 Diego Catalán1
Diego Catalán1 Lilian Soto1,2
Lilian Soto1,2 Óscar Neira3María C. Cuéllar-Gutiérrez3
Óscar Neira3María C. Cuéllar-Gutiérrez3 Roberto Díaz-Peña4,5
Roberto Díaz-Peña4,5 Octavio Aravena1
Octavio Aravena1 Eduard Palou6
Eduard Palou6 Montserrat Carrascal7
Montserrat Carrascal7 Juan C. Aguillón1*†
Juan C. Aguillón1*† Jaxaira Maggi7*†
Jaxaira Maggi7*†- 1Immune Regulation and Tolerance Research Group (IRT Group), Programa Disciplinario de Inmunología, Instituto de Ciencias Biomédicas, Facultad de Medicina, Universidad de Chile, Santiago, Chile
- 2Sección Reumatología, Departamento de Medicina Interna, Hospital Clínico de la Universidad de Chile, Santiago, Chile
- 3Hospital del Salvador, Universidad de Chile, Santiago, Chile
- 4Facultad de Ciencias de la Salud, Universidad Autónoma de Chile, Talca, Chile
- 5Immunogenetics Lab, Fundación Pública Galega de Medicina Xenómica, Servizo Galego de Saúde (SERGAS), Grupo de MedicinaXenómica-Universidade de Santiago de Compostela (USC), Instituto de investigación Sanitaria de Santiago de Compostela, Santiago de Compostela, Spain
- 6Servicio de Inmunología, Hospital Clínic de Barcelona, Barcelona, Spain
- 7Biological and Environmental Proteomics Group, Institute of Biomedical Research of Barcelona, Spanish National Research Council (IIBB-CSIC/IDIBAPS), Barcelona, Spain
Introduction: Rheumatoid arthritis (RA) is an autoimmune disease influenced by genetic factors, particularly HLA-DRB1 alleles. The objective of this study was to characterize the distribution of HLA-DRB1 alleles in Chilean RA patients and healthy controls (HC) and evaluate associations with susceptibility or protection, autoantibody seropositivity, and disease activity.
Methods: We genotyped 367 RA patients and 623 HC for HLA-DRB1 using PCR-SSO. Then, we examined allele frequencies and distribution, including known RA risk alleles of the “Shared Epitope” (SE) of HLA-DRB1 and protective (PR) alleles, using the Chi-square or Fisher’s exact tests. Odds ratios with 95% confidence intervals were calculated to measure the degree of association, and unpaired T-tests were used to compare continuous variables.
Results: The most frequent SE alleles among RA patients were *04:01 (16.1%), *04:04 (13.9%), and *14:02 (11.7%). SE alleles *04:01, *04:04, *04:05, *04:08, and *10:01, along with non-SE alleles *09:01 and *15:02, were associated with RA susceptibility. In addition, allele *14:02 showed an association with the presence of anti-cyclic citrullinated peptides (anti-CCP) antibodies. Meanwhile, PR alleles *11:01 (14.8%) and *16:02 (9.8%) were observed most frequently in HC and RA patients, respectively. PR alleles *11:01, *11:04, and *13:01, as well as the non-PR alleles *15:01, *04:07, *03:01, *07:01, and *08:02, were associated with protection from RA, and showed no significant associations with autoantibody seropositivity.
Discussion: This study provides a comprehensive overview of HLA-DRB1 allele distribution in the Chilean population, identifying both well-known and novel allele associations with RA susceptibility, protection, and disease activity.
1 Introduction
Rheumatoid arthritis (RA) is a systemic autoimmune disease, primarily targeting synovial joints, that affects between 0.1% and 2% of the population worldwide (1), and around 0.6% of the population living in Chile (2). The risk of developing RA, as well as the clinical presentation of the disease, is strongly influenced by genetic factors, which show substantial heterogeneity among different ethnicities (3).
The best-characterized gene associated with RA susceptibility and severity is HLA-DRB1, which encodes the β chain of the class II antigen presenting molecule HLA-DR (4, 5). A group of HLA-DRB1 alleles encoding a conserved amino acid sequence at positions 70-74 (QKRAA, QRRAA, or RRRAA) of the third hypervariable region (HVR3) of the HLA-DR β chain, known as “Shared Epitope” (SE), has been widely associated with RA risk (6). Furthermore, this association is limited to a subset of patients that harbor RA-specific anti-citrullinated protein antibodies (ACPA), accounting for the most severe RA cases (7). It has been proposed that SE-containing HLA-DR molecules can accommodate citrulline but not arginine at their positively charged P4 pocket in HVR3, leading to presentation of citrullinated peptides to CD4+ T cells and ultimately to ACPA responses (7). In extensive studies involving populations with European ancestry, the SE alleles found to be most significantly associated with ACPA-positive RA risk follow an effect size hierarchy and are part of the allelic groups HLA-DRB1*04 (*04:01, *04:04, *04:08), HLA-DRB1*10 (*10:01), and HLA-DRB1*01 (*01:01) (8, 9). Likewise, ACPA-positive RA among East Asian populations is associated with HLA-DRB1*04 SE alleles, showing a prominent effect of the *04:05 allele, with HLA-DRB1*10:01, and to a lesser degree with HLA-DRB1*01:01 (10–12). In addition, among East Asian populations, an association of the non-SE allele HLA-DRB1*09:01 with ACPA-positive RA has also been described (12, 13). Meanwhile, the allele *14:02 has been associated with RA in American populations, including Mexicans (14), Peruvians (15), and Native North Americans (16).
It is equally important to recognize alleles that confer protective effects in RA. In this sense, the best-studied HLA-DRB1 protective (PR) alleles for RA are known to encode an aspartic acid at position 70 (D alleles), leading to the sequences DRRAA and DERAA at positions 70-74 (17–19). The PR alleles include members of the HLA-DR*13 group (*13:01, *13:02, and *13:04) and HLA-DR*11 group (*11:01, *11:02, and *11:04), as well as the alleles *01:03, *04:02 and *12:01, which are associated with a low risk of developing RA and a milder disease (20, 21). It has been suggested that this protective effect could reflect a greater efficiency of these alleles to bind the CLIP peptide from the invariant chain (Ii, CD74), which would prevent the premature occupation of the antigen-binding groove by endogenous peptides (22).
Few reports have examined the distribution of HLA-DRB1 alleles in the Chilean population. In these studies, the most frequent alleles were found to be DRB1*03:01, *07:01 (23, 24), *04:03, *08:02, *14:02, and *16:02 (24). Meanwhile, studies involving Chilean RA patients have shown that the most frequent SE alleles among this group are DRB1*04:01 (25), *01:01, and *04:04 (25, 26). Additionally, the SE alleles *04:01, *10:01 (25), *04:04, and *04:08 (26) have been associated with RA in this population. Of note the non-SE HLA-DR9 alleles have also been found to be significantly associated with RA in Chilean individuals (25), particularly among rheumatoid factor (RF) seropositive patients (27). Regarding potential PR alleles, the non-D alleles *07:01 and *08:02 were found to confer protection against ACPA-positive RA in the Chilean population (25).
In this work, we explored the distribution of HLA-DRB1 alleles in cohorts of Chilean RA patients and healthy controls (HC) and their association with susceptibility or resistance to develop the disease, as well as their association with autoantibodies seropositivity and disease activity. As Chile has a diverse ethnical composition, influenced by Native-American, African, and European ancestries, this study was limited to the central Metropolitan and Maule regions, characterized by the highest European and lowest Native-American ancestry proportions (28).
2 Methods
2.1 Patients and controls
To conduct this cross-sectional study, blood samples from RA patients and HC were collected between 2014 and 2024. In total, 367 RA patients were recruited: 129 from Hospital Clínico de la Universidad de Chile (HCUCH) and Hospital del Salvador (HDS) in the Metropolitan region (RA-cohort-I), and 238 from Hospital de Talca (HT) and the Rheumatology Unit of the Health Network at Pontificia Universidad Católica de Chile (PUCH) in the Maule region (RA-cohort-II). All patients fulfilled the 2010 ACR/EULAR criteria for RA diagnosis (29). Patients receiving biological or targeted synthetic disease-modifying anti-rheumatic drugs (DMARDs) for the last six months before recruitment were excluded from this study. Clinical and laboratory data were collected, including gender, age, and seropositivity for anti-cyclic citrullinated peptides (CCP) antibodies (the whole RA-cohort-II was seropositive for anti-CCP) and RF (only for RA-cohort-I). The Disease Activity Score based on 28 joint counts (DAS28) was assessed in RA-cohort-I at the time of sampling, where a DAS28 value below 2.6 indicates remission (30), 2.6-3.2 corresponds to low disease activity, and values above 5.1 indicate high disease activity. Additionally, blood samples were obtained from 623 HC recruited from hospital staff and voluntary donors who self-reported no history of autoimmune disorders or chronic inflammatory diseases. Medical anamneses and simple physical examinations were conducted. Individuals with suspected symptoms or self-reported for autoimmune or chronic inflammatory diseases were excluded. Of these 623 HC, 213 were from the HCUCH and the Biobanco de Tejidos y Fluidos de la Universidad de Chile (BTUCH) in Metropolitan region (HC-cohort-I), and 410 from La casa del donante (LCD) in Talca, Maule region (HC-cohort-II). All subjects signed an informed consent form in accordance with the Declaration of Helsinki, and the study protocol was approved by the Ethics Committees from all institutions involved (Approval N° 077-2013, N° 24–2028 and N° 27–2022 for HCUCH; N° 121–2018 and N° 63–2022 for HDS; N° 42–2021 for BTUCH; and N° 04/2014 for HT, PUCH and LCD).
2.2 DNA isolation
DNA from RA-cohort-I and HC-cohort-I was isolated from whole blood samples according to the salting-out procedure (31). Briefly, blood was mixed with lysis buffer (10 mM Tris, 10 mM KCl, 10 mM MgCl2, and 2 mM EDTA, pH 7.6, in the presence of 2.5% Triton X-100). After centrifugation, the pellet was washed and incubated at 55 °C in a buffer containing 10 mM Tris, 10 mM KCl, 10 mM MgCl2, 0.4 M NaCl, and 2 mM EDTA (pH 7.6), along with 10% SDS and proteinase K (10 µg/µL). After 10 min, 6 M NaCl was added to precipitate proteins, and the DNA-containing supernatant was transferred to a fresh tube for precipitation using ethanol. Genomic DNA from RA-cohort-II and HC-cohort-II was extracted using the GeneJET Genomic DNA purification kit #K0722 (ThermoFischer Scientific, Waltham, MA, USA), following the manufacturer’s protocols. The concentration of the purified DNA was measured using a BioTek Synergy spectrophotometer (Agilent, Santa Clara, CA, USA). Yields above 100 ng/µL with an A260/A280 ratio greater than 1.8 were used for further analysis. DNA integrity was also confirmed by running an aliquot on a 1% w/v agarose gel.
2.3 PCR-SSO analysis
HLA-DRB1 gene typing was performed on DNA samples by PCR-SSO (Sequence Specific Oligonucleotides) reverse technique, using the Lifecodes HLA-DRB1 SSO Typing Kit and following the manufacturer’s instructions (Immucor-Werfen, Waukesha, WI, USA). Briefly, HLA-DRB1 exon 2 was amplified and subsequently hybridized with DNA probes covering the main polymorphic positions of the gene. A Luminex FLEXMAP 3D instrument and the xPonent software (ThermoFischer Scientific) were used for data acquisition. HLA typing analysis was achieved using the MATCH IT! DNA version 1.3 software suite (Immucor-Werfen). Intermediate resolution HLA typing results obtained by this technique provide an approximation of the most probable alleles (though not offering definitive high-resolution confirmation) in accordance with the classification of an allele as common (frequency ≥1 in 10,000) (32).
2.4 Statistical analysis
Allele frequencies were compared between groups using the Chi-square or Fisher’s exact tests. Odds ratios (ORs) were calculated with 95% confidence intervals (CI) in order to study associations between the presence of the disease or seropositivity in patients, with the presence of a specific HLA-DRB1 allele or a group of HLA-DRB1 alleles. The normality of continuous variables was evaluated by the Kolmogorov-Smirnov test. Comparisons between continuous variables were performed using the unpaired T-test. A p-value less than 0.05 was considered statistically significant. IBM SPSS Statistics version 29.0.2.0 (New York, NY, USA) was used for statistical analysis. GraphPad Prism version 8.3.0 (Boston, MA, USA) was used for the preparation of graphs.
3 Results
3.1 Characteristics of the study populations
The most relevant characteristics of the RA and HC cohorts included in the study are shown in Table 1. Participants were predominantly female (84% and 74% of RA patients and HC, respectively), with ages of 49 ± 12 years for patients and between 39 ± 9 years for controls. While all patients from RA-cohort-II were seropositive for anti-CCP, most patients from RA-cohort-I were seropositive for anti-CCP (54%) and RF (68%) and exhibited an active disease (70% had a DAS28 > 2.6).

Table 1. Main characteristics of RA patients and HC.
3.2 Frequencies of SE alleles in Chilean RA patients and HC
Subjects presenting at least one of the following alleles were considered positive for the SE: HLA-DRB1*01:01, *01:02, *04:01, *04:04, *04:05, *04:08, *04:10, *10:01, *14:02, *14:06, and *14:13 (21). When it was not possible to discriminate between *04:04, *04:05, *04:08, and *04:10 alleles, carriers were nonetheless considered positive for the SE.
A total of 64.8% of RA patients carried at least one copy of the SE, versus 40.8% of HC (OR=2.7; 95% CI=2.1-3.5; p<0.001). When SE allele dosage was analyzed, 44.9% of RA patients and 38.7% of HC carry only one SE copy (OR=1.3; 95% CI=1.0-1.7; p=0.053), while the presence of two SE copies was detected in 19.9% of RA patients and 2.1% of HC (OR=11.7; 95% CI=6.4-21.4; p<0.001) (Figure 1, Table 2).
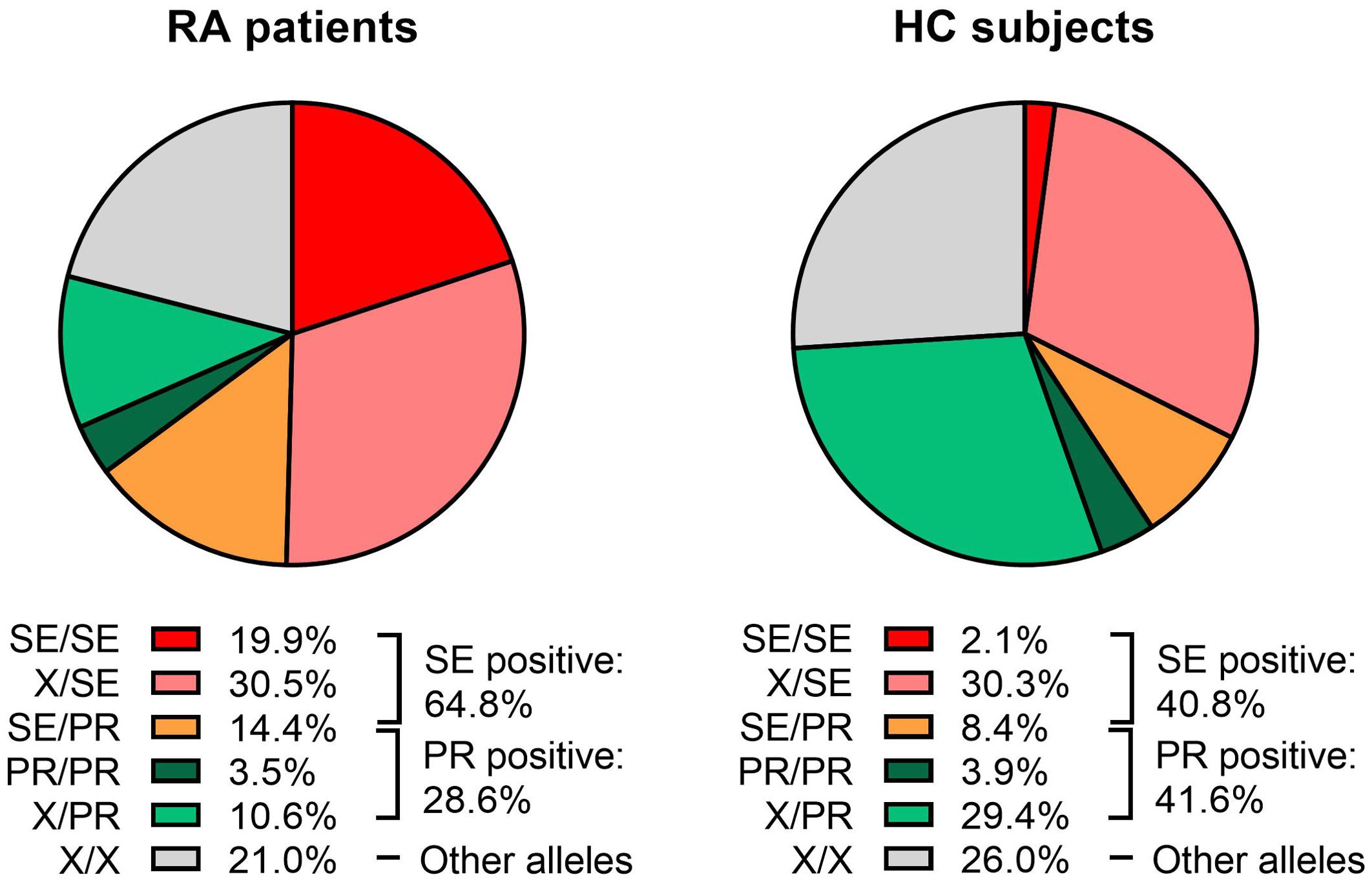
Figure 1. Distribution of HLA-DRB1 genotypes in rheumatoid arthritis (RA) patients and healthy controls (HC). Each pie chart is divided according to the presence or absence of “Shared Epitope” (SE) and Protective (PR) alleles. The following genotype combinations are depicted: SE/SE (red), X/SE (pink), SE/PR (orange), PR/PR (dark green), X/PR (light green), and X/X (gray), where “X” denotes an allele that is neither SE nor PR. Percentages indicate the proportion of subjects within each category. RA patients (left pie chart) exhibited higher frequencies of SE-containing genotypes than HC (right pie chart). The overall percentages of SE-positive and PR-positive individuals are provided to the right of each pie chart.
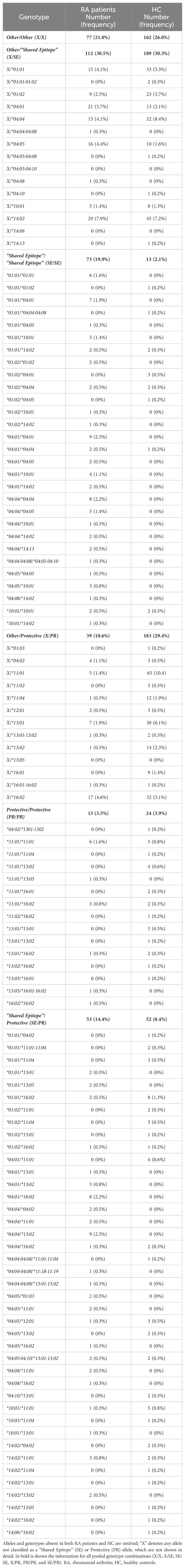
Table 2. HLA-DRB1 genotype frequencies in a Chilean cohort of RA patients and HC.
Specific genotype frequencies in RA patients and HC are shown in Table 2. HLA-DR*04:01 was the most frequent SE allele among RA patients, followed by *04:04 and *14:02 (16.1%, 13.9% and 11.7% of all patients, respectively). Meanwhile, the most frequent SE allele in HC was *04:04, followed by *14:02 and *01:01 (9.2%, 8.8% and 8.2% of all individuals, respectively) (Figure 2, Table 3).
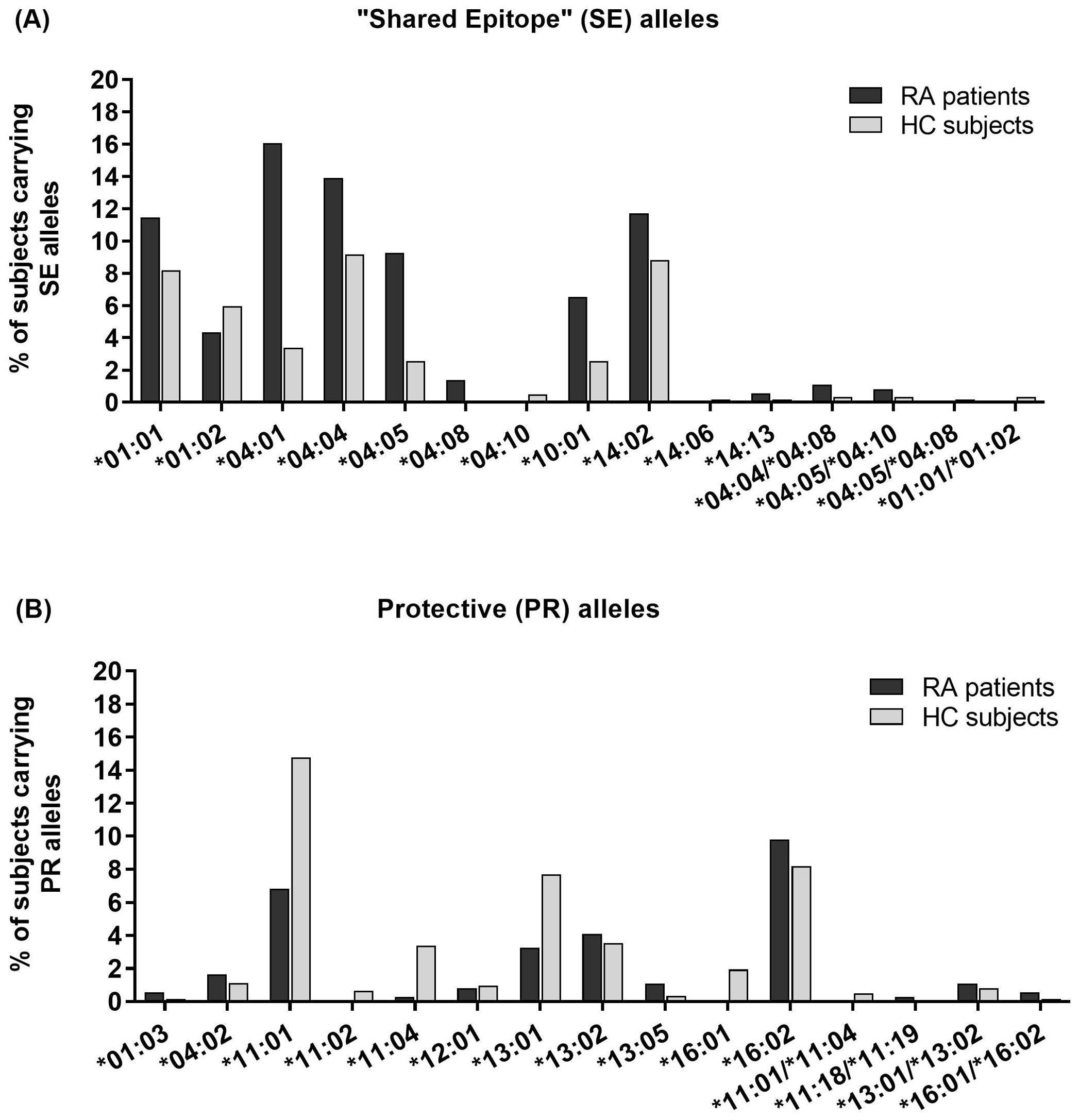
Figure 2. Percentage of rheumatoid arthritis (RA) patients and healthy controls (HC) carrying each (A) “Shared Epitope” (SE) or (B) Protective (PR) HLA-DRB1 allele. The x-axis lists each allele, while the y-axis indicates the percentage of subjects carrying that allele. Black bars represent RA patients, and gray bars represent HC. Alleles not detected in either cohort are excluded.
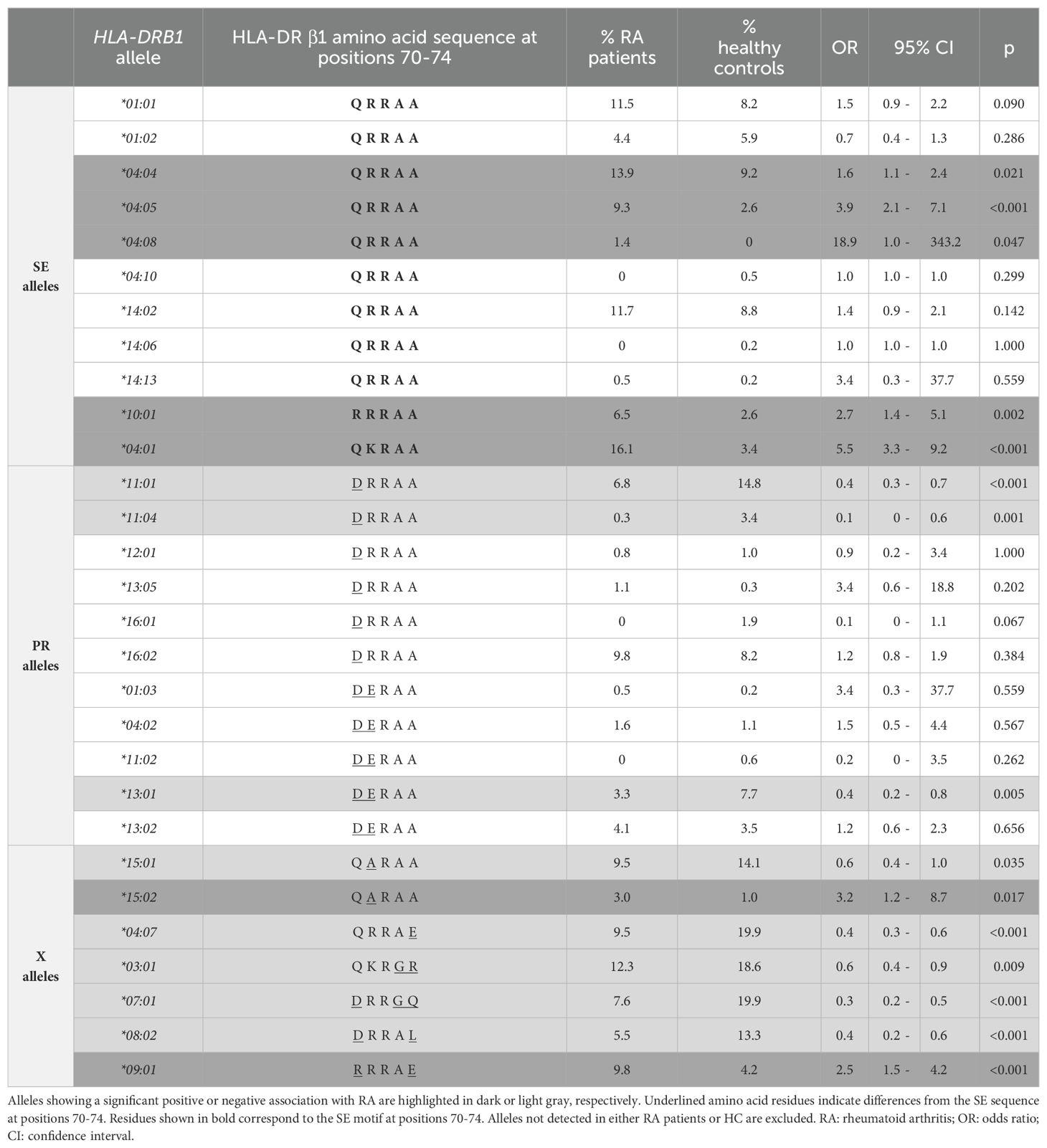
Table 3. Association between RA and HLA-DRB1 genotypes carrying at least one copy of a “Shared Epitope” (SE) allele, protective (PR) allele, or non-SE/non-PR (X) allele, in Chilean individuals.
The association between the disease and the presence of a specific HLA-DRB1 allele was calculated considering the frequency of RA subjects positive and negative for at least one copy of the allele and the correlative data for HC. The SE alleles that showed a significant association with RA were: *04:01 (OR=5.5; 95% CI=3.3-9.2; p<0.001), *04:04 (OR=1.6; 95% CI=1.1-2.4; p=0.021), *04:05 (OR=3.9; 95% CI=2.1-7.1; p<0.001), *04:08 (OR=18.9; 95% CI=1.0-343.2; p=0.047), and *10:01 (OR=2.7; 95% CI=1.4-5.1; p=0.002) (Table 3). In addition, two non-SE alleles showed a significant association with the disease, *09:01 (OR=2.5; 95% CI=1.5-4.2; p<0.001) and *15:02 (OR=3.2; 95% CI=1.2-8.7; p=0.017) (Table 3).
3.3 Frequencies of PR alleles in Chilean RA patients and HC
The frequencies of HLA-DRB1 alleles associated with protection against RA were also analyzed. The following alleles were considered PR alleles: HLA-DRB1*01:03, *04:02, *11:01, *11:02, *11:04, *12:01, *13:01, *13:02, *13:05, *16:01, and *16:02 (21). When it was impossible to discriminate between *13:01, *13:02, *16:01, and *16:02 alleles, carriers were nonetheless considered positive for PR alleles.
At least one copy of a PR allele was found in 28.6% of RA patients and 41.6% of HC (OR=0.6; 95% CI=0.4-0.7; p<0.001). The presence of one PR copy was detected in 25.1% of RA patients and 37.7% of HC (OR=0.6; 95% CI=0.4-0.7; p<0.001), while the frequency of two PR copies was 3.5% for RA patients and 3.9% for HC (OR=0.9; 95% CI=0.5-1.8; p=0.804) (Figure 1, Table 2).
HLA-DRB1*11:01 was the most frequent PR allele among HC, followed by *16:02 and *13:01 (15.2%, 8.2%, and 7.7% of all HC, respectively), while in RA patients the most frequent PR allele was *16:02, followed by *11:01 and *13:02 (9.8%, 6.8%, and 4.1% of all patients, respectively) (Figure 2, Table 2).
The PR alleles that showed a significative association with the absence of the disease (protective effect) were HLA-DRB1*11:01 (OR=0.4; 95% CI=0.3-0.7; p<0.001), *11:04 (OR=0.1; 95% CI=0-0.6; p=0.001), and *13:01 (OR=0.4; 95% CI=0.2-0.8; p=0.005) (Table 3). Notably, some alleles which do not code for the DRRAA or DERAA sequences also showed significant association with protection against RA: *15:01 (OR=0.6; 95% CI=0.4-1.0; p=0.035), *04:07 (OR=0.4; 95% CI=0.3-0.6; p<0.001), *03:01 (OR=0.6; 95% CI=0.4-0.9; p=0.009), *07:01 (OR=0.3; 95% CI=0.2-0.5; p<0.001), and *08:02 (OR=0.4; 95% CI=0.2-0.6; p<0.001) (Table 3).
3.4 Association between HLA-DRB1 alleles and clinical parameters in Chilean RA patients
Because all patients in the RA-cohort-II were anti-CCP positive and only RF and DAS28 data from RA-cohort-I were available, only the latter was used to study the relationship between clinical parameters and the presence of HLA-DRB1 alleles.
The association between circulating autoantibodies and the presence of a specific HLA-DRB1 allele was calculated considering the frequency of seropositivity and seronegativity among RA subjects. No significant associations were found between anti-CCP (OR=1.8; 95% CI=0.6-5.5; p=0.31) or RF seropositivity (OR=0.5; 95% CI=0.2-1.6; p=0.235) and the presence of at least one copy of SE alleles, or for most SE alleles assessed individually (data not shown). However, the SE allele *14:02 showed a significant association with anti-CCP seropositivity among RA patients (OR=1.4; 95% CI=1.2-1.6; p=0.033; Supplementary Table 1). On the other hand, the SE allele *01:01 showed a significant association with RF seronegativity (OR=0.2; 95% CI=0.1-0.8; p=0.024), while the non-SE allele *13:03 was associated with anti-CCP seronegativity (OR=0.1; 95% CI=0-0.6; p=0.017; Supplementary Table 1).
The presence of at least one copy of PR alleles also showed no association with anti-CCP (OR=0.9; 95% CI=0.3-2.8; p=0.767) or RF (OR=2.5; 95% CI=0.7-9.5; p=0.156) seropositivity among RA patients.
Finally, no associations were found between DAS28 values and the presence of at least one copy of SE (p=0.595) or PR (p=0.838) alleles in the analyzed RA cohort. However, we observed that low DAS28 values were significantly associated with the presence of the SE allele *04:04 (mean value of 2.8 versus 4.1 for *04:04 positive and negative RA patients, respectively; p=0.028; Supplementary Table 2).
4 Discussion
Although RA is a multifactorial disease, the presence of HLA-DRB1 alleles encoding the SE remains the most important genetic risk factor (21). To date, a detailed study of the distribution of HLA-DRB1 alleles in Chilean RA patients has not been addressed. This is crucial information to better understand the genetic landscape of RA in the country and the region, and the pathogenic mechanisms underlying disease development in this ethnically mixed population.
In the present study, we observed a higher frequency of SE alleles among Chilean RA patients (64.8%) than HC (40.8%). The proportion of RA patients carrying at least one SE allele was higher than that reported in Asian and African populations (<50%) (10, 33, 34) but lower than in Caucasian patients (>80%) (35, 36), aligning more closely with data from North and South American cohorts (50-75%) (14, 37, 38). We also found that when the SE is expressed in a double dose, the risk of developing the disease is higher, as previously described (10). In our examination of the Chilean population, *04:01 was the most frequent SE allele among RA patients, followed by *04:04 and *14:02, which is in agreement with prior reports of high *04:01 and *04:04 frequencies (25, 26). We also noted significant associations between RA and alleles *04:01, *04:04, *04:05, *04:08, and *10:01. Except for *04:05, these findings align with previous studies in Chilean populations (25, 26). The non-SE allele *09:01 was also linked to RA in our cohort, consistent with studies in Chilean (25) and Asian populations (39). This allele has also been implicated in other autoimmune diseases such as systemic lupus erythematosus (39) and type 1 diabetes (40). Although *09:01 differs from *10:01 by a single amino acid substitution (alanine for glutamic acid at position 74), there is no conclusive evidence that the RRRA sequence at positions 70–73 alone confers susceptibility to RA. However, *09:01 is in linkage disequilibrium with HLA-DRB4, a paralog of HLA-DRB1 that also encodes for an HLA-DRβ chain. HLA-DRB4, found exclusively in DR4, DR7, and DR9 haplotypes, has been associated with a more aggressive RA progression (41). Our cohort also revealed an association between *15:02 and RA. Although DRB1*15 alleles have traditionally been considered as low risk alleles (42), they have been linked to pulmonary complications in RA (43, 44). Future studies with comprehensive records of extra-articular manifestations of the disease would help to clarify this association.
Interestingly, the *14:02 allele emerged as over-represented in our cohorts, ranking as the third most frequent allele in RA patients and the second in HC. Notably, the *14:02 allele has been previously associated with RA in various American populations, including Native Americans (16), Mexicans (14), and Peruvians (15). A study in Indigenous North Americans showed that the *14:02 allele confers substantial risk for RA due to a combination of serine residues at positions 11 and 13, which would allow HLA-DR to accommodate both citrulline and arginine at the P4 pocket, potentially enhancing anti-citrullinated antigen T cell responses and ACPA development (45). In line with these observations, we found that the *14:02 allele was significantly associated with the presence of anti-CCP autoantibodies.
Although many studies attribute the strong association between HLA and anti-CCP RA disease to the presence of the SE in the HLA-DR molecule, other pieces of evidence suggest that the SE hypothesis alone would not fully explain HLA-DRB1 associations with the disease. Indeed, five amino acid positions across three HLA genes (positions 11, 71, and 74 in HLA-DRB1, as well as position 9 in HLA-B and HLA-DPB1) have been shown to collectively explain most of the HLA region’s contribution to seropositive RA in Europeans (8). Conditional haplotype analyses demonstrated that B*08/Asp-9 and/or HLA-DPB1/Phe-9 alleles increase the RA risk on specific HLA-DRB1 backgrounds such as *09:01 and *15:02, with position 11 in HLA-DRB1 being determinant to this effect: while Val-11 and Leu-11 confer high risk, Ser-11 is highly protective for the disease. This is particularly interesting in light of our dataset, where SE alleles *04:04, *04:05, *04:08, *10:01, and *04:01 (all containing Val or Leu at position 11) were associated with RA risk, while *03:01, *08:02, *11:01, *11:04, and *13:01 (all containing Ser-11) showed protective effects. Studies focusing on Asian and European populations with ACPA-positive RA have also linked His-13 to increased RA risk, which is reinforced by our results regarding *04:04, *04:05, and *04:01 alleles (12). The risk effect of Val-11 and His-13, and the protective effect of Ser-11, have been confirmed in autoantibody-positive African RA patients as well (46). In Japanese patients with early RA, *04:05 (encoding Val at position 11) correlated with higher disease risk and anti-CCP seropositivity, whereas *09:01 (encoding Asp at position 11) appeared more commonly in anti-CCP-negative patients, underscoring how position 11 variants can differentially influence seropositivity (34).
On the protective side, PR alleles were more frequent in HC (41.6%) than in RA patients (28.6%), and their presence was negatively correlated with the disease, confirming previous reports (19). Specifically, *11:01, *11:04, and *13:01 alleles were negatively associated with RA, which is consistent with observations in Argentinian (47) and European ACPA-positive cohorts (48). Interestingly, additional alleles not classically categorized as protective (*15:01, *04:07, *03:01, *07:01, and *08:02) also showed negative associations with RA. Previous reports in Caucasian RA populations have attributed the protective role of *07:01 and *15:01 to the presence of an isoleucine at position 67 of the HVR3, which is also present in PR alleles *01:03, *04:02, *11:02, *13:01, *13:02, and *12:01 (49). Our results for *07:01 and *08:02 alleles are consistent with previous evidence showing a negative association with ACPA-positive RA in Latin American admixed populations (25). Although *04:07 has been considered neutral for RA (49), our findings suggest a protective effect, possibly linked to a negatively charged glutamic acid at position 74 of the HVR3, similar to that at position 71 in *04:02, and contrasting with the positively charged residues in SE alleles (50). DRB1*03:01 has been associated with ACPA-negative RA but exhibits a protective effect for ACPA-positive RA (51), paralleling our data (where 54% of the RA cohort was ACPA-positive). These findings imply that HLA-DR-mediated protection against RA may not always depend on DRRAA or DERAA sequences at P4. Interestingly, although we did not find a correlation between PR alleles with seronegativity, as the literature suggests for presence of HLA-DR3 or DERAA-encoding alleles (52), we found a significant association between *13:03 (a non-PR allele) and the absence of anti-CCP autoantibodies. Another study examining ACPA-positive versus ACPA-negative RA in European-descent cohorts revealed that Ser-11 and Leu-11 in HLA-DRB1 and Asp9 in HLA-B can drive risk for ACPA-negative RA (51), potentially explaining our findings for *13:03 which also contains Ser-11. Indeed, HLA-DRB1*13 alleles have been noted to affect the ACPA status, conferring protection for ACPA-positive RA and, in combination with HLA-DRB1*03, reducing the risk of ACPA-negative RA (53). It should be noted that only part of our RA cohort was used to explore associations with autoantibody profiles, as RA-cohort-II was entirely ACPA-positive. Consequently, the associations observed in this study, particularly for seronegative patients, should be interpreted with caution and verified in larger cohorts.
Studying the role of HLA alleles in driving autoantibody production and influencing disease outcomes in RA is not easy. Actually, distinct HLA associations for RA subsets have also been explored by specific autoantibody profiles, where Asp-9 in HLA-B has emerged as a shared risk factor for both ACPA-positive and ACPA-negative RA but with variations according to the presence of different autoantibody clusters (54). In this context, HLA-DRB1*03:01, which was revealed to be protective in our cohort, has been identified as a primary driver of HLA-DRB1 associations in second-generation anti-cyclic citrullinated peptide (CCP2) negative RA patients, but not in CCP2-positive RA patients, demonstrating similar patterns to HLA-B/Asp-9 (51, 54). This illustrates how specific alleles and particular amino acid residues may define diverse autoantibody profiles, further refining the categorization of RA beyond simple seropositivity and seronegativity. Nevertheless, it is widely accepted that the evolving autoimmune response in RA is closely linked to the presence of SE-expressing HLA-DR molecules, which are distinguished as exhibiting a high affinity for citrullinated peptides. Within RA-inflamed joints, citrullinated peptides can be recognized as foreign and preferentially presented by SE-expressing HLA-DR to autoreactive CD4+ T cells (55–58). Subsequently, these activated CD4+ T cells could facilitate the activation of autoreactive B cells, culminating in the production and secretion of autoantibodies, the hallmark of RA (59–62).
Although most of the HLA-DRB1 effect on disease severity is mediated by ACPA, recent evidence has shown that specific HLA-DRB1 amino acids can also modulate inflammatory markers (like C-reactive protein) and clinical parameters (e.g., DAS28 or the Swollen Joint Count) through different pathways, some of which are likely to be independent of ACPA (5, 63). For example, large-scale research in Japanese RA patients found that DRB1*04:05 is significantly associated with radiographic damage (Sharp/van der Heijde score) in anti-CCP-positive RA, even after adjusting for the DAS28, implying subclinical mechanisms not fully captured by conventional disease activity metrics (64, 65). In a separate Caucasian RA cohort, haplotypes containing HLA-DRB1*04:04 and HLA-DRB1*01:01, along with six single-nucleotide polymorphisms in the lymphotoxin alpha and tumor necrosis factor DNA-coding region, correlated strongly with therapeutic response in early RA (66). Although in our dataset HLA-DRB1*04:04 appears to be associated with lower DAS28 values, which is at odds with the literature, this should also be taken with caution given the small sample size of patients with available DAS-28 data in our cohort.
Despite the strengths of our study, several limitations should be acknowledged. First, the HC were selected from a general population cohort that was not screened for autoimmune laboratory tests, potentially allowing for subclinical autoimmune conditions to go undetected. Second, the PCR-SSO method only provided intermediate-resolution typing of HLA-DRB1 alleles, meaning that the reported associations could not be confirmed at the highest possible resolution; however, according to comparative studies, PCR-SSO and next generation sequencing (NSG) methodologies have an acceptable concordance for HLA-DRB1 alleles (67, 68). Third, risk-factor information, including tobacco index and biomass-burning smoke exposure, were unavailable for some participants, thereby restricting the scope of our analyses, in particular given the well-described interaction between tobacco smoking, SE alleles and ACPA in RA (60). A further limitation related to our autoantibody analyses was the lack of quantitative data for anti-CCP and RF antibodies, which prevented us from exploring potential associations between antibody levels and specific HLA-DRB1 alleles. Finally, the cross-sectional design prevented us from performing longitudinal analysis between genetic factors and disease progression. Future investigations incorporating more thoroughly screened populations, high-resolution genotyping approaches, expanded clinical and risk-factor data collection, and longitudinal follow-up will be essential to address these issues.
Considering the clear variations in allele frequencies among different ethnic groups, ongoing research is essential to refine the understanding of how genetics may influence RA establishment and evolution. In this context, our findings provide updated data on HLA-DRB1 distributions in the Chilean population, information that is not only valuable for understanding the disease immunopathogenesis but also holds promise for the development of personalized approaches to RA prevention and treatment.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statement
The studies involving humans were approved by Comité de Ética de Investigación en Seres Humanos, Facultad de Medicina, Universidad de Chile; Comité Ético Científico o de Investigación, Hospital Clínico, Universidad de Chile; Comité de Ética Científico Adultos, Hospital del Salvador. Comité de Ética del Servicio de Salud del Maule. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
DC: Writing – review & editing, Writing – original draft, Formal analysis, Visualization, Validation, Conceptualization, Supervision. LS: Supervision, Investigation, Writing – review & editing, Visualization. ÓN: Supervision, Writing – review & editing, Investigation, Visualization. MC-G: Visualization, Investigation, Writing – review & editing. RD-P: Writing – review & editing, Methodology, Funding acquisition, Resources. OA: Methodology, Investigation, Writing – review & editing. EP: Writing – review & editing, Methodology. MC: Investigation, Writing – review & editing, Visualization. JA: Project administration, Writing – original draft, Visualization, Investigation, Funding acquisition, Writing – review & editing, Supervision, Resources, Conceptualization. JM: Validation, Writing – review & editing, Formal analysis, Software, Supervision, Data curation, Methodology, Writing – original draft, Conceptualization, Visualization, Investigation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by following projects: Fondef-IDeA ID15I10080, ID15I20080, ID18I10243, REDES 180028, Fondecyt 1181853, Fondecyt 1221611, Fondecyt 1220540 and Fondecyt 1240060, all granted by Agencia Nacional de Investigación y Desarrollo (ANID), Chile. Doctoral training of JM was supported by ANID-PFCHA/National Doctoral Scholarship 2018/N&z.ousco;21181538.
Acknowledgments
We acknowledge the patients’ organization Corporación Pro-Ayuda al Enfermo Reumático (VOLAR) and to Biobanco de Tejidos y Fluidos from Universidad de Chile. We also thank Verónica Ramos, Elizabeth Rivas, and Patricio Leiva for their contribution on the standardization of conventional PCRs that laid the foundations for the development of this study, and to Ms. Nancy Fabres, Jennifer Salvatierra, Juana Orellana, and Bárbara Vivanco for their kind assistance to collect samples.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1594723/full#supplementary-material
References
1. Shapira Y, Agmon-Levin N, Shoenfeld Y. Geoepidemiology of autoimmune rheumatic diseases. Nat Rev Rheumatol. (2010) 6:468–76. doi: 10.1038/nrrheum.2010.86
2. Durán J, Massardo L, Llanos C, Iacobelli S, Burgos P, Cisternas M, et al. The Prevalence of rheumatoid arthritis in Chile: A nationwide study performed as part of the national health survey (ENS 2016-17). J Rheumatol. (2019) 47(7):951–8. doi: 10.3899/jrheum.190396
3. Kochi Y, Suzuki A, Yamada R, Yamamoto K. Ethnogenetic heterogeneity of rheumatoid arthritis. Implications pathogenesis. Nat Rev Rheumatol. (2010) 6:290–5. doi: 10.1038/nrrheum.2010.23
4. Eyre S, Bowes J, Diogo D, Lee A, Barton A, Martin P, et al. High-density genetic mapping identifies new susceptibility loci for rheumatoid arthritis. Nat Genet. (2012) 44:1336—1340. doi: 10.1038/ng.2462
5. Viatte S, Plant D, Han B, Fu B, Yarwood A, Thomson W, et al. Association of HLA-DRB1 haplotypes with rheumatoid arthritis severity, mortality, and treatment response. JAMA - J Am Med Assoc. (2015) 313:1645–56. doi: 10.1001/jama.2015.3435
6. Gregersen PK, Silver J, Winchester RJ. The shared epitope hypothesis. An approach to understanding the molecular genetics of susceptibility to rheumatoid arthritis. Arthritis Rheum. (1987) 30:1205–13. doi: 10.1002/art.1780301102
7. van der Helm-van Mil AH, Verpoort KN, Breedveld FC, Huizinga TW, Toes RE, de Vries RR. The HLA-DRB1 shared epitope alleles are primarily a risk factor for anti-cyclic citrullinated peptide antibodies and are not an independent risk factor for development of rheumatoid arthritis. Arthritis Rheum. (2006) 54:1117–21. doi: 10.1002/art.21739
8. Raychaudhuri S, Sandor C, Stahl EA, Freudenberg J, Lee HS, Jia X, et al. Five amino acids in three HLA proteins explain most of the association between MHC and seropositive rheumatoid arthritis. Nat Genet. (2012) 44:291–6. doi: 10.1038/ng.1076
9. van der Woude D, Rantapaa-Dahlqvist S, Ioan-Facsinay A, Onnekink C, Schwarte CM, Verpoort KN, et al. Epitope spreading of the anti-citrullinated protein antibody response occurs before disease onset and is associated with the disease course of early arthritis. Ann Rheum Dis. (2010) 69:1554–61. doi: 10.1136/ard.2009.124537
10. Chun-Lai T, Padyukov L, Dhaliwal JS, Lundström E, Yahya A, Muhamad NA, et al. Shared epitope alleles remain a risk factor for anti-citrullinated proteins antibody (ACPA) - positive rheumatoid arthritis in three Asian ethnic groups. PloS One. (2011) 6:1–9. doi: 10.1371/journal.pone.0021069
11. Tan LK, Too CL, Diaz-Gallo LM, Wahinuddin S, Lau IS, Heselynn H, et al. The spectrum of association in HLA region with rheumatoid arthritis in a diverse Asian population: evidence from the MyEIRA case-control study. Arthritis Res Ther. (2021) 23(1):46. doi: 10.1186/s13075-021-02431-z
12. Okada Y, Kim K, Han B, Pillai NE, Ong RTH, Saw WY, et al. Risk for ACPA-positive rheumatoid arthritis is driven by shared HLA amino acid polymorphisms in Asian and European populations. Hum Mol Genet. (2014) 23:6916–26. doi: 10.1093/hmg/ddu387
13. Okada Y, Suzuki A, Ikari K, Terao C, Kochi Y, Ohmura K, et al. Contribution of a non-classical HLA gene, HLA-DOA, to the risk of rheumatoid arthritis. Am J Hum Genet. (2016) 99:366–74. doi: 10.1016/j.ajhg.2016.06.019
14. Del Rincón I, Escalante A. HLA-DRB1 alleles associated with susceptibility or resistance to rheumatoid arthritis, articular deformities, and disability in Mexican Americans. Arthritis Rheum. (1999) 42:1329–38. doi: 10.1002/1529-0131(199907)42:7<1329::AID-ANR5>3.0.CO;2-1
15. Castro F, Acevedo E, Ciusani E, Angulo JA, Wollheim FA, Sandberg-Wollheim M. Tumour necrosis factor microsatellites and HLA-DRB1*, HLA-DQA1*, and HLA-DQB1* alleles in Peruvian patients with rheumatoid arthritis. Ann Rheum Dis. (2001) 60:791–5. doi: 10.1136/ard.60.8.791
16. Williams RC, Jacobsson LT, Knowler WC, del Puente A, Kostyu D, McAuley JE, et al. Meta-analysis reveals association between most common class II haplotype in full-heritage native americans and rheumatoid arthritis. Hum Immunol. (1995) 42:90–4. doi: 10.1016/0198-8859(94)00079-6
17. Gourraud PA, Boyer JF, Barnetche T, Abbal M, Cambon-Thomsen A, Cantagrel A, et al. A new classification of HLA-DRB1 alleles differentiates predisposing and protective alleles for rheumatoid arthritis structural severity. Arthritis Rheum. (2006) 54:593–9. doi: 10.1002/art.21630
18. Mattey DL, Hassell AB, Plant MJ, Cheung NT, Dawes PT, Jones PW, et al. The influence of HLA-DRB1 alleles encoding the DERAA amino acid motif on radiological outcome in rheumatoid arthritis. Rheumatology. (1999) 38:1221–7. doi: 10.1093/rheumatology/38.12.1221
19. Shadick NA, Heller JE, Weinblatt ME, Maher NE, Cui J, Ginsburg G, et al. Opposing effects of the D70 mutation and the shared epitope in HLA-DR4 on disease activity and certain disease phenotypes in rheumatoid arthritis. Ann Rheum Dis. (2007) 66:1497–502. doi: 10.1136/ard.2006.067603
20. Van Der Helm-Van Mil AHM, Huizinga TWJ, Schreuder GMT, Breedveld FC, De Vries RRP, Toes REM. An independent role of protective HLA class II alleles in rheumatoid arthritis severity and susceptibility. Arthritis Rheum. (2005) 52:2637–44. doi: 10.1002/art.21272
21. Holoshitz J. The rheumatoid arthritis HLA-DRB1 shared epitope. Curr Opin Rheumatol. (2010) 22:293–8. doi: 10.1097/BOR.0b013e328336ba63
22. Kirschmann DA, Duffin KL, Smith CE, Welply JK, Howard SC, Schwartz BD, et al. Naturally processed peptides from rheumatoid arthritis associated and non-associated HLA-DR alleles. J Immunol. (1995) 155:5655. Available online at: http://www.jimmunol.org/content/155/12/5655.abstract (Accessed May 2, 2024).
23. Castro-Santos P, Olloquequi J, Díaz-Peña R. HLA-A, B, C and DRB1 alleles in a Chilean population from Talca. HLA. (2019) 95:200–3. doi: 10.1111/tan.13775
24. Rey D, Juan CP, Cristina M, Arnaiz-villena SAJAA. HLA genetic profile of Mapuche (Araucanian) AmerIndians from Chile. Mol Biol Rep. (2013) 40:4257–67. doi: 10.1007/s11033-013-2509-3
25. Santos PC, Olloquequi J, Verdugo RA, Gutiérrez MA, Pinochet C, Quiñones LA, et al. HLA-DRB1*07:01 and *08:02 alleles confer a protective effect against ACPA-positive rheumatoid arthritis in a Latin American admixed population. Biol (Basel). (2020) 9:1–11. doi: 10.3390/biology9120467
26. González A, Nicovani S, Massardo L, Aguirre V, Cervilla V, Lanchbury JS, et al. Influence of the HLA-DR shared epitope on susceptibility to and clinical expression of rheumatoid arthritis in Chilean patients. Ann Rheum Dis. (1997) 56:191–3. doi: 10.1136/ard.56.3.191
27. Gonzalez A, Nicovani S, Massardo L, Bull P, Rodriguez L, Jacobelli S. Novel genetic markers of rheumatoid arthritis in Chilean patients, by serotyping and restriction fragment length polymorphism analysis. Arthritis Rheum. (1992) 35(3):282–9. doi: 10.1002/art.1780350306
28. Eyheramendy S, Martinez FI, Manevy F, Vial C, Repetto GM. Genetic structure characterization of Chileans reflects historical immigration patterns. Nat Commun. (2015) 6:6472. doi: 10.1038/ncomms7472
29. Kay J, Upchurch KS. ACR/EULAR 2010 rheumatoid arthritis classification criteria. Rheumatology. (2012) 51:vi5–9. doi: 10.1093/rheumatology/kes279
30. Fransen J, Creemers MCW, Van Riel PLCM. Remission in rheumatoid arthritis: Agreement of the disease activity score (DAS28) with the ARA preliminary remission criteria. Rheumatology. (2004) 43:1252–5. doi: 10.1093/rheumatology/keh297
31. Miller SA, Dykes DD, Polesky HF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. (1988) 16:1215. doi: 10.1093/nar/16.3.1215
32. Hurley CK, Kempenich J, Wadsworth K, Sauter J, Hofmann JA, Schefzyk D, et al. Common, intermediate and well-documented HLA alleles in world populations: CIWD version 3.0.0. HLA. (2020) 95:516–31. doi: 10.1111/tan.13811
33. Singwe-Ngandeu M, Finckh A, Bas S, Tiercy JM, Gabay C. Diagnostic value of anti-cyclic citrullinated peptides and association with HLA-DRB1 shared epitope alleles in African rheumatoid arthritis patients. Arthritis Res Ther. (2010) 12(2):R36. doi: 10.1186/ar2945
34. Furuya T, Hakoda M, Ichikawa N, Higami K, Nanke Y, Yago T, et al. Differential association of HLA-DRB1 alleles in Japanese patients with early rheumatoid arthritis in relationship to autoantibodies to cyclic citrullinated peptide. Clin Exp Rheumatol. (2007) 25:219–24.
35. Rønningen KS, Spurkland A, Egeland T, Iwe T, Munthe E, Vartdal F, et al. Rheumatoid arthritis may be primarily associated with HLA-DR4 molecules sharing a particular sequence at residues 67–74. Tissue Antigens. (1990) 36:235–40. doi: 10.1111/j.1399-0039.1990.tb01834.x
36. González-Gay M, García-Porrua C, Hajeer AH. Influence of human leukocyte antigen-DRB1 on the susceptibility and severity of rheumatoid arthritis. Semin Arthritis Rheum. (2002) 31:355–60. doi: 10.1053/sarh.2002.32552
37. Anaya JM, Correa PA, Mantilla RD. Rheumatoid arthritis association in Colombian population is restricted to HLA-DRB1*04 QRRAA alleles. Genes Immun. (2002) 3(1):56–8. doi: 10.1038/sj/gene/6363833
38. Usnayo MJG, Andrade LEC, Alarcon RT, Oliveira JC, Silva GMF, Bendet I, et al. Study of the frequency of HLA-DRB1 alleles in Brazilian patients with rheumatoid arthritis. Rev Bras Reumatol. (2011) 51:465–83. doi: 10.1590/S0482-50042011000500007
39. Shimane K, Kochi Y, Suzuki A, Okada Y, Ishii T, Horita T, et al. An association analysis of HLA-DRB1 with systemic lupus erythematosus and rheumatoid arthritis in a Japanese population : effects of * 09 : 01 allele on disease phenotypes. Rheumatology. (2013) 52(7):1172–82. doi: 10.1093/rheumatology/kes427
40. Yamashita H, Awata T, Kawasaki E, Ikegami H, Tanaka S, Maruyama T, et al. Analysis of the HLA and non-HLA susceptibility loci in Japanese type 1 diabetes. Diabetes Metab Res Rev. (2011) 27:844–8. doi: 10.1002/dmrr
41. Heldt C, Listing J, So O, Bla F, Frischbutter S, Mu B. Differential expression of HLA class II genes associated with disease susceptibility and progression in rheumatoid arthritis. Arthritis Rheum. (2003) 48:2779–87. doi: 10.1002/art.11251
42. Tezenas Du Montcel S, Michou L, Petit-Teixeira E, Osorio J, Lemaire I, Lasbleiz S, et al. New classification of HLA-DRB1 alleles supports the shared epitope hypothesis of rheumatoid arthritis susceptibility. Arthritis Rheum. (2005) 52:1063–8. doi: 10.1002/art.20989
43. Mori S, Koga Y, Sugimoto M. Different risk factors between interstitial lung disease and airway disease in rheumatoid arthritis. Respir Med. (2012) 106:1591–9. doi: 10.1016/j.rmed.2012.07.006
44. Migita K, Nakamura T, Koga T, Eguchi K. HLA-DRB1 alleles and rheumatoid arthritis-related pulmonary fibrosis. J Rheumatol. (2010) 37:205–7. doi: 10.3899/jrheum.090303
45. Scally SW, Law SC, Ting YT, Van Heemst J, Sokolove J, Deutsch AJ, et al. Molecular basis for increased susceptibility of Indigenous North Americans to seropositive rheumatoid arthritis. Ann Rheum Dis. (2017) 76:1915–23. doi: 10.1136/annrheumdis-2017-211300
46. Govind N, Reynolds RJ, Hodkinson B, Ickinger C, Ramsay M, Bridges SL, et al. HLA-DRB1 amino acid positions and residues associated with antibody-positive rheumatoid arthritis in black South Africans. J Rheumatol. (2019) 46:138–44. doi: 10.3899/jrheum.180107
47. Citera G, Padulo LA, Fernandez G, Lazaro MA, Rosemffet MG, Maldonado Cocco JA. Influence of HLA-DR alleles on rheumatoid arthritis: Susceptibility and severity in Argentine patients. J Rheumatol. (2001) 28:1486–91.
48. Van Der Woude D, Lie BA, Lundstro E, Balsa A, Feitsma AL, Houwing-duistermaat JJ, et al. Protection against anti – citrullinated protein antibody – positive rheumatoid arthritis is predominantly associated with A meta-analysis of HLA – DRB1 associations with anti – citrullinated protein antibody – positive and anti – citrullinated protein ant. Arthritis Rheum. (2010) 62:1236–45. doi: 10.1002/art.27366
49. de Vries N, Tijssen H, van Riel PLCM, van de Putte LBA. Reshaping the Shared Epitope Hypothesis: HLA-associated risk for rheumatoid arthritis is encoded by amino acid substitutions at positions 67–74 of the HLA-DRB1 molecule. Arthritis Rheum. (2002) 46:921–8. doi: 10.1002/art.10210
50. Davenport MP, Godkin A, Friede T, Willis AC, Rammensee AVSHH. A distinctive peptide binding motif for HLA-DRB1*0407, an HLA-DR4 subtype not associated with rheumatoid arthritis. Immunogenetics. (1997) 45:229–32. doi: 10.1007/s002510050196
51. Han B, Diogo D, Eyre S, Kallberg H, Zhernakova A, Han B, et al. Fine mapping seronegative and seropositive rheumatoid arthritis to shared and distinct HLA alleles by adjusting for the effects of heterogeneity. Am J Hum Genet. (2014) 94:522–32. doi: 10.1016/j.ajhg.2014.02.013
52. Balsa A, Cabezón A, Orozco G, Cobo T, Miranda-carus E, López- MÁ, et al. Influence of HLA DRB1 alleles in the susceptibility of rheumatoid arthritis and the regulation of antibodies against citrullinated proteins and rheumatoid factor. Arthritis Res Ther. (2010) 12:1–8. doi: 10.1186/ar2975
53. Lundström E, Källberg H, Smolnikova M, Ding B, Rönnelid J, Alfredsson L, et al. Opposing effects of HLA–DRB1*13 alleles on the risk of developing anti–citrullinated protein antibody–positive and anti–citrullinated protein antibody–negative rheumatoid arthritis. Arthritis Rheum. (2009) 60:924–30. doi: 10.1002/art.24410
54. Terao C, Brynedal B, Chen Z, Jiang X, Westerlind H, Hansson M, et al. Distinct HLA associations with rheumatoid arthritis subsets defined by serological subphenotype. Am J Hum Genet. (2019) 105:616–24. doi: 10.1016/j.ajhg.2019.08.002
55. Hill JA, Southwood S, Sette A, Jevnikar AM, Bell DA, Cairns E. Cutting edge: the conversion of arginine to citrulline allows for a high-affinity peptide interaction with the rheumatoid arthritis-associated HLA-DRB1*0401 MHC class II molecule. J Immunol. (2003) 171:538–41. doi: 10.4049/jimmunol.171.2.538
56. Law S, Street S, Yu C-H, Capini C, Ramnoruth S, Nel HJ, et al. T-cell autoreactivity to citrullinated autoantigenic peptides in rheumatoid arthritis patients carrying HLA-DRB1 shared epitope alleles. Arthritis Res Ther. (2012) 14:R118. doi: 10.1186/ar3848
57. Scally SW, Petersen J, Law SC, Dudek NL, Nel HJ, Loh KL, et al. A molecular basis for the association of the HLA-DRB1 locus, citrullination, and rheumatoid arthritis. J Exp Med. (2013) 210:2569–82. doi: 10.1084/jem.20131241
58. van Venrooij WJ, Pruijn GJ. Citrullination: a small change for a protein with great consequences for rheumatoid arthritis. Arthritis Res. (2000) 2:249–51. doi: 10.1186/ar95
59. Furukawa H, Oka S, Shimada K, Hashimoto A, Tohma S. Human leukocyte antigen polymorphisms and personalized medicine for rheumatoid arthritis. J Hum Genet. (2015) 60:691–6. doi: 10.1038/jhg.2015.36
60. Klareskog L, Rönnelid J, Lundberg K, Padyukov L, Alfredsson L. Immunity to citrullinated proteins in rheumatoid arthritis. Annu Rev Immunol. (2008) 26:651–75. doi: 10.1146/annurev.immunol.26.021607.090244
61. Morgan AW, Thomson W, Martin SG, Carter AM, Erlich HA, Barton A, et al. Reevaluation of the interaction between HLA-DRB1 shared epitope alleles, PTPN22, and smoking in determining susceptibility to autoantibody-positive and autoantibody-negative rheumatoid arthritis in a large UK caucasian population. Arthritis Rheum. (2009) 60:2565–76. doi: 10.1002/art.24752
62. Mewar D, Marinou I, Coote AL, Moore DJ, Akil M, Smillie D, et al. Association between radiographic severity of rheumatoid arthritis and shared epitope alleles: Differing mechanisms of susceptibility and protection. Ann Rheum Dis. (2008) 67:980–3. doi: 10.1136/ard.2007.075382
63. Ling SF, Viatte S, Lunt M, Van Sijl AM, Silva-Fernandez L, Symmons DPM, et al. HLA–DRB1 amino acid positions 11/13, 71, and 74 are associated with inflammation level, disease activity, and the health assessment questionnaire score in patients with inflammatory polyarthritis. Arthritis Rheumatol. (2016) 68:2618–28. doi: 10.1002/art.39780
64. Tsuji H, Ikari K, Ohmura K, Yano K, Furu M, Hashimoto M, et al. Significant joint-destructive association of HLA-DRB1∗04:05 independent of DAS28 in rheumatoid arthritis. Ann Rheum Dis. (2019) 78:284–6. doi: 10.1136/annrheumdis-2018-214022
65. Terao C, Yano K, Ikari K, Furu M, Yamakawa N, Yoshida S, et al. Brief report: Main contribution of DRB1∗04:05 among the shared epitope alleles and involvement of DRB1 amino acid position 57 in association with joint destruction in anti-citrullinated protein antibody-positive rheumatoid arthritis. Arthritis Rheumatol. (2015) 67:1744–50. doi: 10.1002/art.39105
66. Criswell LA, Lum RF, Turner KN, Woehl B, Zhu Y, Wang J, et al. The influence of genetic variation in the HLA-DRB1 and LTA-TNF regions on the response to treatment of early rheumatoid arthritis with methotrexate or etanercept. Arthritis Rheum. (2004) 50:2750–6. doi: 10.1002/art.20469
67. Kong D, Lee N, Dela Cruz ID, Dames C, Maruthamuthu S, Golden T, et al. Concurrent typing of over 4000 samples by long-range PCR amplicon-based NGS and rSSO revealed the need to verify NGS typing for HLA allelic dropouts. Hum Immunol. (2021) 82:581–7. doi: 10.1016/j.humimm.2021.04.008
68. Larkins NG, D’Orsogna L, Taverniti A, Sharma A, Chakera A, Chan D, et al. The accuracy of sequence-specific oligonucleotide and real-time polymerase chain reaction HLA typing in determining the presence of pre-transplant donor-specific anti-HLA antibodies and total eplet mismatches for deceased donor kidney transplantation. Front Immunol. (2022) 13:844438. doi: 10.3389/fimmu.2022.844438
Keywords: rheumatoid arthritis, HLA-DRB1 alleles, shared epitope, anti-CCP antibodies, Chilean cohort
Citation: Catalán D, Soto L, Neira Ó, Cuéllar-Gutiérrez MC, Díaz-Peña R, Aravena O, Palou E, Carrascal M, Aguillón JC and Maggi J (2025) HLA-DRB1 allele distribution in Chilean population: insights into rheumatoid arthritis susceptibility and protection. Front. Immunol. 16:1594723. doi: 10.3389/fimmu.2025.1594723
Received: 16 March 2025; Accepted: 22 April 2025;
Published: 13 May 2025.
Edited by:
Seik-Soon Khor, Nanyang Technological University, SingaporeReviewed by:
Ramcés Falfán-Valencia, National Institute of Respiratory Diseases-Mexico (INER), MexicoWataru Hirose, Hirose Clinic of Rheumatology, Japan
Copyright © 2025 Catalán, Soto, Neira, Cuéllar-Gutiérrez, Díaz-Peña, Aravena, Palou, Carrascal, Aguillón and Maggi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jaxaira Maggi, amF4YWlyYS5tYWdnaUBpaWJiLmNzaWMuZXM=; Juan C. Aguillón, amFndWlsbG9AdWNoaWxlLmNs
†ORCID: Juan C. Aguillón, orcid.org/0000-0003-3148-6843
Jaxaira Maggi, orcid.org/0000-0002-1214-8761