 Xiangning Bai1,2,3*
Xiangning Bai1,2,3* Sajan C. Raju3,4
Sajan C. Raju3,4 Andreas Dehlbæk Knudsen5Rebekka Faber Thudium5
Andreas Dehlbæk Knudsen5Rebekka Faber Thudium5 Nicoline Stender Arentoft5Marco Gelpi5,6
Nicoline Stender Arentoft5Marco Gelpi5,6 Safura-Luise Heidari5Ken M. Kunisaki7,8
Safura-Luise Heidari5Ken M. Kunisaki7,8 Karsten Kristiansen9,10
Karsten Kristiansen9,10 Johannes Roksund Hov3,11
Johannes Roksund Hov3,11 Susanne Dam Nielsen5,12,13†Marius Trøseid3,4,14†
Susanne Dam Nielsen5,12,13†Marius Trøseid3,4,14†- 1Department of Microbiology, Division of Laboratory Medicine, Oslo University Hospital, Oslo, Norway
- 2Center for Infectious Medicine, Department of Medicine Huddinge, Karolinska Institutet, Stockholm, Sweden
- 3Research Institute of Internal Medicine, Oslo University Hospital, Oslo, Norway
- 4Institute of Clinical Medicine, University of Oslo, Oslo, Norway
- 5Department of Infectious Diseases, Rigshospitalet, University of Copenhagen, Copenhagen, Denmark
- 6Department of Infectious Diseases, Hvidovre Hospital, University of Copenhagen, Hvidovre, Denmark
- 7Section of Pulmonary, Allergy, Critical Care and Sleep Medicine, Minneapolis Veterans Affairs Health Care System, Minneapolis, Minnesota, MN, United States
- 8Division of Pulmonary, Allergy, Critical Care and Sleep Medicine, University of Minnesota, Minneapolis, Minnesota, MN, United States
- 9BGI-Shenzhen, Shenzhen, China
- 10Laboratory of Integrative Biomedicine, Department of Biology, University of Copenhagen, Copenhagen, Denmark
- 11Norwegian PSC Research Center, Department of Transplantation Medicine, Oslo University Hospital, Oslo, Norway
- 12Department of Clinical Medicine, Faculty of Health and Medical Sciences, University of Copenhagen, Copenhagen, Denmark
- 13Department of Surgical Gastroenterology, Rigshospitalet, Copenhagen University Hospital, Copenhagen, Denmark
- 14Section of Clinical Immunology and Infectious Diseases, Department of Rheumatology, Dermatology and Infectious Diseases, Oslo University Hospital, Oslo, Norway
Background: People with HIV (PWH) have an increased risk of pulmonary comorbidities compared to people without HIV. The gut microbiome regulates host immunity and is altered in PWH. This study aims to determine potential associations between gut microbiome, lung function decline, and airflow limitation in PWH.
Methods: PWH from the Copenhagen Comorbidity in HIV Infection (COCOMO) Study with available lung function testing and microbiome data were included (n=385). The gut microbiome was characterized using shotgun metagenomic sequencing. Associations between gut microbiome, rapid lung function decline, and airflow limitation were analysed in multivariable logistic regressions adjusted for traditional and HIV-associated risk factors for lung disease.
Results: Several bacterial species were significantly enriched in PWH with rapid lung function decline, including opportunistic pathogenic bacterial species Bacteroides coprophilus, Klebsiella michiganensis, and Clostridium perfringens. A gut microbial dysbiosis index based on compositional changes was associated with rapid lung function decline (adjusted odds ratio (aOR) 1.18, 95% confidence interval (CI) [1.11-1.27], p<0.001), and airflow limitation (aOR 1.16, 95% CI [1.04-1.29], p=0.007) in adjusted multivariable logistic regression analyses.
Conclusion: Associations between the gut dysbiosis index and rapid lung function decline and airflow limitation suggest a potential role of certain gut bacterial species in the pathogenesis of pulmonary comorbidities in PWH.
1 Introduction
PWH, even well-treated, have a higher risk of chronic pulmonary comorbidities compared to the background population, in particular chronic obstructive pulmonary disease (COPD) (1, 2). In fact, chronic lung disease was the most prevalent comorbidity in a recent study investigating trends in non-AIDS related comorbidities over a 10-year period in hospitalized PWH in New York (3), and respiratory mortality is one of the few causes of death in PWH that has not declined over the last 20 years (4).
Importantly, a Dutch study showed a decline in forced expiratory volume in one second (FEV1) and forced vital capacity (FVC) in PWH compared with controls (5). Likewise, a recent work from the COCOMO study showed that even well-treated PWH with no history of smoking had faster lung function decline than population-based controls (2). Yet, the underlying mechanisms involved in the pathogenesis of pulmonary comorbidities and faster lung function decline in PWH are poorly understood. Risk factors may include smoking, which is more prevalent in PWH than in the background population, older age, history of pneumonia, oxidative stress, immune cell activation, and chronic airway inflammation (6). Indeed, increased eosinophilic airway inflammation, as measured by higher exhaled nitric oxide level, was observed in PWH in the COCOMO study (7). Furthermore, a recent study showed that elevated interleukin (IL)-1β and IL-10 were independently associated with faster lung function decline in PWH, suggesting that dysregulated systemic inflammation may play a role in the pathogenesis of pulmonary comorbidities (8).
The human gut microbiome has a fundamental impact on immune system and inflammation, contributing to disease or exacerbating pre-existing conditions (9). Recent studies, including our own (10, 11), have shown that the gut microbiome and its metabolic by-products differ in PWH versus controls, even after adjustment for known confounders such as age and sexual orientation, and that these alterations correlate with previous immunodeficiency and present chronic inflammation (10). Changes in the gut microbiome may contribute to pulmonary diseases through effects on systemic inflammation, microbial translocation, or microbial metabolites via a potential gut-lung axis (12, 13). Dysbiosis in the respiratory microbiome has in previous studies been linked to increased risks of adverse pulmonary outcomes in PWH (14, 15). However, little is known about the association between gut microbiome, pulmonary disease, and lung function decline in PWH, as well as the potential of gut-lung microbiome connections in the HIV-associated pulmonary disease.
The primary aim of this study was to determine the potential associations between the gut microbiome including microbial species and their pathways, and lung function decline in PWH. The secondary aim was to determine the associations between the gut microbiome and airflow limitation as defined by Global Initiative for Chronic Obstructive Lung Disease (GOLD) (16).
2 Materials and methods
2.1 Study design and participants
The Copenhagen Comorbidity in HIV Infection (COCOMO) study is an ongoing longitudinal cohort study designed to determine the burden of, and risk factors for, non-AIDS comorbidities in PWH in Copenhagen, Denmark (17). Spirometry was performed at baseline from March 2015 until November 2016, and at subsequent follow-up visits between April 2017 and April 2019 with a median follow-up time of 2.3 years, as described previously (2). Eligibility criteria for this study were participants in COCOMO aged ≥25 years with two valid spirometry tests separated by at least 2 years of follow-up. Only PWH who had received antiretroviral therapy (ART) for a minimum of 6 months at baseline and had available fecal samples were included in the present study. Participants who provided fecal samples were older than participants who did not provide fecal samples (53 vs 49 years old, p<0.001), but they did not differ on any of the other parameters when adjusted for age (all p>0.05).
2.2 Lung function testing
Lung function was assessed by spirometry using an ultrasonic spirometer (EasyOne, ndd Medical, Zürich, Switzerland). Prebronchodilator forced expiratory volume in 1 second (FEV1) and forced vital capacity (FVC) were measured at baseline and at follow-up in accordance with the European Respiratory Society/American Thoracic Society recommendations (18). Predicted values for FEV1 and FVC, along with z-scores and the lower limit of normal for the FEV1/FVC ratio, were determined using the multi-ethnic reference equations from the Global Lung Function Initiative (19). Lung function decline was measured as changes in FEV1 milliliter (mL) per year (8). In relation to each test, a quality grading from A to F was automatically generated by the EasyOne spirometer, spirometry data with grade A-C were defined as high quality and were used for sensitivity analyses.
2.3 Outcomes definition
The primary outcome was rapid lung function decline defined as FEV1 decline >40 mL/year as previously described (2). The secondary outcome was airflow limitation defined as FEV1/FVC <0.70 and FEV1 <80% of the predicted value corresponding to COPD of moderate or worse severity (≥2 COPD grade) according to the GOLD (16).
2.4 Inflammatory markers measurements
Plasma concentrations of pro- and anti-inflammatory cytokines were measured using Luminex immunoassays (R&D systems, Minneapolis, Minnesota), as previously described in detail (8). In brief, the multiplex assay kits included the proinflammatory markers interleukin 1-beta (IL-1β), interleukin 2 (IL-2), IL-6, interferon gamma (IFN-γ), and tumor necrosis factor alpha (TNF-α) and the anti-inflammatory marker interleukin 10 (IL-10). The concentrations of microbial translocation marker soluble CD14 (sCD14) and monocyte activation marker soluble CD163 (sCD163), were measured in plasma by high sensitivity enzyme-linked immunosorbent assay kits (R&D systems, Minneapolis, Minnesota) at the Research Institute of Internal Medicine, Division of Surgery, Inflammatory Diseases and Transplantation, Oslo University Hospital, Rikshospitalet, Oslo, Norway. High-sensitivity CRP (hs-CRP) was measured using a turbidimetric assay at the Department of Clinical Biochemistry, Herlev Hospital, Copenhagen, Denmark.
2.5 Fecal sample collection and processing
At study inclusion, participants were instructed to collect fecal samples using a standardized sampling device and collection tubes with DNA Stabilizer (Stratec Molecular). Samples were frozen at −80°C on arrival and eventually shipped on dry ice to Oslo for microbiome analyses. DNA was extracted using the PSPSpin Stool DNA-Plus Kit (Stratec Molecular), following the manufacturer’s protocol, slightly modified by adding a bead-beating step, as described elsewhere (10).
2.6 Shotgun metagenome sequencing and microbiome profiling
In the COCOMO study, which included 1099 participants, 385 individuals had both lung function measurements and fecal samples collected at baseline. Shotgun metagenome sequencing was performed on these samples using 150 bp paired-end sequencing on the MGISEQ-T7 platform at MGI Tech, Mārupe, Latvia. The pre-processing of the raw reads was conducted using the KneadData pipeline (v0.10.0). Initially, raw reads were trimmed to a Phred score of 20 and reads below the specified minimum length were removed using Trimmomatic (version 0.39), which was integrated within the KneadData pipeline (20). Subsequently, trimmed reads were aligned against the human reference genome using bowtie2 (21), reads aligning to the human genome were eliminated from further analysis. Taxonomic profiling was performed using MetaPhlAn (v3.0.0.0) by default including reference genomes of bacteria, archaea, and eukaryotes (22). HUMAnN3 (v3.0.0) (22) was employed for functional profiling (genes and pathways), incorporating MetaPhlAn, DIAMOND 0.9.36 (23), and the databases uniref90 (v201901) (24) and mpa_v30_CHOCOPhlAn_201901. Additionally, metabolic pathways were identified from the MetaCyc database (metacyc.org).
Taxonomic profiles obtained from MetaPhlAn were imported and analyzed using the phyloseq v1.40.0 (25) package in R. To evaluate taxonomic and functional richness as well as diversity, several alpha diversity indices including the Shannon index and inverse Simpson index were employed. Beta-diversity was assessed by calculating the Bray–Curtis dissimilarity index. Ordinations were constructed using Principal Coordinate Analysis (PCoA). PERMANOVA was performed with adjustment for the significantly different covariates between groups (Table 1), using the vegan::adonis2 function between the groups with 999 permutations. Benjamini-Hochberg (BH) method was used to control the false discovery rate for multiple comparisons.
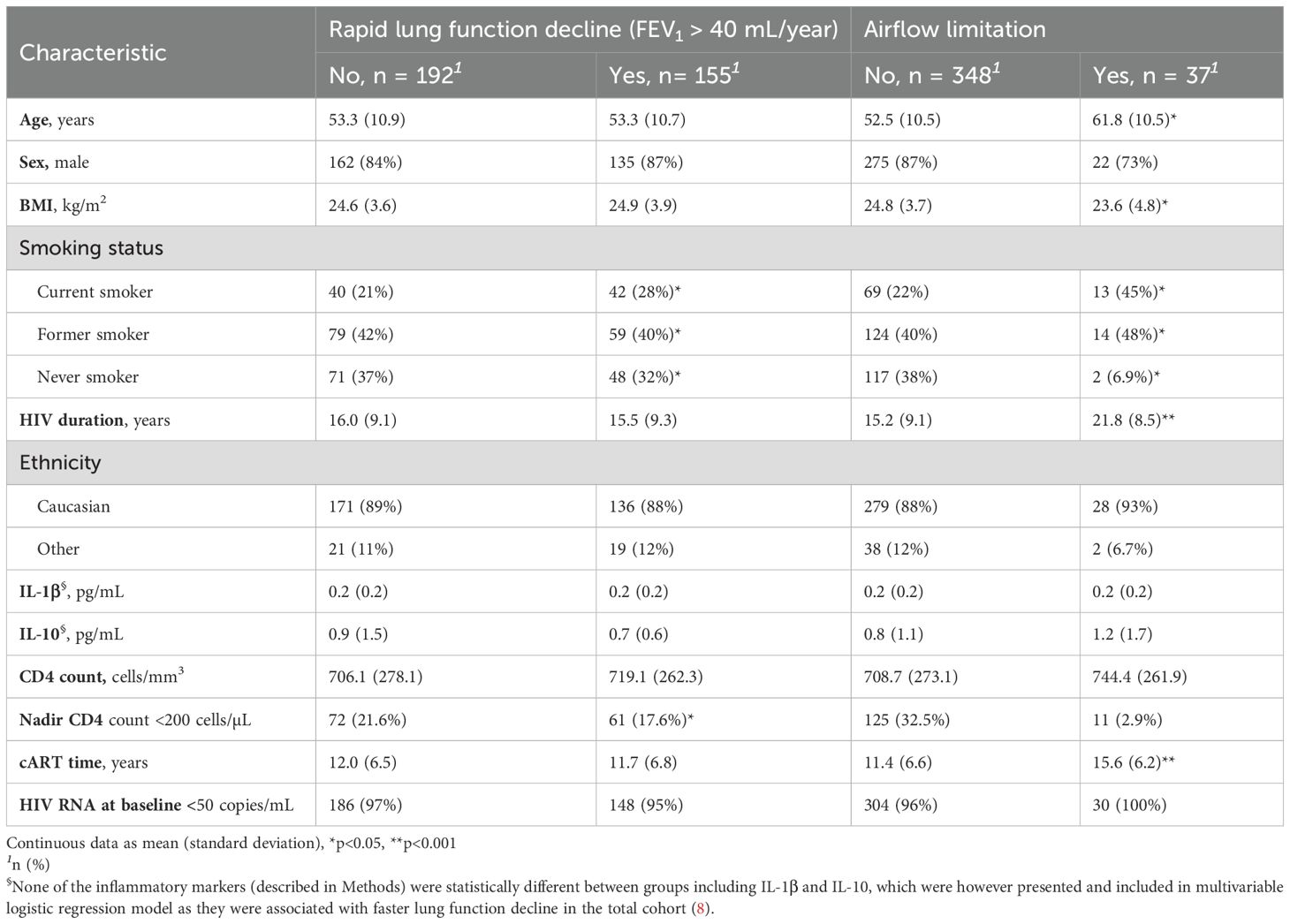
Table 1. Demographic characteristics of PWH in present sub-study of COCOMO.
Differential abundance (DA) analyses were conducted to test the differences in the relative abundance in microbial taxa, genes, and metabolic pathways between groups, using the DESeq2 package with the Wald test and parametric fitting (26), and further adjusted for smoking status, which was associated with rapid lung function decline as well as airflow limitation (Table 1). Log2 (fold-change) and BH-adjusted p values (< 0.05) were extracted for each comparison from the model. Analyses were carried out at different taxonomic (i.e., species, genus, and family), and functional (i.e., genes and pathways) levels. Based on differentially abundant microbial species from the DESeq2 analysis, a microbial dysbiosis index (quantitative measure used to assess the balance or imbalance of microbial communities), was calculated as log10 (sum of the abundances of the species increased in rapid decliners/sum of the abundances of species decreased in rapid decliners), as first reported by Gevers et al. (27), and in accordance with other microbiome studies from the COCOMO cohort (10, 28). In addition, Mann–Whitney U test was performed further as a sensitivity analysis of differentially abundant species identified by DESeq2, and taxa with a p-value >0.10 were removed from the dysbiosis index.
2.7 Statistics
Descriptive statistics were presented by expressing continuous variables as mean ± standard deviation and categorical variables as numbers and percentages. Differences in distributions of continuous variables were tested using Mann–Whitney U tests. DA analyses in DESeq2 were adjusted for significant covariate smoking status at follow-up (current, former, never) for both the primary outcome (rapid lung function decline), and the secondary outcome (airflow limitation).
Associations between the microbial dysbiosis index and primary outcome was further adjusted for other traditional (age, sex, BMI, ethnicity) and HIV-related (transmission mode, nadir CD4 count, HIV duration) risk factors for lung disease in model 1, using multivariable logistic regressions, and further adjusted for soluble levels of IL-1β and IL-10 (previously associated with rapid lung function decline in PWH (8)) in model 2, to verify if the microbial dysbiosis index associated with rapid lung function decline was robust and independent of these potential confounders. Moreover, a sensitivity analysis was performed by excluding those with self-reported history of pneumonia in the decade prior to fecal sampling to rule out its potential confounding effect on rapid lung function decline.
We further tested if the microbial dysbiosis index was also associated with the secondary outcome airflow limitation. Due to the low sample size of airflow limitation, association between the dysbiosis index and airflow limitation was separately adjusted for age, BMI, and a third significant covariate (Table 1) at a time.
Spearman’s correlation analysis was performed between differentially abundant species from DESeq2 analysis and HIV-related clinical parameters, and visualized by the ggcorrplot (Ver 0.1.4.1) package in R.
The co-occurrence network plots were generated for rapid decliners and normal decliners using the iGraph R package (V 2.1.1) using the subset of top abundant taxonomy profiles, including the differentially abundant taxa from DESEq2 analysis, from MetaPhlAn3 to assess the correlations among microbial taxa based on their Spearman’s correlations.
3 Results
3.1 Baseline characteristics of PWH with lung function abnormalities
Among 385 participants who had lung function measurement and metagenome data at baseline, 347 had valid lung function data also at follow-up, thus had measured FEV1 decline, and 155 (44.7%) presented with rapid lung function decline (Table 1). PWH with rapid lung function decline were overrepresented by current smokers, whereas airflow limitation was associated with current smoking, older age, lower BMI, longer HIV duration and cART time.
3.2 Association between gut microbiome and rapid lung function decline in PWH
No statistically significant difference in microbial alpha diversity at species level measured by Shannon and inverse Simpson index was observed between PWH with and without rapid lung function decline (Figures 1A, B). No difference in microbiome composition at species level (beta diversity) was observed between the two groups (Figure 1C). Similarly, no difference in microbial alpha or beta diversity was observed at genus or family level (Supplementary Figures S1A–E, G).

Figure 1. Gut microbiome profiles at species level in PWH with and without rapid lung function decline. (A), Shannon diversity index. (B), inverse Simpson diversity index. (C), Beta-diversity assessed by the Bray–Curtis dissimilarity index, visualized using Principal Coordinate Analysis (PCoA). (D), Bacterial species differentially abundant between PWH with and without rapid lung function decline, analyzed using the DESeq2 package, adjusted for smoking.
On the compositional level, after adjusting for smoking status (current, former, never) in DESeq2, several bacterial species were significantly enriched in PWH with rapid lung function decline compared to those with normal lung function decline (Figure 1D). After removing non-significant taxa from Mann Whitney U test, species enriched in PWH with rapid lung function decline included Bacteroides coprophilus, Klebsiella michiganensis, Catabacter hongkongensis, Clostridium perfringens, Lactobacillus oris, Lactobacillus paragasseri, while the relative abundance of two species, Cloacibacillus porcorum and Absiella dolichum were decreased in rapid decliners. These differentially abundant species between rapid and normal lung function decliners were used to define a microbial dysbiosis index as previously described (27), and explained in the Methods section. At the genus level, the relative abundances of Helicobacter, Catabacter, and Parascardovia were significantly enriched, while the relative abundances of 6 genera were decreased in rapid decliners compared to normal decliners (Supplementary Figure S1F). At the family level, the relative abundance of Helicobacteraceae was significantly increased, while the relative abundance of Spirochaetaceae, Entamoebidae, Hafniaceae were decreased in rapid decliners (Supplementary Figure S1H).
At the functional level, no significant difference in gene richness or diversity was found between rapid and normal decliners (Supplementary Figures S2A–C). No difference in alpha or beta diversity in pathways was found between the two groups (Supplementary Figures S3A–C). No differentially abundant genes or pathways were found between rapid and normal decliners after adjusting for smoking status (current, former, never) at follow-up.
3.3 Gut microbiome profiles in PWH with and without airflow limitation
No statistically significant difference was observed in microbial alpha or beta diversity between PWH with and without airflow limitation (Supplementary Figures S4A–C). After adjusting for smoking status, the relative abundances of five species, including Bacteroides sp CAG 661 which was also enriched in rapid decliners, were significantly enriched in PWH with airflow limitation, while the relative abundances of several species were decreased in this group (Supplementary Figure S4D).
3.4 Prediction of rapid lung function decline by gut microbial dysbiosis
Finally, we assessed whether gut microbial dysbiosis could predict rapid lung function decline independently of relevant confounders in addition to smoking status. In a multivariable logistic regression adjusted for other traditional (age, sex, BMI, ethnicity) and HIV-related (transmission mode, nadir CD4 count, HIV duration) risk factors for lung disease, the dysbiosis index was independently associated with rapid lung function decline (aOR 1.18, 95% CI [1.11-1.27], p<0.001). Restricting analyses to PWH with no self-reported history of pneumonia (62%) did not alter the association between microbial dysbiosis index and rapid lung function decline (aOR 1.25, 95% CI [1.14-1.38], p<0.001). Furthermore, restricting analyses to PWH with predefined high-quality spirometry data (86.2%) did not alter the association between microbial dysbiosis index and rapid lung function decline either (aOR 1.16, 95% CI [1.09-1.25], p<0.001).
The microbial dysbiosis index was not associated with levels of inflammatory markers (described in Methods), and adjusting further for IL-1β and IL-10 which were associated with faster lung function decline in the total cohort (8) did not alter the association between microbial dysbiosis index and rapid lung function decline.
Importantly, the microbial dysbiosis index based on rapid decliners was associated with airflow limitation in multivariable analyses adjusted for significant confounders for airflow limitation (age, BMI, HIV duration, and cART time) (aOR 1.16, 95% CI [1.04-1.29], p=0.007). No significant correlations between microbial taxa within the dysbiosis index (i.e., differentially abundant taxa between rapid and normal decliners) and clinical parameters were found, with a few exceptions showing weak correlations (Spearman’s correlation coeffecient < ±0.2), though correlations were observed between some clinical parameters as anticipated (Supplementary Figure S5).
A number of microbial taxa were closely correlated in co-occurrence plots (Spearman’s correlation coeffecient>0.5 and p<0.05) both in rapid and normal decliners (Supplementary Figure S6), while no significant correlations were observed among most taxa within the microbial dysbiosis index (Supplementary Figure S6, Supplementary Figure S5).
4 Discussion
In this study using a large cohort of well-treated PWH, we found no significant difference in overall gut microbial diversity between PWH with and without rapid lung function decline. However, several microbial species were differentially abundant between PWH with and without rapid lung function decline after adjustment for smoking status. After further adjustment for traditional and HIV-related risk factors for lung disease, as well as cytokines (IL-1β, IL-10), the gut microbial dysbiosis index was independently associated with rapid lung function decline. Importantly, we also found an independent association of the gut microbial dysbiosis index with the airflow limitation in PWH, suggesting that the gut dysbiosis based on the lung function decline may be predictive for airflow limitation. Moreover, we found no overlap at bacterial genus or family levels between microbial dysbiosis associated with rapid lung function decline among PWH in this study and our previously defined HIV-related microbiome dysbiosis using 16S rRNA gene sequencing (10), suggesting that microbiome dysbiosis-associated rapid lung function decline is not related to HIV status per se.
We observed that some opportunistic pathogenic bacterial species were significantly enriched in PWH with rapid lung function decline compared to those without, e.g., Bacteroides coprophilus, Klebsiella michiganensis, Clostridium perfringens. Bacteroides coprophilus has been associated with COVID-19 and multisystem inflammatory syndrome (29). Klebsiella michiganensis, among other Klebsiella species, has been reported to be dominant in gut microbiota among infants with lower respiratory infection (30). Clostridium perfringens produces severe and rapidly fatal enterotoxemia affecting several organs including lungs, and a previous study showed that lung endothelial cells are sensitive to epsilon toxin from C. perfringens (31). The enrichment of these pathogenic bacterial species may play a role in the pulmonary pathology in PWH and contribute to rapid lung function decline, either due to the gut microbiome functioning as a reservoir for the lung microbiome, or through an indirect effect mediated by e.g., systemic inflammation caused by gut microbiota dysbiosis, yet the underlying mechanisms and causal link are to be elucidated.
We investigated further if the microbial dysbiosis index was correlated with soluble levels of IL-1β and IL-10, which in our previous work were associated with rapid lung function decline in the total COCOMO cohort (8). However, the association between the microbial dysbiosis index and lung function decline was not altered when adjusting for these markers. This may indicate that pulmonary pathological effect may be more driven by other mechanisms linked with gut microbial dysbiosis than systemic inflammation. For instance, gut microbiome may serve as a reservoir for the airway microbiome, the dysbiosis in the gut could lead to airway microbiome dysbiosis, which directly contribute to the pulmonary pathology. Further studies on airway microbiome in correlation to pulmonary health are required.
We observed no significant difference in the gut microbiome diversity between PWH with and without airflow limitation. Nevertheless, increased relative abundance of several species were found in PWH with airflow limitation. Of a potential interest, Bacteroides sp CAG 661 was also associated with rapid lung function decliner, suggesting its pathogenic potential to the lung. It should be acknowledged that our sample size regarding airflow limitation at baseline is a major limitation, the observed correlations are to be further validated in future studies.
Our findings are in line with two previous studies of cohorts including PWH and HIV-uninfected controls (32, 33). Both studies, using 16S rRNA gene sequencing, reported no associations between gut microbiota diversity and HIV status or lung function, while one study (32) showed that alterations in the oral microbiome were associated with impaired pulmonary function and systemic inflammation in PWH, but not among HIV-uninfected individuals. Of note, our study provides novel data on the potential role of several bacterial species in the rapid lung function among PWH, which has not been observed in the two earlier studies. This may be due to the variations in the study populations and different microbiome profiling methods. As it is known that 16S rRNA gene-based microbiome profiling detects only part of the microbiome community revealed by shotgun metagenome sequencing, and the less abundant taxa, particularly specific species detected only by shotgun sequencing can be biologically meaningful (34). In the general population, associations between gut microbiome and lung function decline have been reported, particularly in humans with COPD (35). For example, Bowerman et al. found that Bifidobacteriaceae, Eubacteriaceae, Lactobacillaceae, Micrococcaceae, Streptococcaceae, and Veillonellaceae were associated with COPD; while Desulfovibrionaceae, Bacteroidaceae, Gastranaerophilaceae, and Selenomonadaceae were negatively associated with COPD (36). We did not find overlap of bacteria associated with lung function decline in PWH identified in this study, and those commonly reported in the general population. Again, comparisons between different studies should be cautious and consider the variations in populations and microbiome approaches.
We acknowledge limitations and strengths of the study. First, we did not include HIV-negative controls due to lack of HIV-negative individuals in the COCOMO controls with matched microbiome samples and lung function data, further studies using shotgun metagenome sequencing and including both PWH and HIV-negative controls are thus warranted to validate our findings and to assess whether such findings are specific to PWH or not. Second, the sample size of PWH with airflow limitation at baseline was small; thus, the study was underpowered to draw robust conclusions regarding associations between gut microbiome and airflow limitation, larger validation cohorts are needed in future studies. Related to this limitation, our spirometry measures were pre-bronchodilator only, and therefore, we cannot determine how many of participants may have normalized lung function after bronchodilation (suggestive of asthma) versus had persistent airflow limitation (suggestive of COPD). We hence used the term ‘airflow limitation’ rather than COPD. Third, our results do not allow for determination of causal directionality. Furthermore, we acknowledge the lack of virome and resistome analysis from metagenome data, which is however not the focus of this study. The strengths of this study include the large sample size of PWH who had lung function measurements with longitudinal follow-up, and adjustments for a number of known confounders.
5 Conclusions
We report no significant changes in the overall gut microbiome composition and functional potential related to pulmonary comorbidity in PWH. However, changes in the relative abundances of several bacterial species were associated with rapid lung function decline, and the microbial dysbiosis index was independently associated with rapid lung function decline and airflow limitation. Future studies are warranted to validate our findings. In addition, there is a need of studies focusing on the airway microbiome, which may play a more important and more direct role in pulmonary comorbidities in PWH.
Data availability statement
The anonymised shotgun metagenome sequencing datasets presented in this study are deposited in the NCBI Sequence Read Archive (SRA) under the BioProject ID PRJNA1267518. Metadata are subject to restrictions due to approvals from the ethical committee and the Danish Data Protection Agency and cannot be shared publicly. Access to the metadata may be granted upon reasonable request to Dr. Susanne Dam Nielsen (Susanne.Dam.Poulsen@regionh.dk).
Ethics statement
The COCOMO study was approved by the Danish Data Protection Agency and the Committee on Health Research Ethics of the Capital Region of Denmark (H-8-2014-004). All participants provided written informed consent.
Author contributions
XB: Investigation, Methodology, Supervision, Validation, Writing – original draft, Writing – review & editing. SR: Formal Analysis, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. AK: Formal Analysis, Investigation, Methodology, Visualization, Writing – review & editing, Validation. RT: Formal Analysis, Investigation, Methodology, Writing – review & editing, Software. NA: Formal Analysis, Investigation, Methodology, Writing – review & editing. MG: Investigation, Methodology, Writing – review & editing, Formal Analysis. S-LH: Investigation, Methodology, Writing – review & editing. KMK: Investigation, Supervision, Writing – review & editing. KK: Methodology, Supervision, Writing – review & editing, Investigation. JH: Methodology, Software, Supervision, Writing – review & editing. SN: Conceptualization, Data curation, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing. MT: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the South-Eastern Norway Regional Health Authority (2016004), Novo Nordic Foundation and Rigshospitalet Research Council. This material is also the result of work supported with resources and the use of facilities at the Minneapolis Veterans Affairs Medical Center, Minneapolis/USA.
Acknowledgments
The authors thank all the study subjects for their participation. The authors thank the staff at the Department of Infectious Diseases at Rigshospitalet and at Hvidovre Hospital for their dedicated participation. The authors also acknowledge all computational resources for the metagenomics analysis granted by the Norwegian academic high-performance computing and storage facilities managed by Sigma2 under the Norwegian Research Infrastructure Service (NRIS).
Conflict of interest
AK reports grants from the European Commission EU 7th Framework Grant ID:603266, Gilead Sciences Research Scholars Program 2023 and a speaker’s fee from GlaxoSmithKline’; his wife is employed at Novo Nordisk. RT reports a travel grant from Gilead Sciences. NA reports PhD grant from the Research Foundation of Odense University Hospital and Copenhagen University Hospital Rigshospitalet. S-LH reports a research-scholar grant from the Novo Nordic Foundation and a travel grant from GlaxoSmithKline. KMK reports personal fees from Nuvaira for Data and Safety Monitoring Board work. SN reports unrestricted research grants from Novo Nordic Foundation, Rigshospitalet Research Foundation, Independent Research Fund Denmark, Svend Andersen Fonden; she is on advisory board for Gilead, Takeda, and GSK.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1555441/full#supplementary-material
References
1. Bigna JJ, Kenne AM, Asangbeh SL, and Sibetcheu AT. Prevalence of chronic obstructive pulmonary disease in the global population with HIV: a systematic review and meta-analysis. Lancet Glob Health. (2018) 6:e193–202. doi: 10.1016/S2214-109X(17)30451-5
2. Thudium RF, Ronit A, Afzal S, Colak Y, Forman JL, Mendo F, et al. Faster lung function decline in people living with HIV despite adequate treatment: a longitudinal matched cohort study. Thorax. (2023) 78:535–42. doi: 10.1136/thorax-2022-218910
3. Rowell-Cunsolo TL, Hu G, Bellerose M, and Liu JF. Trends in comorbidities among human immunodeficiency virus-infected hospital admissions in New York City from 2006-2016. Clin Infect Dis. (2021) 73:E1957–E63. doi: 10.1093/cid/ciaa1760
4. Trickey A, Mcginnis K, Gill MJ, Abgrall S, Berenguer J, Wyen C, et al. Longitudinal trends in causes of death among adults with HIV on antiretroviral therapy in Europe and North America from 1996 to 2020: a collaboration of cohort studies. Lancet Hiv. (2024) 11:e176–e85. doi: 10.1016/S2352-3018(23)00272-2
5. Verboeket SO, Boyd A, Wit FW, Verheij E, Schim van der Loeff MF, Kootstra N, et al. Changes in lung function among treated HIV-positive and HIV-negative individuals: analysis of the prospective AGE(h)IV cohort study. Lancet Healthy Longev. (2021) 2:e202–e11. doi: 10.1016/S2666-7568(21)00033-7
6. Konstantinidis I, Crothers K, Kunisaki KM, Drummond MB, Benfield T, Zar HJ, et al. HIV-associated lung disease. Nat Rev Dis Primers. (2023) 9:39. doi: 10.1038/s41572-023-00450-5
7. Thudium RF, Hughes NLP, Afzal S, Colak Y, Gelpi M, Knudsen AD, et al. Fraction of exhaled nitric oxide levels are elevated in people living with human immunodeficiency virus compared to uninfected controls, suggesting increased eosinophilic airway inflammation. Clin Infect Dis. (2020) 71:3214–21. doi: 10.1093/cid/ciz1223
8. Thudium RF, Arentoft NS, Hoel H, Afzal S, von Stemann JH, Forman JL, et al. Elevated levels of interleukin-1beta and interleukin-10 are associated with faster lung function decline in people with well-treated human immunodeficiency virus. J Infect Dis. (2023) 228:1080–8. doi: 10.1093/infdis/jiad233
9. Zheng D, Liwinski T, and Elinav E. Interaction between microbiota and immunity in health and disease. Cell Res. (2020) 30:492–506. doi: 10.1038/s41422-020-0332-7
10. Gelpi M, Vestad B, Hansen SH, Holm K, Drivsholm N, Goetz A, et al. Impact of human immunodeficiency virus-related gut microbiota alterations on metabolic comorbid conditions. Clin Infect Dis. (2020) 71:e359–e67. doi: 10.1093/cid/ciz1235
11. Bai X, Narayanan A, Nowak P, Ray S, Neogi U, and Sonnerborg A. Whole-genome metagenomic analysis of the gut microbiome in HIV-1-infected individuals on antiretroviral therapy. Front Microbiol. (2021) 12:667718. doi: 10.3389/fmicb.2021.667718
12. Trompette A, Gollwitzer ES, Yadava K, Sichelstiel AK, Sprenger N, Ngom-Bru C, et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat Med. (2014) 20:159–66. doi: 10.1038/nm.3444
13. Budden KF, Gellatly SL, Wood DL, Cooper MA, Morrison M, Hugenholtz P, et al. Emerging pathogenic links between microbiota and the gut-lung axis. Nat Rev Microbiol. (2017) 15:55–63. doi: 10.1038/nrmicro.2016.142
14. Kayongo A, Bartolomaeus TUP, Birkner T, Marko L, Lober U, Kigozi E, et al. Sputum microbiome and chronic obstructive pulmonary disease in a rural Ugandan cohort of well-controlled HIV infection. Microbiol Spectr. (2023) 11:e0213921. doi: 10.1128/spectrum.02139-21
15. Bai X, Nielsen SD, Kunisaki KM, and Troseid M. Pulmonary comorbidities in people with HIV- the microbiome connection. Curr Opin HIV AIDS. (2024) 19:246–52. doi: 10.1097/COH.0000000000000871
16. Gold. global strategy for prevention, diagnosis and management of COPD: 2024 Report. Global Initiative for Chronic Obstructive Lung Disease (2024). p. 31.
17. Ronit A, Haissman J, Kirkegaard-Klitbo DM, Kristensen TS, Lebech AM, Benfield T, et al. Copenhagen comorbidity in HIV infection (COCOMO) study: a study protocol for a longitudinal, non-interventional assessment of non-AIDS comorbidity in HIV infection in Denmark. BMC Infect Dis. (2016) 16:713. doi: 10.1186/s12879-016-2026-9
18. Miller MR, Hankinson J, Brusasco V, Burgos F, Casaburi R, Coates A, et al. Standardisation of spirometry. Eur Respir J. (2005) 26:319–38. doi: 10.1183/09031936.05.00034805
19. Quanjer PH, Stanojevic S, Cole TJ, Baur X, Hall GL, Culver BH, et al. Multi-ethnic reference values for spirometry for the 3-95-yr age range: the global lung function 2012 equations. Eur Respir J. (2012) 40:1324–43. doi: 10.1183/09031936.00080312
20. Bolger AM, Lohse M, and Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. (2014) 30:2114–20. doi: 10.1093/bioinformatics/btu170
21. Langmead B and Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nat Methods. (2012) 9:357–9. doi: 10.1038/nmeth.1923
22. Beghini F, McIver LJ, Blanco-Miguez A, Dubois L, Asnicar F, Maharjan S, et al. Integrating taxonomic, functional, and strain-level profiling of diverse microbial communities with bioBakery 3. Elife. (2021) 10:e65088. doi: 10.7554/eLife.65088
23. Buchfink B, Xie C, and Huson DH. Fast and sensitive protein alignment using DIAMOND. Nat Methods. (2015) 12:59–60. doi: 10.1038/nmeth.3176
24. Suzek BE, Huang H, McGarvey P, Mazumder R, and Wu CH. UniRef: comprehensive and non-redundant UniProt reference clusters. Bioinformatics. (2007) 23:1282–8. doi: 10.1093/bioinformatics/btm098
25. McMurdie PJ and Holmes S. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One. (2013) 8:e61217. doi: 10.1371/journal.pone.0061217
26. Love MI, Huber W, and Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. (2014) 15:550. doi: 10.1186/s13059-014-0550-8
27. Gevers D, Kugathasan S, Denson LA, Vazquez-Baeza Y, Van Treuren W, Ren B, et al. The treatment-naive microbiome in new-onset Crohn’s disease. Cell Host Microbe. (2014) 15:382–92. doi: 10.1016/j.chom.2014.02.005
28. Troseid M, Molinaro A, Gelpi M, Vestad B, Kofoed KF, Fuchs A, et al. Gut microbiota alterations and circulating imidazole propionate levels are associated with obstructive coronary artery disease in people with HIV. J Infect Dis. (2024) 229:898–907. doi: 10.1093/infdis/jiad604
29. Suskun C, Kilic O, Yilmaz Ciftdogan D, Guven S, Karbuz A, Ozkaya Parlakay A, et al. Intestinal microbiota composition of children with infection with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and multisystem inflammatory syndrome (MIS-C). Eur J Pediatr. (2022) 181:3175–91. doi: 10.1007/s00431-022-04494-9
30. Sjodin KS, Sjodin A, Ruszczynski M, Kristensen MB, Hernell O, Szajewska H, et al. Targeting the gut-lung axis by synbiotic feeding to infants in a randomized controlled trial. BMC Biol. (2023) 21:38. doi: 10.1186/s12915-023-01531-3
31. Dorca-Arevalo J, Dorca E, Torrejon-Escribano B, Blanch M, Martin-Satue M, and Blasi J. Lung endothelial cells are sensitive to epsilon toxin from Clostridium perfringens. Vet Res. (2020) 51:27. doi: 10.1186/s13567-020-00748-2
32. Yang L, Dunlap DG, Qin S, Fitch A, Li K, Koch CD, et al. Alterations in oral microbiota in HIV are related to decreased pulmonary function. Am J Respir Crit Care Med. (2020) 201:445–57. doi: 10.1164/rccm.201905-1016OC
33. Flygel TT, Sovershaeva E, Claassen-Weitz S, Hjerde E, Mwaikono KS, Odland JO, et al. Composition of gut microbiota of children and adolescents with perinatal human immunodeficiency virus infection taking antiretroviral therapy in Zimbabwe. J Infect Dis. (2020) 221:483–92. doi: 10.1093/infdis/jiz473
34. Durazzi F, Sala C, Castellani G, Manfreda G, Remondini D, and De Cesare A. Comparison between 16S rRNA and shotgun sequencing data for the taxonomic characterization of the gut microbiota. Sci Rep-Uk. (2021) 11:3030. doi: 10.1038/s41598-021-82726-y
35. Karakasidis E, Kotsiou OS, and Gourgoulianis KI. Lung and gut microbiome in COPD. J Pers Med. (2023) 13:804. doi: 10.3390/jpm13050804
Keywords: HIV, gut microbiome, pulmonary comorbidity, lung function decline, airflow limitation, spirometry
Citation: Bai X, Raju SC, Knudsen AD, Thudium RF, Arentoft NS, Gelpi M, Heidari S-L, Kunisaki KM, Kristiansen K, Hov JR, Nielsen SD and Trøseid M (2025) Microbiome profiling reveals gut bacterial species associated with rapid lung function decline in people with HIV. Front. Immunol. 16:1555441. doi: 10.3389/fimmu.2025.1555441
Received: 04 January 2025; Accepted: 06 May 2025;
Published: 10 June 2025.
Edited by:
Sonia Moretti, National Institute of Health (ISS), ItalyReviewed by:
Shulin Qin, University of Pittsburgh, United StatesSweta M. Patel, Duke University, United States
Copyright © 2025 Bai, Raju, Knudsen, Thudium, Arentoft, Gelpi, Heidari, Kunisaki, Kristiansen, Hov, Nielsen and Trøseid. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiangning Bai, eGlhbmduaW5nLmJhaUBraS5zZQ==
†These authors share senior authorship