 Petro Booysen1,2*
Petro Booysen1,2* Katalin A. Wilkinson1,2,3
Katalin A. Wilkinson1,2,3 Dylan Sheerin4,5
Dylan Sheerin4,5 Robyn Waters1,2
Robyn Waters1,2 Anna K. Coussens1,4,5
Anna K. Coussens1,4,5 Robert J. Wilkinson1,2,3,6
Robert J. Wilkinson1,2,3,6- 1Centre for Infectious Diseases Research in Africa (CIDRI-Africa), Institute of Infectious Disease and Molecular Medicine, University of Cape Town, Cape Town, South Africa
- 2Department of Medicine, University of Cape Town, Cape Town, South Africa
- 3Tuberculosis Laboratory, The Francis Crick Institute, London, United Kingdom
- 4Infectious Diseases and Immune Defence Division, The Walter & Eliza Hall Institute of Medical Research, Parkville, VIC, Australia
- 5Department of Medical Biology, University of Melbourne, Parkville, VIC, Australia
- 6Department of Infectious Diseases, Imperial College, London, United Kingdom
SARS-CoV-2 and Mycobacterium tuberculosis (Mtb) are major infectious causes of death, with meta-analyses and population-based studies finding increased mortality in co-infected patients simultaneously diagnosed with COVID-19 and tuberculosis (TB). There is a need to understand the immune interaction between SARS-CoV-2 and Mtb which impacts poor outcomes for those co-infected. We performed a PubMed and preprint search using keywords [SARS-CoV-2] AND [tuberculosis] AND [Immune response], including publications after January 2020, excluding reviews or opinions. Abstracts were evaluated by authors for inclusion of data specifically investigating the innate and/or acquired immune responses to SARS-CoV-2 and Mtb in humans and animal models, immunopathological responses in co-infection and both trials and investigations of potential protection against SARS-CoV-2 by Bacille Calmette Guérin (BCG). Of the 248 articles identified, 39 were included. Incidence of co-infection is discussed, considering in areas with a high burden of TB, where reported co-infection is likely underestimated. We evaluated evidence of the clinical association between COVID-19 and TB, discuss differences and similarities in immune responses in humans and in murine studies, and the implications of co-infection. SARS-CoV-2 and Mtb have both been shown to modulate immune responses, particularly of monocytes, macrophages, neutrophils, and T cells. Co-infection may result in impaired immunity to SARS-CoV-2, with an exacerbated inflammatory response, while T cell responses to Mtb may be modulated by SARS-CoV-2. Furthermore, there has been no proven potential COVID-19 clinical benefit of BCG despite numerous large-scale clinical trials.
Introduction
Tuberculosis (TB) and coronavirus disease 2019 (COVID-19) are leading causes of infectious death worldwide (1). As of 14 June 2023, there have been 6,943,390 COVID-19 deaths reported to World Health Organisation (WHO) (2). During the same three-year period, approximately 4.5 million people are estimated to have died of TB. The causative agent of COVID-19, SARS-CoV-2 has undergone various mutations since the start of its pandemic, with several major variants of concern arising and resulting in distinct waves of new infections globally. Since the emergence of the Omicron B.1.1.529 variant, with its attributes of increased transmissibility and reduced risk of mortality, coincident with increasing global vaccine coverage, SARS-CoV-2’s contribution to hospital admissions and overall mortality has been in decline worldwide (3). Notwithstanding COVID-19 remains a highly significant cause of death, TB has again become the leading single infectious cause of death in 2023.
Several recent accounts have shown a detrimental effect of SARS-CoV-2 on TB prevention and care, associating with an increase in reported deaths from TB, a significant decrease in the diagnosis and treatment of TB cases, and diversion of resources allocated for essential TB services and research (4–6). This has now led to a global call to re-establish essential TB services in the wake of widespread disruptions caused by the COVID-19 pandemic.
There are clinical similarities between COVID-19 and TB. Both present predominantly with respiratory signs and symptoms, yet both can also have significant extrapulmonary manifestations (7). Disease severity is greatly influenced by host factors and co-morbidities such as diabetes mellitus, male sex, and HIV-1 co-infection. The purpose of our review was to evaluate clinical evidence of interaction between SARS-CoV-2 and Mycobacterium tuberculosis (Mtb) to determine if co-infection worsens the presentation and outcome of either disease. In addition, we focussed on evidence of potentially adverse immune interaction between the infections that may contribute to worse outcomes for those co-infected.
Method
We performed a PubMed search using keywords [SARS-CoV-2] AND [tuberculosis] AND [Immune response], including publications after January 2020, and MedRxiv search to include preprints. This search delivered 248 publications, which was reduced to 107 abstracts, based on presentation of original clinical, epidemiological, or experimental data, excluding most reviews and viewpoints. Authors evaluated abstracts to include data specifically investigating co-infection prevalence, the impact, acquired and innate immune responses with SARS-CoV-2 and Mtb in humans and animals, immunopathological responses in co-infection and/or trials and investigations of potential BCG protection against SARS-CoV-2. Of the 248 articles identified, 107 abstracts were evaluated and 39 were included (See Supplementary Text 1 for list of articles used in this review).
Results and discussion
Impact of TB on COVID-19 outcomes and vice versa
The COVID-19 pandemic caused global disruptions to health services, with well documented negative impacts on Mtb infected patients and TB-related services, not limited to reduced reporting of active TB cases, difficulty in adequate access to healthcare and health services being overwhelmed by acute COVID-19 cases (1, 4, 5). Co-infection is reported globally with several studies pointing towards increased risk of mortality for co-infected individuals, however studies from high- and low-income countries appear to reflect a marked difference in outcomes (8–12).
Early observational studies of SARS-CoV-2 and Mtb co-infected patients did not suggest TB was a major contributor to increased risk of death in COVID-19 patients, but rather suggested that SARS-CoV-2 infection contributed to a worsening of TB prognosis and/or TB-related death (13, 14). These studies originated from high-income countries with small sample sizes.
Motta et al. (14) reviewed eight cases of co-infected patients in high income countries that died and found SARS-CoV-2 co-infection worsened the prognosis of TB patients and contributed to mortality, with most patients who died acquiring nosocomial SARS-CoV-2 infection. Conversely, an early observational study from China found that patients with asymptomatic latent TB infection (LTBI) or symptomatic active TB were not only potentially more susceptible to SARS-CoV-2 infection, but COVID-19 disease may also progress more rapidly and be more severe in these individuals (15). Although this study was small with only 13 SARS-CoV-2/Mtb co-infected cases, these findings were later supported by large studies from Africa performed in settings of high prevalence of HIV/TB co-infection. These studies surmised that current and previous TB associated with increased COVID-19-related death and were an independent risk factor for mortality (8, 12, 16).
A recent meta-analysis examined the impact of TB on COVID-19 severity and found that overall, COVID-19 patients with TB tended to have an increased risk for more severe disease compared to those without TB (OR = 1.56, 95% CI: 1.13–2.16) (17). As most of the included studies were from Asia, especially from China, the potential generalisability of the findings could be determined through further meta-analyses.
Occurrence of co-infection
Dual presentation was extensively reported early in the COVID-19 pandemic, with TB and COVID-19 co-diagnosis rates ranging between one to four percent (12) although this may be an under ascertainment. A recent evaluation of confirmed co-infected cases reported the prevalence of TB in confirmed COVID-19 patients was 1.1% higher than most reported prevalence in Africa and Asia (18).
Underreporting of SARS-CoV-2 infection, specifically from countries in Africa and other low-income countries, is highly plausible. A study from Zambia identified significant underreporting of COVID-19-related deaths by post-mortem testing of patients (9). They found that most cases died whilst living in the community, where testing facilities were sparse, as opposed to in-hospital deaths. There was evidence of insufficient testing even in hospitals and, despite patient symptoms suggestive of typical COVID-19 disease, SARS-CoV-2 infection was not confirmed (9). Challenges with COVID-19 diagnostic testing and data are not unique to Zambia and have contributed to underreporting in several other African countries. Bradshaw et al. analysed the reported excess deaths data in South Africa during the COVID-19 pandemic and found a near 3-fold increase in excess death from natural causes within timelines corresponding to the peaks of SARS-CoV-2 infection rates, suggesting there was considerable underreporting of SARS-CoV-2 associated deaths (19).
With significant underreporting of SARS-CoV-2 infection in countries with a high TB burden, and decreased reporting of active TB cases in 2020 and 2021 (1), co-infection may also have been far more common than reported. A recent observational study examined the clinical presentation of COVID-19 in an African setting, describing the impact TB and/or HIV-1 infection had on patients admitted with COVID-19 (16). This study included 104 adults, of which 14% had active TB and found clinical features suggestive of either COVID-19 or TB. Chest X-rays in patients with confirmed co-infection were more likely to be classified as non-COVID-19 like, irrespective of HIV status, with a small number having radiological features predominantly suggestive of TB. Although the risk of death due to SARS-CoV-2 infection could not be specifically evaluated, 30/104 (29%) enrolled COVID-19 patients died and 6/15 (40%) of those were co-diagnosed with TB (16).
This study highlighted an important clinical lesson, emphasising that co-infection should be investigated in patients with typical TB presentation in settings with high prevalence of TB (16). This sentiment is echoed by numerous studies reporting similar presentation of signs and symptoms consistent with co-infection across various settings (10, 20) [Summarised in Table 1].
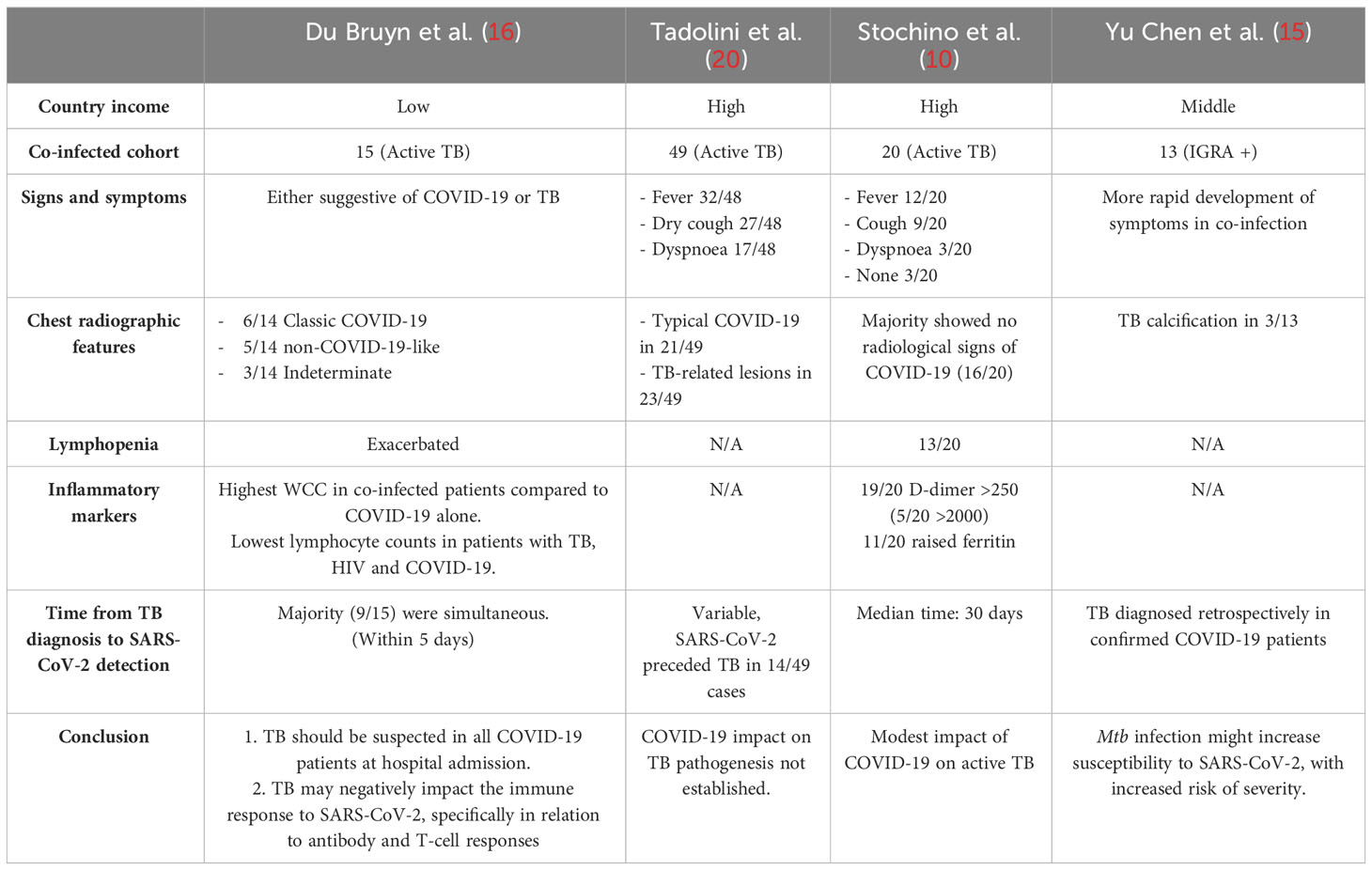
Table 1 Clinical studies of TB and COVID-19 co-diagnosis.
Similarities and differences in the immune response to Mtb and SARS-CoV-2
Both SARS-CoV-2 and Mtb are inhaled as a consequence of infectious aerosols and droplets produced by an infected person. In the case of Mtb, a spectrum of host immunological responses, both innate and acquired, with or without T cell priming either clear the mycobacteria or result in an established Mtb infection. Risk and incidence of infection and disease progression vary greatly depending on population demographics, co-morbidities and environmental factors (21). To establish infection, Mtb must overcome the robust physical barriers of the airway, to reach the lung where alveolar macrophages, neutrophils and dendritic cells are infected, activated and subsequently recruit innate and adaptive lymphocyte populations to aid bacterial containment (22).
Interferon-γ (IFN-γ) activation of alveolar macrophages is the central component of the immune response to Mtb infection. Activation of autophagy results in phagosome maturation and an increase in its acidification which leads to Mtb killing and is a fundamental process Mtb inhibits to maintain its infectious niche (23). Natural killer (NK) cells play a role by recognising and lysing Mtb infected macrophages, increasing IFN-γ production and further secreting cytokines to enhance recruitment of CD8+ T cells and NK T cells. This contributes to the characteristic granuloma formation, consisting of macrophages, neutrophils, Langhans epithelioid giant cells and those formed by fusion of macrophages, surrounded by lymphocytes and a fibrotic cuff (21). Alveolar macrophages use MHC class II molecules to present antigens to CD4+ T cells that are on the outer border of the granuloma, increasing cytokine secretion - notably IFN-γ and tumour necrosis factor (TNF). This will further activate the innate immune response and assist with T cell differentiation and other lymphocyte responses (21, 23). Granuloma morphology and fate are crucial determinants of infection outcome.
SARS-CoV-2 causes an acute infection, with most patients developing symptoms within five to six days after exposure. It predominantly affects the respiratory system; however other organ systems can also be involved. Clinical presentation varies from asymptomatic to severe disease, with symptoms generally being non-specific and includes coughing, fever, headache, and myalgia. SARS-CoV-2 uses angiotensin-converting enzyme 2 (ACE2) receptors to enter target cells. ACE2 can be found in multiple cells, more specifically in lung epithelium, enterocytes, renal and myocardial cells, and oral mucosal epithelium (24).
Whilst ACE2 was first identified as the cell surface receptor for SARS-CoV-2 infection, L-SIGN and DC-SIGN C-type lectins receptors present on various phagocytes and Glucose-regulated protein 78 (GRP78) which translocate to the membrane can also recognise SARS-CoV-2. Binding to receptors is facilitated by proteolytic activation of SARS-CoV-2 S protein by furin-like proteases, transmembrane protease, serine 2 (TMPRSS2) and cathepsin L, whilst viral endocytosis is mediated by clatherin (25–30). Once intracellular, immune cells trigger signalling cascades either by direct endosomal TLR recognition of viral single-stranded (ss)RNA in cells such as plasmacytoid dendritic cells or cytosolic sensing of double-stranded (ds)RNA during viral replication (31). The signalling cascade that results from this recognition triggers transcription factor activation and the production of type I and III IFN and other pro-inflammatory cytokines and chemokines. However, the virus is adept at subverting host IFN responses, leading to lower levels of these cytokines, particularly during severe COVID-19 (32). Type I IFN pathway is important for antiviral responses, and it also plays a key role in TB. Our search, however, did not reveal studies that had investigated this in depth and this important interplay should form the basis for future research.
Alveolar macrophages play a critical role in responding to SARS-CoV-2 in the lungs, but single-cell and spatial transcriptomic studies of BALF and post-mortem lung samples identified depletion of this cell type in the lungs of severe COVID-19 patients as a contributing factor to immunopathology (33). Single cell RNA sequencing (scRNA-seq) has also revealed that profound dysregulation of myeloid cells, specifically increased circulation of various neutrophil subsets, including immature low density neutrophils, immature monocytes or progenitor cells, and myeloid-derived suppressor cells as hallmarks of severe COVID-19, through their contribution to creating an inflammatory cytokines storm (34–37). NK cells exert antiviral activity by clearing infected cells in response to signalling events triggered by SARS-CoV-2 recognition (38).
Clinical markers of COVID-19 deterioration and acute respiratory distress syndrome (ARDS) include elevated lactate dehydrogenase (LDH), C-reactive protein (CRP), interleukin-6 (IL-6), D-dimer, white cell count (WCC), high-sensitivity troponin I, platelet count and renal markers (39). Significant lymphopenia and neutrophilia, creating an elevated neutrophil: lymphocyte ratio is found in critically ill patients (40, 41); a marker not normally associated with viral infection but also associated with severe TB (42). Specific plasma markers: IL-1β, IL-1RA, IL-7, IL-8, IL-9, IL-10, basic FGF, G-CSF, GM-CSF, IFN-γ, CXCL10, CCL2, CCL3, CCL4, PDGF, TNF, and VEGF, show an increased presence in both ICU and non-ICU patients when compared with healthy individuals (43). ICU-admitted patients can also show increased concentrations of G-CSF, CXCL10, CCL2, CCL3, and TNF, hallmarks of the “cytokine storm” associated with COVID-19 disease severity (43).
Having noted an unusual spike in indeterminate Mtb IFN-γ release assay (IGRA) results in their facility, Ward et al. subsequently investigated confirmed SARS-CoV-2-positive hospitalised patients and IFN-γ production. Indeterminate QuantiFERON-TB Gold Plus results in COVID-19 patients, indicative of T cell anergy (positive control PHA-induced IFN-γ production below threshold) seemed to have decreased survival, with higher serum IL-6 and IL-10 levels, however these differences were not statistically significant (44). They also established that this decrease in IFN-γ was not related to lymphopenia or immunosuppressive therapy.
Impact of Mtb and SARS-CoV-2 co-infection on reciprocal immune memory and innate immune responses
Using a rapid, simplified whole blood-based multiparameter assay to quantify and phenotype SARS-CoV-2-specific T cells, Riou et al. examined SARS-CoV-2 antigen-specific CD4+ T cell responses in relation to disease severity in 95 hospitalised COVID-19 patients in South Africa, 38 of whom were HIV and/or Mtb co-infected (45). They found the attributes of SARS-CoV-2-specific CD4+ T cells, and not necessarily the magnitude, were associated with disease severity, characterised by reduced proliferation capacity, and enhanced HLA-DR expression, poor polyfunctional potential and increased proportions of TNF-single positive cells. On the contrary, in non-COVID-19 comparator patients, most SARS-CoV-2-reactive CD4+ T cells were distributed among triple functional cells (IL2+IFN-γ+TNF+) and cells co-producing IFN-γ and TNF.
In the same study, CD4+ T cell depletion resulting from HIV infection, related to suboptimal T cell and humoral immune SARS-CoV-2 responses. In their HIV/TB co-infected COVID-19 cohort consisting of eight patients, only three patients had an antibody response to SARS-CoV-2, and only two had a detectable CD4+ T cell response. Total CD4+ T cell frequency was much higher in SARS-CoV-2 responders compared to non-responders. Furthermore, in the HIV+ cohort, the frequency of total CD4+ T cells was associated with the magnitude of SARS-CoV-2-specific CD4+ T cells. These data suggest that lymphopenia impairs the SARS-CoV-2-specific immune response (45).
When considering the impact of COVID-19 on Mtb-specific responses, it was shown that patients with COVID-19 had a significant 5-fold reduction in the frequency of Mtb-specific CD4+ T cells compared with healthy pre-pandemic LTBI controls, and 2-fold reduction in COVID-19/HIV+ patients compared to HIV+ pre-pandemic controls. As an intact T cell response is essential to control Mtb infection, a decline in Mtb-specific CD4+ T cells could therefore affect the ability of the host to control either existing latent or new Mtb infection (45). Mtb-specific CD4+ T cell activation, previously shown to distinguish active and subclinical TB from those with latent infection, was also found to have a trend towards higher activation in COVID-19/TB patients compared to TB patients without COVID-19, whilst there was no elevation in Mtb-specific CD4+ T cells in COVID-19 patients not co-presenting with TB. Together, this suggests that whilst acute COVID-19 does not immediately reactivate LTBI to subclinical/active disease, it contributes to greater Mtb-specific T cell activation which may exacerbate existing subclinical/active disease.
Looking further into the interaction with LTBI, Rajamanickam et al. (46) examined seropositive, asymptomatic SARS-CoV-2-infected individuals in India and compared immune responses in IGRA-positive (LTBI) and -negative individuals. They showed IGRA-positive individuals had higher levels of humoral, cytokine and acute phase responses compared to IGRA-negative individuals, and thus concluded that LTBI could significantly affect systemic inflammation, as well as cytokine responses and enhanced neutralising antibody capacity in SARS-CoV-2-infected individuals (46). The same investigators also evaluated the effect of SARS-CoV-2 seropositivity on antigen-specific cytokine and chemokine responses in LTBI using QuantiFERON Gold In-tube assay plasma (47). They showed that SARS-CoV-2 seropositive individuals with LTBI had increased cytokine concentrations in both unstimulated and Mtb antigen-stimulated tubes, when compared to those who were SARS-CoV-2 seronegative. These differences were not observed in IGRA-negative individuals who were SARS-CoV-2 seropositive. The authors conclude that both baseline and Mtb antigen-induced cytokine responses are augmented by SARS-CoV-2 sensitisation, suggesting prior SARS-CoV-2 infection augments the immune response to Mtb in LTBI (47).
A highly cited study by Petrone et al. (48) concluded that active TB disease can negatively affect a patient’s ability to generate a SARS-CoV-2-specific immune response, by looking specifically at T cell IFN-γ production in their cohort of co-infected participants. Whole-blood from TB/COVID-19 patients showed the lowest IFN-γ secretion in response to SARS-CoV-2 peptide stimulation compared with COVID-19 patients and to LTBI/COVID-19 patients. They showed that COVID-19 patients with either latent or active TB, still had the ability to respond to Mtb-specific antigens. However only 20% of active TB patients with COVID-19 had a positive response, compared to 64% of COVID-19 patients with LTBI, indicating that active TB depresses the COVID-19-specific host immune response (48), supporting the finding by Riou et al. in COVID-19 with TB/HIV.
A study by Najafi-Fard et al. (49) looked at 119 study participants and compared the plasma immune profile of the 14 TB/COVID-19 co-infected cohort, to the COVID-19 only patients, TB only patients, or 20 healthy controls using a 27-plex multiplex assay. They found that levels of circulating TNF had the strongest association with TB/COVID-19 co-infection compared with COVID-19. They also found that co-infected patients showed a reduced SARS-CoV-2-specific response for several pro-inflammatory cytokines and/or chemokines, anti-inflammatory cytokines, and growth factors and that co-infection negatively affected the Mtb-specific response (49).
Overall, these results (summarised in Table 2), indicate that T cell responses to SARS-CoV-2 and Mtb are both dysregulated by each co-infecting pathogen, resulting in decreased defensive capabilities against both Mtb and HIV-1 in COVID-19 patients, potentially contributing to more unfavourable outcomes and higher mortality in some cases.
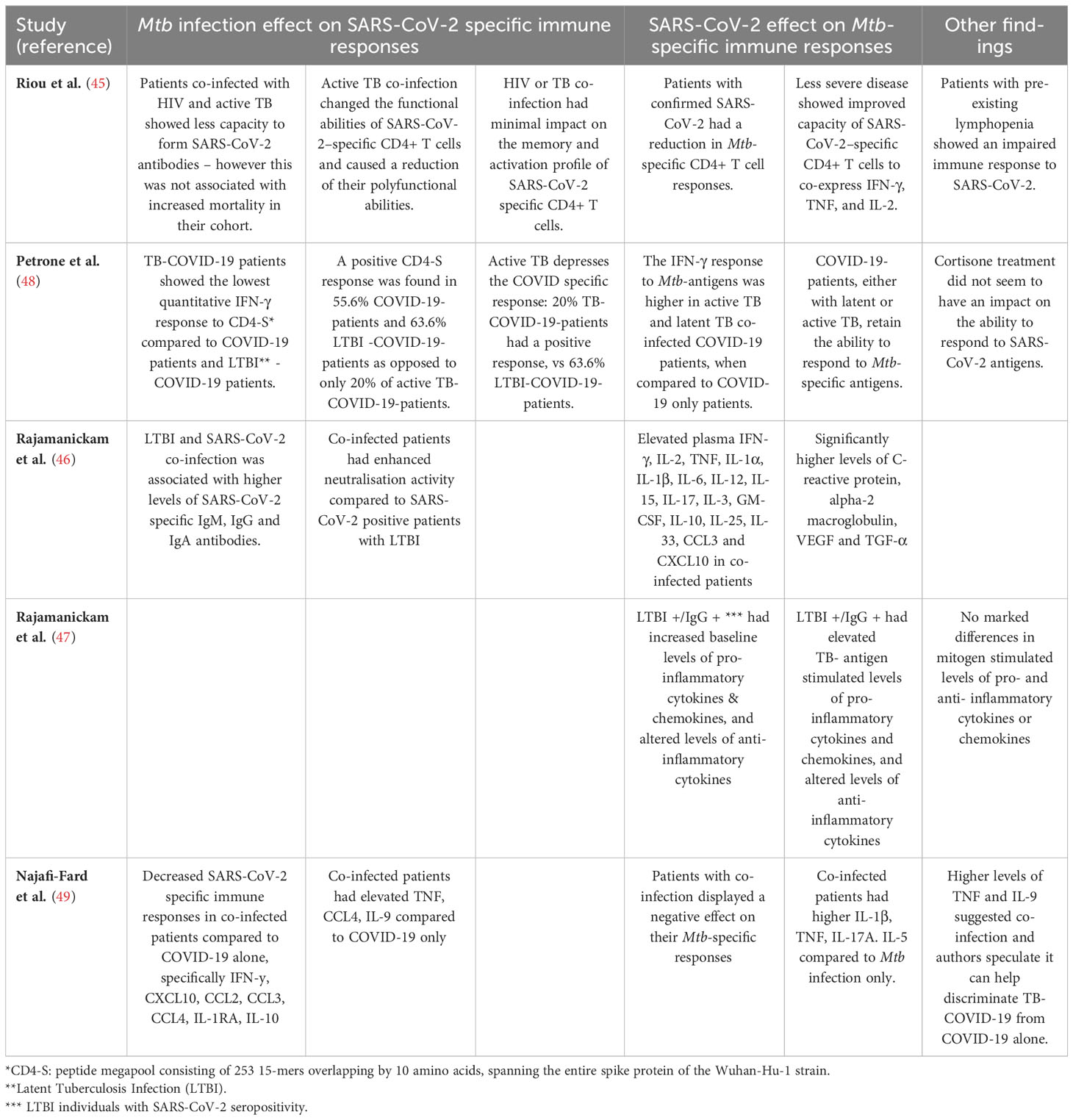
Table 2 Immunological response interactions to Mtb and SARS-CoV-2 in co-infected persons.
Sheerin et al. (50) assessed transcriptional overlap between host immune responses to TB and COVID-19 by profiling scRNA-seq immune cell and severity signatures on bulk RNA-seq data from TB patients across the spectrum of disease, generating “disease risk scores” based on the enrichment of each signature. This analysis indicated that the highest disease risk scores in TB patients were associated with monocyte and neutrophil signatures from severe COVID-19 patients. By summarising gene expression changes at the immunological pathway level for TB, COVID-19 and influenza (as a control for other forms of respiratory infection), it was also shown that IFN-γ and TNF signalling was similarly enriched in COVID-19 and TB patients, but not influenza. Finally, they validated the detrimental interaction between COVID-19 and TB on innate immune cells by comparing the impact of co-culturing human monocyte-derived macrophages (MDM) in the inflammatory milieu from Mtb infected MDM on MDM susceptibility to SARS-CoV-2 infection and inflammatory response. They found co-cultured MDM were more susceptible to SARS-CoV-2 infection and more pro-inflammatory, with increased IFN-α, IFN-γ, TNF, IL-1β and TMPRSS2 expression.
This analysis of blood transcriptional responses from patients and asymptomatic infected persons was followed up by a more thorough exploration of direct co-infection of blood using scRNA-seq; Sheerin et al. (51) infected whole blood from healthy COVID-19 vaccinated donors ex vivo with Mtb, SARS-CoV-2, or both pathogens simultaneously and quantified single cell transcriptome changes, relative to uninfected control samples, across immune cells, 24 and 96 hours post-infection. Distinct neutrophil and monocyte clustering was observed between the three infection conditions. The strongest synergistic co-infection responses were associated with IFN-γ and TNF pathway enrichment 24 hours post-infection. SARS-CoV-2 infection, in the absence of Mtb infection, was associated with enrichment of extrinsic apoptotic signalling, which was negatively regulated by Mtb co-infection, resulting in enhanced cell survival in co-infected verses SARS-CoV-2-only infected cells. SARS-CoV-2 also showed unique enrichment of αβ T cell activation and differentiation not seen in Mtb infection.
TB vaccination with BCG and protection against SARS-CoV-2
The TB vaccine Mycobacterium bovis BCG is known to induce both cellular and humoral immunity in vaccinated individuals (52). The rationale for the potential beneficial effects of BCG in the context of SARS-CoV-2 infection was proposed to include protection via the induction and improved production of pro-inflammatory cytokines through “trained immunity” (53). BCG is thought to provide enhanced protection and/or vaccine responsiveness against a range of pathogens, including Candida albicans, Staphylococcus aureus, Streptococcus pneumoniae, Haemophilus influenzae, vaccinia virus, Bordetella pertussis, and yellow fever virus (54–56); this protection is provided primarily through enhancing monocyte and NK cell production of IL-6, IL-1β, TNF and IFN-γ, and cytokine-induced antigen-specific memory T and B cell activation. BCG enhances innate cytokine production to non-specific pathogens through epigenetic modification and chromatin relaxation at the promoters of these genes, facilitating faster and enhanced cytokine production (57).
There were several suggestions early in the COVID-19 pandemic of epidemiological evidence that prior BCG vaccination correlated with protection against COVID-19 (58), although the evidence became quite mixed as the pandemic progressed (59). Several randomised control trials (RCTs) were set up to test the efficacy of BCG to prevent or decrease the severity of COVID-19 but overall little evidence to support the use of BCG for this purpose has emerged. (A list of all BCG strains used in each of the references is provided in Supplementary Text 2).
A phase III multicentre RCT testing a genetically modified BCG vaccine VPM1002 suggested a prophylactic effect against the development of severe disease in the elderly (60). Another RCT in the elderly reported a reduced rate of new infections after vaccination with standard BCG (61), whereas a larger RCT in the elderly reported no effect on the incidence of disease but noted improved cytokine responses to viral infection (62). An RCT conducted in high-risk adults in India reported that standard BCG reduced the incidence and severity of COVID-19 (63), while a multi-dose BCG phase II/III in diabetic adults claimed an efficacy of 92% for preventing COVID-19 with this regimen (64). Most RCT were conducted in healthcare workers who were among those with the highest risk of exposure to and infection with SARS-CoV-2: an RCT in Brazil reported that re-vaccination with BCG Moscow did not lead to statistically significant reduction in COVID-19 incidence (65), while RCTs conducted in Poland (66), the Netherlands (67) and South Africa (68) also reported no benefit in healthcare workers. A study using samples collected from an Australian RCT investigating the BCG Denmark vaccine in healthcare workers preliminarily reported modulation of cytokines IL‐6, TNF and IL‐10 and CD4+ and CD8+ T cells upon ex vivo stimulation of PBMC, suggesting that this may protect against severe COVID-19 (69), but the same trial recently reported no prevention or reduction in severity of COVID-19 (70). A meta-analysis conducted using these trials revealed no decrease in incidence or hospitalisation from COVID-19 (71).
Experimental models of Mtb and SARS-CoV-2 co-infection and BCG vaccination
Animal studies evaluating immunological responses can contribute to our understanding of host-pathogen interactions and interactions between multiple pathogens within the same host. As summarised in Table 3, Rosas Meija et al. (72) studied mice and the effects of Mtb infection on the immune response to SARS-CoV-2. They used human ACE2 transgenic mice that were chronically infected with Mtb and found these mice to be resistant to secondary infection with SARS-CoV-2. The authors speculated this might be due to the proinflammatory lung environments created by Mtb that are not conducive to SARS-CoV-2 proliferation. Furthermore, SARS-CoV-2 infection did not affect Mtb burden in their experiments.
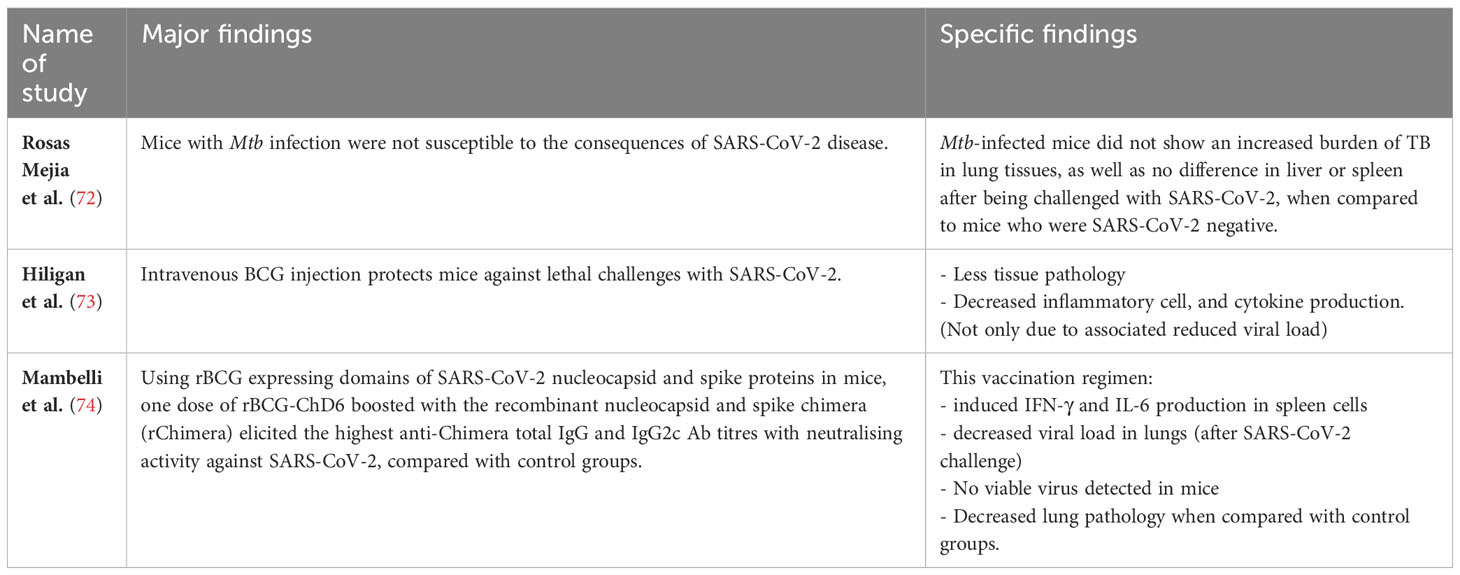
Table 3 Murine studies.
Hilligan et al. (73) also studied human ACE2 transgenic mice to demonstrate that intravenous, but not subcutaneous, inoculation with BCG protected them against lethal challenge with SARS-CoV-2, associated with reduced cytokine production, less tissue pathology and decreased inflammatory cell recruitment, and that was only partially due to the significantly reduced viral load. They speculated that this protection was associated with changes in the composition and function of the pulmonary cellular compartment, likely induced by BCG, providing an experimental model for understanding how a host’s resistance might be promoted by non-specific stimulation of the pulmonary immune response. The protective benefits in this model are in contrast to the lack of clinical efficacy found in RCTs (71). Such discordance may suggest mouse models of Mtb/SARS-CoV-2 co-infection may not reflect the course of human co-infection or could be due to differences in the route of BCG vaccination, as seen in the mouse study discussed above where only the IV route of BCG administration induced protection against a lethal dose of SARS-CoV-2. However, apart from the route of administration (intravenous vs subcutaneous), other factors such as the type of BCG strain or the genetic background of the mouse, might also contribute.
More recently, Mambelli et al. (74) constructed a recombinant BCG (rBCG) that expressed domains of the SARS-CoV-2 nucleocapsid and spike proteins (termed rBCG-ChD6). Using ACE2 transgenic mice, they found that a single dose of rBCG-ChD6 boosted with the recombinant nucleocapsid and spike chimera (rChimera) adjuvanted with alum, resulted in the highest anti-Chimera total IgG and IgG2c Ab titres with neutralising activity against SARS-CoV-2 (specifically the Wuhan strain), compared to their control groups. Furthermore, following SARS-CoV-2 challenge, this vaccination regimen induced IFN-γ and IL-6 production in spleen cells and reduced viral load in the lungs. Moreover, no viable virus was detected in mice immunised with rBCG-ChD6 boosted with rChimera, which was associated with decreased lung pathology when compared with control groups. This study showed the possibility of a prime-boost immunisation system based on an rBCG expressing a chimeric protein derived from SARS-CoV-2.
Mouse models offer numerous useful immunological tools and can be genetically modified. Among mouse strains, the C3HeB/FeJ mouse is the only strain reproducing the pathophysiology of TB, with comparable granuloma encapsulation (75). Although not discussed here, other models, like hamsters and ferrets, and Non-Human Primates (NHP) are also incredibly useful when investigating human pathologies.
Conclusion and consequences
Diversion of healthcare services during the COVID-19 pandemic undoubtedly had an adverse effect on the ongoing TB epidemic. Acute COVID-19 and TB can be coincident and the occurrence of such co-infections in areas of high TB prevalence may have been underestimated. Previous or current TB is a risk factor for death from SARS-CoV-2. Ex vivo studies of blood cells in acutely infected humans suggest the T cell response to Mtb may be modulated by SARS-CoV-2: conversely coincident TB may impair immune responses to SARS-CoV-2 and exacerbate inflammatory responses through enhanced innate and adaptive immune activation. Despite animal studies and epidemiological evidence pointing to potential protection against SARS-CoV-2 by BCG, efficacy has not been borne out in several large-scale clinical evaluations. Further studies of the long-term consequences of SARS-CoV-2 infection on the immune response in, and outcome of latent TB are warranted.
Author contributions
PB: Conceptualisation, Data curation, Investigation, Methodology, Writing – original draft, Writing – review & editing. KW: Writing – original draft, Writing – review & editing, Investigation. DS: Writing – original draft, Writing – review & editing, Investigation. RW: Writing – review & editing, Investigation. AC: Writing – review & editing, Investigation. RJW: Conceptualisation, Funding acquisition, Investigation, Methodology, Supervision, Writing – review & editing.
Funding
This work was supported in part by Wellcome (203135, 222574). RW is supported by the Francis Crick Institute which receives funding from Wellcome (CC2112), Cancer Research UK (CC2112) and the Medical Research Council (CC2112). AC and DS are supported by the NHMRC (GNT2020750) and the Drakensburg Trust.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2023.1254206/full#supplementary-material
References
1. WHO. Global tuberculosis report 2022 (2022). Available at: https://wwwwhoint/teams/global-tuberculosis-programme/tb-reports/global-tuberculosis-report-2022.
2. WHO. Available at: https://covid19.who.int/.
3. Hyams C, Challen R, Marlow R, Nguyen J, Begier E, Southern J, et al. Severity of Omicron (B.1.1.529) and Delta (B.1.617.2) SARS-CoV-2 infection among hospitalised adults: a prospective cohort study in Bristol, United Kingdom. Lancet Regional Health - Europe (2023) 25:100556. doi: 10.1016/j.lanepe.2022.100556
4. Pai M, Kasaeva T, Swaminathan S. Covid-19’s devastating effect on tuberculosis care — A path to recovery. N Engl J Med (2022) 386(16):1490–3. doi: 10.1056/NEJMp2118145
5. Benade M, Long L, Meyer-Rath G, Miot J, Evans D, Tucker J-M, et al. Reduction in initiations of drug-sensitive tuberculosis treatment in South Africa during the COVID-19 pandemic: Analysis of retrospective, facility-level data. PloS Global Public Health (2022) 2(10):e0000559. doi: 10.1371/journal.pgph.0000559
6. Sanduzzi Zamparelli S, Mormile M, Sanduzzi Zamparelli A, Guarino A, Parrella R, Bocchino M. Clinical impact of COVID-19 on tuberculosis. Infez Med (2022) 30(4):495–500. doi: 10.53854/liim-3004-3
7. Gupta A, Madhavan MV, Sehgal K, Nair N, Mahajan S, Sehrawat TS, et al. Extrapulmonary manifestations of COVID-19. Nat Med (2020) 26(7):1017–32. doi: 10.1038/s41591-020-0968-3
8. Jassat W, Cohen C, Tempia S, Masha M, Goldstein S, Kufa T, et al. Risk factors for COVID-19-related in-hospital mortality in a high HIV and tuberculosis prevalence setting in South Africa: a cohort study. Lancet HIV (2021) 8(9):e554–e67. doi: 10.1016/S2352-3018(21)00151-X
9. Mwananyanda L, Gill CJ, MacLeod W, Kwenda G, Pieciak R, Mupila Z, et al. Covid-19 deaths in Africa: prospective systematic postmortem surveillance study. BMJ (2021) 372:n334. doi: 10.1136/bmj.n334
10. Stochino C, Villa S, Zucchi P, Parravicini P, Gori A, Raviglione MC. Clinical characteristics of COVID-19 and active tuberculosis co-infection in an Italian reference hospital. Eur Respir J (2020) 56(1). doi: 10.1183/13993003.01708-2020
11. van der Zalm MM, Lishman J, Verhagen LM, Redfern A, Smit L, Barday M, et al. Clinical experience with severe acute respiratory syndrome coronavirus 2-related illness in children: hospital experience in Cape Town, South Africa. Clin Infect Dis (2021) 72(12):e938–e44. doi: 10.1093/cid/ciaa1666
12. Western Cape Department of Health in collaboration with the National Institute for Communicable Diseases SA. Risk factors for coronavirus disease 2019 (COVID-19) death in a population cohort study from the western cape province, South Africa. Clin Infect Dis (2021) 73(7):e2005–e15. doi: 10.1093/cid/ciaa1198
13. Collins LF, Moran CA, Oliver NT, Moanna A, Lahiri CD, Colasanti JA, et al. Clinical characteristics, comorbidities and outcomes among persons with HIV hospitalized with coronavirus disease 2019 in Atlanta, Georgia. AIDS (2020) 34(12):1789–94. doi: 10.1097/QAD.0000000000002632
14. Motta I, Centis R, D’Ambrosio L, Garcia-Garcia JM, Goletti D, Gualano G, et al. Tuberculosis, COVID-19 and migrants: Preliminary analysis of deaths occurring in 69 patients from two cohorts. Pulmonology (2020) 26(4):233–40. doi: 10.1016/j.pulmoe.2020.05.002
15. Chen Y, Wang Y, Fleming J, Yu Y, Gu Y, Liu C, et al. Active or latent tuberculosis increases susceptibility to COVID-19 and disease severity. medRxiv (2020) 2020:3.
16. du Bruyn E, Stek C, Daroowala R, Said-Hartley Q, Hsiao M, Schafer G, et al. Effects of tuberculosis and/or HIV-1 infection on COVID-19 presentation and immune response in Africa. Nat Commun (2023) 14(1):188. doi: 10.1038/s41467-022-35689-1
17. Wang Y, Feng R, Xu J, Hou H, Feng H, Yang H. An updated meta-analysis on the association between tuberculosis and COVID-19 severity and mortality. J Med Virol (2021) 93(10):5682–6. doi: 10.1002/jmv.27119
18. Daneshvar P, Hajikhani B, Sameni F, Noorisepehr N, Zare F, Bostanshirin N, et al. COVID-19 and tuberculosis coinfection: An overview of case reports/case series and meta-analysis of prevalence studies. Heliyon (2023) 9(2):e13637. doi: 10.1016/j.heliyon.2023.e13637
19. Bradshaw D, Dorrington RE, Laubscher R, Moultrie TA, Groenewald P. Tracking mortality in near to real time provides essential information about the impact of the COVID-19 pandemic in South Africa in 2020. S Afr Med J (2021) 111(8):732–40. doi: 10.7196/SAMJ.2021.v111i8.15809
20. Tadolini M, Codecasa LR, Garcia-Garcia JM, Blanc FX, Borisov S, Alffenaar JW, et al. Active tuberculosis, sequelae and COVID-19 co-infection: first cohort of 49 cases. Eur Respir J (2020) 56(1). doi: 10.1183/13993003.02328-2020
21. Walker NF, Meintjes G, Wilkinson RJ. HIV-1 and the immune response to TB. Future Virol (2013) 8(1):57–80. doi: 10.2217/fvl.12.123
22. Lerner TR, Borel S, Gutierrez MG. The innate immune response in human tuberculosis. Cell Microbiol (2015) 17(9):1277–85. doi: 10.1111/cmi.12480
23. O’Garra A, Redford PS, McNab FW, Bloom CI, Wilkinson RJ, Berry MP. The immune response in tuberculosis. Annu Rev Immunol (2013) 31:475–527. doi: 10.1146/annurev-immunol-032712-095939
24. Yuki K, Fujiogi M, Koutsogiannaki S. COVID-19 pathophysiology: A review. Clin Immunol (2020) 215:108427. doi: 10.1016/j.clim.2020.108427
25. Amraei R, Yin W, Napoleon MA, Suder EL, Berrigan J, Zhao Q, et al. CD209L/L-SIGN and CD209/DC-SIGN act as receptors for SARS-coV-2. ACS Cent Sci (2021) 7(7):1156–65. doi: 10.1021/acscentsci.0c01537
26. Bayati A, Kumar R, Francis V, McPherson PS. SARS-CoV-2 infects cells after viral entry via clathrin-mediated endocytosis. J Biol Chem (2021) 296:100306. doi: 10.1016/j.jbc.2021.100306
27. Hoffmann M, Kleine-Weber H, Schroeder S, Kruger N, Herrler T, Erichsen S, et al. SARS-coV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell (2020) 181(2):271–80 e8. doi: 10.1016/j.cell.2020.02.052
28. Zhao MM, Yang WL, Yang FY, Zhang L, Huang WJ, Hou W, et al. Cathepsin L plays a key role in SARS-CoV-2 infection in humans and humanized mice and is a promising target for new drug development. Signal Transduct Target Ther (2021) 6(1):134. doi: 10.1038/s41392-021-00558-8
29. Ibrahim IM, Abdelmalek DH, Elshahat ME, Elfiky AA. COVID-19 spike-host cell receptor GRP78 binding site prediction. J Infect (2020) 80(5):554–62. doi: 10.1016/j.jinf.2020.02.026
30. Shin W-J, Ha DP, Machida K, Lee AS. The stress-inducible ER chaperone GRP78/BiP is upregulated during SARS-CoV-2 infection and acts as a pro-viral protein. Nat Commun (2022) 13(1):6551. doi: 10.1038/s41467-022-34065-3
31. Madden EA, Diamond MS. Host cell-intrinsic innate immune recognition of SARS-CoV-2. Curr Opin Virol (2022) 52:30–8. doi: 10.1016/j.coviro.2021.11.002
32. Hadjadj J, Yatim N, Barnabei L, Corneau A, Boussier J, Smith N, et al. Impaired type I interferon activity and inflammatory responses in severe COVID-19 patients. Science (2020) 369(6504):718–24. doi: 10.1126/science.abc6027
33. Delorey TM, Ziegler CGK, Heimberg G, Normand R, Yang Y, Segerstolpe Å, et al. COVID-19 tissue atlases reveal SARS-CoV-2 pathology and cellular targets. Nature (2021) 595(7865):107–13. doi: 10.1038/s41586-021-03570-8
34. Liao M, Liu Y, Yuan J, Wen Y, Xu G, Zhao J, et al. Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nat Med (2020) 26(6):842–4. doi: 10.1038/s41591-020-0901-9
35. Schulte-Schrepping J, Reusch N, Paclik D, Baßler K, Schlickeiser S, Zhang B, et al. Severe COVID-19 is marked by a dysregulated myeloid cell compartment. Cell (2020) 182(6):1419–40.e23. doi: 10.1016/j.cell.2020.08.001
36. Carissimo G, Xu W, Kwok I, Abdad MY, Chan Y-H, Fong S-W, et al. Whole blood immunophenotyping uncovers immature neutrophil-to-VD2 T-cell ratio as an early marker for severe COVID-19. Nat Commun (2020) 11(1):5243. doi: 10.1038/s41467-020-19080-6
37. Wu D, Yang XO. TH17 responses in cytokine storm of COVID-19: An emerging target of JAK2 inhibitor Fedratinib. J Microbiol Immunol Infect (2020) 53(3):368–70. doi: 10.1016/j.jmii.2020.03.005
38. Witkowski M, Tizian C, Ferreira-Gomes M, Niemeyer D, Jones TC, Heinrich F, et al. Untimely TGFβ responses in COVID-19 limit antiviral functions of NK cells. Nature (2021) 600(7888):295–301. doi: 10.1038/s41586-021-04142-6
39. Wu C, Chen X, Cai Y, Ja X, Zhou X, Xu S, et al. Risk factors associated with acute respiratory distress syndrome and death in patients with coronavirus disease 2019 pneumonia in Wuhan, China. JAMA Internal Med (2020) 180(7):934–43. doi: 10.1001/jamainternmed.2020.0994
40. Tan L, Wang Q, Zhang D, Ding J, Huang Q, Tang Y-Q, et al. Lymphopenia predicts disease severity of COVID-19: a descriptive and predictive study. Signal Transduct Target Ther (2020) 5(1):33. doi: 10.1038/s41392-020-0148-4
41. Liu J, Liu Y, Xiang P, Pu L, Xiong H, Li C, et al. Neutrophil-to-lymphocyte ratio predicts critical illness patients with 2019 coronavirus disease in the early stage. J Transl Med (2020) 18(1):206. doi: 10.1186/s12967-020-02374-0
42. Panteleev AV, Nikitina IY, Burmistrova IA, Kosmiadi GA, Radaeva TV, Amansahedov RB, et al. Severe tuberculosis in humans correlates best with neutrophil abundance and lymphocyte deficiency and does not correlate with antigen-specific CD4 T-cell response. Front Immunol (2017) 8. doi: 10.3389/fimmu.2017.00963
43. Huang C, Wang Y, Li X, Ren L, Zhao J, Hu Y, et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet (2020) 395(10223):497–506. doi: 10.1016/S0140-6736(20)30183-5
44. Ward JD, Cornaby C, Schmitz JL. Indeterminate quantiFERON gold plus results reveal deficient interferon gamma responses in severely ill COVID-19 patients. J Clin Microbiol (2021) 59(10):e00811–21. doi: 10.1128/JCM.00811-21
45. Riou C, du Bruyn E, Stek C, Daroowala R, Goliath RT, Abrahams F, et al. Relationship of SARS-CoV-2-specific CD4 response to COVID-19 severity and impact of HIV-1 and tuberculosis coinfection. J Clin Invest (2021) 131(12). doi: 10.1172/JCI149125
46. Rajamanickam A, Kumar NP, Padmapriyadarsini C, Nancy A, Selvaraj N, Karunanithi K, et al. Latent tuberculosis co-infection is associated with heightened levels of humoral, cytokine and acute phase responses in seropositive SARS-CoV-2 infection. J Infect (2021) 83(3):339–46. doi: 10.1016/j.jinf.2021.07.029
47. Rajamanickam A, Pavan Kumar N, Chandrasekaran P, Nancy A, Bhavani PK, Selvaraj N, et al. Effect of SARS-CoV-2 seropositivity on antigen - specific cytokine and chemokine responses in latent tuberculosis. Cytokine (2022) 150:155785. doi: 10.1016/j.cyto.2021.155785
48. Petrone L, Petruccioli E, Vanini V, Cuzzi G, Gualano G, Vittozzi P, et al. Coinfection of tuberculosis and COVID-19 limits the ability to in vitro respond to SARS-CoV-2. Int J Infect Dis (2021) 113 Suppl 1:S82–S7. doi: 10.1016/j.ijid.2021.02.090
49. Najafi-Fard S, Aiello A, Navarra A, Cuzzi G, Vanini V, Migliori GB, et al. Characterization of the immune impairment of patients with tuberculosis and COVID-19 coinfection. Int J Infect Dis (2023) 130(Suppl 1):S34–S42. doi: 10.1016/j.ijid.2023.03.021
50. Sheerin D, Abhimanyu, Peton N, Vo W, Allison CC, Wang X, et al. Immunopathogenic overlap between COVID-19 and tuberculosis identified from transcriptomic meta-analysis and human macrophage infection. iScience (2022) 25(6):104464. doi: 10.1016/j.isci.2022.104464
51. Sheerin D, Phan TK, Eriksson EM, Consortium CP, Coussens AK. Distinct and overlapping immunological responses to SARS-CoV-2 and Mycobacterium tuberculosis identified by single-cell RNA-seq of co-infected whole blood. medRxiv (2023), 23290499. doi: 10.1101/2023.05.24.23290499
52. Dennehy M, Williamson A-L. Factors influencing the immune response to foreign antigen expressed in recombinant BCG vaccines. Vaccine (2005) 23(10):1209–24. doi: 10.1016/j.vaccine.2004.08.039
53. O’Neill LAJ, Netea MG. BCG-induced trained immunity: can it offer protection against COVID-19? Nat Rev Immunol (2020) 20(6):335–7. doi: 10.1038/s41577-020-0337-y
54. Moorlag SJCFM, Arts RJW, van Crevel R, Netea MG. Non-specific effects of BCG vaccine on viral infections. Clin Microbiol Infect (2019) 25(12):1473–8. doi: 10.1016/j.cmi.2019.04.020
55. Covián C, Fernández-Fierro A, Retamal-Díaz A, Díaz FE, Vasquez AE, Lay MK, et al. BCG-induced cross-protection and development of trained immunity: implication for vaccine design. Front Immunol (2019) 10. doi: 10.3389/fimmu.2019.02806
56. Arts RJW, Moorlag SJCFM, Novakovic B, Li Y, Wang S-Y, Oosting M, et al. BCG Vaccination Protects against Experimental Viral Infection in Humans through the Induction of Cytokines Associated with Trained Immunity. Cell Host Microbe (2018) 23(1):89–100.e5. doi: 10.1016/j.chom.2017.12.010
57. Mantovani A, Netea MG. Trained innate immunity, epigenetics, and covid-19. N Engl J Med (2020) 383(11):1078–80. doi: 10.1056/NEJMcibr2011679
58. Escobar LE, Molina-Cruz A, Barillas-Mury C. BCG vaccine protection from severe coronavirus disease 2019 (COVID-19). Proc Natl Acad Sci (2020) 117(30):17720–6. doi: 10.1073/pnas.2008410117
59. Fu W, Ho P-C, Liu C-L, Tzeng K-T, Nayeem N, Moore JS, et al. Reconcile the debate over protective effects of BCG vaccine against COVID-19. Sci Rep (2021) 11(1):8356. doi: 10.1038/s41598-021-87731-9
60. Blossey AM, Brückner S, May M, Parzmair GP, Sharma H, Shaligram U, et al. VPM1002 as prophylaxis against severe respiratory tract infections including coronavirus disease 2019 in the elderly: A phase 3 randomized, double-blind, placebo-controlled, multicenter clinical study. Clin Infect Dis (2022) 76(7):1304–10. doi: 10.1093/cid/ciac881
61. Tsilika M, Taks E, Dolianitis K, Kotsaki A, Leventogiannis K, Damoulari C, et al. ACTIVATE-2: A double-blind randomized trial of BCG vaccination against COVID-19 in individuals at risk. Front Immunol (2022) 13. doi: 10.3389/fimmu.2022.873067
62. Moorlag SJCFM, Taks E, ten Doesschate T, van der Vaart TW, Janssen AB, Müller L, et al. Efficacy of BCG vaccination against respiratory tract infections in older adults during the coronavirus disease 2019 pandemic. Clin Infect Dis (2022) 75(1):e938–e46. doi: 10.1093/cid/ciac182
63. Sinha S, Ajayababu A, Thukral H, Gupta S, Guha SK, Basu A, et al. Efficacy of bacillus calmette–guérin (BCG) vaccination in reducing the incidence and severity of COVID-19 in high-risk population (BRIC): a phase III, multi-centre, quadruple-blind randomised control trial. Infect Dis Ther (2022) 11(6):2205–17. doi: 10.1007/s40121-022-00703-y
64. Faustman DL, Lee A, Hostetter ER, Aristarkhova A, Ng NC, Shpilsky GF, et al. Multiple BCG vaccinations for the prevention of COVID-19 and other infectious diseases in type 1 diabetes. Cell Rep Med (2022) 3(9):100728. doi: 10.1016/j.xcrm.2022.100728
65. dos Anjos LRB, da Costa AC, Cardoso A, Guimarães RA, Rodrigues RL, Ribeiro KM, et al. Efficacy and safety of BCG revaccination with M. bovis BCG moscow to prevent COVID-19 infection in health care workers: A randomized phase II clinical trial. Front Immunol (2022) 13. doi: 10.3389/fimmu.2022.841868
66. Czajka H, Zapolnik P, Krzych Ł, Kmiecik W, Stopyra L, Nowakowska A, et al. A Multi-Center, Randomised, Double-Blind, Placebo-Controlled Phase III Clinical Trial Evaluating the Impact of BCG Re-Vaccination on the Incidence and Severity of SARS-CoV-2 Infections among Symptomatic Healthcare Professionals during the COVID-19 Pandemic in Poland-First Results. Vaccines (2022) 10(2):314. doi: 10.3390/vaccines10020314
67. ten Doesschate T, van der Vaart TW, Debisarun PA, Taks E, Moorlag SJCFM, Paternotte N, et al. Bacillus Calmette-Guérin vaccine to reduce healthcare worker absenteeism in COVID-19 pandemic, a randomized controlled trial. Clin Microbiol Infect (2022) 28(9):1278–85. doi: 10.1016/j.cmi.2022.04.009
68. Upton CM, van Wijk RC, Mockeliunas L, Simonsson USH, McHarry K, van den Hoogen G, et al. Safety and efficacy of BCG re-vaccination in relation to COVID-19 morbidity in healthcare workers: A double-blind, randomised, controlled, phase 3 trial. eClinicalMedicine (2022) 48:101414. doi: 10.1016/j.eclinm.2022.101414
69. Messina NL, Germano S, McElroy R, Rudraraju R, Bonnici R, Pittet LF, et al. Off-target effects of bacillus Calmette-Guérin vaccination on immune responses to SARS-CoV-2: implications for protection against severe COVID-19. Clin Transl Immunol (2022) 11(4):e1387. doi: 10.1002/cti2.1387
70. Pittet LF, Messina NL, Orsini F, Moore CL, Abruzzo V, Barry S, et al. Randomized trial of BCG vaccine to protect against covid-19 in health care workers. N Engl J Med (2023) 388(17):1582–96. doi: 10.1056/NEJMoa2212616
71. Wen J, Liu Q, Tang D, He J-Q. Efficacy of BCG vaccination against COVID-19: systematic review and meta-analysis of randomized controlled trials. J Clin Med (2023) 12(3):1154. doi: 10.3390/jcm12031154
72. Rosas Mejia O, Gloag ES, Li J, Ruane-Foster M, Claeys TA, Farkas D, et al. Mice infected with Mycobacterium tuberculosis are resistant to acute disease caused by secondary infection with SARS-CoV-2. PloS Pathog (2022) 18(3):e1010093. doi: 10.1371/journal.ppat.1010093
73. Hilligan KL, Namasivayam S, Clancy CS, O’Mard D, Oland SD, Robertson SJ, et al. Intravenous administration of BCG protects mice against lethal SARS-CoV-2 challenge. J Exp Med (2022) 219(2). doi: 10.1084/jem.20211862
74. Mambelli F, Marinho FV, Andrade JM, de Araujo A, Abuna RPF, Fabri VMR, et al. Recombinant bacillus calmette-guérin expressing SARS-coV-2 chimeric protein protects K18-hACE2 mice against viral challenge. J Immunol (2023) 210(12):1925–37. doi: 10.4049/jimmunol.2200731
Keywords: COVID-19, latent TB, LTBI, Bacille Calmette Guérin, co-infection, immune response, transcriptomics, T cells
Citation: Booysen P, Wilkinson KA, Sheerin D, Waters R, Coussens AK and Wilkinson RJ (2023) Immune interaction between SARS-CoV-2 and Mycobacterium tuberculosis. Front. Immunol. 14:1254206. doi: 10.3389/fimmu.2023.1254206
Received: 06 July 2023; Accepted: 12 September 2023;
Published: 27 September 2023.
Edited by:
Alex Sigal, Africa Health Research Institute (AHRI), South AfricaReviewed by:
Jordi B. Torrelles, Texas Biomedical Research Institute, United StatesAude Remot, Institut National de Recherche pour l’Agriculture, l’Alimentation et l’Environnement (INRAE), France
Copyright © 2023 Booysen, Wilkinson, Sheerin, Waters, Coussens and Wilkinson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Petro Booysen, cGV0cm9AYm9veXNlbi5jb20=