 Hala Ahmad
Hala Ahmad Ahmad Alatshan
Ahmad Alatshan Eduárd Bíró
Eduárd Bíró Szilvia Benkő
Szilvia Benkő- 1Laboratory of Inflammation-Physiology, Department of Physiology, Faculty of Medicine, University of Debrecen, Debrecen, Hungary
- 2Doctoral School of Molecular Cell and Immune Biology, Faculty of Medicine, University of Debrecen, Debrecen, Hungary
Macrophages are indispensable in homeostasis and innate immune responses in multiple tissues, while their polarization and functional characteristics are determined by the activating stimuli and their tissue microenvironment. The vitamin A derivative retinoic acid shows inhomogeneous distribution among the tissues and has an important modulatory role in inflammatory responses. However, its effects on the cytokine secretion induced by the cytosolic pattern-recognition receptors NOD1 and NOD2 are unclear. In our study, we used human monocyte-derived macrophages differentiated in the presence of GM-CSF or M-CSF to generate inflammation inducing (GM-MФ) or inflammation resolving (M-MФ) cells, respectively. We activated the cells with either a NOD1- or NOD2 specific agonist and, using ELISA, we determined the pattern and dynamics of cytokines secreted by the macrophage subpopulations. Furthermore, we studied the effect of all-trans retinoic acid (ATRA) pre-treatment on the NOD1- and NOD2-induced cytokine release. Our comparative analysis shows subpopulation-characteristic pattern of cytokine secretion, as GM-MФ produce significantly higher pro-inflammatory IL-6, IL-8, TNF-α and IL-1β, while M-MФ secret higher anti-inflammatory IL-10. However, IL-18 and IFNβ secretion was comparable between the MФ subpopulations. We also show for the first time that ATRA has marked impact on cytokine secretion triggered by NOD1 and NOD2. Importantly however, the ATRA-induced changes of cytokine secretion follow opposite tendency in two MФ subpopulations. In conclusion, these results show that NOD1/NOD2-induced cytokine secretion by macrophage subsets is highly context-dependent and our results highlight the importance of the retinoic acid content of the local tissue environment in shaping macrophage function in health and disease.
Introduction
Retinoic acid (RA) is the most prevalent and metabolically active vitamin A derivative in the human body that regulates a wide range of biological processes including cell proliferation and differentiation, embryogenesis and development, furthermore it supports the homeostasis of various tissues (1). The pleiotropic effect of RA are mediated by nuclear receptors either via genomic mechanisms by regulating transcription processes; or via non-genomic mechanisms by directly modifying the activity of various signal transduction pathways (2). Importantly, RA has also been shown to play a crucial role in the modulation of various immune functions. These include the differentiation and maturation of immune cells of both the innate- and adaptive immune system, such as lymphocytes, dendritic cells and macrophages (3, 4). Specifically in macrophages, RA has been implicated in various functions including phagocytosis, efferocytosis as well as balancing inflammatory responses through shaping cytokine production (3, 5, 6). Importantly, the modulatory effect of RA on inflammatory responses is highly context dependent as it is affected by the cell type, the stimuli and the tissue microenvironment.
Macrophages (MФs) are indispensable effector cells to modulate inflammatory responses. They form a diverse population of innate immune cells that are present in various tissues and organs where they either provide homeostatic functions as resident MФs, or support protective mechanism as infiltrating MФs by inducing or resolving inflammatory responses (7). Importantly, MФ inflammatory activity involves a complex molecular network including the cooperative crosstalk of pattern recognition receptors, signaling cascades and cytokine release. It is widely recognized that NOD-like receptors (NLRs), a family of cytosolic pattern recognition receptors that regulate various innate immune response, are highly involved in MФs functions (8).
NOD1 and NOD2 are prominent members of the NLR family as they were the earliest to be identified and characterized among the mammalian NLRs, but their roles in the inflammatory responses of MФ subsets are unknown. NOD1 and NOD2 were initially described to sense conserved, but distinct motifs of bacterial peptidoglycan (PGN), however, they can be activated by non-bacterial microorganisms (such as viruses, parasites and fungi) as well as danger-associated molecules (such as sphingosine-1-phosphate released during ER stress (9). Following activation, NOD1 and NOD2 participate in the regulation of cellular processes including metabolism, autophagy, cell death (10, 11) and the secretion of various cytokines (12–14).
Accumulating evidence shows that cytokine secretion by activated myeloid cells is highly modulated by RA (1). Accordingly, ATRA has been reported to enhance anti-inflammatory IL-10 secretion, but suppress IL-12 and TNF-α secretion in LPS-activated THP-1 cell line and splenic MФs (15, 16); while in human alveolar MФs and THP-1 cell line, LPS-induced IL-β expression is enhanced by ATRA (17, 18). Furthermore, we reported that in M-MФs, the LPS–induced IL-6 and IL-1β secretion is enhanced, while IL-10 is down-regulated by ATRA (19). While both NOD1 and NOD2 are important pattern recognition receptors that ignite inflammatory responses, including cytokine secretion in MФs, and the local microenvironment of MФs in several tissues may contain substantial amount of RA, the effect of RA on NOD1- and NOD2-mediated inflammatory cytokine and chemokine release has not been addressed before. Thus, we hypothesized that ATRA may modulate MФ inflammatory responses by interfering with NOD1/NOD2-induced cytokine responses in different MФ subsets.
To this end, we carried out a comparative analysis of typical inflammatory cytokine secretion induced by NOD1 and NOD2 in two distinct subpopulations of human monocyte-derived MФs that are differentiated in the presence of either GM-CSF or M-CSF, and that are commonly used in vitro models of pro-inflammatory- (GM-MФs) or pro-resolving cells (M- MФs), respectively. We focused on a few cytokines that are well-established to mediate the functional responses of different MФ populations as typical pro-inflammatory cytokines (IL-1β/IL-18, IL-6, TNF-α), or anti-inflammatory and pro-resolving cytokines (IL-10, IFNβ) (20, 21). IL-8 was specifically selected as a well-known downstream target of NOD1/NOD2 activation (22, 23). We show that the activation of NOD1 or NOD2 results in the release of different cytokine patterns by the two MФ subpopulations. Furthermore, we report for the first time that ATRA highly modulates cytokine secretion triggered by NOD1 and NOD2. Importantly however, the ATRA-induced changes of cytokine secretion by the two MФ subpopulations follow opposite tendency.
Materials and methods
Ethical statement
Buffy coat from healthy blood donors was provided by the regional National Blood Transfusion Service. The procedure was documentary approved by the Director of the National Blood Transfusion Service (Permit number: OVSZK/1678-2/2024/3090). The study and all experimental protocols were in accordance with the Regional and Institutional Ethics Committee of the University of Debrecen (Debrecen, Hungary). and the Regional and Institutional Ethics Committee of the University of Debrecen, Hungary.
Monocyte isolation
Peripheral human blood mononuclear cells (PBMCs) were extracted from buffy coats using density gradient centrifugation with Ficoll Paque PLUS (GE Healthcare Life Sciences, Little Chalfont, United Kingdom). Then, monocytes were isolated by positive immunomagnetic cell selection using CD14 microbeads according to the manufacturer’s instruction (Miltenyi Biotec, Bergisch Gladbach, Germany).
Macrophage differentiation
The isolated monocytes were cultured in RPMI 1640 medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 2 mM L-glutamine, 10% heat-inactivated FBS, 100 U/mL penicillin-streptomycin and differentiated with either M-CSF (50ng/ml) or GM-CSF (80ng/ml) (PeproTech, Rocky Hill, NJ, USA). Cells were seeded in 24-well plates at a concentration of 1.1 x 106 cells/mL at 37°C and 5% CO2. Half of the culture media was replaced with fresh media containing M-CSF or GM-CSF on day 2. The cells were differentiated for 5 days.
Macrophage treatment
Macrophages were treated on Day 5 with C14-Tri-LAN-Gly (NOD1 specific agonist) (500ng/ml) (InvivoGen, San Diego, CA, USA) or L18-MDP (NOD2 specific agonist) (100ng/ml) (InvivoGen, San Diego, CA, USA) for the indicated time points. Where indicated, cells were pretreated with ATRA (1uM) for 4 hours (Sigma-Aldrich, St. Louis, MO, USA).
Cytokine secretion measurements
Cytokine secretion levels were quantified using enzyme-linked immunosorbent assay (ELISA) according to the manufacturer’s instructions, IL-1β, IL-8 and TNF-α were measured using ELISA kits from BD Biosciences (San Diego, CA, USA). IL-18, IL-6, IL-10 and IFNβ were measured using ELISA kits from R&D Systems (Minneapolis, MN, USA). A microplate reader was used at 450 nm absorbance to determine cytokine levels (FlexStation 3, Molecular Devices, Sunnyvale, CA, USA). The minimum detectable levels were 3.9 pg/mL for IL-1β, 3.1 pg/mL for IL-8, 7.8 pg/mL for TNF-α, 11.7 pg/ml for IL-18, 9.38 pg/mL for IL-6, 31.3 pg/mL for IL-10 and 7.81 pg/mL for IFNβ.
Statistical analysis
Results are presented as mean ± standard deviation (SD). Statistical analysis was conducted using One-way ANOVA followed by Tukey’s post hoc comparison. GraphPad Prism (version 8.4) was used for data analysis (GraphPad Software, San Diego, California, USA). Differences are considered significant at p value less than 0.05. In the figures, the asterisk (*) sign indicates significant difference between the NOD1/NOD2-treated and the control group at one given time-point. The hash symbol (#) indicates statistically significant difference between M- and GM-MФs at a given time-point.
Results
NOD1 differently activates human MФ subpopulations
Activation of NOD1 triggers the induction of various signaling pathways via RIPK2, and results in secretion of different inflammatory cytokines (24). To get a better insight into the dynamics of NOD1-induced cytokine secretion by the human monocyte-derived MФ subpopulations, cells were treated with NOD1 specific agonist (C14-Tri-LAN-Gly), and supernatant was collected at different time-points for ELISA measurements. The comparative analysis between the MФ subpopulations revealed significant differences in cytokine secretion (Figure 1A). While we measured a dynamic increase in IL-6 for both MФ types during the 24 h time interval, in TNF-α secretion we observed a peak at 16 h in both subpopulations. Nevertheless, the secretion of both cytokines was significantly higher in the supernatants of GM-MФs compared to M-MФs. We observed similar tendency in IL-8 chemokine secretion, as we measured gradually increasing, high level of IL-8 production by GM-MФs following NOD1 agonist treatment, while it was significantly less in the supernatant of M-MФs. Importantly, the opposite tendency was observed for the anti-inflammatory IL-10. While IL-10 was induced in both MФ subpopulations, IL-10 release was significantly stronger in M-MФs compared to GM-MФs. Though weakly expressed, but similar tendency was observed for IFNβ, an upstream regulatory cytokine of IL-10 (Supplementary Figure 1A). These results indicate that while NOD1 induces cytokine secretion, it results in a different cytokine profile by the different MФ subsets.
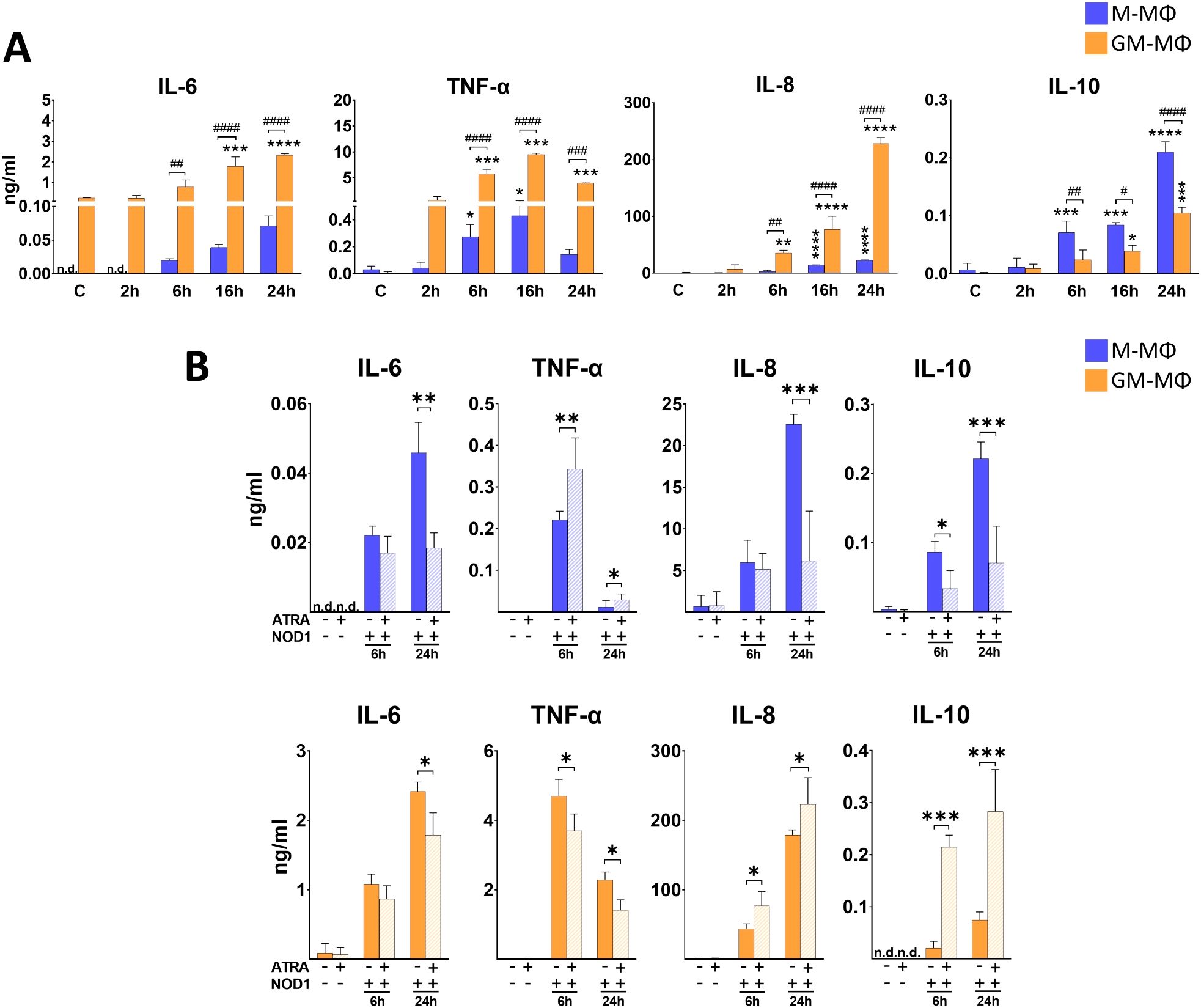
Figure 1. Secretion of IL-6, TNF-α, IL-8 and IL-10 following NOD1 activation. M-MФ and GM-MФ were treated with C14-Tri-LAN-Gly (NOD1 agonist, 500 ng/ml) for the indicated time points. Control cells were treated with the same amount of vehicle as the activated cells. Cytokine secretion was measured from the supernatant using ELISA. (A) Time-kinetics of cytokine secretion. (B) Cells were pretreated with ATRA (1µM) for 4 hours and then stimulated with C14-Tri-LAN-Gly for 6 and 24 hours. Results were obtained from at least four healthy donors. All results are shown as means ± SD. (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001; n.d., not detected).
NOD1-induced cytokine secretion is differently modified by ATRA in the MФ subpopulations
To determine whether ATRA has any modulatory effect on NOD1-induced cytokine secretion, cells were pretreated with ATRA, and supernatant was collected at various time-points following NOD1 activation. Of note, though we detected the same tendency of ATRA effect on cytokine secretion at each time-points (2h, 6h, 16h, 24h), we decided to only present an early (6h) and a late (24h) time point in the manuscript (2h and 16h data are not shown) (Figure 1B). Compared to the non-treated controls, ATRA treatment alone did not change the secretion of cytokines at any time-points. In the presence of ATRA, we found significantly decreased secretion of IL-6 in both NOD1-activated MФ subpopulations. However, surprisingly, TNF-α levels were differently affected in the two MФs. While ATRA increased NOD1-induced TNFα secretion in M-MФs, the secretion of this cytokine was significantly attenuated in GM-MФs by ATRA pretreatment. Interestingly, opposite effect was detected for IL-8 production, as ATRA highly down-regulated the secretion in M-MФs, while significantly enhanced secretion was observed in the supernatant of ATRA-treated NOD1-activated GM-MФs. Surprisingly, ATRA also appeared to polarize release of the anti-inflammatory IL-10 and IFNβ production; (Supplementary Figure 1C) in an opposite direction to IL-6 and TNF-α, and its levels were decreased by ATRA in GM-MФs, while it was increased in M-MФs, similarly to IL-8. Altogether, these results show that ATRA highly modulates NOD1-induced cytokine secretion in human MФs, and this modulation is cytokine- and cell type-dependent.
NOD2 induces cytokine secretion by different human MФ subpopulations with similar tendency as NOD1
While NOD1 specifically recognizes peptidoglycan components of Gram (–) bacteria, NOD2 - a very close family member of NOD1 - is able to recognize peptidoglycan pattern from both Gram (–) and Gram (+) bacteria. Hence, next we aimed to determine the cytokine secretion by the two MФ subpopulations following NOD2 activation. Cells were treated with NOD2 agonist L18-MDP, and supernatant was collected for cytokine measurement (Figure 2A). While we measured higher level of NOD2-induced IL-6 and TNF-α in GM-MФs compared to the M-MФs, NOD2 treatment resulted in a gradually increasing TNF-α secretion during the 24 h treatment in GM-MФs, in contrast to the 12 h peak of TNF-α by the NOD1-induced cells. Similar to NOD1 activation, NOD2 treatment resulted in robust IL-8 secretion in GM-MФs, while it was released at a moderate level by M-MФs. NOD2 activation resulted in higher IL-10 secretion in M- MФs compared to GM- MФs, similar to that observed with NOD1 activation. IFN-β release was gradually increasing and comparable between the MФ populations (Supplementary Figure 1B).
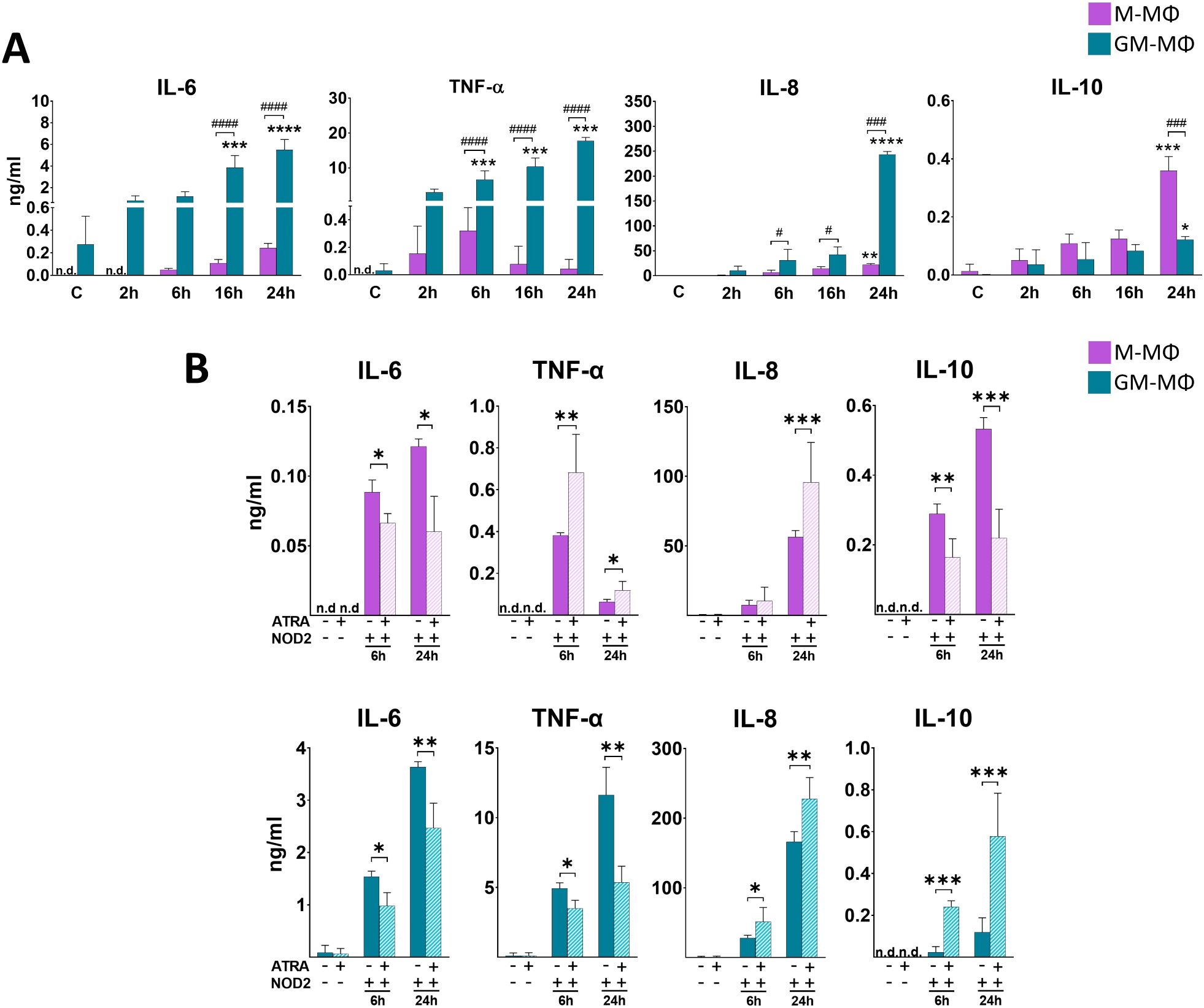
Figure 2. Secretion of IL-6, TNF-α, IL-8 and IL-10 following NOD2 activation. M-MФ and GM-MФ were treated with L-18 MDP (NOD2 agonist, 100 ng/ml) for the indicated time points. Control cells were treated with the same amount of vehicle as the activated cells. Cytokine secretion was measured from the supernatant using ELISA. (A) Time kinetics of cytokine secretion. (B) Cells were pretreated with ATRA (1µM) for 4 hours and then stimulated with L-18 MDP for 6 and 24 hours. Results were obtained from at least four healthy donors. All results are shown as means ± SD. (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, #p < 0.05, ###p < 0.001, ####p < 0.0001; n.d., not detected).
In M-MФs, ATRA differently modifies NOD2-induced IL-8 secretion compared to induction by NOD1
Then, we aimed to see the potential modulatory effect of ATRA on NOD2-induced cytokines production. Similar to NOD1, in the presence of ATRA we detected IL-6 decrease both in M-MФs and GM-MФs following NOD2 activation (Figure 2B). Similar to NOD1, opposite effects were observed for TNF-α and IL-10 secretion between the two cell types; while ATRA enhanced TNF-α and decreased IL-10 secretion in M-MФs, ATRA decreased TNF-α and enhanced IL-10 production in GM-MFs. Interestingly, however, IL-8 chemokine secretion was significantly upregulated by ATRA in both M- and GM-MФs, in contrast to the NOD1-induced IL-8 secretion in M-MФs, where ATRA downregulated the cytokine secretion. Again, similar tendency was measured for IL-10 secretion (and IFNβ production; (Supplementary Figure 1D)); the anti-inflammatory cytokine was decreased by ATRA in M-MФs, while it was increased in GM-MФs.
ATRA differently modifies NOD1 ligand-induced IL-1β and IL-18 secretion in the human MФ subpopulations
In contrast to the other cytokines, the maturation of pro-IL-1β and pro-IL-18 pro-inflammatory cytokines to their active form requires proteolytic cleavage by caspase enzymes. Interestingly, cell activation with NOD1 agonist treatment has been implicated in IL-1β/IL-18 secretion (12). To determine if activation of the monocyte-derived MФs with NOD1 agonist induce their secretion, supernatant was collected for ELISA measurements at different time points following NOD1 agonist treatment. We observed a rapid induction with an early peak (6 h) in IL-1β secretion in GM-MФs, while IL-1β secretion by M-MФs was hardly detectable (Figure 3A). In the case of IL-18, the secretion peaked at 16 h in both MФs, and the secretion level of IL-18 was comparable in the two MФ populations (Figure 3A). However, while in M-MФs both IL-1β and IL-18 secretion showed increasing tendency in the presence of ATRA, we observed an opposite effect in GM-MФs, as the secretion of both IL-1β and IL-18 was significantly attenuated by ATRA pretreatment (Figure 3B).
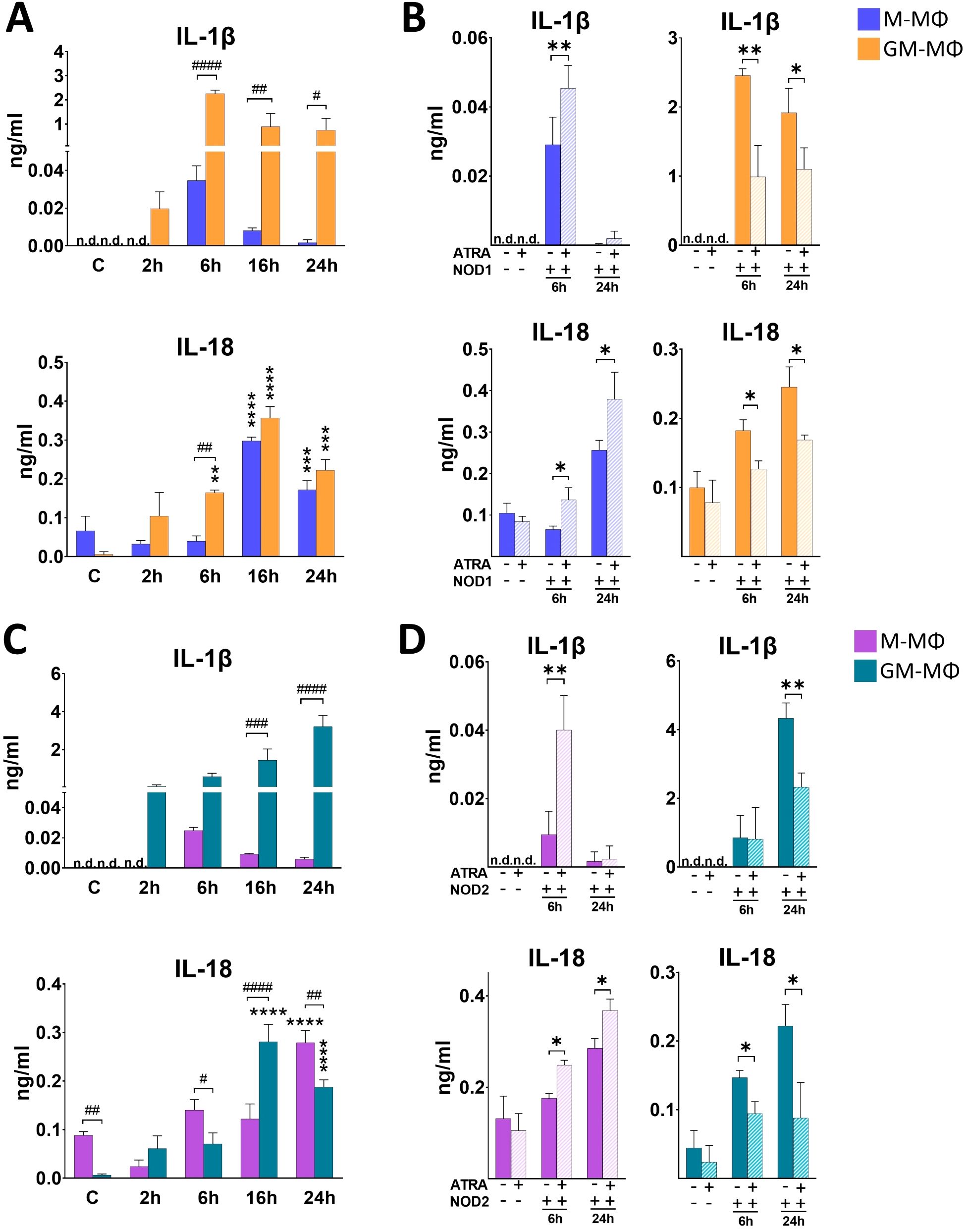
Figure 3. Secretion of IL-1β and IL-18 following NOD1 or NOD2 activation. M-MФ and GM-MФ were treated with (A, B) C14-Tri-LAN-Gly (NOD1 agonist, 500 ng/ml) or (C, D) L-18 MDP (NOD2 agonist, 100 ng/ml) for the indicated time points. Control cells were treated with the same amount of vehicle as the activated cells. Cytokine secretion was measured from the supernatant using ELISA. (A, C)) Time-kinetics of cytokine secretion. (B, D) Cells were pretreated with ATRA (1µM) for 4 hours and then stimulated with NOD1 or NOD2 agonist for 6 and 24 hours. Results were obtained from at least four healthy donors. All results are shown as means ± SD. (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001; n.d., not detected).
NOD2-induced IL-1β and IL-18 secretion are also differently modified by ATRA in the human MФ subpopulations
Then, we aimed to see whether NOD2-activation induces IL-1β and IL-18 secretion, and if the amount and dynamics of these cytokine secretions are comparable with what we observed in the case of NOD1 activation. We found that NOD2 agonist treatment resulted in a gradual increase of both IL-1β and IL-18 in GM-MФs (Figure 3C). Similar to NOD1, however, IL-1β secretion by M-MФs was hardly detectable. Also, similar to NOD1, while ATRA enhanced NOD2-induced IL-1b and IL-18 secretion in M-MФs, the production of both cytokines was decreased in GM-MФs in the presence of ATRA (Figure 3D).
Discussion
Macrophages are remarkably plastic immune cells, whose functional characteristics are highly determined by their origin, and shaped by the tissue microenvironment of their location, as well as the activating stimuli that trigger their effector functions (25, 26). In vitro, differentiation of macrophages from human primary monocytes using GM-CSF or M-CSF are widely used methods to study macrophage polarization and functions, as the generated GM-MФs or M-MФs possess inflammation inducing or inflammation resolving macrophage characteristics, respectively (27, 28). Here we show that the activation of these primary monocyte-derived MФ subpopulations via NOD1 and NOD2 cytosolic receptors results in characteristic profile of secreted cytokines that are typically released during macrophage inflammatory responses, and contribute to pro-inflammatory or pro-resolving/tissue regenerating mechanisms. We detected significantly higher release of the inflammatory IL-6, TNF-α, IL-8 and IL-1β cytokines from the pro-inflammatory GM-MФs compared to the pro-resolving M-MФs. However, the secretion of IL-18 pro-inflammatory cytokine was comparable between the two cell types. Furthermore, although the level of anti-inflammatory IL-10 was more pronounced in the M-MФs compared to the GM-MФs, the level of IL-10 secretion was also in a comparable range between the two subtypes. These results highlight that depending on the type of MФs affected, NOD1/NOD2-mediated cytokine secretion may drive potentially different induction and polarization of cells which is also likely to shape their interactions with cells of adaptive immunity.
NOD1 and NOD2 are the first described members of the cytosolic pattern recognition NOD-like receptor family. Upon recognition of respective ligands by NOD1 or NOD2 induces recruitment of RIPK2 which then mediates the activation of NF-kB and MAPK (ERK, p38, JNK) signaling pathways and drives various cellular processes including cytokine/chemokine secretion (29). RIPK2 activity may also induce the nuclear translocation of IRF7 transcription factor to promote type I IFN release (29, 30) which is an efficient upstream signaling cytokine of IL-10. Furthermore, RIPK2 may also recruit and directly bind caspase-1, which is one of the major regulatory enzymes of IL-1β and IL-18 secretion (10). Besides, NOD1/NOD2 may interact with other, inflammasome forming Nod-like receptors to induce IL-1β and IL-18 secretion (12). However, our comparative analysis also reveals substantial differences in the amount of the secreted typical cytokines between the MФ subtypes, indicating that activation of NOD1/NOD2 results in a subpopulation characteristic pattern of cytokine secretion.
Importantly, these results are in line with our previous findings on LPS-activated MФ subtypes (31, 32). However, while LPS is a harsh activator that is detected by TLR4 and drives high level of IL-10 secretion from M-MФs and high level of pro-inflammatory cytokine from GM-MФs, NOD1/NOD2 induce milder effects and results in a less pronounced secretion of the corresponding cytokines. Importantly, however, we show now that the most highly induced cytokine by NOD1/NOD2 activation is the pro-inflammatory interleukin-8 (IL-8). Surprisingly, NOD1/NOD2-induced IL-8 levels are similar to what we reported following LPS stimulation from these types of cells (26, 27). IL-8 is a crucial regulator of angiogenesis and a potent neutrophil chemotactic factor, hence plays an important role in physiological and pathological conditions (33). It was shown that at low concentrations, IL-8 gradient attracts neutrophils to the inflammatory foci, while at high, receptor-saturating concentration, it triggers neutrophils to release granule proteins and chromatin to form neutrophil extracellular traps (NETs) (33, 34). Hence, we speculate that while both MФ populations possess high capability of neutrophil attraction, NOD1/NOD2-activated GM-MФs may even have good potency to induce NET formation. RA, the most common, physiologically active metabolite of vitamin A, appears as an important tissue-derived signal that contributes to MФ polarization. However, the presence and distribution of RA shows high variability between the organs and even within a tissue, providing different microenvironments to the MФs. While NOD1 and NOD2 are one of the main cytosolic pathogen recognition receptors of MФs, the potential modulatory effect of ATRA on NOD1/NOD2-mediated responses by the different MФ subpopulations has not been addressed before. Here, we show that ATRA significantly modifies cytokine and chemokine release induced by NOD1 and NOD2 activation, and importantly, ATRA treatment results in various effects in the cytokine secretion (Figure 4A). In M-MФs, we observed that the NOD1-induced pro-inflammatory TNF-α, IL-1β and IL-18 were upregulated, while IL-6 and IL-8 were down-regulated by ATRA. Furthermore, the NOD1-induced anti-inflammatory IL-10 was also decreased by ATRA. In GM-MФs, however, under the same conditions, we observed completely the opposite effect of ATRA on cytokine secretion, with the exception of IL-6. Indeed, IL-6 was the only cytokine that, in each condition, was affected the same way by ATRA, as it was down-regulated in both MФ subtypes following both NOD1 and NOD2 activation. Importantly, NOD2-induced cytokine secretions were affected with the same tendency by ATRA as in the case of NOD1 activation, except for IL-8, as the secretion of this cytokine was enhanced both in M-MФs and GM-MФs. Cytokines released by MФs may act in an autocrine or paracrine manner to regulate the secretion of other cytokines. Like IFNβ is an upstream regulator of IL-10 (20), and both IFNβ and IL-10 are negative regulators of caspase-1-mediated IL-1β secretion (35, 36), which may explain the negative correlation between IFNβ/IL-10 and IL-1β/IL-18 secretion by the MФs in the presence of ATRA in our results. Of note, it was reported that ATRA induces the expression of IRF1 transcription factor in human myeloid cell lines and MФs (37). On the other hand, IRF1 was identified as a transcription regulator of Interferon-regulated genes (ISG) by facilitating chromatin accessibility for IRF3 binding which is a major driver of IFNβ expression (38). Nevertheless, ATRA-induced engagement of IRF1 in IFNβ secretion requires further comparative analysis in these MФ subpopulations.
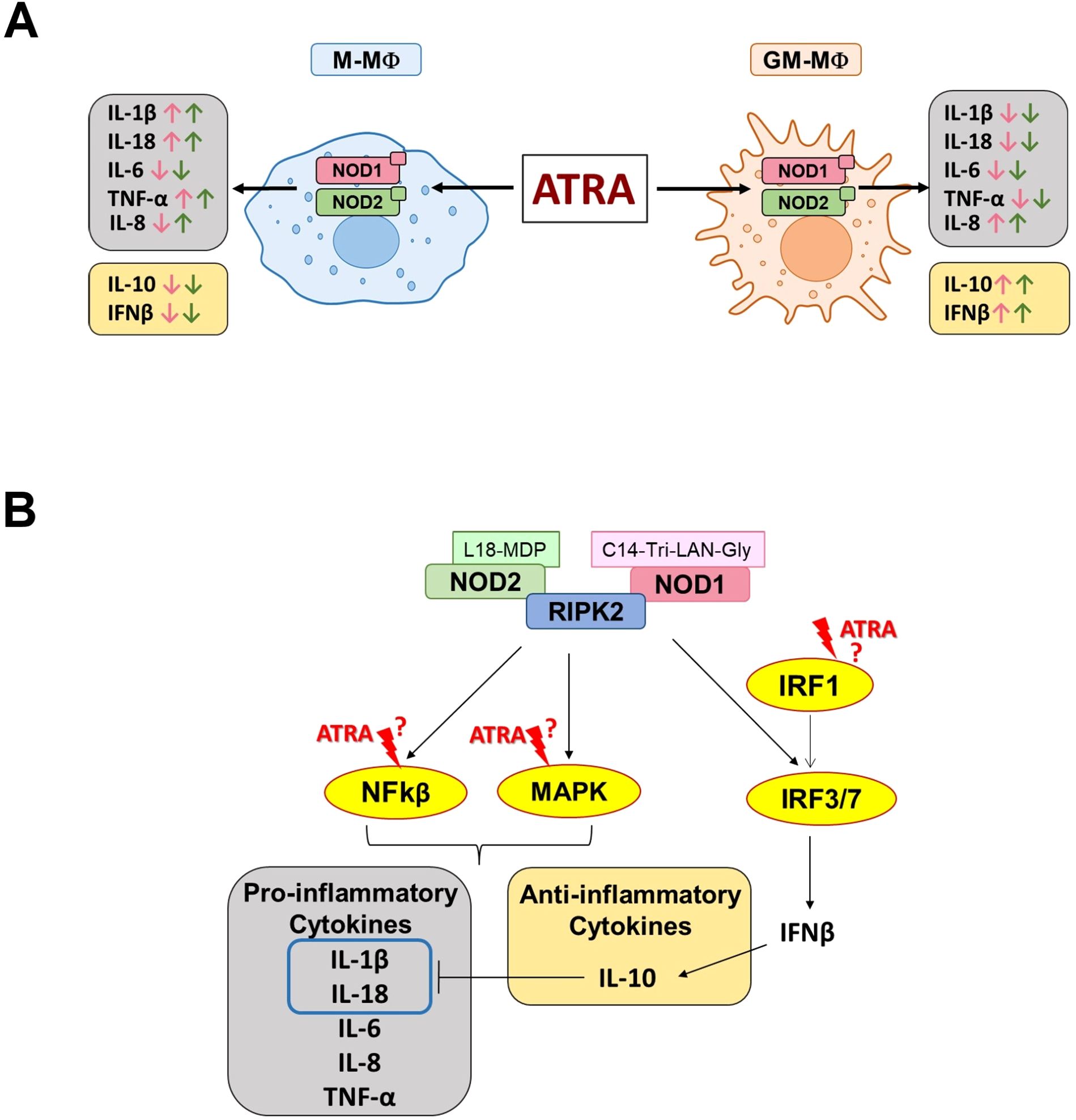
Figure 4. Schematic representation of ATRA-mediated effects on NOD1/NOD2-induced cytokine secretion. (A) Summary of the modulatory effect of ATRA on the NOD1/NOD2-induced cytokine secretion in the different subpolulations of human macrophages. Pink and green arrows indicate NOD1- and NOD2-induced cytokine changes, respectively. M-MФ (M-CSF-differentiated MФ), GM-MФ (GM-CSF-differentiated MФ). (B). Potential intervention points of ATRA on the NOD1/NOD2-induced signaling pathways leading to cytokine secretion. Following activation, NOD1/NOD2 recruits RIPK2 enzyme which may trigger the activation of various signal transduction pathways leading to the secretion of cytokines (29, 30).
ATRA may be recognized by various nuclear receptors (mainly by RAR, but also by PPAR or ROR) that may directly regulate gene expression in the nucleus (39). However, non-genomic effects of nuclear receptors have also been described, as in the cytosol, they can directly interact and modulate proteins of various signal transduction pathways (such as, AKT, NF-κB and ERK) (40). Importantly, many of these pathways overlap with those that are also affected by NOD1/NOD2 activation, rising the potential possibility of potential cross -talk mechanisms. Previous studies have reported context-dependent effect of ATRA on the expression of Toll-like receptors (TLRs) and RIG-like helicases (RLHs). Like ATRA downregulated the expression of TLR2 in monocytes (41) and TLR4 in the inflamed lung (6), while it upregulated TLR5 in THP-1 cells (42) and TLR4 expression in CaCo2 cells (43). ATRA alone upregulated TLR3, TLR4 and RIG-I intestinal epithelial cells, however it suppressed TLR3, TLR7 and RIG-I expression following viral infections (44, 45). Despite these observations, the mechanism of ATRA action has not been revealed. Regarding NLRs, the effect of ATRA has only been studied on the NLRP3 inflammasome activation. It has been reported that ATRA supplementation reduces the expression of NLRP3 inflammasome and caspase-1 in the brain tissue of a chronic alcoholic rat model (46). While, in our previous studies using in silico method, we found putative RAR-response elements in the promoter of NLRP3 and we detected increased NLRP3 protein level following ATRA treatment alone in human M-MФs. Furthermore, we found that ATRA shifts M-MФs metabolism toward glycolysis (19), which can highly effect MФs polarization. In this current study, we didn’t detect changes in the mRNA expression of NOD1/NOD2 (data not included) nor in the cytokine secretion following ATRA treatment alone, which suggests that ATRA acts as a modulator of NOD1/NOD2-induced cytokine production via downstream actions (and/or overall metabolic status of the cell) rather than inducing cytokine production alone.
It has been reported that ATRA treatment modulates several major intracellular signaling pathways, including MAPK signaling (such as ERK, JNK and p38). In human scleral fibroblasts, ATRA decreased ERK1/2 activation, while it increased JNK activation (47), however, in human MФs, ATRA enhanced the phosphorylation of ERK and attenuated that of p38 (19). The ATRA effect on NF-κB is also controversial, as it exhibits an inhibitory effect in THP-1, alveolar MФs and epithelial cells (6, 48), while in human keratinocytes, it induces the activation of both the NF-κB and p38 pathways (49). ATRA can also inhibit the PI3K/AKT signaling in human fibroblasts (50), and induces IRF-1 expression and its nuclear localization in human MФs (37, 51). Importantly, many of these pathways overlap with those that are also affected by NOD1/NOD2 activation, rising the possibility of cross-talk mechanisms (Figure 4B).
Conclusions
In summary, our results show that NOD1/NOD2-induced cytokine secretion by macrophages (MФ) is cell type dependent and it is highly modified by the nuclear receptor agonist ATRA. The implications of these observations are far reaching. NOD1 and NOD2, as sensors of microorganisms and cellular stress, have been identified as potential therapeutic targets in a range of autoimmune and autoinflammatory disorders (52) and in diabetes, asthma, atherosclerosis, various forms of CNS diseases and even cancer (29). In line with this, while it is well established that vitamin A and retinoic acid play important homeostatic and regulatory roles in multiple tissues, based on our data, MФs activated by NOD1/NOD2-dependent or independent stimuli are likely to be profoundly modulated by ATRA. Major immune organs also require a steady dietary intake of vitamin A to maintain their functions and vitamin A deficiency has been associated with dysbiosis, frequent infectious diseases and reduced immune response to vaccines, although the underlying mechanisms have remained unclear to date. Our data shed light on this issue, by showing the potent and diverse modulatory role of ATRA on NOD1/NOD2-induced cytokine secretion. Based on previous reports, we suggest that this modulation is mediated, in part, via interrelated signaling pathways and altered metabolic status of the cell. Hence, we speculate that ATRA may function as a general regulator of cytokine secretion induced by various PRRs. While understanding the precise mechanism of ATRA action on NOD1/NOD2-induced mechanisms in different tissues and MФ populations requires further studies, our results highlight the therapeutic relevance of these pathways in a number of inflammatory conditions, which represent an increasing burden to society.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving humans were approved by National Blood Service, (Budapest); and Regional and Institutional Ethics Committee/REC/IEC) of the Clinical Centre (University of Debrecen). The studies were conducted in accordance with the local legislation and institutional requirements. The human samples used in this study were acquired from a by-product of routine care or industry. Written informed consent for participation was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and institutional requirements.
Author contributions
HA: Investigation, Software, Writing – original draft, Data curation, Formal analysis, Methodology, Validation, Visualization. AA: Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Writing – original draft. EB: Writing – review & editing, Formal analysis, Methodology, Validation, Visualization. SB: Writing – review & editing, Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Software, Supervision, Writing – original draft, Data curation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Hungarian National Scientific Research Fund (NKFIH K-131844 and NKFIH K-147109 to SB), HA is supported by the Richter Talentum Foundation. HA and AA hold a Stipendium Hungaricum Scholarship from the Government of Hungary.
Acknowledgments
We thank Fanni Balogh for the valuable technical assistance. We are grateful to Dr. Ádám Dénes for the critical comments of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1609763/full#supplementary-material
Supplementary Figure 1 | Secretion of IFNβ following NOD1 and NOD2 activation. M-MФ and GM-MФ were treated with C14-Tri-LAN-Gly (NOD1 agonist, 500 ng/ml) and L-18 MDP (NOD2 agonist, 100 ng/ml) for the indicated time points. Control cells were treated with the same amount of vehicle as the activated cells. Cytokine secretion was measured from the supernatant using ELISA. Time kinetics of IFNβ secretion following (A) NOD1 activation and (B) NOD2 activation. Cells were pretreated with ATRA (1µM) for 4 hours and then stimulated with (C) C14-Tri-LAN-Gly or (D) L-18 MDP for 6 and 24 hours. Results were obtained from at least four healthy donors. All results are shown as means ± SD. (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, # p < 0.05, ## p < 0.01, ### p < 0.001, #### p < 0.0001; n.d. – not detected).
References
1. de M Oliveira L, Teixeira FME, and Sato MN. Impact of retinoic acid on immune cells and inflammatory diseases. Mediators Inflammation. (2018) 2018:1–17. doi: 10.1155/2018/3067126
2. Piazza A, Carlone R, and Spencer GE. Non-canonical retinoid signaling in neural development, regeneration and synaptic function. Front Mol Neurosci. (2024) 17:1371135. doi: 10.3389/fnmol.2024.1371135
3. Erkelens MN and Mebius RE. Retinoic acid and immune homeostasis: a balancing act. Trends Immunol. (2017) 38:168–80. doi: 10.1016/j.it.2016.12.006
4. Hall JA, Grainger JR, Spencer SP, and Belkaid Y. The role of retinoic acid in tolerance and immunity. Immunity. (2011) 35:13–22. doi: 10.1016/j.immuni.2011.07.002
5. Pinos I, Yu J, Pilli N, Kane MA, and Amengual J. Functional characterization of interleukin 4 and retinoic acid signaling crosstalk during alternative macrophage activation. Biochim Biophys Acta (BBA)-Molecular Cell Biol Lipids. (2023) 1868:159291. doi: 10.1016/j.bbalip.2023.159291
6. Li S, Lei Y, Lei J, and Li H. All−trans retinoic acid promotes macrophage phagocytosis and decreases inflammation via inhibiting CD14/TLR4 in acute lung injury. Mol Med Rep. (2021) 24:1–11. doi: 10.3892/mmr.2021.12508
7. Guan F, Wang R, Yi Z, Luo P, Liu W, Xie Y, et al. Tissue macrophages: origin, heterogenity, biological functions, diseases and therapeutic targets. Signal Transduct Target Ther. (2025) 10:1–67. doi: 10.1038/s41392-025-02124-y
8. Li D and Wu M. Pattern recognition receptors in health and diseases. Signal Transduct Target Ther. (2021) 6:291. doi: 10.1038/s41392-021-00687-0
9. Pei G, Zyla J, He L, Moura-Alves P, Steinle H, Saikali P, et al. Cellular stress promotes NOD1/2-dependent inflammation via the endogenous metabolite sphingosine-1-phosphate. EMBO J. (2021) 40:e106272. doi: 10.15252/embj.2020106272
10. Kuss-Duerkop SK and Keestra-Gounder AM. NOD1 and NOD2 activation by diverse stimuli: a possible role for sensing pathogen-induced endoplasmic reticulum stress. Infect Immun. (2020) 88:10–1128. doi: 10.1128/IAI.00898-19
11. Zangara MT, Johnston I, Johnson EE, and McDonald C. Mediators of metabolism: An unconventional role for NOD1 and NOD2. Int J Mol Sci. (2021) 22:1156. doi: 10.3390/ijms22031156
12. Moreira LO and Zamboni DS. NOD1 and NOD2 signaling in infection and inflammation. Front Immunol. (2012) 3:328. doi: 10.3389/fimmu.2012.00328
13. Al Nabhani Z, Dietrich G, Hugot J-P, and Barreau F. Nod2: The intestinal gate keeper. PloS Pathog. (2017) 13:e1006177. doi: 10.1371/journal.ppat.1006177
14. Fernández-García V, González-Ramos S, Martín-Sanz P, García-del Portillo F, Laparra JM, and Boscá L. NOD1 in the interplay between microbiota and gastrointestinal immune adaptations. Pharmacol Res. (2021) 171:105775. doi: 10.1016/j.phrs.2021.105775
15. Wang X, Allen C, and Ballow M. Retinoic acid enhances the production of IL-10 while reducing the synthesis of IL-12 and TNF-α from LPS-stimulated monocytes/macrophages. J Clin Immunol. (2007) 27:193–200. doi: 10.1007/s10875-006-9068-5
16. Kang BY, Chung SW, Kim SH, Kang SN, Choe YK, and Kim TS. Retinoid-mediated inhibition of interleukin-12 production in mouse macrophages suppresses Th1 cytokine profile in CD4+ T cells. Br J Pharmacol. (2000) 130:581–6. doi: 10.1038/sj.bjp.0703345
17. Matikainen S, Serkkola E, and Hurme M. Retinoic acid enhances IL-1 beta expression in myeloid leukemia cells and in human monocytes. J Immunol (Baltimore Md 1950). (1991) 147:162–7. doi: 10.4049/jimmunol.147.1.162
18. Hayashi S, Hashimoto S, Kitamura N, Hanazawa S, and Horie T. Retinoic acid regulates differentially the expression of IL-1β and IL-1 receptor antagonist (IL-1ra) in PMA-activated human monocytes. Biochem Biophys Res Commun. (1996) 224:574–8. doi: 10.1006/bbrc.1996.1066
19. Alatshan A, Kovács GE, Aladdin A, Czimmerer Z, Tar K, and Benkő S. All-trans retinoic acid enhances both the signaling for priming and the glycolysis for activation of NLRP3 inflammasome in human macrophage. Cells. (2020) 9:1–21. doi: 10.3390/cells9071591
20. Kumaran Satyanarayanan S, El Kebir D, Soboh S, Butenko S, Sekheri M, Saadi J, et al. IFN-β is a macrophage-derived effector cytokine facilitating the resolution of bacterial inflammation. Nat Commun. (2019) 10:3471. doi: 10.1038/s41467-019-10903-9
21. Liu C, Chu D, Kalantar-Zadeh K, George J, Young HA, and Liu G. Cytokines: from clinical significance to quantification. Adv Sci. (2021) 8:2004433. doi: 10.1002/advs.202004433
22. Takada H and Uehara A. Enhancement of TLR-mediated innate immune responses by peptidoglycans through NOD signaling. Curr Pharm Des. (2006) 12:4163–72. doi: 10.2174/138161206778743510
23. da Silva Correia J, Miranda Y, Leonard N, Hsu J, and Ulevitch RJ. Regulation of Nod1-mediated signaling pathways. Cell Death Differ. (2007) 14:830–9. doi: 10.1038/sj.cdd.4402070
24. Omaru N, Watanabe T, Kamata K, Minaga K, and Kudo M. Activation of NOD1 and NOD2 in the development of liver injury and cancer. Front Immunol. (2022) 13:1004439. doi: 10.3389/fimmu.2022.1004439
25. Locati M, Curtale G, and Mantovani A. Diversity, mechanisms, and significance of macrophage plasticity. Annu Rev Pathol Mech Dis. (2020) 15:123–47. doi: 10.1146/annurev-pathmechdis-012418-012718
26. Ross EA, Devitt A, and Johnson JR. Macrophages: the good, the bad, and the gluttony. Front Immunol. (2021) 12:708186. doi: 10.3389/fimmu.2021.708186
27. Hamilton TA, Zhao C, Pavicic Jr. PG, and Datta S. Myeloid colony-stimulating factors as regulators of macrophage polarization. Front Immunol. (2014) 5:554. doi: 10.3389/fimmu.2014.00554
28. Itoh CY, Gunnarsson C, Babunovic GH, Nibasumba A, Akilimali NA, Wadsworth III MH, et al. GM-CSF differentiation of human monocytes stabilizes macrophage state via oxidative signaling. bioRxiv. (2020), 2009–20. doi: 10.1101/2020.09.29.318352
29. Trindade BC and Chen GY. NOD1 and NOD2 in inflammatory and infectious diseases. Immunol Rev. (2020) 297:139–61. doi: 10.1111/imr.12902
30. Peng XY, Wang KL, Li L, Li B, Wu XY, Zhang ZW, et al. Transcription of NOD1 and NOD2 and their interaction with CARD9 and RIPK2 in IFN signaling in a perciform fish, the Chinese perch, Siniperca chuatsi. Front Immunol. (2024) 15:1374368. doi: 10.3389/fimmu.2024.1374368
31. Budai MM, Tőzsér J, and Benkő S. Different dynamics of NLRP3 inflammasome-mediated IL-1β production in GM-CSF–and M-CSF–differentiated human macrophages. J Leukoc Biol. (2017) 101:1335–47. doi: 10.1189/jlb.3A0716-300RR
32. Kovács EG, Alatshan A, Budai MM, Czimmerer Z, Bíró E, and Benkő S. Caffeine has different immunomodulatory effect on the cytokine expression and NLRP3 inflammasome function in various human macrophage subpopulations. Nutrients. (2021) 13:2409. doi: 10.3390/nu13072409
33. Teijeira A, Garasa S, Ochoa MC, Villalba M, Olivera I, Cirella A, et al. IL8, neutrophils, and NETs in a collusion against cancer immunity and immunotherapy. Clin Cancer Res. (2021) 27:2383–93. doi: 10.1158/1078-0432.CCR-20-1319
34. Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, et al. Neutrophil extracellular traps kill bacteria. Science (80-). (2004) 303:1532–5. doi: 10.1126/science.1092385
35. Díaz-Pino R, Rice GI, San Felipe D, Pepanashvili T, Kasher PR, Briggs TA, et al. Type I interferon regulates interleukin-1beta and IL-18 production and secretion in human macrophages. Life Sci Alliance. (2024) 7:1–14. doi: 10.26508/lsa.202302399
36. Sun Y, Ma J, Li D, Li P, Zhou X, Li Y, et al. Interleukin-10 inhibits interleukin-1β production and inflammasome activation of microglia in epileptic seizures. J Neuroinflamm. (2019) 16:1–13. doi: 10.1186/s12974-019-1452-1
37. Matikainen S, Ronni T, Hurme M, Pine R, and Julkunen I. Retinoic acid activates interferon regulatory factor-1 gene expression in myeloid cells. Blood (1996) 88(1):114–23. doi: 10.1182/blood.V88.1.114.bloodjournal881114
38. Karwacz K, Miraldi ER, Pokrovskii M, Madi A, Yosef N, Wortman I, et al. Critical role of IRF1 and BATF in forming chromatin landscape during type 1 regulatory cell differentiation. Nat Immunol. (2017) 18:412–21. doi: 10.1038/ni.3683
39. Lavudi K, Nuguri SM, Olverson Z, Dhanabalan AK, Patnaik S, and Kokkanti RR. Targeting the retinoic acid signaling pathway as a modern precision therapy against cancers. Front Cell Dev Biol. (2023) 11:1254612. doi: 10.3389/fcell.2023.1254612
40. Alatshan A and Benkő S. Nuclear receptors as multiple regulators of NLRP3 inflammasome function. Front Immunol. (2021) 12:1–25. doi: 10.3389/fimmu.2021.630569
41. Liu PT, Krutzik SR, Kim J, and Modlin RL. Cutting edge: all-trans retinoic acid down-regulates TLR2 expression and function. J Immunol. (2005) 174:2467–70. doi: 10.4049/jimmunol.174.5.2467
42. Cho H-Y, Choi E-K, Lee S-W, Kim K-H, Park S-J, Lee CK, et al. All-trans retinoic acid induces TLR-5 expression and cell differentiation and promotes flagellin-mediated cell functions in human THP-1 cells. Immunol Lett. (2011) 136:97–107. doi: 10.1016/j.imlet.2011.01.001
43. Li Y, Gao Y, Cui T, Yang T, Liu L, Li T, et al. Retinoic acid facilitates toll-like receptor 4 expression to improve intestinal barrier function through retinoic acid receptor beta. Cell Physiol Biochem. (2017) 42:1390–406. doi: 10.1159/000479203
44. Pu J, Chen D, Tian G, He J, Huang Z, Zheng P, et al. All-trans retinoic acid attenuates transmissible gastroenteritis virus-induced inflammation in IPEC-J2 Cells via suppressing the RLRs/NF-κB signaling pathway. Front Immunol. (2022) 13:734171. doi: 10.3389/fimmu.2022.734171
45. Liu T-X, Zhang J-W, Tao J, Zhang R-B, Zhang Q-H, Zhao C-J, et al. Gene expression networks underlying retinoic acid–induced differentiation of acute promyelocytic leukemia cells. Blood J Am Soc Hematol. (2000) 96:1496–504. doi: 10.1182/blood.V96.4.1496
46. Priyanka SH, Thushara AJ, Rauf AA, and Indira M. Alcohol induced NLRP3 inflammasome activation in the brain of rats is attenuated by ATRA supplementation. Brain Behav Immunity-Health. (2020) 2:1–5. doi: 10.1016/j.bbih.2019.100024
47. Huo L, Cui D, Yang X, Gao Z, Trier K, and Zeng J. All-trans retinoic acid modulates mitogen-activated protein kinase pathway activation in human scleral fibroblasts through retinoic acid receptor beta. Mol Vis. (2013) 19:1795.
48. Chen C, Smith AD, Cheung L, Pham Q, Urban JF Jr., and Dawson HD. Potentiation of IL-4 signaling by retinoic acid in intestinal epithelial cells and macrophages—mechanisms and targets. Front Immunol. (2020) 11:605. doi: 10.3389/fimmu.2020.00605
49. Dai X, Yamasaki K, Shirakata Y, Sayama K, and Hashimoto K. All-trans-retinoic acid induces interleukin-8 via the nuclear factor-κB and p38 mitogen-activated protein kinase pathways in normal human keratinocytes. J Invest Dermatol. (2004) 123:1078–85. doi: 10.1111/j.0022-202X.2004.23503.x
50. Liang L, Wang X, Zheng Y, and Liu Y. All-trans-retinoic acid modulates TGF-β-induced apoptosis, proliferation, migration and extracellular matrix synthesis of conjunctival fibroblasts by inhibiting PI3K/AKT signaling. Mol Med Rep. (2019) 20:2929–35. doi: 10.3892/mmr.2019.10507
51. Luo XM and Ross AC. Retinoic acid exerts dual regulatory actions on the expression and nuclear localization of interferon regulatory factor-1. Exp Biol Med. (2006) 231:619–31. doi: 10.1177/153537020623100517
Keywords: retinoic acid, NOD1, NOD2, NOD-like receptor, vitamin A, inflammation, macrophage, cytokine
Citation: Ahmad H, Alatshan A, Bíró E and Benkő S (2025) Retinoic acid differently modulates NOD1/NOD2-mediated inflammatory responses in human macrophage subsets. Front. Immunol. 16:1609763. doi: 10.3389/fimmu.2025.1609763
Received: 10 April 2025; Accepted: 16 June 2025;
Published: 01 July 2025.
Edited by:
Ewa Teresa Marcinkowska, University of Wrocław, PolandReviewed by:
Pandurangan Ramaraj, A.T. Still University, United StatesYordan Sbirkov, Plovdiv Medical University, Bulgaria
Copyright © 2025 Ahmad, Alatshan, Bíró and Benkő. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Szilvia Benkő, YmVua29zekBtZWQudW5pZGViLmh1
†These authors share first authorship