 Eszter Tóth1†
Eszter Tóth1† Máté Posta1,2,3†
Máté Posta1,2,3† Dániel Györffy1,4
Dániel Györffy1,4 Orsolya Oravecz1,5
Orsolya Oravecz1,5 Emese Farkas1,2
Emese Farkas1,2 Andrea Balogh1
Andrea Balogh1 Claudia Escher6Magdalena Bober6
Claudia Escher6Magdalena Bober6 András Szilágyi1Petronella Hupuczi7Lajos Veress8
András Szilágyi1Petronella Hupuczi7Lajos Veress8 Olga Török9
Olga Török9 Sándor Nagy10Oliver Rinner6
Sándor Nagy10Oliver Rinner6 Offer Erez11Zoltán Papp7,12
Offer Erez11Zoltán Papp7,12 Nándor Ács12
Nándor Ács12 Nándor Gábor Than1,7,12*
Nándor Gábor Than1,7,12*- 1Systems Biology of Reproduction Research Group, Institute of Molecular Life Sciences, HUN-REN Research Centre for Natural Sciences, Budapest, Hungary
- 2Doctoral College, Károly Rácz Conservative Medicine Division, Semmelweis University, Budapest, Hungary
- 3Department of Bioinformatics, Semmelweis University, Budapest, Hungary
- 4Faculty of Information Technology and Bionics, Pázmány Péter Catholic University, Budapest, Hungary
- 5Doctoral School of Biology, Institute of Biology, ELTE Eötvös Loránd University, Budapest, Hungary
- 6Biognosys AG, Schlieren, Switzerland
- 7Maternity Private Clinic of Obstetrics and Gynecology, Budapest, Hungary
- 8Department of Pharmacology and Pharmacother, University of Debrecen, Debrecen, Hungary
- 9Department of Obstetrics and Gynecology, Faculty of Medicine, University of Debrecen, Debrecen, Hungary
- 10Department of Obstetrics of Gynecology, Széchenyi István University, Győr, Hungary
- 11Department of Obstetrics and Gynecology, Ben-Gurion University, Beer Sheba, Israel
- 12Department of Obstetrics and Gynecology, Semmelweis University, Budapest, Hungary
Introduction: Miscarriages affect 50-70% of all gestations and 15-20% of clinically recognized pregnancies. Recurrent pregnancy loss (RPL) occurs in 1-5% of clinical pregnancies and has an enormous demographic impact. However, the etiologies and molecular pathways of RPL are scarcely understood, and therefore, reliable diagnostic and preventive methods are not yet available. Here, we aimed to discover novel biomarkers for RPL using next-generation proteomics technology to help develop early and effective diagnostic tools.
Methods: First-trimester blood samples were collected from women with RPL (n=11) and controls with elective termination of pregnancy (n=11) between 6–13 weeks of gestation. After immunodepleting 14 highly abundant proteins, plasma samples were reduced, alkylated, and trypsin digested. For the separation of peptides, nano-flow reversed-phase chromatography was applied, and then mass spectrometric analysis was performed. Differentially abundant (DA) proteins were identified using strict criteria and analyzed by protein network and Gene Ontology (GO) enrichment analyses, and two biomarker candidates (CGB and PAPPA) were validated by immunoassay. Biomarker predictive properties were demonstrated using Receiver Operating Characteristic (ROC) curves. Assessments were performed for all cases and then for two gestational age groups, before and after the start of placental circulation [“early RPL”: gestational weeks (GW) 6–9, “late RPL”: GW 9–13].
Results: Altogether, 651 proteins were identified and quantified across all samples. When comparing “early control” and “late control” samples, 60 proteins [11 predominantly placenta-expressed (PPE)] were DA. When analyzing all cases, 50 DA proteins were found in RPL (top 3 down: PZP, PSG9, CGB; top 3 up: C4BPA, HBA, HBB), among which 11 PPE proteins were found, all downregulated. Enriched GO terms included ‘placental function’, ‘oxidative processes’, ‘immune function’, and ‘blood coagulation’ related biological processes. When cases were split into early and late RPL groups, 40 DA proteins were identified in early RPL (top 3 down: SHBG, CGB, CGA; top 3 up: C4BPA, SAMP, C4BPB) and 90 in late RPL (top 3 down: PZP, PAPPA, PSG9; top 3 up: THBS1, ECM1, HBB), among which only 15 were shared by both RPL groups. In early RPL, only ‘placental function’ and ‘immune function’ related biological processes were enriched, while in late RPL the top enriched GO terms included ‘placental function’, ‘oxidative processes’, ‘immune function’, ‘blood coagulation’, ‘angiogenesis’, ‘cell migration’, and ‘blood circulation’ related biological processes. Among GO terms, only ‘placental function’ related biological processes were enriched when early- and late RPL DA proteins were analyzed together. Furthermore, the areas under the ROC curves were >0.9 for two protein candidates in all RPL, for five proteins in early RPL, and for ten proteins in late RPL. Among these candidates, CGB and PAPPA were validated by immunoassay which showed a good correlation with MS data (RCGB=0.795 and RPAPPA=0.965).
Conclusion: We discovered distinct as well as shared molecular pathways associated with RPL pathogenesis before and after the start of placental circulation and identified novel biomarkers for these pathways which have outstanding discriminative properties. Our results may facilitate a better understanding of the molecular pathways of RPL. However, larger clinical studies are needed to investigate whether the identified biomarkers also have predictive power for RPL before pregnancies fail and to test drugs for the modulation of the identified disease pathways and the prevention of RPL. Our findings highlight the importance of the maternal immune system in maintaining successful pregnancy and suggest that targeting immune pathways may offer novel therapeutic approaches for RPL.
Introduction
Miscarriage is the loss of pregnancy before the 20th week of gestation (1, 2), which affects 50-70% of all conceptions and 15-20% of clinically recognized pregnancies (3–15). Recurrent pregnancy loss (RPL) occurs in 1-5% of clinical pregnancies (6, 10, 16–18) and was defined as the loss of three or more consecutive pregnancies before the 20th week of gestation by the World Health Organization (6, 10, 18–20), and later as two or more failed clinical pregnancies by the American Society for Reproductive Medicine (18, 21). The risk of pregnancy loss during a given gestation was found to increase directly with the number of previous miscarriages (14, 15, 22), however, it is almost the same (i.e. 30% and 33%) after two or three previous miscarriages (17). Therefore, there is an increasing application of the second definition of RPL. In general, miscarriages and RPL have substantial social, psychological, and economic impacts, and in developed countries, where reproductive rates are declining, they may largely influence the demographic processes as well (23, 24).
There are several etiological factors associated with RPL, such as genetic (6, 19, 25–30), anatomical (6, 28, 31–33), immunological (6, 34–40), endocrine (6, 19, 41–45), and thrombotic (6, 46–49) predispositions, including antiphospholipid syndrome (6, 19, 50–53). Moreover, maternal age is also a strong risk factor for subsequent miscarriages (10). Nevertheless, the causes and molecular pathology of about half of the cases are still unknown (6, 13, 18, 21, 54). It is highly probable that causes of RPL vary among different gestational ages, and the timing of miscarriage may have prognostic importance for subsequent pregnancies in each individual case (55–58). However, exact clinical data about gestational ages of miscarriages are not published appropriately in many cases. Therefore, in the future special attention should be paid to the gestational age-specific etiology of RPL. Since the etiologies, molecular pathways, and timing of RPL are not yet comprehensively understood, early and reliable diagnostic and preventive methods are not yet available either.
Pregnancy is a unique immunological state where the maternal immune system must provide tolerance towards the semi-allogenic fetus while maintaining robust defense mechanisms against infections. This complex interaction is orchestrated through immune-modulating mechanisms involving regulatory T cells, cytokines, and trophoblast-derived signals (59–61). Any dysregulation in this immune response can lead to pregnancy complications, including miscarriage and RPL. Altered natural killer cell activity and cytokine profiles can impair maternal-fetal tolerance, leading to placental dysfunction and pregnancy loss (62). Additionally, a heightened inflammatory response, characterized by increased levels of pro-inflammatory cytokines, can contribute to tissue damage and fetal demise (63).
There are several protein biomarker studies in the literature for RPL, either for risk assessment of non-pregnant women (44, 64–70) or for the prediction of the outcome of the current pregnancy (38, 71–84). However, the results of the different studies are often confusing and not comparable because of the heterogeneous patient groups, inadequate definitions, or inappropriate methodology applied (85, 86). Therefore, there is currently no unified protocol for the diagnosis and prediction of RPL.
Although early pregnancy loss is typically defined clinically as a loss before the 13th completed week of gestation (87), in this study we applied a gestational age cut-off at the 9th gestational week to distinguish biologically distinct phases of placental development. Previously, we studied three serum proteins in RPL and defined their exact predictive values (88). Free β-human chorionic gonadotropin (CGB) and pregnancy-associated plasma protein A (PAPPA) were found to be valuable biomarkers for early RPL cases [gestational weeks (GW) 6–9], and their discriminative power was excellent for late RPL cases (GW 9–13), while placental growth factor (PGF) was a good biomarker for late RPL. The more effective diagnosis of early RPL, as well as a better understanding of the pathology of RPL, requires comprehensive molecular studies on the level of the genome, transcriptome, proteome, and metabolome using state-of-the-art techniques and systems biology approaches. Therefore, here we aimed to discover novel biomarkers for RPL using next-generation proteomics technology and proteome-wide analysis of blood plasma samples collected in a case-control study design including women with RPL and those women undergoing elective termination of pregnancy as a gestational age-matched control group. We used strict clinical definitions and homogenous patient groups. Moreover, standardized sample collection, sample storage, treatment, and analysis, as well as data evaluation using different bioinformatics tools and statistical methods were applied.
Materials and methods
Study groups, clinical definitions, and sample collection
The following clinical groups were studied: 1) women with RPL (n=11) and control women with elective termination of pregnancy (n=11). Each RPL sample was paired with gestational age-matched control. Blood samples were collected within the HUN-PER study at the Maternity Private Clinic of Obstetrics and Gynecology (Budapest, Hungary) at the time of surgery. All women were Caucasian. Samples from pregnancies with congenital or chromosomal abnormalities or multiple gestations were not collected. Pregnancies were dated according to ultrasound scans and samples were collected between 6–13 weeks of gestation. Table 1 contains the demographic and clinical data of the two study groups.
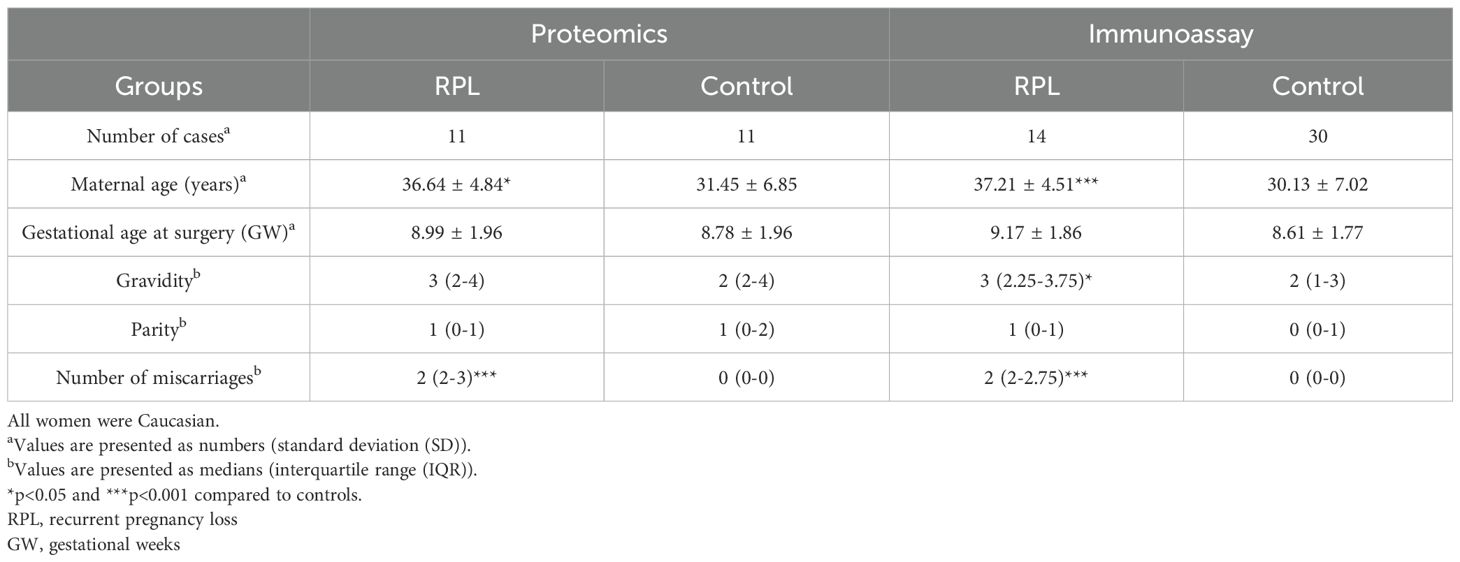
Table 1. Demographic and clinical data of the study groups.
RPL was defined as two or more failed clinical pregnancies by the American Society for Reproductive Medicine (21). RPL cases were recruited from patients with a nonviable intrauterine pregnancy detected by ultrasound [gestational sac containing an embryo or fetus without fetal heart activity within the first 12 6/7 weeks of gestation according to the American College of Obstetricians and Gynecologists Practice Bulletin (87)]. We did not pay special concern whether previous failed first-trimester clinical pregnancies were complete/incomplete spontaneous or missed abortions. In the control group, elective termination of pregnancy was performed at the request of pregnant women for non-medical reasons in all cases.
Processing of blood samples
Blood samples were processed immediately after collection. Plasma and serum samples were separated and stored at −80°C in our Perinatal Biobank at the HUN-REN Research Centre for Natural Sciences.
Sample preparation for proteomics
Plasma samples were depleted using a Human-14 Multi Affinity Removal Spin Cartridge (Agilent Technologies) according to the manufacturer’s instructions. After the addition of Biognosys’ Lysis Buffer, digestion was carried out on single filter units (Sartorius Vivacon 500, 30’000 MWCO HY) following a modified FASP protocol (described by the Max Planck Institute of Biochemistry, Martinsried, Germany).
Samples were reduced and alkylated using Biognosys’ Reduction & Alkylation Solution for 30min at 37°C. Subsequently, digestion to peptides was carried out using trypsin (Promega Corporation, Madison, WI, USA) overnight at 37°C and a protein:protease ratio of 1:100.
Clean-up for mass spectrometry
Peptides were desalted using C18 Midi BioPure columns (NestGroup, Ipswich, MA, USA) according to the manufacturer’s instructions and dried down using a SpeedVac system. Peptides were resuspended in LC solvent A [1% acetonitrile, 0.1% formic acid (FA)] (Sigma-Aldrich, St. Louis, MO, USA). Prior to mass spectrometric analyses, they were spiked with Biognosys’ iRT kit calibration peptides. Peptide concentrations were determined using a UV/VIS Spectrometer (SPECTROstar Nano, BMG Labtech Ortenberg, Germany).
High-Resolution Mass Spectrometry using Tandem Mass Spectrometry
For High-Resolution Mass Liquid Chromatography-Tandem Mass Spectrometry (HRM LC-MS/MS) measurements, 2µg of peptides were injected into an in-house packed C18 column (Dr. Maisch ReproSil Pur, 1.9µm particle size, 120Å pore size; 75µm inner diameter, 50cm length, New Objective) on a Thermo Scientific™ Easy nLC 1200 nano-liquid chromatography system connected to a Thermo Scientific™ Q Exactive™ HF-X mass spectrometer equipped with a standard nano-electrospray source. LC solvents were A: 1% acetonitrile in water with 0.1% FA; B: 15% water in acetonitrile with 0.1% FA (all solvents from Sigma-Aldrich, St. Louis, MO, USA). The nonlinear LC gradient was 1-52% solvent B in 60min followed by 52-90% B in 10sec, 90% B for 10min, 90-1% B in 10sec and 1% B for 5min. A data-independent acquisition (DIA) method with one full-range survey scan and 14 DIA windows was used.
Immunoassay
Three selected biomarker proteins were analyzed applying the same study groups (RPL, n=14; control, n=30) including the same samples as described in our previous study (88). Briefly, serum concentrations of CGB, PAPPA, and PGF proteins were measured with immunoassay, using a BRAHMS plus KRYPTOR Analyzer (ThermoFisher Scientific, Waltham, MA, USA) with monoclonal antibodies targeted to different epitopes of the measured analytes. Here, the results of CGB and PAPPA were used for re-analysis.
Data analysis
Analysis of HRM mass spectrometric data
HRM mass spectrometric data were analyzed using Spectronaut™ X software (Biognosys). The false discovery rate (FDR) on peptide and protein levels was set to 1%, data was filtered using row-based extraction. An assay library (protein inventory) generated previously (combined from plasma and placenta samples) was used for the analysis, and depleted proteins were removed from the dataset. The HRM measurements analyzed with Spectronaut were normalized using local regression normalization, and sparse protein profiles were used for the normalization of the samples (89). Sparse filtered data (no missing values, identification was included if the target was identified in at least one sample with q-value <0.01) were reported. Protein quantities were determined by averaging the intensities of the top 1–3 peptides, ranked by Spectronaut.
Identification of placenta-specific proteins
In this study, we also focused on predominantly placenta-expressed (PPE) proteins, as they serve as molecular markers of placental development and function. PPE proteins were specified as proteins encoded by PPE genes (n=164) as defined by Than et al. (90) and Szilagyi et al. (91). PPE genes’ placental expression is high which is unique or several folds higher than in other tissues, and their dysregulation may reflect altered placental functions.
Identification of differentially abundant proteins
A two-sample unpaired Student’s t-test was performed for the protein intensities between RPL and control samples. The protein regulation was calculated as the log2 ratios of the individual protein intensities averaged over the study groups. P-values were corrected for multiple testing using the q-value approach (92) to control the overall false discovery rate. Up- and down-regulated proteins were selected by two filtering cut-offs: q-values <0.1 and absolute log2 (fold-change) >log2(1.5).
We split samples into two groups according to the gestational age at the time of sampling: one for samples with gestational age <9 weeks (n=7) and another for samples taken at gestational age ≥9 weeks (n=4). DA proteins were calculated, and further analyses were performed also for early RPL and late RPL separately. This cut-off was selected based on the culmination of two critical developmental milestones, the luteal-placental shift and the onset of intervillous circulation. The luteal-placental shift in which progesterone production transitions from the corpus luteum to the placenta typically occurs gradually between the 6th to 9th week of gestation as shown first by the pioneering study of Csapo et al. (58, 93, 94). In addition, trophoblastic plugs that provide hypoxic environment for the developing embryo at the beginning of pregnancy start to dissolve at around 9–10 weeks of gestation, giving place to the start of the maternal circulation of the placenta as evidenced by the rising oxygen tension and change in flow patterns detected with Doppler ultrasound. This is also the time period when the first wave of extravillous trophoblast invasion starts to occur, radically altering the placental milieu (95). Therefore, using 9 weeks as a biological cut-off provides a meaningful demarcation for analyzing gestational age–dependent molecular changes in maternal plasma.
Protein network and Gene Ontology enrichment analyses
Protein networks were retrieved from the STRING database (96) for proteins in the sparse filtered dataset and the following thresholds for candidate identification: q-value <0.1, absolute log2 fold-change >1.5. Gene Ontology (GO) (97) enrichment analysis was performed using the STRING website, and biological processes were checked for over-representation within the set of candidates.
Results are presented using volcano plots showing q-values and fold changes for down- and upregulated DA as well as non-DA proteins, and mosaic plots showing the enrichment of PPE proteins among different groups. We also present the interaction networks of DA proteins retrieved from the STRING database, and the most enriched GO biological processes in the various subgroups.
Classifier to discriminate between RPL and control groups
We trained a Logistic Regression (LR) classifier for each DA protein separately to estimate the predictive power of single proteins. The training process was preceded by hyperparameter optimization. The solver and regularization methods obtained by the optimization process were used for the classifier. An AUC value averaging over 50 runs of five-fold cross-validation procedures was calculated.
To account for potential differences in classification performance at different stages of pregnancy, we trained gestational age-specific classifiers using the same evaluation procedure. Separate classifiers were trained for the two groups of samples, employing an LR classifier for each DA protein. The models were assessed using 50 runs of three-fold cross-validation, and the resulting AUC values were averaged to evaluate predictive performance.
Immunoassay re-analysis and correlation analysis
To validate the proteomic results, we re-analyzed published immunoassay data (88) on the abundance of two clinically established biomarkers – PAPPA and CGB – in a set of plasma samples (total n=44). This included 22 samples that overlapped with the current mass spectrometry cohort, and an additional 22 samples that were independent of this study.
Former immunoassay results of CGB and PAPPA (88) were re-analyzed for the overlapping cohort. Correlation analysis was performed between the immunoassay and proteomic results, and the Pearson correlation coefficients (R) were calculated.
Results
Overall demographic, clinical and proteomic characteristics of the study population
Demographic and clinical information for both the proteomic and validation cohorts is summarized in Table 1. In both sets, the mean maternal age was higher in the RPL than in the control group. Women in the RPL group had experienced two to four spontaneous miscarriages, including the current one, while those in the control group did not have any previous miscarriages.
Using high-resolution mass spectrometry with a strict limit of quantification, we identified and quantified 651 proteins across all plasma samples (RPL, n=11; control, n=11), of which 27 were classified as PPE proteins (Supplementary Table 1).
Given the higher average maternal age in the RPL group, we investigated whether maternal age might act as a potential confounding factor influencing the proteomic data. To assess this, we performed a correlation analysis between maternal age and the abundance of all quantified proteins across the entire cohort. After correcting for multiple hypothesis testing using Storey’s q-value method, none of the proteins showed a statistically significant association with maternal age, suggesting that maternal age did not have a detectable impact on the observed proteomic differences.
Changes in the maternal plasma proteome in the first trimester in control pregnancies
Between early control (n=7) and late control (n=4) samples (Figure 1, Supplementary Table 2), 60 DA proteins were identified, among them 11 were PPE proteins (enrichment factor: 4.59, p=4.31×10-6). 29 proteins were downregulated (one PPE, enrichment non-significant) and 31 upregulated (ten PPE, enrichment factor: 8.08, p=4.11×10-8).
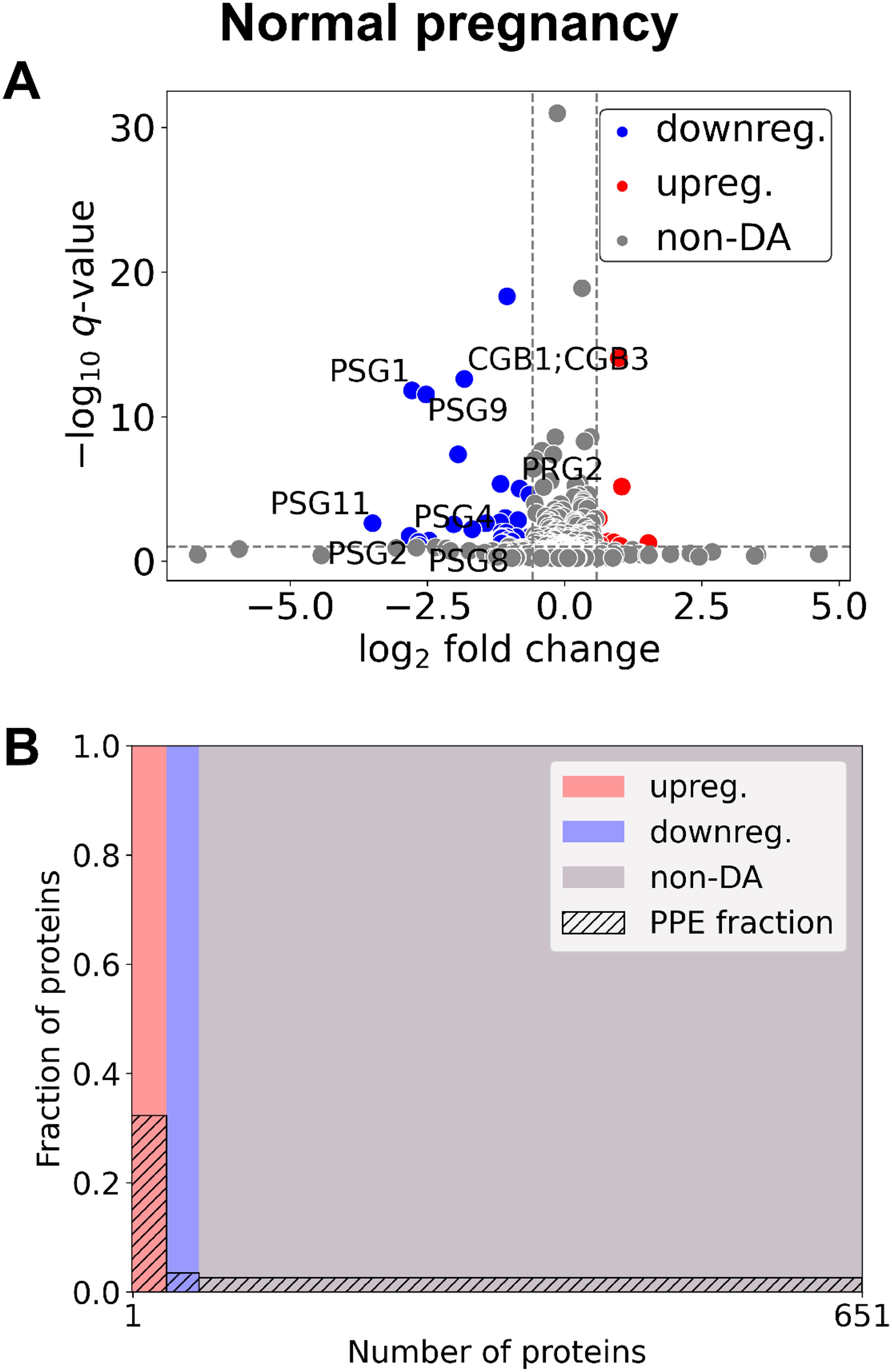
Figure 1. Changes of the maternal plasma proteome in the first trimester of normal pregnancies. (A) Changes in protein intensities among late- (GW 9–13) and early control samples (GW 6–9) are represented using a volcano plot. Proteins are shown in terms of their measured differences in abundance (x-axis) and the significance of the difference (y-axis). Differences in abundance are represented as log2(fold change). The significance is represented as a –log10 (q-value). Dotted lines represent the thresholds used to select the probes for DA proteins: absolute log2 fold change 1.5 and –log10 (q-value) >1 for statistical significance. Downregulated proteins are shown in blue, upregulated proteins are marked in red, and non-DA proteins are shown as grey dots. Gene IDs of PPE DA proteins are shown as well. (B) Mosaic plot shows the enrichment of PPE proteins (striped part). The width of each colored rectangle is proportional to the number of proteins in the corresponding group, and the height of the striped part is proportional to the fraction of PPE proteins within the particular group. Upregulated proteins are marked red, downregulated proteins blue, and non-DA proteins grey. DA, differentially abundant; GW, gestational weeks; PPE, predominantly placenta-expressed.
Changes in the maternal plasma proteome in the first trimester in RPL
In the whole gestational age range (Figure 2, Table 2, Supplementary Tables 2, 3) 50 DA proteins were identified (top 3 down: PZP, PSG9, CGB; top 3 up: C4BPA, HBA, HBB), 36 were downregulated and 14 were upregulated. Eleven PPE proteins were found among DA proteins, corresponding to an enrichment factor of 5.51 and p=5.91×10-7. All of these PPE proteins were down-regulated in RPL (enrichment factor: 7.65, p=1.31×10-8). Significantly enriched GO terms included ‘placental function’ (3/27), ‘oxidative processes’ (4/27), ‘immune function’ (8/27), and ‘blood coagulation’ (1/27) related biological processes.
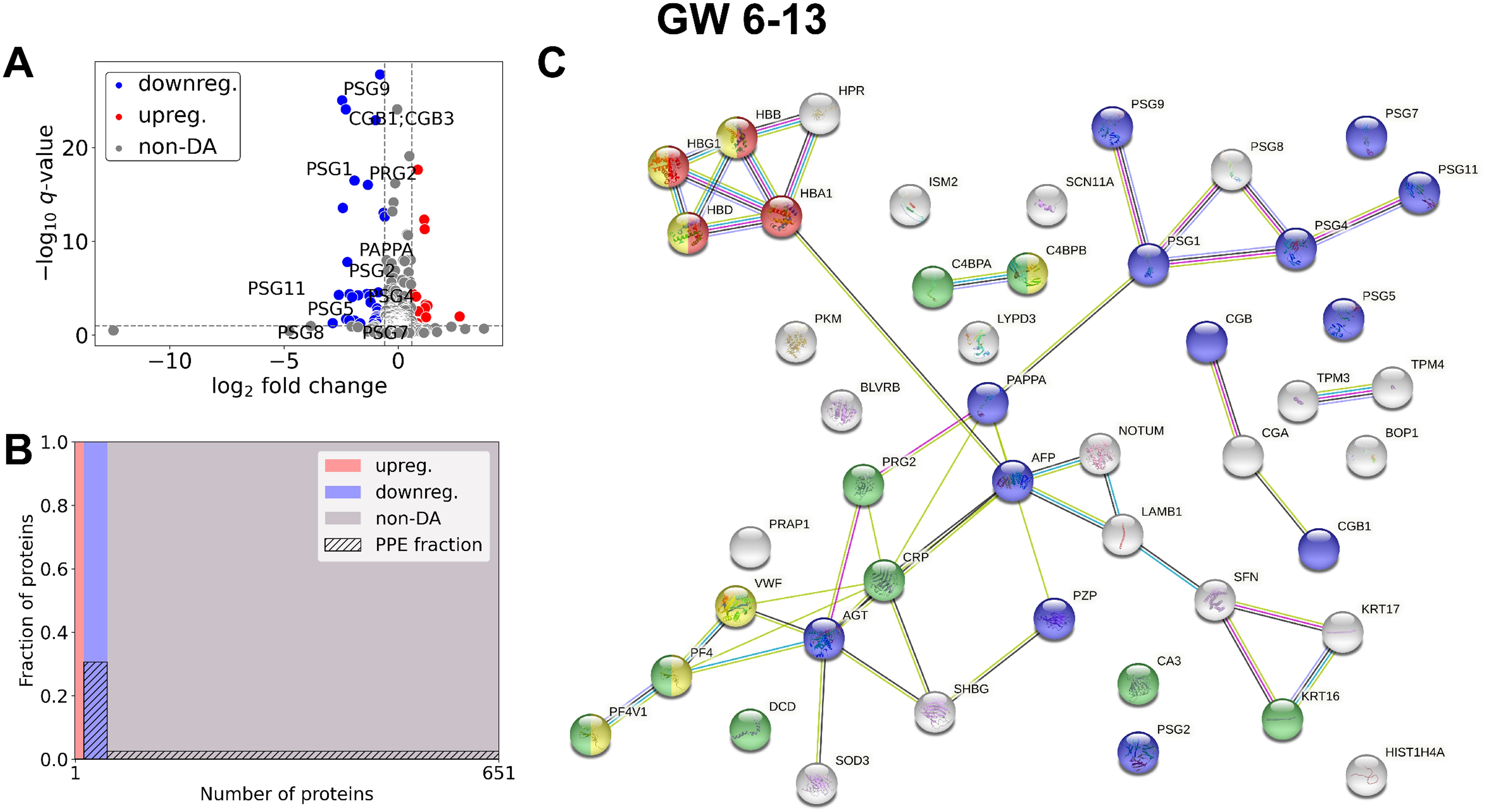
Figure 2. Comparison of RPL and control samples in the whole gestational age range. (A) Changes in protein intensities among RPL (GW 6–13) and control samples (GW 6–13) are represented using a volcano plot. Downregulated proteins are shown in blue, upregulated proteins are marked in red, and non-DA proteins are shown as grey dots. Gene IDs of PPE DA proteins are shown as well. (B) Mosaic plot shows the enrichment of PPE proteins (striped part). Upregulated proteins are marked red, downregulated proteins blue, and non-DA proteins grey. (C) The interaction network of DA proteins and GO biological process analysis was performed using STRING. On the figure components of selected GO terms are marked with different colors: ‘multi-organism reproductive process’ (blue), components of ‘oxygen transport’ (red), ‘response to other organism’ (green), and ‘blood coagulation’ (yellow). DA, differentially abundant; GO, Gene Ontology; GW, gestational weeks; PPE, predominantly placenta-expressed; RPL, recurrent pregnancy loss.
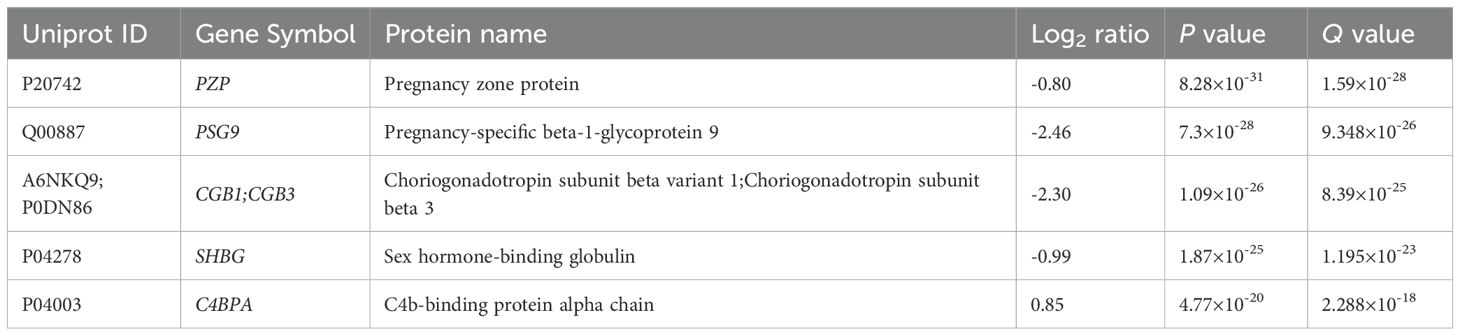
Table 2. Top DA proteins in RPL during GW 6-13.
In early RPL (Figure 3, Table 3, Supplementary Tables 2, 3), 40 DA proteins were identified (top 3 down: SHBG, CGB, CGA; top 3 up: C4BPA, SAMP, C4BPB), 32 were downregulated and 8 were upregulated. Among these, eight PPE proteins were found (enrichment factor: 5.01, p=6.91×10-5). In early RPL, also all PPE proteins were downregulated (enrichment factor: 6.26, p=1.16×10-5) and the fold changes of these proteins were also the lowest with one exception. Placental (3/14), as well as immune function (7/14) -related biological processes were among the top enriched GO terms.
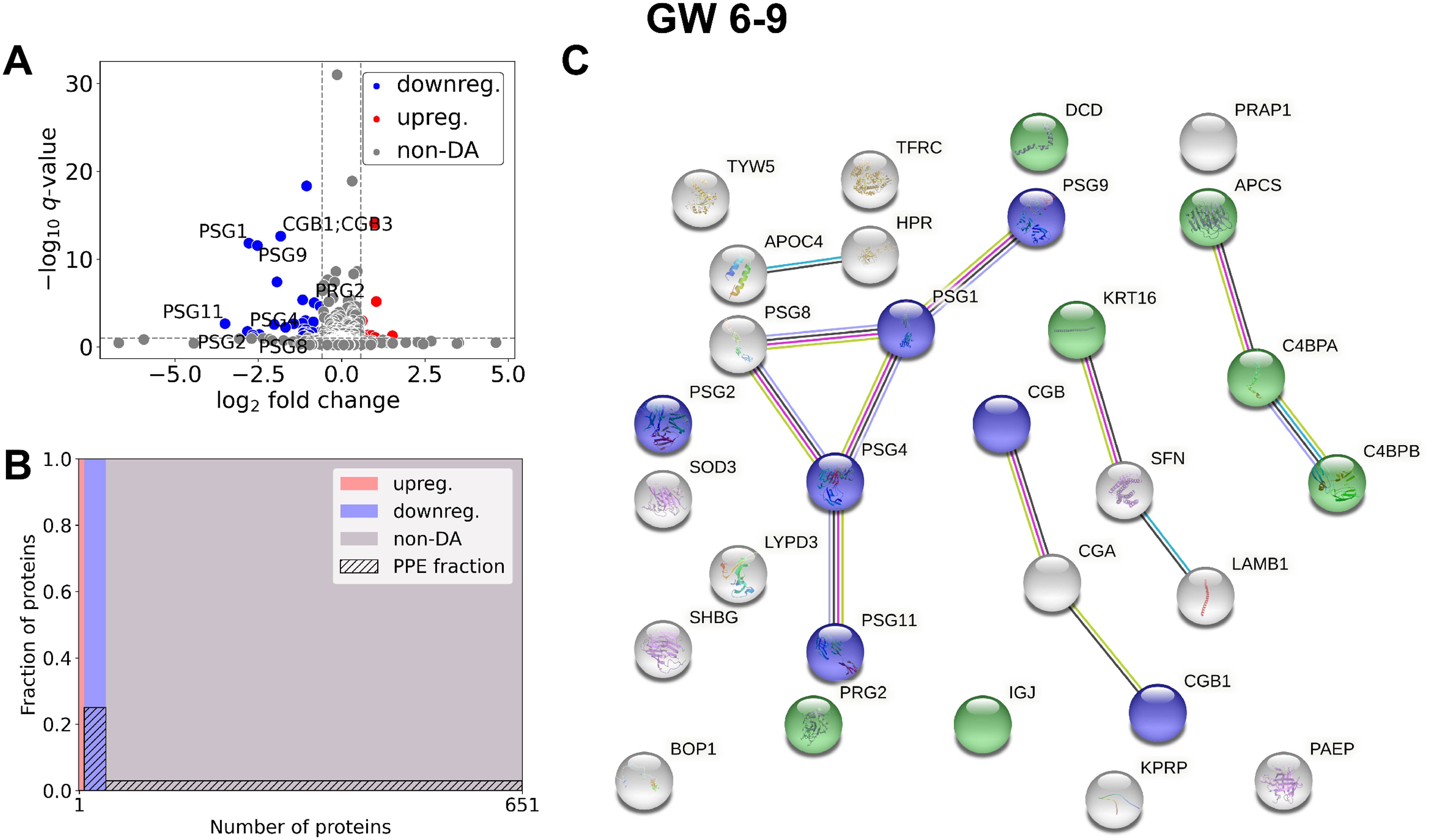
Figure 3. Comparison of RPL and control samples in the early period. (A) Changes in protein intensities among early RPL (GW 6–9) and early control samples (GW 6–9) are represented using a volcano plot. Downregulated proteins are shown in blue, upregulated proteins are marked in red, and non-DA proteins are shown as grey dots. Gene IDs of PPE DA proteins are shown as well. (B) Mosaic plot shows the enrichment of PPE proteins (striped part). Upregulated proteins are marked red, downregulated proteins blue, and non-DA proteins grey. (C) On the interaction networks of DA proteins components of selected GO terms are marked with different colors: ‘multi-organism reproductive process’ (blue) and ‘defense response to other organism’ (green). DA, differentially abundant; GO, Gene Ontology; GW, gestational weeks; PPE, predominantly placenta-expressed; RPL, recurrent pregnancy loss.
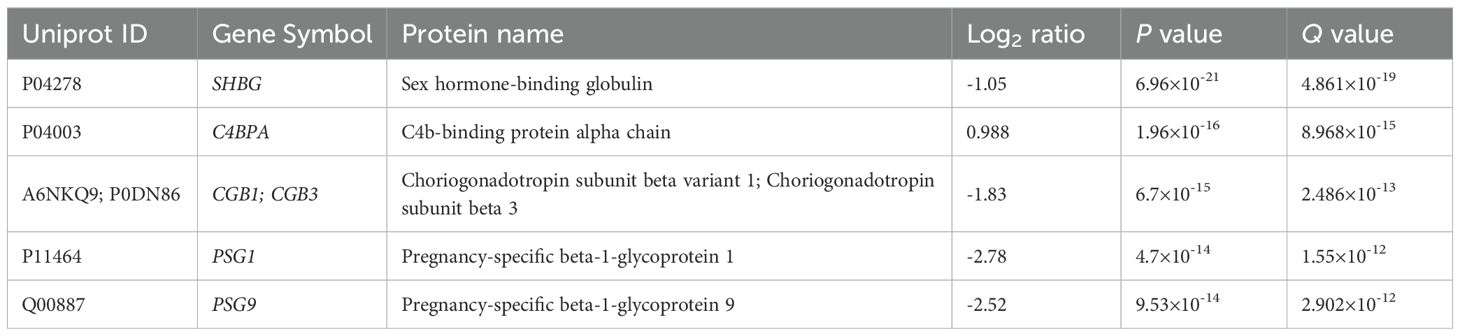
Table 3. Top DA proteins in RPL during GW 6-9.
In late RPL (Figure 4, Table 4, Supplementary Tables 2, 3) 90 DA proteins were found (top 3 down: PZP, PAPPA, PSG9; top 3 up: THBS1, ECM, HBB), among them 51 were downregulated and 39 were upregulated. Ten PPE proteins were found (enrichment factor: 2.78, p=1.25×10-3). In late RPL, similar to early RPL, all of the PPE proteins were downregulated (enrichment factor: 4.91, p=7.36×10-6), and the fold changes of these proteins were also among the lowest ones with two exceptions. The top enriched GO terms included ‘placental function’ (4/206), ‘oxidative processes’ (8/206), ‘immune function’ (55/206), ‘blood coagulation’ (6/206), ‘angiogenesis’ (3/206), ‘cell migration’ (9/206), as well as ‘blood circulation’ (6/206) -related biological processes.
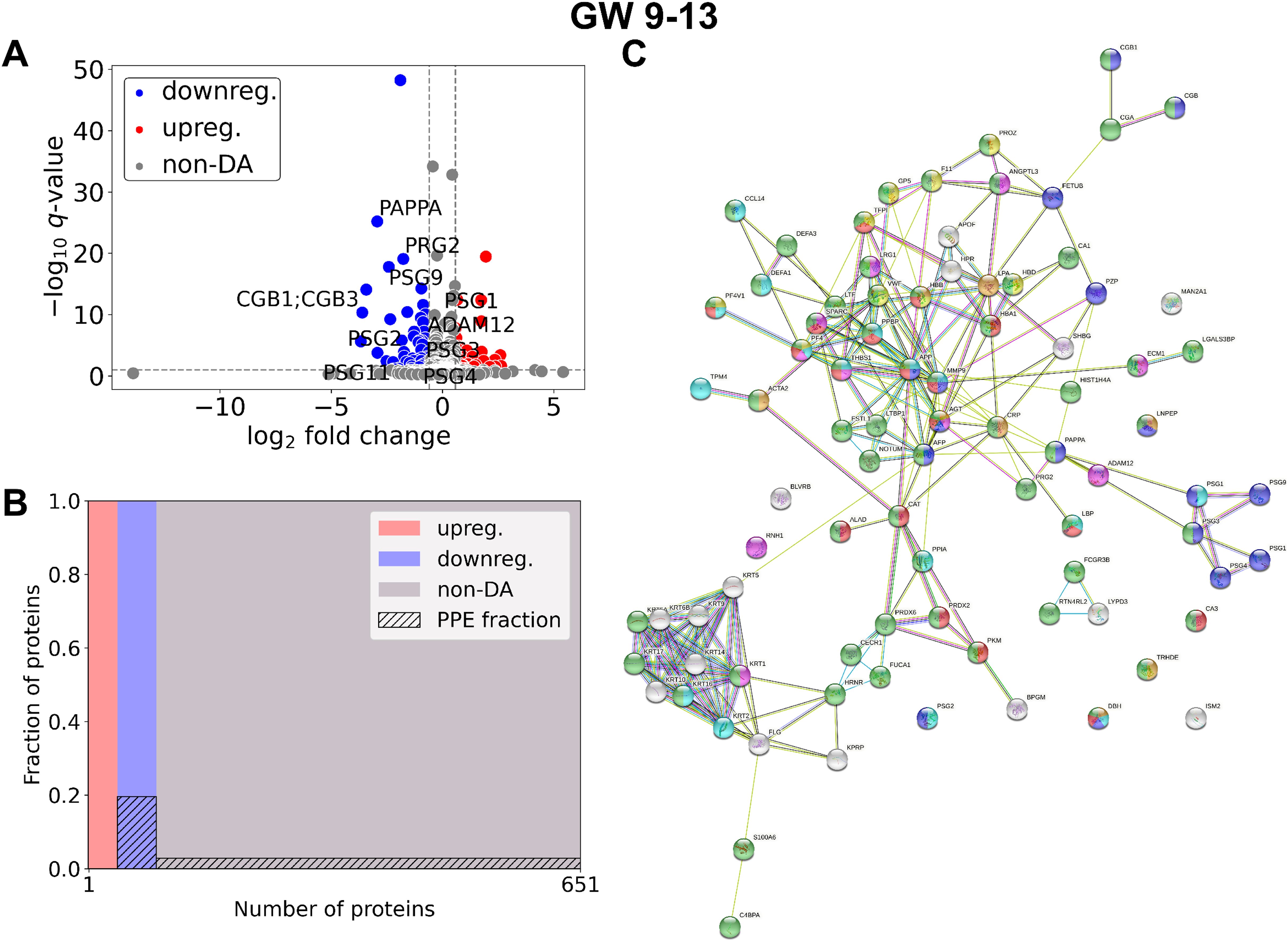
Figure 4. Comparison of RPL and control samples in the late period. (A) Changes in protein intensities among late RPL (GW 9–13) and late control samples (GW 9–13) are represented using a volcano plot. Downregulated proteins are shown in blue, upregulated proteins are marked in red, and non-DA proteins are shown as grey dots. The Gene ID of PPE DA proteins is shown as well. (B) Mosaic plot shows the enrichment of PPE proteins (striped part). Upregulated proteins are marked red, downregulated proteins blue, and non-DA proteins grey. (C) On the interaction networks of DA proteins components of selected GO terms are marked with different colors: ‘reproductive process’ (blue), components of ‘response to oxygen-containing compound’ (red), ‘response to stimulus’ (green), ‘blood coagulation’ (yellow), ‘regulation of angiogenesis’ (purple), ‘movement of cell or subcellular component’ (light blue), and ‘blood circulation’ (orange). DA, differentially abundant; GO, Gene Ontology; GW, gestational weeks; PPE, predominantly placenta-expressed; RPL, recurrent pregnancy loss.
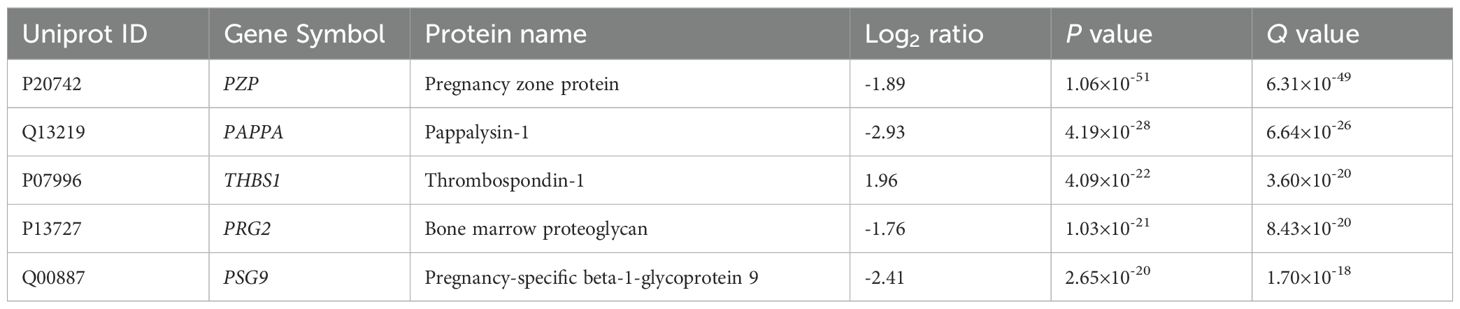
Table 4. Top DA proteins in RPL during GW 9-13.
Among DA proteins (Figure 5, Supplementary Tables 2, 3), 15 were identified in both early and late RPL, including seven PPE proteins (enrichment factor: 4.88, p=3.70×10-5). 25 proteins (one PPE, non-significant) were found only in early RPL, and 75 (three PPE, non-significant) only in late RPL. Importantly, among DA proteins identified in both early and late RPL, only ‘placental function’-related biological processes (3/4) were enriched.
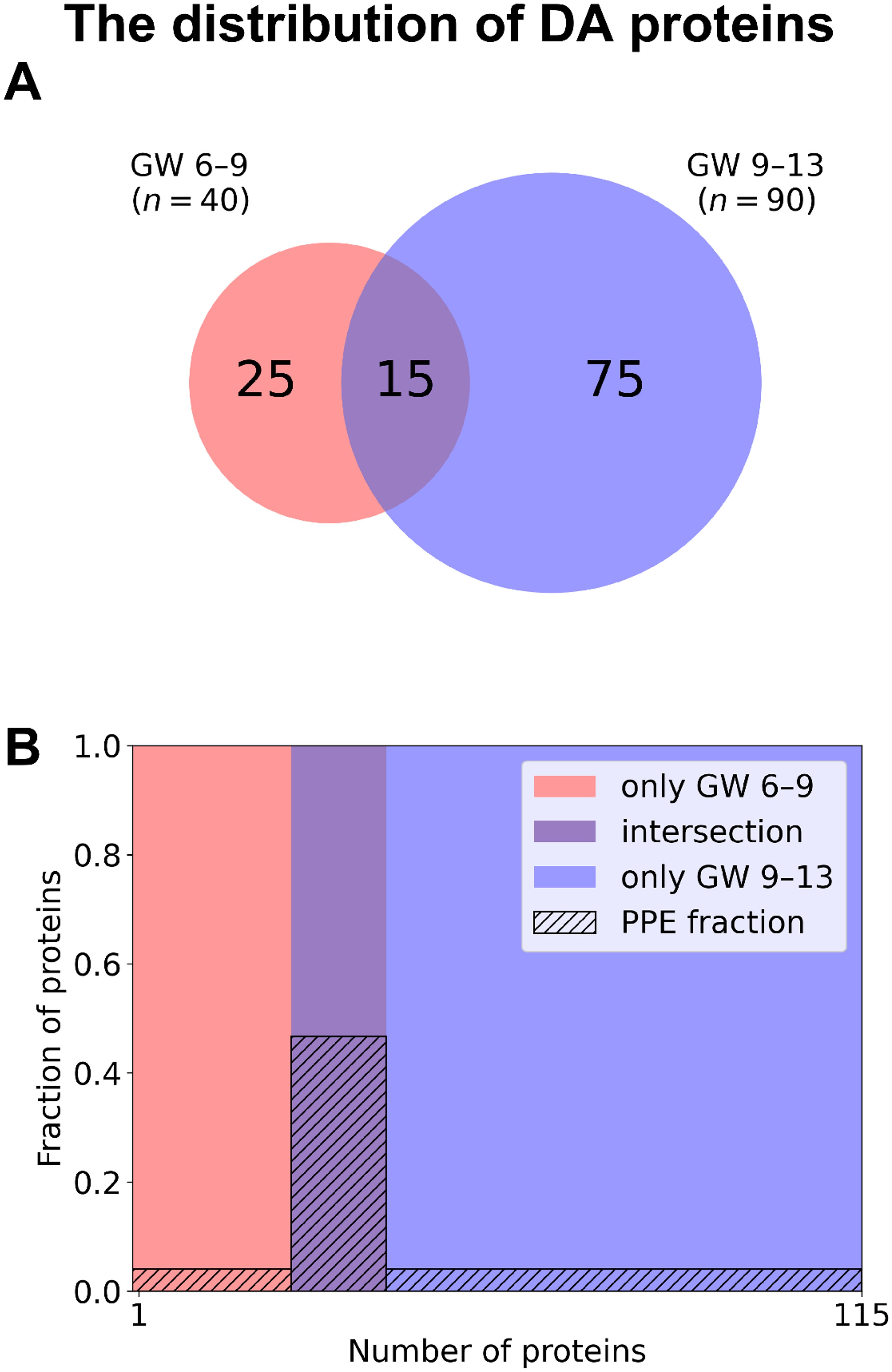
Figure 5. The distribution of DA proteins among early and late samples. (A) The number of DA proteins is displayed on a Venn diagram for early RPL (red, GW 6–9) and for late RPL (blue, GW 9–13) separately, as well as for both sub-groups (purple). (B) Mosaic plot shows the enrichment of PPE proteins (striped part). DA proteins only in early RPL are shown in red, DA proteins only in late RPL in blue, and common DA proteins in purple. DA, differentially abundant; GW, gestational weeks; PPE, predominantly placenta-expressed; RPL, recurrent pregnancy loss.
The discriminative power of plasma proteome and DA proteins with respect to study groups
Principal Component Analysis (PCA) was performed to identify the variance of data in the different sample groups (RPL and control groups) based on all proteins. Principal components 1, 2, and 3 explained 31.7% of the total variance, and little separation between sample groups could be observed (data not shown).
Classification performances were characterized by calculating sensitivity, specificity, and ROC curves for each DA protein in the different gestational age ranges. Scatter plots, box plots, and ROC curves of the best candidate proteins (whole gestational age range: CGA, early RPL: PAEP, late RPL: PAPPA) are presented in Figure 6, respectively.
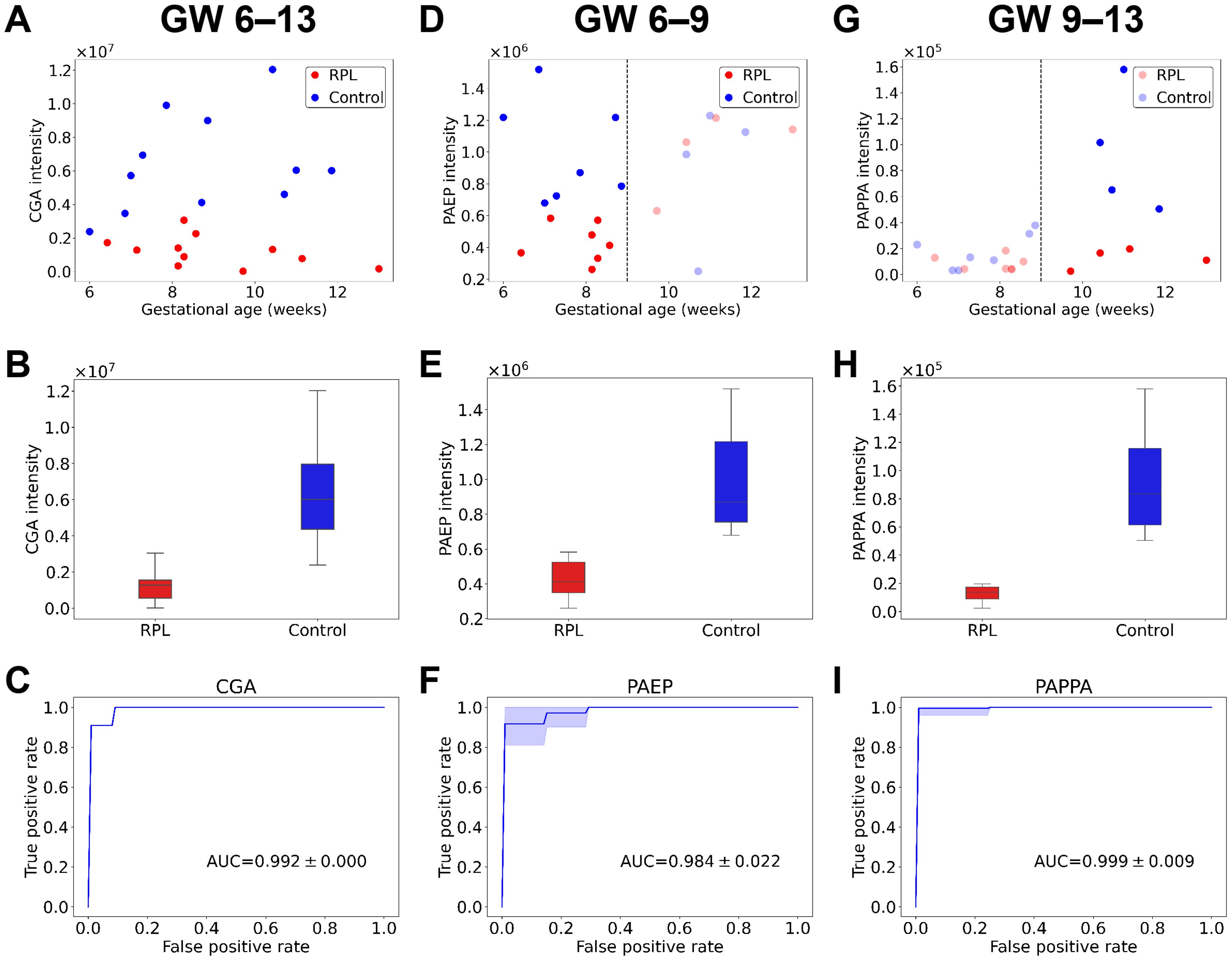
Figure 6. Protein intensities and classification performances of the best biomarker candidates for RPL. Scatter plots show protein intensities vs. gestational age for the best biomarker candidates in the whole gestational age range (A, GW 6–13), for early RPL (D, GW 6–9), and late RPL (G, GW 9–13). Box plots represent the intensities of the best biomarker candidates in the whole gestational age range (B), for early RPL (E), and late RPL (H). ROC curves represent classification performances of the best biomarker candidates in the whole gestational age range (C), for early RPL (F), and late RPL (I). On the plots, the RPL group is marked with red and the control group with blue. AUC, area under the curve; GW, gestational weeks; ROC, receiver operating characteristic; RPL, recurrent pregnancy loss.
Validation of mass spectrometry results with immunoassays
The concentration of CGB and PAPPA was also determined using the same immunoassay as in our previous study (88). In concordance with the proteomic findings, similar trends in biomarker levels were observed in the validation cohort. Both CGB and PAPPA showed significantly lower concentrations in the RPL group compared to the control group, reinforcing the proteomic results (Figure 7).
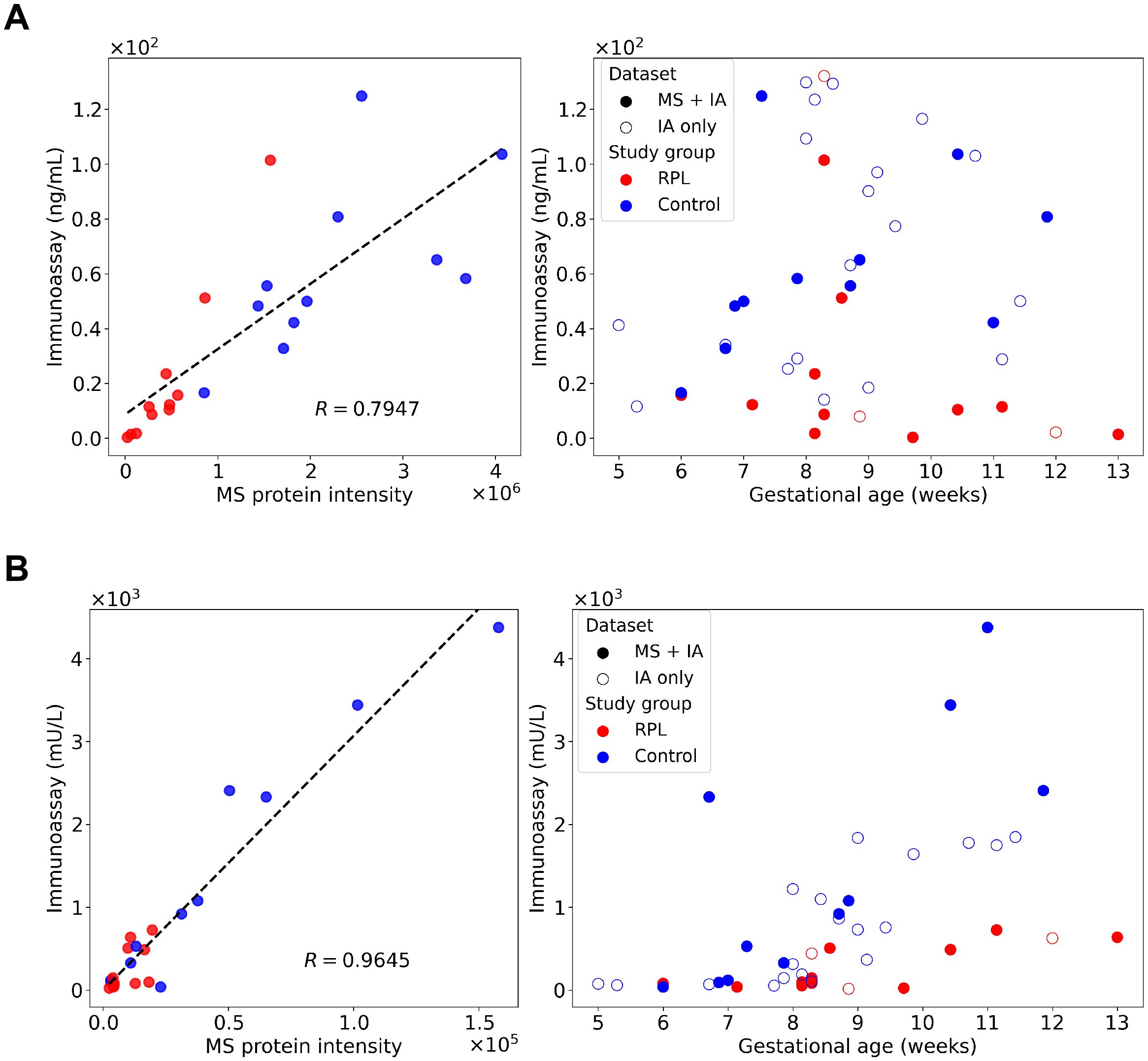
Figure 7. Correlation analysis between immunoassay and mass spectrometry (MS) results and validation of proteomic findings. Concentration of CGB (A) and PAPPA (B) were determined using immunoassay in our previous study (88) and re-evaluated to validate the proteomic results. Correlation between the immunoassay concentration and MS relative intensity results were calculated for both proteins for the identical samples (RPL, n=11; and control group, n=11). Pearson correlation coefficients (R) were 0.7947 for CGB and 0.9645 for PAPPA, respectively. Immunoassay concentrations were plotted against gestational age, with overlapping and distinct samples are denoted with different symbols. CGB, β-human chorionic gonadotropin; MS, mass spectrometry; R, Pearson correlation coefficient; PAPPA, pregnancy-associated plasma protein A; RPL, recurrent pregnancy loss.
The correlation analysis between immunoassay and mass spectrometry data revealed a good correlation (Figure 7) on the overlapping cohort as the Pearson correlation coefficient (R) was 0.795 for CGB and 0.965 for PAPPA.
Discussion
Principal findings of this study
1) Sixty DA proteins had gestational age-related concentration changes in controls. 2) Fifty DA proteins were dysregulated in RPL compared to control samples. 3) Forty DA proteins were found in early RPL and ninety in late RPL. 4) PPE proteins were enriched among DA proteins and showed to be the most downregulated during both periods in RPL. 5) Enriched GO terms in early RPL included ‘immune regulation’, ‘complement activation’, and ‘female pregnancy’ while ‘placental function’, ‘oxidative processes’, ‘immune function’, and ‘blood coagulation’ related biological processes were enriched in late RPL among others. 6) Several biomarker candidates for early and late RPL were identified and two were validated with immunoassay.
Plasma proteome profiling and analysis of PPE proteins in the maternal blood using mass spectrometry
The fetal support and maintenance of pregnancy by providing nutrition, immunological and hormonal regulation as well as gas and waste exchange, are the most important roles of the placenta (98, 99). It has been known for more than half century that protein products of the placenta get into the maternal circulation, and their circulating concentration changes may well reflect changes in placental functions with gestational age in normal pregnancies as well as dysregulation of placental functions in pregnancy complications. Therefore, placental proteins turned out to be of great significance for the monitoring of placental functions through non-invasive sampling of maternal blood (i.e. liquid biopsy) (100–103).
We have recently defined PPE proteins (90, 91), which play a central role in monitoring placental functions in the maternal blood. Also, our previous results showed that placental dysfunction is often associated with the altered expression of PPE proteins (104–113), therefore, special attention should be paid to these proteins in the prediction and treatment of pregnancy complications such as RPL. Recently, with immunoassay, we examined three of these PPE proteins (free β-hCG, PAPP-A, and PlGF) and confirmed their diagnostic significance in RPL (88). Utilizing the recent developments in the field of proteomics and mass spectrometry that enable the deep proteomic investigation of different tissues (114), including the placenta (115), and the liquid biopsy of these tissues via minimal invasive blood draw (116, 117), here we aimed to explore the proteome-wide changes in maternal plasma in RPL. This current analysis had a special attention to PPE proteins in an attempt to provide relevant information about the developmental status and functional condition of the placenta and to support the early detection and/or prediction of failing placental functions.
Changing maternal plasma proteomic signature in normal pregnancy during the first trimester
The placenta undergoes rapid development in the first weeks of pregnancy (118), however, the lack of its connection to the maternal circulation may hinder placental liquid biopsy at the beginning of the pregnancy (119, 120). Indeed, villous placental vessels and circulation start to develop at around gestational weeks 5–6 (121, 122), while the intervillous circulation is only being established after around the 9th gestational week. This is a protective mechanism for the developing embryo and placenta to defend their tissues from excessive and harmful oxygen exposure (95, 123). Therefore, placental products enter into the maternal circulation in much larger quantities only after the opening of the intervillous blood space (124).
To account for this physiological change in placental circulation during the first trimester, we investigated the proteomic changes before and after the supposed start of the intervillous circulation while considering differences in maternal environmental, metabolic, and immunological characteristics. In control pregnancies, our study identified key DA proteins between gestational weeks 6–9 and 9–13, shedding light on the physiological adaptations during early pregnancy. There were 29 proteins with elevated concentrations in gestational weeks 6–9 compared to 9–13. These included FN1 (fibronectin 1) (125), ECM1 (extracellular matrix protein 1) (126), and COMP (cartilage oligomeric matrix protein) (127), which are all associated with extracellular matrix organization, cell adhesion, and tissue remodeling. Their dropping concentration with gestational age suggests a shift from initial implantation and rapid tissue expansion toward the established placental milieu. Other proteins with decreasing concentration over gestational age included PF4 (platelet factor 4), which is a key regulator of vascular homeostasis (128, 129), highlighting the high demands for vascular remodeling during the early phase of placentation. Furthermore, IGFBP2 (insulin-like growth factor-binding protein 2), which regulates the bioavailability of insulin-like growth factors crucial for cellular proliferation and differentiation (130), had a decreasing concentration with advancing gestational age. Several platelet-related proteins (e.g., PF4, PF4V1, PPBP alias CXCL7, THBS1, and VWF) were also found to decrease over gestational age. This may be due to a decrease in mean platelet count gradually throughout gestation; otherwise, upon activation, maternal platelets would provide a source of proinflammatory mediators in the intervillous space of the placenta (129).
Conversely, 31 proteins demonstrated increased concentrations in gestational weeks 9–13 compared to 6–9. These included PZP (pregnancy zone protein) which supports the transition towards immune tolerance (131, 132), as well as PAPPA which increases the bioavailability of insulin-like growth factors, which are crucial for fetal growth and placental development. Also, ADAM12 (a disintegrin and metalloproteinase domain-containing protein 12) is a well-known regulator of trophoblast invasion and extracellular matrix (ECM) remodeling (133). Proteins such as PRG2 (proteoglycan 2, pro eosinophil major basic protein), PSG1, PSG3, and PSG9 (pregnancy-specific beta-1 glycoprotein 1, 3, and 9) are key immunomodulators that help to establish maternal immune tolerance towards the fetus (134). Many of these proteins are also well-known to have increasing concentrations in maternal blood during this timeframe of pregnancy which validates our study (112, 116). These findings provide insights into the dynamic proteomic changes occurring in early pregnancy. The elevated concentrations of ECM-related proteins (FN1, ECM1, COMP) and vascular regulators (PF4) in the earlier phase suggest the importance of ECM reorganization, tissue remodeling, and vascular homeostasis essential for implantation and initial placental formation. ECM proteins not only provide structural scaffolding for invading trophoblasts but also participate in immune regulation by modulating leukocyte adhesion and migration. FN1, for instance, can interact with integrins and Toll-like receptors on maternal immune cells, influencing macrophage activation and cytokine secretion (135).
In contrast, the increased abundance of proteins after week 9, involved in immunomodulation (PZP, PSG1, PSG9, PRG2) and placental development (PAPPA, ADAM12), aligns with the physiological progression toward immune tolerance and increased placental growth.
PZP, a pregnancy-associated protease inhibitor, has been shown to suppress T-cell proliferation and promote a tolerogenic environment by forming complexes with glycodelin-A, and leading to reduced IL-2 production (131).
Pregnancy-specific glycoproteins (PSG1, PSG3, PSG9) are produced by the syncytiotrophoblast (136) and act on maternal monocytes, dendritic cells, and NK cells to induce anti-inflammatory cytokines (e.g., IL-10) and activate TGF-β1 and TGF-β2 (134, 137), which together promote the expansion of regulatory T cells (Tregs) and dampen cytotoxic responses (138).
ADAM12 plays a dual role, facilitating trophoblast invasion through ECM remodeling while also influencing the availability of cytokines and growth factors within the decidua hence contributes to the pathogenesis of tissue inflammation and TH1 differentiation (139). Altogether, these proteins help in reprogramming the maternal immune system, essential for maintaining maternal-fetal tolerance as the intervillous circulation opens and the fetal-derived placenta becomes more exposed to maternal immunity.
Immune dysregulation and impaired maternal-fetal tolerance specific for early RPL
In early RPL cases, we identified the predominant changes in the immune-related proteome. Several proteins involved in immune tolerance and complement regulation were downregulated in early RPL, including C4BPA (complement component 4 binding protein alpha) and C4BPB (complement component 4 binding protein beta), which play crucial roles in modulating complement activation and preventing excessive immune responses at the maternal-fetal interface (140). Additionally, PAEP (progestagen-associated endometrial protein, also known as glycodelin) (141), a critical modulator of maternal immune tolerance that suppresses T-cell activation and supports embryo implantation (142, 143), showed decreased levels in early RPL cases. In addition, GO enrichment analysis further highlighted the disrupted biological processes in early RPL, with enriched terms predominantly linked to ‘immune regulation’, ‘complement activation’, and ‘female pregnancy’. Key terms included ‘multi-organism processes’ (reflecting maternal-fetal interactions), ‘regulation of opsonization’, and ‘negative regulation of complement activation’. These findings may reflect impaired regulation of immune interactions and tolerance mechanisms at the maternal-fetal interface in RPL.
Placental dysregulation specific for late RPL
The proteomic profile in late RPL revealed an increased number of DA proteins associated with angiogenesis and inflammatory responses. ANGPTL3 (angiopoietin-related protein 3), a regulator of lipid metabolism (144), has been implicated in modulating angiogenesis (145) and may contribute to altered placental vascularization. PF4, a chemokine released from activated platelets, is involved in immune cell recruitment and inflammatory signaling (146, 147), and its elevation may reflect heightened immune activation or vascular stress in the decidua. In addition, the expression level of the PZP, a known immunomodulator that neutralizes immune activation (148) was downregulated, similar to anti-microbial peptides, important components of the innate immune system at the maternal-fetal interface (149). Similarly, the intensity of the fetal growth regulator PAPPA (150, 151), was also reduced, suggesting an impairment in placental development function. Furthermore, increased levels of hemoglobin subunits (HBA, HBB, HBD) in late RPL compared may result from hyperviscosity/hemoconcentration, which subsequently increases the risk of miscarriage (152, 153). Interestingly, in these studies, low concentration of hemoglobin in the preconception or early pregnancy period, resulting in anemia, also increased the risk of miscarriage (152, 153).
GO analysis in late RPL revealed enriched terms including ‘immune response’, ‘neutrophil degranulation’, ‘platelet activation’, and ‘inflammatory response’, further emphasizing an excessive and dysregulated immune activation, which may contribute to trophoblast stress and placental dysfunction. The presence of oxidative stress-related terms such as ‘cellular oxidant detoxification’, ‘response to reactive oxygen species’, and ‘positive regulation of reactive oxygen species metabolic process’ further supports the notion that placental oxidative stress (123) is a major contributing factor to late RPL. These findings collectively suggest that late RPL is characterized by a pathological environment marked by impaired angiogenesis, dysregulated immune responses, substantial oxidative stress of the trophoblast and placental dysfunction.
Shared and distinct characteristics in maternal proteome changes in early and late RPL
Among the shared characteristics, we found consistently low levels of CGA across both early and late RPL, indicating a persistent defect in placental function. The pregnancy-specific glycoproteins, such as PSG1, PSG2, PSG4, PSG9, and PSG11, which are essential for immune tolerance by inducing anti-inflammatory cytokines like IL-10 and promoting trophoblast survival (134, 154, 155), were also significantly reduced in both RPL groups. Additionally, we also observed reduced levels of sex hormone-binding globulin (SHBG) in both RPL groups, a key regulator of sex steroid bioavailability. Low SHBG level has been previously associated with early pregnancy loss (67, 156) and not only reflects hormonal or metabolic dysregulation affecting endometrial receptivity and placental development, but may also alter SHBG-mediated estradiol signaling in lymphocytes, potentially disrupting maternal immune adaptation during early pregnancy (157).
However, other findings indicate that immune dysfunction in RPL follows a gestational age-dependent pattern. Early RPL appears to be associated with maternal immune maladaptation characterized by defective complement regulation and insufficient maternal immune tolerance, whereas late RPL shows stronger associations with dysregulation of placental immune function, vascular remodeling, and inflammation. Despite these differences, the persistently low levels of CGA and PSGs across both gestational age groups suggest that placental dysfunction remains a central feature of RPL throughout pregnancy. In addition, in late RPL we found signs of reduced trophoblast invasion and inadequate spiral artery remodeling, which were implicated already in miscarriages (158). The consequently altered placental circulation characterized by turbulent jets damage the villous structure that can also be seen with ultrasound (159). These rheological and mechanical changes also have indirect molecular effects (160), as evidenced by oxidative stress triggered pathological processes (90). Our findings are also consistent with oxidative stress-related molecular changes in the placenta, as reflected in the maternal circulation.
In summary, our results highlight distinct shifts in the maternal immune proteome before and after the establishment of placental circulation. The distinct and overlapping enrichment patterns across gestational ages underscore the dynamic interplay between immune adaptation, placental function, and vascular remodeling during pregnancy, offering insight into the molecular pathways underlying early and late RPL. While these proteomics results offer valuable biological insights and identify potential diagnostic biomarkers, also validated by immunoassay in a small sample, they are exploratory in nature and require further independent validation in larger, external cohorts to confirm their robustness and clinical relevance.
Strengths and limitations of the study
This study has several strengths, including the use of next-generation proteomics with stringent clinical definitions and homogenous patient groups, enabling the identification of gestational age-specific molecular pathways in RPL. Robust methodologies, such as depletion of abundant proteins, mass spectrometry with stringent FDR controls, and validation through immunoassay correlation, enhance the reliability of findings. Indeed, the correlation coefficient between the mass spectrometry and immunoassay results for PAPPA was outstandingly high, while for CGB it was slightly lower, however, even this is entirely sufficient and consistent with the literature (161).
The limitations of the study include the timing of sample collection at the point of miscarriage, which prevents assessing the predictive value of biomarkers before clinical symptoms. The small sample size which inherently reduces statistical power and increases the risk of both type I (false positive) and type II (false negative) errors, and the use of elective termination pregnancies as controls may limit the generalizability of results, while other potential confounders, such as environmental or lifestyle factors, were not addressed. While our use of multiple testing correction, and cross-validation methods aimed to mitigate these risks, the possibility of overfitting and spurious associations cannot be fully excluded. Additionally, the RPL population in our study primarily consisted of women with two or three prior losses, which represents the lower end of the clinical spectrum. It remains to be investigated whether proteomic alterations are more pronounced or distinct in cases with higher-order losses, which may reflect more severe or recurrent pathological processes.
Since one potential confounder that could influence protein levels is maternal age, which was higher in the RPL group, we investigated the potential effect of this confounder by looking into the data focusing on age-related changes in the proteome. It has been described that aging-related changes in the plasma proteome generally occur over much longer timeframes (i.e., decades) and typically result in modest fold changes (162). For instance, in the case of GDF15 - showing the strongest known association between protein abundance and aging from 1301 proteins - the log2 fold change per year of age (β = 0.018) corresponds to only ~1.065-fold change over five years (163). This is minimal compared to the ≥1.5-fold changes observed for DA proteins in our study. Furthermore, correlation analysis between maternal age and proteomic data revealed no significant associations after correcting for multiple comparisons. These findings suggest that maternal age is unlikely to be the primary driver of the observed group differences.
Conclusions
This study provides critical insights into the molecular pathways underlying RPL by leveraging next-generation proteomics and a gestational age-specific approach. We identified distinct and overlapping DA proteins in early and late RPL, highlighting key biological processes such as ‘placental function’, ‘immune modulation’, ‘oxidative stress’, and ‘vascular remodeling’. Several novel biomarkers showed outstanding discriminative performance and may facilitate a better understanding of RPL pathogenesis. The findings emphasize the importance of gestational age in RPL pathogenesis, with early losses mainly linked to immune dysregulation and placental establishment and late losses predominantly associated with oxidative stress and circulatory changes. The study also underscores the significance of placenta-specific proteins as potential liquid biopsy biomarkers for RPL. While the results demonstrate the promise of proteomics in advancing the understanding and diagnosis of RPL, future research with larger cohorts and longitudinal designs will be essential to validate these biomarkers and to test drugs for the modulation of the identified disease pathways and the prevention of RPL. Our findings highlight the importance of the maternal immune system in maintaining successful pregnancy and suggest that targeting immune pathways may offer novel therapeutic approaches for RPL.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving humans were approved by Health Science Board of Hungary (ETT-TUKEB 4834-0/2011-1018EKU). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
ET: Formal analysis, Writing – original draft, Writing – review & editing, Investigation, Validation. MP: Formal analysis, Writing – original draft, Writing – review & editing, Investigation, Validation. DG: Formal analysis, Methodology, Visualization, Writing – original draft, Writing – review & editing. OO: Formal analysis, Writing – original draft, Writing – review & editing. EF: Formal analysis, Writing – original draft, Writing – review & editing. AB: Formal analysis, Writing – original draft, Writing – review & editing. CE: Methodology, Writing – original draft, Writing – review & editing, Investigation. MB: Methodology, Visualization, Writing – original draft, Writing – review & editing, Investigation. AS: Methodology, Visualization, Writing – original draft, Writing – review & editing. PH: Investigation, Writing – original draft, Writing – review & editing, Resources. LV: Validation, Writing – original draft, Writing – review & editing, Investigation. OT: Investigation, Writing – original draft, Writing – review & editing. SN: Investigation, Writing – original draft, Writing – review & editing. OR: Methodology, Writing – original draft, Writing – review & editing, Investigation. OE: Investigation, Writing – original draft, Writing – review & editing. ZP: Investigation, Writing – original draft, Writing – review & editing. NA: Investigation, Writing – original draft, Writing – review & editing. NGT: Conceptualization, Funding acquisition, Project administration, Supervision, Writing – original draft, Writing – review & editing, Resources.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by grants from the Hungarian Academy of Sciences (Momentum LP2014-7/2014 to NGT, Premium postdoc 2019-436 to AB, Young Researcher to ET), the Hungarian Research Network (HUN-REN SA-83/2021 and HUN-REN TECH-2024-042 grant to NGT), and the Ministry of Innovation and Technology of Hungary from the National Research, Development and Innovation Fund, financed under the OTKA-K128262 (to AS), 2024-2-1-1-EKÖP-2024-00004 (to MP), FIEK-16-1-2016-0005, OTKA-K124862, 2020-1.1.2-PIACI-KFI-2021-00273, 2024-1.2.3-HU-RIZONT-2024-00062 and 2019-2.1.7-ERANET-2020-00014 (all to NGT) funding schemes, the latter under the frame of the European Union ERA PerMed (2020-346 grant).
Conflict of interest
Part of the data was submitted by DG and NGT as a patent application in 2025 to describe biomarkers for recurrent pregnancy loss. Other authors declare no conflict of interest.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1621168/full#supplementary-material
References
1. Dugas C and Slane VH. Miscarriage. In: StatPearls. StatPearls Publishing LLC, Treasure Island (FL (2020).
2. WHO: recommended definitions, terminology and format for statistical tables related to the perinatal period and use of a new certificate for cause of perinatal deaths. Modifications recommended by FIGO as amended October 14, 1976. Acta Obstet Gynecol Scand. (1977) 56:247–53. doi: 10.3109/00016347709162009
3. Ammon Avalos L, Galindo C, and Li DK. A systematic review to calculate background miscarriage rates using life table analysis. Birth Defects Res A Clin Mol Teratol. (2012) 94:417–23. doi: 10.1002/bdra.23014
4. Larsen EC, Christiansen OB, Kolte AM, and Macklon N. New insights into mechanisms behind miscarriage. BMC Med. (2013) 11:154. doi: 10.1186/1741-7015-11-154
5. Savitz DA, Hertz-Picciotto I, Poole C, and Olshan AF. Epidemiologic measures of the course and outcome of pregnancy. Epidemiol Rev. (2002) 24:91–101. doi: 10.1093/epirev/mxf006
6. Ford HB and Schust DJ. Recurrent pregnancy loss: etiology, diagnosis, and therapy. Rev Obstet Gynecol. (2009) 2:76–83.
7. Zinaman MJ, Clegg ED, Brown CC, O’Connor J, and Selevan SG. Estimates of human fertility and pregnancy loss. Fertil Steril. (1996) 65:503–9. doi: 10.1016/S0015-0282(16)58144-8
8. Ellish NJ, Saboda K, O’Connor J, Nasca PC, Stanek EJ, and Boyle C. A prospective study of early pregnancy loss. Hum Reprod. (1996) 11:406–12. doi: 10.1093/HUMREP/11.2.406
9. Wilcox AJ, Weinberg CR, O’Connor JF, Baird DD, Schlatterer JP, Canfield RE, et al. Incidence of early loss of pregnancy. New Engl J Med. (1988) 319:189–94. doi: 10.1056/NEJM198807283190401
10. Rai R and Regan L. Recurrent miscarriage. Lancet. (2006) 368:601–11. doi: 10.1016/S0140-6736(06)69204-0
11. Chard T. 11 Frequency of implantation and early pregnancy loss in natural cycles. Baillière’s Clin Obstet Gynaecol. (1991) 5:179–89. doi: 10.1016/S0950-3552(05)80077-X
12. Macklon NS, Geraedts JP, and Fauser BC. Conception to ongoing pregnancy: the ‘black box’ of early pregnancy loss. Hum Reprod Update. (2002) 8:333–43. doi: 10.1093/humupd/8.4.333
13. Brezina PR and Kutteh WH. Classic and cutting-edge strategies for the management of early pregnancy loss. Obstet Gynecol Clin North Am. (2014) 41:1–18. doi: 10.1016/j.ogc.2013.10.011
14. Regan L, Braude PR, and Trembath PL. Influence of past reproductive performance on risk of spontaneous abortion. Br Med J. (1989) 299:541. doi: 10.1136/bmj.299.6698.541
15. Knudsen UB, Hansen V, Juul S, and Secher NJ. Prognosis of a new pregnancy following previous spontaneous abortions. Eur J Obstet Gynecol Reprod Biol. (1991) 39:31–6. doi: 10.1016/0028-2243(91)90138-B
16. Vaiman D. Genetic regulation of recurrent spontaneous abortion in humans. Biomed J. (2015) 38:11–24. doi: 10.4103/2319-4170.133777
17. ACOG. ACOG practice bulletin. Management of recurrent pregnancy loss. Number 24, February 2001. (Replaces Technical Bulletin Number 212, September 1995). American College of Obstetricians and Gynecologists. Int J Gynecol Obstet. (2002) 78:179–90. doi: 10.1016/S0020-7292(02)00197-2
18. Li TC, Makris M, Tomsu M, Tuckerman E, and Laird S. Recurrent miscarriage: aetiology, management and prognosis. Hum Reprod Update. (2002) 8:463–81. doi: 10.1093/humupd/8.5.463
19. Clifford K, Rai R, Watson H, and Regan L. An informative protocol for the investigation of recurrent miscarriage: preliminary experience of 500 consecutive cases. Hum Reprod. (1994) 9:1328–32. doi: 10.1093/oxfordjournals.humrep.a138703
21. ASRM. Evaluation and treatment of recurrent pregnancy loss: a committee opinion. Fertil Steril. (2012) 98:1103–11. doi: 10.1016/j.fertnstert.2012.06.048
22. Risch HA, Weiss NS, Aileen CE, and Miller AB. Risk factors for spontaneous abortion and its recurrence. Am J Epidemiol. (1988) 128:420–30. doi: 10.1093/oxfordjournals.aje.a114982
23. van den Akker OBA. The psychological and social consequences of miscarriage. Expert Rev Obstet Gynecol. (2011) 6:295–304. doi: 10.1586/eog.11.14
24. Murphy S and Cacciatore J. The psychological, social, and economic impact of stillbirth on families. Semin Fetal Neonatal Med. (2017) 22:129–34. doi: 10.1016/j.siny.2017.02.002
25. Giorlandino C, Calugi G, Iaconianni L, Santoro ML, and Lippa A. Spermatozoa with chromosomal abnormalities may result in a higher rate of recurrent abortion. Fertil Steril. (1998) 70:576–7. doi: 10.1016/S0015-0282(98)00192-7
26. Rubio C, Simón C, Vidal F, Rodrigo L, Pehlivan T, Remohí J, et al. Chromosomal abnormalities and embryo development in recurrent miscarriage couples. Hum Reprod. (2003) 18:182–8. doi: 10.1093/humrep/deg015
27. Stirrat GM. Recurrent miscarriage II: clinical associations, causes, and management. Lancet. (1990) 336:728–33. doi: 10.1016/0140-6736(90)92215-4
28. Tulppala M, Palosuo T, Ramsay T, Miettinen A, Salonen R, and Ylikorkala O. A prospective study of 63 couples with a history of recurrent spontaneous abortion: contributing factors and outcome of subsequent pregnancies. Hum Reprod. (1993) 8:764–70. doi: 10.1093/oxfordjournals.humrep.a138137
29. Uehara S, Hashiyada M, Sato K, Sato Y, Fujimori K, and Okamura K. Preferential X-chromosome inactivation in women with idiopathic recurrent pregnancy loss. Fertil Steril. (2001) 76:908–14. doi: 10.1016/S0015-0282(01)02845-X
30. Aldrich CL, Stephenson MD, Karrison T, Odem RR, Branch DW, Scott JR, et al. HLA-G genotypes and pregnancy outcome in couples with unexplained recurrent miscarriage. Mol Hum Reprod. (2001) 7:1167–72. doi: 10.1093/molehr/7.12.1167
31. Grimbizis GF, Camus M, Tarlatzis BC, Bontis JN, and Devroey P. Clinical implications of uterine malformations and hysteroscopic treatment results. Hum Reprod Update. (2001) 7:161–74. doi: 10.1093/humupd/7.2.161
32. Salim R, Regan L, Woelfer B, Backos M, and Jurkovic D. A comparative study of the morphology of congenital uterine anomalies in women with and without a history of recurrent first trimester miscarriage. Hum Reprod. (2003) 18:162–6. doi: 10.1093/humrep/deg030
33. Homer HA, Li T-C, and Cooke ID. The septate uterus: a review of management and reproductive outcome. Fertil Steril. (2000) 73:1–14. doi: 10.1016/S0015-0282(99)00480-X
34. Clifford K, Flanagan AM, and Regan L. Endometrial CD56+ natural killer cells in women with recurrent miscarriage: a histomorphometric study. Hum Reprod. (1999) 14:2727–30. doi: 10.1093/humrep/14.11.2727
35. Quenby S, Bates M, Doig T, Brewster J, Lewis-Jones DI, Johnson PM, et al. Pre-implantation endometrial leukocytes in women with recurrent miscarriage. Hum Reprod. (1999) 14:2386–91. doi: 10.1093/humrep/14.9.2386
36. Lachapelle MH, Miron P, Hemmings R, and Roy DC. Endometrial T, B, and NK cells in patients with recurrent spontaneous abortion altered profile pregnancy outcome. J Immunol. (1996) 156:4027. doi: 10.4049/jimmunol.156.10.4027
37. Eblen AC, Gercel-Taylor C, Shields LBE, Sanfilippo JS, Nakajima ST, and Taylor DD. Alterations in humoral immune responses associated with recurrent pregnancy loss. Fertil Steril. (2000) 73:305–13. doi: 10.1016/S0015-0282(99)00505-1
38. Wilson R, Maclean MA, Jenkins C, Kinnane D, Mooney J, and Walker JJ. Abnormal immunoglobulin subclass patterns in women with a history of recurrent miscarriage. Fertil Steril. (2001) 76:915–7. doi: 10.1016/S0015-0282(01)02857-6
39. Makhseed M, Raghupathy R, Azizieh F, Omu A, Al-Shamali E, and Ashkanani L. Th1 and Th2 cytokine profiles in recurrent aborters with successful pregnancy and with subsequent abortions. Hum Reprod. (2001) 16:2219–26. doi: 10.1093/humrep/16.10.2219
40. Quack KC, Vassiliadou N, Pudney J, Anderson DJ, and Hill JA. Leukocyte activation in the decidua of chromosomally normal and abnormal fetuses from women with recurrent abortion. Hum Reprod. (2001) 16:949–55. doi: 10.1093/humrep/16.5.949
41. Hirahara F, Andoh N, Sawai K, Hirabuki T, Uemura T, and Minaguchi H. Hyperprolactinemic recurrent miscarriage and results of randomized bromocriptine treatment trials. Fertil Steril. (1998) 70:246–52. doi: 10.1016/S0015-0282(98)00164-2
42. Garzia E, Borgato S, Cozzi V, Doi P, Bulfamante G, Persani L, et al. Lack of expression of endometrial prolactin in early implantation failure: a pilot study. Hum Reprod. (2004) 19:1911–6. doi: 10.1093/humrep/deh350
43. Craig LB, Ke RW, and Kutteh WH. Increased prevalence of insulin resistance in women with a history of recurrent pregnancy loss. Fertil Steril. (2002) 78:487–90. doi: 10.1016/S0015-0282(02)03247-8
44. Li TC, Spuijbroek MDEH, Tuckerman E, Anstie B, Loxley M, and Laird S. Endocrinological and endometrial factors in recurrent miscarriage. BJOG: Int J Obstet Gynaecol. (2000) 107:1471–9. doi: 10.1111/j.1471-0528.2000.tb11670.x
45. Bussen S, Sütterlin M, and Steck T. Endocrine abnormalities during the follicular phase in women with recurrent spontaneous abortion. Hum Reprod. (1999) 14:18–20. doi: 10.1093/humrep/14.1.18
46. Kovalevsky G, Gracia CR, Berlin JA, Sammel MD, and Barnhart KT. Evaluation of the association between hereditary thrombophilias and recurrent pregnancy loss: A meta-analysis. Arch Internal Med. (2004) 164:558–63. doi: 10.1001/archinte.164.5.558
47. Rey E, Kahn SR, David M, and Shrier I. Thrombophilic disorders and fetal loss: a meta-analysis. Lancet. (2003) 361:901–8. doi: 10.1016/S0140-6736(03)12771-7
48. Rai R, Backos M, Elgaddal S, Shlebak A, and Regan L. Factor V Leiden and recurrent miscarriage—prospective outcome of untreated pregnancies. Hum Reprod. (2002) 17:442–5. doi: 10.1093/humrep/17.2.442
49. Rai R, Shlebak A, Cohen H, Backos M, Holmes Z, Marriott K, et al. Factor V Leiden and acquired activated protein C resistance among 1000 women with recurrent miscarriage. Hum Reprod. (2001) 16:961–5. doi: 10.1093/humrep/16.5.961
50. Rai RS, Clifford K, Cohen H, and Regan L. High prospective fetal loss rate in untreated pregnancies of women with recurrent miscarriage and antiphospholipid antibodies. Hum Reprod. (1995) 10:3301–4. doi: 10.1093/oxfordjournals.humrep.a135907
51. Rai RS, Regan L, Clifford K, Pickering W, Dave M, Mackie I, et al. Immunology: Antiphospholipid antibodies and β2-glycoprotein-I in 500 women with recurrent miscarriage: results of a comprehensive screening approach. Hum Reprod. (1995) 10:2001–5. doi: 10.1093/oxfordjournals.humrep.a136224
52. Greaves M, Cohen H, Machin SJ, and Mackie I. Guidelines on the investigation and management of the antiphospholipid syndrome. Br J Haematol. (2000) 109:704–15. doi: 10.1046/j.1365-2141.2000.02069.x
53. Tanimura K, Saito S, Tsuda S, Ono Y, Deguchi M, Nagamatsu T, et al. Low-dose aspirin and heparin treatment improves pregnancy outcome in recurrent pregnancy loss women with anti-beta2-glycoprotein I/HLA-DR autoantibodies: a prospective, multicenter, observational study. Front Immunol. (2024) 15:1445852. doi: 10.3389/fimmu.2024.1445852
54. Christiansen OB, Steffensen R, Nielsen HS, and Varming K. Multifactorial etiology of recurrent miscarriage and its scientific and clinical implications. Gynecol Obstet Invest. (2008) 66:257–67. doi: 10.1159/000149575
55. Heuser C, Dalton J, Macpherson C, Branch DW, Porter TF, and Silver RM. Idiopathic recurrent pregnancy loss recurs at similar gestational ages. Am J Obstet Gynecol. (2010) 203:343.e1–5. doi: 10.1016/j.ajog.2010.05.010
56. Yan J, Saravelos SH, Ma N, Ma C, Chen ZJ, and Li TC. Consecutive repeat miscarriages are likely to occur in the same gestational period. Reprod Biomed Online. (2012) 24:634–8. doi: 10.1016/j.rbmo.2012.03.003
57. Ticconi C, Giuliani E, Sorge R, Patrizi L, Piccione E, and Pietropolli A. Gestational age of pregnancy loss in women with unexplained recurrent miscarriage. J Obstet Gynaecol Res. (2016) 42:239–45. doi: 10.1111/jog.12903
58. Baird DD. The gestational timing of pregnancy loss: adaptive strategy? Am J Hum Biol. (2009) 21:725–7. doi: 10.1002/ajhb.20935
59. Piccinni MP, Raghupathy R, Saito S, and Szekeres-Bartho J. Cytokines, hormones and cellular regulatory mechanisms favoring successful reproduction. Front Immunol. (2021) 12:717808. doi: 10.3389/fimmu.2021.717808
60. Blois SM, Klapp BF, and Barrientos G. Decidualization and angiogenesis in early pregnancy: unravelling the functions of DC and NK cells. J Reprod Immunol. (2011) 88:86–92. doi: 10.1016/j.jri.2010.11.002
61. Blois SM, Freitag N, Tirado-Gonzalez I, Cheng SB, Heimesaat MM, Bereswill S, et al. NK cell-derived IL-10 is critical for DC-NK cell dialogue at the maternal-fetal interface. Sci Rep. (2017) 7:2189. doi: 10.1038/s41598-017-02333-8
62. Vomstein K, Feil K, Strobel L, Aulitzky A, Hofer-Tollinger S, Kuon RJ, et al. Immunological risk factors in recurrent pregnancy loss: guidelines versus current state of the art. J Clin Med. (2021) 10:869. doi: 10.3390/jcm10040869
63. Yang X, Tian Y, Zheng L, Luu T, and Kwak-Kim J. The update immune-regulatory role of pro- and anti-inflammatory cytokines in recurrent pregnancy losses. Int J Mol Sci. (2022) 24:132. doi: 10.3390/ijms24010132
64. Trout SW and Seifer DB. Do women with unexplained recurrent pregnancy loss have higher day 3 serum FSH and estradiol values? Fertil Steril. (2000) 74:335–7. doi: 10.1016/s0015-0282(00)00625-7
65. Gürbüz B, Yalti S, Ozden S, and Ficicioglu C. High basal estradiol level and FSH/LH ratio in unexplained recurrent pregnancy loss. Arch Gynaecol Obstet. (2004) 270:37–9. doi: 10.1007/s00404-003-0490-0
66. Pils S, Promberger R, Springer S, Joura E, and Ott J. Decreased ovarian reserve predicts inexplicability of recurrent miscarriage? A retrospective analysis. PloS One. (2016) 11:e0161606. doi: 10.1371/journal.pone.0161606
67. Okon MA, Laird SM, Tuckerman EM, and Li T-C. Serum Androgen Levels in Women who Have Recurrent Miscarriages and their Correlation with Markers of Endometrial Function 11Supported by a research grant from the Special Trustees of the former United Sheffield Hospitals, Royal Hallamshire Hospital, Sheffield, Yorkshire, United Kingdom. Fertil Steril. (1998) 69:682–90. doi: 10.1016/S0015-0282(98)00007-7
68. Kim M-S, Gu B-H, Song S, Choi B-C, Cha D-H, and Baek K-H. ITI-H4, as a biomarker in the serum of recurrent pregnancy loss (RPL) patients. Mol BioSystems. (2011) 7:1430–40. doi: 10.1039/c0mb00219d
69. Ogasawara M, Kajiura S, Katano K, Aoyama T, and Aoki K. Are serum progesterone levels predictive of recurrent miscarriage in future pregnancies? Fertil Steril. (1997) 68:806–9. doi: 10.1016/s0015-0282(97)00328-2
70. Wu Y, He J, Guo C, Zhang Y, Yang W, Xin M, et al. Serum biomarker analysis in patients with recurrent spontaneous abortion. Mol Med Rep. (2017) 16:2367–78. doi: 10.3892/mmr.2017.6890
71. Cui Y, He L, Yang C-Y, and Ye Q. iTRAQ and PRM-based quantitative proteomics in early recurrent spontaneous abortion: biomarkers discovery. Clin Proteomics. (2019) 16:36. doi: 10.1186/s12014-019-9256-y
72. Al-Azemi M, Ledger WL, Diejomaoh M, Mousa M, Makhseed M, and Omu A. Measurement of inhibin A and inhibin pro-alphaC in early human pregnancy and their role in the prediction of pregnancy outcome in patients with recurrent pregnancy loss. Fertil Steril. (2003) 80:1473–9. doi: 10.1016/S0015-0282(03)02215-5
73. Liu Y, Liu Y, Li X, Jiao X, Zhang R, and Zhang J. Predictive value of serum β-hCG for early pregnancy outcomes among women with recurrent spontaneous abortion. Int J Gynecology Obstetrics. (2016) 135:16–21. doi: 10.1016/j.ijgo.2016.03.007
74. Muttukrishna S, Jauniaux E, Greenwold N, McGarrigle H, Jivraj S, Carter S, et al. Circulating levels of inhibin A, activin A and follistatin in missed and recurrent miscarriages. Hum Reprod. (2002) 17:3072–8. doi: 10.1093/humrep/17.12.3072
75. Rull K and Laan M. Expression of β-subunit of HCG genes during normal and failed pregnancy. Hum Reprod. (2005) 20:3360–8. doi: 10.1093/humrep/dei261
76. Darwish A, Ghorab N, El-Ashmawy HM, Kamal MM, and Soliman AMS. Biochemical markers for prediction of pregnancy outcome in cases of recurrent pregnancy loss. Conf Proc. (2005) 10:59–62.
77. Heinig J, Steinhard J, Schmitz R, Nofer JR, Kiesel L, and Klockenbusch W. Maternal serum free ß-hCG and PAPP-A in patients with habitual abortion-influence on first-trimester screening for chromosomal abnormalities. Prenat Diagn. (2007) 27:814–6. doi: 10.1002/pd.1779
78. Kato K, Mostafa MH, Mann K, Schindler AE, and Hoermann R. Human chorionic gonadotropin exhibits normal biological activity in patients with recurrent pregnancy loss. Gynecol Endocrinol. (2002) 16:179–86. doi: 10.1080/gye.16.3.179.186
79. Prakash A, Laird S, Tuckerman E, Li TC, and Ledger WL. Inhibin A and activin A may be used to predict pregnancy outcome in women with recurrent miscarriage. Fertil Steril. (2005) 83:1758–63. doi: 10.1016/j.fertnstert.2004.11.072
80. Bao SH, Shuai W, Tong J, Wang L, Chen P, and Duan T. Increased Dickkopf-1 expression in patients with unexplained recurrent spontaneous miscarriage. Clin Exp Immunol. (2013) 172:437–43. doi: 10.1111/cei.12066
81. Rull K, Tomberg K, Kõks S, Männik J, Möls M, Sirotkina M, et al. Increased placental expression and maternal serum levels of apoptosis-inducing TRAIL in recurrent miscarriage. Placenta. (2013) 34:141–8. doi: 10.1016/j.placenta.2012.11.032
82. Pang L, Wei Z, Li O, Huang R, Qin J, Chen H, et al. An increase in vascular endothelial growth factor (VEGF) and VEGF soluble receptor-1 (sFlt-1) are associated with early recurrent spontaneous abortion. PloS One. (2013) 8:e75759. doi: 10.1371/journal.pone.0075759
83. Laird SM, Quinton ND, Anstie B, Li TC, and Blakemore AIF. Leptin and leptin-binding activity in women with recurrent miscarriage: correlation with pregnancy outcome. Hum Reprod. (2001) 16:2008–13. doi: 10.1093/humrep/16.9.2008
84. Hassan MF. Soluble fms-like tyrosine kinase-1 and vascular endothelial growth factor: Novel markers for unexplained early recurrent pregnancy loss. Asian Pacific J Reprod. (2014) 3:30–4. doi: 10.1016/S2305-0500(13)60181-5
85. Christiansen OB, Nybo Andersen AM, Bosch E, Daya S, Delves PJ, Hviid TV, et al. Evidence-based investigations and treatments of recurrent pregnancy loss. Fertil Steril. (2005) 83:821–39. doi: 10.1016/j.fertnstert.2004.12.018
86. Christiansen OB. Evidence-based investigations and treatments of recurrent pregnancy loss. Curr Opin Obstet Gynecol. (2006) 18:304–12. doi: 10.1097/01.gco.0000193011.73405.07
87. ACOG. The American College of Obstetricians and Gynecologists Practice Bulletin no. 150. Early pregnancy loss. Obstet Gynecol. (2015) 125:1258–67. doi: 10.1097/01.AOG.0000465191.27155.25
88. Toth E, Gyorffy D, Posta M, Hupuczi P, Balogh A, Szalai G, et al. Decreased expression of placental proteins in recurrent pregnancy loss: functional relevance and diagnostic value. Int J Mol Sci. (2024) 25:1865. doi: 10.3390/ijms25031865
89. Callister SJ, Barry RC, Adkins JN, Johnson ET, Qian W-J, Webb-Robertson B-JM, et al. Normalization approaches for removing systematic biases associated with mass spectrometry and label-free proteomics. J Proteome Res. (2006) 5:277–86. doi: 10.1021/pr050300l
90. Than NG, Romero R, Tarca AL, Kekesi KA, Xu Y, Xu Z, et al. Integrated systems biology approach identifies novel maternal and placental pathways of preeclampsia. Front Immunol. (2018) 9:1661. doi: 10.3389/fimmu.2018.01661
91. Szilagyi A, Gelencser Z, Romero R, Xu Y, Kiraly P, Demeter A, et al. Placenta-specific genes, their regulation during villous trophoblast differentiation and dysregulation in preterm preeclampsia. Int J Mol Sci. (2020) 21:628. doi: 10.3390/ijms21020628
92. Storey JD. A direct approach to false discovery rates. J R Stat Society: Ser B (Statistical Methodology). (2002) 64:479–98. doi: 10.1111/1467-9868.00346
93. Tal R and Taylor HS. Endocrinology of pregnancy. In: Feingold KR, Ahmed SF, Anawalt B, Blackman MR, Boyce A, Chrousos G, et al, editors. Endotext. South Dartmouth (MA (2000).
94. Csapo AI and Pulkkinen M. Indispensability of the human corpus luteum in the maintenance of early pregnancy. Luteectomy evidence. Obstet Gynecol Surv. (1978) 33:69–81. doi: 10.1097/00006254-197802000-00001
95. Jauniaux E, Watson AL, Hempstock J, Bao YP, Skepper JN, and Burton GJ. Onset of maternal arterial blood flow and placental oxidative stress. A possible factor in human early pregnancy failure. Am J Pathology. (2000) 157:2111–22. doi: 10.1016/S0002-9440(10)64849-3
96. Szklarczyk D, Kirsch R, Koutrouli M, Nastou K, Mehryary F, Hachilif R, et al. The STRING database in 2023: protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. (2023) 51:D638–D46. doi: 10.1093/nar/gkac1000
97. Gene Ontology C, Aleksander SA, Balhoff J, Carbon S, Cherry JM, Drabkin HJ, et al. The gene ontology knowledgebase in 2023. Genetics. (2023) 224. doi: 10.1093/genetics/iyad031
98. Burton GJ and Jauniaux E. What is the placenta? Am J Obstet Gynecol. (2015) 213:S6.e1, S6–8. doi: 10.1016/j.ajog.2015.07.050
99. Khorami-Sarvestani S, Vanaki N, Shojaeian S, Zarnani K, Stensballe A, Jeddi-Tehrani M, et al. Placenta: an old organ with new functions. Front Immunol. (2024) 15:1385762. doi: 10.3389/fimmu.2024.1385762
100. Crowley E, Di Nicolantonio F, Loupakis F, and Bardelli A. Liquid biopsy: monitoring cancer-genetics in the blood. Nat Rev Clin Oncol. (2013) 10:472–84. doi: 10.1038/nrclinonc.2013.110
101. Alix-Panabieres C and Pantel K. Circulating tumor cells: liquid biopsy of cancer. Clin Chem. (2013) 59:110–8. doi: 10.1373/clinchem.2012.194258
102. Heitzer E, Ulz P, and Geigl JB. Circulating tumor DNA as a liquid biopsy for cancer. Clin Chem. (2015) 61:112–23. doi: 10.1373/clinchem.2014.222679
103. Alix-Panabieres C and Pantel K. Clinical applications of circulating tumor cells and circulating tumor DNA as liquid biopsy. Cancer Discov. (2016) 6:479–91. doi: 10.1158/2159-8290.CD-15-1483
104. Karaszi K, Szabo S, Juhasz K, Kiraly P, Kocsis-Deak B, Hargitai B, et al. Increased placental expression of Placental Protein 5 (PP5)/Tissue Factor Pathway Inhibitor-2 (TFPI-2) in women with preeclampsia and HELLP syndrome: Relevance to impaired trophoblast invasion? Placenta. (2019) 76:30–9. doi: 10.1016/j.placenta.2019.01.011
105. Than NG, Balogh A, Romero R, Kárpáti É, Erez O, Szilágyi A, et al. Placental protein 13 (PP13) – A placental immunoregulatory galectin protecting pregnancy. Front Immunol. (2014) 5. doi: 10.3389/fimmu.2014.00348
106. Balogh A, Pozsgay J, Matkó J, Dong Z, Kim CJ, Várkonyi T, et al. Placental protein 13 (PP13/galectin-13) undergoes lipid raft-associated subcellular redistribution in the syncytiotrophoblast in preterm preeclampsia and HELLP syndrome. Am J Obstet Gynecol. (2011) 205:156.e1–.e14. doi: 10.1016/j.ajog.2011.03.023
107. Than NG, Abdul Rahman O, Magenheim R, Nagy B, Fule T, Hargitai B, et al. Placental protein 13 (galectin-13) has decreased placental expression but increased shedding and maternal serum concentrations in patients presenting with preterm pre-eclampsia and HELLP syndrome. Virchows Archiv. (2008) 453:387–400. doi: 10.1007/s00428-008-0658-x
108. Than NG, Romero R, Balogh A, Karpati E, Mastrolia SA, Staretz-Chacham O, et al. Galectins: double-edged swords in the cross-roads of pregnancy complications and female reproductive tract inflammation and neoplasia. J Pathol Trans Med. (2015) 49:181–208. doi: 10.4132/jptm.2015.02.25
109. Romero R, Kusanovic JP, Than NG, Erez O, Gotsch F, Espinoza J, et al. First-trimester maternal serum PP13 in the risk assessment for preeclampsia. Am J Obstet Gynecol. (2008) 199:122.e1–.e11. doi: 10.1016/j.ajog.2008.01.013
110. Than NG, Romero R, Meiri H, Erez O, Xu Y, Tarquini F, et al. PP13, maternal ABO blood groups and the risk assessment of pregnancy complications. PloS One. (2011) 6:e21564–e. doi: 10.1371/journal.pone.0021564
111. Balogh A, Toth E, Romero R, Parej K, Csala D, Szenasi NL, et al. Placental galectins are key players in regulating the maternal adaptive immune response. Front Immunol. (2019) 10:1240. doi: 10.3389/fimmu.2019.01240
112. Than GN, Bohn H, and Szabo DG. Advances in Pregnancy-Related Protein Research. Boca Raton: CRC Press (1993).
113. Vokalova L, Balogh A, Toth E, Van Breda SV, Schäfer G, Hoesli I, et al. Placental protein 13 (Galectin-13) polarizes neutrophils toward an immune regulatory phenotype. Front Immunol. (2020) 11:145–. doi: 10.3389/fimmu.2020.00145
114. Aebersold R and Mann M. Mass-spectrometric exploration of proteome structure and function. Nature. (2016) 537:347–55. doi: 10.1038/nature19949
115. Manna S, Scheel J, Noone A, McElwain CJ, Scaife C, Gupta S, et al. A proteomic profile of the healthy human placenta. Clin Proteomics. (2023) 20:1. doi: 10.1186/s12014-022-09388-4
116. Tarca AL, Romero R, Bhatti G, Gotsch F, Done B, Gudicha DW, et al. Human plasma proteome during normal pregnancy. J Proteome Res. (2022) 21:2687–702. doi: 10.1021/acs.jproteome.2c00391
117. Michela B. Liquid biopsy: A family of possible diagnostic tools. Diagnostics. (2021) 11:391. doi: 10.3390/diagnostics11081391
118. Masserdotti A, Gasik M, Grillari-Voglauer R, Grillari J, Cargnoni A, Chiodelli P, et al. Unveiling the human fetal-maternal interface during the first trimester: biophysical knowledge and gaps. Front Cell Dev Biol. (2024) 12:1411582. doi: 10.3389/fcell.2024.1411582
119. Lo YM, Tein MS, Lau TK, Haines CJ, Leung TN, Poon PM, et al. Quantitative analysis of fetal DNA in maternal plasma and serum: implications for noninvasive prenatal diagnosis. Am J Hum Genet. (1998) 62:768–75. doi: 10.1086/301800
120. Sarker S, Scholz-Romero K, Perez A, Illanes SE, Mitchell MD, Rice GE, et al. Placenta-derived exosomes continuously increase in maternal circulation over the first trimester of pregnancy. J Trans Med. (2014) 12:204. doi: 10.1186/1479-5876-12-204
121. Mayhew TM, Charnock-Jones DS, and Kaufmann P. Aspects of human fetoplacental vasculogenesis and angiogenesis. III. Changes in complicated pregnancies. Placenta. (2004) 25:127–39. doi: 10.1016/j.placenta.2003.10.010
122. Kaufmann P, Mayhew TM, and Charnock-Jones DS. Aspects of human fetoplacental vasculogenesis and angiogenesis. II. Changes during normal pregnancy. Placenta. (2004) 25:114–26. doi: 10.1016/j.placenta.2003.10.009
123. Burton GJ and Jauniaux E. Placental oxidative stress: from miscarriage to preeclampsia. J Soc Gynecol Invest. (2004) 11:342–52. doi: 10.1016/j.jsgi.2004.03.003
124. Gude NM, Roberts CT, Kalionis B, and King RG. Growth and function of the normal human placenta. Thromb Res. (2004) 114:397–407. doi: 10.1016/j.thromres.2004.06.038
125. Singh P, Carraher C, and Schwarzbauer JE. Assembly of fibronectin extracellular matrix. Annu Rev Cell Dev Biol. (2010) 26:397–419. doi: 10.1146/annurev-cellbio-100109-104020
126. Smits P, Ni J, Feng P, Wauters J, Van Hul W, Boutaibi ME, et al. The human extracellular matrix gene 1 (ECM1): genomic structure, cDNA cloning, expression pattern, and chromosomal localization. Genomics. (1997) 45:487–95. doi: 10.1006/geno.1997.4918
127. Posey KL, Coustry F, and Hecht JT. Cartilage oligomeric matrix protein: COMPopathies and beyond. Matrix Biol. (2018) 71-72:161–73. doi: 10.1016/j.matbio.2018.02.023
128. Aidoudi S and Bikfalvi A. Interaction of PF4 (CXCL4) with the vasculature: a role in atherosclerosis and angiogenesis. Thromb Haemostasis. (2010) 104:941–8. doi: 10.1160/TH10-03-0193
129. Forstner D, Guettler J, and Gauster M. Changes in maternal platelet physiology during gestation and their interaction with trophoblasts. Int J Mol Sci. (2021) 22:10732. doi: 10.3390/ijms221910732
130. Nawathe AR, Christian M, Kim SH, Johnson M, Savvidou MD, and Terzidou V. Insulin-like growth factor axis in pregnancies affected by fetal growth disorders. Clin Epigenetics. (2016) 8:11. doi: 10.1186/s13148-016-0178-5
131. Skornicka EL, Kiyatkina N, Weber MC, Tykocinski ML, and Koo PH. Pregnancy zone protein is a carrier and modulator of placental protein-14 in T-cell growth and cytokine production. Cell Immunol. (2004) 232:144–56. doi: 10.1016/j.cellimm.2005.03.007
132. Lob S, Vattai A, Kuhn C, Schmoeckel E, Mahner S, Wockel A, et al. Pregnancy Zone Protein (PZP) is significantly upregulated in the decidua of recurrent and spontaneous miscarriage and negatively correlated to Glycodelin A (GdA). J Reprod Immunol. (2021) 143:103267. doi: 10.1016/j.jri.2020.103267
133. Aghababaei M, Perdu S, Irvine K, and Beristain AG. A disintegrin and metalloproteinase 12 (ADAM12) localizes to invasive trophoblast, promotes cell invasion and directs column outgrowth in early placental development. Mol Hum Reprod. (2014) 20:235–49. doi: 10.1093/molehr/gat084
134. Moore T, Williams JM, Becerra-Rodriguez MA, Dunne M, Kammerer R, and Dveksler G. Pregnancy-specific glycoproteins: evolution, expression, functions and disease associations. Reprod. (2022) 163:R11–23. doi: 10.1530/REP-21-0390
135. Fei D, Meng X, Yu W, Yang S, Song N, Cao Y, et al. Fibronectin (FN) cooperated with TLR2/TLR4 receptor to promote innate immune responses of macrophages binding to integrin beta1. Virulence. (2018) 9:1588–600. doi: 10.1080/21505594.2018.1528841
136. Horne CH, Towler CM, Pugh-Humphreys RG, Thomson AW, and Bohn H. Pregnancy specific beta1-glycoprotein–a product of the syncytiotrophoblast. Experientia. (1976) 32:1197. doi: 10.1007/BF01927624
137. Snyder SK, Wessner DH, Wessells JL, Waterhouse RM, Wahl LM, Zimmermann W, et al. Pregnancy-specific glycoproteins function as immunomodulators by inducing secretion of IL-10, IL-6 and TGF-beta1 by human monocytes. Am J Reprod Immunol. (2001) 45:205–16. doi: 10.1111/j.8755-8920.2001.450403.x
138. Wan YY and Flavell RA. ‘Yin-Yang’ functions of transforming growth factor-beta and T regulatory cells in immune regulation. Immunol Rev. (2007) 220:199–213. doi: 10.1111/j.1600-065X.2007.00565.x
139. Liu Y, Bockermann R, Hadi M, Safari I, Carrion B, Kveiborg M, et al. ADAM12 is a costimulatory molecule that determines Th1 cell fate and mediates tissue inflammation. Cell Mol Immunol. (2021) 18:1904–19. doi: 10.1038/s41423-020-0486-8
140. Teirila L, Heikkinen-Eloranta J, Kotimaa J, Meri S, and Lokki AI. Regulation of the complement system and immunological tolerance in pregnancy. Semin Immunol. (2019) 45:101337. doi: 10.1016/j.smim.2019.101337
141. Lee CL, Lam KK, Vijayan M, Koistinen H, Seppala M, Ng EH, et al. The pleiotropic effect of glycodelin-A in early pregnancy. Am J Reprod Immunol. (2016) 75:290–7. doi: 10.1111/aji.12471
142. Rachmilewitz J, Borovsky Z, Riely GJ, Miller R, and Tykocinski ML. Negative regulation of T cell activation by placental protein 14 is mediated by the tyrosine phosphatase receptor CD45. J Biol Chem. (2003) 278:14059–65. doi: 10.1074/jbc.M211716200
143. Seppala M, Taylor RN, Koistinen H, Koistinen R, and Milgrom E. Glycodelin: a major lipocalin protein of the reproductive axis with diverse actions in cell recognition and differentiation. Endocrine Rev. (2002) 23:401–30. doi: 10.1210/er.2001-0026
144. Sylvers-Davie KL and Davies BSJ. Regulation of lipoprotein metabolism by ANGPTL3, ANGPTL4, and ANGPTL8. Am J Physiol Endocrinol Metab. (2021) 321:E493–508. doi: 10.1152/ajpendo.00195.2021
145. Camenisch G, Pisabarro MT, Sherman D, Kowalski J, Nagel M, Hass P, et al. ANGPTL3 stimulates endothelial cell adhesion and migration via integrin alpha vbeta 3 and induces blood vessel formation in vivo. J Biol Chem. (2002) 277:17281–90. doi: 10.1074/jbc.M109768200
146. Greinacher A and Warkentin TE. Platelet factor 4 triggers thrombo-inflammation by bridging innate and adaptive immunity. Int J Lab Hemost. (2023) 45 Suppl 2:11–22. doi: 10.1111/ijlh.14075
147. Ji Y, Zhang Q, Li H, Chen L, Wu Y, and Lin S. Platelet factor 4: A mysterious chemokine in inflammatory regulation diseases. J Inflam Res. (2025) 18:4481–95. doi: 10.2147/JIR.S504673
148. Wu Y, Zhao Z, Deng X, Jia J, and Yuan G. Pregnancy zone protein, a potential research target in multiple diseases. Gene. (2025) 935:149013. doi: 10.1016/j.gene.2024.149013
149. Erez O, Romero R, Tarca AL, Chaiworapongsa T, Kim YM, Than NG, et al. Differential expression pattern of genes encoding for anti-microbial peptides in the fetal membranes of patients with spontaneous preterm labor and intact membranes and those with preterm prelabor rupture of the membranes. J Matern Fetal Neonatal Med. (2009) 22:1103–15. doi: 10.3109/14767050902994796
150. Lob S, Vattai A, Kuhn C, Mittelberger J, Herbert SL, Wockel A, et al. The Pregnancy Zone Protein (PZP) is significantly downregulated in the placenta of preeclampsia and HELLP syndrome patients. J Reprod Immunol. (2022) 153:103663. doi: 10.1016/j.jri.2022.103663
151. Papamichail M, Fasoulakis Z, Daskalakis G, Theodora M, Rodolakis A, and Antsaklis P. Importance of low pregnancy associated plasma protein-A (PAPP-A) levels during the first trimester as a predicting factor for adverse pregnancy outcomes: A prospective cohort study of 2636 pregnant women. Cureus. (2022) 14:e31256. doi: 10.7759/cureus.31256
152. Xu Q, Yang Y, Liu F, Wang L, Wang Q, Shen H, et al. Preconception Hb concentration with risk of spontaneous abortion: a population-based cohort study in over 3.9 million women across rural China. Public Health Nutr. (2020) 23:2963–72. doi: 10.1017/S1368980019003811
153. Diaz-Lopez A, Ribot B, Basora J, and Arija V. High and low haemoglobin levels in early pregnancy are associated to a higher risk of miscarriage: A population-based cohort study. Nutrients. (2021) 13:1578. doi: 10.3390/nu13051578
154. Schumacher A. Human chorionic gonadotropin as a pivotal endocrine immune regulator initiating and preserving fetal tolerance. Int J Mol Sci. (2017) 18:2166. doi: 10.3390/ijms18102166
155. Martinez FF, Cervi L, Knubel CP, Panzetta-Dutari GM, and Motran CC. The role of pregnancy-specific glycoprotein 1a (PSG1a) in regulating the innate and adaptive immune response. Am J Reprod Immunol. (2013) 69:383–94. doi: 10.1111/aji.12089
156. Takeuchi T, Nishii O, Okamura T, Yaginuma T, and Kawana T. Free testosterone and abortion in early pregnancy. Int J Gynaecol Obstet. (1993) 43:151–6. doi: 10.1016/0020-7292(93)90322-N
157. Balogh A, Karpati E, Schneider AE, Hetey S, Szilagyi A, Juhasz K, et al. Sex hormone-binding globulin provides a novel entry pathway for estradiol and influences subsequent signaling in lymphocytes via membrane receptor. Sci Rep. (2019) 9:4. doi: 10.1038/s41598-018-36882-3
158. Pijnenborg R, Vercruysse L, and Hanssens M. The uterine spiral arteries in human pregnancy: facts and controversies. Placenta. (2006) 27:939–58. doi: 10.1016/j.placenta.2005.12.006
159. Burton GJ, Woods AW, Jauniaux E, and Kingdom JC. Rheological and physiological consequences of conversion of the maternal spiral arteries for uteroplacental blood flow during human pregnancy. Placenta. (2009) 30:473–82. doi: 10.1016/j.placenta.2009.02.009
160. Jauniaux E and Burton GJ. Pathophysiology of histological changes in early pregnancy loss. Placenta. (2005) 26:114–23. doi: 10.1016/j.placenta.2004.05.011
161. Parker CE and Borchers CH. Mass spectrometry based biomarker discovery, verification, and validation–quality assurance and control of protein biomarker assays. Mol Oncol. (2014) 8:840–58. doi: 10.1016/j.molonc.2014.03.006
162. Lehallier B, Gate D, Schaum N, Nanasi T, Lee SE, Yousef H, et al. Undulating changes in human plasma proteome profiles across the lifespan. Nat Med. (2019) 25:1843–50. doi: 10.1038/s41591-019-0673-2
Keywords: clinical proteomics, diagnostics, high-dimensional, miscarriage, personalized medicine, prevention, reproduction, systems biology
Citation: Tóth E, Posta M, Györffy D, Oravecz O, Farkas E, Balogh A, Escher C, Bober M, Szilágyi A, Hupuczi P, Veress L, Török O, Nagy S, Rinner O, Erez O, Papp Z, Ács N and Than NG (2025) Novel proteomics biomarkers of recurrent pregnancy loss reflect the dysregulation of immune interactions at the maternal-fetal interface. Front. Immunol. 16:1621168. doi: 10.3389/fimmu.2025.1621168
Received: 30 April 2025; Accepted: 11 July 2025;
Published: 28 August 2025.
Edited by:
Michael Eikmans, Leiden University Medical Center (LUMC), NetherlandsReviewed by:
Panicos Shangaris, King’s College London, United KingdomDanai Bagkou Dimakou, University of Birmingham, United Kingdom
Copyright © 2025 Tóth, Posta, Györffy, Oravecz, Farkas, Balogh, Escher, Bober, Szilágyi, Hupuczi, Veress, Török, Nagy, Rinner, Erez, Papp, Ács and Than. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nándor Gábor Than, dGhhbi5nYWJvckB0dGsuaHU=; dGhhbi5nYWJvckBzZW1tZWx3ZWlzLmh1
†These authors have contributed equally to this work and share first authorship