 Laura Edith Martínez-Gómez1
Laura Edith Martínez-Gómez1 Carlos Martinez-Armenta1
Carlos Martinez-Armenta1 Teresa Tusie-Luna2,3
Teresa Tusie-Luna2,3 Paola Vázquez-Cárdenas4
Paola Vázquez-Cárdenas4 Rosa P. Vidal-Vázquez4
Rosa P. Vidal-Vázquez4 Juan P. Ramírez-Hinojosa4
Juan P. Ramírez-Hinojosa4 Diana Gómez-Martín5
Diana Gómez-Martín5 Gilberto Vargas-Alarcón6
Gilberto Vargas-Alarcón6 Rosalinda Posadas-Sánchez6
Rosalinda Posadas-Sánchez6 José Manuel Fragoso6
José Manuel Fragoso6 Aurora de la Peña7
Aurora de la Peña7 José Manuel Rodríguez-Pérez6
José Manuel Rodríguez-Pérez6 Mónica M. Mata-Miranda8
Mónica M. Mata-Miranda8 Gustavo J. Vázquez-Zapién8
Gustavo J. Vázquez-Zapién8 Adriana Martínez-Cuazitl8
Adriana Martínez-Cuazitl8 Felipe de J. Martínez-Ruiz9Dulce M. Zayago-Angeles9
Felipe de J. Martínez-Ruiz9Dulce M. Zayago-Angeles9 Luis Ramos-Tavera5Alberto Méndez-Aguilera1
Luis Ramos-Tavera5Alberto Méndez-Aguilera1 María del C. Camacho-Rea5María L. Ordoñez-Sánchez2Yayoi Segura-Kato2
María del C. Camacho-Rea5María L. Ordoñez-Sánchez2Yayoi Segura-Kato2 Carlos Suarez-Ahedo1Jessel Olea-Torres1
Carlos Suarez-Ahedo1Jessel Olea-Torres1 Brígida Herrera-López1
Brígida Herrera-López1 Carlos Pineda1
Carlos Pineda1 Gabriela A. Martínez-Nava1*
Gabriela A. Martínez-Nava1* Alberto López-Reyes1*
Alberto López-Reyes1*- 1Laboratorio de Gerociencias, Dirección General, Departamento de Reconstrucción Articular, Laboratorio Facilitador, Instituto Nacional de Rehabilitación Luis Guillermo Ibarra Ibarra, Secretaría de Salud, Mexico City, Mexico
- 2Unidad de Biología Molecular y Medicina Genómica, Instituto Nacional de Ciencias Médicas y Nutrición, Salvador, Zubirán, Mexico City, Mexico
- 3Instituto de Investigaciones Biomédicas Universidad Nacional Autónoma de México, Mexico City, Mexico
- 4Centro de Innovación Médica Aplicada, Hospital General Dr. Manuel Gea González, Mexico City, Mexico
- 5Department of Immunology and Rheumatology, Departamento de Inmunogenética, Departamento de Nutrición Animal, Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán, Secretaría de Salud, Mexico City, Mexico
- 6Departamento de Biología Molecular y Endocrinología, Instituto Nacional de Cardiología Ignacio Chávez, Mexico City, Mexico
- 7Departamento de Farmacología, Facultad de Medicina, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 8Laboratorio de Biología Celular y Tisular, Laboratorio de Embriología, Escuela Médico Militar, Universidad del Ejército y Fuerza Aérea, Mexico City, Mexico
- 9Nuevo Hospital General Delegación Regional Sur de la Ciudad de México Instituto de Seguridad y Servicios Sociales de los Trabajadores del Estado (ISSSTE), Mexico City, Mexico
Introduction: Serine proteases play a critical role during SARS-CoV-2 infection. Therefore, polymorphisms of transmembrane protease serine 2 (TMPRSS2) and serpine family E member 1 (SERPINE1) could help to elucidate the contribution of variability to COVID-19 outcomes.
Methods: To evaluate the genetic variants of the genes previously associated with COVID-19 outcomes, we performed a cross-sectional study in which 1536 SARS-CoV-2-positive participants were enrolled. TMPRSS2 (rs2070788, rs75603675, rs12329760) and SERPINE1 (rs2227631, rs2227667, rs2070682, rs2227692) were genotyped using the Open Array Platform. The association of polymorphisms with disease outcomes was determined by logistic regression analysis adjusted for covariates (age, sex, hypertension, type 2 diabetes, and obesity).
Results: According to our codominant model, the GA genotype of rs2227667 (OR=0.55; 95% CI = 0.36-0.84; p=0.006) and the AG genotype of rs2227667 (OR=0.59; 95% CI = 0.38-0.91; p=0.02) of SERPINE1 played a protective role against disease. However, the rs2227692 T allele and TT genotype SERPINE1 (OR=1.45; 95% CI = 1.11-1.91; p=0.006; OR=2.08; 95% CI = 1.22-3.57; p=0.007; respectively) were associated with a decreased risk of death. Similarly, the rs75603675 AA genotype TMPRSS2 had an OR of 1.97 (95% CI = 1.07-3.6; p=0.03) for deceased patients. Finally, the rs2227692 T allele SERPINE1 was associated with increased D-dimer levels (OR=1.24; 95% CI = 1.03-1.48; p=0.02).
Discussion: Our data suggest that the rs75603675 TMPRSS2 and rs2227692 SERPINE1 polymorphisms are associated with a poor outcome. Additionally, rs2227692 SERPINE1 could participate in hypercoagulable conditions in critical COVID-19 patients, and this genetic variant could contribute to the identification of new pharmacological targets and treatment strategies to block the inhibition of TMPRSS2 entry into SARS-CoV-2.
1 Introduction
Serine protease cascades control coagulation, and innate immune responses are increased during severe SARS-CoV-2 infection (1, 2). Different enzymes with serine protease activity, such as transmembrane protease serine 2 (TMPRSS2), have been described as critical determinants of spike (S) protein shedding in the SARS-CoV-2 virus and therefore trigger the infection process via the endosomal route or by membrane fusion with the host cell (3, 4). TMPRSS2 expression is crucial for the spread and pathogenesis of SARS-CoV-2. The spike protein of SARS-CoV-2 can be cleaved by circulating thrombotic proteases, thrombin and Factor X, as well as the thrombotic protease plasmin, which contributes to severe COVID-19 complications (1, 5).
The TMPRSS2 gene is located on chromosome 21q22.3 and comprises 14 exons and 13 introns; its transcriptional activity is controlled by androgen receptors, which play roles in carcinogenesis (6). The enzymatic activity of TMPRSS2 in viral infection and its inhibition have been proposed as novel mechanisms to reduce mortality associated with SARS-CoV-2 infection (7, 8). In this sense, a nonpharmacological strategy to regulate the activity of TMPRSS2 by its endogenous inhibitor plasminogen activator inhibitor-1 (PAI-1) for influenza and coronavirus infections has recently been reported (9).
The serpine family E member 1 (SERPINE1) gene encodes plasminogen activator inhibitor-1 (PAI-1), whose principal physiological activity is to inhibit urokinase plasminogen activator (uPA) and tissue-type plasminogen activator (tPA) to further regulate the breakdown of blood clots. Therefore, the modulation of uPA and tPA could control the production of plasmin, D-dimer, and ferritin, which are associated with coagulopathies and adverse outcomes in patients with COVID-19 (10). The presence of D-dimer guarantees that coagulation is amplified and that fibrin deposits are ultimately stabilized (11, 12). SERPINE1 and other serine protease inhibitors have the potential to inhibit TMPRSS2 (5).
The SERPINE1 gene is located on chromosome 7q22.1, and some polymorphisms in the SERPINE1 promoter region have been associated with severe COVID-19 (13, 14) as well as with a suboptimal fibrinolytic response (15, 16).
Genetic polymorphisms in these genes could modulate genetic predisposition to infection and virus clearance in the host (17). The rs12329760 polymorphism is present in the exonic splicing enhancer site srp40 and could increase the chance of expression due to potential disruption of the exonic splicing enhancer site. rs2070788 and rs75603675 were reported to have higher levels of TMPRSS2 expression and structural changes (13, 17). rs2227631 is located in the promoter and is implicated in PAI regulation. rs5557667 is located in the intronic region between exons 3-4, rs2070682 is located in introns 5-6, and rs222692 is located in introns 7-8 (14, 18).
Given the critical participation of TMPRSS2 and SERPINE1 in SARS-CoV-2 infection, it is relevant to investigate whether their genetic variants could be associated with the severity of clinical manifestations and/or fatal outcomes in COVID-19 patients. The aim of this study was to determine the associations of the polymorphisms rs2070788, rs75603675 and rs12329760 of the TMPRSS2 gene and the polymorphisms rs2227631, rs2227667, rs2070682 and rs2227692 of the SERPINE1 gene with COVID-19 severity and their relationships with inflammatory biomarkers.
2 Materials and methods
2.1 Setting and participants
We conducted a cross-sectional study including 1536 patients from different Mexican institutions of governmental health care from June 2020 to March 2021. Nonprobability sampling was performed for unvaccinated patients. The inclusion criteria were as follows: individuals who were not familiar with COVID-19, independent of their sex, were aged ≥18 years with clinical features of COVID-19, and had a positive qRT–PCR test from a nasopharyngeal swab. Participant enrollment was performed in the following public hospitals of the Mexican governmental health system located in Mexico City: Instituto Nacional de Rehabilitación Luis Guillermo Ibarra Ibarra (17/20 AC); Instituto Nacional de Cardiología Ignacio Chávez (20–1202); Hospital Central Militar (045/2020); Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán (REF 3340); and Hospital General Dr. Manuel Gea González (CONBIOETICA09-CEI-024-20161215).
The exclusion criteria were pregnancy and incomplete clinical records. The participants were classified according to disease severity as previously described (19). All demographic and clinical data were obtained from the clinical records of each included patient.
This research complied with the Declaration of Helsinki and was approved by the participating health institutions’ ethics and research committees. In addition, all participants provided written informed consent before agreeing to participate in the study.
2.2 Blood, serum, and DNA sample processing
Peripheral blood samples were collected from each participant at the hospital’s triage for DNA and serum isolation. Genomic DNA was isolated using a specialized commercial kit (QIAmp DNA Blood Mini Kit, part number 51106, Qiagen, Hilden, Germany). The quality of the DNA samples was evaluated by the 260/280 nm absorbance ratio, and 1% agarose gels were stained with SYBR® Green (Invitrogen, CA, USA). Then, the DNA concentration was quantified using a Thermo Scientific NanoDrop spectrophotometer to measure the absorbance at wavelengths ranging from 260 to 280 nm; the quality of the samples ranged from 1.8-2.0, and the concentration was adjusted to 20 ng/μl. In addition, a vacutainer tube with SST II Advance gel was used for serum isolation. Serum samples were separated and stored at -80°C until further use.
2.3 Selection of single nucleotide polymorphisms (SNPs)
The polymorphisms of TMPRSS2 (rs2070788, rs75603675, rs12329760) and SERPINE1 (rs2227631, rs2227667, rs2070682, rs2227692) were selected on the basis of their previous scientific evidence of associations with different diseases in any population that included independent genetic studies from 2003–2020. The included polymorphisms had to present a minor allele frequency (MAF) ≥5%, according to the 1000 Genomes Project or Hap map in the Mexican population (MXL) or the Iberia (IBS) population (20).
For genotyping, 10 ng/μl genomic DNA was transferred into genotyping OpenArray plates, which previously contained the specific genotyping primers and probes, using the AccuFill system. Real-time PCR amplification was performed according to the supplier’s protocol using the Open Array Platform through a Quant Studio 12 K Flex System (Thermo Fisher Scientific, Waltham, MA, USA), and the results were analyzed using TaqMan Genotyper v1.6 software.
2.4 Statistical analysis
We performed an exploratory bivariate analysis. Nonparametric variables are reported as medians (p50) with interquartile ranges (IQRs). We used the Kruskal−Wallis test for continuous variables, while categorical variables were evaluated with the chi-square test. Hardy−Weinberg equilibrium was tested for all SNPs with a mild outcome. Linkage disequilibrium estimations between SNPs and haplotypes were performed with Haploview V4.2 Software (Broad Institute of Massachusetts Institute of Technology and Harvard University, Cambridge, MA, USA).
Binary logistic regression analysis was applied to determine the genetic associations with the outcomes of patients with COVID-19. The main inherence models were considered and adjusted for risk confounding variables such as age, sex, obesity, type 2 diabetes, and hypertension. In addition, the final models were assessed using the Hosmer−Lemeshow goodness-of-fit test.
To determine the association of SERPINE1 polymorphisms with increased D-dimer levels, we conducted a logistic analysis adjusted for age, sex, obesity, type 2 diabetes, and hypertension, stratifying D-dimer according to serum levels. The cutoff points for high and low concentrations were ≥500 ng/mL and <500 ng/mL, respectively (21).
The statistical analysis used STATA v.16 (StataCorp, Texas, USA). A P value < 0.05 indicated statistical significance.
3 Results
3.1 Patients and clinical traits
We enrolled 1,728 patients; however, subjects with incomplete data were excluded (n=172). For the final analysis, 1,536 patients were classified according to disease severity into mild (35%), severe (33%), critical (18%) and deceased (14%) groups. Sixty-four percent of the total population were males, and the median age was 55 years (IQR=45-65). However, the median age of the deceased group was 67 years (IQR=57-75.5). Further relevant clinical and laboratory features are depicted in Table 1.
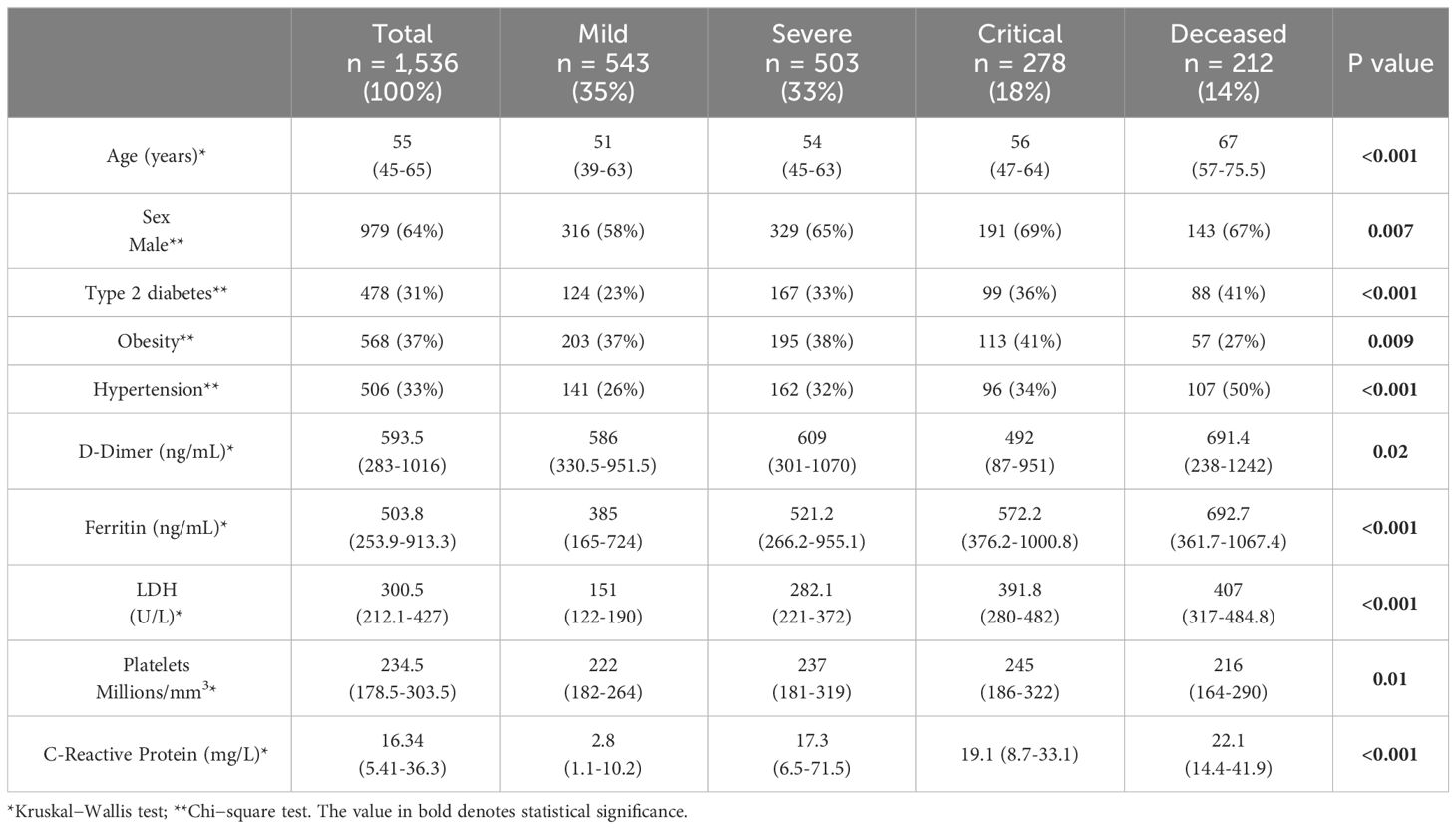
Table 1 Clinical parameters and anthropometric characteristics of the population.
Our data revealed that the highest levels of D-dimer (p50 = 691.41 ng/mL (IQR=238-1242)), ferritin (p50 = 692.7 ng/mL (IQR=361.7-1067.4)), LDH (p50 = 407 ng/mL (IQR=317-484.8)) and C-reactive protein (p50 = 22.1 mg/L (IQR=14.4-41.9)) were detected in the deceased group (Table 1). Although the laboratory parameters tended to increase with disease severity, this was not the case for D-dimer in the critical group.
3.2 Genotypes and allelic frequencies by disease severity and linkage disequilibrium
The genotypic and allelic frequencies among the COVID-19 groups were assessed for each TMPRSS2 (rs2070788, rs75603675, rs12329760) and SERPINE1 (rs2227631, rs2227667, rs2070682, rs2227692) polymorphism (Supplementary 1). We identified two genetic variants out of seven with a statistically significant difference in allelic frequency distribution among COVID-19 patients, corresponding to rs2070788 (P=0.003) and rs7560375 (P=0.04) of the TMPRSS2 gene. Nevertheless, only rs2070788 was also significant for genotype distribution (P=0.009).
Our data showed that only the TMPRSS2 rs75603675 and SERPINE1 rs2227667 genotypes were not in Hardy−Weinberg equilibrium (P=0.03 and P=0.02, respectively). The SERPINE1 polymorphisms rs2227667, rs2070682 and rs2227692 displayed linkage disequilibrium (LD), with D´ values of 0.99 and r2 = 0.74 (Figure 1). The frequencies of the haplotypes were 27% for ATC, 27% for ATT, 13% for GTC, 13% for GCC, 10% for GTT and 10% for GCT. The TMPRSS2 polymorphisms did not show an LD.
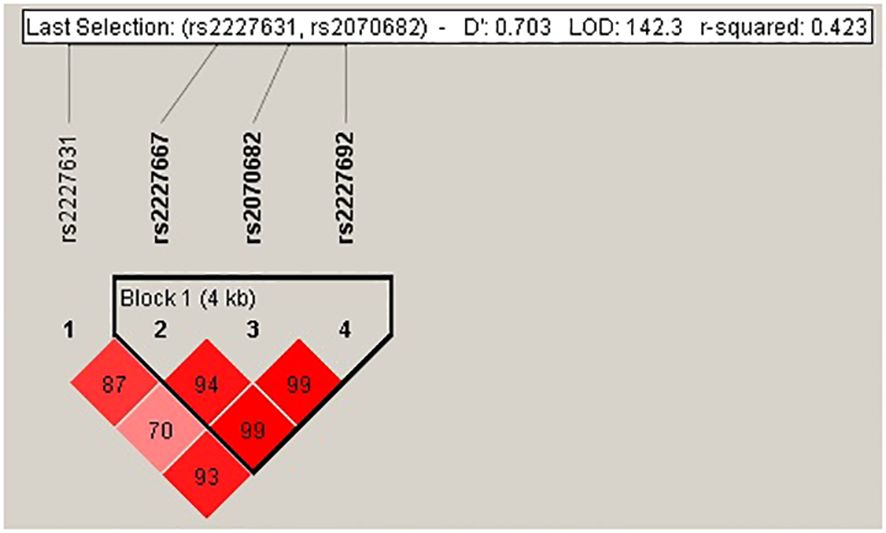
Figure 1 Linkage disequilibrium of rs2227631, rs2227667, rs2070682 and rs2227692 of the SERPINE1 gene. The D´ value showed a LD of rs2227667, rs2070682 and rs2227692.
3.3 Logistic regression analysis of TMPRSS2 and SERPINE1 polymorphisms in COVID-19 patients
Logistic regression analysis adjusted for age, sex, and comorbidities (hypertension, type 2 diabetes, and obesity) revealed a statistically significant association between the TMPRSS2 rs2070788 and rs75603675 genetic variants and between the SERPINE1 rs2227631, rs222667 and rs2227692 and critical and deceased outcomes. Table 2 summarizes the significance of each TMPRSS2 and SERPINE1 polymorphism with respect to COVID-19 outcomes.
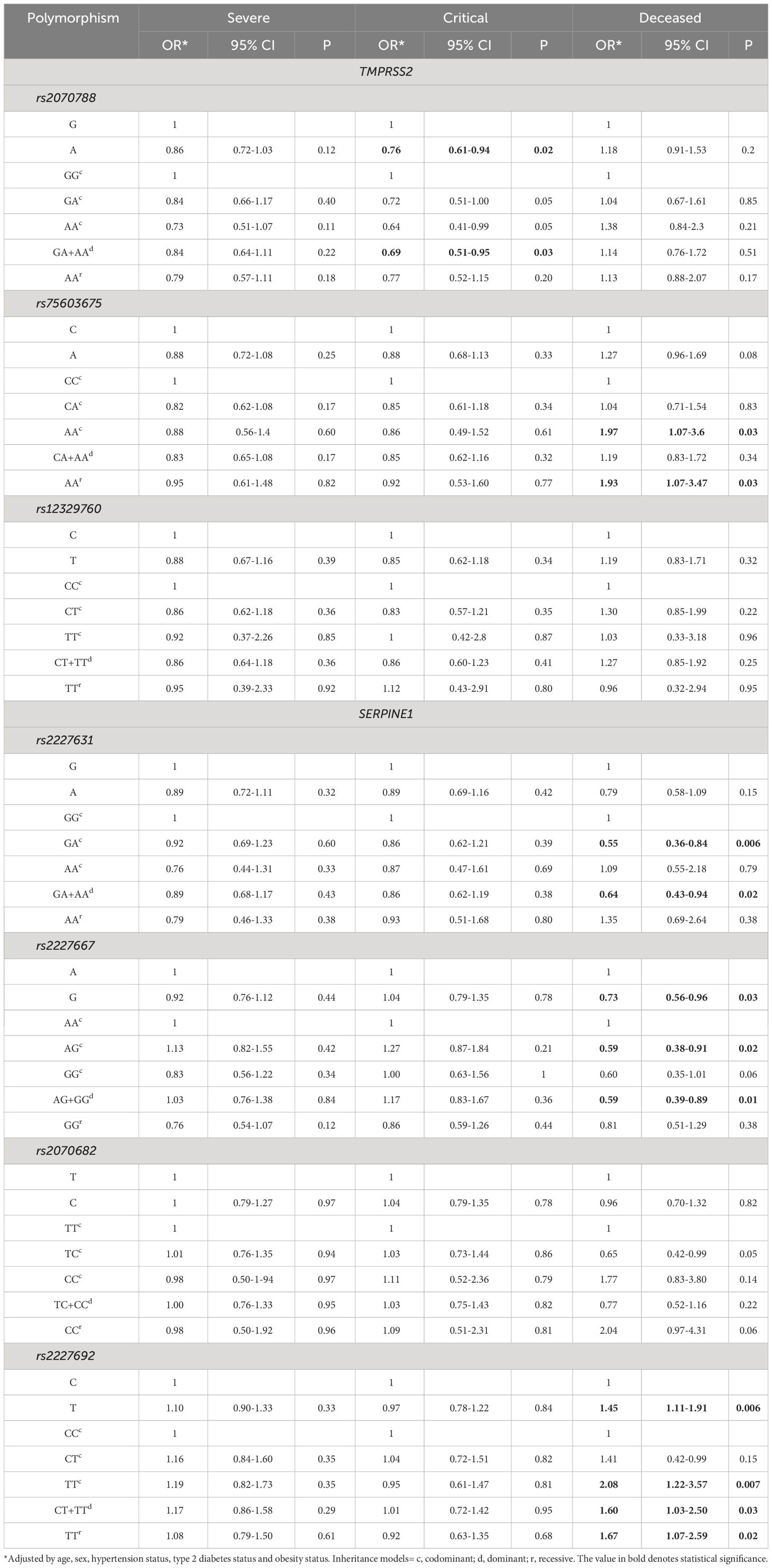
Table 2 Associations of TMPRSS2 and SERPINE1 polymorphisms with COVID-19 severity.
3.4 Polymorphisms associated with protection
The protective effects of the TMPRSS2 and SERPINE1 genetic variants are shown in Table 2. For rs2070788, the A allele had an OR of 0.76 (95% CI = 0.61-0.94; P=0.02), indicating a critical COVID-19 outcome. Moreover, the dominant model (GA+AA) exerted a protective effect, with an OR of 0.69 (95% CI 0.51-0.95; P=0.03) for critical outcomes.
Interestingly, patients who died from COVID-19 were the main group in which rs2227631 and rs2227667 of SERPINE1 showed a significant protective effect. Similarly, the rs2227631 GA genotype had an OR of 0.55 (95% CI = 0.36-0.84; P=0.006), and similar results were found for the dominant model (AG+GG), with an OR of 0.64 (95% CI = 0.43-0.94; P=0.02). Regarding the rs2227667 G allele, a significant protective association was also observed with decreased COVID-19 incidence (OR=0.73; 95% CI = 0.65-0.96; P=0.03). Similar results were obtained for the GA genotype, with an OR of 0.59 (95% CI = 0.38-0.91; P=0.02) in the deceased group.
3.5 Risk polymorphisms
According to the main inherent genetics models of TMPRSS2 rs75603675, a statistically significant association was found between the AA genotype and decreased COVID-19 incidence (OR=1.97; 95% CI 1.07-3.6; P=0.03). Similarly, a significant association was observed for the rs2227692 T allele of the SERPINE1 gene, with the deceased outcome showing an OR of 1.45 (95% CI = 1.11-1.91; P=0.006). The main inherence genetics models were associated with the deceased group (Table 2). Interestingly, the codominant model for the TT genotype had an OR of 2.08 (95% CI=1.22-3.57; P=0.007) for the deceased outcome, while the dominant (TC+TT) and recessive models had ORs of 1.60 (95% CI=1.03-2.50; P=0.03) and 1.67 (95% CI=1.07-2.59; P=0.02), respectively.
We performed a logistic regression of SERPINE1 haplotypes and only found a statistically significant association between the ATT haplotype and the deceased group, with an OR of 1.6 (95% CI = 1.08-2.25; P=0.02).
3.6 Associations of TMPRSS2 and SERPINE1 polymorphisms with clinical laboratory parameters stratified by outcome
To understand the impact of each polymorphism on the immune-hematological response to SARS-CoV-2, we further studied the relationships of each polymorphism with diverse laboratory features among COVID-19 patients. In this sense, the analysis revealed a significant difference between the minor allele carriers of SERPINE1 rs2227667, which had a decreased LDH concentration compared to the major allele (p50 = 306 U/L; IQR=205-431) vs. p50 = 322 U/L; IQR=222-453, respectively) (P=0.03). Although the minor allele of rs2227631 also tended to decrease LDH concentration, there was no statistically significant difference from the major allele. On the other hand, the risk variant of the SERPINE1 gene (rs2227692) had increased D-dimer levels compared with those of the minor and major alleles (p50 = 613.5 ng/mL; IQR=281.6-1035 vs. p50 = 545.8 ng/mL; IQR=229-1001, respectively) (P=0.02). This was also observed for the LDH concentration (p50 = 317 U/L; IQR=222-453 vs. p50 = 294 U/L; IQR=202-426.1) (P=0.01) (Table 3).
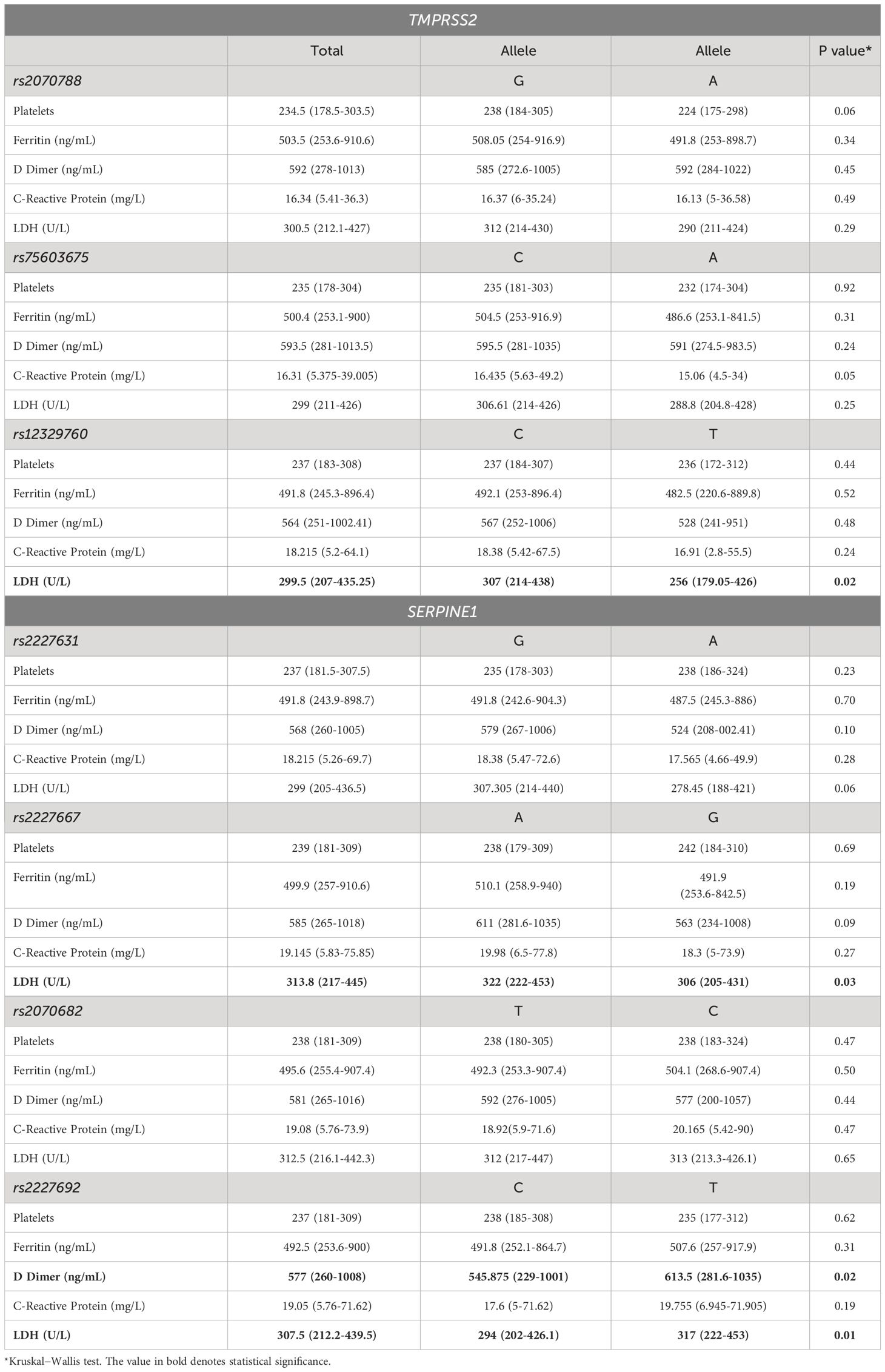
Table 3 Laboratory parameters of COVID-19 patients stratified by polymorphism alleles.
Due to the possible relationship between D-dimer and SERPINE1 gene risk polymorphisms, we further evaluated the association of each genetic variant with D-dimer, which was dichotomized based on the cutoff value of 500 ng/mL. Our population study revealed a significant association between the T allele of the rs2227692 polymorphism and a high D-dimer concentration (>500 ng/mL) (OR=1.24; 95% CI=1.03-1.48; P=0.02, Table 4). Interestingly, when COVID-19 outcomes were stratified, rs2227692 maintained its association in the deceased patient group (OR=2.04; 95% CI=1.23-3.37; p=0.006) (Table 5).
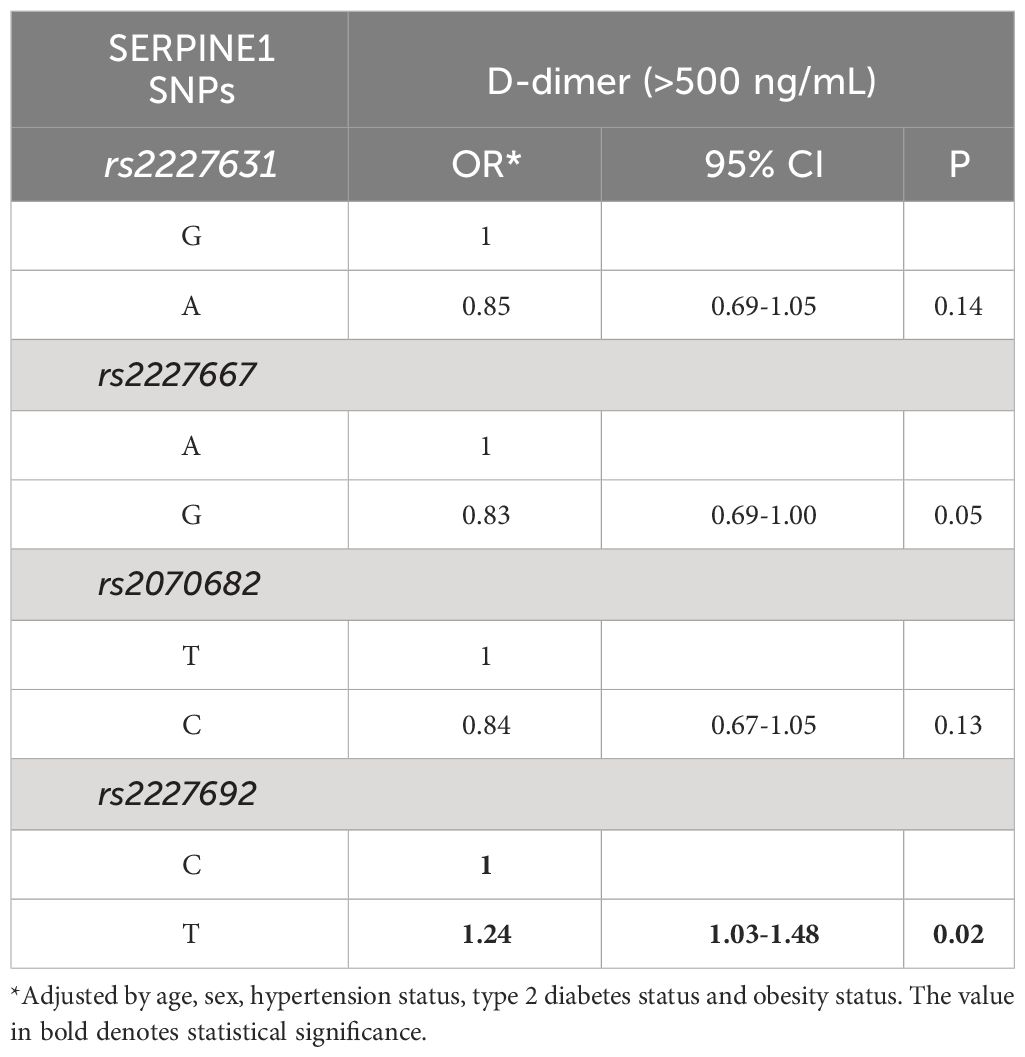
Table 4 Association of high D-dimer concentrations with SERPINE1 risk polymorphisms.
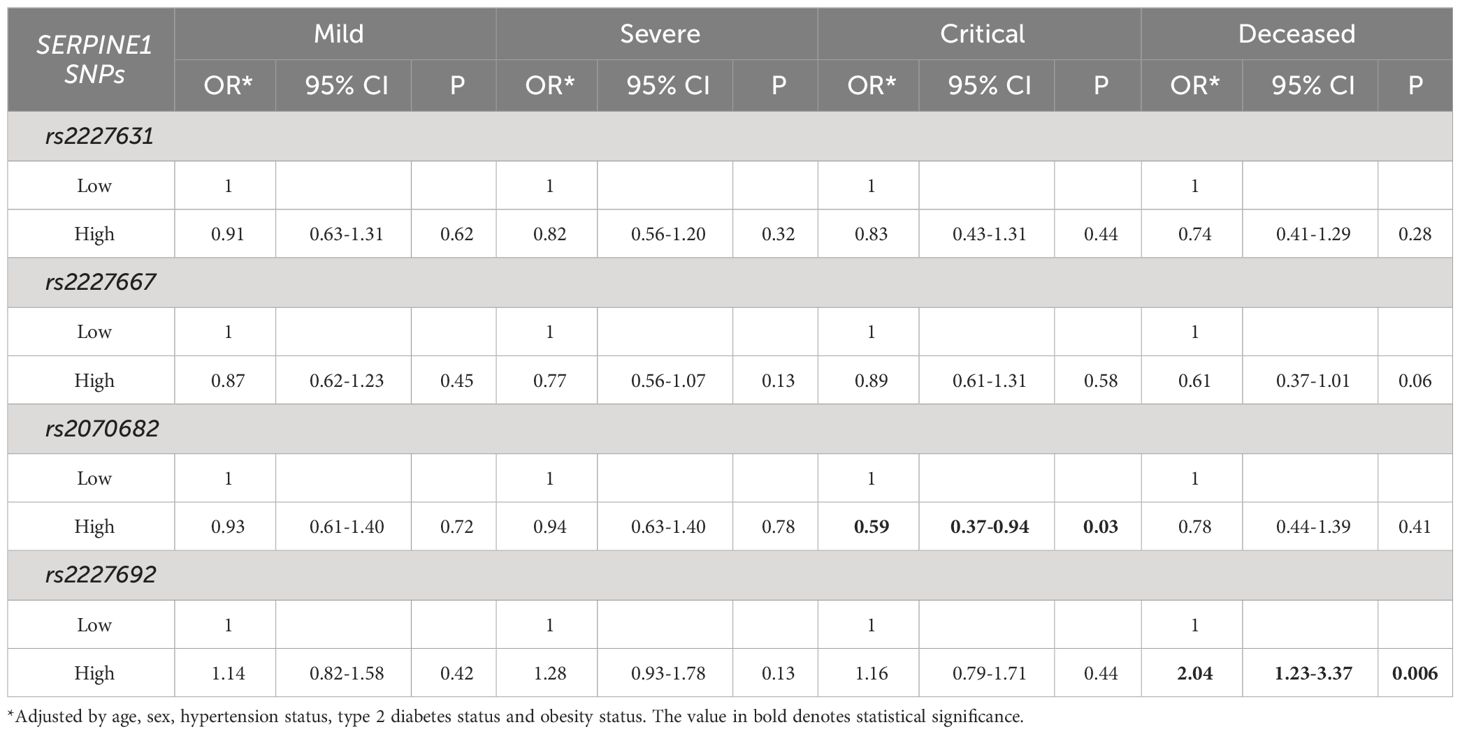
Table 5 Laboratory parameters of COVID-19 patients stratified by polymorphism alleles.
4 Discussion
Since COVID-19 emerged, people infected with SARS-CoV-2 have experienced different clinical outcomes. Nevertheless, the development of fatal COVID-19 cases has been strongly associated with comorbidities, including obesity, type 2 diabetes, and cardiovascular diseases, which can exacerbate the inflammatory state (21–25).
In COVID-19 pathogenesis, many factors contribute to viral pathogenesis. Several authors have described three points: (1) recognition of the virus by cellular receptors; (2) suppression of the antiviral response; and (3) the ability to evade the immune system (23, 26).
SARS-CoV-2 uses the ACE2 receptor and TMPRSS2 to promote cellular entry by cleaving the S protein into S1 and S2 (27–29). TMPRSS2 is used by diverse viruses to infect humans, and it has been associated with physiological processes such as digestion, tissue remodeling, blood coagulation, fertility, inflammatory responses, and pain, among others, and the expression of TMPRSS2 is regulated by aging (30). Recent studies have explained the possible role of structural and regulatory variants of TMPRSS2 in susceptibility to COVID-19. In this sense, the variants p.Gly8Val/c.23G>T (rs75603675) and p.Val197Met/c.589G>A (rs12329760) have been reported to influence its interaction with ACE2 and the S protein (29, 31, 32). rs2070788 has been reported to be highly expressed in the lungs of patients at risk of developing severe COVID-19 (33), suggesting that these variants could play an important role in the severity of SARS-CoV-2 infection (34–36). Nevertheless, the study of TMPRSS2 polymorphisms has been described in some populations with contradictory results (37–39).
Our study explored the association of TMPRSS2 genetic variants with COVID-19 severity; interestingly, we observed that rs75603675 increased the risk of death due to COVID-19. rs75603675 is a missense variant, c.23G>T, which modifies protein structure to decrease specificity or induce impaired interaction with viral proteins. The isoforms of TMPRSS2 are composed of 492 amino acids and 22 cysteine residues (40, 41). Some reports have described different strains of SARS-CoV-2 with the TMPRSS2 receptor; of the different SARS-CoV-2 variants, the omicron is the least dependent on TMPRSS2 (42). In the present study, we did not characterize the viral strain; however, according to epidemiological data reported in Mexico from the first, second, and third waves (summer 2020-summer 2021), the SARS-CoV-2 variants were alpha-gamma, which is associated with a high mortality rate (43). In this sense, Sabyasachi Senapati et al. reported that rs75603675 could disrupt the local protein structure, increasing the stability of TMPRSS2, while rs12329760 increased the number of S protein domains (31). However, other SNPs in TMPRSS2 influence its expression (29). According to these findings, using a molecular docking approach, Sabayasachi Senapati et al. identified some phytochemicals that could bind to TMPRSS2 during host−interactions. However, the authors recommended in silico and in vitro studies to validate the efficacy of these phytochemicals.
The rs75603675 23G>T (Gly8Val) and rs12329760 (589 G>A, Val197Met) are theoretically considered to be responsible for changes in the interaction of TMPRSS2 with the S protein of the virus (26).
Posadas-Sánchez et al. previously studied the associations of the TMPRSS2 rs462574, rs456298, rs2298659, and rs12329760 (pV197M) polymorphisms with the risk of infection with SARS-CoV-2 in a Mexican population and reported that only rs462574 and rs456298 were associated with this association (44). Consistent with our results, rs12329760 (pV197M) does not show an association with either the risk or protection of infection. rs12329760 (pV197M) has been described to protect against COVID-19 in different populations because it is present in an exonic splicing enhancer site associated with protein malformation (45). This result also agrees with the study of Schönfelder et al. (2021), who also concluded that there is no association between rs12329760 and the risk of infection or COVID-19 severity (46).
In some studies, TMPRSS2 expression of the G allele of rs2070788 was associated with increased protein expression in lung tissue, which could lead to an association with increased susceptibility to COVID-19 (35, 41). Recently, a bioinformatics analysis performed by Mujalli et al. (2022) revealed the overexpression of genes implicated in the ACE2-TMPRSS2 signaling pathway in COVID-19 patients with severe and fatal phenotypes. Moreover, SERPINE1 was identified as a drug target gene of TMPRSS2, with a similarity score of 0.54, which could suggest an interaction with the spike protein of SARS-CoV-2 (47).
The SERPINE1 gene encodes PAI-1, which is implicated in coagulopathy and regulates the balance between coagulation and fibrinolytic systems. In some studies, coagulopathies have been reported in patients with severe COVID-19 (48). The mechanism of thrombosis in patients with COVID-19 may involve a cytokine syndrome that activates the coagulation process. For the imbalance of PAI-1, the coagulation process, among other factors, has been implicated as a genetic risk factor. SERPINE1 has been associated with thrombosis in diseases such as ischemic stroke, cancer, and, more recently, COVID-19 (10, 49–52).
In that sense, COVID-19 patients exhibit variable states of coagulopathy, with a marked thrombotic tendency among nonsurviving individuals. Salem N. et al. reported that 31% of COVID-19 patients exhibited increased hypercoagulability with hypofibrinolytic capacity (53, 54). These findings indicate the potential implications of SERPINE1 genetic variants for COVID-19 outcomes. Moreover, in infectious diseases, PAI-1 plays a role in the inflammatory process as a mediator of the early host defense response to combat pathogens and inhibits fibrinolysis and could be related to thrombophilia (55).
The association of several SERPINE1 polymorphisms with previously described pathologies has been reported and could represent a risk factor for severe COVID-19. The SERPINE1 polymorphisms evaluated in this study included rs2227631 (-1844 G/A), which is located in the promoter region and has been implicated as a regulatory region variant with possible functional loci; however, functional studies are necessary to explore the specific effects on COVID-19 severity (18). Furthermore, rs2227667, rs2070682, and rs2227692, located in intronic regions, were also analyzed (14).
In the present study, we observed a greater D-dimer concentration (>500 ng/mL) in COVID-19 patients, similar to that reported by others from different populations, who reported higher D-dimer concentrations in patients with COVID-19 and critical illness (21, 56). Chocron et al. (2021) reported that D-dimer is one of the measures used to detect COVID-19 severity; increased D-dimer levels could be a risk marker of thrombotic events (21). However, other authors found no association between D-dimer levels and death in COVID-19 patients (57, 58).
In addition, it has been reported that in patients with COVID-19, hyperinflammation can induce dysfunction and damage in endothelial cells, resulting in increased D-dimer levels (59). Lange et al. (2008) reported that SERPINE1 genetic variants were associated with D-dimer concentrations in older European and African-American populations and that rs2227667 was associated with higher D-dimer levels and fibrin deposits (11); however, gene SNPs explained ≈2% of the total variation in D-dimer levels (60).
Coagulation biomarkers such as D-dimer, which are associated with SERPINE1 genetic variants, could increase the risk of complications due to coagulation activity in patients with severe COVID-19. Identifying patients at risk of complications in clinical practice could improve treatment and outcomes to optimize health services. Lopez-Castaneda et al. (2021) suggested the use of low-molecular-weight heparin, as a prophylactic treatment for COVID-19 patients, to limit the hypercoagulable state (13).
Dittmann et al. (2015) showed that some SERPINE1 SNPs could inhibit TMPRSS2, showing an antiviral effect against the influenza A virus (61). rs2227631 and rs2227667 of the SERPINE1 gene could have antiviral effects by preventing viral membrane fusion of SARS-CoV-2, leading to the inhibition of TMPRSS2-mediated S protein cleavage. In this sense, Rosendal et al. (2022) showed that SERPINE1 prevents the cleavage of the S protein by binding to TMPRSS2 (5). Moreover, the authors found that the antiviral effects of SERPINA1, SERPINE1, SERPINE2, and SERPINF1 were observed during the first steps of infection in HBEC ALI cultures, revealing reduced SARS-CoV-2 entry into target cells (5).
Treatment with serpentine targets could decrease lung inflammation and modify thrombotic protease and complement levels. PEGSerp-1 has been developed as a new anti-inflammatory therapeutic or biologic for vascular damage, coagulation disorders, and inflammation damage (1). The highlights of the present study contribute to the knowledge of the specific polymorphisms that could affect COVID-19 severity through the identification of new pharmacological targets and treatment strategies to block SARS-CoV-2 entry via TMPRSS2 inhibition, such as argatroban and famotidine, which act as new scaffolds for TMPRSS2 inhibition (62), and other future treatments that could be used for other similar diseases.
SERPINE has been identified as an endogenous antiviral molecule against SARS-CoV-2 and could represent a possible treatment option due to its biological role in inhibiting the entry of the virus into host cells (22). Polymorphisms of TMPRSS2 and SERPINE1 could be associated with COVID-19 severity, modifying the susceptibility to fatal outcomes. However, it is necessary to elucidate the genetic susceptibility to severe disease caused by SARS-CoV-2.
In response to the recent COVID-19 pandemic, the search for therapeutic targets to combat the severity and complications of infection caused by SARS-CoV-2 has led to the identification of SERPINE1 as a natural inhibitor of the TMPRSS2 protease that enhances the viral infection process.
In conclusion, our study revealed that the TMPRSS2 rs75603675 gene variant may induce an amino acid change from glycine to valine, which is more frequent in patients with COVID-19 who die. Similarly, we were able to demonstrate the association of the SERPINE1 rs2227692 variant not only with a decreased outcome but also with an increase in D-dimer, which could influence the altered processes of fibrinolysis development in patients with a hypercoagulable state, as observed in critical pathology outcomes (Figure 2).
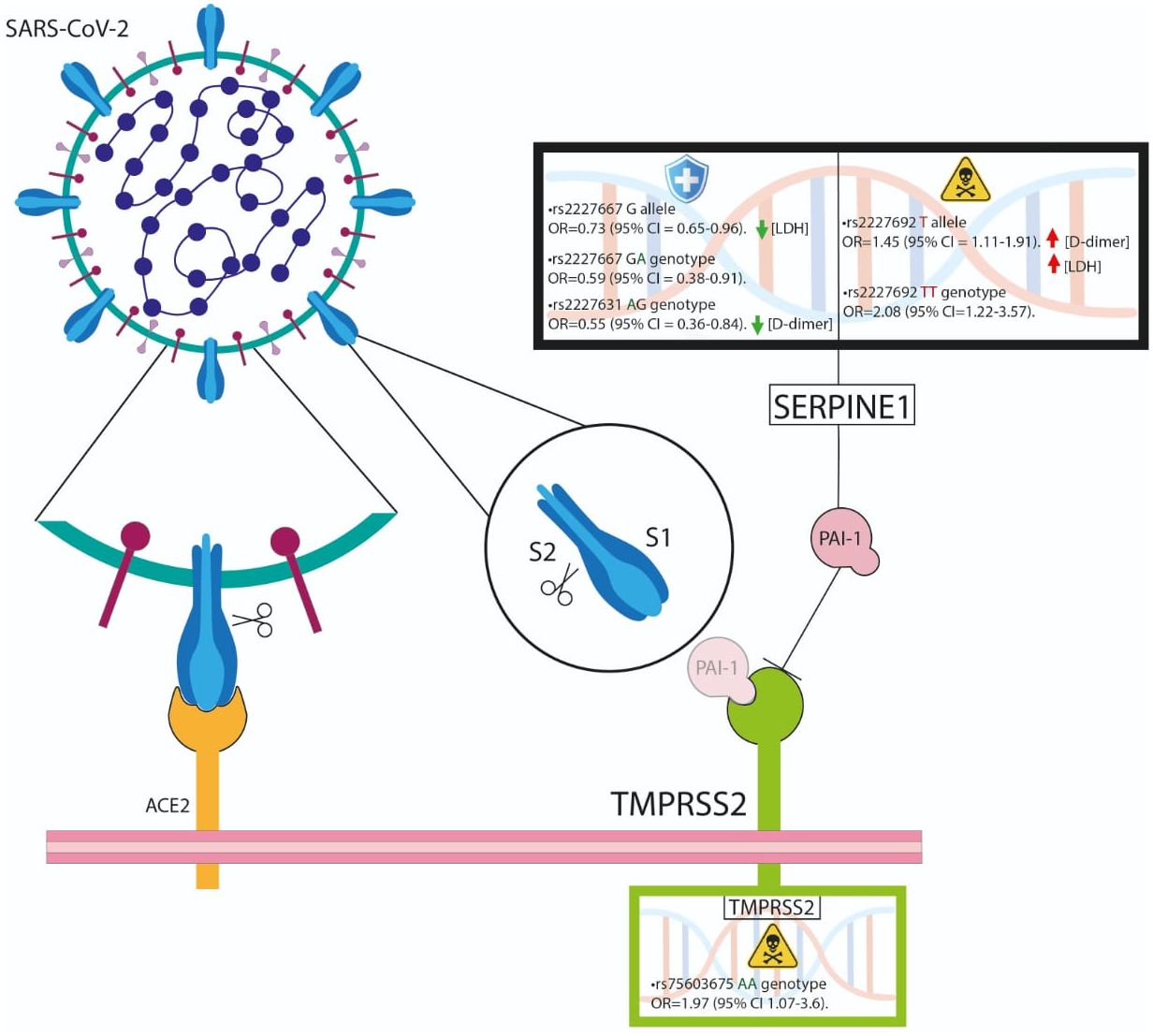
Figure 2 The TMPRSS2 rs75603675 and SERPINE1 rs2227692 variants are associated with poor outcomes. SERPINE1 could influence the altered processes of fibrinolysis development in hypercoagulable patients.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Ethics statement
The study was approved by the ethics committee of (INR-LGII: 17/20). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
LM-G: Formal analysis, Methodology, Software, Writing – original draft. CM-A: Methodology, Writing – original draft. TT-L: Conceptualization, Supervision, Validation, Writing – review & editing. PV-C: Visualization, Writing – review & editing. RV-V: Resources, Writing – review & editing. JR-H: Data curation, Writing – review & editing. DG-M: Visualization, Writing – review & editing. GV-A: Data curation, Writing – review & editing. RP-S: Data curation, Writing – review & editing. JF: Data curation, Writing – review & editing. AP: Validation, Writing – review & editing. JR-P: Data curation, Investigation, Writing – review & editing. MM-M: Resources, Writing – review & editing. GV-Z: Resources, Writing – review & editing. AM-C: Resources, Writing – review & editing. FM-R: Resources, Writing – review & editing. DZ-A: Resources, Writing – review & editing. LR-T: Investigation, Resources, Writing – review & editing. AM-A: Visualization, Writing – review & editing. MC-R: Investigation, Resources, Writing – review & editing. MO-S: Methodology, Validation, Writing – review & editing. YS-K: Resources, Writing – review & editing. CS-A: Data curation, Writing – review & editing. JO-T: Investigation, Methodology, Writing – review & editing. BH-L: Investigation, Methodology, Writing – review & editing. CP: Project administration, Writing – review & editing. GM-N: Supervision, Writing – review & editing. AL-R: Funding acquisition, Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was funded by the Consejo Nacional de Ciencia y Tecnología; CONACYT 312513 SARS-COV 2.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2024.1335963/full#supplementary-material
Supplementary 1 | TMPRSS2 and SERPINE1 polymorphism allelic and genotype frequencies in the population study.
References
1. Zhang L, Li YH, Kibler K, Kraberger S, Varsani A, Turk J, et al. Viral anti-inflammatory serpin reduces immuno-coagulopathic pathology in SARS-CoV-2 mouse models of infection. EMBO Mol Med. (2023) 15:e17376. doi: 10.15252/emmm.202317376
2. Rahbar Saadat Y, Hosseiniyan Khatibi SM, Zununi Vahed S, Ardalan M. Host serine proteases: A potential targeted therapy for COVID-19 and influenza. Front Mol Biosci. (2021) 8:725528. doi: 10.3389/fmolb.2021.725528
3. Luan B, Huynh T, Cheng X, Lan G, Wang HR. Targeting proteases for treating COVID-19. J Proteome Res. (2020) 19:4316–26. doi: 10.1021/acs.jproteome.0c00430
4. Nejat R, Torshizi MF, Najafi DJ, Protein S. ACE2 and host cell proteases in SARS-coV-2 cell entry and infectivity; is soluble ACE2 a two blade sword? A narrative review. Vaccines (Basel). (2023) 11(2):204. doi: 10.3390/vaccines11020204
5. Rosendal E, Mihai IS, Becker M, Das D, Frängsmyr L, Persson BD, et al. Serine protease inhibitors restrict host susceptibility to SARS-coV-2 infections. mBio. (2022) 13:e0089222. doi: 10.1128/mbio.00892-22
6. Wettstein L, Kirchhoff F, Münch J. The transmembrane protease TMPRSS2 as a therapeutic target for COVID-19 treatment. Int J Mol Sci. (2022) 23(3):1351. doi: 10.3390/ijms23031351
7. Mahoney M, Damalanka VC, Tartell MA, Chung DH, Lourenço AL, Pwee D, et al. A novel class of TMPRSS2 inhibitors potently block SARS-CoV-2 and MERS-CoV viral entry and protect human epithelial lung cells. Proc Natl Acad Sci U.S.A. (2021) 118(43):e2108728118. doi: 10.1073/pnas.2108728118
8. Shapira T, Monreal IA, Dion SP, Buchholz DW, Imbiakha B, Olmstead AD, et al. A TMPRSS2 inhibitor acts as a pan-SARS-CoV-2 prophylactic and therapeutic. Nature. (2022) 605:340–8. doi: 10.1038/s41586-022-04661-w
9. Shen LW, Mao HJ, Wu YL, Tanaka Y, Zhang W. TMPRSS2: A potential target for treatment of influenza virus and coronavirus infections. Biochimie. (2017) 142:1–10. doi: 10.1016/j.biochi.2017.07.016
10. Wang T, Zhai Y, Xue H, Zhou W, Ding Y, Nie H. Regulation of epithelial sodium transport by SARS-coV-2 is closely related with fibrinolytic system-associated proteins. Biomolecules. (2023) 13(4):578. doi: 10.3390/biom13040578
11. Mosesson MW, Siebenlist KR, Meh DA. The structure and biological features of fibrinogen and fibrin. Ann N Y Acad Sci. (2001) 936:11–30. doi: 10.1111/j.1749-6632.2001.tb03491.x
12. Udovenko A, Makogonenko Y, Korolova D, Druzhyna N, Chernyshenko V, Komisarenko S. Formation and elimination of soluble fibrin and D-dimer in the bloodstream. Croat Med J. (2023) 64:421–9. doi: 10.3325/cmj.2023.64.421
13. Lopez-Castaneda S, García-Larragoiti N, Cano-Mendez A, Blancas-Ayala K, Damian-Vázquez G, Perez-Medina AI, et al. Inflammatory and prothrombotic biomarkers associated with the severity of COVID-19 infection. Clin Appl Thromb Hemost. (2021) 27:1076029621999099. doi: 10.1177/1076029621999099
14. Tziastoudi M, Dardiotis E, Pissas G, Filippidis G, Golfinopoulos S, Siokas V, et al. Serpin family E member 1 tag single-nucleotide polymorphisms in patients with diabetic nephropathy: an association study and meta-analysis using a genetic model-free approach. Genes (Basel). (2021) 12(12):1887. doi: 10.3390/genes12121887
15. Khan SS. The central role of PAI-1 in COVID-19: thrombosis and beyond. Am J Respir Cell Mol Biol. (2021) 65:238–40. doi: 10.1165/rcmb.2021-0208ED
16. Whyte CS, Simpson M, Morrow GB, Wallace CA, Mentzer AJ, Knight JC, et al. The suboptimal fibrinolytic response in COVID-19 is dictated by high PAI-1. J Thromb Haemost. (2022) 20:2394–406. doi: 10.1111/jth.15806
17. Singh H, Choudhari R, Nema V, Khan AA. ACE2 and TMPRSS2 polymorphisms in various diseases with special reference to its impact on COVID-19 disease. Microb Pathog. (2021) 150:104621. doi: 10.1016/j.micpath.2020.104621
18. Raman S, Dada R, Nag TC, Yadav CS. Plasminogen activator inhibitor-1 promoter sequence variations in idiopathic osteonecrosis of head of femur. Indian J Med Res. (2021) 154:849–56. doi: 10.4103/ijmr.IJMR_496_19
19. Martínez-Gómez LE, Ibarra-González I, Fernández-Lainez C, Tusie T, Moreno-Macías H, Martinez-Armenta C, et al. Metabolic reprogramming in SARS-coV-2 infection impacts the outcome of COVID-19 patients. Front Immunol. (2022) 13:936106. doi: 10.3389/fimmu.2022.936106
20. Auton A, Brooks LD, Durbin RM, Garrison EP, Kang HM, Korbel JO, et al. A global reference for human genetic variation. Nature. (2015) 526:68–74. doi: 10.1038/nature15393
21. Chocron R, Duceau B, Gendron N, Ezzouhairi N, Khider L, Trimaille A, et al. D-dimer at hospital admission for COVID-19 are associated with in-hospital mortality, independent of venous thromboembolism: Insights from a French multicenter cohort study. Arch Cardiovasc Dis. (2021) 114:381–93. doi: 10.1016/j.acvd.2021.02.003
22. Humphreys SJ, Whyte CS, Mutch NJ. "Super" SERPINs-A stabilizing force against fibrinolysis in thromboinflammatory conditions. Front Cardiovasc Med. (2023) 10:1146833. doi: 10.3389/fcvm.2023.1146833
23. Jackson CB, Farzan M, Chen B, Choe H. Mechanisms of SARS-CoV-2 entry into cells. Nat Rev Mol Cell Biol. (2022) 23:3–20. doi: 10.1038/s41580-021-00418-x
24. López-Reyes A, Martinez-Armenta C, Espinosa-Velázquez R, Vázquez-Cárdenas P, Cruz-Ramos M, Palacios-Gonzalez B, et al. NLRP3 inflammasome: the stormy link between obesity and COVID-19. Front Immunol. (2020) 11:570251. doi: 10.3389/fimmu.2020.570251
25. Serebrovska ZO, Chong EY, Serebrovska TV, Tumanovska LV, Xi L. Hypoxia, HIF-1α, and COVID-19: from pathogenic factors to potential therapeutic targets. Acta Pharmacol Sin. (2020) 41:1539–46. doi: 10.1038/s41401-020-00554-8
26. Minashkin MM, Grigortsevich NY, Kamaeva AS, Barzanova VV, Traspov AA, Godkov MA, et al. The role of genetic factors in the development of acute respiratory viral infection COVID-19: predicting severe course and outcomes. Biomedicines. (2022) 10(3):549. doi: 10.3390/biomedicines10030549
27. Hartog N, Faber W, Frisch A, Bauss J, Bupp CP, Rajasekaran S, et al. SARS-CoV-2 infection: molecular mechanisms of severe outcomes to suggest therapeutics. Expert Rev Proteomics. (2021) 18:105–18. doi: 10.1080/14789450.2021.1908894
28. Hou Y, Zhao J, Martin W, Kallianpur A, Chung MK, Jehi L, et al. New insights into genetic susceptibility of COVID-19: an ACE2 and TMPRSS2 polymorphism analysis. BMC Med. (2020) 18:216. doi: 10.1186/s12916-020-01673-z
29. Senapati S, Banerjee P, Bhagavatula S, Kushwaha PP, Kumar S. Contributions of human ACE2 and TMPRSS2 in determining host-pathogen interaction of COVID-19. J Genet. (2021) 100(1):12. doi: 10.1007/s12041-021-01262-w
30. Thunders M, Delahunt B. Gene of the month: TMPRSS2 (transmembrane serine protease 2). J Clin Pathol. (2020) 73:773–6. doi: 10.1136/jclinpath-2020-206987
31. Jeon S, Blazyte A, Yoon C, Ryu H, Jeon Y, Bhak Y, et al. Regional TMPRSS2 V197M allele frequencies are correlated with COVID-19 case fatality rates. Mol Cells. (2021) 44:680–7. doi: 10.14348/molcells.2021.2249
32. Sharma S, Singh I, Haider S, Malik MZ, Ponnusamy K, Rai E. ACE2 homo-dimerization, human genomic variants and interaction of host proteins explain high population specific differences in outcomes of COVID19. bioRxiv. (2020), 050534. doi: 10.1101/2020.04.24.050534
33. Irham LM, Chou WH, Calkins MJ, Adikusuma W, Hsieh SL, Chang WC. Genetic variants that influence SARS-CoV-2 receptor TMPRSS2 expression among population cohorts from multiple continents. Biochem Biophys Res Commun. (2020) 529:263–9. doi: 10.1016/j.bbrc.2020.05.179
34. Adimulam T, Arumugam T, Gokul A, Ramsuran V. Genetic variants within SARS-coV-2 human receptor genes may contribute to variable disease outcomes in different ethnicities. Int J Mol Sci. (2023) 24(10):8711. doi: 10.3390/ijms24108711
35. Kehdy FSG, Pita-Oliveira M, Scudeler MM, Torres-Loureiro S, Zolini C, Moreira R, et al. Human-SARS-CoV-2 interactome and human genetic diversity: TMPRSS2-rs2070788, associated with severe influenza, and its population genetics caveats in Native Americans. Genet Mol Biol. (2021) 44:e20200484. doi: 10.1590/1678-4685-gmb-2020-0484
36. Pandey RK, Srivastava A, Singh PP, Chaubey G. Genetic association of TMPRSS2 rs2070788 polymorphism with COVID-19 case fatality rate among Indian populations. Infect Genet Evol. (2022) 98:105206. doi: 10.1016/j.meegid.2022.105206
37. Monticelli M, Hay Mele B, Benetti E, Fallerini C, Baldassarri M, Furini S, et al. Protective role of a TMPRSS2 variant on severe COVID-19 outcome in young males and elderly women. Genes (Basel). (2021) 12(4):596. doi: 10.3390/genes12040596
38. Ravikanth V, Sasikala M, Naveen V, Latha SS, Parsa KVL, Vijayasarathy K, et al. A variant in TMPRSS2 is associated with decreased disease severity in COVID-19. Meta Gene. (2021) 29:100930. doi: 10.1016/j.mgene.2021.100930
39. Rokni M, Heidari Nia M, Sarhadi M, Mirinejad S, Sargazi S, Moudi M, et al. Association of TMPRSS2 gene polymorphisms with COVID-19 severity and mortality: a case-control study with computational analyses. Appl Biochem Biotechnol. (2022) 194:3507–26. doi: 10.1007/s12010-022-03885-w
40. Karimian A, Behjati M, Karimian M. Molecular mechanisms involved in anosmia induced by SARS-CoV-2, with a focus on the transmembrane serine protease TMPRSS2. Arch Virol. (2022) 167:1931–46. doi: 10.1007/s00705-022-05545-0
41. Villapalos-García G, Zubiaur P, Rivas-Durán R, Campos-Norte P, Arévalo-Román C, Fernández-Rico M, et al. Transmembrane protease serine 2 (TMPRSS2) rs75603675, comorbidity, and sex are the primary predictors of COVID-19 severity. Life Sci Alliance. (2022) 5(10):e202201545. doi: 10.26508/lsa.202201545
42. Zhao H, Lu L, Peng Z, Chen LL, Meng X, Zhang C, et al. SARS-CoV-2 Omicron variant shows less efficient replication and fusion activity when compared with Delta variant in TMPRSS2-expressed cells. Emerg Microbes Infect. (2022) 11:277–83. doi: 10.1080/22221751.2021.2023329
43. Ascencio-Montiel IJ, Ovalle-Luna OD, Rascón-Pacheco RA, Borja-Aburto VH, Chowell G. Comparative epidemiology of five waves of COVID-19 in Mexico, March 2020-August 2022. BMC Infect Dis. (2022) 22:813. doi: 10.1186/s12879-022-07800-w
44. Posadas-Sánchez R, Fragoso JM, Sánchez-Muñoz F, Rojas-Velasco G, Ramírez-Bello J, López-Reyes A, et al. Association of the transmembrane serine protease-2 (TMPRSS2) polymorphisms with COVID-19. Viruses. (2022) 14(9):1976. doi: 10.3390/v14091976
45. Paniri A, Hosseini MM, Akhavan-Niaki H. First comprehensive computational analysis of functional consequences of TMPRSS2 SNPs in susceptibility to SARS-CoV-2 among different populations. J Biomol Struct Dyn. (2021) 39:3576–93. doi: 10.1080/07391102.2020.1767690
46. Schönfelder K, Breuckmann K, Elsner C, Dittmer U, Fistera D, Herbstreit F, et al. Transmembrane serine protease 2 Polymorphisms and Susceptibility to Severe Acute Respiratory Syndrome Coronavirus Type 2 Infection: A German Case-Control Study. Front Genet. (2021) 12:667231. doi: 10.3389/fgene.2021.667231
47. Mujalli A, Alghamdi KS, Nasser KK, Al-Rayes N, Banaganapalli B, Shaik NA, et al. Bioinformatics insights into the genes and pathways on severe COVID-19 pathology in patients with comorbidities. Front Physiol. (2022) 13:1045469. doi: 10.3389/fphys.2022.1045469
48. Mohamed MFH, Al-Shokri SD, Shunnar KM, Mohamed SF, Najim MS, Ibrahim SI, et al. Prevalence of venous thromboembolism in critically ill COVID-19 patients: systematic review and meta-analysis. Front Cardiovasc Med. (2020) 7:598846. doi: 10.3389/fcvm.2020.598846
49. Fricke-Galindo I, Buendia-Roldan I, Chavez-Galan L, Pérez-Rubio G, Hernández-Zenteno RJ, Ramos-Martinez E, et al. SERPINE1 rs6092 variant is related to plasma coagulation proteins in patients with severe COVID-19 from a tertiary care hospital. Biol (Basel). (2022) 11(4):595. doi: 10.3390/biology11040595
50. Oh J, An HJ, Kim JO, Jun HH, Kim WR, Kim EJ, et al. Association between five common plasminogen activator inhibitor-1 (PAI-1) gene polymorphisms and colorectal cancer susceptibility. Int J Mol Sci. (2020) 21(12):4334. doi: 10.3390/ijms21124334
51. Rinde LB, Morelli VM, Småbrekke B, Mathiesen EB, Løchen ML, Njølstad I, et al. Effect of prothrombotic genotypes on the risk of venous thromboembolism in patients with and without ischemic stroke. Tromsø Study J Thromb Haemost. (2019) 17:749–58. doi: 10.1111/jth.14410
52. Skille H, Paulsen B, Hveem K, Gabrielsen ME, Brumpton B, Hindberg K, et al. Combined effects of five prothrombotic genotypes and cancer on the risk of a first venous thromboembolic event. J Thromb Haemost. (2020) 18:2861–9. doi: 10.1111/jth.15011
53. Salem N, Atallah B, El Nekidy WS, Sadik ZG, Park WM, Mallat J. Thromboelastography findings in critically ill COVID-19 patients. J Thromb Thrombolysis. (2021) 51:961–5. doi: 10.1007/s11239-020-02300-7
54. Ahmad S, Manzoor S, Siddiqui S, Mariappan N, Zafar I, Ahmad A, et al. Epigenetic underpinnings of inflammation: Connecting the dots between pulmonary diseases, lung cancer and COVID-19. Semin Cancer Biol. (2022) 83:384–98. doi: 10.1016/j.semcancer.2021.01.003
55. Sillen M, Declerck PJ. A narrative review on plasminogen activator inhibitor-1 and its (Patho)Physiological role: to target or not to target? Int J Mol Sci. (2021) 22(5):2721. doi: 10.3390/ijms22052721
56. Esmailian M, Vakili Z, Nasr-Esfahani M, Heydari F, Masoumi B. D-dimer levels in predicting severity of infection and outcome in patients with COVID-19. Tanaffos. (2022) 21:419–33.
57. Chen R, Liang W, Jiang M, Guan W, Zhan C, Wang T, et al. Risk factors of fatal outcome in hospitalized subjects with coronavirus disease 2019 from a nationwide analysis in China. Chest. (2020) 158:97–105. doi: 10.1016/j.chest.2020.04.010
58. Xie J, Covassin N, Fan Z, Singh P, Gao W, Li G, et al. Association between hypoxemia and mortality in patients with COVID-19. Mayo Clin Proc. (2020) 95:1138–47. doi: 10.1016/j.mayocp.2020.04.006
59. Li Y, Deng Y, Ye L, Sun H, Du S, Huang H, et al. Clinical significance of plasma D-dimer in COVID-19 mortality. Front Med (Lausanne). (2021) 8:638097. doi: 10.3389/fmed.2021.638097
60. Lange LA, Reiner AP, Carty CL, Jenny NS, Cushman M, Lange EM. Common genetic variants associated with plasma fibrin D-dimer concentration in older European- and African-American adults. J Thromb Haemost. (2008) 6:654–9. doi: 10.1111/j.1538-7836.2008.02906.x
61. Dittmann M, Hoffmann HH, Scull MA, Gilmore RH, Bell KL, Ciancanelli M, et al. A serpin shapes the extracellular environment to prevent influenza A virus maturation. Cell. (2015) 160:631–43. doi: 10.1016/j.cell.2015.01.040
Keywords: COVID-19, SERPINE1, TMPRSS2, Polymorphism, SARS-CoV-2
Citation: Martínez-Gómez LE, Martinez-Armenta C, Tusie-Luna T, Vázquez-Cárdenas P, Vidal-Vázquez RP, Ramírez-Hinojosa JP, Gómez-Martín D, Vargas-Alarcón G, Posadas-Sánchez R, Fragoso JM, de la Peña A, Rodríguez-Pérez JM, Mata-Miranda MM, Vázquez-Zapién GJ, Martínez-Cuazitl A, Martínez-Ruiz FdJ, Zayago-Angeles DM, Ramos-Tavera L, Méndez-Aguilera A, Camacho-Rea MdC, Ordoñez-Sánchez ML, Segura-Kato Y, Suarez-Ahedo C, Olea-Torres J, Herrera-López B, Pineda C, Martínez-Nava GA and López-Reyes A (2024) The fatal contribution of serine protease-related genetic variants to COVID-19 outcomes. Front. Immunol. 15:1335963. doi: 10.3389/fimmu.2024.1335963
Received: 13 November 2023; Accepted: 14 March 2024;
Published: 27 March 2024.
Edited by:
Maria Antonia De Francesco, University of Brescia, ItalyReviewed by:
Georgios S. Markopoulos, University of Ioannina, GreeceFlora De Conto, University of Parma, Italy
Copyright © 2024 Martínez-Gómez, Martinez-Armenta, Tusie-Luna, Vázquez-Cárdenas, Vidal-Vázquez, Ramírez-Hinojosa, Gómez-Martín, Vargas-Alarcón, Posadas-Sánchez, Fragoso, de la Peña, Rodríguez-Pérez, Mata-Miranda, Vázquez-Zapién, Martínez-Cuazitl, Martínez-Ruiz, Zayago-Angeles, Ramos-Tavera, Méndez-Aguilera, Camacho-Rea, Ordoñez-Sánchez, Segura-Kato, Suarez-Ahedo, Olea-Torres, Herrera-López, Pineda, Martínez-Nava and López-Reyes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alberto López-Reyes, YWxsb3JleUB5YWhvby5jb20=; Gabriela A. Martínez-Nava, YW1lcmlhLmp1c3RpY2VAZ21haWwuY29t