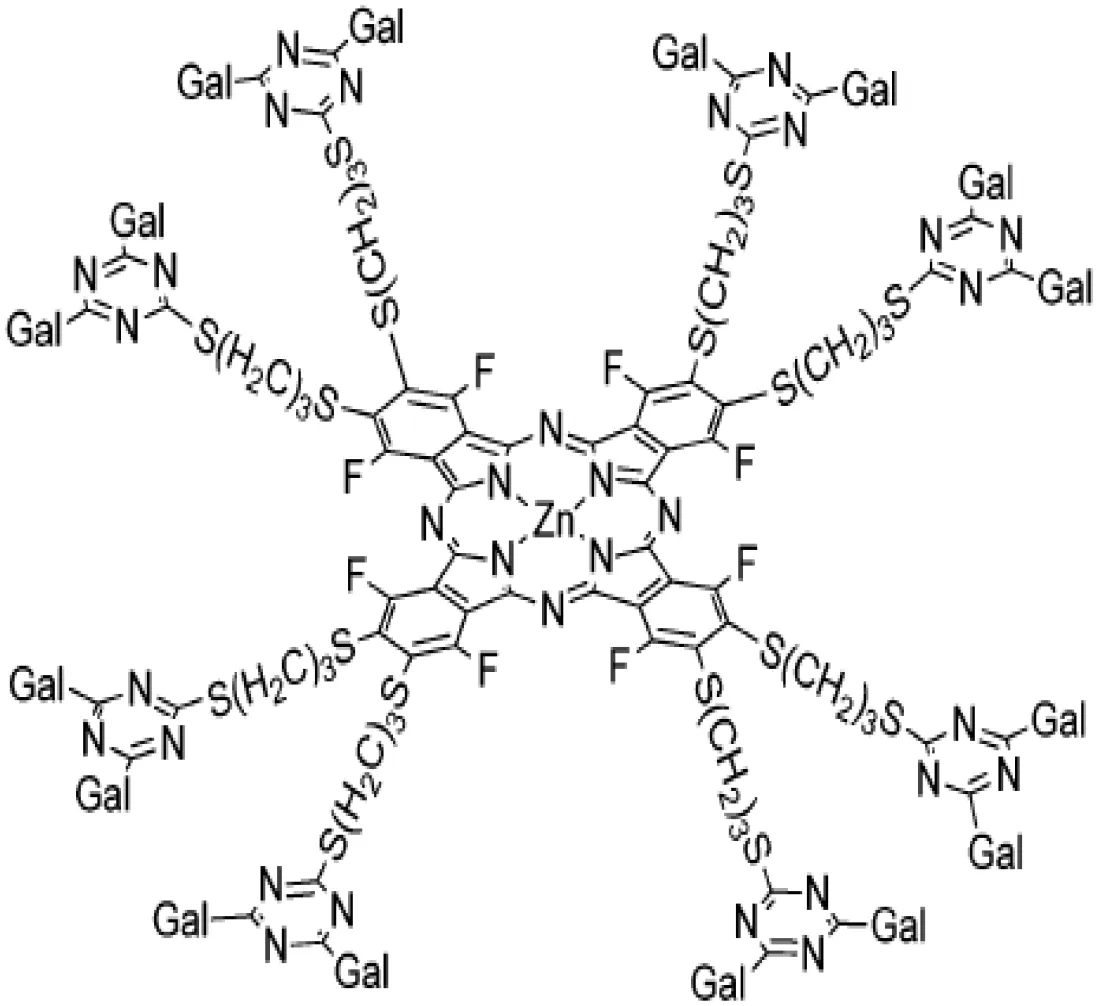 | Photodynamic anti-tumor therapy Bladder cancer model Radiation-induced fibrosarcoma | (57) (58) |
| Modified citrus pectin (MCP) | Heterogenous chemical definition, with the following general structure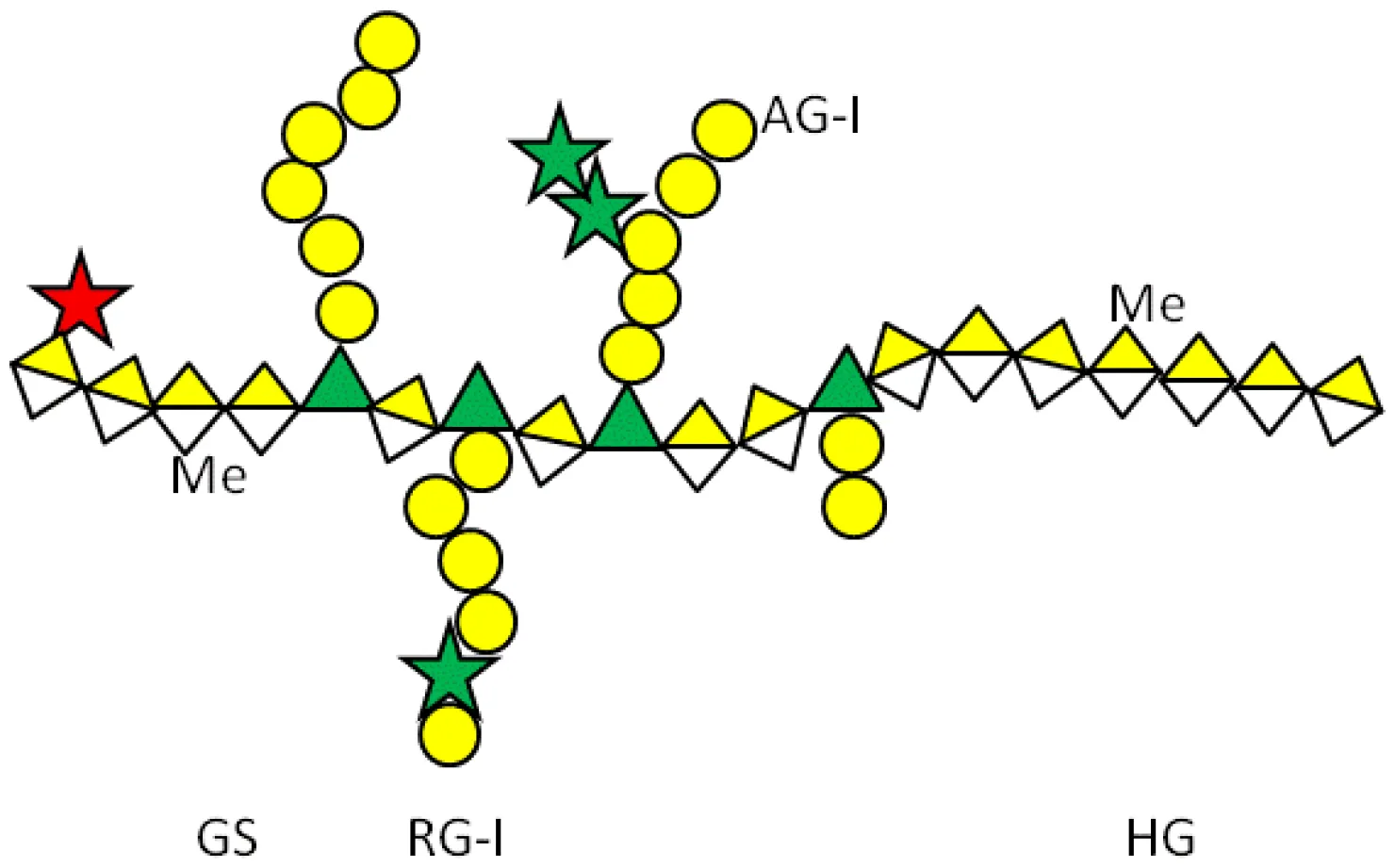 Several methods of preparation: US7491708B1, ES2537936B1, US 2016/0030467 A1 patents | Melanoma Thyroid cancer Breast and colon cancers Prostate cancer | (59) (60) (61) (62) |
| PectaSol-C | Derived from MCP Low molecular weight, 5 % galacturonic acid US 2011/0294755A1 patent, EcoNugenics | Not in vivo pre-clinical studies in animals found (only original MCP) | |
| GCS-100 | derived from MCP US8877263B2 patent, La Jolla Pharmaceutical Company | Mastocytoma | (63) |
| GM-CT-01 or DAVANAT | 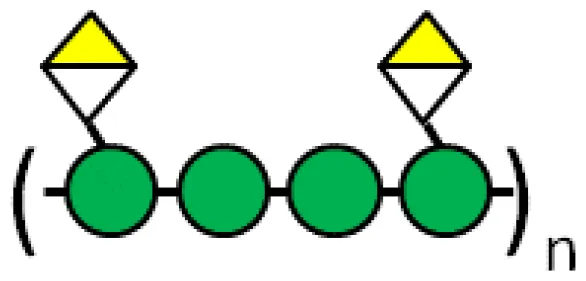 US 2014/0235571 A1 patent, Galectin Therapeutics Inc | Toxicity studies on mice, rats and dogs Colon Cancer | (64) (64) |
| GR-MD-02 (belapectin) | 1,4-linked (methyl) galacturonic acid backbone interspersed with α-1,2 linked rhamnose, the rhamnose carrying 1,4-β-D-galactose residues or 1,5-α-L-arabinose oligomers.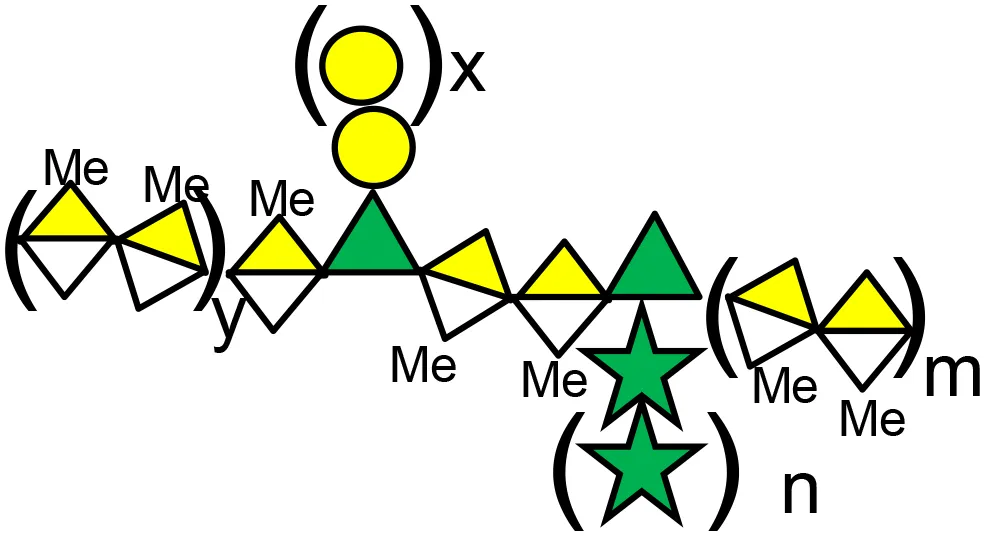 US8871925B2 patent, Galectin Therapeutics Inc. | Sarcoma, breast, and prostate cancer | (65) |
| Carbohydrate-complexed nanoparticles | Citrus pectin-nanoparticles Galactose-Tuftsin peptide-nanoparticles | Colon cancer Melanoma | (66) (67) |
| b) Peptides, peptidomimetics and proteins | |||
| Anginex peptide | ANIKLSVQMKLFKRHLKWKIIVKLNDGRELSLD | In vivo angiogenesis | (68) |
| Teratocarcinoma | (69) | ||
| Melanoma, Ovarian and breast carcinoma | (26, 32, 43, 44) | ||
| Peptidomimetics: 6DBF7 dibenzofuran (DBF)-modified peptide | [DBF]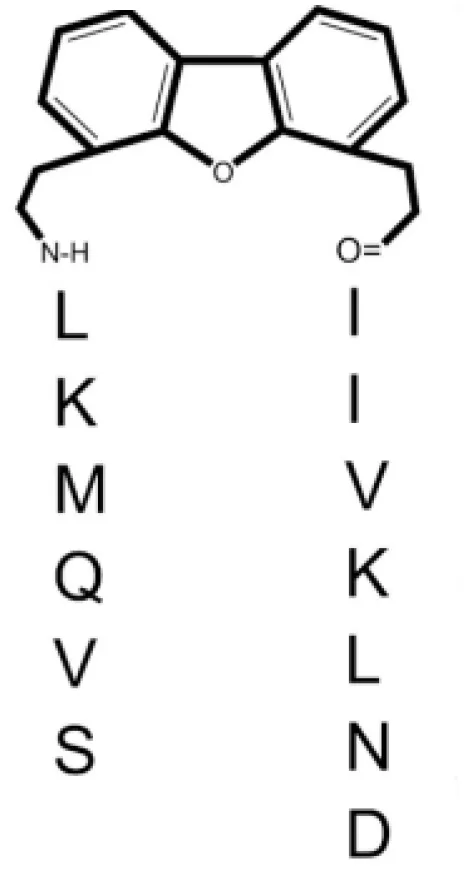 | Melanoma, lung, and ovarian carcinoma | (70, 71) |
| DB16 | SVQMKL-[DBF]-AIVKLNA | Melanoma, lung, and ovarian carcinoma | (71) |
| DB21 | SVQNvaKL-[DBF]-IIVKLNA | Melanoma, lung, and ovarian carcinoma | (71) |
| OTX008 | 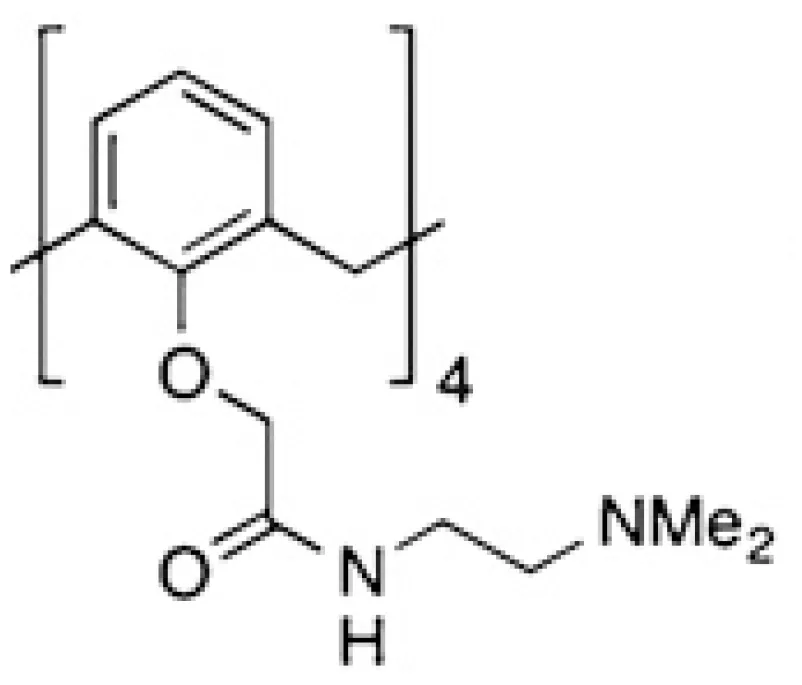 | Melanoma, glioblastoma, thyroid and ovarian carcinoma | (40, 43, 72, 73) |
| PTX013 | 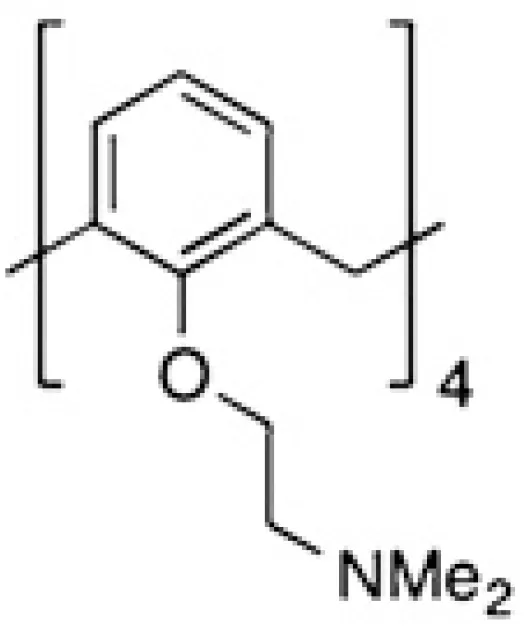 | Melanoma | (74) |
| Dominant negative mutants | Gal-3C (lacks N terminal) | Multiple myeloma | (75) |
| Breast cancer | (76) | ||
| Ameliorates heart failure after myocardial infarction | (77) | ||
| Gal-3 (Ser6-->Glu Ser6-->Ala) mutant unable to phosphorylate | Breast cancer | (78) | |
| Neutralizing antibodies | anti-galectin-1-mAb | Head and neck cancer | (45) |
| Lung carcinoma and melanoma | (79) | ||
| Kaposi' s sarcoma | (80) | ||
| anti-galectin-3-mAb | Breast and ovarian cancers | (81) | |
| anti-galectin-9 mAb | Colon adenocarcinoma | (82) | |
| Breast cancer | (83) | ||
| Pancreatic carcinoma | (84) | ||
| Myeloid Leukemia | (85) | ||
| c) Oligonucleotides | |||
| Aptamers | AP-74 M-545 DNA aptamer (galectin-1 specific) | Lung cancer | (86) |
| siRNA and shRNA-coding vectors (few exemples cited) | galectin-1 shRNA | Hepatocellular carcinoma | (87) |
| Peripheral nerve sheath tumors | (88) | ||
| Gastric cancer | (89) | ||
| Osteosarcoma | (90) | ||
| Lung carcinoma | (91) | ||
| Glioblastoma | (37, 92–95) | ||
| Prostate cancer | (96) | ||
| Melanoma | (97, 98) | ||
| Kaposi's sarcoma | (80) | ||
| galectin-3 shRNA | Hepatocellular carcinoma | (99) | |
| Melanoma | (100) | ||
| Pancreatic cancer | (101) | ||
| Prostate cancer | (102) | ||
| galectin-8 shRNA | Prostate cancer | (103) | |
| galectin-4 shRNA | Colorectal cancer | (104) | |
| Regulation of mi-RNA | miR-424-3p (galectin-3) using resveratrol | Ovarian and colorectal cancers | (105) |
| d) Compounds from chemical synthesis | |||
| Benzimidazole compounds | LLS30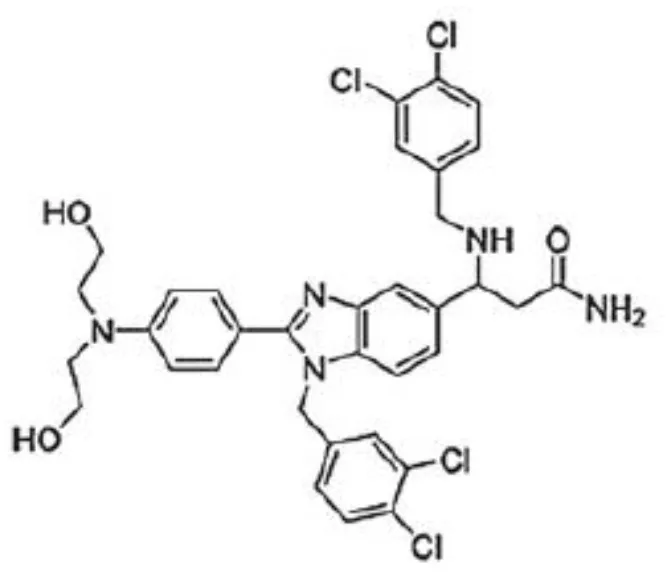 | Ovarian cancer Prostate cancer | (106) (107) |
LLS2 | Peripheral nerve sheath tumors | (88) | |
In vivo pre-clinical studies with galectin inhibitors.
Glycans symbols (according to https://www.ncbi.nlm.nih.gov/glycans/snfg.html).
 D-galacturonic acid,
D-galacturonic acid,  D-galactose,
D-galactose,  L-rhamnose,
L-rhamnose,  L-arabinose,
L-arabinose,  D-xylose, Me: methyl ester
D-xylose, Me: methyl ester
Table 2
| Clinical trial # | Phase | Inhibitor | Combinatory treatment | Targeted-galectin (reported) | Disease | Last Update | Status (mention if the results are available) |
|---|---|---|---|---|---|---|---|
| Healthy subjects | |||||||
| NCT03809052 | I | GB1211 | None | gal-3 | Healthy subjects | March 17, 2021 | Completed, with results |
| Cancers | |||||||
| NCT05240131 | I/II | GB1211 | Atezolizumab | gal-3 | Non-small cell lung cancer | October 3, 2022 | Recruiting |
| NCT01681823 | II | PectaSol-C | None | gal-1/-3 | Biochemical relapsed prostate cancer | January 29, 2020 | Completed (108, 109), |
| NCT00514696 | II | GCS-100 | None | gal-3 | Chronic lymphocytic leukemia | June 17, 2013 | Completed, unreported results |
| NCT00776802 | I/II | GCS-100 | Etoposide/Dexamethasone | gal-3 | Relapsed/refractory diffuse large B-cell lymphoma | June 25, 2013 | Withdrawn (Lack of funding), unreported results |
| NCT00609817 | I | GCS-100 | Bortezomib/Dexamethasone | gal-3 | Relapsed/refractory multiple myelome | June 25, 2013 | Terminated (Lack of funding), unreported results |
| NCT00054977 | I | GM-CT-01 | 5-Fluorouracil | gal-1/-3 | Advanced solid cancers: colorectal, lung, head and neck, and prostate cancers | March 12, 2012 | Completed, unreported results |
| NCT00388700 | II | GM-CT-01 | 5-Fluorouracil, Leucovorin, bevacizumab | gal-1/-3 | Colorectal cancer | February 14, 2018 | Withdrawn (Financing and re-organization), unreported results |
| NCT00110721 | II | GM-CT-01 | 5-Fluorouracil | gal-1/-3 | Colorectal cancer | March 6, 2012 | Terminated (study protocol amended to a new treatment regimen: study DAVFU-006.), unreported results |
| NCT00386516 | II | GM-CT-01 | 5-Fluorouracil | gal-1/-3 | Advanced gall bladder and bile duct cancer | August 1, 2017 | Withdrawn (Financing and re-organization), unreported results |
| NCT01723813 | I/II | GM-CT-01 | Peptide vaccination | gal-1/-3 | Metastatic melanoma | March 12, 2019 | Terminated due to end of validity of the peptide vaccine; no reported results, unreported results |
| NCT02117362 | I | GR-MD-02 | Ipilimumab | gal-1/-3 | Metastatic melanoma | March 21, 2019 | Completed, unreported results |
| NCT00054977 | I | GR-MD-02 | 5-fluorouracil | gal-1/-3 | Advanced solid tumors: colorectal, lung, breast, head and neck, prostate | March 12, 2012 | Completed, unreported results |
| NCT02575404 | I | GR-MD-02 | Pembrolizumab | gal-1/-3 | Advanced melanoma, non-small cell lung cancer, and head and neck squamous cell cancer | July 15, 2022 | Active, not recruiting (110) |
| NCT04987996 | II | GR-MD-02 | Pembrolizumab | gal-1/-3 | Metastatic melanoma, head and neck squamous cell carcinoma | September 10, 2022 | Suspended (Study delayed due to ongoing discussions with the owner of one of the investigational agents), unreported results |
| NCT02117362 | I | GR-MD-02 | Ipilimumab | gal-1/-3 | Metastatic melanoma | March 21, 2019 | Completed, unreported results |
| NCT01724320 | I | OTX008 | None | gal-1 | Advanced solid tumors | November 9, 2012 | Unknown, unreported results |
| NCT04666688 | I/II | Lyt-200 | Chemotherapy, Anti-PD-1 | gal-9 | Relapsed/refractory metastatic solid tumors | March 11, 2022 | Recruiting |
| Non-cancer diseases | |||||||
| NCT02257177 | I/II | TD139 | gal-3/others | Idiopathic pulmonary fibrosis | April 8, 2021 | Completed, with results | |
| NCT03832946 | II | TD139 | gal-3/others | Idiopathic pulmonary fibrosis | May 24, 2022 | Active, not recruiting | |
| NCT04473053 | I/II | TD139 | gal-3/others | COVID-19 | September 16, 2021 | Active, not recruiting | |
| NCT04607655 | I/II | GB1211 | gal-3 | Non-alcoholic steatohepatitis (NASH) and liver fibrosis | February 4, 2021 | Withdrawn (Due to COVID-19 pandemic and change in the clinical development strategy for the GB1211 compound), unreported results | |
| NCT05009680 | I/II | GB1211 | gal-3 | Hepatic impairment | August 3, 2022 | Active, not recruiting | |
| NCT01960946 | I/II | MCP/PectaSol C | gal-1/-3 | Hypertension | February 21, 2021 | Completed, results in (111) | |
| NCT01717248 | I | GCS-100 | gal-3 | Chronic kidney disease | June 20, 2013 | Completed, unreported results | |
| NCT01843790 | II | GCS-100 | gal-3 | Chronic kidney disease | September 1, 2015 | Completed, unreported results | |
| NCT02312050 | II | GCS-100 | gal-3 | Chronic kidney disease | May 19, 2015 | Unknown, unreported results | |
| NCT02155673 | II | GCS-100 | gal-3 | Chronic kidney disease | December 26, 2016 | Completed, unreported results | |
| NCT02333955 | II | GCS-100 | gal-3 | Chronic kidney disease | January 15, 2015 | Withdrawn (Corporate decision), unreported results | |
| NCT01899859 | I | GR-MD-02 | gal-1/-3 | Non-alcoholic steatohepatitis, portal hypertension, and advanced liver fibrosis | February 23, 2015 | Completed, unreported results | |
| NCT02462967 | II | GR-MD-02 | gal-1/-3 | Portal hypertension, and advanced liver fibrosis | October 8, 2020 | Completed, with results | |
| NCT02421094 | II | GR-MD-02 | gal-1/-3 | Liver fibrosis | October 8, 2020 | Completed, with results | |
| NCT02407041 | II | GR-MD-02 | gal-1/-3 | Psoriasis | September 7, 2020 | Completed, with results | |
| NCT04332432 | I | GR-MD-02 | gal-1/-3 | Subjects with normal hepatic function and subjects with hepatic impairment | March 28, 2022 | Completed, unreported results | |
| NCT04365868 | IIb/III | GR-MD-02 | gal-1/-3 | Esophageal varices in NASH cirrhosis | September 22, 2022 | Recruiting | |
Clinical trials with galectin inhibitors.
Data from www.clinicaltrials.gov. [Accessed November 24, 2022].
Figure 1

Galectin inhibitory strategies.
Galectin inhibitors affecting carbohydrate recognition
Competitive inhibitors of carbohydrate-binding to galectins
The lectin functions of this family of proteins are the most widely studied. Indeed, galectins bind to β-galactosides through their CRD. For instance, considering its canonical ligand lactose, the C4’ and C6’ hydroxyls of the galactose and C2 and C3 of glucose are primarily responsible for the hydrogen-bond interactions with conserved residues of CRD in galectin-3 (117) and galectin-1 (118). Basis of the molecular glycan-protein interactions has also been described for other galectins (119, 120). The fine specificity of galectins for different oligosaccharides stems from residues surrounding this main binding site. Consequently, each galectin has a different glycan-binding preference contributing to its specific biological activities (121). The first described galectin inhibitors are molecules capable of binding to the CRD and preventing further ligand binding. Galectin inhibitors based on these competitive interactions consist of chemically modified mono or disaccharides structured around galactose (58, 122–125), lactose (58, 125–127), thiodigalactose (TDG) (34, 128–132), talose (133, 134) and lactulose (135). One of the first tempts to use this type of inhibitor in cancer consisted of administering a β-D-lactosyl-steroid. This treatment significantly increased the survival of mice grafted with lymphoma and glioblastoma cells (47, 136). Moreover, this compound increases the anti-tumor cytotoxic effects of cisplatin in mice (47).
Several chemical modifications of glycans have been developed to improve these molecules’ inhibitory properties. For example, introducing a sulfur atom into the glycoside linkage in TDG makes the molecule more resistant to glycosidases (137). The in vivo anti-tumor properties of some of these compounds were challenged in pre-clinical studies. For instance, TDG administration reduces pulmonary metastasis in murine breast and colon cancer models (48). TDG promotes immune infiltration, reduces angiogenesis, and protects cells against oxidative stress (49). The most advanced TDG in clinical studies is TD139 (also named as GB0139), developed by Galecto Biotech (Copenhagen, Denmark). TD139 recognizes galectin-3 CDR with high affinity (Kd 68 nM) (138). However, its absolute selectivity for galectin-3 is relative since it also binds to galectin-1 CDR (Kd 220 nM) and other galectins with lower affinities (138). This compound was initially evaluated in pre-clinical models of lung fibrosis (50, 51). Interestingly, TD139 was also evaluated in a clinical trial as a potential therapeutic for idiopathic pulmonary fibrosis (NCT02257177; www.clinicaltrials.gov [accessed November 24, 2022]; Table 2) (139).
More recently, a series of monosaccharide galectin-3 inhibitors with high affinities and good selectivity over other galectins have been described (140). From this series, GB1107 (3,4-dichlorophenyl 3-deoxy-3-[4(3,4,5-trifluorophenyl)-1H-1,2,3- triazol-1-yl]-1-thio-α-D-galactopyranoside) from Galecto Biotech; has good affinity (Kd 37 nM) and bind to the CRD of galectin-3. Both, TD139 and GB1107 are membrane-permeable small molecules (141). GB1107 is characterized by good biodisponibility upon oral administration and low clearance (52). It was demonstrated that the oral administration of GB1107 reduced human and mouse lung adenocarcinoma growth and blocked metastasis in murine models (52). Mechanistically, treatment with GB1107 promotes tumor M1 macrophage polarization and CD8(+) T-cell infiltration (52). Moreover, GB1107 potentiated the effects of a PD-L1 immune checkpoint inhibitor to increase expression of cytotoxic (IFNgamma, granzyme B, perforin-1, Fas ligand) and apoptotic (cleaved caspase-3) effector molecules (52, 53). In addition, GB1107 and cetuximab displayed a synergistic inhibitory effect on the growth of oral squamous cell carcinoma (54). Phase I studies with GB1211 (which shares a chemical template with GB1107) have been completed (NCT03809052, Table 2), and Galecto Biotech initiated safety and efficacy clinical studies with GB1211 combined with atezolizumab in the treatment of non-small-cell lung cancer (NCT05240131, Table 2).
Finally, it should be mentioned that chemical modifications of galactosides and their evaluation as galectin inhibitors in cancer are an intense field of research. First, synthetic glycoamines evidenced anti-tumor activity (55, 56, 142, 143). Indeed, lactulose-L-leucine mimics cancer-associated Thomsen-Friedenreich glycoantigen and binds to galectin-3. At a molecular level, it was demonstrated that this compound binds to the CRD of galectins-1 and -3 with higher affinity than lactose and TDG (135). In a murine breast cancer model, the administration of lactulose-L-leucine (and fructosyl-D-leucine) inhibited spontaneous metastasis in nude mice (56). The same group demonstrated the beneficial effects of lactulose-L-leucine in controlling and preventing prostate cancer metastasis to the bone (55). Other inhibitory molecules arising from chemical modifications of galactosides can also be cited (122, 144–146); however, they do not reach the level of in vivo evaluation.
To improve galectin inhibitors’ properties, inspiration was found in the clustering nature of galectin glycan interactions. Indeed, the synthesis of multivalent glyco-clusters with improved galectin inhibitory potential has been reported (147–152). Interestingly, cell aggregation can either be inhibited or enhanced depending on the number of lactose groups in functionalized dendrimers (153). Unfortunately, no evaluation of their in vivo biological effects in pre-clinical models was yet reported. Another strategy based on the same conceptual framework tested dendrimers obtained by galactose conjugation to the porphyrin derivatives (154). In this case, a photodynamic anti-tumor therapy was successfully reported in a pre-clinical in vivo bladder cancer model (57).
Pectins are another group of galectin-binding, inhibitory compounds. Natural pectins are large and heterogeneous polysaccharides found in plants which constitutes fiber components of our diet. Pectins have molecular weights ranging from 60 to 130 kDa and are constituted by three main polysaccharides: homogalacturonan (HG), rhamnogalacturonan-I (RG-I), and substituted galacturonans (GS) (155, 156). Pectins must be modified by pH and heat to gain solubility and biological effects. Indeed, hydrolysis induces galactoside exposure, and now, modified pectins bind galectins (157, 158). Contrary to what was often supposed, some experimental data prompt the existence of non-conventional sites of pectin binding in galectins (159–162). On the contrary, other in vitro data support that pectin-mediated biological effects are (or partially are) mediated by glycans (163–166). Adding complexity to the field, modified pectins are generally administered orally. Nevertheless, pectins are not digestible in the human intestinal tract, and their modifications are believed to increase their absorbability (167, 168). Moreover, it has been postulated that products of pectin fermentation by the human microbiota should contribute to their systemic in vivo biological effects (169). It should also be mentioned that pectins induce galectin-independent biological effects (170, 171). Altogether, these arguments indicate that more basic research is needed to clarify the fine mechanisms through which pectins induce their biological effects.
In this context, one of the most studied galectin inhibitors is the modified citrus pectin (MCP), which is obtained by partial hydrolysis of citrus pectin. In vitro studies demonstrated that MCP binds galectin-3 through galactoside residues (59, 62). Functionally, MCP inhibits galectin-3 binding to endothelial cells, and more importantly, the adhesion of breast tumors to endothelial cells (61). In addition, MCP treatment induces important metabolic changes in tumor-associated macrophages, which impacts on tumor growth and metastasis (172, 173). Interestingly, these MCP biological effects are carbohydrate dependent (59). In vivo administration of MCP inhibits melanoma (59), thyroid (60), breast and colon tumor growth, angiogenesis and metastasis (61, 173), and spontaneous metastasis in a rat prostate cancer model (62). Due to the high chemical variability of dietary MCP supplements on the market, more defined MCP variants have been described: PectaSol-C, GCS-100, GM-CT-01 and GR-MD-02. PectaSol-C has a molecular weight ranging from 5-10 kDa with 5% of monogalacturonic acid content (174). In vitro studies demonstrated the potential interest of PectaSol-C MCP in prostate (174, 175), breast (175) and ovarian cancers (176, 177), particularly if used combined with other therapies (175, 177). Interestingly, phase II pilot studies demonstrated the tolerability and encouraging biological results obtained by the use of this inhibitor in prostate patients (108, 109) (NCT01681823, Table 2).
GCS-100 is a complex polysaccharide prepared from modified citrus pectin. Mechanistically, GCS-100 detaches galectin-3 from CD4+ and CD8+ tumor-infiltrating lymphocytes, boosts cytotoxicity and restores IFN-gamma secretion (63). Similar effects were obtained by using N-acetyllactosamine, suggesting GCS-100 effects are carbohydrate-dependent (63). Interestingly, GCS-100 induces tumor rejection only when associated with vaccination in pre-clinical model of mastocytoma secretion (63), implying GCS-100 modulates the tumor immune attack. Altogether, these promising results prompt La Jolla Pharmaceuticals to launch GCS-100-based clinical trials. Following a phase I dose escalation safety study in patients with refractory solid tumors (178), a phase II study was completed in patients with chronic lymphocytic leukemia (179) (NCT00514696, Table 2). In these exploratory trials, GCS-100 was well tolerated, and 25% of patients showed a partial response (179). In addition, the use of GCS-100 has also been evaluated in chronic kidney disease (Phase I NCT01717248 and phase IIa NCT01843790, Table 2). In 2015, La Jolla Pharmaceuticals announced that they were discontinuing the development of GCS-100 after the Food and Drug Administration (FDA) required a more complex characterization of the compound to advance into late-stage development (NCT00776802 and NCT00609817, Table 2).
Another pectin-derived polysaccharide able to inhibit galectins is GM-CT-01 or DAVANAT®. This polysaccharide is extracted from guar seeds and subjected to controlled partial chemical degradation (developed by Galectin Therapeutics, formerly Pro-Pharmaceuticals). A backbone of the galactomannan is composed of (1→4)-linked β-D-mannopyranosyl units, to which single α-D-galactopyranosyl is attached by (1→6)-linkage (64). The average repeating unit of GM-CT-01 consists of seventeen β-D-Man residues and ten α-D-Gal residues (Man/Gal ratio is 1.7), and an average polymeric molecule contains approximately 12 of such repeating units (for the average molecular weight of 51,000 Da). In vitro, GM-CT-01 boosts the cytotoxic properties of CD8(+) tumor-infiltrating lymphocytes and their ability to produce IFN-gamma (180). Indeed, this pectin prevents glycosylated cytokines (IFNγ between others) be captured by galectin-3 and therefore allowing the chemokine gradient needed to attract lymphocytes towards the tumor (181). Pre-clinical studies in mice defined GM-CT-01 non-toxic doses (alone or combined with other chemotherapies) (64). Moreover, such studies demonstrated GM-CT-01 beneficial effects in colon cancer models (64). Interestingly, a phase I clinical trial was completed in cancer patients with advanced solid tumors by administration of DAVANAT® combined with 5-fluorouracil treatment (NCT00054977, Table 2). Combinatory treatment was well-tolerated. While phase II trials were announced, these trials were never initiated, having a “withdrawn/terminated status” in www.clinicaltrials.gov (NCT00388700, NCT00110721, NCT00386516, Table 2). In addition, melanoma peptide vaccination plus GM-CT-01 was evaluated in melanoma (NCT01723813). This clinical trial was “terminated due to end of validity of peptide vaccine” with no reported results.
Finally, GR-MD-02 (belapectin) is a galactoarabino-rhamnogalacturonan-rich polysaccharide obtained through chemical processing from apple pectin (developed by Galectin Therapeutics, Norcross, Georgia, USA). GR-MD-02 is a galectin-3 inhibitor which synergizes with anti-OX40 treatment to promote tumor regression and increases survival of tumor-bearing mice (65). This occurs through a CD8(+) T cell-dependent mechanism, reducing the immunosuppresion mediated by myeloid-derived suppressor and regulatory Foxp3(+)CD4(+)T cells (65). GR-MD-02 administration induced a significant reduction of liver fibrosis in experimental models of non-alcoholic steatohepatitis (182, 183). GR-MD-02 is being evaluated in melanoma, squamous head and neck, and non-small cell lung cancer patients combined with the negative immune checkpoint inhibitors pembrolizumab (anti-PD-1, NCT02575404, and NCT04987996, this last suspended) and ipilimumab (anti-CTLA-4, NCT02117362; Table 2). No results are available yet from those clinical studies. Interestingly, this compound has also been evaluated in clinical trials for non-alcoholic steatohepatitis, portal hypertension, and advanced liver fibrosis (NCT01899859 and NCT02462967, Table 2). In this case, GR-MD-02 was safe but not associated with significantly ameliorating hepatic disease (184).
An area under intense investigation tries to achieve formulations with improved pharmacokinetic properties for this type of carbohydrate-based inhibitors. This is the case of lactose-, galactose- or pectins-complexed nanoparticles (185–187). Apart from improving the pharmacokinetic properties of the inhibitor, these nanoparticles can also serve as delivery carriers of cytotoxic drugs toward the tumor (66, 188, 189). Moreover, attempts are being made with nanoparticle modifications to improve selective targeting of the tumor (or tumor-associated stroma) (67, 190).
Interestingly, non-carbohydrate inhibitors for galectins have also been proposed. First, the anti-tumor properties of several synthetic heterocyclic compounds able to bind galectin-1 have been evaluated. Molecular docking experiments described fine interactions between these molecules and the CRD domain of galectin-1 (191–194). Moreover, in vitro results indicate these compounds have anti-tumor cytotoxic properties (191–194). However, in vivo anti-tumor pre-clinical evaluations of such compounds remain to be performed. Second, bacteriophage display library systems for interaction screening allowed the discovery of galectin-binding peptides. For instance, a Thomsen-Friedenreich antigen-specific peptide (P-30) able to bind galectin-3 has been described (195). This peptide modulates breast and prostate tumor homotypic aggregation and tumor cell adhesion to the endothelium (195). Using similar technological approaches, stapled-peptides ligands binding galectin-3 were described (196). These peptides bind to the CRD of galectin-3 and the best one has an intermediate affinity (Kd 0.45 μM) (196). However, no functional studies have been reported for these peptides. As already mentioned, formulations with improved pharmacokinetics are being evaluated. In this context, nanoparticles combining carbohydrates (inhibitor) and peptides (addressers) have been described, a strategy that significantly improves their biodistribution and the biological effects (67).
Finally, genetic engineering methods are used to inhibit the glycan-dependent functions of galectins. For instance, a dominant negative mutant formed by the last 143 carboxyl-terminal amino acid residues and lacking the N-terminal domain of galectin-3 (named Gal-3C) has been described. This Gal-3C molecule preserves the CRD but lacks cooperative binding and crosslinking properties of the wild-type galectin-3 (197). Indeed, it is hypothesized that the administration of an excess of soluble Gal-3C competes with endogenous galectin-3 for carbohydrate binding sites (76). In this context, Gal-3C reduces angiogenesis by abrogating extracellular galectin-3 interaction with αvβ3 integrin through its carbohydrate recognition domain (198). Interestingly, Gal-3C inhibits CXCL12-induced leukocyte migration in (non-cancer) inflammatory conditions (199). Gal-3C also inhibits tumor cell motility and invasion (75, 200). Hence, Gal-3C alone or combined with other chemotherapies can reduce ovarian, breast cancer, and multiple myeloma growth and drug resistance (75, 76, 200). Interestingly, Gal-3C can be used in vivo without toxic effects (76); this treatment ameliorates heart failure after myocardial infarction (77). Galectin-9 mutants have also been described. Indeed, mutations in galectin-9 CRD abolish its binding to the negative checkpoint Tim-3; this interaction occurs via the carbohydrates (201). Dominant negative mutants can also interfere with nuclear partners in a glycan-dependent manner. This is the case of the interactions between galectin-1 and Foxp3. This transcription factor functions as a master controller of regulatory T cells (Treg). Moreover, the interaction between galectin-1 and Foxp3 controls a panoply of genes and functions in breast cancer cells (202). Consequently, galectin-1 mutants that lack the N-terminus and do not bind Foxp3 can be used to inhibit breast tumor proliferative and invasive properties (202). These results show that negative dominants could be interesting tools to inhibit galectins.
Non-competitive allosteric inhibitors of carbohydrate-binding to galectins
Some inhibitors do not directly interact with the CRD of galectins, but their inhibitory effects are still glycan-dependent. Indeed, these molecules function as allosteric inhibitors, interacting outside the CRD but inducing changes in this region, thereby inhibiting glycan binding and biological effects. For instance, in vivo galectin-1 inhibition through the administration of lactose-conjugated purpurinimide photosensitizers reduced the growth of radiation-induced fibrosarcoma (58). Molecular modeling analysis indicated that this compound does not interfere with the CRD (203). Similar photodynamic strategies with galactose-bound porphyrin demonstrated anti-tumor effects in bladder cancers (57). In this case, galectin-1 inhibition generates oxidative stress and apoptosis of tumor cells over-expressing this lectin (57).
However, allosteric inhibition can also be performed using non-carbohydrate molecules. Based on the significant role of galectins in the interaction between tumor and endothelial cells during tumorigenesis, a cytokine-like peptide named anginex was described as a potent anti-angiogenic tool (68). This biological effect is mediated through galectin-1 binding (69), although this peptide also binds other galectins (204). The anti-tumor effects of anginex were demonstrated in several experimental cancer models (26, 32, 43, 44). Anginex’s angiostatic beta-sheet-forming structure inspired the design of the 6DBF7, a peptidomimetic that also interacts with galectin-1 (70, 71). This 6DBF7 molecule inhibits glycan binding of galectin-1 in a noncompetitive, allosteric manner (71). Based on these studies, other potent analogs (DB16 and DB21) have also been described (71). These peptides inhibit angiogenesis and tumor growth significantly better than 6DBF7 or anginex (71). To overcome the susceptibility of these peptides to hydrolysis by proteases, Dings et al. designed a non-peptidic topomimetic of anginex and 6DBF7 based on a calixarene scaffold. Indeed, calix[4]arene compound 0118/OTX008/PTX008 binds to galectin-1 at a site away from the lectin’s carbohydrate binding site, thereby attenuating lactose binding to the lectin (205). It should be mentioned that the specificity of this compound is relative since it also binds to galectin-3, albeit more weakly (206). Pharmacokinetics and anti-tumor activity of OTX008 alone or combined with other treatments were evaluated in melanoma, glioblastoma, thyroid and ovarian carcinoma (40, 43, 72, 73, 207). A phase I study of OXT008 in patients with advanced solid tumors was reported (NCT01724320, Table 2). Unfortunately, this study is listed with an “unknown recruitment status”; no updates have been posted since 2012. Chemical modifications of PTX008 were also described; it is interesting to mention the PTX013 compound. This compound is more potent as a cytotoxic tumor agent than the parenteral PTX008. This higher inhibitory potency of PTX013 was demonstrated both in vitro (head and neck, breast, ovarian, renal, lung, and prostate cancer lines, several of them radiation resistant), and importantly in vivo (melanoma) (74).
Galectin inhibition can also be achieved using specific neutralizing monoclonal antibodies (mAb). It must be noted that, for this strategy, mechanisms of galectin inhibition (competition or allosteric inhibition) depends on each antibody. In the case of galectin-1, one of these antibodies (Gal-1-mAb3) has been characterized, and the epitope recognized by this mAb localizes outside the CRD although it is still capable of inhibiting N-acetyllactosamine-galectin-1 interaction (208). This antibody recognizes specifically galectin-1 with high affinity (EC50 = 523nM). This neutralizing antibody reproduces the anti-angiogenic and immunopotentiating activities observed with other types of inhibitors (208, 209). In particular, blockade of galectin-1 (Clone 25C1; Novo Castra) significantly reduces the in vitro inhibitory effects of human and mouse CD4+CD25+ Treg cells (210). Moreover, another anti-galectin-1 neutralizing mAb ameliorates the negative immune checkpoint (PD1) response in irradiated mice carrying oral cancer cells (45).
In the case of galectin-3, earlier studies described mAbs recognizing non-CRD domains but causing a profound modulation of its lectin activities (211). On the other hand, a galectin-3-specific mAb (14D11) competes with lactose for the carbohydrate-binding pocket of galectin-3 (81). This antibody inhibits invasion of Mucin-16-expressing cancer cells, prolonging overall survival in animal tumor models (81). However, inhibition of galectin-3 also impacts the tumor stroma cells. Indeed, the use of an anti-galectin-3 mAb (B2C10) promotes IFN-γ secretion by in vitro stimulated CD8+ tumor-infiltrating T lymphocytes (63).
The scientific interest in developing anti-galectin-9 mAb is major since this protein participates in various mechanisms of immune escape by tumors: control of T cell survival (212), T cell effector exhaustion and differentiation (82, 201, 213, 214), lymphocyte migration towards the tumor via an endothelial cell reprogramming (45), Treg function (215–220), regulation of antigen presentation (221–223), and myeloid suppressive cells (224). Confirmation of these functions by the use of blocking antibodies is becoming very frequent. Such is the case of two antibodies (clones 292-13 and 292-18A) reacting with high affinity with the N-CRD of human galectin-9; their use protects T cells from galectin-9 mediated cell death and promotes tumor-cell killing by T cells (225). The same group, but using a commercial anti-galectin-9 mAb (RG9-1 from InVivoMAb), demonstrated prevention of CD8+T cell exhaustion and near complete Treg depletion when this mAb is combined with anti-GITR (glucocorticoid-induced tumor necrosis factor receptor-related protein)-specific antibody (82). Two other anti-galectin-9 mAb have also been reported (Gal-Nab1 and Gal-Nab2). In this case, antibodies recognize an epitope comprising 213-224 amino-acid sequence with high affinity (in the order of nM) (226). Again, these antibodies protect T cells from galectin-9-mediated cell death (226). An anti-galectin-9 was combined with anti-Tim-3 mAb to improve taxane-based chemotherapy in breast cancer (83). Apart from the direct effects on adaptive immunity, blockade of galectin-9 by antibodies potentiates immune attack in pancreatic carcinoma through modulation of macrophage function (84). Nevertheless, galectin-9 blockade by antibodies also acts directly on tumor cells. Indeed, leukemia stem cells secrete galectin-9, which through the interaction with Tim-3 constitutes an autocrine loop critical for leukemic self-renewal and development (85). Indeed, galectin-9 neutralization is a potent way to prevent the reconstitution and the self-renewal of human acute myeloid leukemia cells in a xenogeneic transplantation model (85). Finally, an anti-galectin-9 mAb (Lyt-200) is currently under clinical investigation in phase I/II trial for its safety and efficacy in patients with relapsed/refractory metastatic solid tumors (NCT04666688, Table 2). In this clinical trial, Lyt-200 is evaluated alone and in combination with chemotherapy or anti-PD-1.
The use of neutralizing antibodies to block other galectin members in cancer is more incipient, and in most cases, polyclonal antibodies are evaluated. For instance, neutralizing surface-bound galectin-4 in human colorectal cancer induces significant transcriptional changes and chemokines production in tumor cells (227).
While neutralizing antibodies carry several benefits over small inhibitory carbohydrate molecules, they also have several drawbacks. Some of the concerns are related to their selectivities and biodistributions. Antibodies inhibit extra-cellular galectins, and lack restricted biodistribution in the body. These characteristics imply that antibody-mediated inhibition of galectins could act as partial inhibitors (lack of intracellular effects), and do not discriminate between non-transformed and transformed cells resulting in adverse effects. More studies are needed to fully understand the effects induced by galectin-neutralizing antibodies and their potential transfer to the clinic.
Finally, nucleotide-based molecules are a different family of galectin inhibitors. In this sense, a single-stranded DNA aptamer (AP-74 M-545) has been described as an antagonist of galectin-1 (86). This aptamer shows higher affinity (KD = 3.7 nM) and specificity than the previous inhibitors. Administration of this compound induces in vivo anti-tumor effects through activation of the immune system. Indeed, this aptamer prevents T cells from apoptosis and restores T cell-mediated immunity (86). This study did not evaluate aptamer dependence on glycans, so this point remains to be clarified.
Carbohydrate-independent galectin inhibitors
Apart from their extracellular glycan-dependent functions, galectins also display intracellular functions, most of which are glycan-independent. Therefore, the development of molecules inhibiting these functions may be convenient. In this respect, small benzimidazole compounds (LLS2 and the improved LLS30) bind to the interface between the dimeric galectin-1 subunits within 6 Å from the β-galactoside binding pocket (106). The binding of these compounds to galectin-1 decreased membrane-associated H-Ras and K-Ras and contributed to the suppression of CXCR4, pErk, and AKT signaling pathways (88, 106, 107). Interestingly, pre-treatment of prostate tumor cells with LLS30 reduced their adhesion on collagen-, fibronectin-, and laminin-coated surfaces (107). In vivo administration of these compounds promotes anti-cancer effects in ovarian (106), hepatic (87), malignant peripheral nerve sheath (88), and prostate (107) pre-clinical cancer models. Importantly, combining these compounds with taxanes in in vitro and in vivo experiments resulted in synergistic cytotoxicity against several human cancer cell lines (ovarian, pancreatic, prostatic, and breast cancer cells) (106). These compounds have a direct cytotoxic effect on tumor cells and the cancer-associated stroma (e.g., fibroblasts) (87).
In addition, two tetrahydroisoquinoline natural products (DX-52-1 and HUK-921) inhibit cell migration through interactions with galectin-3 (228). This interaction occurs outside the β-galactoside-binding site of galectin-3. While this compound’s exact mechanism of action remains to be understood, experiments demonstrated that this effect is glycan-independent (228).
While the use of dominant negative mutants for in vivo therapies is still way off, this type of inhibitor allowed us to understand several aspects of the glycan-independent intracellular signaling of galectins. For example, galectins-1 and -3 are constituents of the pre-mRNA splicing machinery (229–233). This interaction is glycan-independent (234), and a N-terminal galectin-3 polypeptide exhibited a dominant negative effect on splicing (231). Interestingly, silencing of galectin-3 was sufficient to alter the splicing patterns of several genes, including the transcripts coding for the SET nuclear oncogene (235). Moreover, galectin-3 regulates promoter activity of different genes highly involved in malignant transformation such as cyclin D1 (236), FOXD1 (237), the thyroid-specific transcription factor TTF-1 (238), and MUC2 (239). A galectin-3 mutant that cannot be phosphorylated at the Ser6 site demonstrated that this post-translational modification is critical for galectin-3 function as a modulator of gene expression (78, 240).
At the cytoplasm, galectins-1 and -3 are recruited by the small GTPase Ras, which become integral parts of plasma membrane nanoclusters (241). Indeed, mutations in a hydrophobic pocket of the galectin-1 CRD induce a dominant negative mutant that cannot interact with H-Ras anymore and, therefore, abrogates signal output (242). Nevertheless, the biological interaction between galectins and Ras does not depend on carbohydrate binding (242, 243). Inspired by that observation, a galectin-3 dominant negative was also created. Similarly, this galectin-3 dominant negative does not interact with K-Ras anymore and abrogates signal output from the Raf/mitogen-activated protein (MAP)/extracellular signal-regulated kinase (ERK; MEK) pathway (241, 244, 245). This initial molecular model of galectin-Ras interactions was then revised by demonstrating that galectin-1 does not directly bind to H-Ras, but instead to the Ras binding domain of Ras effectors, such as Raf (246). Whatever the exact interactor in Ras signaling, galectin-1 and -3 dominant negative mutants reduce cell growth and transformation (243–245). Finally, dominant negative galectins interfere with another type of cytoplasmic interactions with regulatory potential for tumorigenesis. Indeed, galectin-3 bears the NWGR conserved motif with several members of the Bcl-2 family, and using a galectin-3 mutant modifies this delicate balance between cell survival and death (247). In conclusion, several reports have shown the utility of inhibiting the carbohydrate-independent functions of galectins. No report is yet found on their use in pre-clinical as well as clinical trials.
Negative control of galectin gene expression (ablation of all its functions)
Since the description and widespread use of RNA interference to control gene expression, its use to inhibit galectins has been intensive. RNA interference strategies include transient (siRNA) or stable (shRNA-encoding vectors) effectors. Interestingly, this strategy should affect galectin functions more than former inhibitors since it modulates glycan-mediated and -independent effects, and with higher specificity since the nucleotide sequence is highly different between galectins’ members. It is impossible to cite all the publications that have used this approach to downregulate galectins in this review; we only mention a few examples. Indeed, RNA interference was often used to confirm basic aspects of tumor biology (which includes intrinsic effects on the transformed cells themselves (88–91, 99–101, 103, 237, 248–258), the modulation of the tumor-associated stroma (80, 96, 97, 201, 259–265) and, importantly, as a synergic therapy option for cancer (37, 42, 92, 98, 101, 102, 104, 266–270). Several properties of this gene control strategy deserve to be highlighted compared to the aforementioned galectin inhibitors. First, these inhibitory molecules have the highest reported affinities for their messenger RNA target. Indeed, siRNA concentrations in the picomolar range can induce efficient gene expression knockdown, and intracellular amounts of less than 2,000 siRNAs molecules per cell were demonstrated to induce potent biological effects (271). Second, the actions of this type of inhibitor are highly specific. Indeed, siRNAs can downregulate the expression of mRNA transcripts through a highly specific nucleotide hybridization process; it can differentiate single base changes in genes (272, 273). These two properties (affinity and selectivity) make siRNA (and their chemical modifications) an efficient approach to inhibit any target through their gene expression knockdown, and their evaluation in clinical trials is promising [reviewed in (274–277)]. Although protein-based drugs, including monoclonal antibodies, are highly specific, their targets are primarily limited to cell surface receptors or circulating proteins. On the contrary, specific degradation of the galectin transcript by siRNA leads to significant protein downregulation, affecting all the functions galectins are involved in, independently of their glycan dependence. However, various hurdles must be resolved before bringing siRNA into clinical use. First, a selective biodistribution (it would be highly desirable to address siRNA towards the tumor or the tumor-associated stroma, avoiding a non-specific biodistribution that would be responsible for adverse effects). Second, it is needed to improve siRNA stability and reduce their clearance to increase their half-life in the biological fluids. Finally, it is necessary to prevent off-target effects including nucleotide-based immune activation (278, 279). To do this, delivery systems have been developed to protect siRNA from nuclease degradation and facilitate cellular uptake at target sites [chemically modified RNAs (280, 281), nanoparticles (37, 92, 93, 282) and lipoplexes (283)]. These strategies have demonstrated effectiveness to some extent. However, all these approaches face different problems concerning safety, production costs, and often poor correlation between in vitro and in vivo efficacy, making their development a significant challenge.
On the other hand, the endogenous expression of several galectins is subject to gene control by miRNAs. It has been reported that miRNA-22 and -2467 regulate the expression of galectin-1 (284–286), miR-424-3p, -873 and -128 regulate galectin-3 (105, 287–290), miR-1236-3p regulates galectin-8 (291) and miR -455-5p and -22 regulate galectin-9 (292, 293). This finding offers another level of intervention that could be of great interest as therapeutical strategies for various cancers. For example, the utility of miR-424-3p modulation has been demonstrated for ovarian and colorectal cancers (105, 287, 288). In this regard, it has been shown that resveratrol stimulates the transcription of miR-424-3p, which suppresses the expression of galectin-3 (105). In the future, it is expected that the development of gene control strategies through miRNAs will provide new means for controlling galectin levels in the tumor microenvironment.
Finally, developing genome editing strategies such as CRISPR Cas-9 for galectins in the clinic is confronted with ethical obstacles (induction of genome alterations in non-targeted cells) (294, 295). Indeed, the safe and effective delivery of genome editing enzymes represents a substantial challenge that must be tackled to enable the next generation of genetic therapies. However, such genetic strategies will probably contribute to a better fundamental understanding of the role of galectins in cancer. Despite this limitation regarding their direct in vivo use in cancer patients, these strategies could represent real options for in vitro approaches (development of cell-based anti-tumor vaccines or cell conditioning before being infused into patients) (296).
Challenges for clinical application of galectin inhibitors
This chapter itemizes the properties that differentiate galectin inhibitors from each other, and that should be taken into account when scaling up their use in the clinic:
1-Affinity: this is one of the most distinctive parameters of current inhibitors. In general, molecules with higher affinity will require lower doses to obtain in vivo biological effects and, therefore, may induce fewer adverse effects (297, 298). However, it should be noted that affinity calculations are performed in vitro; these molecules have IC50 (inhibitory concentration 50) ranging from μM to pM (as discussed throughout the review for each type of inhibitor). Routine methods used to measure affinity and selectivity include fluorescence polarization binding (299), competitive binding enzyme-linked immunosorbent assays (300), isothermal titration calorimetry (301), biolayer interferometry (138), and surface plasmon resonance (302). These binding assays primarily focus on the CRD, although other non-CRD interactions can also be detected (159). In the case of genetic-based strategies, inhibitors are evaluated by determining galectin transcript or protein levels and functional assays. This methodological heterogeneity makes assessing inhibition potency a real challenge. In addition, although these in vitro determinations allow the compounds to be compared with each other in controlled conditions, they do not define their real inhibitory capacity in vivo. Indeed, in addition to affinity determination in controlled conditions, several other parameters will determine their in vivo inhibitory potential. We can cite their abilities to diffuse across membranes (which determine their tissue biodistribution and extra/intra-cellular localization), the properties of the local microenvironment, and the presence of other biological competitive interactors (141).
2-Specificity for a galectin member (and isoform): this is another fundamental challenge in the field of galectin inhibitors due to the high amino acid sequence homology in the core site between the different members of the galectins (303, 304). Compounds should recognize the correct galectin member. Moreover, several galectin members display multiple isoforms generated from alternative splicing [we can cite galectins-8 (305), -9 (306), and -12 ((307, 308) LGALS12 galectin 12 [Homo sapiens (human)]-Gene-NCBI)]. In this context, gene inhibition strategies are compelling alternatives in terms of specificity. However, other post-translational modifications generate galectin variants such as the cleaved or phosphorylated forms of galectin-3 (240, 309–311) and the O-GlcNAcylation of galectins; this last modification plays a major role in their secretion (312–315). Furthermore, it is worth noting that the quaternary structural conformations of galectins are highly dependent on the properties of the microenvironment. For example, the balance between galectin-1 monomers and dimers depends on the redox state of the cellular microenvironment (316).
Inhibitor specificity is a major point since different galectin members (and even different isoforms) often induce opposite biological effects (317–319) (240, 311, 320, 321). Therefore, the in vivo biological results can be complex if compounds simultaneously inhibit different galectin members (or different isoforms). Furthermore, many galectins play relevant physiological roles (13, 322). Thus, the ideal galectin inhibitor should alter tumor pathology without affecting physiological processes. These inhibitory molecules should be as selective as possible for a particular galectin member (appropriate isoform).
With state-of-the-art, it is not easy to establish a pecking order as to which galectin member should be inhibited to obtain maximal anti-cancer effects. All scientific reports that focus on individual galectins extol their experimental findings. However, to our best knowledge, no systematic study compared the anti-cancer effects obtained by inhibiting multiple galectins (individually or combined) using the same experimental design, especially considering the in vivo complexity. In addition, this scenario is complex since each type of cancer has particularities, so this study must be carried out for each cancer.
3-Galectin function(s) that should be inhibited in cancer: galectin-mediated biological processes in cancer involve interactions more complex than initially proposed and not only restricted to glycan-dependent ones (Figure 1). In this context, there is a lot of information about the glycan-dependent functions of galectins. On the contrary, our comprehension of the glycan-independent ones is more limited. At this level, an implicit question in selecting the best galectin inhibitory strategy for cancer is: what function(s) of these proteins should be preferentially inhibited? Is it sufficient to inhibit the lectin-mediated functions of galectins, or should the non-lectin functions also be inhibited for maximum anti-tumor activity? Noteworthy, complete inhibition of galectins by RNA interference-based approaches was generally used to confirm already-known biological functions of galectins (Table 1). To the best of our knowledge, no new biological functions have been reported by using these approaches. Therefore, more research is needed to clarify this point and to define which galectin functions should be targeted for cancer treatments.
4-Where galectin inhibition should be accomplished: This point is closely related to the previous one. Since galectins play relevant physiological functions, it would be highly advantageous to inhibit them selectively where they play a role in tumorigenesis. In this sense, we have some clues for certain galectins. For instance, galectin-1 downregulation in transformed (17, 123, 131–135) and tumor-associated stroma cells (46, 47, 125, 126, 136) have demonstrated beneficial effects in pre-clinical studies. Therefore, these reports clarify the cellular targets where galectin-1 should be inhibited to obtain beneficial anti-tumor effects. In addition, the sub-cellular localization where galectins play their functional roles must also be considered. For instance, galectin-3 was described at different sub-cellular compartments; inhibition of this protein in each of these localizations often causes opposite biological effects (182). These questions should be addressed for all the galectin members.
5- Appropriate pharmacokinetics; specific biodistribution towards the cell targets: Several of these inhibitors are polar molecules, of low molecular weight, with different capabilities to diffuse through the plasma membrane and, therefore, acting inside the cell (141). On the contrary, large molecules such as inhibitory antibodies are predicted only to engage extracellular galectins. Moreover, like most molecules, galectin inhibitors are trapped in organs with high blood flow, such as the liver, and inactivated through metabolic processes. Moreover, small molecules generally suffer rapid renal clearance (323). Such phenomena reduces the half-life of these molecules, and in consequence, their inhibitory efficiency. Furthermore, other pharmacokinetic properties may also be taken into consideration. In particular, many of the inhibitors are sensitive to enzymatic hydrolysis by glycosidases (324), proteases (325) or nucleases (326). Additionally, inhibitors’ random biodistribution can generate adverse effects due to the inhibition of galectins in tumor-unrelated cells. Therefore, developing degradation-resistant molecules with tumor (and its stroma)-selective biodistribution would be highly desirable.
6- Not expensive and easy translation to clinics should also be addressed.
7- Development of resistance to inhibitory treatments: tumors are highly dynamic biological entities capable of surviving by inducing resistance mechanisms. In the case of inhibiting the lectin functions of galectins, it is worth noting that the glycome is highly adjustable (by enzymatic remodeling without requiring neosynthesis). Thus, we might think that tumor cells would be capable of changing the glycan structures through sialylations (327) or sulfations (328); modifications which have a high impact on galectin biological effects. Otherwise, the same reasoning applies to glycan-independent functions of galectins and resistance development. In this context, it has been shown that the synergism between different treatments allows the use of lower doses of compounds and thus avoids the development of resistance (329). Therefore, this topic represents a significant issue for their transfer to the clinics.
Faced with the critical challenges of galectin inhibitors, regulating the cell glycosylation pattern appears as an alternative option [reviewed in (330, 331)]. Indeed, the creation of glycan ligands for galectins depends on the activities of various glycosyltransferases and glycosidases in the cell (332). In pre-clinical studies, glycome regulation is obtained through control of glycosyltransferases and glycosidases-coding genes (333–339), the use of metabolic inhibitors of glycan biosynthesis (340, 341), or carbohydrate-specific and blocking antibodies (342–344). While such biological disruptions are easily obtained at a pre-clinical level, their therapeutic implementation in patients must also overcome important challenges. In particular, as the glycome is a major determinant of multiple physiological processes, it is essential to avoid side effects. Once again, this type of intervention should be tumor (or tumor-associated stroma)-selective. Moreover, it is pertinent to point out that glycome regulation would only affect some galectin functions (those glycan-dependent). On the other hand, certain galectin inhibitors affect broader functions (including glycan-independent ones such as gene control). The authors consider that both strategies (galectin and glycome regulations) should be evaluated more in-depth, and synergistic or additive anti-tumor effects could be obtained through their combinations.
Final considerations
The first reports about the usefulness of galectin inhibitors appeared in the early 2000s. Since then, a remarkable compendium of basic studies supports their potential utility in cancer, especially in synergy with other treatments (Table 1). However, none of the described galectin inhibitors have achieved clinical success; most did not go beyond the initial phases of clinical trials (Table 2). A detailed analysis of this Table 2 shows that most studies did not translate into better treatments for patients, not even in a better fundamental understanding, as results are often not reported. Therefore, clinical and pre-clinical results must be communicated (even if the observed results differ from those expected) since they contribute to the continuous amelioration of these strategies.
Analyzing all the inhibition strategies reported so far, the authors opine that molecular biology techniques (e.g., RNA interference) offer attractive advantages in affinity and member specificity compared to inhibitors with a carbohydrate nature or those obtained from chemical synthesis. In the case of blocking antibodies, there are important biodistribution drawbacks, which limit galectin inhibition in specific cellular compartments. Despite these particular aspects, much remains to be understood about the pharmacokinetic parameters, toxicity, and tumor resistance mechanisms for all galectin inhibitors.
Finally, since the available literature indicates that galectin inhibition induces effective anti-tumor effects, especially when combined with other strategies (e.g., irradiation, anti-angiogenic, chemotherapies, etc.), this concept should also be considered when designing therapeutic approaches. We conclude that many basic studies are still needed for an efficient clinical translation of galectin inhibitors.
Statements
Author contributions
DL and DC writing—review and editing. All authors contributed to the article and approved the submitted version.
Funding
Research at the Molecular and Functional Glyco-Oncology laboratory was funded by Worldwide Cancer Research (Scotland, UK) and the Argentinean National Cancer Institute. Currently, it is funded by the Argentinean National Agency for Promotion of Science and Technology (PICT2019-01451 andPICT 2020-00298).
Acknowledgments
We apologize to the many authors whose papers could not be cited owing to space limitations. We would like to thank all the members of the Molecular and Functional Glyco-Oncology laboratory for the constant and fruitful scientific discussions. We are also grateful to the staff of the Flow Cytometry and Animal Core Facilities for helpful assistance in the experimental procedures of our projects. DL and DC are members of the scientific career of the National Research Council (CONICET, Argentina).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1
CummingsRDLiuFTRabinovichGAStowellSRVastaGR. Galectins. In: VarkiACummingsRDEskoJDStanleyPHartGWAebiMMohnenDKinoshitaTPackerNHPrestegardJHSchnaarRLSeebergerPH, editors. Essentials of glycobiology, 4th edition, vol. Chapter 36 . Cold Spring Harbor (NY: Cold Spring Harbor Laboratory Press (2022). doi: 10.1101/glycobiology.4e.36
2
GabiusH-J. Galectins: (Much) more than Ga(Lactose-Binding)Lectins. Glycoforum (2021) 24(1):A1. doi: 10.32285/glycoforum.24A1
3
WatanabeMNakamuraOMuramotoKOgawaT. Allosteric regulation of the carbohydrate-binding ability of a novel conger eel galectin by d-mannoside. J Biol Chem (2012) 287(37):31061–72. doi: 10.1074/jbc.M112.346213
4
WellsVMallucciL. Identification of an autocrine negative growth factor: Mouse beta-Galactoside-Binding protein is a cytostatic factor and cell growth regulator. Cell (1991) 64(1):91–7. doi: 10.1016/0092-8674(91)90211-G
5
CompagnoDJaworskiFMGentiliniLContrufoGGonzalez PerezIElolaMTet al. Galectins: Major signaling modulators inside and outside the cell. Curr Mol Med (2014) 14(5):630–51. doi: 10.2174/1566524014666140603101953
6
LiuFTPattersonRJWangJL. Intracellular functions of galectins. Biochim Biophys Acta (2002) 1572(2-3):263–73. doi: 10.1016/S0304-4165(02)00313-6
7
SanjurjoLBroekhuizenECKoenenRRThijssenV. Galectokines: The promiscuous relationship between galectins and cytokines. Biomolecules (2022) 12(9). doi: 10.3390/biom12091286
8
LaderachDJCompagnoD. Unraveling how tumor-derived galectins contribute to anti-cancer immunity failure. Cancers (Basel) (2021) 13(18). doi: 10.3390/cancers13184529
9
VilenZJoehECritcherMParkerCGHuangML. Proximity tagging identifies the glycan-mediated glycoprotein interactors of galectin-1 in muscle stem cells. ACS Chem Biol (2021) 16(10):1994–2003. doi: 10.1021/acschembio.1c00313
10
JoehEO'LearyTLiWHawkinsRHungJRParkerCGet al. Mapping glycan-mediated galectin-3 interactions by live cell proximity labeling. Proc Natl Acad Sci U.S.A. (2020) 117(44):27329–38. doi: 10.1073/pnas.2009206117
11
ObermannJPriglingerCSMerl-PhamJGeerlofAPriglingerSGotzMet al. Proteome-wide identification of glycosylation-dependent interactors of galectin-1 and galectin-3 on mesenchymal retinal pigment epithelial (Rpe) cells. Mol Cell Proteomics (2017) 16(8):1528–46. doi: 10.1074/mcp.M116.066381
12
ElolaMTChiesaMEAlbertiAFMordohJFinkNE. Galectin-1 receptors in different cell types. J BioMed Sci (2005) 12(1):13–29. doi: 10.1007/s11373-004-8169-5
13
LaderachDJCompagnoDToscanoMACrociDODergan-DylonSSalatinoMet al. Dissecting the signal transduction pathways triggered by galectin-glycan interactions in physiological and pathological settings. IUBMB Life (2010) 62(1):1–13. doi: 10.1002/iub.281
14
ThijssenVLHeusschenRCaersJGriffioenAW. Galectin expression in cancer diagnosis and prognosis: A systematic review. Biochim Biophys Acta (2015) 1855(2):235–47. doi: 10.1016/j.bbcan.2015.03.003
15
CompagnoDTiraboschiCGarciaJDRondonYCorapiEVelazquezCet al. Galectins as checkpoints of the immune system in cancers, their clinical relevance, and implication in clinical trials. Biomolecules (2020) 10(5). doi: 10.3390/biom10050750
16
GirottiMRSalatinoMDalotto-MorenoTRabinovichGA. Sweetening the hallmarks of cancer: Galectins as multifunctional mediators of tumor progression. J Exp Med (2020) 217(2). doi: 10.1084/jem.20182041
17
ElolaMTWolfenstein-TodelCTroncosoMFVastaGRRabinovichGA. Galectins: Matricellular glycan-binding proteins linking cell adhesion, migration, and survival. Cell Mol Life Sci (2007) 64(13):1679–700. doi: 10.1007/s00018-007-7044-8
18
LiuFTRabinovichGA. Galectins as modulators of tumour progression. Nat Rev Cancer (2005) 5(1):29–41. doi: 10.1038/nrc1527
19
MirceaAZinovkinDPranjolZI. Vascular modulation of antitumor immunity: A crosstalk between immune cells and the tumor vasculature. In: RezaeiN, editor. Handbook of cancer and immunology. Springer Nature Switzerland (2022). p. 1–27. AG 2022. doi: 10.1007/978-3-030-80962-1_273-1
20
GriffioenAWThijssenVL. Galectins in tumor angiogenesis. Ann Transl Med (2014) 2(9):90. doi: 10.3978/j.issn.2305-5839.2014.09.01
21
Martinez-BoschNNavarroP. Galectins in the tumor microenvironment: Focus on galectin-1. Adv Exp Med Biol (2020) 1259:17–38. doi: 10.1007/978-3-030-43093-1_2
22
van den BruleFAWaltregnyDCastronovoV. Increased expression of galectin-1 in carcinoma-associated stroma predicts poor outcome in prostate carcinoma patients. J Pathol (2001) 193(1):80–7. doi: 10.1002/1096-9896(2000)9999:9999<::AID-PATH730>3.0.CO;2-2
23
SzokeTKayserKBaumhakelJDTrojanIFurakJTiszlaviczLet al. Prognostic significance of endogenous Adhesion/Growth-regulatory lectins in lung cancer. Oncology (2005) 69(2):167–74. doi: 10.1159/000087841
24
Dube-DelarosbilCSt-PierreY. The emerging role of galectins in high-fatality cancers. Cell Mol Life Sci (2018) 75(7):1215–26. doi: 10.1007/s00018-017-2708-5
25
UpretiMJyotiAJohnsonSESwindellEPNapierDSethiPet al. Radiation-enhanced therapeutic targeting of galectin-1 enriched malignant stroma in triple negative breast cancer. Oncotarget (2016) 7(27):41559–74. doi: 10.18632/oncotarget.9490
26
DingsRPWilliamsBWSongCWGriffioenAWMayoKHGriffinRJ. Anginex synergizes with radiation therapy to inhibit tumor growth by radiosensitizing endothelial cells. Int J Cancer (2005) 115(2):312–9. doi: 10.1002/ijc.20850
27
KuoPLeQT. Galectin-1 links tumor hypoxia and radiotherapy. Glycobiology (2014) 24(10):921–5. doi: 10.1093/glycob/cwu062
28
UpretiMJamshidi-ParsianAApanaSBerridgeMFologeaDAKoonceNAet al. Radiation-induced galectin-1 by endothelial cells: A promising molecular target for preferential drug delivery to the tumor vasculature. J Mol Med (Berl) (2013) 91(4):497–506. doi: 10.1007/s00109-012-0965-1
29
HuangEYChenYFChenYMLinIHWangCCSuWHet al. A novel radioresistant mechanism of galectin-1 mediated by h-Ras-Dependent pathways in cervical cancer cells. Cell Death Dis (2012) 3:e251. doi: 10.1038/cddis.2011.120
30
GriffinRJKoonceNADingsRPSiegelEMorosEGBrauer-KrischEet al. Microbeam radiation therapy alters vascular architecture and tumor oxygenation and is enhanced by a galectin-1 targeted anti-angiogenic peptide. Radiat Res (2012) 177(6):804–12. doi: 10.1667/rr2784.1
31
JiaDKoonceNAHalakattiRLiXYaccobySSwainFLet al. Repression of multiple myeloma growth and preservation of bone with combined radiotherapy and anti-angiogenic agent. Radiat Res (2010) 173(6):809–17. doi: 10.1667/RR1734.1
32
DingsRPLorenMHeunHMcNielEGriffioenAWMayoKHet al. Scheduling of radiation with angiogenesis inhibitors anginex and avastin improves therapeutic outcome Via vessel normalization. Clin Cancer Res (2007) 13(11):3395–402. doi: 10.1158/1078-0432.CCR-06-2441
33
AmanoMSuzukiMAndohSMonzenHTeraiKWilliamsBet al. Antiangiogenesis therapy using a novel angiogenesis inhibitor, anginex, following radiation causes tumor growth delay. Int J Clin Oncol (2007) 12(1):42–7. doi: 10.1007/s10147-006-0625-y
34
LinCIWhangEEDonnerDBJiangXPriceBDCarothersAMet al. Galectin-3 targeted therapy with a small molecule inhibitor activates apoptosis and enhances both chemosensitivity and radiosensitivity in papillary thyroid cancer. Mol Cancer Res (2009) 7(10):1655–62. doi: 10.1158/1541-7786.MCR-09-0274
35
LeungZKoFCFTeySKKwongEMLMaoXLiuBHMet al. Galectin-1 promotes hepatocellular carcinoma and the combined therapeutic effect of Otx008 galectin-1 inhibitor and sorafenib in tumor cells. J Exp Clin Cancer Res (2019) 38(1):423. doi: 10.1186/s13046-019-1402-x
36
WangFLvPGuYLiLGeXGuoG. Galectin-1 knockdown improves drug sensitivity of breast cancer by reducing p-glycoprotein expression through inhibiting the raf-1/Ap-1 signaling pathway. Oncotarget (2017) 8(15):25097–106. doi: 10.18632/oncotarget.15341
37
Van WoenselMMathivetTWauthozNRosiereRGargADAgostinisPet al. Sensitization of glioblastoma tumor micro-environment to chemo- and immunotherapy by galectin-1 intranasal knock-down strategy. Sci Rep (2017) 7(1):1217. doi: 10.1038/s41598-017-01279-1
38
NamKSonSHOhSJeonDKimHNohDYet al. Binding of galectin-1 to integrin Beta1 potentiates drug resistance by promoting survivin expression in breast cancer cells. Oncotarget (2017) 8(22):35804–23. doi: 10.18632/oncotarget.16208
39
SuYCDavuluriGVChenCHShiauDCChenCCChenCLet al. Galectin-1-Induced autophagy facilitates cisplatin resistance of hepatocellular carcinoma. PloS One (2016) 11(2):e0148408. doi: 10.1371/journal.pone.0148408
40
ZucchettiMBonezziKFrapolliRSalaFBorsottiPZangariniMet al. Pharmacokinetics and antineoplastic activity of galectin-1-Targeting Otx008 in combination with sunitinib. Cancer Chemother Pharmacol (2013) 72(4):879–87. doi: 10.1007/s00280-013-2270-2
41
TarighatSSFeiFJooEJAbdel-AzimHYangLGengHet al. Overcoming microenvironment-mediated chemoprotection through stromal galectin-3 inhibition in acute lymphoblastic leukemia. Int J Mol Sci (2021) 22(22). doi: 10.3390/ijms222212167
42
WangDYouDLiL. Galectin-3 regulates chemotherapy sensitivity in epithelial ovarian carcinoma Via regulating mitochondrial function. J Toxicol Sci (2019) 44(1):47–56. doi: 10.2131/jts.44.47
43
DingsRPVan LaarESWebberJZhangYGriffinRJWatersSJet al. Ovarian tumor growth regression using a combination of vascular targeting agents anginex or topomimetic 0118 and the chemotherapeutic irofulven. Cancer Lett (2008) 265(2):270–80. doi: 10.1016/j.canlet.2008.02.048
44
DingsRPYokoyamaYRamakrishnanSGriffioenAWMayoKH. The designed angiostatic peptide anginex synergistically improves chemotherapy and antiangiogenesis therapy with angiostatin. Cancer Res (2003) 63(2):382–5.
45
NambiarDKAguileraTCaoHKwokSKongCBloomsteinJet al. Galectin-1-Driven T cell exclusion in the tumor endothelium promotes immunotherapy resistance. J Clin Invest (2019) 129(12):5553–67. doi: 10.1172/JCI129025
46
DingsRPVangKBCastermansKPopescuFZhangYOude EgbrinkMGet al. Enhancement of T-Cell-Mediated antitumor response: Angiostatic adjuvant to immunotherapy against cancer. Clin Cancer Res (2011) 17(10):3134–45. doi: 10.1158/1078-0432.CCR-10-2443
47
IngrassiaLNshimyumukizaPDewelleJLefrancFWlodarczakLThomasSet al. A lactosylated steroid contributes in vivo therapeutic benefits in experimental models of mouse lymphoma and human glioblastoma. J Med Chem (2006) 49(5):1800–7. doi: 10.1021/jm050971v
48
ItoKRalphSJ. Inhibiting galectin-1 reduces murine lung metastasis with increased Cd4(+) and Cd8 (+) T cells and reduced cancer cell adherence. Clin Exp Metastasis (2012) 29(6):561–72. doi: 10.1007/s10585-012-9471-7
49
ItoKScottSACutlerSDongLFNeuzilJBlanchardHet al. Thiodigalactoside inhibits murine cancers by concurrently blocking effects of galectin-1 on immune dysregulation, angiogenesis and protection against oxidative stress. Angiogenesis (2011) 14(3):293–307. doi: 10.1007/s10456-011-9213-5
50
DelaineTCollinsPMacKinnonASharmaGStegmayrJRajputVKet al. Galectin-3-Binding glycomimetics that strongly reduce bleomycin-induced lung fibrosis and modulate intracellular glycan recognition. Chembiochem (2016) 17(18):1759–70. doi: 10.1002/cbic.201600285
51
MackinnonACGibbonsMAFarnworthSLLefflerHNilssonUJDelaineTet al. Regulation of transforming growth factor-Beta1-Driven lung fibrosis by galectin-3. Am J Respir Crit Care Med (2012) 185(5):537–46. doi: 10.1164/rccm.201106-0965OC
52
VuongLKouverianouERooneyCMMcHughBJHowieSEMGregoryCDet al. An orally active galectin-3 antagonist inhibits lung adenocarcinoma growth and augments response to pd-L1 blockade. Cancer Res (2019) 79(7):1480–92. doi: 10.1158/0008-5472.CAN-18-2244
53
ZhangHLiuPZhangYHanLHuZCaiZet al. Inhibition of galectin-3 augments the antitumor efficacy of pd-L1 blockade in non-Small-Cell lung cancer. FEBS Open Bio (2021) 11(3):911–20. doi: 10.1002/2211-5463.13088
54
YinPCuiSLiaoXYaoX. Galectin3 blockade suppresses the growth of cetuximabresistant human oral squamous cell carcinoma. Mol Med Rep (2021) 24(4). doi: 10.3892/mmr.2021.12325
55
GlinskiiOVSudSMossineVVMawhinneyTPAnthonyDCGlinskyGVet al. Inhibition of prostate cancer bone metastasis by synthetic tf antigen Mimic/Galectin-3 inhibitor lactulose-L-Leucine. Neoplasia (2012) 14(1):65–73. doi: 10.1593/neo.111544
56
GlinskyGVPriceJEGlinskyVVMossineVVKiriakovaGMetcalfJB. Inhibition of human breast cancer metastasis in nude mice by synthetic glycoamines. Cancer Res (1996) 56(23):5319–24.
57
PereiraPMSilvaSRamalhoJSGomesCMGiraoHCavaleiroJAet al. The role of galectin-1 in in vitro and in vivo photodynamic therapy with a galactodendritic porphyrin. Eur J Cancer (2016) 68:60–9. doi: 10.1016/j.ejca.2016.08.018
58
PandeyKRDoughertyTJ. Galectin recognized photosensitizers for photodynamic therapy. Washington, DC: U.S. Patent and Trademark Office. Patent US6849607B2 (2005).
59
PlattDRazA. Modulation of the lung colonization of B16-F1 melanoma cells by citrus pectin. J Natl Cancer Inst (1992) 84(6):438–42. doi: 10.1093/jnci/84.6.438
60
MenachemABodnerOPastorJRazAKloogY. Inhibition of malignant thyroid carcinoma cell proliferation by ras and galectin-3 inhibitors. Cell Death Discovery (2015) 1:15047. doi: 10.1038/cddiscovery.2015.47
61
Nangia-MakkerPHoganVHonjoYBaccariniSTaitLBresalierRet al. Inhibition of human cancer cell growth and metastasis in nude mice by oral intake of modified citrus pectin. J Natl Cancer Inst (2002) 94(24):1854–62. doi: 10.1093/jnci/94.24.1854
62
PientaKJNaikHAkhtarAYamazakiKReplogleTSLehrJet al. Inhibition of spontaneous metastasis in a rat prostate cancer model by oral administration of modified citrus pectin. J Natl Cancer Inst (1995) 87(5):348–53. doi: 10.1093/jnci/87.5.348
63
DemotteNWieersGvan der SmissenPMoserMSchmidtCThielemansKet al. A galectin-3 ligand corrects the impaired function of human Cd4 and Cd8 tumor-infiltrating lymphocytes and favors tumor rejection in mice. Cancer Res (2010) 70(19):7476–88. doi: 10.1158/0008-5472.CAN-10-0761
64
KlyosovAZomerEDavid PlattD. Davanat® (Gm-Ct-01) and colon cancer: Preclinical and clinical (Phase I and ii) studies. ACS Symposium Series: Glycobiology Drug Design (2012) 1102:89–130. doi: 10.1021/bk-2012-1102.ch004
65
SturgillERRoligASLinchSNMickCKasiewiczMJSunZet al. Galectin-3 inhibition with belapectin combined with anti-Ox40 therapy reprograms the tumor microenvironment to favor anti-tumor immunity. Oncoimmunology (2021) 10(1):1892265. doi: 10.1080/2162402X.2021.1892265
66
SubudhiMBJainAHurkatPShilpiSGulbakeAJainSK. Eudragit S100 coated citrus pectin nanoparticles for colon targeting of 5-fluorouracil. Materials (Basel) (2015) 8(3):832–49. doi: 10.3390/ma8030832
67
GuYZhaoYZhangZHaoJZhengYLiuQet al. An antibody-like polymeric nanoparticle removes intratumoral galectin-1 to enhance antitumor T-cell responses in cancer immunotherapy. ACS Appl Mater Interfaces (2021) 13(19):22159–68. doi: 10.1021/acsami.1c02116
68
GriffioenAWvan der SchaftDWBarendsz-JansonAFCoxAStruijker BoudierHAHillenHFet al. Anginex, a designed peptide that inhibits angiogenesis. Biochem J (2001) 354(Pt 2):233–42. doi: 10.1042/bj3540233
69
ThijssenVLPostelRBrandwijkRJDingsRPNesmelovaISatijnSet al. Galectin-1 is essential in tumor angiogenesis and is a target for antiangiogenesis therapy. Proc Natl Acad Sci U.S.A. (2006) 103(43):15975–80. doi: 10.1073/pnas.0603883103
70
MayoKHDingsRPFladerCNesmelovaIHargittaiBvan der SchaftDWet al. Design of a partial peptide mimetic of anginex with antiangiogenic and anticancer activity. J Biol Chem (2003) 278(46):45746–52. doi: 10.1074/jbc.M308608200
71
DingsRPKumarNMillerMCLorenMRangwalaHHoyeTRet al. Structure-based optimization of angiostatic agent 6dbf7, an allosteric antagonist of galectin-1. J Pharmacol Exp Ther (2013) 344(3):589–99. doi: 10.1124/jpet.112.199646
72
GheysenLSoumoyLTrelcatAVersetLJourneFSaussezS. New treatment strategy targeting galectin-1 against thyroid cancer. Cells (2021) 10(5). doi: 10.3390/cells10051112
73
DingsRPChenXHellebrekersDMvan EijkLIZhangYHoyeTRet al. Design of nonpeptidic topomimetics of antiangiogenic proteins with antitumor activities. J Natl Cancer Inst (2006) 98(13):932–6. doi: 10.1093/jnci/djj247
74
DingsRPLevineJIBrownSGAstorgues-XerriLMacDonaldJRHoyeTRet al. Polycationic calixarene Ptx013, a potent cytotoxic agent against tumors and drug resistant cancer. Invest New Drugs (2013) 31(5):1142–50. doi: 10.1007/s10637-013-9932-0
75
MirandolaLYuYChuiKJenkinsMRCobosEJohnCMet al. Galectin-3c inhibits tumor growth and increases the anticancer activity of bortezomib in a murine model of human multiple myeloma. PloS One (2011) 6(7):e21811. doi: 10.1371/journal.pone.0021811
76
JohnCMLefflerHKahl-KnutssonBSvenssonIJarvisGA. Truncated galectin-3 inhibits tumor growth and metastasis in orthotopic nude mouse model of human breast cancer. Clin Cancer Res (2003) 9(6):2374–83.
77
WangXGaurMRodriguezHJMounzihKQiuHChenMet al. Galectin-3 inhibition reduces cardiac fibrosis and prevents progressive heart failure following myocardial infarction. Circulation (2019) 140(Abstract 10591).
78
MazurekNSunYJPriceJERamdasLSchoberWNangia-MakkerPet al. Phosphorylation of galectin-3 contributes to malignant transformation of human epithelial cells Via modulation of unique sets of genes. Cancer Res (2005) 65(23):10767–75. doi: 10.1158/0008-5472.CAN-04-3333
79
CrociDOCerlianiJPDalotto-MorenoTMendez-HuergoSPMascanfroniIDDergan-DylonSet al. Glycosylation-dependent lectin-receptor interactions preserve angiogenesis in anti-vegf refractory tumors. Cell (2014) 156(4):744–58. doi: 10.1016/j.cell.2014.01.043
80
CrociDOSalatinoMRubinsteinNCerlianiJPCavallinLELeungHJet al. Disrupting galectin-1 interactions with n-glycans suppresses hypoxia-driven angiogenesis and tumorigenesis in kaposi's sarcoma. J Exp Med (2012) 209(11):1985–2000. doi: 10.1084/jem.20111665
81
StasenkoMSmithEYekuOParkKJLasterILeeKet al. Targeting galectin-3 with a high-affinity antibody for inhibition of high-grade serous ovarian cancer and other Muc16/Ca-125-Expressing malignancies. Sci Rep (2021) 11(1):3718. doi: 10.1038/s41598-021-82686-3
82
YangRSunLLiCFWangYHYaoJLiHet al. Galectin-9 interacts with pd-1 and Tim-3 to regulate T cell death and is a target for cancer immunotherapy. Nat Commun (2021) 12(1):832. doi: 10.1038/s41467-021-21099-2
83
de Mingo PulidoAGardnerAHieblerSSolimanHRugoHSKrummelMFet al. Tim-3 regulates Cd103(+) dendritic cell function and response to chemotherapy in breast cancer. Cancer Cell (2018) 33(1):60–74 e6. doi: 10.1016/j.ccell.2017.11.019
84
DaleyDManiVRMohanNAkkadNOchiAHeindelDWet al. Dectin 1 activation on macrophages by galectin 9 promotes pancreatic carcinoma and peritumoral immune tolerance. Nat Med (2017) 23(5):556–67. doi: 10.1038/nm.4314
85
KikushigeYMiyamotoTYudaJJabbarzadeh-TabriziSShimaTTakayanagiSet al. A Tim-3/Gal-9 autocrine stimulatory loop drives self-renewal of human myeloid leukemia stem cells and leukemic progression. Cell Stem Cell (2015) 17(3):341–52. doi: 10.1016/j.stem.2015.07.011
86
TsaiYTLiangCHYuJHHuangKCTungCHWuJEet al. A DNA aptamer targeting galectin-1 as a novel immunotherapeutic strategy for lung cancer. Mol Ther Nucleic Acids (2019) 18:991–8. doi: 10.1016/j.omtn.2019.10.029
87
TsaiYTLiCYHuangYHChangTSLinCYChuangCHet al. Galectin-1 orchestrates an inflammatory tumor-stroma crosstalk in hepatoma by enhancing Tnfr1 protein stability and signaling in carcinoma-associated fibroblasts. Oncogene (2022) 41(21):3011–23. doi: 10.1038/s41388-022-02309-7
88
ShihTCFanYKissSLiXDengXNLiuRet al. Galectin-1 inhibition induces cell apoptosis through dual suppression of Cxcr4 and ras pathways in human malignant peripheral nerve sheath tumors. Neuro Oncol (2019) 21(11):1389–400. doi: 10.1093/neuonc/noz093
89
YouXWangYWuJLiuQChenDTangDet al. Galectin-1 promotes metastasis in gastric cancer through a sphingosine-1-Phosphate receptor 1-dependent mechanism. Cell Physiol Biochem (2018) 51(1):11–30. doi: 10.1159/000495157
90
MiaoJHWangSQZhangMHYuFBZhangLYuZXet al. Knockdown of galectin-1 suppresses the growth and invasion of osteosarcoma cells through inhibition of the Mapk/Erk pathway. Oncol Rep (2014) 32(4):1497–504. doi: 10.3892/or.2014.3358
91
HsuYLWuCYHungJYLinYSHuangMSKuoPL. Galectin-1 promotes lung cancer tumor metastasis by potentiating integrin Alpha6beta4 and Notch1/Jagged2 signaling pathway. Carcinogenesis (2013) 34(6):1370–81. doi: 10.1093/carcin/bgt040
92
DanhierFMessaoudiKLemaireLBenoitJPLagarceF. Combined anti-Galectin-1 and anti-egfr sirna-loaded chitosan-lipid nanocapsules decrease temozolomide resistance in glioblastoma: In vivo evaluation. Int J Pharm (2015) 481(1-2):154–61. doi: 10.1016/j.ijpharm.2015.01.051
93
Van WoenselMWauthozNRosiereRMathieuVKissRLefrancFet al. Development of sirna-loaded chitosan nanoparticles targeting galectin-1 for the treatment of glioblastoma multiforme Via intranasal administration. J Control Release (2016) 227:71–81. doi: 10.1016/j.jconrel.2016.02.032
94
CambyIBelotNLefrancFSadeghiNde LaunoitYKaltnerHet al. Galectin-1 modulates human glioblastoma cell migration into the brain through modifications to the actin cytoskeleton and levels of expression of small gtpases. J Neuropathol Exp Neurol (2002) 61(7):585–96. doi: 10.1093/jnen/61.7.585
95
Le MercierMMathieuVHaibe-KainsBBontempiGMijatovicTDecaesteckerCet al. Knocking down galectin 1 in human Hs683 glioblastoma cells impairs both angiogenesis and endoplasmic reticulum stress responses. J Neuropathol Exp Neurol (2008) 67(5):456–69. doi: 10.1097/NEN.0b013e318170f892
96
LaderachDJGentiliniLDGiribaldiLDelgadoVCNugnesLCrociDOet al. A unique galectin signature in human prostate cancer progression suggests galectin-1 as a key target for treatment of advanced disease. Cancer Res (2013) 73(1):86–96. doi: 10.1158/0008-5472.CAN-12-1260
97
StortiPMarchicaVAiroldiIDonofrioGFioriniEFerriVet al. Galectin-1 suppression delineates a new strategy to inhibit myeloma-induced angiogenesis and tumoral growth in vivo. Leukemia (2016) 30(12):2351–63. doi: 10.1038/leu.2016.137
98
MathieuVLe MercierMDe NeveNSauvageSGrasTRolandIet al. Galectin-1 knockdown increases sensitivity to temozolomide in a B16f10 mouse metastatic melanoma model. J Invest Dermatol (2007) 127(10):2399–410. doi: 10.1038/sj.jid.5700869
99
SerizawaNTianJFukadaHBaghyKScottFChenXet al. Galectin 3 regulates hcc cell invasion by rhoa and mlck activation. Lab Invest (2015) 95(10):1145–56. doi: 10.1038/labinvest.2015.77
100
BraeuerRRZiglerMKamiyaTDobroffASHuangLChoiWet al. Galectin-3 contributes to melanoma growth and metastasis Via regulation of Nfat1 and autotaxin. Cancer Res (2012) 72(22):5757–66. doi: 10.1158/0008-5472.CAN-12-2424
101
KobayashiTShimuraTYajimaTKuboNArakiKWadaWet al. Transient silencing of galectin-3 expression promotes both in vitro and in vivo drug-induced apoptosis of human pancreatic carcinoma cells. Clin Exp Metastasis (2011) 28(4):367–76. doi: 10.1007/s10585-011-9376-x
102
TiraboschiCGentiliniLVelazquezCCorapiEJaworskiFMGarciaJDet al. Combining inhibition of galectin-3 with and before a therapeutic vaccination is critical for the prostate-tumor free outcome. J ImmunoTherapy Cancer (2020) 8(2):e001535. doi: 10.1136/jitc-2020-001535
103
GentiliniLDJaworskiFMTiraboschiCPerezIGKotlerMLChauchereauAet al. Stable and high expression of galectin-8 tightly controls metastatic progression of prostate cancer. Oncotarget (2017) 8(27):44654–68. doi: 10.18632/oncotarget.17963
104
KimSWParkKCJeonSMOhnTBKimTIKimWHet al. Abrogation of galectin-4 expression promotes tumorigenesis in colorectal cancer. Cell Oncol (Dordr) (2013) 36(2):169–78. doi: 10.1007/s13402-013-0124-x
105
El-KottAFShatiAAAli Al-KahtaniMAlharbiSA. The apoptotic effect of resveratrol in ovarian cancer cells is associated with downregulation of galectin-3 and stimulating mir-424-3p transcription. J Food Biochem (2019) 43(12):e13072. doi: 10.1111/jfbc.13072
106
ShihTCLiuRFungGBhardwajGGhoshPMLamKS. A novel galectin-1 inhibitor discovered through one-bead two-compound library potentiates the antitumor effects of paclitaxel in vivo. Mol Cancer Ther (2017) 16(7):1212–23. doi: 10.1158/1535-7163.MCT-16-0690
107
ShihTCLiuRWuCTLiXXiaoWDengXet al. Targeting galectin-1 impairs castration-resistant prostate cancer progression and invasion. Clin Cancer Res (2018) 24(17):4319–31. doi: 10.1158/1078-0432.CCR-18-0157
108
KeizmanDFrenkelMPeerAKushnirIRosenbaumESaridDet al. Modified citrus pectin treatment in non-metastatic biochemically relapsed prostate cancer: Results of a prospective phase ii study. Nutrients (2021) 13(12). doi: 10.3390/nu13124295
109
GuessBWScholzMCStrumSBLamRYJohnsonHJJennrichRI. Modified citrus pectin (Mcp) increases the prostate-specific antigen doubling time in men with prostate cancer: A phase ii pilot study. Prostate Cancer Prostatic Dis (2003) 6(4):301–4. doi: 10.1038/sj.pcan.4500679
110
CurtiBDKoguchiYLeidnerRSRoligASSturgillERSunZet al. Enhancing clinical and immunological effects of anti-Pd-1 with belapectin, a galectin-3 inhibitor. J Immunother Cancer (2021) 9(4). doi: 10.1136/jitc-2021-002371
111
LauESLiuEPaniaguaSMSarmaAAZampierolloGLopezBet al. Galectin-3 inhibition with modified citrus pectin in hypertension. JACC Basic Transl Sci (2021) 6(1):12–21. doi: 10.1016/j.jacbts.2020.10.006
112
SethiASanamSAlvalaM. Non-carbohydrate strategies to inhibit lectin proteins with special emphasis on galectins. Eur J Med Chem (2021) 222:113561. doi: 10.1016/j.ejmech.2021.113561
113
ChanYCLinHYTuZKuoYHHsuSDLinCH. Dissecting the structure-activity relationship of galectin-ligand interactions. Int J Mol Sci (2018) 19(2). doi: 10.3390/ijms19020392
114
WdowiakKFrancuzTGallego-ColonERuiz-AgamezNKubeczkoMGrocholaIet al. Galectin targeted therapy in oncology: Current knowledge and perspectives. Int J Mol Sci (2018) 19(1). doi: 10.3390/ijms19010210
115
CagnoniAJPerez SaezJMRabinovichGAMarinoKV. Turning-off signaling by siglecs, selectins, and galectins: Chemical inhibition of glycan-dependent interactions in cancer. Front Oncol (2016) 6:109. doi: 10.3389/fonc.2016.00109
116
ObergCTLefflerHNilssonUJ. Inhibition of galectins with small molecules. Chimia (Aarau) (2011) 65(1-2):18–23. doi: 10.2533/chimia.2011.18
117
BlanchardHBum-ErdeneKHugoMW. Inhibitors of galectins and implications for structure-based design of galectin-specific therapeutics. Aust J Chem (2014) 67(12):1763–79. doi: 10.1071/CH14362
118
BlanchardHBum-ErdeneKBohariMHYuX. Galectin-1 inhibitors and their potential therapeutic applications: A patent review. Expert Opin Ther Pat (2016) 26(5):537–54. doi: 10.1517/13543776.2016.1163338
119
RoyRMurphyPVGabiusHJ. Multivalent carbohydrate-lectin interactions: How synthetic chemistry enables insights into nanometric recognition. Molecules (2016) 21(5). doi: 10.3390/molecules21050629
120
LefflerHNilssonUJ. Low-molecular weight inhibitors of galectins. In: Galectins and disease implications for targeted therapeutics. American Chemical Society (ACS) Symposium Series, vol. 1115. (2012) 47–59. doi: 10.1021/bk-2012-1115.ch002
121
HirabayashiJHashidateTArataYNishiNNakamuraTHirashimaMet al. Oligosaccharide specificity of galectins: A search by frontal affinity chromatography. Biochim Biophys Acta (2002) 1572(2-3):232–54. doi: 10.1016/S0304-4165(02)00311-2
122
TejlerJSalamehBLefflerHNilssonUJ. Fragment-based development of triazole-substituted O-galactosyl aldoximes with fragment-induced affinity and selectivity for galectin-3. Org Biomol Chem (2009) 7(19):3982–90. doi: 10.1039/b909091f
123
ÖbergCTNoressonA-LLefflerHNilssonUJ. Synthesis of 3-Amido-3-De-Oxy-B-D-Talopyranosides: All-Cis-Substituted pyranosides as lectin inhibitors. Tetrahedron (2011) 67(47):9164–72. doi: 10.1016/j.tet.2011.09.098
124
ÖbergCTBlanchardHLefflerHNilssonUJ. Protein subtype-targeting through ligand epimerization: Talose-selectivity of galectin-4 and galectin-8. Bioorg Med Chem Lett (2008) 18(13):3691–4. doi: 10.1016/j.bmcl.2008.05.066
125
GiguereDBoninMACloutierPPatnamRSt-PierreCSatoSet al. Synthesis of stable and selective inhibitors of human galectins-1 and -3. Bioorg Med Chem (2008) 16(16):7811–23. doi: 10.1016/j.bmc.2008.06.044
126
BaillyCThuruXQuesnelB. Modulation of the gal-9/Tim-3 immune checkpoint with alpha-lactose. does anomery of lactose matter? Cancers (Basel) (2021) 13(24). doi: 10.3390/cancers13246365
127
GiguereDSatoSSt-PierreCSiroisSRoyR. Aryl O- and s-galactosides and lactosides as specific inhibitors of human galectins-1 and -3: Role of electrostatic potential at O-3. Bioorg Med Chem Lett (2006) 16(6):1668–72. doi: 10.1016/j.bmcl.2005.12.010
128
van KlaverenSDernovsekJJakopinZAnderluhMLefflerHNilssonUJet al. Design and synthesis of novel 3-Triazolyl-1-Thiogalactosides as galectin-1, -3 and -8 inhibitors. RSC Adv (2022) 12(29):18973–84. doi: 10.1039/d2ra03163a
129
van HattumHBranderhorstHMMoretEENilssonUJLefflerHPietersRJ. Tuning the preference of thiodigalactoside- and lactosamine-based ligands to galectin-3 over galectin-1. J Med Chem (2013) 56(3):1350–4. doi: 10.1021/jm301677r
130
SalamehBACumpsteyISundinALefflerHNilssonUJ. 1h-1,2,3-Triazol-1-Yl thiodigalactoside derivatives as high affinity galectin-3 inhibitors. Bioorg Med Chem (2010) 18(14):5367–78. doi: 10.1016/j.bmc.2010.05.040
131
van ScherpenzeelMMoretEEBallellLLiskampRMNilssonUJLefflerHet al. Synthesis and evaluation of new thiodigalactoside-based chemical probes to label galectin-3. Chembiochem (2009) 10(10):1724–33. doi: 10.1002/cbic.200900198
132
CumpsteyISalomonssonESundinALefflerHNilssonUJ. Double affinity amplification of galectin-ligand interactions through arginine-arene interactions: Synthetic, thermodynamic, and computational studies with aromatic diamido thiodigalactosides. Chemistry (2008) 14(14):4233–45. doi: 10.1002/chem.200701932
133
Bum-ErdeneKCollinsPMHugoMWTarighatSSFeiFKishorCet al. Novel selective galectin-3 antagonists are cytotoxic to acute lymphoblastic leukemia. J Med Chem (2022) 65(8):5975–89. doi: 10.1021/acs.jmedchem.1c01296
134
CollinsPMObergCTLefflerHNilssonUJBlanchardH. Taloside inhibitors of galectin-1 and galectin-3. Chem Biol Drug Des (2012) 79(3):339–46. doi: 10.1111/j.1747-0285.2011.01283.x
135
KishorCRossRLBlanchardH. Lactulose as a novel template for anticancer drug development targeting galectins. Chem Biol Drug Des (2018) 92(4):1801–8. doi: 10.1111/cbdd.13348
136
IngrassiaLCambyILefrancFMathieuVNshimyumukizaPDarroFet al. Anti-galectin compounds as potential anti-cancer drugs. Curr Med Chem (2006) 13(29):3513–27. doi: 10.2174/092986706779026219
137
DriguezH. Thiooligosaccharides as tools for structural biology. Chembiochem (2001) 2(5):311–8. doi: 10.1002/1439-7633(20010504)2:5<311::AID-CBIC311>3.0.CO;2-L
138
HsiehTJLinHYTuZLinTCWuSCTsengYYet al. Dual thio-Digalactoside-Binding modes of human galectins as the structural basis for the design of potent and selective inhibitors. Sci Rep (2016) 6:29457. doi: 10.1038/srep29457
139
HiraniNMacKinnonACNicolLFordPSchambyeHPedersenAet al. Target inhibition of galectin-3 by inhaled Td139 in patients with idiopathic pulmonary fibrosis. Eur Respir J (2021) 57(5). doi: 10.1183/13993003.02559-2020
140
ZetterbergFRPetersonKJohnssonREBrimertTHakanssonMLoganDTet al. Monosaccharide derivatives with low-nanomolar lectin affinity and high selectivity based on combined fluorine-amide, phenyl-arginine, sulfur-pi, and halogen bond interactions. ChemMedChem (2018) 13(2):133–7. doi: 10.1002/cmdc.201700744
141
StegmayrJZetterbergFCarlssonMCHuangXSharmaGKahl-KnutsonBet al. Extracellular and intracellular small-molecule galectin-3 inhibitors. Sci Rep (2019) 9(1):2186. doi: 10.1038/s41598-019-38497-8
142
GlinskyVVKiriakovaGGlinskiiOVMossineVVMawhinneyTPTurkJRet al. Synthetic galectin-3 inhibitor increases metastatic cancer cell sensitivity to taxol-induced apoptosis in vitro and in vivo. Neoplasia (2009) 11(9):901–9. doi: 10.1593/neo.09594
143
RabinovichGACumashiABiancoGACiavardelliDIurisciID'EgidioMet al. Synthetic lactulose amines: Novel class of anticancer agents that induce tumor-cell apoptosis and inhibit galectin-mediated homotypic cell aggregation and endothelial cell morphogenesis. Glycobiology (2006) 16(3):210–20. doi: 10.1093/glycob/cwj056
144
DahlqvistAMandalSPetersonKHakanssonMLoganDTZetterbergFRet al. 3-substituted 1-Naphthamidomethyl-C-Galactosyls interact with two unique Sub-sites for high-affinity and high-selectivity inhibition of galectin-3. Molecules (2019) 24(24). doi: 10.3390/molecules24244554
145
TejlerJTullbergEFrejdTLefflerHNilssonUJ. Synthesis of multivalent lactose derivatives by 1,3-dipolar cycloadditions: Selective galectin-1 inhibition. Carbohydr Res (2006) 341(10):1353–62. doi: 10.1016/j.carres.2006.04.028
146
TejlerJSkogmanFLefflerHNilssonUJ. Synthesis of galactose-mimicking 1h-(1,2,3-Triazol-1-Yl)-Mannosides as selective galectin-3 and 9n inhibitors. Carbohydr Res (2007) 342(12-13):1869–75. doi: 10.1016/j.carres.2007.03.012
147
WangGNAndreSGabiusHJMurphyPV. Bi- to tetravalent glycoclusters: Synthesis, structure-activity profiles as lectin inhibitors and impact of combining both valency and headgroup tailoring on selectivity. Org Biomol Chem (2012) 10(34):6893–907. doi: 10.1039/c2ob25870f
148
GiguereDAndreSBoninMABellefleurMAProvencalACloutierPet al. Inhibitory potential of chemical substitutions at bioinspired sites of beta-D-Galactopyranose on Neoglycoprotein/Cell surface binding of two classes of medically relevant lectins. Bioorg Med Chem (2011) 19(10):3280–7. doi: 10.1016/j.bmc.2011.03.022
149
GouinSGGarcia FernandezJMVanquelefEDupradeauFYSalomonssonELefflerHet al. Multimeric lactoside "Click clusters" as tools to investigate the effect of linker length in specific interactions with peanut lectin, galectin-1, and -3. Chembiochem (2010) 11(10):1430–42. doi: 10.1002/cbic.201000167
150
AndreSSansoneFKaltnerHCasnatiAKopitzJGabiusHJet al. Calix[N]Arene-based glycoclusters: Bioactivity of thiourea-linked Galactose/Lactose moieties as inhibitors of binding of medically relevant lectins to a glycoprotein and cell-surface glycoconjugates and selectivity among human Adhesion/Growth-regulatory galectins. Chembiochem (2008) 9(10):1649–61. doi: 10.1002/cbic.200800035
151
AndreSKaltnerHFuruikeTNishimuraSGabiusHJ. Persubstituted cyclodextrin-based glycoclusters as inhibitors of protein-carbohydrate recognition using purified plant and mammalian lectins and wild-type and lectin-Gene-Transfected tumor cells as targets. Bioconjug Chem (2004) 15(1):87–98. doi: 10.1021/bc0340666
152
AndreSPietersRJVrasidasIKaltnerHKuwabaraILiuFTet al. Wedgelike glycodendrimers as inhibitors of binding of mammalian galectins to glycoproteins, lactose maxiclusters, and cell surface glycoconjugates. Chembiochem (2001) 2(11):822–30. doi: 10.1002/1439-7633(20011105)2:11<822::AID-CBIC822>3.0.CO;2-W
153
MichelAKNangia-MakkerPRazACloningerMJ. Lactose-functionalized dendrimers arbitrate the interaction of galectin-3/Muc1 mediated cancer cellular aggregation. Chembiochem (2014) 15(14):2106–12. doi: 10.1002/cbic.201402134
154
SilvaSPereiraPMSilvaPPazFAFaustinoMACavaleiroJAet al. Porphyrin and phthalocyanine glycodendritic conjugates: Synthesis, photophysical and photochemical properties. Chem Commun (Camb) (2012) 48(30):3608–10. doi: 10.1039/c2cc17561d
155
ZhangWXuPZhangH. Pectin in cancer therapy: A review. Trends Food Sci Technol (2015) 44(2):258–71. doi: 10.1016/j.tifs.2015.04.001
156
GaoXZhiYZhangTXueHWangXFodayADet al. Analysis of the neutral polysaccharide fraction of mcp and its inhibitory activity on galectin-3. Glycoconj J (2012) 29(4):159–65. doi: 10.1007/s10719-012-9382-5
157
LeclereLCutsemPVMichielsC. Anti-cancer activities of ph- or heat-modified pectin. Front Pharmacol (2013) 4:128. doi: 10.3389/fphar.2013.00128
158
JacksonCLDreadenTMTheobaldLKTranNMBealTLEidMet al. Pectin induces apoptosis in human prostate cancer cells: Correlation of apoptotic function with pectin structure. Glycobiology (2007) 17(8):805–19. doi: 10.1093/glycob/cwm054
159
StegmayrJLepurAKahl-KnutsonBAguilar-MoncayoMKlyosovAAFieldRAet al. Low or no inhibitory potency of the canonical galectin carbohydrate-binding site by pectins and galactomannans. J Biol Chem (2016) 291(25):13318–34. doi: 10.1074/jbc.M116.721464
160
MillerMCIppelHSuylenDKlyosovAATraberPGHackengTet al. Binding of polysaccharides to human galectin-3 at a noncanonical site in its carbohydrate recognition domain. Glycobiology (2016) 26(1):88–99. doi: 10.1093/glycob/cwv073
161
MillerMCKlyosovAAMayoKH. Structural features for alpha-galactomannan binding to galectin-1. Glycobiology (2012) 22(4):543–51. doi: 10.1093/glycob/cwr173
162
MillerMCKlyosovAMayoKH. The alpha-galactomannan davanat binds galectin-1 at a site different from the conventional galectin carbohydrate binding domain. Glycobiology (2009) 19(9):1034–45. doi: 10.1093/glycob/cwp084
163
ZhangTMillerMCZhengYZhangZXueHZhaoDet al. Macromolecular assemblies of complex polysaccharides with galectin-3 and their synergistic effects on function. Biochem J (2017) 474(22):3849–68. doi: 10.1042/BCJ20170143
164
ShiHYuLShiYLuJTengHZhouYet al. Structural characterization of a rhamnogalacturonan I domain from ginseng and its inhibitory effect on galectin-3. Molecules (2017) 22(6). doi: 10.3390/molecules22061016
165
GaoXZhiYSunLPengXZhangTXueHet al. The inhibitory effects of a rhamnogalacturonan I (Rg-I) domain from ginseng pectin on galectin-3 and its structure-activity relationship. J Biol Chem (2013) 288(47):33953–65. doi: 10.1074/jbc.M113.482315
166
GunningAPBongaertsRJMorrisVJ. Recognition of galactan components of pectin by galectin-3. FASEB J (2009) 23(2):415–24. doi: 10.1096/fj.08-106617
167
CummingsJHSouthgateDABranchWJWigginsHSHoustonHJenkinsDJet al. The digestion of pectin in the human gut and its effect on calcium absorption and Large bowel function. Br J Nutr (1979) 41(3):477–85. doi: 10.1079/bjn19790062
168
SandbergASAhderinneRAnderssonHHallgrenBHultenL. The effect of citrus pectin on the absorption of nutrients in the small intestine. Hum Nutr Clin Nutr (1983) 37(3):171–83.
169
PedrosaLFRazAFabiJP. The complex biological effects of pectin: Galectin-3 targeting as potential human health improvement? Biomolecules (2022) 12(2). doi: 10.3390/biom12020289
170
LefflerH. Letter by leffler regarding article, "Modified citrus pectin prevents blood-brain barrier disruption in mouse subarachnoid hemorrhage by inhibiting galectin-3". Stroke (2019) 50(5):e136. doi: 10.1161/STROKEAHA.119.024744
171
FanYSunLYangSHeCTaiGZhouY. The roles and mechanisms of homogalacturonan and rhamnogalacturonan I pectins on the inhibition of cell migration. Int J Biol Macromol (2018) 106:207–17. doi: 10.1016/j.ijbiomac.2017.08.004
172
WangLZhaoLGongFLSunCDuDDYangXXet al. Modified citrus pectin inhibits breast cancer development in mice by targeting tumor-associated macrophage survival and polarization in hypoxic microenvironment. Acta Pharmacol Sin (2022) 43(6):1556–67. doi: 10.1038/s41401-021-00748-8
173
WangLLiYSYuLGZhangXKZhaoLGongFLet al. Galectin-3 expression and secretion by tumor-associated macrophages in hypoxia promotes breast cancer progression. Biochem Pharmacol (2020) 178:114113. doi: 10.1016/j.bcp.2020.114113
174
YanJKatzA. Pectasol-c modified citrus pectin induces apoptosis and inhibition of proliferation in human and mouse androgen-dependent and- independent prostate cancer cells. Integr Cancer Ther (2010) 9(2):197–203. doi: 10.1177/1534735410369672
175
JiangJEliazISlivaD. Synergistic and additive effects of modified citrus pectin with two polybotanical compounds, in the suppression of invasive behavior of human breast and prostate cancer cells. Integr Cancer Ther (2013) 12(2):145–52. doi: 10.1177/1534735412442369
176
HosseinGHalvaeiSHeidarianYDehghani-GhobadiZHassaniMHosseiniHet al. Pectasol-c modified citrus pectin targets galectin-3-Induced Stat3 activation and synergize paclitaxel cytotoxic effect on ovarian cancer spheroids. Cancer Med (2019) 8(9):4315–29. doi: 10.1002/cam4.2334
177
HosseinGKeshavarzMAhmadiSNaderiN. Synergistic effects of pectasol-c modified citrus pectin an inhibitor of galectin-3 and paclitaxel on apoptosis of human skov-3 ovarian cancer cells. Asian Pac J Cancer Prev (2013) 14(12):7561–8. doi: 10.7314/apjcp.2013.14.12.7561
178
GrousJJRedfernCHMahadevanmDSchindlerJ. Gcs-100, a galectin-3 antagonist, in refractory solid tumors: A phase I study. J Clin Oncol ASCO Annu Meeting Proc (2006) 24(18 suppl):13023. doi: 10.1200/jco.2006.24.18_suppl.13023
179
CotterFSmithDABoydTERichardsDAAlemanyCLoeschDet al. Single-agent activity of gcs-100, a first-in-Class galectin-3 antagonist, in elderly patients with relapsed chronic lymphocytic leukemia. J Clin Oncol 2009 ASCO Annu Meeting (2009) 27(15 suppl):7006. doi: 10.1200/jco.2009.27.15_suppl.7006
180
DemotteNBigirimanaRWieersGStroobantVSquiffletJLCarrascoJet al. A short treatment with galactomannan gm-Ct-01 corrects the functions of freshly isolated human tumor-infiltrating lymphocytes. Clin Cancer Res (2014) 20(7):1823–33. doi: 10.1158/1078-0432.CCR-13-2459
181
Gordon-AlonsoMHirschTWildmannCvan der BruggenP. Galectin-3 captures interferon-gamma in the tumor matrix reducing chemokine gradient production and T-cell tumor infiltration. Nat Commun (2017) 8(1):793. doi: 10.1038/s41467-017-00925-6
182
FukumoriTTakenakaYYoshiiTKimHRHoganVInoharaHet al. CD29 and CD7 mediate galectin-3-induced type II T-cell apoptosis. Cancer Res (2003) 63(23):8302–11.
183
TraberPGChouHZomerEHongFKlyosovAFielMIet al. Regression of fibrosis and reversal of cirrhosis in rats by galectin inhibitors in thioacetamide-induced liver disease. PloS One (2013) 8(10):e75361. doi: 10.1371/journal.pone.0075361
184
ChalasaniNAbdelmalekMFGarcia-TsaoGVuppalanchiRAlkhouriNRinellaMet al. Effects of belapectin, an inhibitor of galectin-3, in patients with nonalcoholic steatohepatitis with cirrhosis and portal hypertension. Gastroenterology (2020) 158(5):1334–45 e5. doi: 10.1053/j.gastro.2019.11.296
185
LiuQSaccoPMarsichEFurlaniFAribCDjakerNet al. Lactose-modified chitosan Gold(Iii)-pegylated complex-bioconjugates: From synthesis to interaction with targeted galectin-1 protein. Bioconjug Chem (2018) 29(10):3352–61. doi: 10.1021/acs.bioconjchem.8b00520
186
BesfordQAWojnilowiczMSumaTBertleff-ZieschangNCarusoFCavalieriF. Lactosylated glycogen nanoparticles for targeting prostate cancer cells. ACS Appl Mater Interfaces (2017) 9(20):16869–79. doi: 10.1021/acsami.7b02676
187
BiswasSMedinaSHBarchiJJJr. Synthesis and cell-selective antitumor properties of amino acid conjugated tumor-associated carbohydrate antigen-coated gold nanoparticles. Carbohydr Res (2015) 405:93–101. doi: 10.1016/j.carres.2014.11.002
188
KongYLiXLiuXPangJMuXLiuW. Galactosylated chitosan modified magnetic mesoporous silica nanoparticles loaded with nedaplatin for the targeted chemo-photothermal synergistic therapy of cancer. J Nanosci Nanotechnol (2021) 21(9):4553–64. doi: 10.1166/jnn.2021.19142
189
LiuWWangFZhuYLiXLiuXPangJet al. Galactosylated chitosan-functionalized mesoporous silica nanoparticle loading by calcium leucovorin for colon cancer cell-targeted drug delivery. Molecules (2018) 23(12). doi: 10.3390/molecules23123082
190
Garcia CalaviaPChambrierICookMJHainesAHFieldRARussellDA. Targeted photodynamic therapy of breast cancer cells using lactose-phthalocyanine functionalized gold nanoparticles. J Colloid Interface Sci (2018) 512:249–59. doi: 10.1016/j.jcis.2017.10.030
191
SethiASasikalaKJakkulaPGaddeDSanamSQureshiIAet al. Design, synthesis and computational studies involving indole-coumarin hybrids as galectin-1 inhibitors. Chem Papers (2021) 75:2791–805. doi: 10.1007/s11696-021-01534-w
192
Sridhar GoudNPooladandaVMuni ChandraKLakshmi SoukyaPSAlvalaRKumarPet al. Novel benzimidazole-triazole hybrids as apoptosis inducing agents in lung cancer: Design, synthesis, (18)F-radiolabeling & galectin-1 inhibition studies. Bioorg Chem (2020) 102:104125. doi: 10.1016/j.bioorg.2020.104125
193
GoudNSPooladandaVMahammadGSJakkulaPGatreddiSQureshiIAet al. Synthesis and biological evaluation of morpholines linked coumarin-triazole hybrids as anticancer agents. Chem Biol Drug Des (2019) 94(5):1919–29. doi: 10.1111/cbdd.13578
194
GoudNSGhouseMSVishnuJPranayJAlvalaRTallaVet al. Synthesis and biological evaluation of novel heterocyclic imines linked coumarin- thiazole hybrids as anticancer agents. Anticancer Agents Med Chem (2019) 19(4):557–66. doi: 10.2174/1871520619666190207140120
195
GlinskyVVHuflejtMEGlinskyGVDeutscherSLQuinnTP. Effects of thomsen-friedenreich antigen-specific peptide p-30 on beta-Galactoside-Mediated homotypic aggregation and adhesion to the endothelium of mda-Mb-435 human breast carcinoma cells. Cancer Res (2000) 60(10):2584–8.
196
AnananuchatkulTChangIVMikiTTsutsumiHMiharaH. Construction of a stapled alpha-helix peptide library displayed on phage for the screening of galectin-3-Binding peptide ligands. ACS Omega (2020) 5(11):5666–74. doi: 10.1021/acsomega.9b03461
197
SarabojiKHakanssonMGenhedenSDiehlCQvistJWeiningerUet al. The carbohydrate-binding site in galectin-3 is preorganized to recognize a sugarlike framework of oxygens: Ultra-High-Resolution structures and water dynamics. Biochemistry (2012) 51(1):296–306. doi: 10.1021/bi201459p
198
MarkowskaAILiuFTPanjwaniN. Galectin-3 is an important mediator of vegf- and bfgf-mediated angiogenic response. J Exp Med (2010) 207(9):1981–93. doi: 10.1084/jem.20090121
199
EckardtVMillerMCBlanchetXDuanRLeberzammerJDucheneJet al. Chemokines and galectins form heterodimers to modulate inflammation. EMBO Rep (2020) 21(4):e47852. doi: 10.15252/embr.201947852
200
MirandolaLYuYCannonMJJenkinsMRRahmanRLNguyenDDet al. Galectin-3 inhibition suppresses drug resistance, motility, invasion and angiogenic potential in ovarian cancer. Gynecol Oncol (2014) 135(3):573–9. doi: 10.1016/j.ygyno.2014.09.021
201
ZhuCAndersonACSchubartAXiongHImitolaJKhourySJet al. The Tim-3 ligand galectin-9 negatively regulates T helper type 1 immunity. Nat Immunol (2005) 6(12):1245–52. doi: 10.1038/ni1271
202
GaoYLiXShuZZhangKXueXLiWet al. Nuclear galectin-1-Foxp3 interaction dampens the tumor-suppressive properties of Foxp3 in breast cancer. Cell Death Dis (2018) 9(4):416. doi: 10.1038/s41419-018-0448-6
203
ZhengGGrahamAShibataMMissertJROseroffARDoughertyTJet al. Synthesis of beta-Galactose-Conjugated chlorins derived by enyne metathesis as galectin-specific photosensitizers for photodynamic therapy. J Org Chem (2001) 66(26):8709–16. doi: 10.1021/jo0105080
204
SalomonssonEThijssenVLGriffioenAWNilssonUJLefflerH. The anti-angiogenic peptide anginex greatly enhances galectin-1 binding affinity for glycoproteins. J Biol Chem (2011) 286(16):13801–4. doi: 10.1074/jbc.C111.229096
205
DingsRPMillerMCNesmelovaIAstorgues-XerriLKumarNSerovaMet al. Antitumor agent calixarene 0118 targets human galectin-1 as an allosteric inhibitor of carbohydrate binding. J Med Chem (2012) 55(11):5121–9. doi: 10.1021/jm300014q
206
MillerMCZhengYSuylenDIppelHCanadaFJBerbisMAet al. Targeting the crd f-face of human galectin-3 and allosterically modulating glycan binding by angiostatic Ptx008 and a structurally optimized derivative. ChemMedChem (2021) 16(4):713–23. doi: 10.1002/cmdc.202000742
207
Astorgues-XerriLRiveiroMETijeras-RaballandASerovaMRabinovichGABiecheIet al. Otx008, a selective small-molecule inhibitor of galectin-1, downregulates cancer cell proliferation, invasion and tumour angiogenesis. Eur J Cancer (2014) 50(14):2463–77. doi: 10.1016/j.ejca.2014.06.015
208
Perez SaezJMHocklPFCagnoniAJMendez HuergoSPGarciaPAGattoSGet al. Characterization of a neutralizing anti-human galectin-1 monoclonal antibody with angioregulatory and immunomodulatory activities. Angiogenesis (2021) 24(1):1–5. doi: 10.1007/s10456-020-09749-3
209
OuyangJPlutschowAPogge von StrandmannEReinersKSPonaderSRabinovichGAet al. Galectin-1 serum levels reflect tumor burden and adverse clinical features in classical Hodgkin lymphoma. Blood (2013) 121(17):3431–3. doi: 10.1182/blood-2012-12-474569
210
GarinMIChuCCGolshayanDCernuda-MorollonEWaitRLechlerRI. Galectin-1: A key effector of regulation mediated by Cd4+Cd25+ T cells. Blood (2007) 109(5):2058–65. doi: 10.1182/blood-2006-04-016451
211
LiuFTHsuDKZuberiRIHillPNShenhavAKuwabaraIet al. Modulation of functional properties of galectin-3 by monoclonal antibodies binding to the non-lectin domains. Biochemistry (1996) 35(19):6073–9. doi: 10.1021/bi952716q
212
LuLHNakagawaRKashioYItoAShojiHNishiNet al. Characterization of galectin-9-Induced death of jurkat T cells. J Biochem (2007) 141(2):157–72. doi: 10.1093/jb/mvm019
213
ShahbazSDunsmoreGKolevaPXuLHoustonSElahiS. Galectin-9 and vista expression define terminally exhausted T cells in hiv-1 infection. J Immunol (2020) 204(9):2474–91. doi: 10.4049/jimmunol.1901481
214
ZhouQMungerMEVeenstraRGWeigelBJHirashimaMMunnDHet al. Coexpression of Tim-3 and pd-1 identifies a Cd8+ T-cell exhaustion phenotype in mice with disseminated acute myelogenous leukemia. Blood (2011) 117(17):4501–10. doi: 10.1182/blood-2010-10-310425
215
MadireddiSEunSYMehtaAKBirtaAZajoncDMNikiTet al. Regulatory T cell-mediated suppression of inflammation induced by Dr3 signaling is dependent on galectin-9. J Immunol (2017) 199(8):2721–8. doi: 10.4049/jimmunol.1700575
216
OomizuSArikawaTNikiTKadowakiTUenoMNishiNet al. Cell surface galectin-9 expressing Th cells regulate Th17 and Foxp3+ treg development by galectin-9 secretion. PloS One (2012) 7(11):e48574. doi: 10.1371/journal.pone.0048574
217
OomizuSArikawaTNikiTKadowakiTUenoMNishiNet al. Galectin-9 suppresses Th17 cell development in an il-2-Dependent but Tim-3-Independent manner. Clin Immunol (2012) 143(1):51–8. doi: 10.1016/j.clim.2012.01.004
218
SekiMOomizuSSakataKMSakataAArikawaTWatanabeKet al. Galectin-9 suppresses the generation of Th17, promotes the induction of regulatory T cells, and regulates experimental autoimmune arthritis. Clin Immunol (2008) 127(1):78–88. doi: 10.1016/j.clim.2008.01.006
219
WangFWanLZhangCZhengXLiJChenZK. Tim-3-Galectin-9 pathway involves the suppression induced by Cd4+Cd25+ regulatory T cells. Immunobiology (2009) 214(5):342–9. doi: 10.1016/j.imbio.2008.10.007
220
WuCThalhamerTFrancaRFXiaoSWangCHottaCet al. Galectin-9-Cd44 interaction enhances stability and function of adaptive regulatory T cells. Immunity (2014) 41(2):270–82. doi: 10.1016/j.immuni.2014.06.011
221
NobumotoAOomizuSArikawaTKatohSNagaharaKMiyakeMet al. Galectin-9 expands unique macrophages exhibiting plasmacytoid dendritic cell-like phenotypes that activate nk cells in tumor-bearing mice. Clin Immunol (2009) 130(3):322–30. doi: 10.1016/j.clim.2008.09.014
222
NagaharaKArikawaTOomizuSKontaniKNobumotoATatenoHet al. Galectin-9 increases Tim-3+ dendritic cells and Cd8+ T cells and enhances antitumor immunity Via galectin-9-Tim-3 interactions. J Immunol (2008) 181(11):7660–9. doi: 10.4049/jimmunol.181.11.7660
223
YamauchiADaiSYNakagawaRKashioYAbeHKatohSet al. [Galectin-9 induces maturation of human monocyte-derived dendritic cells]. Nihon Rinsho Meneki Gakkai Kaishi (2005) 28(6):381–8. doi: 10.2177/jsci.28.381
224
LimagneERichardCThibaudinMFumetJDTruntzerCLagrangeAet al. Tim-3/Galectin-9 pathway and mmdsc control primary and secondary resistances to pd-1 blockade in lung cancer patients. Oncoimmunology (2019) 8(4):e1564505. doi: 10.1080/2162402X.2018.1564505
225
YangRSunLLiCFWangYHXiaWLiuBet al. Development and characterization of anti-Galectin-9 antibodies that protect T cells from galectin-9-Induced cell death. J Biol Chem (2022) 298(4):101821. doi: 10.1016/j.jbc.2022.101821
226
LhuillierCBarjonCBalocheVNikiTGelinAMustaphaRet al. Characterization of neutralizing antibodies reacting with the 213-224 amino-acid segment of human galectin-9. PloS One (2018) 13(9):e0202512. doi: 10.1371/journal.pone.0202512
227
RaoUSRaoPS. Surface-bound galectin-4 regulates gene transcription and secretion of chemokines in human colorectal cancer cell lines. Tumour Biol (2017) 39(3):1010428317691687. doi: 10.1177/1010428317691687
228
KahsaiAWCuiJKaniskanHUGarnerPPFenteanyG. Analogs of tetrahydroisoquinoline natural products that inhibit cell migration and target galectin-3 outside of its carbohydrate-binding site. J Biol Chem (2008) 283(36):24534–45. doi: 10.1074/jbc.M800006200
229
HaudekKCVossPGWangJLPattersonRJ. A 10s galectin-3-U1 snrnp complex assembles into active spliceosomes. Nucleic Acids Res (2016) 44(13):6391–7. doi: 10.1093/nar/gkw303
230
WangWParkJWWangJLPattersonRJ. Immunoprecipitation of spliceosomal rnas by antisera to galectin-1 and galectin-3. Nucleic Acids Res (2006) 34(18):5166–74. doi: 10.1093/nar/gkl673
231
ParkJWVossPGGrabskiSWangJLPattersonRJ. Association of galectin-1 and galectin-3 with Gemin4 in complexes containing the smn protein. Nucleic Acids Res (2001) 29(17):3595–602. doi: 10.1093/nar/29.17.3595
232
VyakarnamADagherSFWangJLPattersonRJ. Evidence for a role for galectin-1 in pre-mrna splicing. Mol Cell Biol (1997) 17(8):4730–7. doi: 10.1128/MCB.17.8.4730
233
DagherSFWangJLPattersonRJ. Identification of galectin-3 as a factor in pre-mrna splicing. Proc Natl Acad Sci U.S.A. (1995) 92(4):1213–7. doi: 10.1073/pnas.92.4.1213
234
VossPGGrayRMDickeySWWangWParkJWKasaiKet al. Dissociation of the carbohydrate-binding and splicing activities of galectin-1. Arch Biochem Biophys (2008) 478(1):18–25. doi: 10.1016/j.abb.2008.07.003
235
FritschKMernbergerMNistAStieweTBrehmAJacobR. Galectin-3 interacts with components of the nuclear ribonucleoprotein complex. BMC Cancer (2016) 16:502. doi: 10.1186/s12885-016-2546-0
236
LinHMPestellRGRazAKimHR. Galectin-3 enhances cyclin D(1) promoter activity through Sp1 and a camp-responsive element in human breast epithelial cells. Oncogene (2002) 21(52):8001–10. doi: 10.1038/sj.onc.1205820
237
LiCHChangYCHsiaoMLiangSM. Foxd1 and gal-3 form a positive regulatory loop to regulate lung cancer aggressiveness. Cancers (Basel) (2019) 11(12). doi: 10.3390/cancers11121897
238
ParonIScaloniAPinesABachiALiuFTPuppinCet al. Nuclear localization of galectin-3 in transformed thyroid cells: A role in transcriptional regulation. Biochem Biophys Res Commun (2003) 302(3):545–53. doi: 10.1016/S0006-291X(03)00151-7
239
SongSByrdJCMazurekNLiuKKooJSBresalierRS. Galectin-3 modulates Muc2 mucin expression in human colon cancer cells at the level of transcription Via ap-1 activation. Gastroenterology (2005) 129(5):1581–91. doi: 10.1053/j.gastro.2005.09.002
240
YoshiiTFukumoriTHonjoYInoharaHKimHRRazA. Galectin-3 phosphorylation is required for its anti-apoptotic function and cell cycle arrest. J Biol Chem (2002) 277(9):6852–7. doi: 10.1074/jbc.M107668200
241
Shalom-FeuersteinRPlowmanSJRotblatBAriottiNTianTHancockJFet al. K-Ras nanoclustering is subverted by overexpression of the scaffold protein galectin-3. Cancer Res (2008) 68(16):6608–16. doi: 10.1158/0008-5472.CAN-08-1117
242
RotblatBNivHAndreSKaltnerHGabiusHJKloogY. Galectin-1(L11a) predicted from a computed galectin-1 farnesyl-binding pocket selectively inhibits ras-gtp. Cancer Res (2004) 64(9):3112–8. doi: 10.1158/0008-5472.can-04-0026
243
PazAHaklaiRElad-SfadiaGBallanEKloogY. Galectin-1 binds oncogenic h-ras to mediate ras membrane anchorage and cell transformation. Oncogene (2001) 20(51):7486–93. doi: 10.1038/sj.onc.1204950
244
Shalom-FeuersteinRLevyRMakovskiVRazAKloogY. Galectin-3 regulates Rasgrp4-mediated activation of n-ras and h-ras. Biochim Biophys Acta (2008) 1783(6):985–93. doi: 10.1016/j.bbamcr.2008.03.009
245
Shalom-FeuersteinRCooksTRazAKloogY. Galectin-3 regulates a molecular switch from n-ras to K-ras usage in human breast carcinoma cells. Cancer Res (2005) 65(16):7292–300. doi: 10.1158/0008-5472.CAN-05-0775
246
BlazevitsOMideksaYGSolmanMLigabueAAriottiNNakhaeizadehHet al. Galectin-1 dimers can scaffold raf-effectors to increase h-ras nanoclustering. Sci Rep (2016) 6:24165. doi: 10.1038/srep24165
247
AkahaniSNangia-MakkerPInoharaHKimHRRazA. Galectin-3: A novel antiapoptotic molecule with a functional Bh1 (Nwgr) domain of bcl-2 family. Cancer Res (1997) 57(23):5272–6.
248
ArcoliaVJourneFWattierALeteurtreERenaudFGabiusHJet al. Galectin-1 is a diagnostic marker involved in thyroid cancer progression. Int J Oncol (2017) 51(3):760–70. doi: 10.3892/ijo.2017.4065
249
ParkGBChungYHKimD. Induction of galectin-1 by tlr-dependent Pi3k activation enhances epithelial-mesenchymal transition of metastatic ovarian cancer cells. Oncol Rep (2017) 37(5):3137–45. doi: 10.3892/or.2017.5533
250
ShenKHLiCFChienLHHuangCHSuCCLiaoACet al. Role of galectin-1 in urinary bladder urothelial carcinoma cell invasion through the jnk pathway. Cancer Sci (2016) 107(10):1390–8. doi: 10.1111/cas.13016
251
CimminoFSchulteJHZolloMKosterJVersteegRIolasconAet al. Galectin-1 is a major effector of trkb-mediated neuroblastoma aggressiveness. Oncogene (2009) 28(19):2015–23. doi: 10.1038/onc.2009.70
252
CambyIDecaesteckerCLefrancFKaltnerHGabiusHJKissR. Galectin-1 knocking down in human U87 glioblastoma cells alters their gene expression pattern. Biochem Biophys Res Commun (2005) 335(1):27–35. doi: 10.1016/j.bbrc.2005.07.037
253
CoppinLJanninAAit YahyaEThuillierCVillenetCTardivelMet al. Galectin-3 modulates epithelial cell adaptation to stress at the er-mitochondria interface. Cell Death Dis (2020) 11(5):360. doi: 10.1038/s41419-020-2556-3
254
LaSHKimSJKangHGLeeHWChunKH. Ablation of human telomerase reverse transcriptase (Htert) induces cellular senescence in gastric cancer through a galectin-3 dependent mechanism. Oncotarget (2016) 7(35):57117–30. doi: 10.18632/oncotarget.10986
255
LuHLiuYWangDWangLZhouHXuGet al. Galectin-3 regulates metastatic capabilities and chemotherapy sensitivity in epithelial ovarian carcinoma Via nf-kappab pathway. Tumour Biol (2016) 37(8):11469–77. doi: 10.1007/s13277-016-5004-3
256
QiaoLLiangNXieJLuoHZhangJDengGet al. Gene silencing of galectin-3 changes the biological behavior of Eca109 human esophageal cancer cells. Mol Med Rep (2016) 13(1):160–6. doi: 10.3892/mmr.2015.4543
257
SatelliARaoPSThirumalaSRaoUS. Galectin-4 functions as a tumor suppressor of human colorectal cancer. Int J Cancer (2011) 129(4):799–809. doi: 10.1002/ijc.25750
258
MeinohlCBarnardSJFritz-WolfKUngerMPorrAHeipelMet al. Galectin-8 binds to the farnesylated c-terminus of K-Ras4b and modifies Ras/Erk signaling and migration in pancreatic and lung carcinoma cells. Cancers (Basel) (2019) 12(1). doi: 10.3390/cancers12010030
259
ZhouWZhouYChenXNingTChenHGuoQet al. Pancreatic cancer-targeting exosomes for enhancing immunotherapy and reprogramming tumor microenvironment. Biomaterials (2021) 268:120546. doi: 10.1016/j.biomaterials.2020.120546
260
Martinez-BoschNFernandez-BarrenaMGMorenoMOrtiz-ZapaterEMunne-ColladoJIglesiasMet al. Galectin-1 drives pancreatic carcinogenesis through stroma remodeling and hedgehog signaling activation. Cancer Res (2014) 74(13):3512–24. doi: 10.1158/0008-5472.CAN-13-3013
261
HeXJTaoHQHuZMMaYYXuJWangHJet al. Expression of galectin-1 in carcinoma-associated fibroblasts promotes gastric cancer cell invasion through upregulation of integrin Beta1. Cancer Sci (2014) 105(11):1402–10. doi: 10.1111/cas.12539
262
NorlingLVSampaioALCooperDPerrettiM. Inhibitory control of endothelial galectin-1 on in vitro and in vivo lymphocyte trafficking. FASEB J (2008) 22(3):682–90. doi: 10.1096/fj.07-9268com
263
KubachJLutterPBoppTStollSBeckerCHuterEet al. Human Cd4+Cd25+ regulatory T cells: Proteome analysis identifies galectin-10 as a novel marker essential for their anergy and suppressive function. Blood (2007) 110(5):1550–8. doi: 10.1182/blood-2007-01-069229
264
WilliamsSPOdellAFKarnezisTFarnsworthRHGouldCMLiJet al. Genome-wide functional analysis reveals central signaling regulators of lymphatic endothelial cell migration and remodeling. Sci Signal (2017) 10(499). doi: 10.1126/scisignal.aal2987
265
WuMHYingNWHongTMChiangWFLinYTChenYL. Galectin-1 induces vascular permeability through the neuropilin-1/Vascular endothelial growth factor receptor-1 complex. Angiogenesis (2014) 17(4):839–49. doi: 10.1007/s10456-014-9431-8
266
GaoJWangW. Knockdown of galectin-1 facilitated cisplatin sensitivity by inhibiting autophagy in neuroblastoma cells. Chem Biol Interact (2019) 297:50–6. doi: 10.1016/j.cbi.2018.10.014
267
ZhangPShiBZhouMJiangHZhangHPanXet al. Galectin-1 overexpression promotes progression and chemoresistance to cisplatin in epithelial ovarian cancer. Cell Death Dis (2014) 5:e991. doi: 10.1038/cddis.2013.526
268
Le MercierMLefrancFMijatovicTDebeirOHaibe-KainsBBontempiGet al. Evidence of galectin-1 involvement in glioma chemoresistance. Toxicol Appl Pharmacol (2008) 229(2):172–83. doi: 10.1016/j.taap.2008.01.009
269
LeeYKLinTHChangCFLoYL. Galectin-3 silencing inhibits epirubicin-induced atp binding cassette transporters and activates the mitochondrial apoptosis pathway Via beta-Catenin/Gsk-3beta modulation in colorectal carcinoma. PloS One (2013) 8(11):e82478. doi: 10.1371/journal.pone.0082478
270
CheongTCShinJYChunKH. Silencing of galectin-3 changes the gene expression and augments the sensitivity of gastric cancer cells to chemotherapeutic agents. Cancer Sci (2010) 101(1):94–102. doi: 10.1111/j.1349-7006.2009.01364.x
271
WittrupAAiALiuXHamarPTrifonovaRCharisseKet al. Visualizing lipid-formulated sirna release from endosomes and target gene knockdown. Nat Biotechnol (2015) 33(8):870–6. doi: 10.1038/nbt.3298
272
SchwarzDSDingHKenningtonLMooreJTSchelterJBurchardJet al. Designing sirna that distinguish between genes that differ by a single nucleotide. PloS Genet (2006) 2(9):e140. doi: 10.1371/journal.pgen.0020140
273
MartinezLANaguibnevaILehrmannHVervischATchenioTLozanoGet al. Synthetic small inhibiting rnas: Efficient tools to inactivate oncogenic mutations and restore P53 pathways. Proc Natl Acad Sci U.S.A. (2002) 99(23):14849–54. doi: 10.1073/pnas.222406899
274
FriedrichMAignerA. Therapeutic sirna: State-of-the-Art and future perspectives. BioDrugs (2022) 36(5):549–71. doi: 10.1007/s40259-022-00549-3
275
KuSHJoSDLeeYKKimKKimSH. Chemical and structural modifications of rnai therapeutics. Adv Drug Delivery Rev (2016) 104:16–28. doi: 10.1016/j.addr.2015.10.015
276
BattistellaMMarsdenPA. Advances, nuances, and potential pitfalls when exploiting the therapeutic potential of rna interference. Clin Pharmacol Ther (2015) 97(1):79–87. doi: 10.1002/cpt.8
277
BornaHImaniSImanMAzimzadeh JamalkandiS. Therapeutic face of rnai: In vivo challenges. Expert Opin Biol Ther (2015) 15(2):269–85. doi: 10.1517/14712598.2015.983070
278
MarquesJTWilliamsBR. Activation of the mammalian immune system by sirnas. Nat Biotechnol (2005) 23(11):1399–405. doi: 10.1038/nbt1161
279
WinklerJStesslMAmarteyJNoeCR. Off-target effects related to the phosphorothioate modification of nucleic acids. ChemMedChem (2010) 5(8):1344–52. doi: 10.1002/cmdc.201000156
280
BerkCCivenniGWangYSteuerCCatapanoCVHallJ. Pharmacodynamic and pharmacokinetic properties of full phosphorothioate small interfering rnas for gene silencing in vivo. Nucleic Acid Ther (2021) 31(3):237–44. doi: 10.1089/nat.2020.0852
281
IrieASatoKHaraRIWadaTShibasakiF. An artificial cationic oligosaccharide combined with phosphorothioate linkages strongly improves sirna stability. Sci Rep (2020) 10(1):14845. doi: 10.1038/s41598-020-71896-w
282
WangYXieYKilchristKVLiJDuvallCLOupickyD. Endosomolytic and tumor-penetrating mesoporous silica nanoparticles for Sirna/Mirna combination cancer therapy. ACS Appl Mater Interfaces (2020) 12(4):4308–22. doi: 10.1021/acsami.9b21214
283
YousefiABourajjajMBabaeNNoortPISchaapveldRQBeijnumJRet al. Anginex lipoplexes for delivery of anti-angiogenic sirna. Int J Pharm (2014) 472(1-2):175–84. doi: 10.1016/j.ijpharm.2014.06.028
284
QiuBQZhangPFXiongDXuJJLongXZhuSQet al. Circrna fibroblast growth factor receptor 3 promotes tumor progression in non-small cell lung cancer by regulating galectin-1-Akt/Erk1/2 signaling. J Cell Physiol (2019) 234(7):11256–64. doi: 10.1002/jcp.27783
285
YouYTanJXDaiHSChenHWXuXJYangAGet al. Mirna-22 inhibits oncogene galectin-1 in hepatocellular carcinoma. Oncotarget (2016) 7(35):57099–116. doi: 10.18632/oncotarget.10981
286
ZhangPFWuJLuoJHLiKSWangFHuangWet al. Snhg22 overexpression indicates poor prognosis and induces chemotherapy resistance Via the mir-2467/Gal-1 signaling pathway in epithelial ovarian carcinoma. Aging (Albany NY) (2019) 11(19):8204–16. doi: 10.18632/aging.102313
287
BiegDSypniewskiDNowakEBednarekI. Mir-424-3p suppresses galectin-3 expression and sensitizes ovarian cancer cells to cisplatin. Arch Gynecol Obstet (2019) 299(4):1077–87. doi: 10.1007/s00404-018-4999-7
288
LuWWangJYangGYuNHuangZXuHet al. Posttranscriptional regulation of galectin-3 by mir-128 contributes to colorectal cancer progression. Oncotarget (2017) 8(9):15242–51. doi: 10.18632/oncotarget.14839
289
YuWMaBZhaoWLiuJYuHTianZet al. The combination of circrna-Umad1 and galectin-3 in peripheral circulation is a Co-biomarker for predicting lymph node metastasis of thyroid carcinoma. Am J Transl Res (2020) 12(9):5399–415.
290
XieXJiJChenXXuWChenHZhuSet al. Human umbilical cord mesenchymal stem cell-derived exosomes carrying hsa-Mirna-128-3p suppress pancreatic ductal cell carcinoma by inhibiting galectin-3. Clin Transl Oncol (2022) 24(3):517–31. doi: 10.1007/s12094-021-02705-7
291
XuYPDongZNWangSWZhengYMZhangCZhouYQet al. Circhmgcs1-016 reshapes immune environment by sponging mir-1236-3p to regulate Cd73 and gal-8 expression in intrahepatic cholangiocarcinoma. J Exp Clin Cancer Res (2021) 40(1):290. doi: 10.1186/s13046-021-02095-2
292
YangQHouCHuangDZhuangCJiangWGengZet al. Mir-455-5p functions as a potential oncogene by targeting galectin-9 in colon cancer. Oncol Lett (2017) 13(3):1958–64. doi: 10.3892/ol.2017.5608
293
YangQJiangWZhuangCGengZHouCHuangDet al. Microrna-22 downregulation of galectin-9 influences lymphocyte apoptosis and tumor cell proliferation in liver cancer. Oncol Rep (2015) 34(4):1771–8. doi: 10.3892/or.2015.4167
294
KhuranaASayedNSinghVKhuranaIAllawadhiPRawatPSet al. A comprehensive overview of Crispr/Cas 9 technology and application thereof in drug discovery. J Cell Biochem (2022) 123(10):1674–98. doi: 10.1002/jcb.30329
295
RaguramABanskotaSLiuDR. Therapeutic in vivo delivery of gene editing agents. Cell (2022) 185(15):2806–27. doi: 10.1016/j.cell.2022.03.045
296
HirakawaMPKrishnakumarRTimlinJACarneyJPButlerKS. Gene editing and crispr in the clinic: Current and future perspectives. Biosci Rep (2020) 40(4). doi: 10.1042/BSR20200127
297
IurisciICumashiAShermanAATsvetkovYETinariNPiccoloEet al. Synthetic inhibitors of galectin-1 and -3 selectively modulate homotypic cell aggregation and tumor cell apoptosis. Anticancer Res (2009) 29(1):403–10.
298
BlanchardHYuXCollinsPMBum-ErdeneK. Galectin-3 inhibitors: A patent review (2008-present). Expert Opin Ther Pat (2014) 24(10):1053–65. doi: 10.1517/13543776.2014.947961
299
SormePKahl-KnutssonBHuflejtMNilssonUJLefflerH. Fluorescence polarization as an analytical tool to evaluate galectin-ligand interactions. Anal Biochem (2004) 334(1):36–47. doi: 10.1016/j.ab.2004.06.042
300
InoharaHRazA. Functional evidence that cell surface galectin-3 mediates homotypic cell adhesion. Cancer Res (1995) 55(15):3267–71.
301
PetersonKKumarRStenstromOVermaPVermaPRHakanssonMet al. Systematic tuning of fluoro-Galectin-3 interactions provides thiodigalactoside derivatives with single-digit nm affinity and high selectivity. J Med Chem (2018) 61(3):1164–75. doi: 10.1021/acs.jmedchem.7b01626
302
ZhangTZhengYZhaoDYanJSunCZhouYet al. Multiple approaches to assess pectin binding to galectin-3. Int J Biol Macromol (2016) 91:994–1001. doi: 10.1016/j.ijbiomac.2016.06.058
303
CooperDN. Galectinomics: Finding themes in complexity. Biochim Biophys Acta (2002) 1572(2-3):209–31. doi: 10.1016/s0304-4165(02)00310-0
304
CooperDNBarondesSH. God Must love galectins; he made so many of them. Glycobiology (1999) 9(10):979–84. doi: 10.1093/glycob/9.10.979
305
Bidon-WagnerNLe PennecJP. Human galectin-8 isoforms and cancer. Glycoconj J (2002) 19(7-9):557–63. doi: 10.1023/B:GLYC.0000014086.38343.98
306
HeusschenRGriffioenAWThijssenVL. Galectin-9 in tumor biology: A jack of multiple trades. Biochim Biophys Acta (2013) 1836(1):177–85. doi: 10.1016/j.bbcan.2013.04.006
307
HottaKFunahashiTMatsukawaYTakahashiMNishizawaHKishidaKet al. Galectin-12, an adipose-expressed galectin-like molecule possessing apoptosis-inducing activity. J Biol Chem (2001) 276(36):34089–97. doi: 10.1074/jbc.M105097200
308
YangRYHsuDKYuLNiJLiuFT. Cell cycle regulation by galectin-12, a new member of the galectin superfamily. J Biol Chem (2001) 276(23):20252–60. doi: 10.1074/jbc.M010914200
309
SaraswatiSBlockASDavidsonMKRankRGMahadevanMDiekmanAB. Galectin-3 is a substrate for prostate specific antigen (Psa) in human seminal plasma. Prostate (2011) 71(2):197–208. doi: 10.1002/pros.21236
310
Nangia-MakkerPRazTTaitLHoganVFridmanRRazA. Galectin-3 cleavage: A novel surrogate marker for matrix metalloproteinase activity in growing breast cancers. Cancer Res (2007) 67(24):11760–8. doi: 10.1158/0008-5472.CAN-07-3233
311
BalanVNangia-MakkerPKhoDHWangYRazA. Tyrosine-phosphorylated galectin-3 protein is resistant to prostate-specific antigen (Psa) cleavage. J Biol Chem (2012) 287(8):5192–8. doi: 10.1074/jbc.C111.331686
312
MathewMPAbramowitzLKDonaldsonJGHanoverJA. Nutrient-responsive O-glcnacylation dynamically modulates the secretion of glycan-binding protein galectin 3. J Biol Chem (2022) 298(3):101743. doi: 10.1016/j.jbc.2022.101743
313
GatieMISpiceDMGarhaAMcTagueAAhmerMTimoshenkoAVet al. O-Glcnacylation and regulation of galectin-3 in extraembryonic endoderm differentiation. Biomolecules (2022) 12(5). doi: 10.3390/biom12050623
314
TazhitdinovaRTimoshenkoAV. The emerging role of galectins and O-glcnac homeostasis in processes of cellular differentiation. Cells (2020) 9(8). doi: 10.3390/cells9081792
315
HartCChaseLGHajivandiMAgnewB. Metabolic labeling and click chemistry detection of glycoprotein markers of mesenchymal stem cell differentiation. Methods Mol Biol (2011) 698:459–84. doi: 10.1007/978-1-60761-999-4_33
316
GuardiaCMCarameloJJTrujilloMMendez-HuergoSPRadiREstrinDAet al. Structural basis of redox-dependent modulation of galectin-1 dynamics and function. Glycobiology (2014) 24(5):428–41. doi: 10.1093/glycob/cwu008
317
TribulattiMVFiginiMGCarabelliJCattaneoVCampetellaO. Redundant and antagonistic functions of galectin-1, -3, and -8 in the elicitation of T cell responses. J Immunol (2012) 188(7):2991–9. doi: 10.4049/jimmunol.1102182
318
StowellSRQianYKarmakarSKoyamaNSDias-BaruffiMLefflerHet al. Differential roles of galectin-1 and galectin-3 in regulating leukocyte viability and cytokine secretion. J Immunol (2008) 180(5):3091–102. doi: 10.4049/jimmunol.180.5.3091
319
StillmanBNHsuDKPangMBrewerCFJohnsonPLiuFTet al. Galectin-3 and galectin-1 bind distinct cell surface glycoprotein receptors to induce T cell death. J Immunol (2006) 176(2):778–89. doi: 10.4049/jimmunol.176.2.778
320
AanhaneESchulkensIAHeusschenRCastricumKLefflerHGriffioenAWet al. Different angioregulatory activity of monovalent galectin-9 isoforms. Angiogenesis (2018) 21(3):545–55. doi: 10.1007/s10456-018-9607-8
321
CagnoniAJTroncosoMFRabinovichGAMarinoKVElolaMT. Full-length galectin-8 and separate carbohydrate recognition domains: The whole is greater than the sum of its parts? Biochem Soc Trans (2020) 48(3):1255–68. doi: 10.1042/BST20200311
322
ArthurCMBaruffiMDCummingsRDStowellSR. Evolving mechanistic insights into galectin functions. Methods Mol Biol (2015) 1207:1–35. doi: 10.1007/978-1-4939-1396-1_1
323
ErnstBMagnaniJL. From carbohydrate leads to glycomimetic drugs. Nat Rev Drug Discovery (2009) 8(8):661–77. doi: 10.1038/nrd2852
324
CagnoniAJKovenskyJUhrigML. Design and synthesis of hydrolytically stable multivalent ligands bearing thiodigalactoside analogues for peanut lectin and human galectin-3 binding. J Org Chem (2014) 79(14):6456–67. doi: 10.1021/jo500883v
325
LaiXTangJElSayedMEH. Recent advances in proteolytic stability for peptide, protein, and antibody drug discovery. Expert Opin Drug Discovery (2021) 16(12):1467–82. doi: 10.1080/17460441.2021.1942837
326
SajidMIMoazzamMKatoSYeseom ChoKTiwariRK. Overcoming barriers for sirna therapeutics: From bench to bedside. Pharm (Basel) (2020) 13(10). doi: 10.3390/ph13100294
327
AmanoMErikssonHManningJCDetjenKMAndreSNishimuraSet al. Tumour suppressor P16(Ink4a) - anoikis-favouring decrease in N/O-Glycan/Cell surface sialylation by down-regulation of enzymes in sialic acid biosynthesis in tandem in a pancreatic carcinoma model. FEBS J (2012) 279(21):4062–80. doi: 10.1111/febs.12001
328
HonkeKTsudaMKoyotaSWadaYIida-TanakaNIshizukaIet al. Molecular cloning and characterization of a human beta-Gal-3'-Sulfotransferase that acts on both type 1 and type 2 (Gal beta 1-3/1-4glcnac-R) oligosaccharides. J Biol Chem (2001) 276(1):267–74. doi: 10.1074/jbc.M005666200
329
Bayat MokhtariRHomayouniTSBaluchNMorgatskayaEKumarSDasBet al. Combination therapy in combating cancer. Oncotarget (2017) 8(23):38022–43. doi: 10.18632/oncotarget.16723
330
MereiterSBalmanaMCamposDGomesJReisCA. Glycosylation in the era of cancer-targeted therapy: Where are we heading? Cancer Cell (2019) 36(1):6–16. doi: 10.1016/j.ccell.2019.06.006
331
DimitroffCJ. Galectin-binding O-glycosylations as regulators of malignancy. Cancer Res (2015) 75(16):3195–202. doi: 10.1158/0008-5472.CAN-15-0834
332
ThiemannSBaumLG. Galectins and immune responses-just how do they do those things they do? Annu Rev Immunol (2016) 34:243–64. doi: 10.1146/annurev-immunol-041015-055402
333
JonesRBDorsettKAHjelmelandABBellisSL. The St6gal-I sialyltransferase protects tumor cells against hypoxia by enhancing hif-1alpha signaling. J Biol Chem (2018) 293(15):5659–67. doi: 10.1074/jbc.RA117.001194
334
KawashimaH. Roles of the gel-forming Muc2 mucin and its O-glycosylation in the protection against colitis and colorectal cancer. Biol Pharm Bull (2012) 35(10):1637–41. doi: 10.1248/bpb.b12-00412
335
AnGWeiBXiaBMcDanielJMJuTCummingsRDet al. Increased susceptibility to colitis and colorectal tumors in mice lacking core 3-derived O-glycans. J Exp Med (2007) 204(6):1417–29. doi: 10.1084/jem.20061929
336
PartridgeEALe RoyCDi GuglielmoGMPawlingJCheungPGranovskyMet al. Regulation of cytokine receptors by golgi n-glycan processing and endocytosis. Science (2004) 306(5693):120–4. doi: 10.1126/science.1102109
337
IharaSMiyoshiEKoJHMurataKNakaharaSHonkeKet al. Prometastatic effect of n-acetylglucosaminyltransferase V is due to modification and stabilization of active matriptase by adding beta 1-6 glcnac branching. J Biol Chem (2002) 277(19):16960–7. doi: 10.1074/jbc.M200673200
338
DemetriouMGranovskyMQuagginSDennisJW. Negative regulation of T-cell activation and autoimmunity by Mgat5 n-glycosylation. Nature (2001) 409(6821):733–9. doi: 10.1038/35055582
339
GranovskyMFataJPawlingJMullerWJKhokhaRDennisJW. Suppression of tumor growth and metastasis in Mgat5-deficient mice. Nat Med (2000) 6(3):306–12. doi: 10.1038/73163
340
Cedeno-LaurentFOppermanMJBarthelSRHaysDSchattonTZhanQet al. Metabolic inhibition of galectin-1-Binding carbohydrates accentuates antitumor immunity. J Invest Dermatol (2012) 132(2):410–20. doi: 10.1038/jid.2011.335
341
BrownJRFusterMMWhisenantTEskoJD. Expression patterns of alpha 2,3-sialyltransferases and alpha 1,3-fucosyltransferases determine the mode of sialyl Lewis X inhibition by disaccharide decoys. J Biol Chem (2003) 278(26):23352–9. doi: 10.1074/jbc.M303093200
342
Haji-GhassemiOBlacklerRJMartin YoungNEvansSV. Antibody recognition of carbohydrate epitopesdagger. Glycobiology (2015) 25(9):920–52. doi: 10.1093/glycob/cwv037
343
SmaletzODizMDdo CarmoCCSabbagaJCunha-JuniorGFAzevedoSJet al. A phase ii trial with anti-Lewis-Y monoclonal antibody (Hu3s193) for the treatment of platinum Resistant/Refractory ovarian, fallopian tube and primary peritoneal carcinoma. Gynecol Oncol (2015) 138(2):272–7. doi: 10.1016/j.ygyno.2015.05.023
344
SakaiKYuasaNTsukamotoKTakasaki-MatsumotoAYajimaYSatoRet al. Isolation and characterization of antibodies against three consecutive tn-antigen clusters from a phage library displaying human single-chain variable fragments. J Biochem (2010) 147(6):809–17. doi: 10.1093/jb/mvq014
Summary
Keywords
galectins, galectin inhibitors, cancer treatments, tumor microenvironment, medical intervention for cancer
Citation
Laderach DJ and Compagno D (2023) Inhibition of galectins in cancer: Biological challenges for their clinical application. Front. Immunol. 13:1104625. doi: 10.3389/fimmu.2022.1104625
Received
21 November 2022
Accepted
16 December 2022
Published
10 January 2023
Volume
13 - 2022
Edited by
Dmitry Aleksandrovich Zinovkin, Gomel State Medical University, Belarus
Reviewed by
Victor L. Thijssen, VU Medical Center, Netherlands; Alexander Timoshenko, Western University, Canada; Eldar Nadyrov, Gomel State Medical University, Belarus; Jale Yuzugulen, Eastern Mediterranean University, Turkey; Charles J. Dimitroff, Florida International University, United States
Updates
Copyright
© 2023 Laderach and Compagno.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Diego José Laderach, diegoladerach@qb.fcen.uba.ar
This article was submitted to Cancer Immunity and Immunotherapy, a section of the journal Frontiers in Immunology
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.