 Alicia Simón-Rueda1,2,3,4
Alicia Simón-Rueda1,2,3,4 Clara Sánchez-Menéndez1,2
Clara Sánchez-Menéndez1,2 Guiomar Casado1,4
Guiomar Casado1,4 Daniel Fuertes5
Daniel Fuertes5 María Aránzazu Murciano-Antón2,6Elena Mateos1,3Susana Domínguez-Mateos2,7
María Aránzazu Murciano-Antón2,6Elena Mateos1,3Susana Domínguez-Mateos2,7 Francisco Pozo8,9Javier García-Pérez3,10
Francisco Pozo8,9Javier García-Pérez3,10 Mayte Pérez-Olmeda3,11
Mayte Pérez-Olmeda3,11 Miguel Cervero12
Miguel Cervero12 Marta Massanella3,13,14
Marta Massanella3,13,14 Gemma Moncunill3,14,15,16
Gemma Moncunill3,14,15,16 Montserrat Torres1,3,14*
Montserrat Torres1,3,14* Mayte Coiras1,3,14*
Mayte Coiras1,3,14*- 1Immunopathology and Viral Reservoir Unit, National Center of Microbiology, Instituto de Salud Carlos III, Madrid, Spain
- 2PhD Program in Biomedical Sciences and Public Health, Universidad Nacional de Educación a Distancia (UNED), Madrid, Spain
- 3Biomedical Research Center Network in Infectious Diseases (CIBERINFEC), Instituto de Salud Carlos III, Madrid, Spain
- 4Infectious Diseases Unit, Internal Medicine Service, Hospital Clínico San Carlos, Madrid, Spain
- 5School of Telecommunications Engineering, Universidad Politécnica de Madrid, Madrid, Spain
- 6Family Medicine Service, Primary Healthcare Center Doctor Pedro Laín Entralgo, Madrid, Spain
- 7Family Medicine Service, Primary Healthcare Center Arroyomolinos, Madrid, Spain
- 8Respiratory Viruses Unit, National Center of Microbiology, Instituto de Salud Carlos III, Madrid, Spain
- 9Biomedical Research Center Network in Epidemiology and Public Health (CIBERESP), Instituto de Salud Carlos III, Madrid, Spain
- 10AIDS Immunopathology Unit, National Center of Microbiology, Instituto de Salud Carlos III, Madrid, Spain
- 11Serology Service, National Center of Microbiology, Instituto de Salud Carlos III, Madrid, Spain
- 12School of Medicine, Universidad Alfonso X El Sabio, Madrid, Spain
- 13IrsiCaixa AIDS Research Institute, Germans Trias i Pujol Research Institute (IGTP), Can Ruti Campus, Barcelona, Spain
- 14REICOP, Spanish Network for Persistent COVID Research, Madrid, Spain
- 15Instituto de Salud Global de Barcelona (ISGLOBAL), Barcelona, Spain
- 16Facultat de Medicina i Ciències de la Salut, Universitat de Barcelona (UB), Barcelona, Spain
Introduction: Long COVID (LC) affects approximately 10% of individuals post-SARS-CoV-2 infection, with symptoms persisting beyond 12 weeks. The underlying mechanisms remain unclear, and current models often focus on pre-existing comorbidities.
Methods: This cohort study aimed to identify robust biomarkers and clarify LC pathogenesis through a comprehensive analysis performed in 32 LC individuals 26 months post-infection compared with 35 fully recovered individuals recruited between March and July 2022. Blood and fecal samples were collected, and multiple parameters associated with immune dysfunction, endothelial damage, bacterial translocation, and coagulation alterations, alongside signs of viral persistence and sociodemographic and clinical features, were analyzed.
Results: Although viral RNA was undetected on blood or stool, elevated plasma IgG against the nucleocapsid may indicate frequent reinfections, greater infection severity, or delayed immune normalization. Increased levels of prothrombin, thrombin, fibrinogen, sEPCR, and CRP pointed to persistent endothelial dysfunction and coagulation imbalance. Lower levels of the bactericidal protein REG3A suggest potential disruptions in mucosal immune response. We found no major differences in traditional comorbidities, highlighting that LC may stem from distinct pathogenic mechanisms beyond pre-existing conditions. Importantly, our study revealed impaired humoral immunity and identified an association between vaccine heterogeneity and increased LC risk, emphasizing the relevance of consistent vaccination strategies. A Random Forest model using the measured biomarkers achieved 100% accuracy in classifying LC individuals, reinforcing their diagnostic potential.
Discussion: These findings support a multifactorial model of LC involving immune dysregulation and persistent endothelial damage that led to coagulation abnormalities and a pro-thrombotic profile, supporting that LC is more closely related to a sustained, uncontrolled inflammatory response rather than immunodeficiency, and underscoring the value of multidimensional biomarker profiling for guiding clinical management and prevention strategies.
1 Introduction
The COVID-19 pandemic, caused by SARS-CoV-2, has profoundly impacted global health systems and societies. Since the World Health Organization (WHO) declared it a pandemic in 2020, over 770 million confirmed infections and 7 million deaths have been reported, though the actual numbers are likely higher (1).
Effective vaccination campaigns reduced viral spread and mitigated severe infections, but Long COVID (LC) remains a significant consequence. Also known as Post-acute COVID-19 Syndrome (PACS) or Post-COVID-19 Condition (PCC), LC is characterized by symptoms persisting beyond 12 weeks after infection, affecting 10–20% of individuals (2). Symptoms vary widely, including fatigue, breathlessness, myalgia, palpitations, gastrointestinal issues, cognitive impairment, and mental health disorders such as anxiety and depression (3). It is unclear whether these mental health effects result from underlying physiological conditions or impaired quality of life (4). Risk factors include female sex, older age, and comorbidities like obesity and diabetes (5). There is conflicting evidence on whether vaccination affects the risk of LC. While some reports suggest an association between LC and vaccination (Post-COVID-19 Vaccination Syndrome) (6), most evidence indicates that vaccination, whether pre- or post-infection, provides protection against LC (7).
Over 200 symptoms across multiple organ systems complicate the establishment of a diagnostic framework for this heterogeneous syndrome (8). Several hypotheses attempt to explain LC’s underlying mechanisms. One leading theory is viral persistence, where SARS-CoV-2 remains in hidden reservoirs, driving chronic inflammation and immune dysregulation (9). LC shares similarities with autoimmune conditions like fibromyalgia or chronic fatigue syndrome, which involve persistent inflammation and exaggerated immune responses (10). Evidence of viral RNA and proteins has been found in various tissues, including the respiratory and cardiovascular systems, kidneys, gastrointestinal tract, muscles, brain, and lymph nodes (2). Prolonged viral shedding in stool samples has been detected months after diagnosis, even with negative nasopharyngeal RT-PCR results (11), suggesting that viral persistence in the gut could contribute to LC-related gastrointestinal symptoms (12).
Intestinal epithelium damage and compromised barrier integrity may enable bacterial translocation, altering the gut microbiome and triggering sustained immune activation (13). Persistent endothelial dysfunction has also been observed in LC patients, evidenced by elevated markers of endothelial damage and activation (14). This dysfunction may stem from endothelial inflammation during acute infection, as SARS-CoV-2 infects endothelial cells via ACE2 (15). Chronic endothelial damage could underlie the systemic inflammation in LC, characterized by elevated levels of proinflammatory markers such as interleukin-6 (IL-6), C-reactive protein (CRP), and tumor necrosis factor alpha (TNFα) (16, 17). Since endothelial cells are crucial for coagulation regulation through anticoagulant factor production, including thrombin inhibitors and elements of the protein C pathway such as thrombomodulin and endothelial protein C receptor (EPCR) (18, 19), this persistent endothelial inflammation and damage may disrupt this regulation, contributing to coagulopathy in LC. In fact, while acute COVID-19 is known to cause disseminated intravascular coagulation (20), LC is associated with coagulopathies such as thrombotic endotheliitis, hyperactivated platelets, and fibrinaloid microclots, which may exacerbate the syndrome’s symptoms (17). However, although dysbiosis, persistent endothelial damage, altered coagulability, autoimmunity, or even the reactivation of latent herpesvirus such as Epstein-Barr virus (EBV) could contribute to long-lasting inflammation in people with LC (21, 22), these findings remain controversial, and their role in LC pathogenesis is not fully understood. The search for biomarkers linked to these mechanisms is critical for diagnosis, identifying therapeutic targets, and developing curative treatments.
In this cohort study, we evaluated multiple parameters associated with immune dysfunction, endothelial damage, bacterial translocation, and coagulation alterations, alongside signs of viral persistence and sociodemographic and clinical features, in a cohort of individuals with LC. Our objective was to gain insights into the mechanisms driving LC symptomatology and to identify biomarkers that may aid in its diagnosis. This knowledge may also inform the development of novel therapeutic strategies and help refine clinical guidelines for managing LC patients.
2 Materials and methods
2.1 Study subjects
Sixty-seven individuals with symptomatic COVID-19 during the first pandemic waves in Spain were recruited at the Primary Healthcare Center (PHC) Doctor Pedro Laín Entralgo (Alcorcón, Madrid, Spain) and PHC Arroyomolinos (Arroyomolinos, Madrid, Spain) in March-July 2022. Inclusion criteria required participants to be over 18 years old and have a confirmed diagnosis of acute mild COVID-19, verified either by a positive RT-qPCR test for SARS-CoV-2 in a nasopharyngeal swab or by the presence of virus-specific IgM in plasma. Participants were categorized into two groups based on the duration of symptoms’ resolution. Those who experienced at least eight clinical signs and symptoms consistent with LC more than 12 weeks after their positive diagnosis, as defined by the National Institute for Health and Care Excellence (NICE) guidelines (23), were included in the LC cohort (n=32). Conversely, individuals who fully recovered within the first four weeks post-diagnosis were included in the Recovered cohort (n=35). All participants were recruited with the collaboration of the non-profit Spanish Association of Patients with Long-COVID (Long COVID-ACTS). Cases and controls were matched on age. In order to participate, all individuals completed a comprehensive, structured questionnaire covering clinical, cognitive, and systemic symptoms. This was used to confirm eligibility for recruitment and to ensure consistent documentation of self-reported manifestations across the cohort. All participants were followed for a total of 2 years to record breakthrough infections from primary infection to blood sample collection, confirmed by SARS-CoV-2 antigens test. Sample size was calculated to achieve 80% power with an alpha level of 0.05.
2.2 Ethical statement
All participants provided informed written consent prior to their inclusion in the study. The study protocol (CEI PI 72_2022) was developed in accordance with the Declaration of Helsinki and received previous review and approval from the Ethics Committee of the Instituto de Salud Carlos III (IRB IORG0006384) and the Primary Care Management Commission of the Comunidad de Madrid (Spain). Participant confidentiality and anonymity were safeguarded in compliance with current Spanish and European Data Protection regulations.
2.3 Blood and fecal samples
Blood samples from all participants were collected using BD Vacutainer tubes containing EDTA K2 (Becton Dickinson, Franklin Lakes, NJ). These samples were promptly processed to isolate peripheral blood mononuclear cells (PBMCs) and plasma via Ficoll-Hypaque density gradient centrifugation (Corning Inc, Corning, NY). Fecal samples were collected in specialized containers with preservative buffer provided by Palex Medical (Barcelona, Spain). All samples were cryopreserved until analysis. Due to sample limitations, not all determinations were conducted for every sample.
2.4 Detection of SARS-CoV-2 RNA by RT-qPCR
The presence of SARS-CoV-2 RNA in plasma and feces was assessed using an RT-qPCR assay targeting the envelope (E) and nucleocapsid (N) genes, following the guidelines outlined in the WHO Interim Guidance for the diagnostic testing of SARS-CoV-2 (24). Viral RNA was extracted from plasma samples using the QIAamp MinElute Virus Spin Kit and from fecal samples using the QIAamp Viral RNA Kit (Qiagen Iberia, Madrid, Spain). Samples were classified as positive if the quantification cycle (Cq) value was below 42.
2.5 SARS-CoV-2 serology
IgG antibodies against subunit 1 (S1) from Spike (S) protein of SARS-CoV-2 were analyzed in plasma using Euroimmun Anti-SARS-CoV-2 ELISA Assay (Euroimmun, Lübeck, Germany), according to manufacturer’s instructions. Semi-quantitative results were analyzed by calculating the ratio of optical density (OD) of each sample over the calibrator. Samples were considered positive when this ratio was ≥ 0.8. In addition, IgG against the receptor binding domain (RBD), S1, subunit 2 (S2), and N proteins of SARS-CoV-2 were analyzed by chemiluminescence immunoassay (CLIA) using BioPlex 2200 SARS-CoV-2 IgG Panel (BioRad, Hercules, CA), according to the manufacturer’s instructions. Samples were considered positive as follows: S1 ≥22 binding antibody units (BAU)/mL; S2 ≥10 U/mL; RBD ≥13 BAU/mL; and N ≥24 BAU/mL.
2.6 SARS-CoV-2 pseudovirus neutralization assay
One single-cycle, pseudotyped SARS-CoV-2 virus (pNL4-3Δenv_SARS-CoV-2-SΔ19(G614)_Ren) was synthesized as previously described (25). Co-transfection with vector pcDNA-VSV-G was used as a control of specificity. Briefly, neutralization activity of heat-decomplemented plasma was measured by pre-incubation of pNL4-3Δenv_SARS-CoV-2-SΔ19(G614)_Ren pseudovirus (10ng p24 Gag per well) with serial dilutions of plasma (1/32 to 1/8192) for 1 hour at 37°C (25). This mixture was then added to a monolayer of Vero E6 cells and incubated for 48 hours. Vero E6 cell line (ECACC 85020206) was kindly provided by Dr Antonio Alcamí (CBM Severo Ochoa, Madrid, Spain) and it was cultured in DMEM supplemented with 10% fetal calf serum (FCS), 100U/ml penicillin/streptomycin, and 2mM L-Glutamin (Lonza, Basil, Switzerland). After incubation, cells were lysed and viral infectivity was assessed by measuring Renilla luciferase activity (Renilla Luciferase Assay, Promega, Madison, WI) in a 96-well plate luminometer Centro XS3 LB 960 with MikroWin 2010 software (Berthold Technologies, Baden-Württemberg, Germany). Titers of neutralizing IgG were calculated as 50% neutralizing dose (NT50) using non-linear regression analysis in GraphPad Prism Software v10.2.1. (GraphPad, Inc., San Diego, CA). NT50 was defined as the highest dilution of plasma that caused 50% reduction of luciferase activity, in comparison with control without plasma.
2.7 Herpesvirus serology
The levels of IgG against cytomegalovirus (CMV), Varicella Zoster virus (VZV), and Herpes Simplex virus 1/2 (HSV-1/2) were analyzed in plasma on the LIASON automated platform (Diasorin) with LIASON CMV IgG II, LIASON VZV IgG, and LIASON HSV-1/2 IgG CLIA assays, respectively (DiaSorin, Saluggia, Italy). The levels of IgG against EBV viral capsid antigens (VCA) were tested using LIAISON VCA IgG CLIA assay (DiaSorin). CMV IgG and VCA IgG antibody titers were calculated and expressed as U/mL; VZV IgG titers were expressed as mU/ml; and HSV-1/2 IgG titers were expressed as a ratio. Samples were considered positive or negative according to the cutoffs established by the manufacturer.
2.8 Analysis of herpesvirus reactivation by qPCR
Total DNA was extracted from plasma samples using QIAamp MinElute Virus Spin Kit (Qiagen Iberia). To detect the presence of DNA from EBV and CMV, used as markers of viral reactivation, DNA was amplified by qPCR with StepOnePlus Real-Time PCR System (Applied Biosystems; Thermo Fisher, Waltham, MA), using EBV R-GENE and CMV R-GENE kits (bioMérieux. Lyon, France), respectively. Data was analyzed with StepOne v2.3 Software (Thermo Fisher) and samples were considered positive when showing a calculated value of CT (Threshold cycle).
2.9 Detection of bacterial translocation, endothelial damage, and coagulation markers in plasma by ELISA
Plasma levels of parameters associated with bacterial translocation such as regenerating islet derived 3 alpha (REG3A) and fatty acid binding protein 2 (FABP2) were measured using Abcam ELISA Kits (Cambridge, UK), while occludin, lipopolysaccharide-binding protein (LBP), and lipopolysaccharides (LPS) were measured using Cusabio ELISA kits (Wuhan, China). Markers for coagulation parameters and endothelial damage such as thrombin, thrombomodulin, tissue-type plasminogen activator (tPA), intercellular adhesion molecule 1 (ICAM-1), and soluble EPCR (sEPCR) were measured using Abcam ELISA Kits (Cambridge, UK), while activated protein C (APC) was quantified using Abyntek Biopharma ELISA kit (Bizkaia, Spain). Data was acquired in a Tecan Sunrise Basic Microplate reader (Tecan, Männedorf, Switzerland).
2.10 Analysis of cytokines and coagulation factors in plasma by Luminex assay
Customized Human Magnetic Luminex Assay kit (Thermo Fisher) was used for the simultaneous detection of the following cytokines and chemokines in plasma: interleukin (IL)-1β, IL-6, IL-8, IL-10, IL-12p70, monocyte chemoattractant protein (MCP)-4, monokine induced by interferon-gamma (MIG), macrophage inflammatory protein (MIP)-3α, and myeloid progenitor inhibitor factor (MPIF). ProcartaPlex™ Human Coagulation Panel (Thermo Fisher) was used for the detection of prothrombin, Factor XI, and Factor XIII. ProcartaPlex™ Human Simplex Kits were also used for the detection of D-dimer, fibrinogen, and CRP. Data acquisition was performed with Luminex 200 Instrument System (Thermo Fisher).
2.11 Random forest algorithm
A Random Forest algorithm (26) was applied to evaluate the accuracy of the qualitative and quantitative variables that showed significant differences (p<0.05) between the LC and Recovered groups, aiming to identify the most important features associated with LC. To avoid bias in selecting the training, testing, and validation sets, a nested 5-fold cross-validation procedure was performed for each competing algorithm, as previously described (27, 28). The relative importance for each feature in classifying participants was determined using the Gini Variable Importance Measure (VIM) method (29).
2.12 Statistical analysis
Statistical analysis was conducted using GraphPad Prism v10.2.1 (GraphPad Software Inc.) and STATA 14.2 (StataCorp LLC, College Station, TX). Quantitative variables were reported as the median with interquartile range (IQR), and qualitative variables were expressed as absolute or relative frequencies. The normality of the samples was assessed using the Shapiro-Wilk test. Significance between the two cohorts was determined using the unpaired t-test or nonparametric Mann-Whitney test, based on the normality of the data. Qualitative data were compared using Fisher’s exact test or the chi-square test, as appropriate. Linear and logistic regressions were used to estimate the odds ratio (OR) and 95% confidence interval (CI) for associations between the levels of quantitative variables and the development of LC, compared to healthy donors. Logistic regression was also applied to estimate the OR and CI for qualitative variables. A p-value of < 0.05 was considered statistically significant for all comparisons.
3 Results
3.1 Sociodemographic and clinical characteristics of the participants
This single-center, cohort study recruited 67 participants. Thirty-two participants were diagnosed with LC (LC cohort). All participants in LC cohort experienced symptomatic acute COVID-19 following initial SARS-CoV-2 infection, and 3 (9.4%) required hospitalization for a median of 5 days (IQR 5-5) during the acute phase of infection. Thirty-five individuals who fully recovered from symptomatic acute SARS-CoV-2 infection were included as controls in the Recovered cohort. 57% participants with LC were diagnosed by RT-qPCR on nasopharyngeal swabs, while 43% were diagnosed by the presence of virus-specific IgM in plasma. In Recovered participants, 66% were diagnosed by PCR and 34% by serology. Most participants were women (97% in LC cohort, 71% in Recovered cohort) and the median age was 49 (IQR 45-52) and 50 years (IQR 38-52), respectively. The median time from COVID-19 diagnosis to sample collection was 26 months (IQR 23-27) for the LC cohort and 24 months (IQR 24-24) for the Recovered cohort. Key sociodemographic and clinical characteristics are in Table 1, with further details in Supplementary Table 1 and Supplementary Table 2.
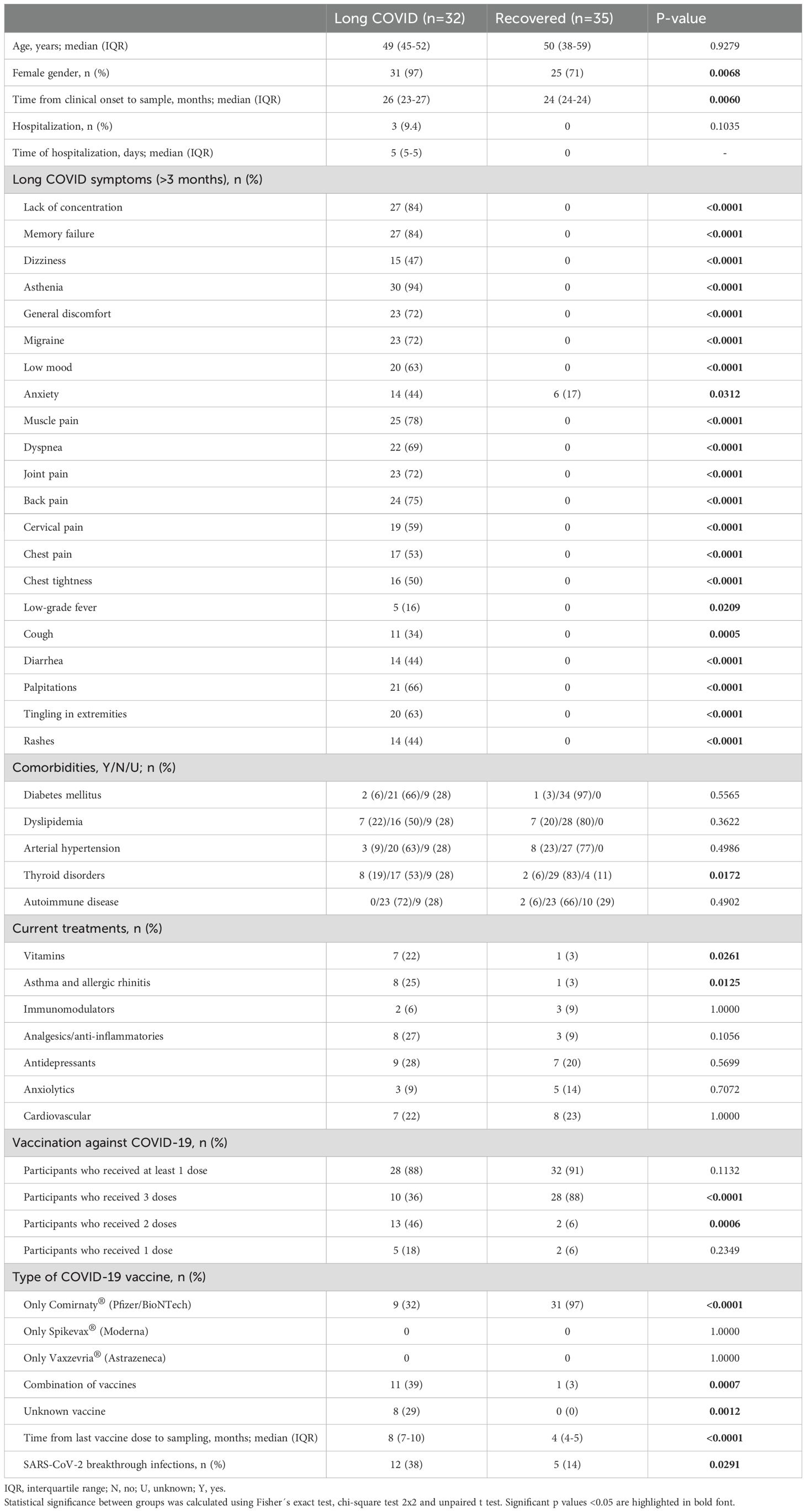
Table 1. Clinical and sociodemographic characteristics of all participants in this study.
At the time of sampling, the most frequent symptoms reported by participants in the LC cohort were fatigue (94%), memory loss (84%), and difficulty concentrating (84%). Other common symptoms included back pain (75%), joint pain (72%), migraine (72%), general discomfort (72%), dyspnea (69%), palpitations (66%), tingling in the extremities (63%), low mood (63%), and neck pain (59%). LC participants also reported chest pain (53%), chest tightness (50%), dizziness (47%), skin rashes (44%), diarrhea (44%), and anxiety (44%). Less common symptoms include persistent cough (34%) and low-grade fever (16%). People with LC reported that these symptoms persisted for more than 3 months after COVID-19 diagnosis. In contrast, participants in the Recovered cohort did not report any of these symptoms for more than 3 months, except for anxiety, which was reported by 6 (17%) participants.
Comorbidities such as diabetes, dyslipidemia, and hypertension were recorded in both groups, with no significant differences observed. However, thyroid disorders were more common among LC participants compared to Recovered individuals (19% versus 6%; p=0.0172). Participants from both cohorts were receiving a variety of treatments, including immunomodulators, analgesics/anti-inflammatory drugs, antidepressants, anxiolytics, and/or cardiovascular treatments at sampling, with no significant differences between groups. A higher proportion of LC participants were on treatment for asthma and allergic rhinitis compared to the Recovered group (25% versus 3%; p=0.0096). Similarly, more LC participants were taking vitamin supplements than those in the Recovered group (22% versus 3%; p<0.0211).
Most participants in both LC and Recovered groups (88% and 91%, respectively) had received at least one dose of an authorized COVID-19 vaccine, with most having received two or three doses (94% and 82%, respectively). Among Recovered participants, the most common vaccine was Comirnaty® (Pfizer/BioNTech) (97%; p<0.0001), while LC participants received a combination of different vaccines (39%; p=0.0007). The LC cohort reported a higher incidence of breakthrough SARS-CoV-2 infections compared to the Recovered cohort (38% versus 14%; p=0.0291). Time from last vaccine dose to sampling was 8 (IQR 7-10) and 4 (IQR 4-5) months in LC and Recovered participants, respectively (p<0.0001). All breakthrough infections were mild, and no participant required hospitalization.
3.2 Detection of SARS-CoV-2 RNA in plasma and feces
The presence of RNA from SARS-CoV-2 was not detected in the plasma or feces of any participant in the study (data not shown).
3.3 Changes in the levels of antibodies against SARS-CoV-2 in people with LC
We observed lower levels of IgG against the S1 protein in participants from the LC cohort compared to the Recovered cohort (-1.2-fold; p=0.0153) (Figure 1A). The neutralizing capacity of antibodies against SARS-CoV-2 was reduced 1.8-fold (p=0.0067) in LC cohort (Figure 1B). Analysis of the proportion of participants with detectable IgG against different viral proteins revealed that 15.6% of participants in the LC cohort had undetectable levels of IgG against the S2 protein, compared to the Recovered cohort (p=0.0151) (Figure 1C). In contrast, the number of participants in the LC cohort with detectable levels of IgG against the N protein was 1.7-fold higher than in the Recovered cohort (p=0.0381). No significant differences were found in the proportion of participants with detectable IgG against S1 and RBD.
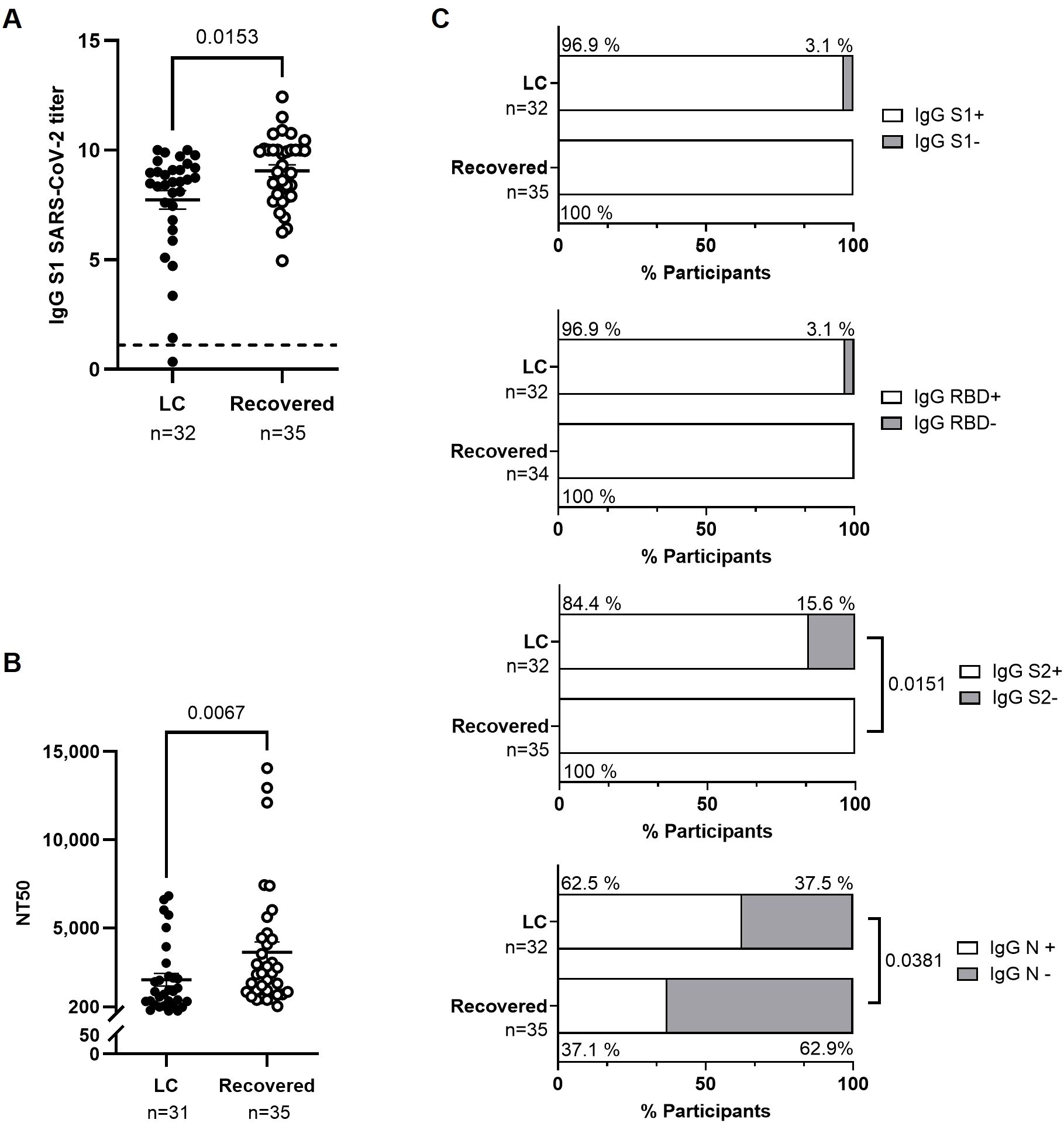
Figure 1. Characterization of the humoral response against SARS-CoV-2 in individuals with LC compared to Recovered. (A) Analysis by ELISA of the levels of IgG against SARS-CoV-2 spike protein subunit 1 (S1) in plasma from individuals with LC or Recovered. The horizontal line at 1.1 represents the positivity threshold. (B) Titers of neutralizing anti-S1 IgG in plasma calculated as NT50 from individuals with LC or Recovered. Each dot corresponds to one sample (LC, closed circles; Recovered, open circles) and vertical lines represent mean ± standard error of the mean (SEM). Statistical significance was obtained using non-parametric Mann-Whitney test. (C) Analysis by CLIA of the levels of IgG against S1, RBD, S2, and N proteins from SARS-CoV-2 in plasma of participants with LC or Recovered. Bars depict the percentage of participants considered producers (open bar) or non-producers (grey bar) of each IgG. Statistical significance was calculated using the Chi-square test.
3.4 Levels of plasma proteins related to intestinal barrier integrity and bacterial translocation
Plasma levels of the bactericidal intestinal protein REG3A were 1.2-fold lower (p=0.0153) in the LC cohort compared to the Recovered cohort (Figure 2A). No significant differences were observed between groups in the levels of plasma proteins associated with intestinal injury, such as FABP2 and occludin (Figure 2B) or markers of bacterial translocation such as LPS and LBP (Figure 2C).
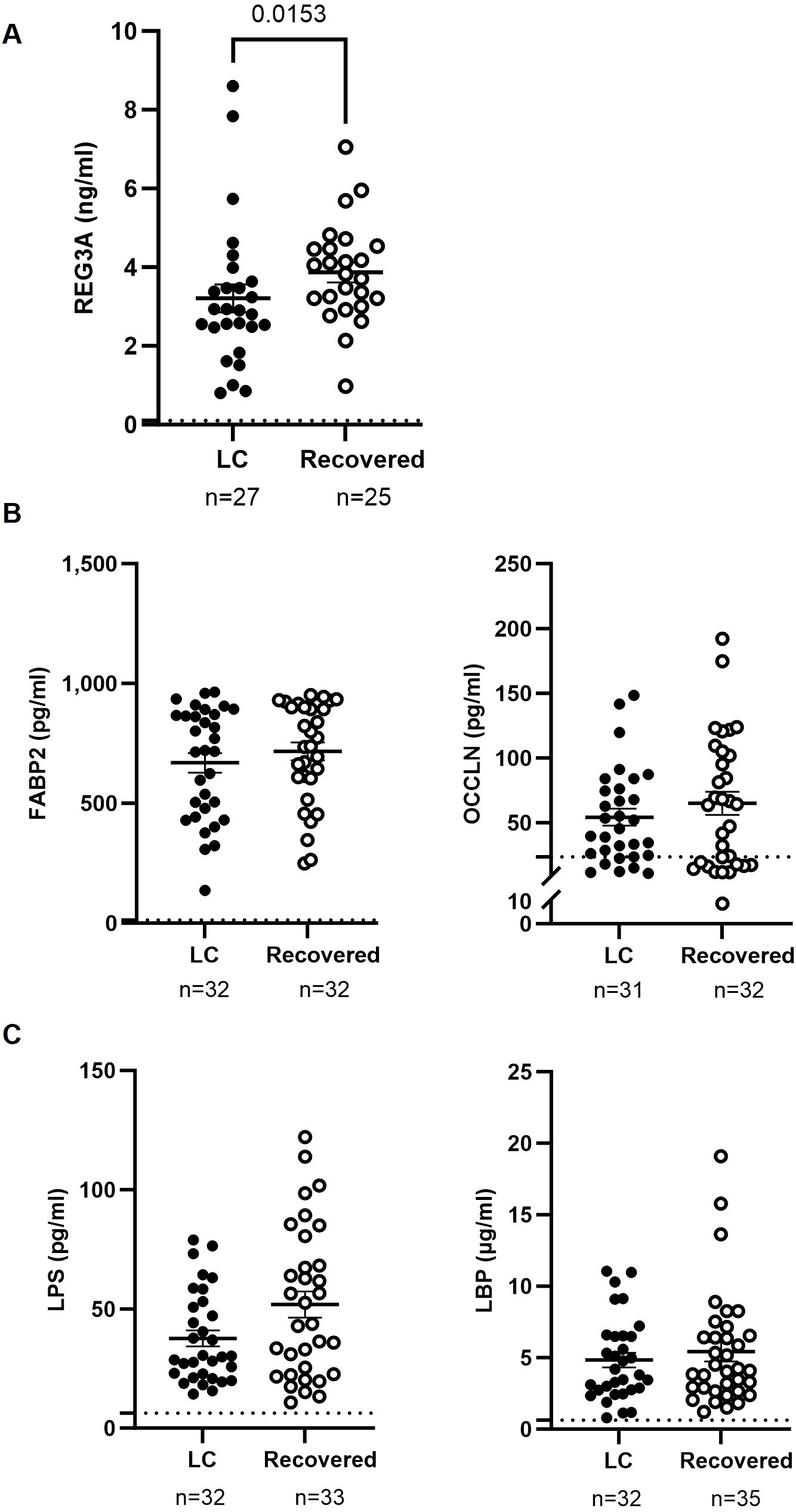
Figure 2. Analysis of factors related to intestinal injury and bacterial translocation in plasma of individuals with LC and Recovered. Plasma levels of the gut bactericidal protein REG3A (A), markers of intestinal barrier integrity FABP2 and OCCLN (B), and markers associated with bacterial translocation LPS and LBP (C), were analyzed by ELISA. Each dot corresponds to one sample (LC, closed circles; Recovered, open circles) and vertical lines represent mean±SEM. Horizontal dashed lines mark the limit of detection. Statistical significance was calculated using non-parametric Mann-Whitney test.
3.5 Plasma levels of cytokines and chemokines
Similar levels of pro-inflammatory (IL-1β, IL-12, IL-6) (Figure 3A) and anti-inflammatory (IL-10) cytokines (Figure 3B) were observed in plasma of participants from both groups. However, participants in the LC cohort had 2.7- and 1.3-fold lower levels of the chemokines MIG/CXCL9 and MPIF/CCL23, respectively, compared to the Recovered cohort (p=0.0069 and p=0.0423), while levels of the chemokines MCP-4/CCL13, MIP-3α/CCL20, and IL-8/CXCL8 were similar between both groups (Figure 3C).
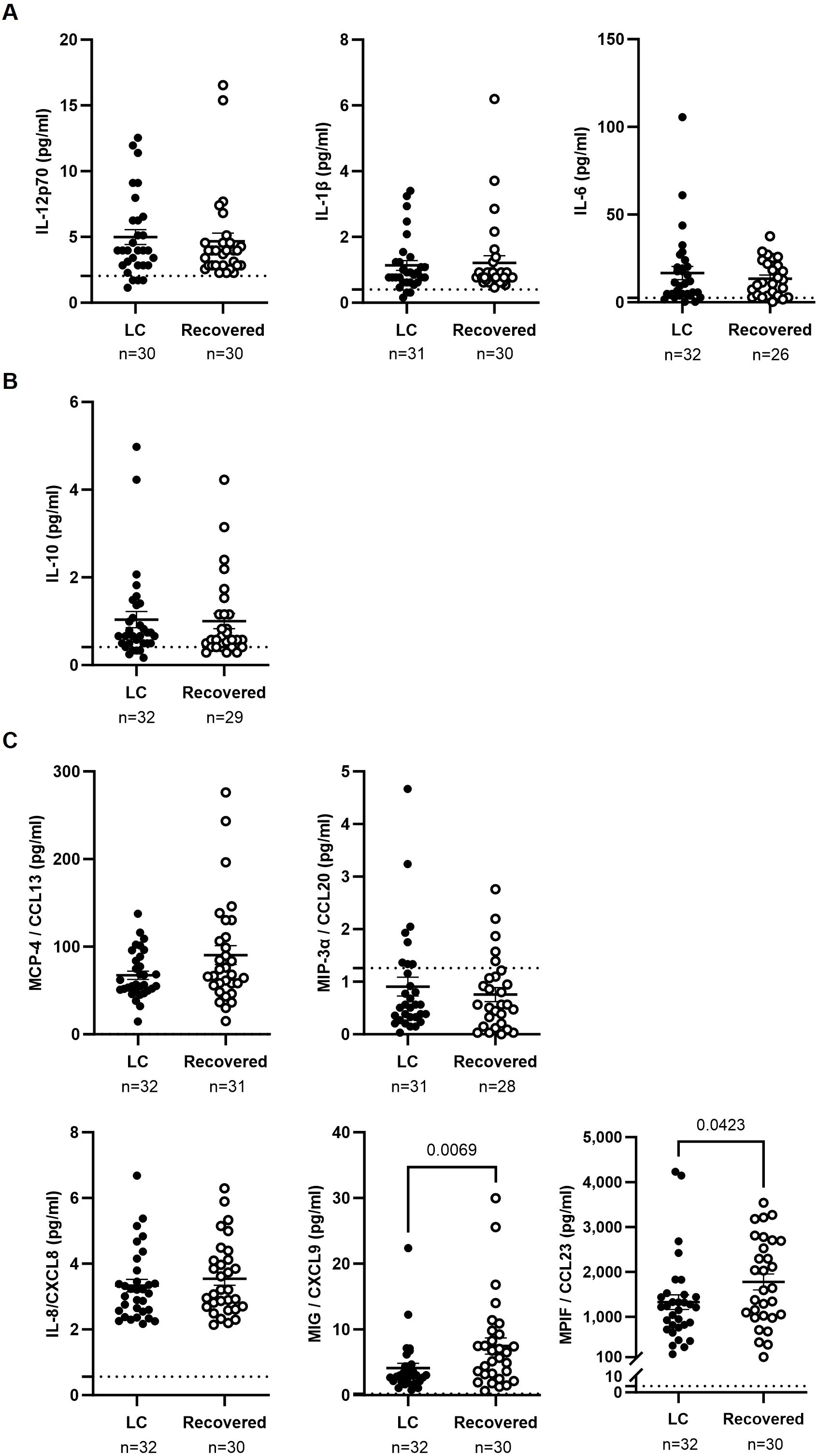
Figure 3. Analysis of cytokines in plasma of individuals with LC and Recovered. Plasma levels of proinflammatory cytokines IL-12p70, IL-1β, and IL-6 (A), anti-inflammatory cytokine IL-10 (B), and chemokines MCP-4/CCL1, MIP-3α/CCL20, IL-8/CXCL8, MIG/CXCL9, and MPIF/CCL23 (C) were analyzed by Luminex. Each dot corresponds to one sample (LC, closed circles; Recovered, open circles) and vertical lines represent mean±SEM. Horizontal dashed lines mark the limit of detection. Statistical significance was calculated using non-parametric Mann-Whitney test.
3.6 Serology and reactivation of herpesvirus
No significant differences were observed in plasma IgG titers against CMV, VZV, HSV-1/2, and EBV, nor in the total number of individuals who tested positive for each IgG between the two cohorts (Figure 4A). However, EBV DNA was detectable in the plasma of both the Recovered and LC cohorts (74.1% versus 40.8%, respectively; p=0.0133) (Figure 4B). Two individuals from each cohort (7.4%) had detectable CMV DNA in plasma. Reactivation of EBV or CMV did not result in any clinical consequences.
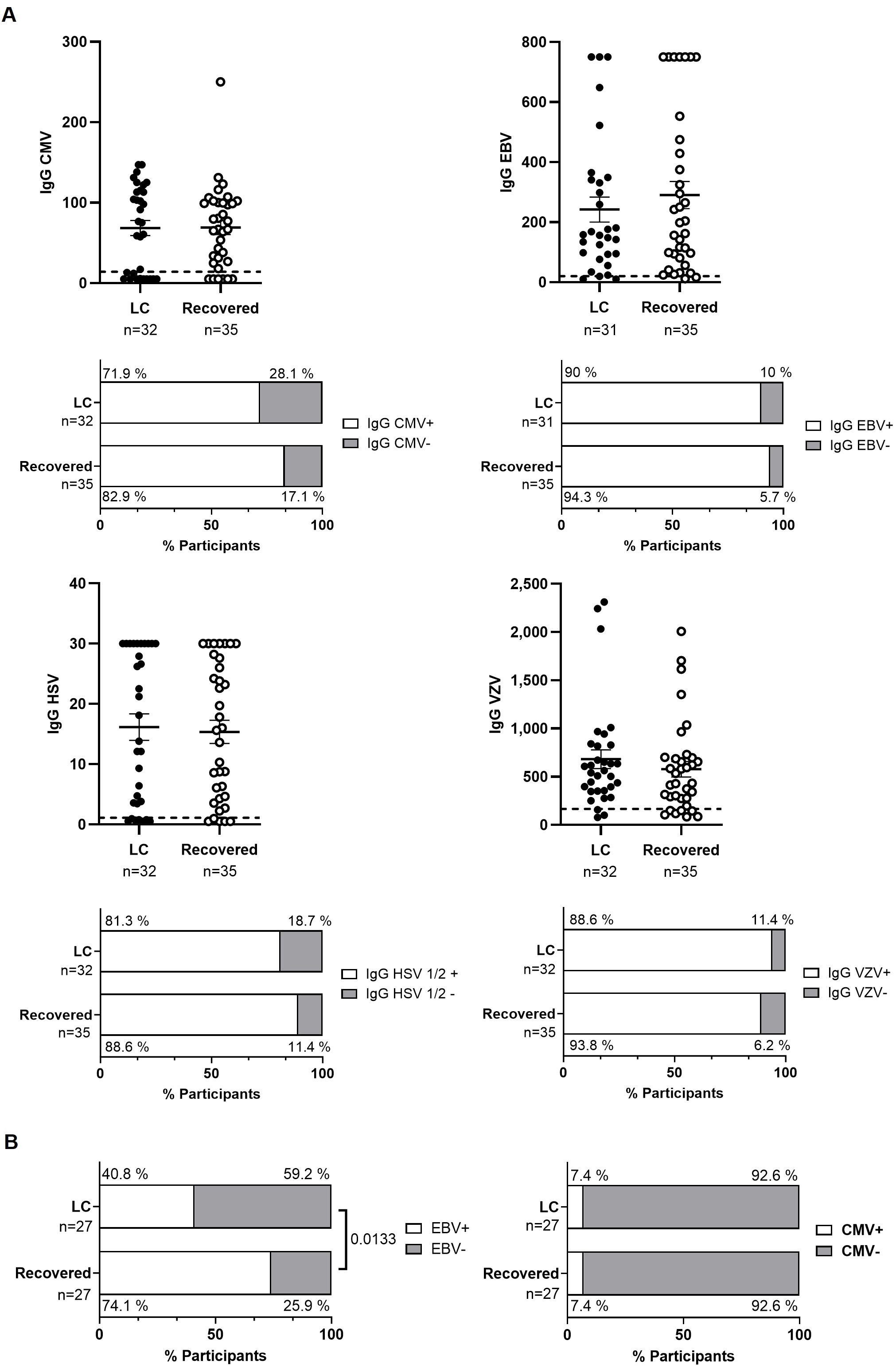
Figure 4. Analysis of herpesvirus reactivation in plasma from individuals with LC or Recovered. (A) Dot graphs show levels of IgG against CMV, EBV, HSV-1/2 and VZV analyzed by CLIA. Each dot represents data from one individual (LC, closed circles; Recovered, open circles) and vertical lines represent mean±SEM. Horizontal dashed line marks the limit of detection. Statistical significance was calculated using non-parametric Mann-Whitney test. Bar graphs show the count of individuals producers (open bar) or non-producers (grey bars) of antibodies against herpesvirus. Statistical significance was calculated using the Chi-square test. (B) Detection of provirus reactivation of EBV and CMV was analyzed by qPCR in plasma from participants with LC or Recovered. Graph bars depict the percentage of individuals in which these herpesviruses were reactivated (open bars) or not reactivated (grey bars). Statistical significance was calculated using the Chi-square test.
3.7 Evidence of endothelial dysfunction and altered coagulation in LC cohort
The expression of several proteins related to endothelial dysfunction and coagulation was analyzed in plasma using thrombin as a central factor (Figure 5A). Individuals from the LC cohort showed significantly higher levels of prothrombin (1.3-fold; p=0.0038) and thrombin (1.2-fold; p=0.0121) compared to the Recovered cohort (Figure 5B). Since thrombin mediates activation of Factors XI and XIII (30), we also assessed the levels of these factors, but no significant changes were observed between the cohorts (Figure 5C). Thrombin cleaves fibrinogen to form fibrin monomers, which then polymerize into a fibrin clot (31). As a result, fibrinogen levels were 1.3-fold lower (p=0.0270), while CRP levels were 3.1-fold higher (p=0.0051) in LC participants, alterations that have been previously linked to a higher risk of venous thromboembolism (32) (Figure 5D). No significant differences were observed in tPA levels, which mediate plasminogen conversion during fibrinolysis (33), or in D-dimer levels, a degradation product of cross-linked fibrin (34) (Figure 5E).
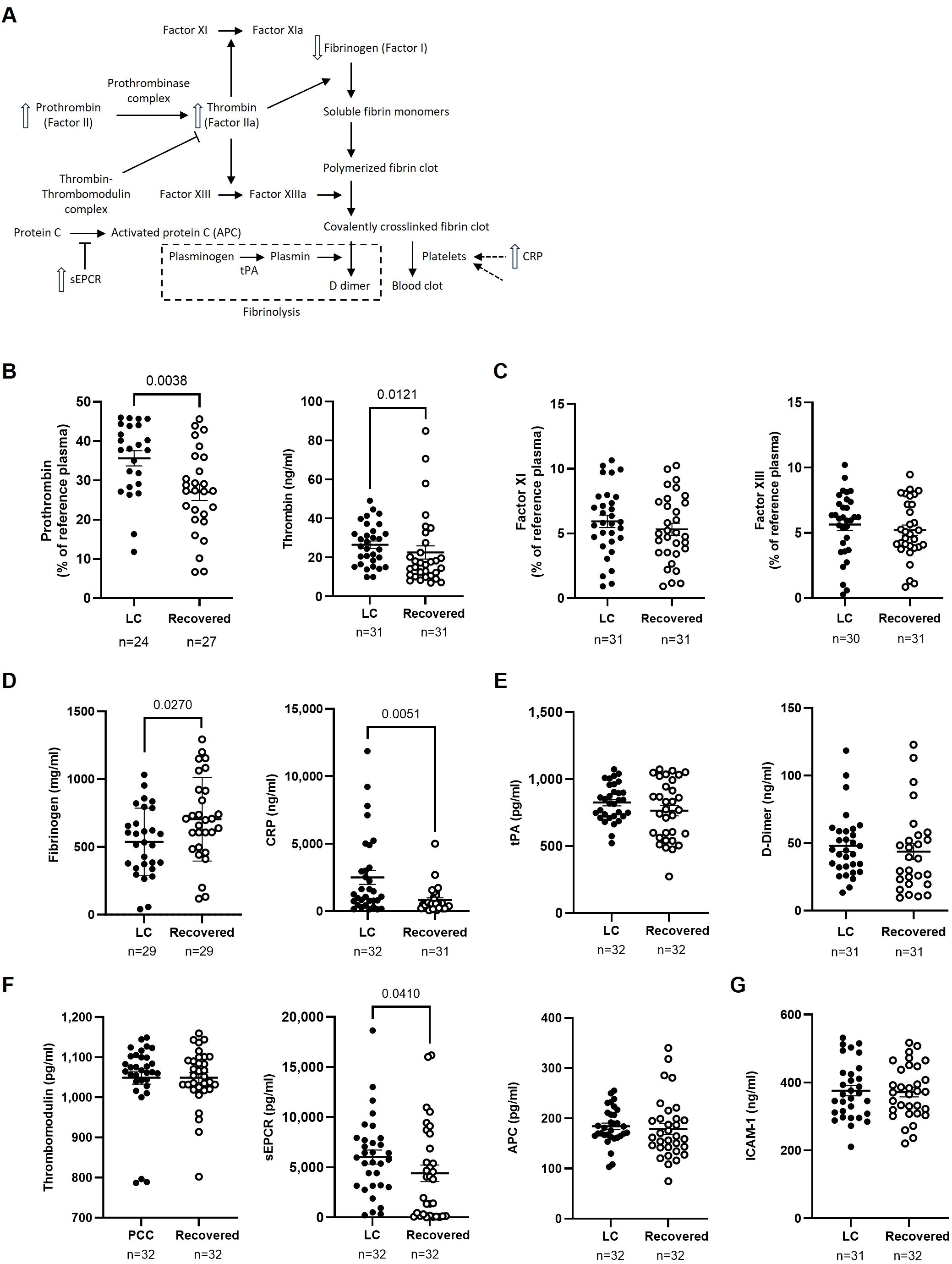
Figure 5. Analysis of markers of endothelial damage, thrombotic risk, and coagulation factors in the plasma of participants with LC or Recovered. (A) Schematic diagram of the interactions between coagulation proteins using thrombin as a central factor. Up and down arrows show those parameters that were increased or decreased, respectively, in plasma of individuals with LC compared to Recovered. (B) Analysis of plasma levels of prothrombin and thrombin as key proteins in the blood clotting process. (C) Analysis of plasma concentration of Factor XI, which is essential for the amplification phase of coagulation, and Factor XIII that is a fibrin-stabilizing factor. (D) Analysis of plasma concentration of acute-phase proteins fibrinogen and CRP whose levels increase during inflammation and acute responses. (E) Analysis of plasma levels of fibrinolysis markers tPA (tissue plasminogen activator) and D-dimer, which are associated with the breakdown of clots. (F) Analysis of plasma levels of anticoagulant regulators thrombomodulin, sEPCR (soluble Endothelial Protein C Receptor), and APC (Activated Protein C), which are involved in the regulation of anticoagulation and maintaining hemostatic balance. (G) Analysis of plasma concentration of endothelial activator ICAM-1 that can influence coagulation processes in the context of inflammation and thrombosis. Each dot represents data from one individual (LC, closed circles; Recovered, open circles) and vertical lines represent mean±SEM. Statistical significance was obtained using non-parametric Mann-Whitney test or unpaired t-test, as appropriate.
Since the thrombin-thrombomodulin complex activates PC to downregulate coagulation and inflammation (35), we also analyzed plasma levels of thrombomodulin and APC and found no differences between the two groups. However, sEPCR, which inhibits APC activity and has been reported as a marker associated with higher thrombotic risk (36), was 1.4-fold higher (p=0.0410) in the LC cohort compared to the Recovered cohort (Figure 5F).
Lastly, there were no differences in the plasma levels of ICAM-1 (Figure 5G), an endothelial injury marker (37), between the two groups.
3.8 Association of qualitative and quantitative variables with the persistence of COVID-19
The association of the qualitative variables in the development of LC was analyzed using binary logistic regression analysis (OR) (Table 2A). This analysis revealed that female gender (OR 12.400; 95% CI 1.485 to 103.520; p=0.020), receiving doses of a combination of different vaccines rather than the same type (OR 5.775; 95% CI 1.991 to 16.748; p=0.001), high levels of antibodies against the SARS-CoV-2 N protein (OR 2.821; 95% CI 1.047 to 7.599; p=0.040), and the development of asthma and/or allergic rhinitis (OR 10.909; 95% CI 1.270 to 93.692; p=0.029) were positively correlated with the occurrence of LC. Additionally, LC participants were more likely to take vitamin supplements compared to the Recovered cohort (OR 9.130; 95% CI 1.048 to 79.532; p=0.045).
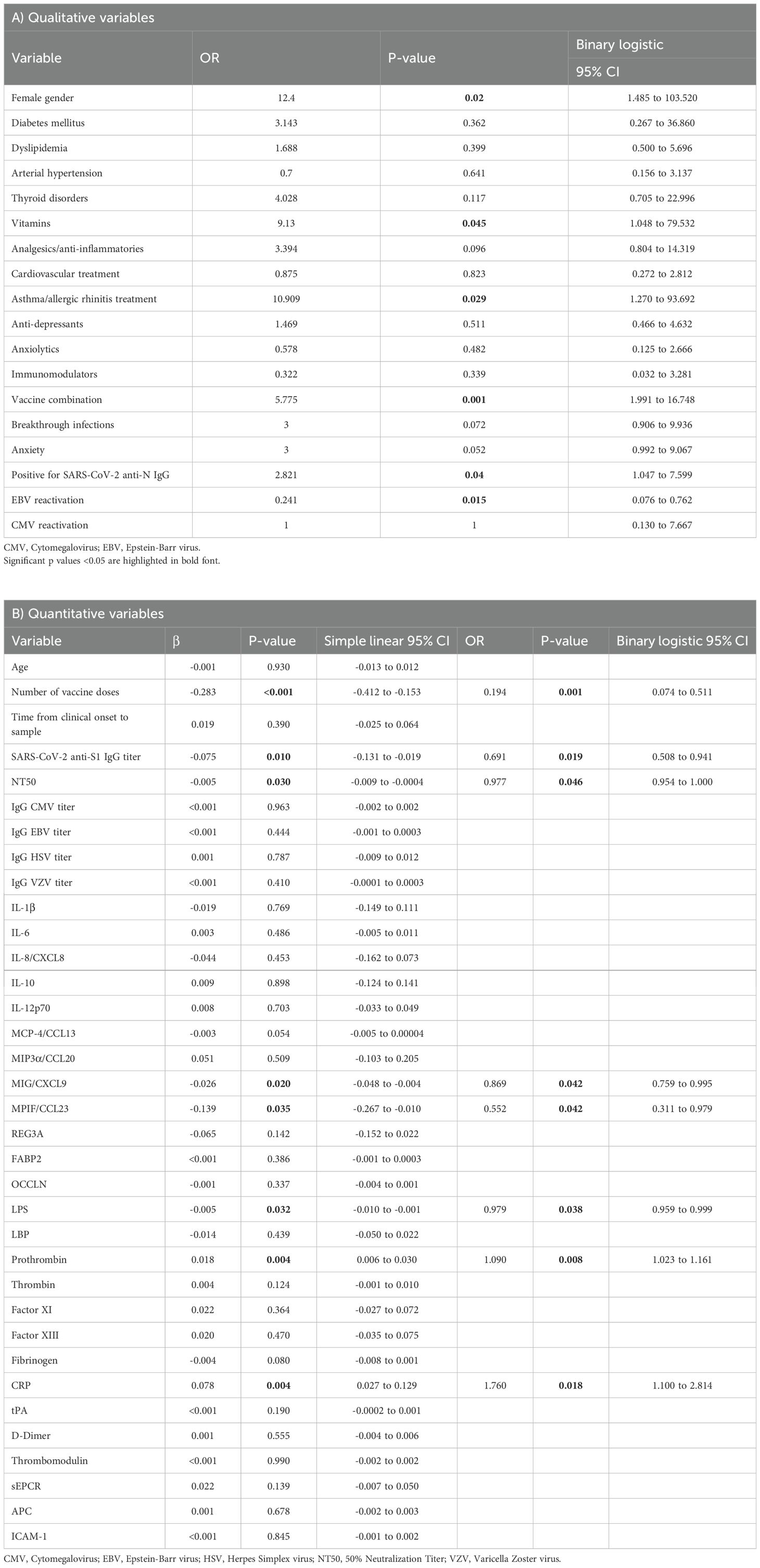
Table 2. Association between qualitative (A) and quantitative variables (B) with the development of LC was assessed using simple linear regression analysis and subsequent binary logistic regression analysis to calculate odds ratio (OR) for quantitative variables and using logistic regression analysis for qualitative variables.
The association of the quantitative variables in the development of LC was assessed through linear regression followed by binary logistic regression analysis (Table 2B). Linear regression revealed a trend toward an association between the number of COVID-19 vaccine doses, the neutralizing capacity of IgG against SARS-CoV-2, the titers of IgG against SARS-CoV-2 S1 protein, and plasma levels of CXCL9, CCL23, prothrombin, and CRP with the development and/or persistence of LC, compared to healthy donors. This trend was confirmed by binary logistic regression, indicating that low number of vaccine doses (OR 0.194; 95% CI 0.074 to 0.511; p=0.001), IgG levels against SARS-CoV-2 S1 protein (OR 0.691; 95% CI 0.508 to 0.941; p=0.019), neutralizing capacity (NT50) of IgG against SARS-CoV-2 (OR 0.977; 95% CI 0.954 to 1.000; p=0.046), MIG/CXCL9 levels (OR 0.869; 95% CI 0.759 to 0.995; p=0.042), MPIF/CCL23 levels (OR 0.552; 95% CI 0.311 to 0.979; p=0.042) were correlated with the occurrence of LC. Conversely, high prothrombin levels (OR 1.090; 95% CI 1.023 to 1.161; p=0.008) and CRP levels (OR 1.760; 95% CI 1.100 to 2.814; p=0.018) were correlated with the occurrence of LC.
3.9 Application of random forest for the evaluation of the importance of the assayed variables for LC
An accuracy of 97.03% ± 3.64% was achieved across the 5 iterations of the outer loop in the nested K-fold cross-validation for each competing algorithm (Figure 6A). As a result, all 35 participants (100%) in the LC cohort were correctly classified, while 30 of the 32 participants (93.75%) in the Recovered group were accurately assigned to their respective group (Figure 6B). The Gini VIM method identified several clinical variables as crucial for classification into the LC group, including memory failure and lack of concentration, muscle and back pain, paresthesia in extremities, asthenia, and malaise (Figure 6C). Key quantitative variables contributing to LC group classification included the neutralizing capacity of IgGs against SARS-CoV-2, the levels of CRP, thrombin, prothrombin, and REG3A, as well as the number of vaccine doses and having received a combination of vaccine types rather than a single vaccine type. Conversely, variables with lower importance for LC group assignment included gender, low-grade fever, thyroid disorders, and having had breakthrough infections.
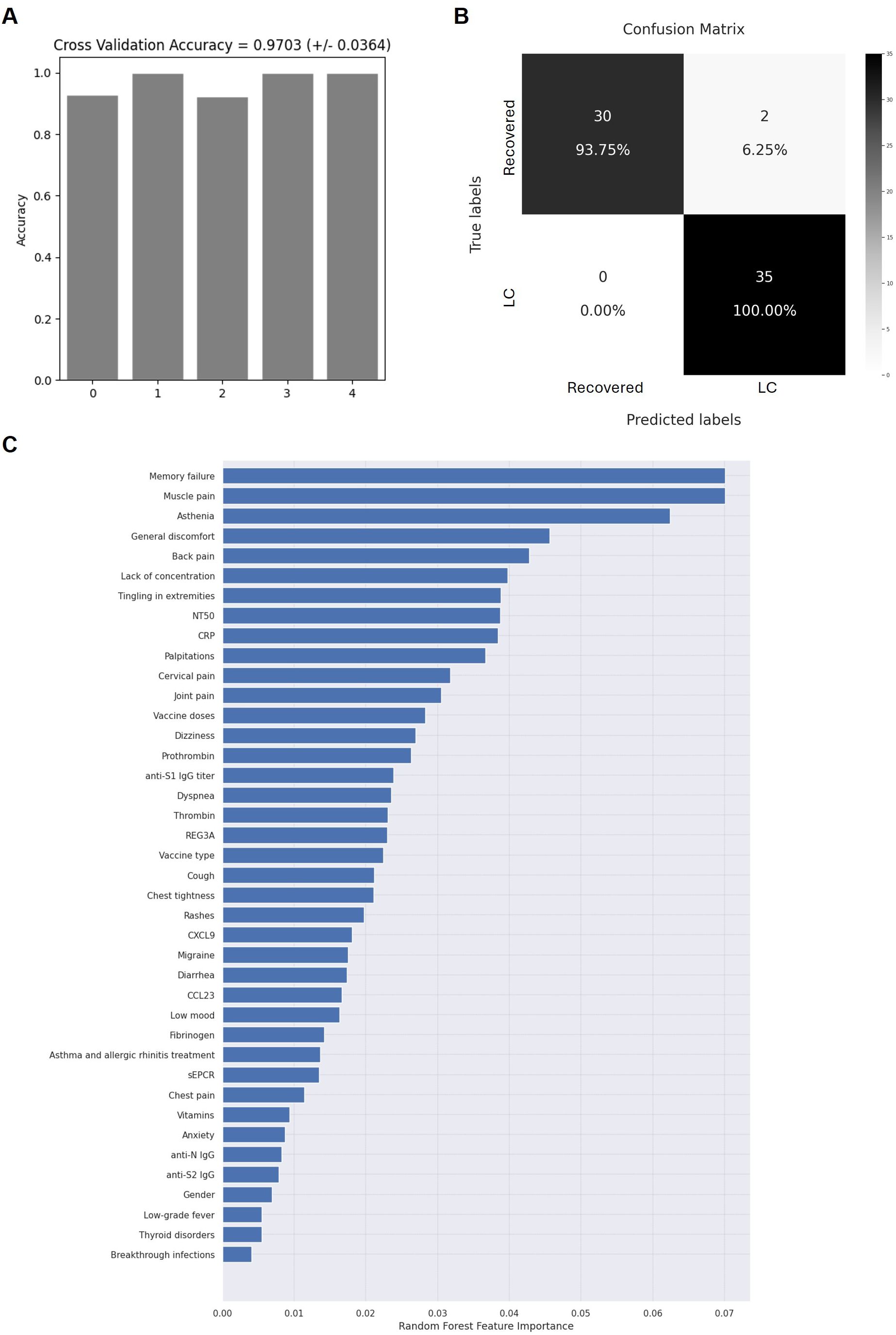
Figure 6. Application of random forest algorithm and Gini VIM method to evaluate the accuracy and importance of the selected biomarkers. Accuracy for the 5 iterations of the outer loop of the nested K-fold cross validation (A) and confusion matrix confronting the conditions calculated by the algorithm and the true chronicity-related conditions of individuals with LC or Recovered (B, C) Relative importance of the selected parameters for the categorization of individuals with LC or Recovered, according to Gini VIM method.
4 Discussion
Long COVID (LC) has emerged as a major sequela of the COVID-19 pandemic, presenting challenges for diagnosis and treatment. Despite extensive research, the precise causes remain unclear, and among the more than 200 reported symptoms, debate persists regarding which are most critical for diagnosis and prognosis. Identifying key biomarkers is essential to understanding LC mechanisms and guiding effective treatments.
Key contributors to LC include host-related and external factors such as SARS-CoV-2 persistence, herpesvirus reactivation, and dysbiosis due to endothelial damage. Demographic factors like middle age and female gender have been consistently linked to LC susceptibility (3, 5, 38). In our cohort, 97% of LC participants were women (median age: 49 years). Most (90.6%) did not require hospitalization during acute infection, confirming that LC is not limited to severe cases. While comorbidities like diabetes, dyslipidemia, and hypertension have been associated with LC (5), we found no significant differences in their prevalence between LC and Recovered individuals, except for thyroid disorders (19% vs. 6%; p=0.0172). This finding aligns with previous reports, but the lack of baseline data raises questions about whether thyroid dysfunction results from an autoimmune response triggered by SARS-CoV-2 infection, vaccination, or treatments (39, 40). While both thyroid disorders and LC are more common among women, studies that controlled for age and gender still reported a higher prevalence of thyroid dysfunction in LC cohorts (39). Finally, key symptoms included asthenia, memory failure, concentration issues, muscle and joint pain, migraines, dyspnea, and palpitations; and 44% of LC participants reported low mood and anxiety, compared to 17% in the Recovered group (4).
An effective immune response is crucial for controlling SARS-CoV-2 primary infection and preventing reinfections. Impairments in both humoral and cellular immunity have been documented in COVID-19 (25, 41, 42) and LC (43–45). Our LC cohort exhibited lower SARS-CoV-2-specific IgG levels and reduced neutralization capacity. One participant had undetectable IgG levels despite a positive qPCR diagnosis, and another had IgG just above the threshold despite receiving two Comirnaty® doses. This impaired humoral response may increase susceptibility to reinfections, potentially exacerbating or perpetuating the condition. LC individuals from our cohort experienced a higher frequency of breakthrough infections compared to Recovered participants, which may contribute to the persistence of symptoms characteristic of LC (46, 47). This finding remark that an adequate vaccination schedule is essential to develop a protective humoral immunity, although not all antibodies are equally produced in vaccinated individuals. Elevated anti-N IgG levels are not directly related to vaccination and they suggest either recent reinfections or persistent viral proteins in LC individuals (48). Although these results remain controversial (49), high levels of anti-N IgG have been previously associated with LC and related neurological symptoms (50), as well as with the persistence of viral proteins in the host (51). Vaccination predominantly induces IgG against the S1 subunit and RBD, with lower induction of anti-S2 antibodies, likely because the S2 subunit is not fully exposed until the protein interacts with the ACE2 receptor (52). Interestingly, individuals vaccinated with booster doses exhibit increased levels of anti-S2 IgG compared to those with natural infection alone (53). In our LC cohort, not all individuals tested positive for anti-S2 IgG, and some presented low levels of anti-S1 IgG and reduced neutralization capacity. This implied that vaccination may have induced a less effective humoral response in LC individuals, increasing their susceptibility to reinfections. Although reinfections should also reinforce immunity against SARS-CoV-2 better than vaccination, the reinfections themselves pose additional risks, often contributing to further sequelae in multiple organ systems beyond those observed during the initial infection (54). Remarkably, 39% of LC participants had received mixed vaccine types, compared to only 3% of Recovered individuals, identifying vaccine heterogeneity as a potential LC risk factor. Together, these findings emphasize the critical role of an effective and consistent vaccination strategy in reducing the risk of LC and associated complications.
LC-associated immune dysregulation may stem from suboptimal vaccination, unresolved infection, or persistent immune activation due to viral reservoirs (9). While prolonged viral shedding in feces has been documented (11), we did not detect viral RNA in plasma or stool, likely due to the long interval (26 months) between infection and sample collection. Although viral proteins may persist in tissues or extracellular vesicles (55–57), intestinal dysbiosis may also contribute to sustained immune activation (58, 59). In our cohort, 44% of LC individuals reported persistent diarrhea. Though no direct signs of increased gut permeability were detected, REG3A levels, critical for epithelial regeneration, were significantly reduced in LC individuals (60, 61). REG3A deficiency promotes inflammation and alters microbiota (62), potentially exacerbating LC symptoms and contributing to persistent inflammation (13, 58). This correlates with the lower capacity of CD4+ Th22 cells to release IL-13 and IL-22 in people with LC (44). Th22 cells play a re-epithelializing role through IL-22 production (63) and both IL-13 and IL-22 are protective factors during acute and persistent COVID-19, promoting tissue protection and regeneration (64). Additionally, LC participants exhibited lower levels of MIG/CXCL9, MPIF/CCL23, and MPIF/CCL2, suggesting impaired immune recruitment and delayed inflammation resolution (65, 66). These deficits align with a detrimental Th1 antiviral response reported in LC (44). Psychological factors, including anxiety and depression (reported by 50% of LC cohort), could also contribute to gastrointestinal symptoms (67). Overall, these findings suggest that, while we did not observe direct signs of bacterial translocation or dysbiosis, an impaired mucosal immune axis may underlie gastrointestinal dysregulation in LC.
Persistent immune activation in LC could lead to herpesvirus reactivation (9), yet we found no significant EBV, CMV, VZV, or HSV-1/2 reactivation in LC individuals, possibly due to their heightened cytotoxic response (45). Thus, LC appears more closely associated with a sustained, uncontrolled inflammatory response rather than immunodeficiency. When the levels of pro- and anti-inflammatory markers were analyzed, CRP, a marker elevated in response to infections, tissue damage, and various inflammatory conditions (68), was increased in plasma of LC participants. CRP can remain elevated for months after acute COVID-19 (16, 69) and it has been associated with cardiovascular events due to its interaction with endothelial cells and the coagulation system (70). COVID-19 has been linked to cardiovascular disorders like myocardial injury, arrhythmia, and venous thromboembolism, due to the virus's affinity for endothelial cells (71). Several reports have identified markers of endothelial damage, microclots, complement dysregulation, thromboinflammation, and hypercoagulability in LC individuals (17, 72). In our study, prothrombin and thrombin levels were increased in plasma of LC individuals (73). Both thrombin and CRP are associated with endothelial activation and dysfunction during acute COVID-19 (74). In LC individuals, elevated thrombin levels coincided with decreased fibrinogen, a pattern that has been previously associated with an increased thrombotic risk in acute COVID-19 (17, 75). Our analysis revealed no compensatory mechanisms for this alteration, as fibrinolytic activity and the levels of the marker of fibrin degradation D-dimer remained unchanged. Additionally, LC participants showed higher plasma levels of sEPCR, which reduces the anticoagulant and anti-inflammatory effects of APC by competing with membrane-bound EPCR (36). Elevated sEPCR levels have been reported in acute severe COVID-19 and other chronic inflammatory conditions such as systemic lupus erythematosus (76), but this is the first report of increased sEPCR in plasma of LC individuals. Higher sEPCR levels may result from vascular injury or regulated proteolytic release, possibly through thrombin-induced endothelial stimulation and subsequent metalloproteinase activity (77). These findings support the persistent endothelial and coagulation dysfunctions in LC.
Although no pre-pandemic uninfected control group was available to establish basal levels of inflammatory markers, most of these markers typically normalize within 8–10 months after acute infection (78–81). Therefore, our Recovered group can be considered an adequate control, as a median of 24 months had elapsed since the acute infection. In addition, reinfections reported in some participants occurred more than 6 months before sampling, making it unlikely that they influenced the immunological and endothelial alterations detected in the LC cohort.
Among the potential limitations of this study, we must consider the differences in gender distribution, since LC predominantly affects women (53). However, as this was an observational cohort rather than a case-control study, we chose to preserve the natural female predominance in LC and to recruit a control group representative of the broader post-COVID population, rather than artificially adjusting its sex composition. This approach avoids selection bias and provides a realistic comparator group, while still allowing meaningful identification of LC-associated biological features. Other limitation is the fact that some symptoms were self-reported, which could be especially important for the cognitive manifestations that were not recorded through standardized neurocognitive tests, which may limit their objective quantification. However, the use of self-reported symptoms is broadly accepted in LC research, given that many manifestations of the syndrome (e.g., fatigue, brain fog, malaise, or pain) are inherently subjective and often not fully captured by standardized tests available in routine practice. In addition, some clinical variables used in the Random Forest analysis could be considered part of the original criteria for classifying participants as LC, which may introduce circularity and limit the interpretation of the model as an independent predictive tool. Nevertheless, including these clinical variables is important, as it allows the model to capture the central features defining LC and to identify biological markers that cluster with them, providing an integrated view of the syndrome's multidimensional profile.
In conclusion, the novelty of this study lies in its long-term, multidimensional analysis of LC-related parameters. By examining immune dysregulation and persistent endothelial damage, we identified a combined mechanism associated with coagulation abnormalities that may contribute to a pro-thrombotic profile in LC. These findings suggest that, despite impaired humoral and mucosal immunity, LC could be more strongly linked to a sustained, uncontrolled inflammatory response rather than to immunodeficiency, and appears to illuminate underlying pathogenic processes that could go beyond traditional comorbidities and challenge earlier models based on isolated pathways. Uniquely, our work also uncovers an association between vaccine heterogeneity and LC, suggesting that inconsistent vaccination strategies may compromise immune protection and contribute to the syndrome, thus emphasizing the need for uniform vaccination policies. Furthermore, the study's strength is underscored by using an exploratory Random Forest model that achieved 100% accuracy in distinguishing LC from recovered individuals, emphasized the association between key clinical features and related biological markers. This multidimensional approach not only advances our understanding of LC's complex pathogenesis but also establishes a solid foundation for enhanced diagnostic tools, ultimately leading to better prevention and clinical management strategies for affected individuals.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statement
The studies involving humans were approved by Ethics Committee of the Instituto de Salud Carlos III (IRB IORG0006384). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
AS-R: Investigation, Conceptualization, Methodology, Formal Analysis, Writing – original draft. CS-M: Writing – original draft, Investigation, Formal Analysis, Methodology. GC: Formal Analysis, Writing – original draft, Methodology. DF: Writing – original draft, Data curation, Formal Analysis, Methodology. MM-A: Writing – original draft, Resources, Methodology. EM: Investigation, Writing – original draft, Methodology. SD-M: Methodology, Resources, Writing – original draft. FP: Investigation, Methodology, Writing – original draft. JG-P: Investigation, Methodology, Writing – original draft. MP-O: Investigation, Writing – original draft, Methodology. MiC: Data curation, Formal Analysis, Writing – original draft. MM: Resources, Investigation, Project administration, Methodology, Writing – original draft. GM: Project administration, Resources, Methodology, Investigation, Writing – original draft. MT: Conceptualization, Validation, Formal Analysis, Investigation, Writing – original draft, Writing – review & editing, Data curation. MaC: Data curation, Investigation, Conceptualization, Resources, Validation, Writing – review & editing, Project administration, Supervision, Writing – original draft, Funding acquisition, Formal Analysis.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was funded by Strategic Action in Health of the Instituto de Salud Carlos III (ISCIII) (grant PI22CIII/00059) and intramural research grant (CB21/13/00034) from CIBERINFEC (Centro de Investigación Biomédica en Red Enfermedades Infecciosas, CB 2021), Instituto de Salud Carlos III, Ministerio de Ciencia e Innovación and Unión Europea – NextGenerationEU. The work of AS-R is financed by a pre-doctoral grant funded by CIBERINFEC (CB21/13/00126). The work of CS-M is financed by a pre-doctoral grant funded by the Community of Madrid (CAM), Spain (PIPF-2023_SAL-GL-30376). The work of GC is financed by the Consejería de Educación, Universidades, Ciencia y Portavocía of the Comunidad de Madrid, Spain (PEJ-2021-AI_BMD-22752). The work of MT is financed by CIBERINFEC (CB21/13/00015).
Acknowledgments
We greatly appreciate all individuals who participated in this study. We also thank the physicians who contributed with the recruitment of the participants and the collection of blood samples. We thank María Dolores Fernández García for her contribution with the extraction of DNA from stool.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that Generative AI was used in the creation of this manuscript. During the preparation of this work the authors used chatGPT in order to enhance clarity and refine complex sentence structures. After using this tool, the authors reviewed and edited the content as needed and take full responsibility for the content of the publication.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1613195/full#supplementary-material
References
1. WHO. Coronavirus (COVID-19) dashboard. Available online at: https://covid19.who.int/ (Accessed April 19, 2024).
2. Fernández-de-las-Peñas C, Raveendran AV, Giordano R, and Arendt-Nielsen L. Long COVID or post-COVID-19 condition: past, present and future research directions. Microorganisms. (2023) 11:2959. doi: 10.3390/microorganisms11122959
3. Chen C, Haupert SR, Zimmermann L, Shi X, Fritsche LG, and Mukherjee B. Global prevalence of post-coronavirus disease 2019 (COVID-19) condition or long COVID: A meta-analysis and systematic review. J Infect Dis. (2022) 226:1593–607. doi: 10.1093/infdis/jiac136
4. Rudenstine S, Schulder T, Bhatt KJ, McNeal K, Ettman CK, and Galea S. Long-COVID and comorbid depression and anxiety two years into the COVID-19 pandemic. Psychiatry Res. (2022) 317:114924. doi: 10.1016/j.psychres.2022.114924
5. Tsampasian V, Elghazaly H, Chattopadhyay R, Debski M, Naing TKP, Garg P, et al. Risk factors associated with post-COVID-19 condition: A systematic review and meta-analysis. JAMA Intern Med. (2023) 183:566–80. doi: 10.1001/jamainternmed.2023.0750
6. Shrestha Y and Venkataraman R. The prevalence of post-COVID-19 vaccination syndrome and quality of life among COVID-19-vaccinated individuals. Vacunas. (2024) 25:7–18. doi: 10.1016/j.vacune.2024.02.010
7. Chow NKN, Tsang CYW, Chan YH, Telaga SA, Ng LYA, Chung CM, et al. The effect of pre-COVID and post-COVID vaccination on long COVID: A systematic review and meta-analysis. J Infect. (2024) 89:106358. doi: 10.1016/j.jinf.2024.106358
8. Tan KWA and Koh D. Long COVID—Challenges in diagnosis and managing return-to-work. J Occup Health. (2023) 65:e12401. doi: 10.1002/1348-9585.12401
9. Chen B, Julg B, Mohandas S, Bradfute SB, and Mechanistic Pathways Task Force. Viral persistence RECOVER. reactivation, and mechanisms of long COVID. eLife. (2023) 12:e86015. doi: 10.7554/eLife.86015
10. Twomey R, DeMars J, Franklin K, Culos-Reed SN, Weatherald J, and Wrightson JG. Chronic fatigue and postexertional malaise in people living with long COVID: an observational study. Phys Ther. (2022) 102:pzac005. doi: 10.1093/ptj/pzac005
11. Natarajan A, Zlitni S, Brooks EF, Vance SE, Dahlen A, Hedlin H, et al. Gastrointestinal symptoms and fecal shedding of SARS-CoV-2 RNA suggest prolonged gastrointestinal infection. Med N Y N. (2022) 3:371–387.e9. doi: 10.1016/j.medj.2022.04.001
12. Lamers MM, Beumer J, van der Vaart J, Knoops K, Puschhof J, Breugem TI, et al. SARS-CoV-2 productively infects human gut enterocytes. Science. (2020) 369:50–4. doi: 10.1126/science.abc1669
13. Alenazy MF, Aljohar HI, Alruwaili AR, Daghestani MH, Alonazi MA, Labban RS, et al. Gut microbiota dynamics in relation to long-COVID-19 syndrome: role of probiotics to combat psychiatric complications. Metabolites. (2022) 12:912. doi: 10.3390/metabo12100912
14. Vassiliou AG, Vrettou CS, Keskinidou C, Dimopoulou I, Kotanidou A, and Orfanos SE. Endotheliopathy in acute COVID-19 and long COVID. Int J Mol Sci. (2023) 24:8237. doi: 10.3390/ijms24098237
15. Varga Z, Flammer AJ, Steiger P, Haberecker M, Andermatt R, Zinkernagel AS, et al. Endothelial cell infection and endotheliitis in COVID-19. Lancet Lond Engl. (2020) 395:1417–8. doi: 10.1016/S0140-6736(20)30937-5
16. Lai YJ, Liu SH, Manachevakul S, Lee TA, Kuo CT, and Bello D. Biomarkers in long COVID-19: A systematic review. Front Med. (2023) 10:1085988. doi: 10.3389/fmed.2023.108598
17. Turner S, Khan MA, Putrino D, Woodcock A, Kell DB, and Pretorius E. Long COVID: pathophysiological factors and abnormalities of coagulation. Trends Endocrinol Metab TEM. (2023) 34:321–44. doi: 10.1016/j.tem.2023.03.002
19. van Hinsbergh VWM. Endothelium–role in regulation of coagulation and inflammation. Semin Immunopathol. (2012) 34:93–106. doi: 10.1007/s00281-011-0285-5
20. Gando S and Akiyama T. Disseminated intravascular coagulation is associated with poor prognosis in patients with COVID-19. Sci Rep. (2024) 14:1–13. doi: 10.1038/s41598-024-63078-9
21. Gold JE, Okyay RA, Licht WE, and Hurley DJ. Investigation of long COVID prevalence and its relationship to epstein-barr virus reactivation. Pathog Basel Switz. (2021) 10:763. doi: 10.3390/pathogens10060763
22. Tandon P, Abrams ND, Avula LR, Carrick DM, Chander P, Divi RL, et al. Unraveling links between chronic inflammation and long COVID: workshop report. J Immunol Baltim Md 1950. (2024) 212:505–12. doi: 10.4049/jimmunol.2300804
23. National Institute for Health and Care Excellence (NICE). COVID-19 rapid guideline: Managing the Long-Term effects of COVID-19. (2024). Available online at: http://www.ncbi.nlm.nih.gov/books/NBK567261/ (Accessed April 20, 2024).
24. World Health Organization. Diagnostic testing for SARS-CoV-2: Interim Guidance (2020). Available online at: https://iris.who.int/handle/10665/334254 (Accessed April 30, 2024).
25. Vigón L, Fuertes D, García-Pérez J, Torres M, Rodríguez-Mora S, Mateos E, et al. Impaired cytotoxic response in PBMCs from patients with COVID-19 admitted to the ICU: biomarkers to predict disease severity. Front Immunol. (2021) 12:665329. doi: 10.3389/fimmu.2021.665329
27. Meyer HV and Birney E. PhenotypeSimulator: A comprehensive framework for simulating multi-trait, multi-locus genotype to phenotype relationships. Bioinformatics. (2018) 34:2951–6. doi: 10.1093/bioinformatics/bty197
28. Sarica A, Cerasa A, and Quattrone A. Random forest algorithm for the classification of neuroimaging data in alzheimer’s disease: A systematic review. Front Aging Neurosci. (2017) 9:329. doi: 10.3389/fnagi.2017.00329
29. Nembrini S, König IR, and Wright MN. The revival of the Gini importance? Bioinforma Oxf Engl. (2018) 34:3711–3718. doi: 10.1093/bioinformatics/bty373
30. Jadhav MA, Goldsberry WN, Zink SE, Lamb KN, Simmons KE, Riposo CM, et al. Screening cleavage of factor XIII V34X activation peptides by thrombin mutants: A strategy for controlling fibrin architecture. Biochim Biophys Acta. (2017) 1865:1246. doi: 10.1016/j.bbapap.2017.07.001
31. Weisel JW and Litvinov RI. Fibrin formation, structure and properties. Subcell Biochem. (2017) 82:405–56. doi: 10.1007/978-3-319-49674-0_13
32. Grimnes G, Isaksen T, Tichelaar YIGV, Brox J, Brækkan SK, and Hansen JB. C-reactive protein and risk of venous thromboembolism: results from a population-based case-crossover study. Haematologica. (2018) 103:1245–50. doi: 10.3324/haematol.2017.186957
33. Yepes M. Tissue-type plasminogen activator. In: Choi S, editor. Encyclopedia of signaling molecules. Springer, New York, NY (2016).
34. Kangro K, Wolberg AS, and Flick MJ. Fibrinogen, fibrin, and fibrin degradation products in COVID-19. Curr Drug Targets. (2022) 23:1593–602. doi: 10.2174/1389450123666220826162900
35. Okamoto T, Tanigami H, Suzuki K, and Shimaoka M. Thrombomodulin: A bifunctional modulator of inflammation and coagulation in sepsis. Crit Care Res Pract. (2012) 2012:614545. doi: 10.1155/2012/614545
36. Liaw PC, Neuenschwander PF, Smirnov MD, and Esmon CT. Mechanisms by which soluble endothelial cell protein C receptor modulates protein C and activated protein C function. J Biol Chem. (2000) 275:5447–52. doi: 10.1074/jbc.275.8.5447
37. Bui TM, Wiesolek HL, and Sumagin R. ICAM-1: A master regulator of cellular responses in inflammation, injury resolution, and tumorigenesis. J Leukoc Biol. (2020) 108:787–99. doi: 10.1002/JLB.2MR0220-549R
38. Bai F, Tomasoni D, Falcinella C, Barbanotti D, Castoldi R, Mulè G, et al. Female gender is associated with long COVID syndrome: a prospective cohort study. Clin Microbiol Infect Off Publ Eur Soc Clin Microbiol Infect Dis. (2022) 28:611.e9–611.e16. doi: 10.1016/j.cmi.2021.11.002
39. Rossini A, Cassibba S, Perticone F, Benatti SV, Venturelli S, Carioli G, et al. Increased prevalence of autoimmune thyroid disease after COVID-19: A single-center, prospective study. Front Endocrinol. (2023) 14:1126683. doi: 10.3389/fendo.2023.1126683
40. Caron P. Autoimmune and inflammatory thyroid diseases following vaccination with SARS-CoV-2 vaccines: from etiopathogenesis to clinical management. Endocrine. (2022) 78:406–17. doi: 10.1007/s12020-022-03118-4
41. Vigón L, García-Pérez J, Rodríguez-Mora S, Torres M, Mateos E, Castillo de la Osa M, et al. Impaired antibody-dependent cellular cytotoxicity in a spanish cohort of patients with COVID-19 admitted to the ICU. Front Immunol. (2021) 12:742631. doi: 10.3389/fimmu.2021.742631
42. Casado-Fernández G, Corona M, Torres M, Saez AJ, Ramos-Martín F, Manzanares M, et al. Sustained cytotoxic response of peripheral blood mononuclear cells from unvaccinated individuals admitted to the ICU due to critical COVID-19 is essential to avoid a fatal outcome. Int J Environ Res Public Health. (2023) 20:1947. doi: 10.3390/ijerph20031947
43. Yin K, Peluso MJ, Luo X, Thomas R, Shin MG, Neidleman J, et al. Long COVID manifests with T cell dysregulation, inflammation and an uncoordinated adaptive immune response to SARS-CoV-2. Nat Immunol. (2024) 25:218–25. doi: 10.1038/s41590-023-01724-6
44. Sánchez-Menéndez C, de la Calle-Jiménez O, Mateos E, Vigón L, Fuertes D, Antón MAM, et al. Different polarization and functionality of CD4+ T helper subsets in people with post-COVID condition. Front Immunol. (2024) 15:1431411. doi: 10.3389/fimmu.2024.1431411
45. Galán M, Vigón L, Fuertes D, Murciano-Antón MA, Casado-Fernández G, Domínguez-Mateos S, et al. Persistent overactive cytotoxic immune response in a spanish cohort of individuals with long-COVID: identification of diagnostic biomarkers. Front Immunol. (2022) 13:848886. doi: 10.3389/fimmu.2022.848886
46. Barrera Martínez Y, Boillat Oriani GA, Vega Montes P, Martínez Moreno E, Pérez Pérez A, Casajuana Pérez RJ, et al. Long-term persistence of post-COVID-19 symptoms: A two-year follow-up of a Primary Care cohort. Sist Sanit Navar. (2025) 48:e1101. doi: 10.23938/ASSN.1101
47. Liu D, Chen B, Liao X, Zhang Z, Wei S, Han X, et al. Specific persistent symptoms of COVID-19 and associations with reinfection: a community-based survey study in southern China. Front Public Health. (2024) 12:1452233. doi: 10.3389/fpubh.2024.1452233
48. Kervevan J, Staropoli I, Slama D, Jeger-Madiot R, Donnadieu F, Planas D, et al. Divergent adaptive immune responses define two types of long COVID. Front Immunol. (2023) 14:1221961. doi: 10.3389/fimmu.2023.1221961
49. Phetsouphanh C, Jacka B, Ballouz S, Jackson KJL, Wilson DB, Manandhar B, et al. Improvement of immune dysregulation in individuals with long COVID at 24-months following SARS-CoV-2 infection. Nat Commun. (2024) 15:1–15. doi: 10.1038/s41467-024-47720-8
50. Su Y, Yuan D, Chen DG, Ng RH, Wang K, Choi J, et al. Multiple early factors anticipate post-acute COVID-19 sequelae. Cell. (2022) 185:881–895.e20. doi: 10.1016/j.cell.2022.01.014
51. Pang NYL, Pang ASR, Chow VT, and Wang DY. Understanding neutralising antibodies against SARS-CoV-2 and their implications in clinical practice. Mil Med Res. (2021) 8:47. doi: 10.1186/s40779-021-00342-3
52. Altmann DM, Whettlock EM, Liu S, Arachchillage DJ, and Boyton RJ. The immunology of long COVID. Nat Rev Immunol. (2023) 23:618–34. doi: 10.1038/s41577-023-00904-7
53. Meyers J, Windau A, Schmotzer C, Saade E, Noguez J, Stempak L, et al. SARS-CoV-2 antibody profile of naturally infected and vaccinated individuals detected using qualitative, semi-quantitative and multiplex immunoassays. Diagn Microbiol Infect Dis. (2022) 104:115803. doi: 10.1016/j.diagmicrobio.2022.115803
54. Bowe B, Xie Y, and Al-Aly Z. Acute and postacute sequelae associated with SARS-CoV-2 reinfection. Nat Med. (2022) 28:2398–405. doi: 10.1038/s41591-022-02051-3
55. Goh D, Lim JCT, Fernaíndez SB, Joseph CR, Edwards SG, Neo ZW, et al. Case report: Persistence of residual antigen and RNA of the SARS-CoV-2 virus in tissues of two patients with long COVID. Front Immunol. (2022) 13:939989. doi: 10.3389/fimmu.2022.939989
56. Craddock V, Mahajan A, Spikes L, Krishnamachary B, Ram AK, Kumar A, et al. Persistent circulation of soluble and extracellular vesicle-linked Spike protein in individuals with postacute sequelae of COVID-19. J Med Virol. (2023) 95:e28568. doi: 10.1002/jmv.28568
57. Peluso MJ, Deeks SG, Mustapic M, Kapogiannis D, Henrich TJ, Lu S, et al. SARS-coV-2 and mitochondrial proteins in neural-derived exosomes of COVID-19. Ann Neurol. (2022) 91:772–81. doi: 10.1002/ana.26350
58. Zhang D, Zhou Y, Ma Y, Chen P, Tang J, Yang B, et al. Gut microbiota dysbiosis correlates with long COVID-19 at one-year after discharge. J Korean Med Sci. (2023) 38:e120. doi: 10.3346/jkms.2023.38.e120
59. Bernard-Raichon L, Venzon M, Klein J, Axelrad JE, Zhang C, Sullivan AP, et al. Gut microbiome dysbiosis in antibiotic-treated COVID-19 patients is associated with microbial translocation and bacteremia. Nat Commun. (2022) 13:5926. doi: 10.1038/s41467-022-33395-6
60. Zhao D, Kim YH, Jeong S, Greenson JK, Chaudhry MS, Hoepting M, et al. Survival signal REG3α prevents crypt apoptosis to control acute gastrointestinal graft-versus-host disease. J Clin Invest. (2018) 128:4970–9. doi: 10.1172/JCI99261
61. Shin JH and Seeley RJ. Reg3 proteins as gut hormones? Endocrinology. (2019) 160:1506–14. doi: 10.1210/en.2019-00073
62. Vaishnava S, Yamamoto M, Severson KM, Ruhn KA, Yu X, Koren O, et al. The antibacterial lectin RegIIIgamma promotes the spatial segregation of microbiota and host in the intestine. Science. (2011) 334:255–8. doi: 10.1126/science.1209791
63. Arshad T, Mansur F, Palek R, Manzoor S, and Liska V. A double edged sword role of interleukin-22 in wound healing and tissue regeneration. Front Immunol. (2020) 11:2148. doi: 10.3389/fimmu.2020.02148
64. Pathinayake PS, Awatade NT, and Wark PAB. Type 2 immunity and its impact on COVID-19 infection in the airways. Viruses. (2023) 15:402. doi: 10.3390/v15020402
65. Baresi G, Giacomelli M, Moratto D, Chiarini M, Conforti IC, Padoan R, et al. Case report: analysis of inflammatory cytokines IL-6, CCL2/MCP1, CCL5/RANTES, CXCL9/MIG, and CXCL10/IP10 in a cystic fibrosis patient cohort during the first wave of the COVID-19 pandemic. Front Pediatr. (2021) 9:645063. doi: 10.3389/fped.2021.645063
66. Arruda-Silva F, Bianchetto-Aguilera F, Gasperini S, Polletti S, Cosentino E, Tamassia N, et al. Human neutrophils produce CCL23 in response to various TLR-agonists and TNFα. Front Cell Infect Microbiol. (2017) 7:176. doi: 10.3389/fcimb.2017.00176
67. Freedberg DE and Chang L. Gastrointestinal symptoms in COVID-19: the long and the short of it. Curr Opin Gastroenterol. (2022) 38:555–61. doi: 10.1097/MOG.0000000000000876
68. Luan YY, Yin CH, and Yao YM. Update advances on C-reactive protein in COVID-19 and other viral infections. Front Immunol. (2021) 12. doi: 10.3389/fimmu.2021.720363
69. Espín E, Yang C, Shannon CP, Assadian S, He D, and Tebbutt SJ. Cellular and molecular biomarkers of long COVID: a scoping review. eBioMedicine. (2023) 91:104552. doi: 10.1016/j.ebiom.2023.104552
70. Dix C, Zeller J, Stevens H, Eisenhardt SU, Shing KSCT, Nero TL, et al. C-reactive protein, immunothrombosis and venous thromboembolism. Front Immunol. (2022) 13:1002652. doi: 10.3389/fimmu.2022.1002652
71. Conway EM, Mackman N, Warren RQ, Wolberg AS, Mosnier LO, Campbell RA, et al. Understanding COVID-19-associated coagulopathy. Nat Rev Immunol. (2022) 22:639–49. doi: 10.1038/s41577-022-00762-9
72. Cervia-Hasler C, Brüningk SC, Hoch T, Fan B, Muzio G, Thompson RC, et al. Persistent complement dysregulation with signs of thromboinflammation in active Long Covid. Science. (2024) 383:eadg7942. doi: 10.1126/science.adg7942
73. Fan BE, Wong SW, Sum CLL, Lim GH, Leung BP, Tan CW, et al. Hypercoagulability, endotheliopathy, and inflammation approximating 1 year after recovery: Assessing the long-term outcomes in COVID-19 patients. Am J Hematol. (2022) 97:915–23. doi: 10.1002/ajh.26575
74. Zhang J, Tecson KM, and McCullough PA. Endothelial dysfunction contributes to COVID-19-associated vascular inflammation and coagulopathy. Rev Cardiovasc Med. (2020) 21:315–9. doi: 10.31083/j.rcm.2020.03.126
75. Gorog DA, Storey RF, Gurbel PA, Tantry US, Berger JS, Chan MY, et al. Current and novel biomarkers of thrombotic risk in COVID-19: a Consensus Statement from the International COVID-19 Thrombosis Biomarkers Colloquium. Nat Rev Cardiol. (2022) 19:475. doi: 10.1038/s41569-021-00665-7
76. Bayrakci N, Ozkan G, Mutlu LC, Erdem L, Yildirim I, Gulen D, et al. Relationship between serum soluble endothelial protein C receptor level and COVID-19 findings. Blood Coagul Fibrinolysis. (2021) 32:550–5. doi: 10.1097/MBC.0000000000001070
77. Xu J, Qu D, Esmon NL, and Esmon CT. Metalloproteolytic release of endothelial cell protein C receptor. J Biol Chem. (2000) 275:6038–44. doi: 10.1074/jbc.275.8.6038
78. Ghaffarpour S, Ghazanfari T, Ardestani SK, Naghizadeh MM, Vaez Mahdavi MR, Salehi M, et al. Cytokine profiles dynamics in COVID-19 patients: a longitudinal analysis of disease severity and outcomes. Sci Rep. (2025) 15:14209. doi: 10.1038/s41598-025-98505-y
79. Phetsouphanh C, Darley DR, Wilson DB, Howe A, Munier CML, Patel SK, et al. Immunological dysfunction persists for 8 months following initial mild-to-moderate SARS-CoV-2 infection. Nat Immunol. (2022) 23:210–6. doi: 10.1038/s41590-021-01113-x
80. Sommen SL, Havdal LB, Selvakumar J, Einvik G, Leegaard TM, Lund-Johansen F, et al. Inflammatory markers and pulmonary function in adolescents and young adults 6 months after mild COVID-19. Front Immunol. (2023) 13:1081718. doi: 10.3389/fimmu.2022.1081718
Keywords: Long COVID, immunity, COVID-19 vaccines, endothelium, blood coagulation disorders, biomarkers
Citation: Simón-Rueda A, Sánchez-Menéndez C, Casado G, Fuertes D, Murciano-Antón MA, Mateos E, Domínguez-Mateos S, Pozo F, García-Pérez J, Pérez-Olmeda M, Cervero M, Massanella M, Moncunill G, Torres M and Coiras M (2025) Immune dysregulation and endothelial dysfunction associate with a pro-thrombotic profile in Long COVID. Front. Immunol. 16:1613195. doi: 10.3389/fimmu.2025.1613195
Received: 16 April 2025; Accepted: 30 September 2025;
Published: 16 October 2025.
Edited by:
Mariza Gonçalves Morgado, Oswaldo Cruz Foundation (Fiocruz), BrazilReviewed by:
Giulia Unali, Boston University, United StatesEugenio D. Hottz, Juiz de Fora Federal University, Brazil
Copyright © 2025 Simón-Rueda, Sánchez-Menéndez, Casado, Fuertes, Murciano-Antón, Mateos, Domínguez-Mateos, Pozo, García-Pérez, Pérez-Olmeda, Cervero, Massanella, Moncunill, Torres and Coiras. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mayte Coiras, bWNvaXJhc0Bpc2NpaWkuZXM=; Montserrat Torres, bS50b3JyZXNAZXh0ZXJub3MuaXNjaWlpLmVz