 Luciele Guerra Minuzzi1,2,3*
Luciele Guerra Minuzzi1,2,3* Alexandre Abilio De Souza Teixeira4
Alexandre Abilio De Souza Teixeira4 Caique Figueiredo1
Caique Figueiredo1 Gilson Dorneles5,6
Gilson Dorneles5,6 Anna Cláudia Castelo Branco7
Anna Cláudia Castelo Branco7 Bruna Spolador de Alencar Silva1
Bruna Spolador de Alencar Silva1 Pedro L. Valenzuela8,9
Pedro L. Valenzuela8,9 Alessandra Peres6
Alessandra Peres6 Alejandro Lucia10,11
Alejandro Lucia10,11 Maria Notomi Sato7
Maria Notomi Sato7 José Cesar Rosa Neto4
José Cesar Rosa Neto4 Karsten Krüger2
Karsten Krüger2 Fabio Santos Lira1,3
Fabio Santos Lira1,3- 1Exercise and Immunometabolism Research Group, Postgraduation Program in Movement Sciences, Department of Physical Education, Universidade Estadual Paulista “Julio de Mesquita Filho”, Sao Paulo, Brazil
- 2Department of Exercise Physiology and Sports Therapy, Institute of Sport Science, Justus-Liebig-University Giessen, Giessen, Germany
- 3CIPER, Faculty of Sport Sciences and Physical Education, University of Coimbra, Coimbra, Portugal
- 4Immunometabolism Research Group, Institute of Biomedical Science, University of São Paulo, São Paulo, Brazil
- 5Escritório de Projetos, Hospital Moinhos de Vento, Porto Alegre, Brazil
- 6Laboratory of Cellular and Molecular Immunology, Federal University of Health Sciences of Porto Alegre (UFCSPA), Porto Alegre, RS, Brazil
- 7Laboratory of Medical Investigation-56, Department of Dermatology, Faculty of Medicine, University of São Paulo, São Paulo, Brazil
- 8GENUD Toledo Research Group, Centro de Investigación Biomédica en Red Fragilidad y Envejecimiento Saludable (CIBERFES), Instituto de Salud Carlos III, Madrid, Spain
- 9Grupo Mixto de Fragilidad y Envejecimiento Exitoso UCLM-SESCAM (IDISCAM), Faculty of Sport Sciences, University of Castilla-La Mancha, Toledo, Spain
- 10Physical Activity and Health Research Group (PaHerg), Research Institute of Hospital 12 de Octubre (imas12), Madrid, Spain
- 11Department of Sports Sciences, Faculty of Medicine, Health and Sports, Universidad Europea de Madrid, Madrid, Spain
Background: Lifelong exercise is associated with beneficial immune adaptations, but the extent to which these adaptations manifest during an acute inflammatory challenge remains unclear. Therefore, we aimed to compare the inflammatory responses to ex vivo whole blood and peripheral blood mononuclear cells [PBMCs] cultures from young and master athletes, before and after a single bout of moderate-intensity exercise.
Methods: Young (n=7; 22 ± 4 years) and master (n=12; 52 ± 9 years) female and male athletes with similar performance levels performed a 30-minute bout of moderate-intensity exercise. Blood samples were collected before and post-exercise to assess cytokine production in whole blood and PBMCs after stimulation with lipopolysaccharide [LPS] and a cocktail with phorbol 12-myristate 13-acetate [PMA] plus ionomycin.
Results: In whole blood, LPS induced higher interleukin [IL]-6 release in both groups, with a greater increase in young athletes at post-exercise (p=0.014). Tumor necrosis factor [TNF]-α levels increased only in young athletes (p<0.0001). In PBMCs, master athletes showed lower LPS-induced TNF-α release, increasing only post-exercise (p<0.034), whereas young athletes responded at both baseline (p<0.001) and post-exercise (p=0.003). Under PMA/ionomycin stimulation, TNF-α (p<0.0001) and interferon (IFN)-γ (p=0.007) release increased only in young athletes, while IL-6 production decreased in young athletes at baseline (p=0.002) and post-exercise (p=0.003).
Conclusion: Young athletes exhibit a stronger cytokine response to ex vivo inflammatory stimuli, while master athletes demonstrate a more controlled and regulated inflammatory profile.
1 Introduction
The long-term benefits of regular exercise, particularly concerning immune function and inflammation, are mediated—at least partly—by the cumulative effects of repeated bouts of ‘acute’ exercise (1). Of note, although acute unaccustomed exercise can be considered a pro-inflammatory stimulus, it stimulates the release of circulating components—the so-called exerkines—that can exert an anti-inflammatory function (2). A well-known example is the exercise-induced increase in IL-6, which induces the release of other circulating cytokines that have anti-inflammatory properties, such as IL-10 (3), while decreasing the circulating levels of the pro-inflammatory cytokine TNF-α (4), thereby ultimately contributing to re-establishing inflammatory homeostasis (5).
Lifelong athletes with consistent and regular years (≥20 years) of exercise practice (also known as ‘master athletes’) represent the paradigm of youthful immunomodulation (as a result of repeated acute exercise bouts in the long term) as they show, among others, similar (e.g., IL-10) if not higher plasma levels of anti-inflammatory cytokines than much younger adults, together with reduced senescent T cell numbers (6–8). Regular exercise may also improve macrophage function by preventing age-related dysregulation in Toll-like receptors (TLRs, which recognize pathogens and activate immune responses to defend against infections) (9). This implies that regular exercise might increase the threshold for immune activation (i.e., the level of stress or stimulus required to activate the innate immune system) (10), potentially making lifelong exercisers more resilient to stress and less prone to excessive or inappropriate immune responses.
However, there is currently no evidence as to whether lifelong exercise also affects the response to a pro-inflammatory stress stimulus, such as exposure to lipopolysaccharide [LPS], a potent endotoxin and widely recognized activator of the monocyte/macrophage arm of the innate immune system—to stimulate cells ex vivo (11, 12). LPS is a component of the outer membrane of Gram-negative bacteria that activates an innate immune response by interacting with Toll-like receptor 4 (TLR4). This interaction initiates a signaling cascade via nuclear factor kappa B (NF-kB), leading to the production of pro-inflammatory cytokines like TNF-α, IL-18, and macrophage inflammatory protein-1α [MIP-1α] required for a robust immune response against bacterial infections (13–15). The normal response to an LPS challenge is characterized by a transient, robust upregulation of inflammatory activity, that resolves once the threat has passed (16). This temporal restriction of inflammation (i.e., activating and resolving the inflammatory response in a controlled manner) is crucial for maintaining overall health, as prolonged or dysregulated inflammatory responses can lead to a host of chronic diseases and health conditions (17). In addition to LPS stimulation, phorbol ester was used in combination with ionomycin, a calcium ionophore, to broadly activate lymphocytes (18). Phorbol 12-myristate 13-acetate (PMA)/ionomycin through protein kinase C (PKC) activates mononuclear cells, including T cells and B cells, and, to some extent, NK cells by mimicking intracellular signaling events that lead to cytokine production and other activation markers (e.g., IFN-γ, CD69).
The primary aim of this study was, therefore, to determine whether master athletes exhibit an inflammatory response comparable to that of younger trained counterparts, with a particular focus on cytokine production. To address this, we compared the cytokine production to pro-inflammatory triggers by stimulating whole blood and PBMC cultures at baseline and after an acute exercise session. By integrating gene expression profiling and detailed flow cytometric phenotyping, we further explored whether differences in cytokine production could be attributable to transcriptional regulation, alterations in immune cell frequency, phenotype, or both. We hypothesized that while younger athletes would demonstrate a robust cytokine release in response to inflammatory stimuli, master athletes would exhibit a comparatively tightly regulated (i.e., controlled yet sufficient) inflammatory response, possibly reflecting adaptive immunomodulatory mechanisms accumulated over long-term regular training.
2 Methods
The study was conducted in accordance with the ethical standards of the Declaration of Helsinki. All participants were explained about the purpose and procedures of the study and gave their written informed consent. The Ethical Institutional Review Board approved the study (approval number: 38701820.0.0000.5402).
An overview of the design study is illustrated in Figure 1.
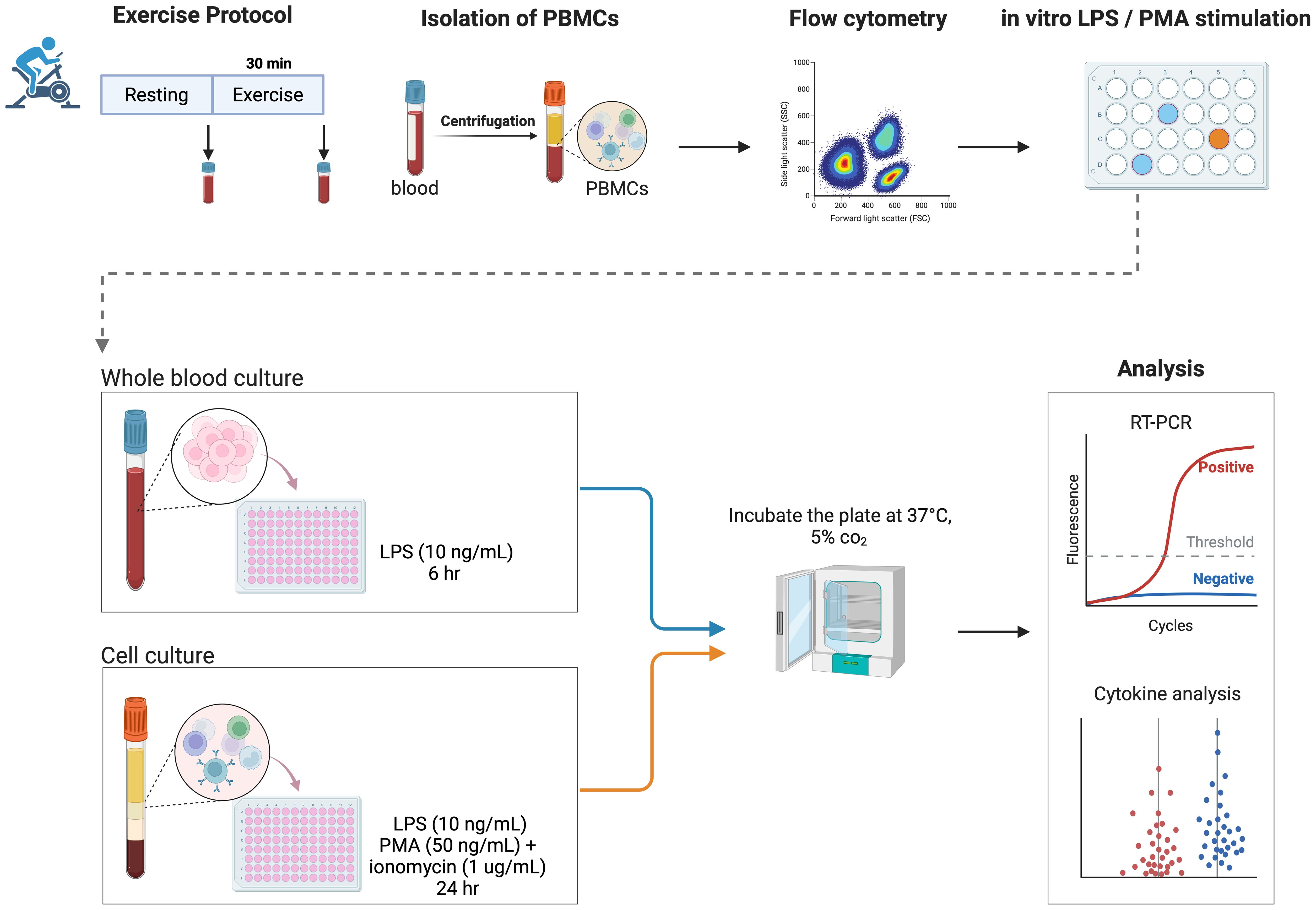
Figure 1. Study design. Overview of the study design, including an acute bout of moderate exercise (30 minutes on the cycle ergometer), blood sampling, isolation of PBMCs, and an in vitro inflammatory response protocol in both whole blood and PBMCs. LPS, lipopolysaccharide; PBMCs, peripheral blood mononuclear cells; PMA, phorbol 12-myristate 13-acetate. Created in BioRender. Guerra minuzzi, L. (2025) https://BioRender.com/z58z271.
2.1 Participants
Master athletes (n = 12; 9 men, 3 women; 51.8 ± 9.0 years) included eight cyclists, two track-and-field athletes, and two swimmers with 28.1 ± 13.8 years of continuous training (8.8 ± 4.4 h·wk-¹, 11.9 ± 0.3 months·yr-¹), whereas young athletes (n = 7; 5 men, 2 women; 22.0 ± 4.1 years) comprised five runners, one cyclist, and one team-sport participant with 8.7 ± 4.1 years of continuous training (9.8 ± 4.9 h·wk-¹, 10.8 ± 1.0 months·yr-¹). Exclusion criteria included smokers, excessive consumption of alcohol (>2 drinks/day), and any known cardiovascular, musculoskeletal, or neurological condition or regular medication use. The sports experience history of master and young athletes was assessed by self-reporting their athletic life, sports modalities practiced, and the duration dedicated to each category, including the start and end of the year, hours/week, months/year, injuries episodes, and participation in competitions and performances (Supplementary File 1). A second questionnaire focused on the development of the other activities, namely type, category, hours/week, and classification of effort intensity, was applied.
Nutritional information was measured at the initial screening using a three-day food diary that consisted of two weekdays and one weekend day to reflect typical intake. Total and relative energy, protein, carbohydrates and lipids intake, and other nutrients (vitamins, minerals, and different types of lipids) were analyzed using NutWinsoftware, version 1.5 (Nutrition Support Program, Federal University of São Paulo, Brazil, 2002) (Supplementary File 2).
2.2 Exercise protocol
Participants underwent a maximal incremental exercise test on a cycle ergometer (Inbrasport CG-04, Embramed: Porto Alegre, Brazil) linked to an automated breath-by-breath gas analyzer (Quark PFT, Cosmed®; Rome, Italy) for peak oxygen uptake (VO2peak) and ventilatory threshold (VT) determination. The protocol started with an initial load of 75 W and was increased by 25 W every 3 minutes until volitional exhaustion or until cadence fell below 60 rpm. The VO2 peak was determined as the mean VO2 consumption in the last 30 seconds of the test. The VT was determined based on the relationship between ventilation (VE) and workload during each stage of the maximal exercise test. Specifically, VE/workload was plotted against workload, and the point at which VE/workload began to rise disproportionately (i.e., a distinct break in the slope) was identified as the VT, in accordance with the method described previously (19). Participants were asked to refrain from vigorous exercise, alcohol consumption, and caffeine for 24 hours before the test. Participants returned to the laboratory at least 24 hours after the aforementioned maximal incremental test to perform a 30-minute exercise bout on the cycle ergometer at a power output slightly above (by 5-15%) the one eliciting the VT in the previous maximal test. The chosen exercise trial is recognized for a selective redeployment of immune cells, notably those subsets with a high differentiation phenotype (20).
2.3 Blood measures
All participants arrived at the laboratory following an overnight fast. Upon arrival, baseline venous blood samples were obtained from the forearm vein using Vacutainer tubes (Becton Dickinson, Juiz de Fora, Brazil) containing ethylenediaminetetraacetic acid (EDTA) for subsequent whole blood sampling and isolation of PBMCs, as well as using dry Vacutainer tubes (without anticoagulant agent) for serum separation. Participants then consumed a standardized breakfast that they had brought with them, thereby replicating their typical pre-training meal, and undergoing body composition assessments. Following this period (approximately 1 hour), they completed the acute exercise session. A second blood sample was collected immediately (≤5 minutes for maximal leukocyte mobilization (21)) post-exercise in accordance with the same procedure. Blood samples were centrifuged at 3,000 rpm for 15 minutes at 4°C and the serum supernatant was stored at −80°C until further analysis. Serum levels of IFN-α (i.e., a marker of antiviral response), IL-10, IL-18, MIP-1α, and cytomegalovirus (CMV) were determined by enzyme-linked immunosorbent assay (ELISA) (Duoset R&D System, Minneapolis, MN). All athletes were tested for CMV since studies have shown that CMV influences the immune response (22–24).
2.4 Inflammatory response
2.4.1 Whole blood stimulated with LPS
We employed a protocol similar to that described by Barry et al. (25) for the whole blood-stimulated ex vivo assay. Fresh EDTA blood samples were diluted 10-fold with serum-free RPMI media (Sigma Aldrich, MA) supplemented with penicillin (100 U/mL) and streptomycin (0.1 mg/mL), and seeded into a 24-well plate, either in the absence (i.e., ‘non-stimulated’ condition) or adding 10 ng/mL of LPS (Escherichia coli, type: 0111: B4; Sigma-Aldrich, St. Louis, MO) (i.e., ‘LPS-stimulated’ condition). Following incubation for 6 hours, the culture supernatants for both non-stimulated and LPS-stimulated conditions were collected and stored at −80°C until further analysis of the inducible IL-6 and TNF-α levels by ELISA (Duoset R&D System, Minneapolis, MN).
2.4.2 Peripheral blood mononuclear cell culture
The EDTA-coated blood samples were added to Histopaque®-1077 (Sigma–Aldrich Co. LLC) (1:1) for PBMC isolation and centrifuged at 400×g for 30 min at room temperature. After centrifugation, the PBMCs were washed in phosphate-buffered saline and re-suspended in a supplemented RPMI medium [glutamine (2mM), HEPES (20mM), 10% fetal bovine serum and antibiotics penicillin (100 U/mL) and streptomycin (0.1 mg/mL)] in 12-well plates (Kasvi; PR, Brazil). A total of 1x106 cells were plated either without (non-stimulated) and with (LPS–stimulated) 10 ng/mL of LPS, or with PMA [50 ng/mL] (Sigma, St. Louis, MO) plus ionomycin [1 μg/mL] (Sigma, St. Louis, MO) in a final volume of 1 mL of supplemented RPMI medium. After 24-hour incubation, supernatants of non-stimulated, LPS-stimulated, and ‘PMA/ionomycin-stimulated’ conditions, respectively, were collected and stored at −80°C for subsequent analysis. LPS-stimulated samples were analyzed for IL-6, IL-10, and TNF-α levels, and PMA/ionomycin-stimulated samples were analyzed for IL-6, IFN-γ, and TNF-α levels via ELISA kits (Duoset R&D System, Minneapolis, MN).
2.5 Real-time-polymerase chain reaction
Polymerase chain reaction (PCR) was used to amplify and quantify specific genes relevant to inflammation and immune regulation to determine whether exercise status influenced their transcriptional responses following stimulation. Thus, total RNA was extracted from PBMCs using the Brazol reagent (LGC Biotechnology Ltda.; Cotia, Brazil) following the manufacturer’s recommendations and used for real-time polymerase chain reaction (RT-PCR) analyses. The High-Capacity cDNA Reverse Transcription kit (Applied Biosystems, Thermo Fisher Scientific; Foster, CA) was used to perform the reverse transcription to complementary DNA (cDNA), which was stored at -80°C for subsequent analysis of genes by RT-PCR with Power SYBR Green PCR Master Mix (Applied Biosystems). Primer sequences are presented in Supplementary File 3.
The gene expression was quantified using β-actin as an internal control. We performed relative quantification (using the 2-ΔΔCT formula) of the expression levels of ADRB1 (adrenergic receptor β1), CD62L (L-selectin), CCR7 (C-C chemokine receptor type 7), FAS (Fas cell surface death receptor), ICOS (inducible T-cell co-stimulator), PDCD1 (programmed cell death protein 1) and TNF-α genes.
2.6 Flow cytometry
Flow cytometry allowed us to identify and quantify specific immune cell subsets (such as monocyte and NK cell populations) and to assess the expression of surface markers (e.g., CD69), thus providing insight into how the composition or activation state of immune cells may differ between age groups and across exercise status. PBMCs were isolated using the above-mentioned protocol and then frozen at −80°C in a solution containing 90% fetal bovine serum and 10% dimethyl sulfoxide (DMSO) until analysis. Cells were thawed by diluting them in pre-warmed RPMI 1640 medium containing 5% FBS and spun at 1500 rpm for 5 min. After centrifugation, the cells were re-suspended, and their viability (>98%) was confirmed using trypan-blue staining (Gibco, Grand Island; New York, NJ).
Subsequently, 2 × 105 PBMCs were stained with monoclonal antibodies conjugated to specific fluorochromes for cell phenotyping: anti-CD14+ FITC, anti-CD16+ Pe, anti-CD69+ Percp-Cy5.5, anti-CD3+ FITC, anti-CD56+ Pe, anti-CD16+ Percp-Cy5.5, anti-CD279+ APC, anti-CXCR3 APC, anti-KLRG1 AlexaFluor 633 (BioLegend, San Diego, CA). Flow cytometry analysis was performed using CellQuest Pro Software (BD Bioscience; Franklin Lakes, NJ) on a FACSCalibur flow cytometer (BD Bioscience), with a minimum of 20,000 events per tube acquired. Lymphocytes were identified based on forward and side scatter profiles. The NK-cells were defined as CD3−CD56+, and the two major subsets of NK-cells were identified as CD56+CD16− and CD56+CD16+ (CD56bright and CD56dim, respectively). The frequencies of activated and terminally differentiated NK cell subsets were defined by early activation markers CD69 and CD57, respectively. NK cells subsets were also identified and characterized according to exhaustion [programmed cell death protein 1 (PD1)], senescence [killer-cell lectin-like receptor G1 (KLRG1)], and migratory [CXC chemokine receptor 3 (CXCR3)] markers. Classical (CD14+CD16−) and non-classical (CD14+CD16+) monocyte subsets were identified, and the frequency of CD69+ cells was determined in each subset.
2.7 Statistical analysis
Data are shown as mean and standard deviation (SD) unless otherwise stated. Baseline comparisons between groups were assessed using the Mann–Whitney U test, given the small sample size and potential non-normality of the data. For the main outcomes, between-group and within-group effects were determined using a mixed-design ANOVA with ‘group’ (young, older) as the between-subject factor and ‘condition’ (baseline, exercise) as the within-subject factor. An additional within-subject factor, ‘stimulus’ [i.e., stimulation (or not) with LPS or stimulation (or not) with PMA plus ionomycin], was included to analyze the differences in response to inflammatory stimuli. To verify the assumptions for ANOVA, we applied Levene’s test to assess homogeneity of variance and Mauchly’s test for sphericity. When sphericity was violated, the Greenhouse–Geisser correction was applied. The Bonferroni post-hoc test was only performed within groups when a significant interaction effect was found. Analyses were performed using SPSS (v29, IBM, Armonk, NY), with the significance level set as p ≤ 0.05.
3 Results
Master athletes reported 2.4 ± 1.7 competitive events in the past two years, 83.3% engaged in at least one additional sport (4.6 ± 1.6 h·wk-¹ of alternative activities), and 50% had experienced an injury in the last two years. Young athletes completed 3.2 ± 1.7 events, 85.7% participated in another sport (7.6 ± 2.6 h·wk-¹), and 66.7% reported a recent injury (Supplementary File 1). Young and master athletes showed similar values of body mass index (24.1 ± 3.4 and 25.4 ± 2.1 kg/m2, respectively, p=0.303), VO2peak (53.0 ± 13.2 and 52.2 ± 10.3 ml/kg/min, p=0.442) and power output at the VT (139 ± 24 and 135 ± 37 watts, p=0.786; Supplementary File 4). Regarding dietary intake, young athletes consumed more protein than master athletes (p=0.019). No differences were observed in the consumption of carbohydrates, lipids, cholesterol, fiber, or other vitamins and minerals (Supplementary File 3).
3.1 Serum cytokine profile
All participants were CMV seropositive. Figure 2 presents the serum levels of IFN-α (A), IL-10 (B), IL-18 (C) and MIP-1α (D) measured at baseline and after exercise. A significant group (<0.001), condition (p<0.007), and group*condition interaction (p=0.045) effect was found for MIP-1α (Figure 2). Specifically, master athletes had significantly lower MIP-1α levels than young athletes both at baseline (p<0.001) and after exercise (p<0.001). Additionally, in the younger group, MIP-1α levels increased significantly from baseline to post-exercise (p=0.004), whereas no change was observed in the master athletes. In contrast, no significant group, condition, or group*condition interaction effect was observed for IFN-α (p=0.243, p=0.198 and p=0.122, respectively), IL-18 (p=0.148, p=0.150 and p=0.316) or IL-10 (p=0.309, p=0.293 and p=0.823) (Figure 2).
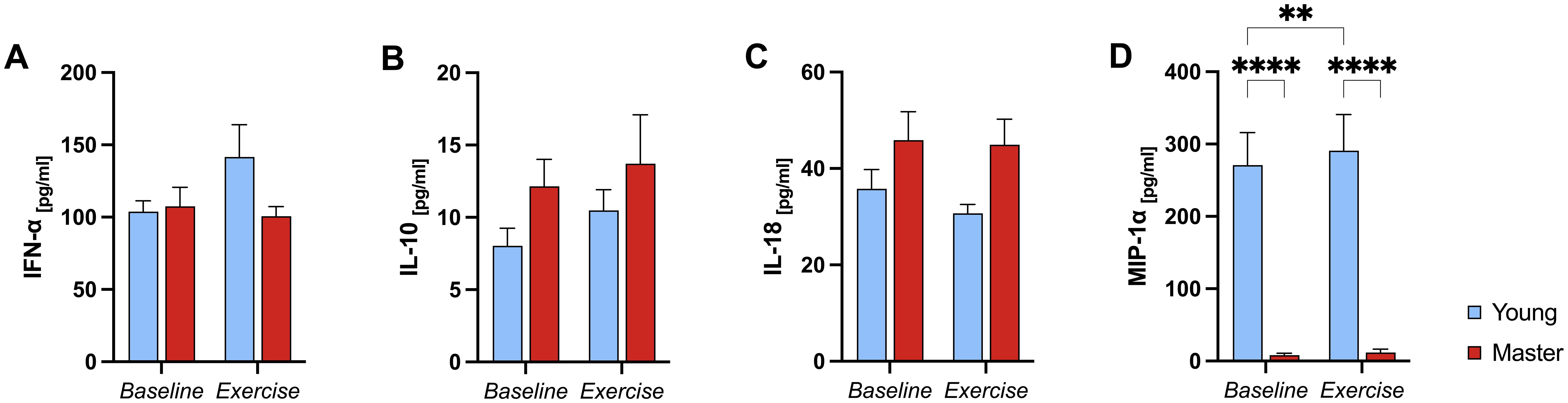
Figure 2. Serum cytokine levels in young and master athletes at baseline and post-exercise. Serum IFN-α (A), IL-10 (B), IL-18 (C), and MIP-1α (D) concentrations were measured before (baseline) and immediately after a 30-minute moderate-intensity cycling session (exercise) in young (n = 7) and master (n = 12) athletes. Data are mean ± SEM. (A–C): No significant main or interaction effects. (D): **** p<0.0001, significant Bonferroni post-hoc analysis, two-way ANOVA (group*condition) interaction effect. IFN-α, Interferon-alpha; IL, Interleukin; MIP-1α, Macrophage inflammatory protein-1 alpha.
In summary, MIP-1α was the only cytokine to show significant differences across groups and exercise conditions—rising in the younger group but remaining consistently lower in master athletes—while IFN-α, IL-18, and IL-10 levels were unchanged regardless of age or exercise.
3.2 Frequency of NK cells, classical and non-classical monocytes, and subsets
A significant condition effect was observed for NK cells (p<0.001), but no group (p=0.772) or group*condition interaction effect (p=0.291), indicating that both groups exhibited a similar change from baseline to post-exercise. Similarly, a significant condition effect was observed for PD-1 expression in both NK (p=0.007) and CD56+CD16+ cells (p=0.020), but no group (p=0.682, p=0.665) or group*condition interaction effect (p=0.731, p=0.258). In contrast, there was no significant group*condition interaction, group or condition effect for CD56+CD16− (p=0.393, p=0.558, and p=0.478, respectively) and PD1+CD56+CD16− (p=0.525, p=0.858, p=0.064) cells. No significant group*condition interaction effects were observed for the expression of CD69, CXCR3, and KLRG1 in NK cells or NK cell subsets (p>0.05) (Supplementary File 5). Additionally, there were no significant group, condition, or group*condition interaction effects for the frequency of classical [CD14+CD16−] (p=0.286, p=0.937, and p=0.255, respectively) or non-classical [CD14+CD16+] (p=0.253, p=0.937, and p=0.284), monocytes. Likewise, CD69 expression in both classical (p=0.274, p=0.141, and p=0.705) and non-classical (p=0.295, p=0.103, and p=0.762) monocytes did not change with exercise and did not differ between groups (Supplementary File 5).
In summary, while total NK cell frequency and PD-1 expression on NK cells and CD56dim subsets showed a significant shift from baseline to post-exercise (indicating a ‘condition’ effect), there were no significant differences between the groups or interactions between group and condition for any cell frequencies or activation markers. Classical and non-classical monocytes, as well as activation markers, remained unchanged across time and between groups.
3.3 Whole blood stimulated with LPS
A significant group*condition*stimulus (p=0.009), group*condition (p=0.003), and group*stimulus (p<0.0001) interaction effect was observed for TNF-α release in whole blood upon LPS stimulation. Specifically, the younger group showed a robust increase in TNF-α levels in response to LPS at both baseline (p<0.0001) and post-exercise (p<0.0001). In contrast, no significant change was observed in master athletes (p>0.999 for both baseline and exercise). Moreover, master athletes consistently exhibited lower LPS-stimulated TNF-α levels than the younger group a both baseline and exercise conditions (p<0.0001) (Figure 3).
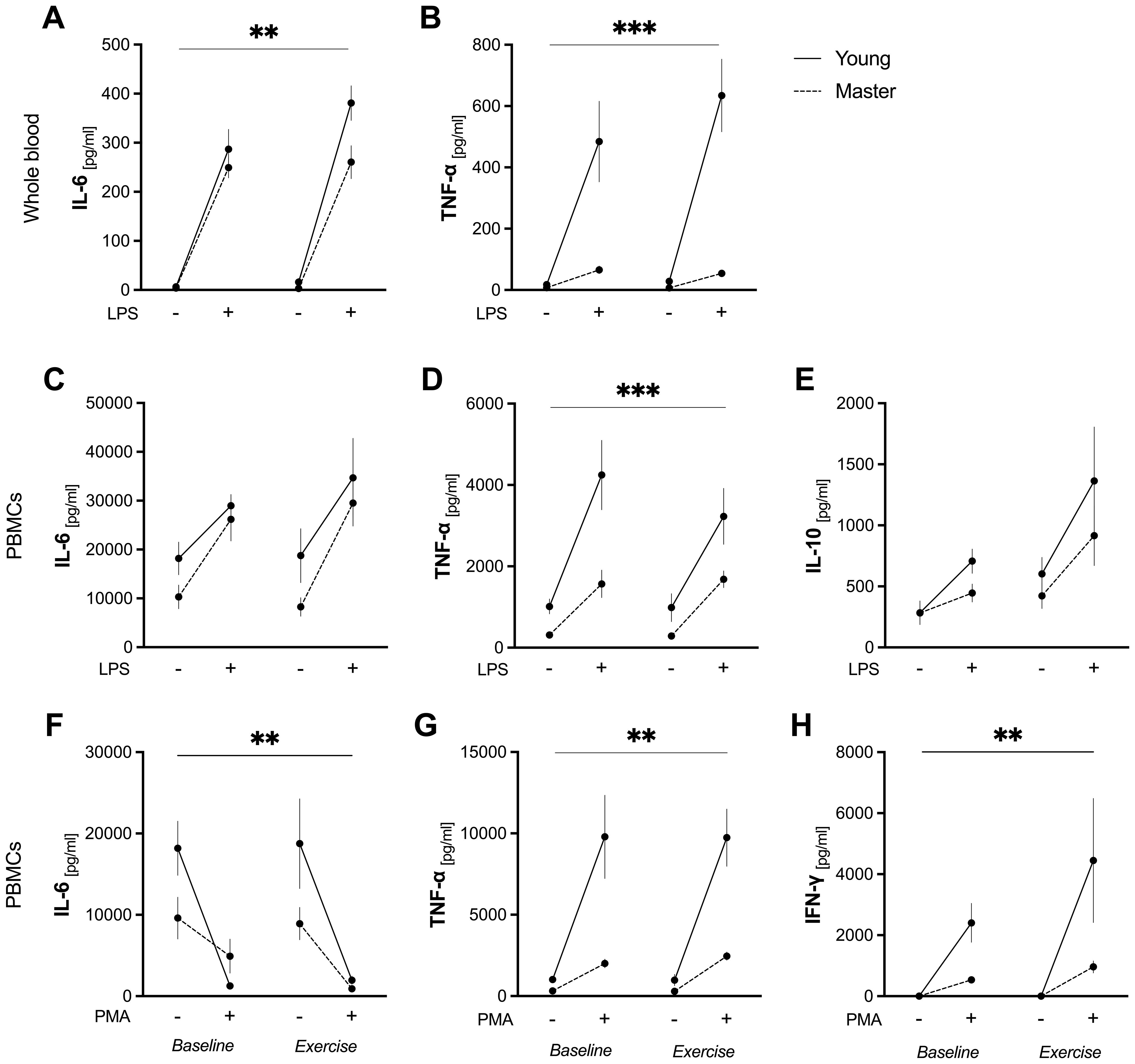
Figure 3. Ex vivo cytokine production in whole blood and PBMCs before and after acute exercise. Whole blood was incubated for 6 h with (+) or without LPS (10 ng/mL) (A–B). PBMCs were stimulated for 24 h with LPS (10 ng/mL) (C–E) or PMA (50 ng/mL) + ionomycin (1 μg/mL) (F–H). Data are mean ± SEM for non-stimulated (–) and stimulated (+) conditions in young (n = 7) and master (n = 12) athletes at baseline and immediately after a 30-min moderate cycling bout (“exercise”). (A): LPS-induced IL-6 release in whole blood. ** p<0.05, significant two-way ANOVA (group*condition) interaction effect. (B, D): LPS-induced TNF-α in whole blood (B) and in PBMCs (D). *** p<0.05, significant three-way ANOVA (group*condition*stimulus) interaction effect. (C, E): LPS-stimulated IL-6 (C) and IL-10 (E) release by PBMCs. No significant group, condition, or interaction effects were observed. (F–H): PMA/ionomycin-induced IL-6 (F), TNF-α (G), and IFN-γ (H) release by PBMCs. ** p<0.05, significant two-way ANOVA (group*stimulus) interaction effect. IFN-γ, interferon-γ; IL, interleukin; LPS, lipopolysaccharide; PBMCs, peripheral blood mononuclear cells; PMA, phorbol 12-myristate 13-acetate; SEM, standard error of the mean; TNF-α, tumor necrosis factor-α.
No significant group*condition*stimulus interaction effect was observed for IL-6 (p=0.519). However, a significant group*stimulus interaction (p=0.039) effect indicated differential responses between groups when stimulated with LPS. LPS induced a significant increase in IL-6 in both groups at baseline and post-exercise (p<0.0001 for all comparisons). Notably, the younger group showed higher LPS-stimulated IL-6 levels than master athletes in post-exercise (p=0.014) (Figure 3).
In summary, young adults exhibited a marked increase in LPS-induced TNF-α at both baseline and exercise, while master athletes showed no significant change and maintained consistently lower TNF-α values. Although LPS similarly increased IL-6 in both groups, the younger athletes had higher IL-6 levels post-exercise.
3.4 PBMC culture
3.4.1 LPS-stimulated cytokine release
A significant group*stimulus (p<0.034), group (p=0.0001), and stimulus (p<0.0001) effect was observed for TNF-α release by PBMCs upon LPS stimulation, although there was no group*condition*stimulus interaction (p=0.144). In the younger group, LPS significantly increased TNF-α release at baseline (p<0.001) and post-exercise (p=0.003). In contrast, master athletes showed an increase only in the exercise condition (p=0.043). Moreover, LPS-stimulated TNF-α levels were significantly lower in master athletes than in the younger group at baseline (p<0.001), with a trend toward significance post-exercise (p=0.051) (Figure 3). These findings suggest that although LPS stimulates TNF release in both groups, young athletes have a more pronounced response, particularly at baseline.
No significant group*condition*stimulus interaction effect was found for LPS-stimulated IL-6 release by PBMCs (p=0.976), and no group*stimulus, group*condition, or condition*stimulus interaction effects were observed (p=0.484, p=0.612, p=0.288, respectively). However, LPS significantly increased IL-6 release in both groups compared to unstimulated conditions (stimulus effect, p=0.0001), and overall IL-6 levels were similar in young compared to the master athletes (group effect, p=0.083). Meanwhile, there were no notable differences when comparing baseline to exercise conditions (condition effect, p=0.444). These data suggest that, while LPS robustly induced IL-6 production, the magnitude of this response differed between the two groups yet remained unaffected by acute exercise. Similarly, there was no significant group*condition*stimulus interaction effect for IL-10 release (p=0.984), and no group*stimulus, group*condition, or condition*stimulus interaction effect (p=0.366, p=0.436, p=0.158) were found. Nevertheless, LPS stimulation significantly increased IL-10 production in both groups (stimulus effect, p=0.013), and IL-10 levels also differed between baseline and post-exercise across all participants (condition effect, p=0.002). However, the pattern of IL-10 release did not differ by group (p=0.231), indicating similar responses to both LPS and exercise (Figure 3).
3.4.2 PMA/ionomycin-stimulated cytokine release
No significant group*condition*stimulus interaction effect was found for TNF-α (p=0.724). However, a significant group*stimulus interaction (p<0.0001), group (p<0.0001), and stimulus (p<0.0001) effect were observed. PMA/ionomycin markedly increased TNF-a release in PBMCs from the younger group (p<0.0001), resulting in higher levels than those of master athletes at both baseline and post-exercise (p<0.0001). A similar pattern emerged for IL-6, with no group*condition*stimulus interaction (p=0.506), but a significant group*stimulus (p=0.020) and stimulus (p<0.0001) effect. There was no group (p=0.075) effect. In the younger group, IL-6 production decreased with PMA/ionomycin at both baseline (p=0.0008) and post-exercise (p=0.0009). Conversely, master athletes showed no significant change in IL-6 upon PMA/ionomycin stimulation (p>0.999 for baseline; p=0.134 for exercise conditions; Figure 3).
PMA/ionomycin-stimulated IFN-γ release showed no significant group*condition*stimulus interaction effects (p=0.212). However, significant group*stimulus (p=0.007), stimulus (p<0.001), and group (p=0.007) effects were observed. PMA/ionomycin significantly increased IFN-γ release only in young athletes (p<0.001), resulting in higher levels compared to master athletes in post-exercise (p=0.001). No group*condition (p=0.221) or condition*stimulus (p=0.067) interaction effects were found (Figure 3).
In summary, while LPS stimulation induced increased TNF-α, IL-6, and IL-10 production by PBMCs in both groups to varying degrees, the younger group generally demonstrated a more robust cytokine response. Under PMA/ionomycin stimulation, the younger group again produced higher TNF-α and IFN-γ than master athletes.
3.5 Gene expression
A significant group*condition*stimulus interaction effect (p=0.038) was observed for CD62L gene expression in PBMCs following LPS stimulation. Specifically, the young group exhibited higher LPS-stimulated CD62L mRNA levels at baseline compared to master athletes (p=0.015) and their own post-exercise levels (p=0.049). By contrast, master athletes did not show a significant change in CD62L expression from baseline to post-exercise.
No significant group*condition*stimulus interaction was found for PMA/ionomycin-stimulated CD62L expression (p=0.411). However, there were significant group*stimulus (p=0.035), group (p=0.013) and stimulus (p=0.019) effects. In the younger group, CD62L gene expression increased at baseline following PMA/ionomycin stimulation (p=0.013), resulting in higher values than those of master athletes (p=0.010). No significant group*condition (p=0.487), condition*stimulus (p=0.258), or condition (p=0.540) effects were observed, suggesting that acute exercise did not influence PMA/ionomycin-induced CD62L expression in PBMCs (Figure 4).
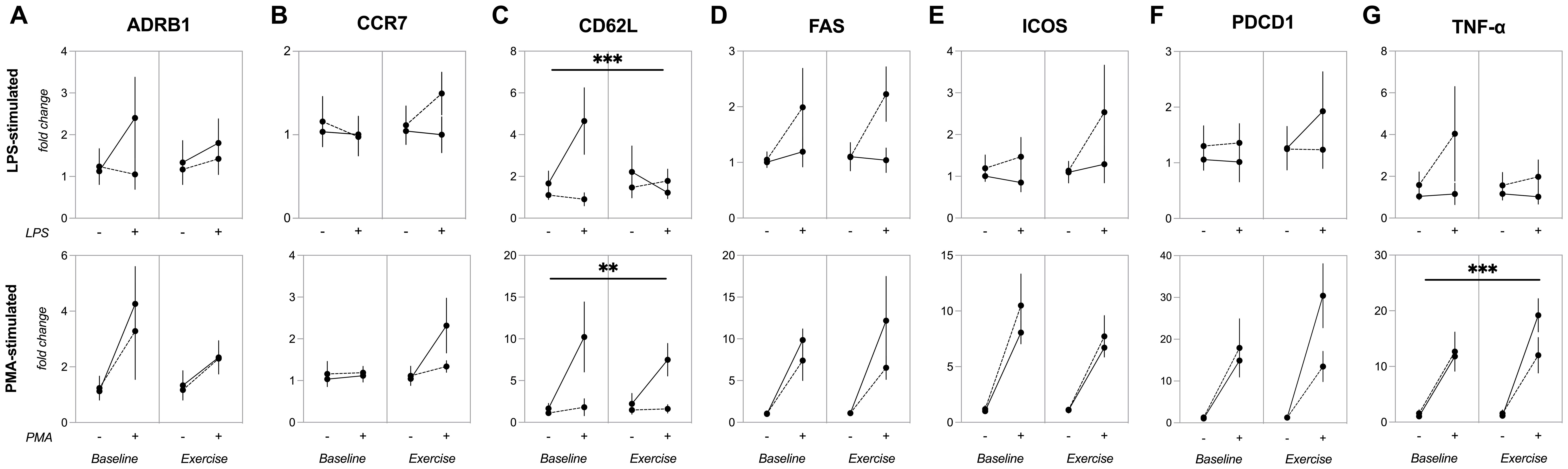
Figure 4. Gene expression of inflammatory and regulatory markers in PBMCs before and after acute exercise. Expression of (A) ADRB1, (B) CCR7, (C) CD62L, (D) FAS, (E) ICOS, (F) PDCD1, (G) TNF-α genes in PBMCs stimulated by 24-hour incubation with LPS (upper panels) or PMA plus ionomycin (lower panels). Data are mean ± SEM for non-stimulated (–) or stimulated (+) conditions in young (n=7) and master (n=12) athletes before (baseline) and immediately after a 30-minute moderate cycling session (exercise). (A, B, D–F): No significant main or interaction effects. (C) (CD62L): LPS-stimulated CD62L gene expression in PBMCs (upper). *** p<0.005, significant three-way ANOVA (group*condition*stimulus) interaction effect. PMA-stimulated CD62L gene expression in PBMCs (lower). ** p<0.05, significant two-way ANOVA (group*condition) interaction effect. (G) (TNF-α): PMA-stimulated TNF-α gene expression in PBMCs. *** p< 0.005, significant three-way ANOVA (group*condition*stimulus) interaction effect. ADRβ1, adrenergic receptor beta-1; CCR7, C-C chemokine receptor type 7; CD62L, CD62L (L-selectin); FAS, cell surface death receptor; ICOS, inducible T-cell costimulatory; LPS, lipopolysaccharide; PBMCs, peripheral blood mononuclear cells; PDCD1, programmed cell death protein 1; PMA, phorbol 12-myristate 13-acetate; SEM, standard error of the mean; TNF-α, tumor necrosis factor-alpha.
A significant group*condition*stimulus interaction (p=0.035) effect was found for TNF-α gene expression in PBMCs stimulated with PMA/ionomycin, along with a group*condition interaction (p=0.030) and stimulus (p<0.0001) effect. Under PMA/ionomycin stimulation, TNF-α gene expression increased for both young and master athletes at baseline (p=0.018 and p=0.053, respectively) and exercise (p<0.0001 and p=0.010). Furthermore, in the younger group, PMA-induced TNF-α gene expression was higher in exercise compared to baseline (p=0.015). By contrast, no group*condition*stimulus interaction (p=0.512) was observed for TNF-α gene expression when PBMCs were stimulated with LPS (Figure 4).
A significant stimulus effect was observed for ADRB1 (p=0.018), FAS (p<0.001), ICOS (p<0.0001), and PDCD1 (p<0.0001) gene expression in PBMCs stimulated with PMA/ionomycin, but not for CCR7 (p=0.074). Under LPS stimulation, only FAS showed a significant stimulus effect (p=0.043), while CCR7 (p=0.885), ADRB1 (0.269), or PDCD1 (p=0.643) were unaffected. Moreover, there were no significant group*condition*stimulus effects for ADRB1 (LPS: p=0.303, PMA/ionomycin: p=0.603), CCR7 (LPS: p=0.186, PMA/ionomycin: p=0.190), FAS (LPS: p=0.0.691, PMA/ionomycin: p=0.507), ICOS (LPS: p=0.609, PMA/ionomycin: p=0.680), or PDCD1 (LPS: p=0.348, PMA/ionomycin: p=0.0.068) (Figure 4).
In summary, these results indicate that while certain genes (ADRB1, FAS, ICOS, and PDCD1) are responsive to PMA/ionomycin stimulation, and FAS is also responsive to LPS, neither acute exercise nor group differences appear to modulate these effects.
4 Discussion
Our findings indicate a distinct inflammatory profile between young and master athletes following acute exercise when evaluated using ex vivo stimulation models, including activation of TLR4 (via LPS) and PKC (via PMA/ionomycin). Overall, young athletes showed a stronger stimulated cytokine response, with higher production of IL-6 and TNF-α in both whole blood and PBMCs cultures. In contrast, master athletes showed lower baseline TNF-α levels across both models but demonstrated an increase in TNF-α production by PBMCs following exercise when stimulated with LPS—an effect not observed at rest—suggesting that the exercise bout modulated their inflammatory responsiveness. These results indicate that master athletes exhibit a more controlled inflammatory response, in contrast to the stronger stimulated cytokine production observed in younger athletes.
Yet, the lower production of TNF-α [a powerful pro-inflammatory agent that is rapidly released after stressors (e.g., LPS) and is one of the main early mediators in inflamed tissues] observed in master athletes could be attributed to a “hypo-responsiveness” state of PBMCs to pro-inflammatory stimuli, such as the so-called ‘endotoxin tolerance’ (15, 26, 27). Endotoxin tolerance, typically characterized by reduced production of pro-inflammatory cytokines upon repeated LPS encounters, can protect against excessive inflammation but may also increase the susceptibility to secondary infections due to immunosuppression. Aging is known to impact immune function, including alterations in cell-type-specific signaling pathways and expression of receptors (14, 28, 29), thereby impairing endotoxin tolerance (15). Although older adults can exhibit insufficient or exaggerated inflammatory responses in different contexts, our master athletes—who maintain decades of exercise training—seem to exhibit a more regulated inflammatory response rather than an outright deficiency, as suggested by their capacity to upregulate TNF-α post-exercise.
While specific studies on the effects of physical exercise on endotoxin tolerance in aging are limited, these findings align with the literature indicating that regular exercise training can modulate TLR expression and dampen excessive TNF-α release following LPS stimulation (30–32). A 3-week running intervention of moderate intensity prevented excessive immune system activation under LPS-induced inflammatory conditions in mice (32). The training status also exhibits effects on inflammatory response. Antunes et al. found that the anti-inflammatory response is linked with higher physical fitness and may be explained by an efficient cellular framework for the PPAR-γ responses in monocytes (33). In this context, Olesen et al. observed that exposure to LPS-induced inflammation in humans led to a significant suppression of LPS-induced TNF-α levels in trained individuals compared to untrained individuals (31). In the context of acute exhaustive exercise, a decrease in plasma TNF-α levels in mice in response to LPS was attributed to the suppressive impact of this type of effort on the inflammatory response (34, 35). Here, despite a similar proportion of circulating monocytes at baseline and after the acute exercise session, master athletes showed an increased release of TNF-α by PBMCs when stimulated with LPS only in the exercise condition, suggesting that acute exercise impacts the production of cytokines in response to LPS in master athletes.
MIP-1α (CCL3) levels, which attract monocytes and other immune cells to inflammatory sites, were consistently lower in master athletes at both baseline and exercise conditions. On the other hand, MIP-1α levels increased only in the younger group after acute exercise. Although high MIP-1α levels may help recruit immune cells rapidly to sites of damage or infection, chronically elevated levels could contribute to tissue damage or prolonged inflammation. Previous work suggests that regular exercise may reprogram macrophages toward an anti-inflammatory phenotype (e.g. M2-like, typically involved in the removal of tissue debris, tissue remodeling and repair and responsible for large production of IL-10-associated genes), potentially decreasing LPS-induced NF-κB activation and reducing pro-inflammatory gene expression (36). Furthermore, monocytes from individuals with higher physical fitness levels showed a more efficient cellular framework than those with lower physical fitness levels (33). This shift in master athletes might explain the relatively restrained TNF-α and MIP-1α responses in the present study, without completely compromising the ability to mount an inflammatory response when needed (37).
Corroborating with this scenario, we also observed that LPS stimulation increased IL-6 in whole blood from both age groups, but this response was larger in young athletes at post-exercise. Muscle-derived IL-6 is known to shift macrophages toward an anti-inflammatory phenotype (38–40). Hence, the attenuated rise in master athletes could reflect an adaptative, more balanced inflammatory milieu honed over years of regular training (6, 9, 41–46). Similarly, under PMA/ionomycin stimulation, young athletes showed greater IFN-γ and TNF-α production in PBMCs, whereas master athletes showed minimal changes. While such a reduced cytokine release might raise concerns about “exhaustion” (47), neither IL-10 nor PD-1 (PDCD1 gene) was significantly elevated in master athletes. Immune exhaustion is characterized by reduced levels of effector cytokines (such as IFN-γ and TNF-α) and an increase in inhibitory signals [such as IL-10 and PD-1—an inhibitory receptor that can inhibit cell activity upon engagement with its ligands PD-L1 or PD-L2, leading to immune exhaustion (48). We found that IL-10 production by PBMCs, as well as the gene expression of TNF-α and PDCD1 (gene coding PD-1) were similar between groups. Furthermore, NK-cell markers of senescence or terminal differentiation (such as KLRG1 and PD-1) did not differ between groups, thereby suggesting that the attenuated inflammatory response in master athletes need not imply a pathologic immune exhaustion. Taken together, these findings suggest a comparable capacity to counteract inflammation across ages, which is crucial to prevent ‘excessive’ immune responses (8, 44).
Interestingly, an exercise-specific increase was noted in PD-1 expression on NK cells and CD56+CD16+ cells, irrespective of age, indicating that acute exercise mobilizes not only effector lymphocytes but also those with a higher cytotoxic profile. As reviewed by Simpson (49) and supported by others (20, 50–52), acute exercise preferentially mobilizes senescent and exhausted T-cells which undergo apoptosis in peripheral tissues during the recovery period (53, 54). This creates an “immunological space” for naïve lymphocytes. Moreover, we observed a stimulus-specific effect on gene expression of FAS—a cell surface-bound receptor that triggers apoptosis (55)—in PBMCs. Whether the exercise-induced increase in PD-1 levels on NK cells causes the apoptosis of these cells during the recovery period (e.g. 1h, 2h) or downregulates NK functionality through enhanced PD-1 signaling needs to be further investigated.
4.1 Limitations and future perspectives
Although our strict inclusion criteria have eliminated potential confounding factors (e.g., age-related physical inactivity and elevated fat mass) on the inflammatory response, we acknowledge that the small sample size and unequal distribution of sex and sport disciplines between groups may have limited the generalizability of the findings and introduced variability that could affect assumptions of statistical tests. Future studies with larger and more balanced cohorts are warranted to validate these results. We recognize that the absence of an age-matched untrained control group limits our ability to attribute the observed responses solely to lifelong training without considering the effects of aging in the absence of training. However, from the present results, it cannot be excluded that regular exercise training in master athletes could optimize the ability to respond to stressors (like LPS and PMA stimulation), resulting in lower baseline TNF-α levels compared to young athletes but at the same time increasing TNF-α and IL-6 compared to baseline as part of the inflammatory response. We used a 24-hour stimulation period which may not be optimal for measuring IL-6 levels because it might have resulted in missing differences in the initiation or duration of cytokine secretion (56). This could explain why IL-6 release decreased in PBMCs following stimulation with PMA/ionomycin. Another point which needed be point out is the higher values IL-6, we are acknowledging this limitation and suggesting future experiments to compare PBMC activation markers (e.g., CD69, HLA-DR) and IL-6 mRNA expression post-isolation to determine whether early cell activation contributes to the observed findings.
While individual environmental exposures were not quantified (57, 58), our cohort trained under broadly similar mild-temperature, low-pollution conditions, and all sampling occurred at 22 ± 1°C and 50 ± 5% relative humidity—thus reducing the likelihood that external stimuli (heat, cold, or airborne irritants) systematically influenced our assays. Although our master athletes were free of diagnosed inflammatory diseases and showed no elevations in baseline IFN-, IL-18, or IL-10—and even lower MIP-1α compared to young athletes—subclinical low-grade inflammation cannot be entirely excluded. Future work incorporating other aging-related biomarkers would further delineate the interplay between chronological aging and exercise-induced immune adaptations. Further investigations with larger cohorts and including untrained young and older adults would benefit from the present findings to be able to compare age-related vs. training-related immunological adaptations.
5 Conclusion
In summary, our findings indicate that master athletes exhibit a generally lower pro-inflammatory cytokine response to ex vivo stimulation compared to younger athletes, while still maintaining the capacity to upregulate certain mediators following exercise (e.g., maintaining IL-6 release and increasing LPS-induced TNF-α post-exercise). These results suggest a pattern of more controlled inflammatory response in master athletes, balancing the need to mount an appropriate immune response against the risk of excessive inflammation. Additionally, phenotypic markers of natural killer cell differentiation in master athletes were comparable to those of younger trained individuals. Future research with larger cohorts and age-matched controls is warranted to clarify whether these effects are unique to lifelong exercise rather than a feature of aging.
Data availability statement
The original contributions presented in the study are publicly available. This data can be found here: https://hdl.handle.net/11449/183294.
Ethics statement
The Ethical Institutional Review Board of the Universidade Estadual Paulista “Julio de Mesquita Filho” – Campus Presidente Prudente approved the study (approval number: 38701820.0.0000.5402). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
LM: Writing – review & editing, Conceptualization, Software, Funding acquisition, Writing – original draft, Resources, Investigation, Validation, Formal analysis, Project administration, Methodology, Supervision, Data curation, Visualization. AD: Formal analysis, Data curation, Writing – review & editing. CF: Writing – review & editing, Formal analysis, Data curation. GD: Data curation, Formal analysis, Writing – review & editing. AB: Writing – review & editing, Formal analysis. BS: Formal analysis, Data curation, Writing – review & editing. PV: Writing – review & editing. AP: Writing – review & editing, Resources. AL: Writing – review & editing. MS: Writing – review & editing, Resources. JR: Writing – review & editing, Funding acquisition. KK: Writing – review & editing. FL: Funding acquisition, Project administration, Conceptualization, Formal analysis, Supervision, Resources, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was funded by the Coordination for the Improvement of Higher Education Personnel (CAPES), Brazil (code 001). FSL was granted a research scholarship (PQ2) from the CNPq (302714/2021-9) and Regular Grant FAPESP (2022/13978-8). AAST (2023/07162-8, 2021/00352-0), CF (2019/26378-6), BSAs (2021/11932-8 and 2023/09216-8), PLV is supported by a postdoctoral contract granted by University of Castilla la Mancha and Fondo Social Europeo Plus (FSE+) (2024-UNIVERS-12850), and LM (2022/02723–9 and 2019/25626-6) were granted a research scholarship from São Paulo Research Foundation (FAPESP)/Brazil.
Acknowledgments
The authors thank all the volunteers and the students of the Laboratory of Exercise Cell Physiology (LAFICE) at UNESP, Presidente Prudente, Brazil, for their assistance and contribution to the development of this research.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1601405/full#supplementary-material
References
1. Fiuza-Luces C, Valenzuela PL, Gálvez BG, Ramírez M, López-Soto A, Simpson RJ, et al. The effect of physical exercise on anticancer immunity. Nat Rev Immunol. (2024) 24(4):282–93. doi: 10.1038/s41577-023-00943-0
2. Chow LS, Gerszten RE, Taylor JM, Pedersen BK, van Praag H, Trappe S, et al. Exerkines in health, resilience and disease. Nat Rev Endocrinol. (2022) 18:273–89. doi: 10.1038/s41574-022-00641-2
3. Steensberg A, Fischer CP, Keller C, Møller K, and Pedersen BK. IL-6 enhances plasma IL-1ra, IL-10, and cortisol in humans. Am J Physiol Endocrinol Metab. (2003) 285:E433–437. doi: 10.1152/ajpendo.00074.2003
4. Starkie R, Ostrowski SR, Jauffred S, Febbraio M, and Pedersen BK. Exercise and IL-6 infusion inhibit endotoxin-induced TNF-alpha production in humans. FASEB J Off Publ Fed Am Soc Exp Biol. (2003) 17:884–6. doi: 10.1096/fj.02-0670fje
5. Beiter T, Zügel M, Hudemann J, Schild M, Fragasso A, Burgstahler C, et al. The acute, short-, and long-term effects of endurance exercise on skeletal muscle transcriptome profiles. Int J Mol Sci. (2024) 25:2881. doi: 10.3390/ijms25052881
6. Minuzzi LG, Rama L, Bishop NC, Rosado F, Martinho A, Paiva A, et al. Lifelong training improves anti-inflammatory environment and maintains the number of regulatory T cells in masters athletes. Eur J Appl Physiol. (2017) 117:1131–40. doi: 10.1007/s00421-017-3600-6
7. Minuzzi LG, Rama L, Chupel MU, Rosado F, Martinho A, Paiva A, et al. Effects of lifelong training on senescence and mobilization of T lymphocytes in response to acute exercise. Exerc Immunol Rev. (2018) 24:72–84.
8. Minuzzi LG, Chupel MU, Rama L, Rosado F, Muñoz VR, Gaspar RC, et al. Lifelong exercise practice and immunosenescence: Master athletes cytokine response to acute exercise. Cytokine. (2019) 115:1–7. doi: 10.1016/j.cyto.2018.12.006
9. Perkins RK, Lavin KM, Raue U, Jemiolo B, Trappe SW, and Trappe TA. Effects of aging and lifelong aerobic exercise on expression of innate immune components in human skeletal muscle. J Appl Physiol. (2020) 129:1483–92. doi: 10.1152/japplphysiol.00615.2020
10. Simpson RJ, Campbell JP, Gleeson M, Krüger K, Nieman DC, Pyne DB, et al. Can exercise affect immune function to increase susceptibility to infection? Exerc Immunol Rev. (2020) 26:8–22.
11. Padilha CS, Figueiredo C, Minuzzi LG, Chimin P, Deminice R, Krüger K, et al. Immunometabolic responses according to physical fitness status and lifelong exercise during aging: New roads for exercise immunology. Ageing Res Rev. (2021) 68:101341. doi: 10.1016/j.arr.2021.101341
12. Rosa-Neto JC, Lira FS, Little JP, Landells G, Islam H, Pyne DB, et al. Immunometabolism-fit: How exercise and training can modify T cell and macro- phage metabolism in health and disease. Exerc Immunol Rev. (2022) 28:29–46.
13. Kawai T and Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol. (2010) 11:373–84. doi: 10.1038/ni.1863
14. Collins PE and Carmody RJ. The regulation of endotoxin tolerance and its impact on macrophage activation. Crit Rev Immunol. (2015) 35:293–323. doi: 10.1615/critrevimmunol.2015015495
15. Wiesenthal AA, Legroux TM, Richter C, Junker BH, Hecksteden A, Kessler SM, et al. Endotoxin tolerance acquisition and altered hepatic fatty acid profile in aged mice. Biology. (2023) 12:530. doi: 10.3390/biology12040530
16. Furman D, Campisi J, Verdin E, Carrera-Bastos P, Targ S, Franceschi C, et al. Chronic inflammation in the etiology of disease across the life span. Nat Med. (2019) 25:1822–32. doi: 10.1038/s41591-019-0675-0
17. Franceschi C, Garagnani P, Parini P, Giuliani C, and Santoro A. Inflammaging: a new immune–metabolic viewpoint for age-related diseases. Nat Rev Endocrinol. (2018) 14:576–90. doi: 10.1038/s41574-018-0059-4
18. Olsen I and Sollid LM. Pitfalls in determining the cytokine profile of human T cells. J Immunol Methods. (2013) 390:106–12. doi: 10.1016/j.jim.2013.01.015
19. Simões HG, Moreira SR, Moffatt RJ, and Campbell CSG. Methods to identify the anaerobic threshold for type-2 diabetic and non-diabetic subjects. Arq Bras Cardiol. (2010) 94:71–8. doi: 10.1590/s0066-782x2010000100012
20. Bigley AB, Rezvani K, Pistillo M, Reed J, Agha N, Kunz H, et al. Acute exercise preferentially redeploys NK-cells with a highly-differentiated phenotype and augments cytotoxicity against lymphoma and multiple myeloma target cells. Part II: Impact of latent cytomegalovirus infection and catecholamine sensitivity. Brain Behav Immun. (2015) 49:59–65. doi: 10.1016/j.bbi.2014.12.027
21. Adammek F, Wences Chirino TY, Walzik D, Trebing S, Belen S, Renpening D, et al. Kinetics of immune cell mobilization during acute aerobic exercise in healthy adults. Int J Sports Med. (2024) 45:908–16. doi: 10.1055/a-2338-5397
22. Poznanski SM, Barra NG, Ashkar AA, and Schertzer JD. Immunometabolism of T cells and NK cells: metabolic control of effector and regulatory function. Inflammation Res. (2018) 67:813–28. doi: 10.1007/s00011-018-1174-3
23. O’Brien KL and Finlay DK. Immunometabolism and natural killer cell responses. Nat Rev Immunol. (2019) 19:282–90. doi: 10.1038/s41577-019-0139-2
24. Bigley AB, Lowder TW, Spielmann G, Rector JL, Pircher H, Woods JA, et al. NK-cells have an impaired response to acute exercise and a lower expression of the inhibitory receptors KLRG1 and CD158a in humans with latent cytomegalovirus infection. Brain Behav Immun. (2012) 26:177–86. doi: 10.1016/j.bbi.2011.09.004
25. Barry JC, Simtchouk S, Durrer C, Jung ME, Mui AL, and Little JP. Short-term exercise training reduces anti-inflammatory action of interleukin-10 in adults with obesity. Cytokine. (2018) 111:460–9. doi: 10.1016/j.cyto.2018.05.035
26. Sun Y, Li H, Yang M-F, Shu W, Sun M-J, and Xu Y. Effects of aging on endotoxin tolerance induced by lipopolysaccharides derived from Porphyromonas gingivalis and Escherichia coli. PloS One. (2012) 7:e39224. doi: 10.1371/journal.pone.0039224
27. Kinoshita M, Nakashima M, Nakashima H, and Seki S. Immune mechanisms underlying susceptibility to endotoxin shock in aged hosts: implication in age-augmented generalized shwartzman reaction. Int J Mol Sci. (2019) 20:3260. doi: 10.3390/ijms20133260
28. Carapeto PV and Aguayo-Mazzucato C. Effects of exercise on cellular and tissue aging. Aging. (2021) 13:14522–43. doi: 10.18632/aging.203051
29. Young P, Rauch C, Russo I, Gaskell S, Davidson Z, and Costa RJS. Plasma endogenous endotoxin core antibody response to exercise in endurance athletes. Int J Sports Med. (2022) 43:1023–32. doi: 10.1055/a-1827-3124
30. Stewart LK, Flynn MG, Campbell WW, Craig BA, Robinson JP, McFarlin BK, et al. Influence of exercise training and age on CD14+ cell-surface expression of toll-like receptor 2 and 4. Brain Behav Immun. (2005) 19:389–97. doi: 10.1016/j.bbi.2005.04.003
31. Olesen J, Biensø RS, Meinertz S, van Hauen L, Rasmussen SM, Gliemann L, et al. Impact of training status on LPS-induced acute inflammation in humans. J Appl Physiol Bethesda Md. (2015) 1985:118:818–829. doi: 10.1152/japplphysiol.00725.2014
32. Scheffer D da L, Ghisoni K, Aguiar AS, and Latini A. Moderate running exercise prevents excessive immune system activation. Physiol Behav. (2019) 204:248–55. doi: 10.1016/j.physbeh.2019.02.023
33. Antunes BM, Rosa-Neto JC, Batatinha HAP, Franchini E, Teixeira AM, and Lira FS. Physical fitness status modulates the inflammatory proteins in peripheral blood and circulating monocytes: role of PPAR-gamma. Sci Rep. (2020) 10:14094. doi: 10.1038/s41598-020-70731-6
34. Tanaka Y, Kawanishi N, Shiva D, Tsutsumi N, Uchida M, Kitamura H, et al. Exhaustive exercise reduces tumor necrosis factor-alpha production in response to lipopolysaccharide in mice. Neuroimmunomodulation. (2010) 17:279–86. doi: 10.1159/000290044
35. Abbasi A, Hauth M, Walter M, Hudemann J, Wank V, Niess AM, et al. Exhaustive exercise modifies different gene expression profiles and pathways in LPS-stimulated and un-stimulated whole blood cultures. Brain Behav Immun. (2014) 39:130–41. doi: 10.1016/j.bbi.2013.10.023
36. Murugathasan M, Jafari A, Amandeep A, Hassan SA, Chihata M, and Abdul-Sater AA. Moderate exercise induces trained immunity in macrophages. Am J Physiol-Cell Physiol. (2023) 325:C429–42. doi: 10.1152/ajpcell.00130.2023
37. Swift ME, Burns AL, Gray KL, and DiPietro LA. Age-related alterations in the inflammatory response to dermal injury. J Invest Dermatol. (2001) 117:1027–35. doi: 10.1046/j.0022-202x.2001.01539.x
38. Kanda H, Tateya S, Tamori Y, Kotani K, Hiasa K, Kitazawa R, et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J Clin Invest. (2006) 116:1494–505. doi: 10.1172/JCI26498
39. Oliveira AG, Araujo TG, Carvalho BM, Guadagnini D, Rocha GZ, Bagarolli RA, et al. Acute exercise induces a phenotypic switch in adipose tissue macrophage polarization in diet-induced obese rats. Obes Silver Spring Md. (2013) 21:2545–56. doi: 10.1002/oby.20402
40. Chazaud B. Inflammation and skeletal muscle regeneration: leave it to the macrophages! Trends Immunol. (2020) 41:481–92. doi: 10.1016/j.it.2020.04.006
41. Lavin KM, Perkins RK, Jemiolo B, Raue U, Trappe SW, and Trappe TA. Effects of aging and lifelong aerobic exercise on basal and exercise-induced inflammation. J Appl Physiol. (2020) 128:87–99. doi: 10.1152/japplphysiol.00495.2019
42. Gao H-E, Wu D-S, Sun L, Yang L-D, Qiao Y-B, Ma S, et al. Effects of lifelong exercise on age-related body composition, oxidative stress, inflammatory cytokines, and skeletal muscle proteome in rats. Mech Ageing Dev. (2020) 189:111262. doi: 10.1016/j.mad.2020.111262
43. de Souza Teixeira AA, Minuzzi LG, Lira FS, Gonçalves ASVP, Martinho A, Rosa Neto JC, et al. Improvement in the anti-inflammatory profile with lifelong physical exercise is related to clock genes expression in effector-memory CD4+ T cells in master athletes. Exerc Immunol Rev. (2021) 27:67–83.
44. Hayes LD, Herbert P, Sculthorpe NF, and Grace FM. Short-term and lifelong exercise training lowers inflammatory mediators in older men. Front Physiol. (2021) 12:702248. doi: 10.3389/fphys.2021.702248
45. Aguilera JA, Pourhashemi N, Sharpe CJ, and Friesen B. Use it or lose it: protecting ageing muscles with lifelong recreational exercise. J Physiol. (2022) 600:3397–8. doi: 10.1113/JP283338
46. Dias MR de J, Correia TML, Borges J de O, Santos LSD, Pereira R, da SRAA, et al. Therapeutic or lifelong training effects on pancreatic morphological and functional parameters in an animal model of aging and obesity. Exp Gerontol. (2023) 175:112144. doi: 10.1016/j.exger.2023.112144
47. Brooks DG, Ha S-J, Elsaesser H, Sharpe AH, Freeman GJ, and Oldstone MBA. IL-10 and PD-L1 operate through distinct pathways to suppress T-cell activity during persistent viral infection. Proc Natl Acad Sci U.S.A. (2008) 105:20428–33. doi: 10.1073/pnas.0811139106
48. Philips EA, Liu J, Kvalvaag A, Mørch AM, Tocheva AS, Ng C, et al. Transmembrane domain–driven PD-1 dimers mediate T cell inhibition. Sci Immunol. (2024) 9:eade6256. doi: 10.1126/sciimmunol.ade6256
49. Simpson RJ. Aging, persistent viral infections, and immunosenescence: can exercise “make space”? Exerc Sport Sci Rev. (2011) 39:23–33. doi: 10.1097/JES.0b013e318201f39d
50. Wadley AJ, Cullen T, Vautrinot J, Keane G, Bishop NC, and Coles SJ. High intensity interval exercise increases the frequency of peripheral PD-1+ CD8+ central memory T-cells and soluble PD-L1 in humans. Brain Behav Immun - Health. (2020) 3:100049. doi: 10.1016/j.bbih.2020.100049
51. Schenk A, Esser T, Belen S, Gunasekara N, Joisten N, Winker MT, et al. Distinct distribution patterns of exercise-induced natural killer cell mobilisation into the circulation and tumor tissue of patients with prostate cancer. Am J Physiol-Cell Physiol. (2022). doi: 10.1152/ajpcell.00243.2022
52. Proschinger S, Schenk A, Weßels I, Donath L, Rappelt L, Metcalfe AJ, et al. Intensity- and time-matched acute interval and continuous endurance exercise similarly induce an anti-inflammatory environment in recreationally active runners: focus on PD-1 expression in Tregs and the IL-6/IL-10 axis. Eur J Appl Physiol. (2023) 123:2575–84. doi: 10.1007/s00421-023-05251-y
53. Krüger K, Alack K, Ringseis R, Mink L, Pfeifer E, Schinle M, et al. Apoptosis of T-cell subsets after acute high-intensity interval exercise. Med Sci Sports Exerc. (2016) 48:2021–9. doi: 10.1249/MSS.0000000000000979
54. Alack K, Krüger K, Weiss A, Schermuly R, Frech T, Eggert M, et al. Aerobic endurance training status affects lymphocyte apoptosis sensitivity by induction of molecular genetic adaptations. Brain Behav Immun. (2019) 75:251–7. doi: 10.1016/j.bbi.2018.10.001
55. Schneider P and Tschopp J. Apoptosis induced by death receptors. Pharm Acta Helv. (2000) 74:281–6. doi: 10.1016/s0031-6865(99)00038-2
56. Mandala W, Harawa V, Munyenyembe A, Soko M, and Longwe H. Optimization of stimulation and staining conditions for intracellular cytokine staining (ICS) for determination of cytokine-producing T cells and monocytes. Curr Res Immunol. (2021) 2:184–93. doi: 10.1016/j.crimmu.2021.10.002
57. Kurowski M, Jurczyk J, Moskwa S, Jarzębska M, Krysztofiak H, and Kowalski ML. Winter ambient training conditions are associated with increased bronchial hyperreactivity and with shifts in serum innate immunity proteins in young competitive speed skaters. Arch Med Sci. (2017) 14:60–8. doi: 10.5114/aoms.2017.69438
Keywords: inflammation, athletes, moderate exercise, natural killer cells, monocytes
Citation: Minuzzi LG, De Souza Teixeira AA, Figueiredo C, Dorneles G, Branco ACC, Silva BSdA, Valenzuela PL, Peres A, Lucia A, Sato MN, Rosa Neto JC, Krüger K and Lira FS (2025) Differential inflammatory responses to acute exercise and ex vivo immune challenge in young and master athletes. Front. Immunol. 16:1601405. doi: 10.3389/fimmu.2025.1601405
Received: 27 March 2025; Accepted: 07 July 2025;
Published: 31 July 2025.
Edited by:
Malen Massot-Cladera, University of Barcelona, SpainReviewed by:
Nick M. Filipov, University of Georgia, United StatesMarcin Kurowski, Medical University of Lodz, Poland
Copyright © 2025 Minuzzi, De Souza Teixeira, Figueiredo, Dorneles, Branco, Silva, Valenzuela, Peres, Lucia, Sato, Rosa Neto, Krüger and Lira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fabio Santos Lira, ZmFiaW8ubGlyYUB1bmVzcC5icg==; Luciele Guerra Minuzzi, bHVjaWVsZWdtQGdtYWlsLmNvbQ==