 Breno Luiz Pimenta1,2
Breno Luiz Pimenta1,2 Daniela Pagliara Lage1Camila Simões de Freitas1Danniele Luciana Vale1Karolina Oliveira de Melo Falcão3Saulo Samuel Gonçalves Dias3Raquel Soares Bandeira Câmara1Isabela Amorim Gonçalves Pereira1Ana Luiza Silva1Lauro de Assis Duarte Júnior2,4
Daniela Pagliara Lage1Camila Simões de Freitas1Danniele Luciana Vale1Karolina Oliveira de Melo Falcão3Saulo Samuel Gonçalves Dias3Raquel Soares Bandeira Câmara1Isabela Amorim Gonçalves Pereira1Ana Luiza Silva1Lauro de Assis Duarte Júnior2,4 Miguel Angel Chavez-Fumagalli5Rodrigo Fernando Bianchi4
Miguel Angel Chavez-Fumagalli5Rodrigo Fernando Bianchi4 Eduardo Antonio Ferraz Coelho1,3*
Eduardo Antonio Ferraz Coelho1,3* André Talvani1,2*
André Talvani1,2*- 1Programa de Pós-Graduação em Ciências da Saúde: Infectologia e Medicina Tropical, Faculdade de Medicina, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil
- 2Laboratório de Imunobiologia da Inflamação, Departamento de Ciências Biológicas, Instituto de Ciências Exatas e Biológicas (ICEB), Universidade Federal de Ouro Preto, Ouro Preto, Brazil
- 3Departamento de Patologia Clínica, Colégio Técnico da Universidade Federal de Minas Gerais, Belo Horizonte, Brazil
- 4Laboratório de Polímeros e Propriedades Eletrônicas de Materiais, Instituto de Ciências Exatas e Biológicas (ICEB), Universidade Federal de Ouro Preto, Ouro Preto, Brazil
- 5Computational Biology and Chemistry Research Group, Vicerrectorado de Investigación, Universidad Católica de Santa María, Arequipa, Peru
The treatment of tegumentary leishmaniasis (TL) faces significant challenges, including drug toxicity, high costs, and the emergence of resistant strains. These limitations highlight the urgent need for novel antileishmanial agents and therapeutic strategies. This study evaluated blue light-emitting diode (LED) phototherapy as an alternative approach to inhibit Leishmania stationary promastigotes and treat infected mammalian models. In vitro assays using Leishmania amazonensis, L. braziliensis, and L. infantum demonstrated that blue LED significantly inhibited parasite growth during and after treatment, with inhibition levels comparable to those achieved with amphotericin B (AmpB). Treatment of infected macrophages with blue LED substantially reduced infection rates and amastigote recovery across all three parasite species. Ultrastructural analyses revealed the destruction of internal organelles and alterations to the surface membranes of all Leishmania species following blue LED exposure. In in vivo experiments, L. amazonensis-infected BALB/c mice were treated with AmpB, blue LED alone, combination of blue LED plus AmpB, or saline as a control. Animals treated with blue LED, particularly in combination with AmpB, exhibited significant reductions in parasite loads in infected tissues such as lesions, spleens, livers, and draining lymph nodes, as confirmed by limiting dilution assays and qPCR. Additionally, these treatments induced a robust antileishmanial Th1-type immune response, characterized by increased production of IFN-γ, IL-12, nitrite, and IgG2a antibodies. These findings suggest that blue LED phototherapy holds promise as a potential therapeutic strategy for TL and warrants further investigation in future studies.
1 Introduction
Leishmaniases are endemic diseases caused by more than 20 species of Leishmania parasites (1), which are distributed geographically across the world with different species able to cause distinct clinical manifestations ranging from self-limiting cutaneous lesions to life-threatening visceral disease (2). The infection outcome is mainly determined by factors related to parasite infectivity, vector biology, host immune responses and nutritional status (3). In this context, the main clinical forms of the disease are tegumentary leishmaniasis (TL) and visceral leishmaniasis (VL) (4); where TL is presented in the cutaneous, diffuse cutaneous and mucosal forms, being characterized by one or more lesions on the skin, which can cause destruction of the mucosa and cartilage leading to respiratory compromise in the patient (5). It can be caused by several parasite species, such as Leishmania braziliensis, L. major, L. amazonensis, L. tropica, and others. VL is the most serious form of the disease and can lead to death, causing symptoms such as fever, anorexia, diarrhea, hepatosplenomegaly, lymphadenopathy, and vascular problems (6).
Treatment for the disease is based on the use of pentavalent antimonials, free or liposomal amphotericin B (AmpB), pentamidine, miltefosine, among others; although they cause organ toxicity, are high in cost and/or lead to the emergence of resistant strains (7, 8). Given these limitations, there is a pressing need to develop safe and alternative approaches to conventional treatment. Besides new and promising synthetic molecules, delivery systems and therapeutic protocols, photodynamic therapy based on light interaction at a suitable wavelength with a photosensitive antileishmanial compound and oxygen can trigger a photochemical reaction and generate reactive oxygen species leading to cell death (9, 10). This strategy has been used as an adjuvant therapy to treat pulmonary, respiratory tract, neural, and urinary tract tumors, as well as vitreoretinal disease (11, 12). Different compounds, such as carbaporphyrin ketals, methylene blue, aminolevulinic acid, chloroaluminum phthalocyanine, and aluminum phthalocyanine tetrasulfonate have been employed as photosensitizers, because the investigation of new photosensitizers is important to improve the effectiveness of the therapy. Photodynamic therapy (PDT) has also emerged as an alternative for the treatment of TL, with studies on its effectiveness being conducted in humans and animal models (13, 14). In another promising experimental strategy, blue light-emitting diode (LED) phototherapy has also been proposed as a treatment for diseases (15, 16). Blue light is part of the natural light received from the sun, which has been shown to inactivate microorganisms such as bacteria and fungi (17, 18). The proposed action is similar to PDT, since it is based on the photoexcitation of endogenous porphyrins that increase the level of reactive oxygen species (19), causing damage to proteins and lipids in the microorganism’s membrane, thereby affecting intracellular transport (20). An additional effect of blue light is photo-immunomodulation because light, in general, affects the release of hormones and cytokines (21).
In a recent study, Ivanova et al. (22) proposed an innovative approach to controlling Trypanosoma cruzi infection using blue LED phototherapy. In vitro assays with axenic cultures of Y and CL strains of T. cruzi demonstrated a 50% reduction in epimastigote replication after five days of blue light exposure. In vivo experiments in C57BL/6 mice infected with the Y strain of T. cruzi revealed that blue LED phototherapy reduced trypomastigote levels in blood and cardiac tissue. Additionally, it lowered plasma levels of IL-6, TNF, and IL-10 (but not CCL2), suggesting a light-mediated parasite control. The authors claimed their study was the first to explore blue LED phototherapy for T. cruzi infection. However, no studies have evaluated this therapy for treating tegumentary leishmaniasis (TL).
So, the present study investigated the antileishmanial effects of blue LED phototherapy, both alone and in combination with amphotericin B (AmpB), against stationary-phase promastigotes of L. infantum, L. braziliensis, and L. amazonensis. The effects were also evaluated in vitro (axenic and peritoneal macrophage cultures) and in vivo (BALB/c mice). Additionally, the study assessed how chemotherapy and phototherapy influenced parasite ultrastructure and modulated the mammalian immune response.
2 Materials and methods
2.1 Ethical committee on animal research
The study was approved by the Ethical Committee on Animal Research (CEUA) of the Federal University of Minas Gerais (UFMG; Belo Horizonte, Minas Gerais, Brazil), with the protocol number 056/2022. Female BALB/c mice, aged 8 weeks, were obtained from the Center for Animal Facilities at the Institute of Biological Sciences of UFMG, and were kept under pathogen-free conditions.
2.2 In vitro activity against distinct Leishmania species
L. amazonensis (IFLA/BR/1967/PH-8), L. braziliensis (MHOM/BR/1975/M2904) and L. infantum (MHOM/BR/1970/BH46) were used. Stationary-phase promastigotes were cultured in complete Schneider’s medium (Sigma-Aldrich, USA), which was supplemented with 20% heat-inactivated fetal bovine serum (FBS; Sigma-Aldrich, USA), 20 mM L-glutamine, 200 U/mL penicillin, and 100 μg/mL streptomycin, at pH 7.4, at 24°C (23). All technical procedures were carried out under sterile conditions. Parasites (1 x 106 cells/mL) were plated in complete Schneider’s medium with a final volume of 500 μL per well and submitted to blue LED phototherapy for 12 h per day over 5 days at 24°C. Daily, the emitted irradiance was checked with a radiometer to ensure consistency throughout the treatment. Some wells were untreated (background control) or treated with AmpB (Cat Y0001361 – European Pharmacopoeia. 0.1 μg/mL/well, 1 day at 24°C). Cell viability was assessed daily by counting viable parasites in a Neubauer chamber over five days (12h/day), as trypanosomatids exhibit partial resistance to phototherapy in the absence of mammalian immune cells (22).
2.3 Transmission electron microscopy
Parasite ultrastructural alterations were evaluated using transmission electron microscopy (TEM). Samples from the three Leishmania species, previously exposed to white and blue light for 5 days (12 hours/day), were fixed in a solution containing 2.5% glutaraldehyde, 4% paraformaldehyde, and 0.1 M phosphate buffer. The samples were then adhered to glass slides coated with 0.1% poly-L-lysine for 30 minutes at 37°C.
After fixation, the slides were washed twice with 0.1 M phosphate buffer, and postfixed in osmium tetroxide (OsO4) solution for 1 hour at room temperature, then washed again twice with 0.1 M phosphate buffer. The samples were gradually dehydrated through an ethanol series and analyzed using a Tecnai G2-12 FEI Spirit Biotwin 120 kV electron microscope at the Centro de Microscopia, UFMG, Belo Horizonte, Brazil.
2.4 Treatment using blue LED light phototherapy in Leishmania-infected macrophages
Treatment of infected murine peritoneal macrophages was performed by culturing cells (5 x 105 per well) in complete RPMI 1640 medium (Sigma-Aldrich, USA), which was supplemented with 20% heat-inactivated fetal bovine serum (FBS; Sigma-Aldrich, USA), 20 mM L-glutamine, 200 U/mL penicillin, and 100 μg/mL streptomycin, for 2 h at 37°C with 5% (v/v) CO2. Next, non-adherent macrophages were removed by washing with a complete RPMI 1640 medium. L. amazonensis, L. braziliensis and L. infantum stationary-phase promastigotes (at a ratio of 10 parasites per macrophage) were added, and a new incubation was carried out for 24 h at 37°C with 5% CO2. Parasites were washed to remove non-adherent or non-phagocytized ones, and infected macrophages were treated with blue LED phototherapy or AmpB (1.0 μg/mL/well) for 24 or 48 h, at 37°C. Since parasites within peritoneal macrophages are subjected to immune pressure, the blue LED phototherapy protocol was adjusted accordingly. Unlike in axenic cultures, phototherapy was applied continuously for 24 or 48 h. After incubation, cells were stained using the panoptic method and, the percentage of infected macrophages and the number of intracellular amastigotes were determined by counting 200 cells per cover glass in triplicate under an optical microscope.
2.5 In vivo infection and treatment schedules
Two independent experiments were performed, and similar results were obtained. One representative experiment is shown in this work. BALB/c mice (n=4 per group) were infected subcutaneously on the dorsum with 106 L. amazonensis stationary-phase promastigotes. After lesion development (at 50 to 60 days after infection), treatment was initiated with animals exposed to blue LED phototherapy for 12 h per day over 10 days, with the light positioned above the cage, directed onto the lesion. The phototherapy device was custom-built by an electrical engineer in our laboratory. It consists of a wooden box equipped with four blue LED lights positioned at the top, 30 cm above the polypropylene cages containing the mice. The intensity (7 μW/cm2) of blue light was verified daily with a portable radiometer, inside each cage, to guarantee uniform radiance for all animals. Each device was connected to an electrical power source, which accommodated four cages simultaneously. The animals remained in their usual confined environment within the cages, with food pellets provided inside to minimize any resistance to light exposure. Additionally, cooling fans were installed at the top of the device to prevent temperature increases inside the box, despite the room being temperature-controlled. A 24-hour surveillance camera was also placed on top of the device to monitor the animals’ environment and detect any potential disturbances.
Mice treated with AmpB (1.0 mg/kg/dose) or receiving saline were handled through intraperitoneal injections. The study design was as follows: (i) control group: uninfected mice received blue LED phototherapy for 12 h per day for 10 days; (ii) saline group: mice were infected and received 100 μL PBS 1x in 5 doses with one-day intervals; (iii) blue LED phototherapy group: mice were infected and treated with blue LED phototherapy for 12 h per day for 10 days; (iv) AmpB group: mice were infected and treated with AmpB (1 mg/kg/dose in 5 doses with one-day intervals); (v) blue LED phototherapy plus AmpB group: mice were infected and treated with blue LED phototherapy for 12 h per day for 10 days and AmpB (1 mg/kg/dose in 5 doses with one-day intervals). The development of nodules, metastasis and other clinical signs were monitored during and after treatment.
2.6 Cellular response and nitrite secretion
Cytokine production was evaluated in splenic cultures from infected and treated animals, as previously described (24). Briefly, splenic cell suspensions (1 x 106 cells/well) were plated in duplicate in 24-well plates (Nunc) and incubated in complete RPMI 1640 (Sigma-Aldrich, USA). Cells were either unstimulated (medium) or stimulated with soluble Leishmania antigen - SLA (25 μg/mL) for 48 h at 37°C in 5% CO2. IFN-γ, IL-4, IL-10 and IL-12p70 levels were measured in the culture supernatant by capture ELISA using commercial kits (BD Pharmingen®, San Diego, CA, USA), according to manufacturer’s instructions. Nitrite production was assessed in the same culture supernatant, as previously described (25).
2.7 Antibody production evaluation
The humoral response was evaluated through the measurement of levels of total anti-SLA IgG and the ratio between IgG2a and IgG1 antibodies through an indirect ELISA, as previously described (24). After previous titration curves, SLA was added to the plates (1.0 μg per well), which were incubated for 16 h at 4°C. Then, 250 μL of a solution composed of 2% casein diluted in PBS-T (PBS 1x and 0.05% Tween 20) were added to the wells, and incubated for 1 h at 37°C. Plates were washed with PBS-T and mouse serum samples were added (1:50 diluted in PBS-T), and plates were again incubated for 1 h at 37°C. Plates were washed with PBS-T, and incubated with anti-mouse IgG total (goat anti-mouse IgG total secondary antibody, catalog SA1-31430, Invitrogen, USA), IgG1 (rat anti-mouse IgG1 secondary antibody, catalog SA1-35640, Invitrogen, USA) or IgG2a (rat anti-mouse IgG2a secondary antibody, catalog SA1-35646, Invitrogen, USA) antibodies. A new incubation was then performed for 1 h at 37°C, after which plates were washed with PBS-T and reactions were developed using H2O2, ortho-phenylenediamine, and a citrate-phosphate buffer pH 5.0 for 30 min in the dark. Next, they were stopped by adding 2N H2SO4 and optical density (OD) values were read in a spectrophotometer (Molecular Devices, Spectra Max Plus, Concord, Canada), at 492 nm.
2.8 Parasite load estimation
The parasite load was evaluated in the lesion, spleen, liver and draining lymph nodes (dLN) of infected and treated animals after therapy using a limiting dilution technique (26). Briefly, the skin lesions and organs were collected, weighed, and homogenized in a glass tissue grinder in sterile PBS 1x, and debris was removed by centrifugation at 150 x g. The suspension was plated in flat-bottom 96-well microplates (Nunc), and serial dilutions were made in complete Schneider’s medium from 10¹ to 10¹². Pipette tips were discarded after each dilution to prevent cross-contamination. Each sample was plated and incubated at 24°C and the presence of viable parasites was assessed seven days after culture initiation. Results were expressed as the negative log of the titer, which corresponded to the dilution in the last positive well, adjusted for tissue or organ weight. The L. amazonensis burden was also evaluated through a qPCR technique in skin lesions of infected mice, as previously described (27). Results were calculated by interpolation from a standard curve included in the same run, which was done in duplicate and expressed as the number of parasites per total DNA.
2.9 Statistical analysis
Results were analyzed using GraphPad Prism™ (version 6.0 for Windows), and one-way analysis of variance (ANOVA) and Student’s t-test were used for comparisons among groups. Differences were considered significant at P < 0.05. The experiment was performed twice, and similar results were obtained. The data shown are representative of one whole experiment.
3 Results
3.1 Blue LED phototherapy presents in vitro antileishmanial activity against distinct parasite species
We evaluated the in vitro activity of blue LED phototherapy against distinct Leishmania species, i.e., L. amazonensis, L. infantum and L. braziliensis, which received the treatment for 8 hours per day for 5 consecutive days. Controls were treated with AmpB. Results showed that exposure to natural white light (NL) for five days led to an increase in Leishmania numbers across all tested species. In contrast, blue LED phototherapy induced a peak in parasite replication on the third day, followed by a significant proportional reduction on the fourth and fifth days, as well as one day after the end of treatment. Interestingly, the antileishmanial efficacy of amphotericin B (AmpB) was more pronounced against L. amazonensis and L. braziliensis, but it also effectively reduced L. infantum numbers within the first three days of treatment in culture. However, when comparing chemotherapy and phototherapy over five days and one day post-treatment, blue LED phototherapy showed superior leishmanicidal effects (Table 1). Scanning electron micrographs of parasites revealed that exposure to white (natural) light for 5 days did not induce ultrastructural changes in L. amazonensis (Figure 1A), L. infantum (Figure 1C), or L. braziliensis (Figure 1E). In contrast, blue LED light therapy caused alterations in the surface membrane structure and disruption of intracellular organelles in all three species: L. amazonensis (Figure 1B), L. infantum (Figure 1D), and L. braziliensis (Figure 1F). Blue LED treatment applied for 24h in infected macrophages reduced the infection percentage by 42.4%, 66.8%, and 42.5% when L. amazonensis, L. infantum and L. braziliensis promastigotes were used, respectively, and by 50.6%, 80.7%, and 71.8%, respectively, after 48h of treatment (Table 2). Using AmpB, reductions were 34.6%, 71.8%, and 53.0%, respectively, after 24h of treatment; and 54.8%, 87.5%, and 73.5%, respectively, after 48h of treatment. Regarding the reduction in the number of recovered amastigotes, values were 47.0%, 75.6%, and 44.3%, respectively, when blue LED phototherapy was used for 24h in L. amazonensis, L. infantum and L. braziliensis, respectively; and 76.8%, 86.8%, and 89.5%, when treatment was performed by 48h, respectively. Using AmpB, reductions were 35.8%, 78.0%, and 76.0%, after 24h of treatment; and 78.8%, 93.4%, and 89.7%, when treatment was performed by 48h, respectively (Table 2).
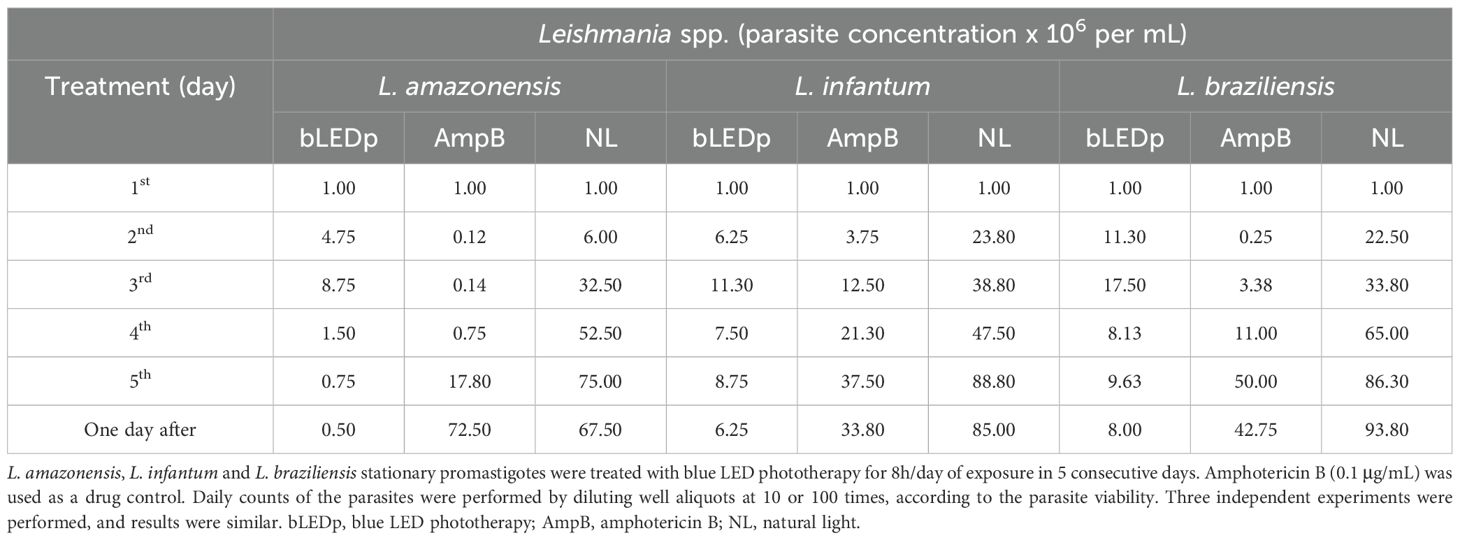
Table 1. In vitro antileishmanial activity after treatment with blue LED phototherapy.
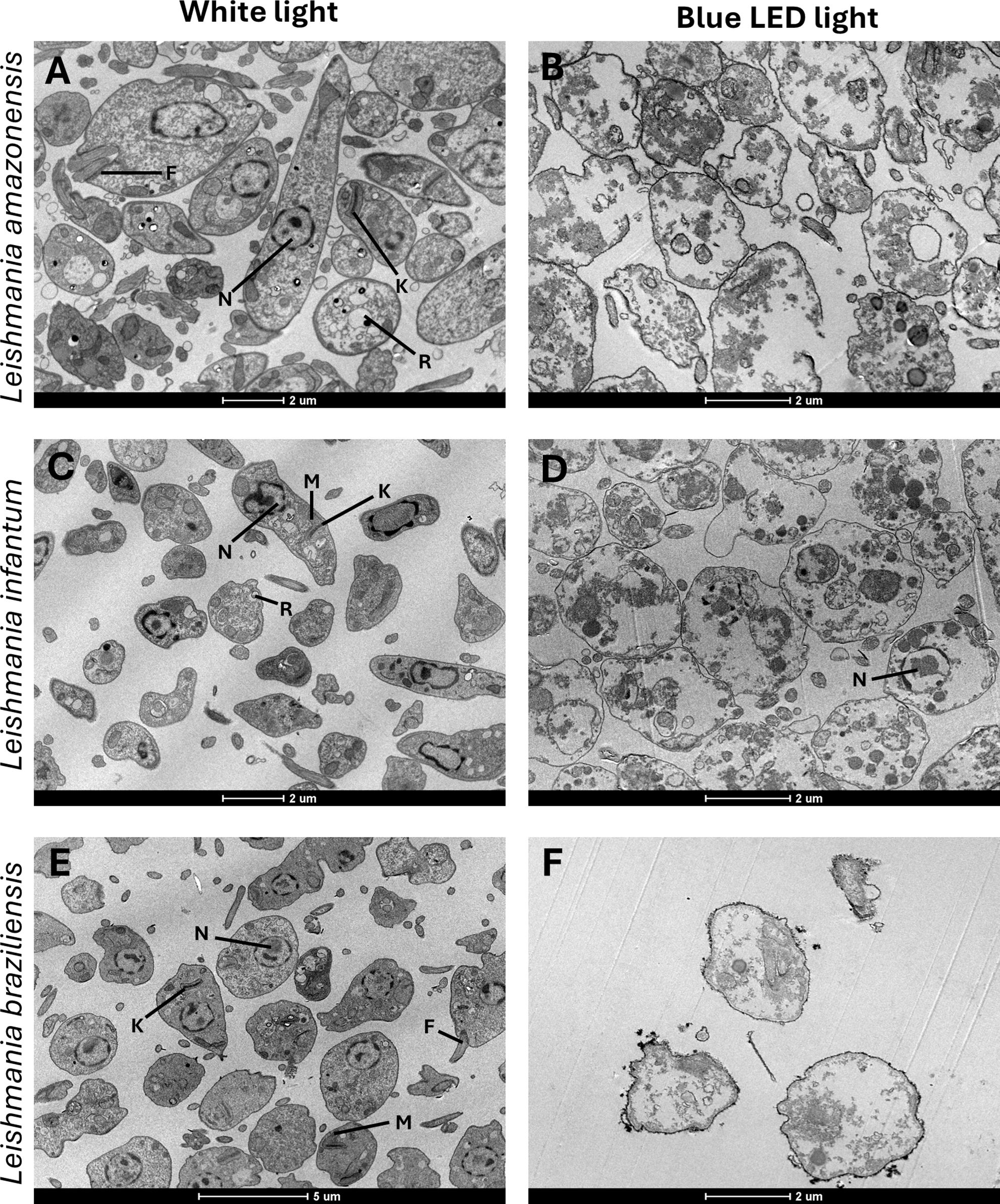
Figure 1. Ultrastructural comparison of Leishmania under white light and blue LED phototherapy conditions. The left column shows representative ultrastructural images of promastigotes of Leishmania amazonensis (IFLA/BR/1967/PH-8) (A), L. braziliensis (MHOM/BR/1975/M2904) (C), and L. infantum (MHOM/BR/1970/BH46) (E) cultured in Schneider’s medium at 24°C without treatment, under white light exposure. The right column depicts promastigotes of L. amazonensis (B), L. braziliensis (D), and L. infantum (F) following daily exposure to blue LED phototherapy (12 hours/day) for 5 days. Structural features include the flagellum (F), mitochondrion (M), reservosome (R), nucleus (N), and kinetoplast (K). Scale bars denote magnification (2um).
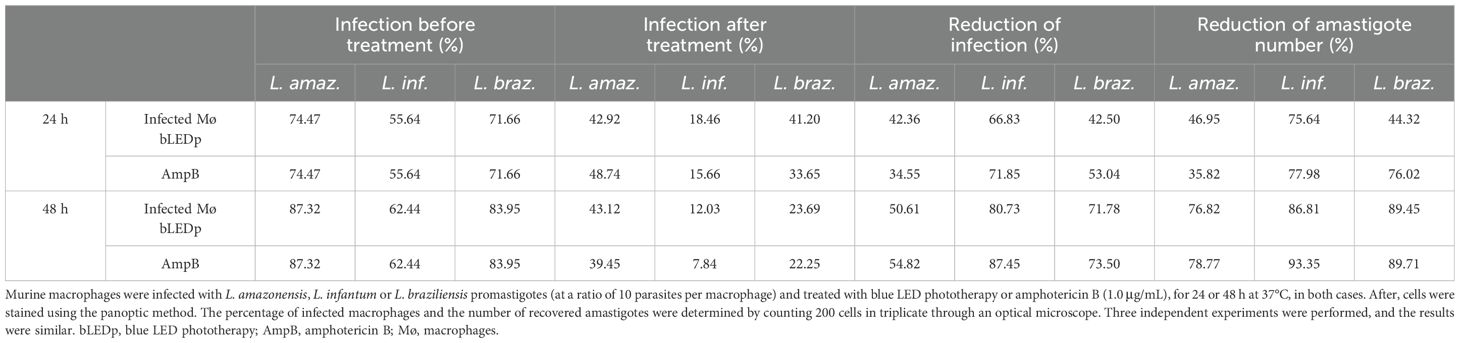
Table 2. Treatment of infected macrophages using blue LED phototherapy.
3.2 Blue LED phototherapy induces the development of a Th1-type immune response in treated and L. amazonensis-infected mice
We evaluated the therapeutic efficacy induced by blue LED phototherapy in L. amazonensis-infected mice. To do so, levels of Th1-and Th2-type cytokines were measured in the supernatant of SLA-stimulated spleen cell cultures, and results showed that treatment with blue LED induced the production of higher levels of IFN-γ and IL-12, which were associated with lower IL-4 and IL-10 levels (Figure 2A). A Th1-type response was found in mice treated with an association of blue LED and AmpB, when compared to values obtained in the other groups. To evaluate the parasite-specific activation of macrophages in the treated and infected animals, the nitrite secretion was investigated in the culture supernatant, and results corroborated those described for the cytokine levels, since the treatment with blue LED plus AmpB induced higher levels of antileishmanial nitrite compared to the others (Figure 2B). We also evaluated antibody production after treatment (Figure 3) and, mice receiving AmpB, blue LED and, mainly, blue LED plus AmpB produced higher levels of anti-SLA IgG2a antibodies compared to IgG1 levels. Otherwise, infected and untreated mice (saline group) produced higher levels of IgG1 antibodies, in comparison to IgG2a levels (Figure 3).
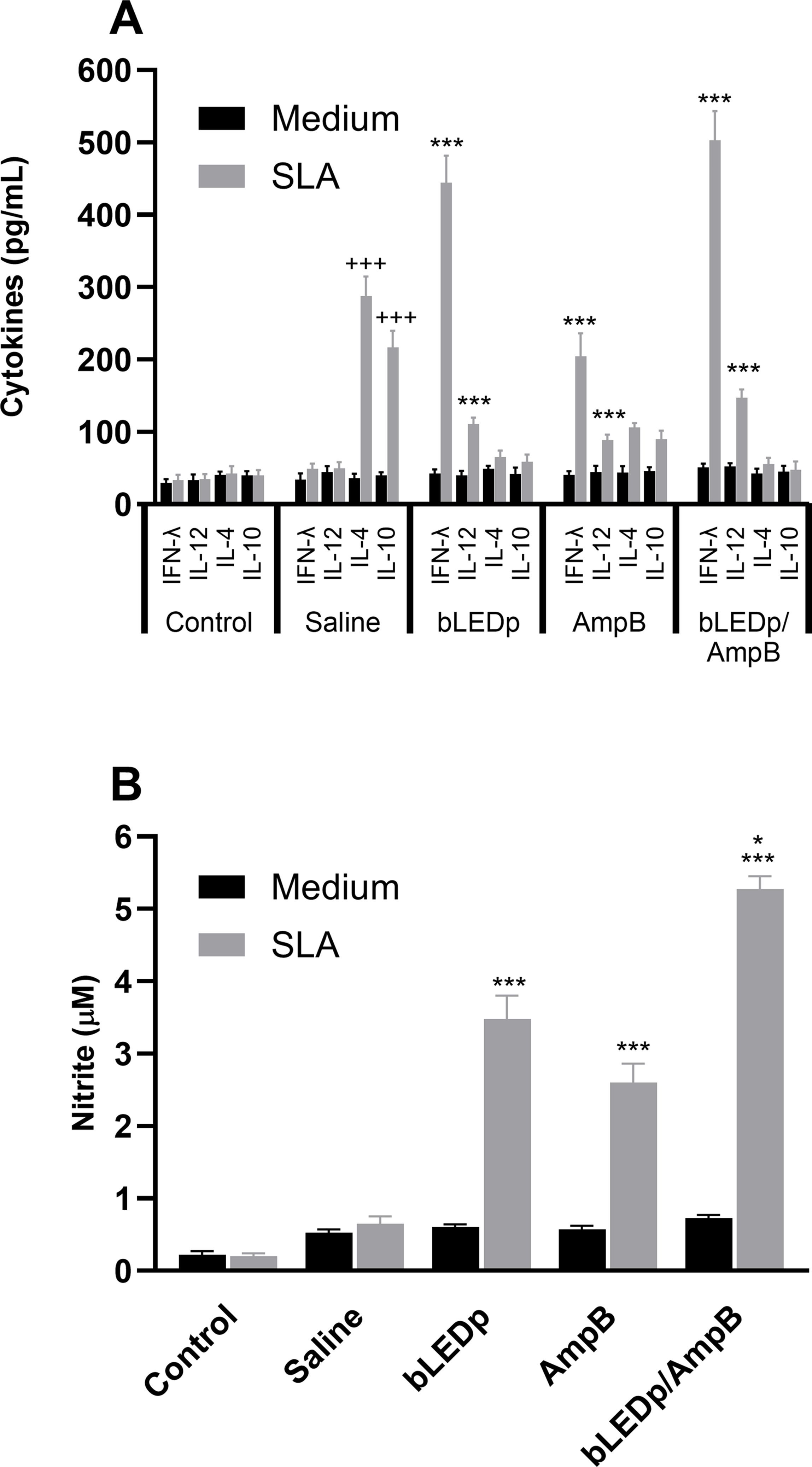
Figure 2. Cytokine production and nitrite secretion after treatment with blue LED phototherapy. Mice were infected with L. amazonensis promastigotes and treated with amphotericin B (AmpB), blue LED phototherapy (bLEDp) or associated with AmpB (bLEDp/AmpB), or they received only saline. Then, one day after treatment, animals were euthanized, and spleen cells were cultured in RPMI medium and stimulated with soluble Leishmania antigen (SLA) for 48 h at 37°C at 5% CO2. Levels of IFN-γ, IL-4, IL-10 and IL-12p70 cytokines were measured in culture supernatant through a capture ELISA, and results are shown (A). The same cell supernatant was used to measure the levels of antileishmanial nitrite, and results are also shown (B). Two independent experiments were performed, and bars indicate the mean ± standard deviation of the groups. (*) indicates a statistically significant difference between the bLEDp and AmpB groups (P < 0.05). (***) indicates a statistically significant difference between the control and saline groups (P < 0.001). (+++) indicates a statistically significant difference among bLEDp, AmpB and bLEDp/AmpB groups (P < 0.05).
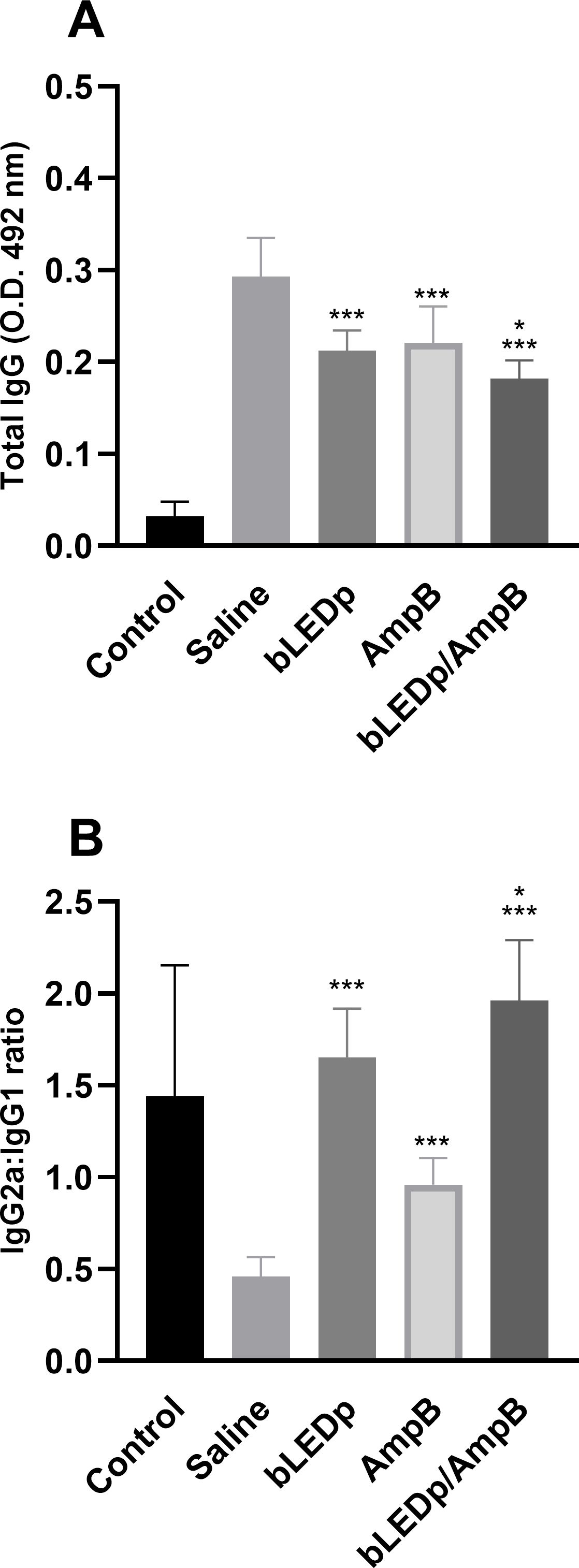
Figure 3. Antibody production after treatment with blue LED phototherapy. Mice were infected with L. amazonensis promastigotes and treated with saline, amphotericin B (AmpB), blue LED phototherapy (bLEDp) or in association with AmpB (bLEDp/AmpB. One day after treatment, sera samples were collected and used to evaluate the production of anti-soluble Leishmania antigen (SLA) IgG total (A) and the IgG2a/IgG1 ratio (B) antibodies, through an indirect ELISA. Two independent experiments were performed, and bars indicate the mean ± standard deviation of the groups. (*) indicates a statistically significant difference in the bLEDp and AmpB groups (P < 0.05). (***) indicates a statistically significant difference in the control and saline groups (P < 0.001).
3.3 Blue LED phototherapy induces a reduction in the L. amazonensis burden after treatment of infected mice
The parasite burden was evaluated in livers, spleens, dLNs and skin lesions of treated and infected animals, and results showed significant reductions in parasitism in mice receiving blue LED alone or in combination with AmpB, compared to values found in the saline group mice, with reductions of 4.0- and 4.8-log, respectively, in their lesions; 1.8- and 2.3-log, respectively, in their livers; 2.3- and 2.7-log, respectively, in their spleens; and 3.2- and 4.0-log, respectively, in their dLNs (Figure 4). A qPCR assay was also performed on the skin lesions, and results showed that mice receiving blue LED associated with AmpB presented greater reductions in parasite load, of 67.0% and 74.0%, respectively, compared to data described for the saline group (Figure 5).
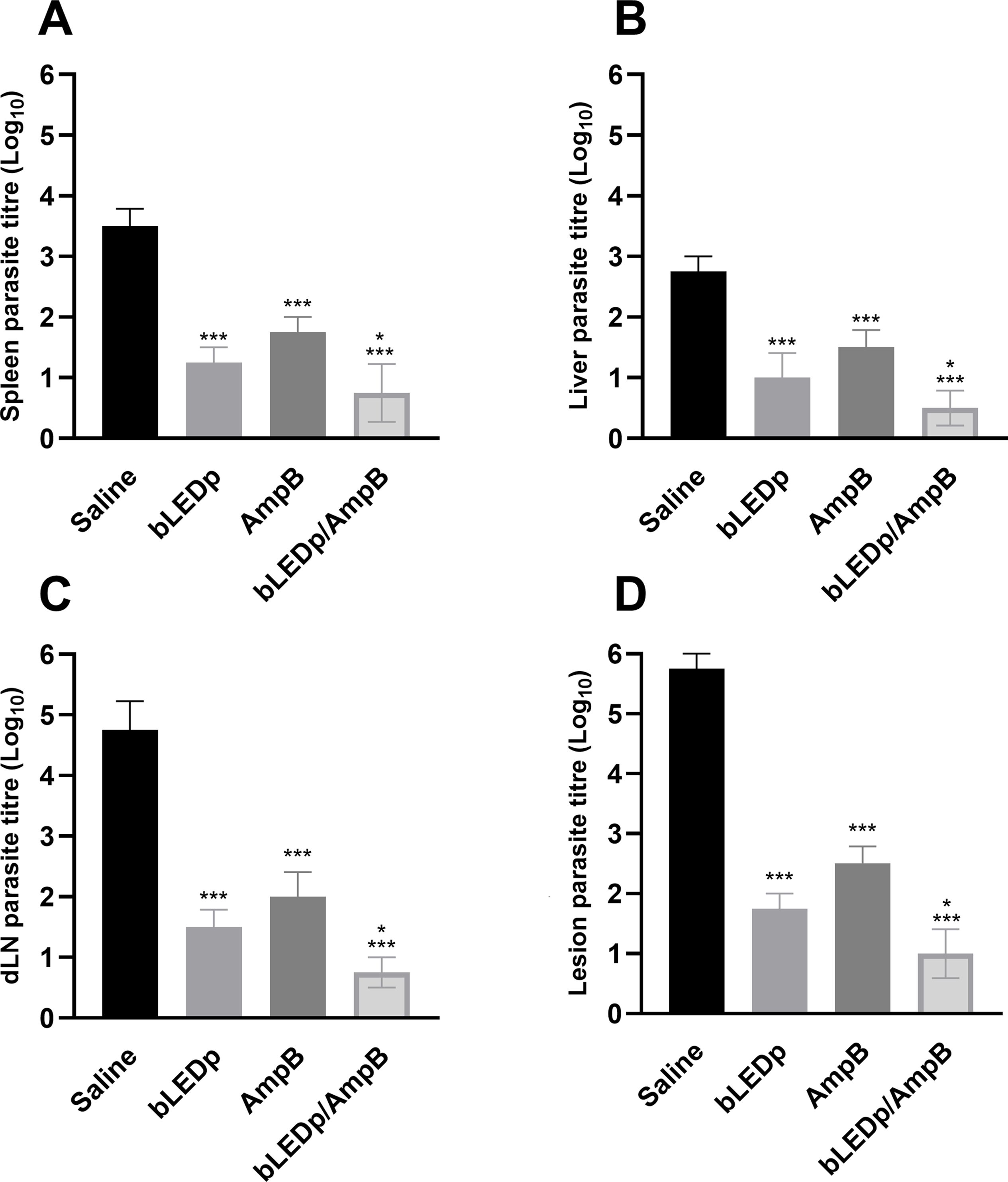
Figure 4. Leishmania amazonensis burden evaluated after blue LED phototherapy by a limiting dilution technique. L. amazonensis-infected mice were treated with amphotericin B (AmpB), blue LED phototherapy (bLEDp) alone or associated with AmpB (bLEDp/AmpB). Other animals were infected and received only saline. Three days after treatment, animals were euthanized and spleen (A), liver (B), draining lymph node (dLN) (C), and skin lesion (D) were collected to evaluate the parasite load, through a limiting dilution technique (A). In addition, their skin lesions were used in a qPCR assay to estimate the parasite load, with results being normalized by number of parasites per total DNA. Two independent experiments were performed, and bars indicate the mean ± standard deviation of the groups. (*) indicates a statistically significant difference between bLEDp and AmpB groups (P < 0.05). (***) indicates a statistically significant difference in the control and saline groups (P < 0.001).
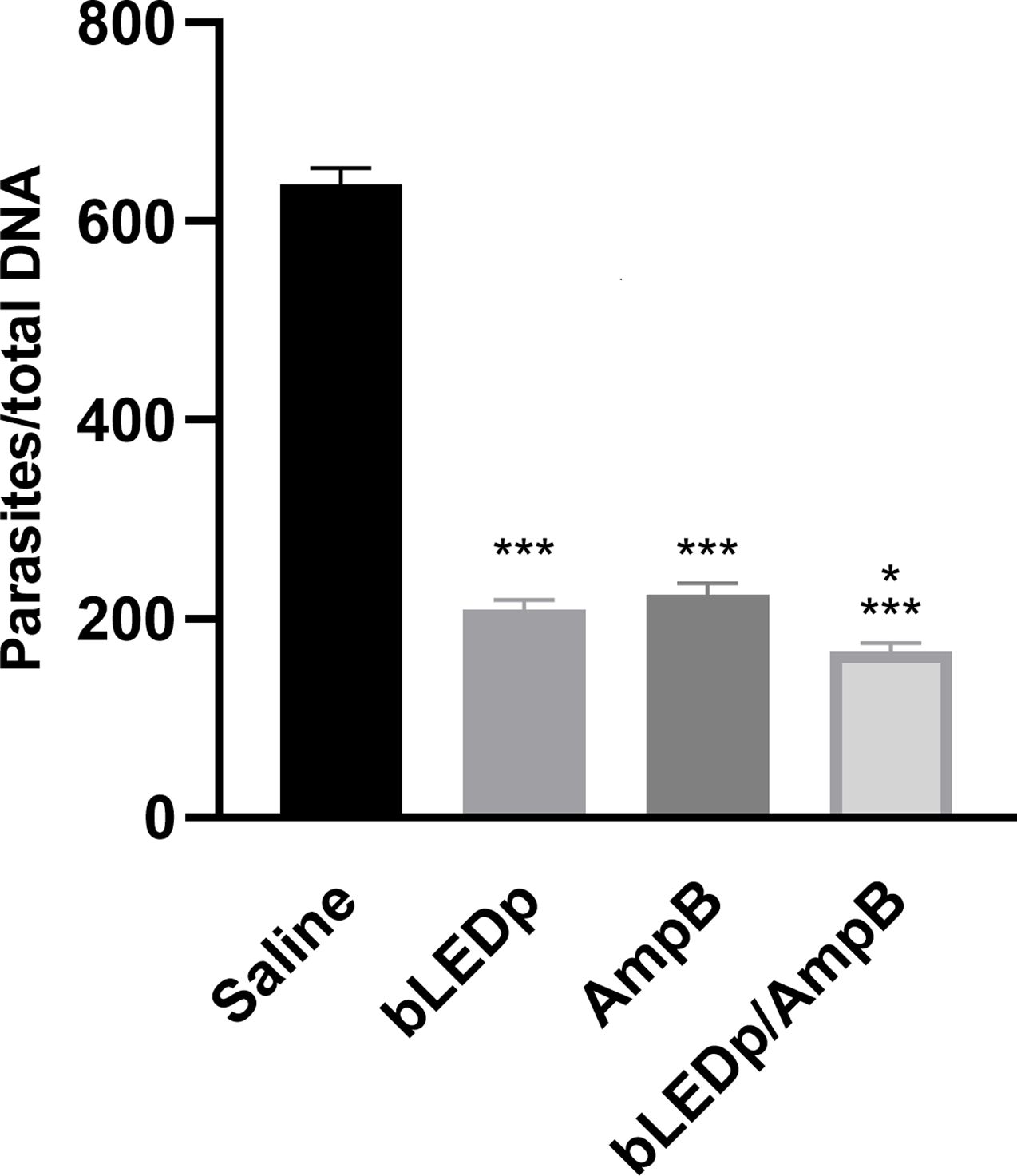
Figure 5. Skin lesion parasitism estimated through a qPCR assay. L. amazonensis-infected mice received saline or were treated with blue LED phototherapy (bLEDp) alone, with amphotericin B (AmpB), or with association of both treatments (bLEDp/AmpB). One day after treatment, animals were euthanized, and skin lesions were collected, macerated, and used to estimate the parasite load through a qPCR technique. Results were normalized by number of parasites per total DNA. Two independent experiments were performed, and bars indicate the mean plus standard deviation of the groups. (*) indicates a statistically significant difference in the bLEDp and AmpB groups (P < 0.05). (***) indicates a statistically significant difference in the control and saline groups (P < 0.001).
4 Discussion
Treatment for leishmaniasis faces significant challenges, including drug toxicity, high costs, and the emergence of drug-resistant strains (28). In this context, the discovery of new antileishmanial targets and strategies is urgently needed (7). In the present study, we explored blue LED phototherapy as a novel antileishmanial approach through in vitro and in vivo experiments. The therapy proved effective in reducing parasite viability in in vitro cultures and decreasing parasite burden in infected macrophages. Additionally, in vivo experiments, blue LED phototherapy helped Leishmania-infected mice control the infection and develop a therapeutic phenotype, offering protection against the disease. To the best of our knowledge, this is the first study to demonstrate the efficacy of blue LED phototherapy against Leishmania spp.
Our initial experiments evaluated the antileishmanial effects of blue LED phototherapy in vitro by applying the therapy directly to parasite cultures at a single frequency without photosensitizing molecules. A significant reduction in parasite viability was observed after three days of treatment, suggesting a cumulative effect (29, 30). Similar findings were reported by Ivanova et al. (22), who showed that blue LED light inhibited T. cruzi cultures. The exposure of microorganisms - including bacteria, protozoa, and viruses - to distinct light wavelengths has demonstrated efficacy in inactivating these pathogens, supporting their potential for managing infectious diseases (19, 31).
The mechanisms underlying blue LED phototherapy’s effects remain under investigation. One proposed mechanism involves intracellular photosensitizing chromophores, which generate reactive oxygen species (ROS), leading to pathogen death (32, 33). Another mechanism suggests disruption of lipid and protein structures, impairing intracellular transport and organelle organization (20, 34). Additionally, blue LED light may modulate the host’s immune response, further contributing to its therapeutic effects (35, 36). Notably, the biological effects of blue LED phototherapy can vary across microorganisms due to differences in environmental conditions, light wavelength, and exposure duration (37, 38).
Following promising in vitro results, we selected L. amazonensis for the in vivo experiments, as it is a well-established model for cutaneous leishmaniasis (CL) and generates visible lesions at the site of infection, which is crucial for evaluating phototherapy’s effects. The ability to directly target the lesions with blue LED light is an advantage when studying therapeutic strategies. While L. braziliensis also causes cutaneous leishmaniasis, we opted to focus on L. amazonensis for the in vivo experiments due to the limitations of the BALB/c mouse model. BALB/c mice are not typically susceptible to L. braziliensis, as they do not develop significant lesions and tend to spontaneously resolve the infection, which would not allow for meaningful evaluation of phototherapy’s impact (39, 40).
Leishmania infections in mammals are often associated with the development of a Th2-type immune response. This response is characterized by the production of cytokines such as IL-4, IL-5, IL-6, IL-10, and IL-13, which deactivate infected macrophages and allow parasite dissemination at infection sites and internal organs (41, 42). The Th2 cytokine environment promotes the recruitment of anti-inflammatory cells, facilitating parasite persistence. In contrast, a Th1-type response - mediated by cytokines such as IFN-γ, IL-2, GM-CSF, and IL-12 - induces inflammation and enhances parasite elimination (43, 44).
In our study, blue LED phototherapy increased levels of pro-inflammatory cytokines IFN-γ and IL-12, which synergized with nitric oxide production to eliminate intracellular parasites (27, 45). In contrast, control group mice exhibited elevated levels of IL-4 and IL-10, which suppressed macrophage activation and promoted disease progression (46). The immunostimulatory effects of blue LED phototherapy likely contributed to the reduced parasite burden in treated animals, as evidenced by lower IL-10 levels and increased IFN-γ and IL-12 production. This suggests that blue LED phototherapy not only exerts direct antileishmanial effects but also modulates the host immune response, offering a dual mechanism for controlling the infection.
Distinct Leishmania species are responsible for various clinical manifestations of leishmaniasis. For example, species such as L. major, L. braziliensis, L. amazonensis, and L. tropica cause cutaneous leishmaniasis (CL), while L. donovani and L. infantum are associated with visceral leishmaniasis (VL) (47). In our study, we verified that blue LED phototherapy exhibited antileishmanial activity against three Leishmania species, significantly reducing cell viability in vitro experiments. The leishmanicidal effects of blue LED phototherapy were partially attributed to the disruption of intracellular organelles and cell membranes. This disruption impairs the parasites’ ability to regulate their metabolic environment, ultimately leading to cell death.
Although the current study did not investigate the precise mechanisms underlying these structural alterations, previous research on Staphylococcus aureus offers valuable insights. Blue light phototherapy has been shown to induce changes in membrane potential and stimulate free radical production via photo-acceptor molecules in bacteria (48). These findings may serve as a foundation for further exploration of similar mechanisms in Leishmania.
Currently, drugs such as pentavalent antimonials, miltefosine, pentamidine, and AmpB are commonly used to treat leishmaniasis; however, these treatments often cause significant toxic effects in patients (49). In our study, combining AmpB with blue LED phototherapy enhanced antileishmanial activity compared to AmpB alone. Similar synergistic effects have been reported in other studies, where AmpB was combined with immunogenic antigens in immunotherapeutic strategies tested in Leishmania-infected mice (26, 50, 51). Future studies may explore the potential of combining blue LED phototherapy with immunogenic candidates as part of such immunotherapeutic protocols.
To the best of our knowledge, this is the first study to demonstrate that blue LED phototherapy can control in vitro infections of Leishmania cultures following daily exposure. Additionally, our results indicate that this strategy may contribute to modulating the immune response in infected mice and reducing parasite burden in tissues and internal organs. While further research is needed to confirm the in vivo antileishmanial effects of blue LED phototherapy, the findings presented here provide proof-of-concept evidence supporting its potential use for the treatment of TL.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Ethical Committee in Animal Research (CEUA) from Federal University of Minas Gerais (UFMG; Belo Horizonte, Minas Gerais, Brazil), with protocol number 056/2022. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
BP: Conceptualization, Formal Analysis, Methodology, Resources, Visualization, Writing – original draft. DL: Investigation, Resources, Supervision, Validation, Writing – original draft. Cd: Formal Analysis, Investigation, Resources, Supervision, Writing – original draft. DV: Investigation, Methodology, Resources, Writing – original draft. KF: Formal Analysis, Investigation, Resources, Writing – review & editing. SD: Methodology, Resources, Writing – review & editing. RC: Writing – review & editing. IP: Writing – review & editing. AS: Methodology, Resources, Writing – review & editing. LJ: Methodology, Resources, Software, Writing – review & editing. MC: Investigation, Methodology, Writing – review & editing. RB: Conceptualization, Project administration, Supervision, Writing – review & editing. EC: Conceptualization, Funding acquisition, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing. AT: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Validation, Writing – review & editing, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by grants APQ-02167-21, APQ-01646-23, APQ-00720-23, RED-0067-23 and RED-00198-23 from the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG), as well as by the grants 402417/2023-2, 405946/2021-0 and 442782/2024-1 from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). The authors also thank the Brazilian agencies Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), FAPEMIG and CNPq for the scholarships. RB, EC and, AT express their gratitude to the CNPq for the research productivity fellowship.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. WHO. Leishmaniasis (2024). Available online at: https://www.who.int/health-topics/leishmaniasistab=tab_1 (Accessed March 25 2024).
2. Burza S, Croft SL, Boelaert M. Leishmaniasis. Lancet. (2018) 392:951–70. doi: 10.1016/S0140-6736(18)31204-2
3. Mathison BA, Bradley BT. Review of the clinical presentation, pathology, diagnosis, and treatment of Leishmaniasis. Lab Med. (2023) 54:363–71. doi: 10.1093/labmed/lmac134
4. Mann S, Frasca K, Scherrer S, Henao-Martínez AF, Newman S, Ramanan P, et al. A Review of leishmaniasis: current knowledge and future directions. Curr Trop Med Rep. (2021) 8:121–32. doi: 10.1007/s40475-021-00232-7
5. Torres-Guerrero E, Quintanilla-Cedillo MR, Ruiz-Esmenjaud J, Arenas R. Leishmaniasis: a review. F1000Res. (2017) 6:750. doi: 10.12688/f1000research
6. Costa CHN, Chang KP, Costa DL, Cunha FVM. From infection to death: an overview of the pathogenesis of visceral leishmaniasis. Pathogens. (2023) 12:969. doi: 10.3390/pathogens12070969
7. Shmueli M, Ben-Shimol S. Review of leishmaniasis treatment: can we see the forest through the trees? Pharm (Basel). (2024) 12:30. doi: 10.3390/pharmacy12010030
8. Sundar S, Singh VK, Agrawal N, Singh OP, Kumar R. Investigational new drugs for the treatment of leishmaniasis. Expert Opin Investig Drugs. (2024) 33:1029–46. doi: 10.1080/13543784.2024.2400139
9. Varzandeh M, Mohammadinejad R, Esmaeilzadeh-Salestani K, Dehshahri A, Zarrabi A, Aghaei-Afshar A. Photodynamic therapy for leishmaniasis: recent advances and future trends. Photod Photodyn Ther. (2021) 36:102609. doi: 10.1016/j.pdpdt.2021.102609
10. Vital-Fujii DG, Baptista MS. Progress in the photodynamic therapy treatment of Leishmaniasis. Braz J Med Biol Res. (2021) 54:e11570. doi: 10.1590/1414-431x2021e11570
11. Glisinski K, Kurman JS, Spandorfer A, Pastis NJ, Murgu S, Cheng GZ. Photodynamic therapy for the treatment of tracheobronchial papillomatosis: a multicenter experience. Photod Photodyn Ther. (2020) 30:101711. doi: 10.1016/j.pdpdt.2020.101711
12. Sen P, Manayath G, Shroff D, Salloju V, Dhar P. Polypoidal choroidal vasculopathy: an update on diagnosis and treatment. Clin Ophthalmol. (2023) 17:53–70. doi: 10.2147/OPTH.S385827
13. Pinto SML, Muehlmann LA, Ojeda LLM, Vera AM, Cordero MVR, Santos MFMA, et al. Nanoemulsions with Chloroaluminium Phthalocyanine and paromomycin for combined photodynamic and antibiotic therapy for cutaneous leishmaniasis. Infect Chemother. (2021) 53:342–54. doi: 10.3947/ic.2021.0010
14. Çalişkan E, Uncu HB, Akoğlu G. Cutaneous leishmaniasis treated with daylight methylene blue photodynamic therapy. Int J Dermatol. (2024) 63:e65–7. doi: 10.1111/ijd.16977
15. Lodi G, Del Re C, Nisticò SP, Bennardo L, Cannarozzo G, Sannino M. Blue light-emitting diodes for the treatment of localized vitiligo: a retrospective study. J Cosmet Dermatol. (2023) 22:1273–8. doi: 10.1111/jocd.15567
16. Takeuchi Y, Aoki A, Hiratsuka K, Chui C, Ichinose A, Aung N, et al. Application of different wavelengths of led lights in antimicrobial photodynamic therapy for the treatment of periodontal disease. Antibiotics (Basel). (2023) 12:1676. doi: 10.3390/antibiotics12121676
17. Trzaska WJ, Wrigley HE, Thwaite JE, May RC. Species-specific antifungal activity of blue light. Sci Rep. (2017) 7:4605. doi: 10.1038/s41598-017-05000-0
18. Lewis AJ, Zhang X, Griepentrog JE, Yuan D, Collage RD, Waltz PK, et al. Blue light enhances bacterial clearance and reduces organ injury during sepsis. Crit Care Med. (2018) 46:e779–87. doi: 10.1097/CCM.0000000000003190
19. Wu F, Yang M, Zhang J, Zhu S, Shi M, Wang K. Metalloporphyrin-indomethacin conjugates as new photosensitizers for photodynamic therapy. J Biol Inorg Chem. (2019) 24:53–60. doi: 10.1007/s00775-018-1626-9
20. Alves E, Santos N, Melo T, Maciel E, Dória ML, Faustino MA, et al. Photodynamic oxidation of Escherichia coli membrane phospholipids: new insights based on lipidomics. Rapid Commun Mass Spectrom. (2013) 27:2717–28. doi: 10.1002/rcm.v27.23
21. Yuan A, Huan W, Liu X, Zhang Z, Zhang Y, Wu J, et al. NIR light-activated drug release for synergetic chemo-photothermal therapy. Mol Pharm. (2017) 14:242–51. doi: 10.1021/acs.molpharmaceut.6b00820
22. Ivanova N, Leite ALJ, Vieira MB, Silva OHCE, Mota LWR, Costa GP, et al. New insights into blue light phototherapy in experimental Trypanosoma cruzi infection. Front Cell Infect Microbiol. (2021) 11:673070. doi: 10.3389/fcimb.2021.673070
23. Coelho EAF, Tavares CA, Carvalho FA, Chaves KF, Teixeira KN, Rodrigues RC, et al. Immune responses induced by the Leishmania (Leishmania) donovani A2 antigen, but not by the LACK antigen, are protective against experimental Leishmania (Leishmania) amazonensis infection. Inf Immun. (2003) 71:3988–94. doi: 10.1128/IAI.71.7.3988-3994.2003
24. Lage DP, Vale DL, Linhares FP, Freitas CS, MaChado AS, Cardoso JMO, et al. A recombinant chimeric protein-based vaccine containing T-cell epitopes from amastigote proteins and combined with distinct adjuvants, induces immunogenicity and protection against Leishmania infantum infection. Vaccines (Basel). (2022) 10:1146. doi: 10.3390/vaccines10071146
25. Talvani A, MaChado FS, Santana GC, Klein A, Barcelos L, Silva JS, et al. Leukotriene B(4) induces nitric oxide synthesis in Trypanosoma cruzi-infected murine macrophages and mediates resistance to infection. Infect Immun. (2002) 70:4247–53. doi: 10.1128/IAI.70.8.4247-4253.2002
26. Vale DL, Freitas CS, Martins VT, Moreira GJL, MaChado AS, Ramos FF, et al. Efficacy of an immunotherapy combining immunogenic chimeric protein plus adjuvant and amphotericin B against murine visceral leishmaniasis. Biol (Basel). (2023) 12:851. doi: 10.3390/biology12060851
27. MaChado AS, Lage DP, Vale DL, Freitas CS, Linhares FP, Cardoso JMO, et al. Leishmania LiHyC protein is immunogenic and induces protection against visceral leishmaniasis. Par Immunol. (2022) 44:e12921. doi: 10.1111/pim.12921
28. Chakravarty J, Sundar S. Current and emerging medications for the treatment of leishmaniasis. Expert Opin Pharmacother. (2019) 20:1251–65. doi: 10.1080/14656566.2019.1609940
29. Sankhuan D, Niramolyanun G, Kangwanrangsan N, Nakano M, Supaibulwatana K. Variation in terpenoids in leaves of Artemisia annua grown under different LED spectra resulting in diverse antimalarial activities against Plasmodium falciparum. BMC Plant Biol. (2022) 22:128. doi: 10.1186/s12870-022-03528-6
30. Cabral FV, Cerone M, Persheyev S, Lian C, Samuel IDW, Ribeiro MS, et al. New insights in photodynamic inactivation of Leishmania amazonensis: a focus on lipidomics and resistance. PLoS One. (2023) 18:e0289492. doi: 10.1371/journal.pone.0289492
31. Jager TL, Cockrell AE, Du Plessis SS. Ultraviolet light induced generation of reactive oxygen species. Adv Exp Med Biol. (2017) 996:15–23. doi: 10.1007/978-3-319-56017-5_2
32. Bouillaguet S, Owen B, Wataha JC, Campo MA, Lange N, Schrenzel J. Intracellular reactive oxygen species in monocytes generated by photosensitive chromophores activated with blue light. Dent Mater. (2008) 24:1070–6. doi: 10.1016/j.dental.2007.12.004
33. Bumah VV, Morrow BN, Cortez PM, Bowman CR, Rojas P, Masson-Meyers DS, et al. The importance of porphyrins in blue light suppression of Streptococcus agalactiae. J Photochem Photobiol B. (2020) 212:111996. doi: 10.1016/j.jphotobiol.2020.111996
34. Wang Y, Murray CK, Hamblin MR, Hooper DC, Dai T. Antimicrobial blue light inactivation of pathogenic microbes: state of the art. Drug Resist Updat. (2017) 35:1–22. doi: 10.1016/j.drup.2017.10.002
35. Dimmer J, Cabral FV, Sabino CP, Silva CR, Núñez-Montoya SC, Cabrera JL, et al. Natural anthraquinones as novel photosentizers for antiparasitic photodynamic inactivation. Hytomedicine. (2019) 61:152894. doi: 10.1016/j.phymed.2019.152894
36. Yu W, Wang Y, Zhu J, Jin L, Liu B, Xia K, et al. Autophagy inhibitor enhance ZnPc/BSA nanoparticle induced photodynamic therapy by suppressing PD-L1 expression in osteosarcoma immunotherapy. Biomaterials. (2019) 192:128–39. doi: 10.1016/j.biomaterials.2018.11.019
37. Rapacka-Zdonczyk A, Wozniak A, Pieranski M, Woziwodzka A, Bielawski KP, Grinholc M. Development of Staphylococcus aureus tolerance to antimicrobial photodynamic inactivation and antimicrobial blue light upon sub-lethal treatment. Sci Rep. (2019) 9:9423. doi: 10.1038/s41598-019-45962-x
38. Cabral FV, Sabino CP, Dimmer JA, Sauter IP, Cortez MJ, Ribeiro MS. Preclinical investigation of methylene blue-mediated antimicrobial photodynamic therapy on Leishmania parasites using real-time bioluminescence. Photochem Photobiol. (2020) 96:604–10. doi: 10.1111/php.13188
39. de Moura TR, Novais FO, Oliveira F, Clarêncio J, Noronha A, Barral A, et al. Toward a novel experimental model of infection to study American cutaneous leishmaniasis caused by Leishmania Braziliensis. Infect Immun. (2005) 73:5827–34. doi: 10.1128/IAI.73.9.5827-5834.2005
40. de Oliveira CI, Brodskyn CI. The immunobiology of Leishmania Braziliensis infection. Front Immunol. (2012) 3:145. doi: 10.3389/fimmu.2012.00145
41. Awasthi A, Mathur RK, Saha B. Immune response to Leishmania infection. Indian J Med Res. (2004) 119:238–58.
42. Lage DP, Martins VT, Vale DL, Freitas CS, Pimenta BL, Moreira GJL, et al. The association between rLiHyp1 protein plus adjuvant and amphotericin B is an effective immunotherapy against visceral leishmaniasis in mice. Acta Tropica. (2023) 246:106986. doi: 10.1016/j.actatropica.2023.106986
43. Rostami MN, Khamesipour A. Potential biomarkers of immune protection in human leishmaniasis. Med Microbiol Immunol. (2021) 210:81–100. doi: 10.1007/s00430-021-00703-8
44. Seyed N, Rafati S. Th1 concomitant immune response mediated by IFN-gamma protects against sand fly delivered Leishmania infection: implications for vaccine design. Cytokine. (2021) 147:155247. doi: 10.1016/j.cyto.2020.155247
45. Soyer TG, Câmara RSB, Pereira IAG, Ramos FF, Jesus MM, Ludolf F, et al. Immunotherapy combining mimotopes selected by phage display plus amphotericin B is effective for treatment against visceral leishmaniasis. Par Immunol. (2024) 46:e13037. doi: 10.1111/pim.13037
46. Kumar R, Bhatia M, Pai K. Role of chemokines in the pathogenesis of visceral leishmaniasis. Curr Med Chem. (2022) 29:5441–61. doi: 10.2174/0929867329666220509171244
47. Grimaldi G, Tesh RB. Leishmaniases of the New World: current concepts and implications for future research. Clin Microbiol Rev. (1993) 6:230–50. doi: 10.1128/CMR.6.3.230
48. Bowman C, Bumah VV, Niesman IR, Cortez P, Enwemeka CS. Structural membrane changes induced by pulsed blue light on methicillin-resistant Staphylococcus aureus (MRSA). J Photochem Photobiol B. (2021) 216:112150. doi: 10.1016/j.jphotobiol.2021.112150
49. Ponte-Sucre A, Gamarro F, Dujardin JC, Barrett MP, López-Vélez R, García-Hernández R, et al. Drug resistance and treatment failure in leishmaniasis: a 21st century challenge. PLoS Negl Trop Dis. (2017) 11:e0006052. doi: 10.1371/journal.pntd.0006052
50. Singh PK, Pawar VK, Jaiswal AK, Singh Y, Srikanth CH, Chaurasia M, et al. Chitosan coated Pluronic F127 micelles for effective delivery of amphotericin B in experimental visceral leishmaniasis. Int J Biol Macromol. (2017) 105:1220–31. doi: 10.1016/j.ijbiomac.2017.07.161
Keywords: blue LED phototherapy, tegumentary leishmaniasis, treatment, amphotericin B, immune response, antileishmanial activity
Citation: Pimenta BL, Lage DP, Freitas CS, Vale DL, Falcão KOM, Dias SSG, Câmara RSB, Pereira IAG, Silva AL, Duarte-Júnior LA, Chavez-Fumagalli MA, Bianchi RF, Coelho EAF and Talvani A (2025) Blue light-emitting diode phototherapy presents in vitro efficacy against distinct Leishmania species and is therapeutic against tegumentary leishmaniasis in BALB/c mice. Front. Immunol. 16:1554051. doi: 10.3389/fimmu.2025.1554051
Received: 31 December 2024; Accepted: 08 April 2025;
Published: 05 May 2025.
Edited by:
Ramona Hurdayal, University of Cape Town, South AfricaReviewed by:
Eduardo René Perez Gonzalez, São Paulo State University, BrazilSonali Das, National Heart, Lung, and Blood Institute (NIH), United States
Raimunda Nonata Ribeiro Sampaio, University of Brasilia, Brazil
Copyright © 2025 Pimenta, Lage, Freitas, Vale, Falcão, Dias, Câmara, Pereira, Silva, Duarte-Júnior, Chavez-Fumagalli, Bianchi, Coelho and Talvani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eduardo Antonio Ferraz Coelho, ZWR1YXJkb2ZlcnJhemNvZWxob0B5YWhvby5jb20uYnI=; André Talvani, dGFsdmFuaUB1Zm9wLmVkdS5icg==