 Tünde Fekete
Tünde Fekete Kitti Pázmándi
Kitti Pázmándi- Department of Immunology, Faculty of Medicine, University of Debrecen, Debrecen, Hungary
Dendritic cells (DCs) constitute a heterogeneous population of immune cells that acting as antigen presenting cells link innate and adaptive immune responses. Their functions are mainly dictated by microenvironmental cues, enabling them to either maintain immune tolerance or initiate robust humoral and cellular immune responses. While DCs are important for orchestrating immune responses, accumulating evidence suggests that aberrant DC activation contributes to the pathogenesis of autoimmune and chronic inflammatory diseases, making them promising targets for therapeutic modulation. Modulating DC functionality therefore represents a potent strategy to attenuate excessive inflammation in such conditions. Plant-derived bioactive compounds, or phytochemicals, are structurally diverse secondary metabolites with established anti-inflammatory and immunomodulatory properties. This review consolidates current in vitro, in vivo, and in silico findings on ten well-characterized phytochemicals including curcumin, 6-gingerol, 6-shogaol, resveratrol, epigallocatechin-3-gallate, quercetin, apigenin, capsaicin, berberine and ginsenosides, which have the capacity to modulate DC phenotype and function. Notably, these phytochemicals can skew DCs toward a tolerogenic phenotype, characterized by reduced expression of antigen presenting and co-stimulatory molecules, diminished pro-inflammatory cytokine secretion, and enhanced regulatory T cell induction. Mechanistic insights reveal convergence on key signaling pathways such as nuclear factor-kappa B (NF-κB), mitogen activated protein kinase (MAPK) and mammalian target of rapamycin (mTOR) in DCs. In silico studies further predict interactions of these compounds with various molecular targets, providing a structural basis for their immunoregulatory effects. Furthermore, studies using preclinical models of autoimmune and inflammatory diseases have demonstrated that these phytochemicals can attenuate disease severity, likely through DC modulation. Given their multifaceted immunomodulatory capacity, phytochemicals hold promise both as adjuvant therapies in DC-mediated autoimmune diseases and as agents for generating tolerogenic DCs for cell-based immunotherapies.
1 Introduction
Dendritic cells (DCs) are a specialized group of immune cells with a pivotal role in orchestrating both innate and adaptive immune responses. Beyond their well-known function as professional antigen presenting cells, DCs are essential for maintaining peripheral tolerance and immune homeostasis under physiological conditions. As primary sentinels of the immune system, they rapidly detect invading pathogens or endogenous danger signals and initiate appropriate immune responses. Their inherent plasticity enables them to dynamically respond to environmental stimuli and to adopt either immunostimulatory or tolerogenic phenotypes, thus contributing to both initiation of inflammatory responses and the induction of immune tolerance. Owing to their exceptional immunomodulatory capacity, they are considered promising therapeutic targets for the treatment of various inflammatory and autoimmune diseases (1).
In recent years, increasing attention has been directed toward plant-derived bioactive compounds as potential immunomodulatory agents (2). Plants are rich in diverse secondary metabolites, commonly categorized into phenolics, terpenoids, and nitrogen-containing compounds based on their chemical structures, biosynthetic origin, and biological functions (3). These phytochemicals are not only central to the defense mechanisms of the plants, but also exert a wide range of pharmacological effects in humans. Many of them have been recognized for their anti-inflammatory, antioxidant antimicrobial, anticancer, neuroprotective, and cardioprotective activities. Their immunomodulatory properties, in particular, have gained attention in the context of chronic inflammation and immune mediated disorders (4).
Several in vitro and preclinical studies have demonstrated that specific phytochemicals can modulate DC function by promoting a tolerogenic phenotype. These compounds are often derived from medicinal and culinary plants such as herbs, spices, fruits, and vegetables, which have long been used in traditional medicine (4). By interfering with signaling pathways, cytokine production, and costimulatory molecule expression, these molecules may inhibit DC maturation or promote the development of DC subsets with a tolerogenic phenotype. Consequently, they hold therapeutic promise in shaping immune responses toward tolerance, especially when applied in ex vivo DC-based immunotherapies.
In this review, we aimed to compile current evidence on the immunomodulatory effects of plant-derived bioactive compounds on DCs, with a particular focus on their potential use in treating autoimmune and inflammatory conditions. Specifically, we selected 10 well-characterized phytochemicals including curcumin, 6-gingerol, 6-shogaol, resveratrol, epigallocatechin-3-gallate, quercetin, apigenin, capsaicin, berberine and ginsenosides. These bioactive compounds were selected based on literature data demonstrating their immunosuppressive properties and ability to promote tolerogenic DC differentiation. Scientific publications from the past two decades were collected from databases such as PubMed and Web of Science, using combinations of terms such as “dendritic cell,” “tolerogenic,” “immunosuppressive,” and the specific compound names. Retrieved articles were manually screened to ensure relevance, and reviewed to provide a comprehensive overview of the current findings and to highlight potential directions for future therapeutic applications. In addition, during the manual selection process, preference was given to peer-reviewed articles published in reputable scientific journals to ensure data quality. Studies using purified compounds were included, while those employing crude extracts were excluded. The chemical classification, structural features, and biological functions of the selected compounds are presented in Table 1.
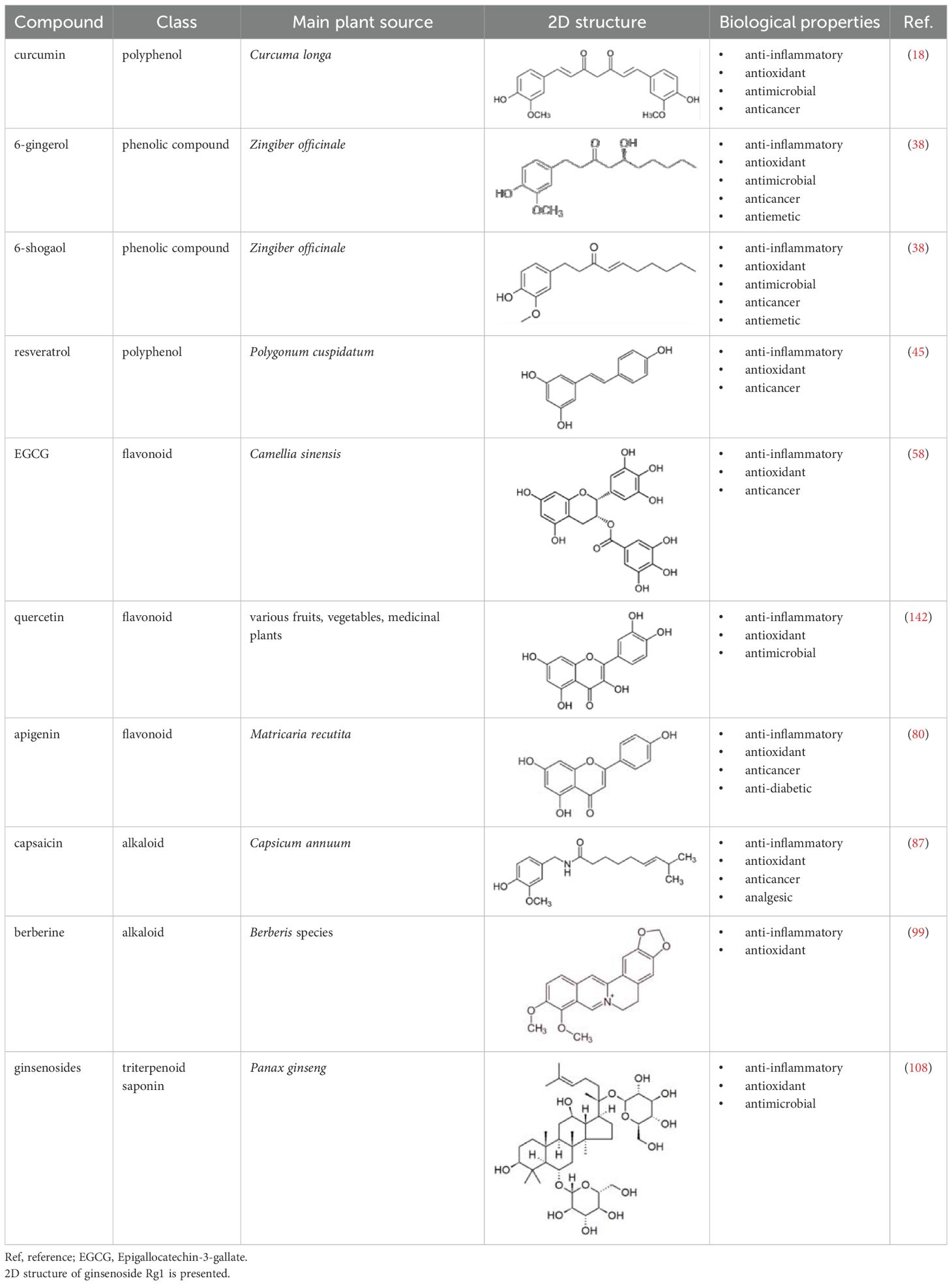
Table 1. Characteristics and main biological properties of selected phytochemicals.
2 DCs at a glance
In the early 1970’s a special stellate shaped cell type was discovered in mouse spleen by Steinmann and Cohn (5, 6). These cells were named DCs for their tree-like processes after the Greek word, dendron, meaning tree. The large surface area enables DCs to interact with numerous surrounding cells, and simultaneously present antigens to multiple T cells. While DCs are known for their high antigen presenting capacity, their role goes far beyond antigen presentation. In addition to interacting with T cells, DCs communicate with innate immune cells such as natural killer (NK) cells and neutrophils, and convey information to non-immune cell types as well. As a result, DCs are able to exhibit a multitude of tasks from initiating, coordinating and regulating immune responses to maintaining tissue homeostasis and self-tolerance (1).
DCs constitute a heterogeneous population of immune cells originating from the bone marrow (7). They are widely distributed throughout the body, circulate in the blood and are present in both lymphoid and non-lymphoid tissues. Based on their ontogeny, phenotype and transcriptional profile DCs can been categorized into distinct subsets. In humans, DCs comprise approximately 1% of peripheral blood mononuclear cells (PBMCs) and are generally classified into two major groups: plasmacytoid DCs (pDCs) and conventional DCs (cDCs), each endowed with distinct effector functions (7). pDCs majorly sense viral infections and as a response rapidly produce type I interferons (IFNs) to create an immediate phase of antiviral state in the surrounding cells (8). The cDC compartment consists of several subtypes that express a broad range of pattern recognition receptors such as Toll-like receptors (TLRs) enabling them to detect a wide variety of pathogens (9). Importantly, under inflammatory conditions, DCs might also arise from monocytes that are usually termed as inflammatory or monocyte-derived DCs (moDCs) (7). In 1994, Sallusto and Lanzavecchia described that human monocytes treated with granulocyte-macrophage colony-stimulating factor (GM-CSF) and interleukin-4 (IL-4) acquire DC-like properties (10). Since then, moDC has become the most commonly used in vitro model for studying DC biology and for DC-based immunotherapy approaches. For a long time, Langerhans cells (LCs) in the skin were also considered DCs due to their migratory capacity and antigen presenting functions. However, recent findings have revealed their distinct developmental origin and LCs have been reclassified as macrophages (11). The major DC subsets are highly conserved between mice and humans. Although, several phenotypic differences exist between the same DC subsets across the two species, functional equivalences can often be drawn (12). Therefore, mouse models are the most widely used systems for studying DC biology and evaluating the therapeutic potential of DC-based vaccines (13). The classification, ontogeny and functional specialization of DC subsets have been comprehensively reviewed elsewhere (1, 7).
In general, DCs exist in two functional states: a resting state and an activated state (1). In their resting or quiescent state, DCs act as sentinels of the immune system, continuously surveilling tissues for invading pathogens, damaged or cancerous cells. Resting DCs are characterized by high endocytic capacity, low expression of costimulatory molecules and rapid turnover of major histocompatibility complex II (MHC II) molecules. Importantly, resting DCs constitutively express both MHC I and MHC II molecules, but the dynamics of these molecules on the cell surface differ substantially. MHC II-peptide complexes exhibit rapid turnover on the surface of resting DCs to ensure constant sampling and presentation of extracellular antigens to CD4+ T cells (14). In contrast, MHC I-peptide complexes are more stable and long-lived on the plasma membrane that is sufficient for continuous surveillance of intracellular proteins and presentation to CD8+ T cells (15). Upon sensing changes in their local environment, DCs become activated, enabling them to migrate to draining lymph nodes and engage with other immune cell types such as B and T cells. Activated DCs are characterized by reduced endocytosis, upregulation of the homing receptor C-C chemokine receptor type 7 (CCR7), upregulation of MHC I, stabilization of MHC II expression, and increased capacity to prime naive T cells. According to the latest concept, DC activation is a prerequisite for communication with other cells, and depending on the stimuli encountered, they can initiate either tolerogenic or immunogenic immune responses (1). Immunogenic DCs promote the differentiation of naive T cells into effector subsets, such as T helper (Th) 1 and Th17 cells, thereby they are highly efficient in facilitating adaptive immunity against invading pathogens and tumors. In contrast, tolerogenic DCs support the development of regulatory T cells (Tregs), which exert suppressive functions, and are essential for maintaining immune tolerance and prevention of autoimmunity (16).
Nevertheless, altered distribution and aberrant activation of DCs might lead to the breakdown of immune tolerance against self-antigens and result in the induction of autoimmune disorders such as systemic lupus erythematosus (SLE), rheumatoid arthritis (RA), and psoriasis (17). Consequently, modulation of DC functionality has emerged as a promising therapeutic approach to treat autoimmune conditions.
3 Effects of plant-derived compounds on DCs
Natural bioactive compounds are commonly found in healthy foods such as fruits and vegetables, as well as in beverages like herbal and green teas, which are consumed daily by millions of people (4). This chapter focuses on ten well-studied plant-derived compounds that have gained attention due to their anti-inflammatory, antimicrobial and antioxidant properties. Numerous studies have investigated the impact of these phytochemicals on various compartments of the immune system. Here, we aimed to highlight the effects of these selected natural compounds on DCs and introduce the general mechanisms underlying their activities. Their immunomodulatory effects, mechanisms of action on DCs, and predicted targets based on in silico studies are summarized in Table 2.
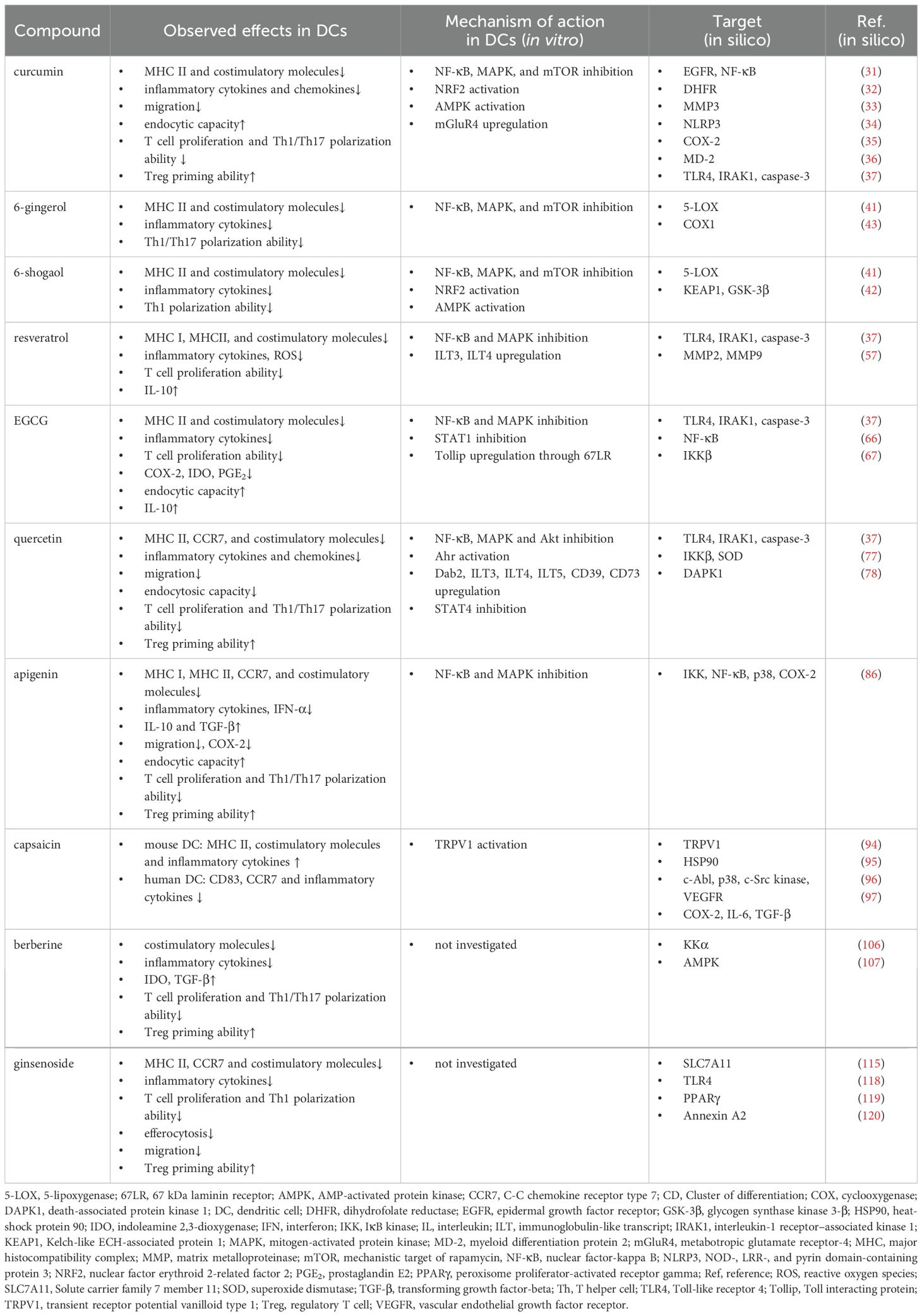
Table 2. The immunomodulatory effects, the mechanisms of action on DCs, and predicted molecular targets of selected phytochemicals.
3.1 Curcumin
Turmeric (Curcuma longa), a member of the Zingiberaceae family, has been used as both a culinary spice and traditional herbal medicine for thousands of years in India and Eastern Asia (18). Curcumin, the main bioactive compound in its rhizome, has been extensively studied for its diverse pharmacological properties over the last few decades. Several in vitro and in vivo animal studies, as well as clinical trials, have demonstrated its anticancer, anti-inflammatory, and radioprotective properties, as summarized in a recent review (19).
Curcumin is one of the most thoroughly investigated phytochemicals, with its effect on DCs first reported in 2005 (20). In murine bone marrow-derived DCs (BM-DCs) stimulated with the TLR4 ligand lipopolysaccharide (LPS), curcumin significantly decreased the expression of MHC II and the costimulatory molecules CD80 and CD86 (20). Furthermore, it suppressed the LPS-triggered production of various inflammatory cytokines such as interleukin (IL)-12, IL-1β, IL-6 and tumor necrosis factor (TNF), and reduced the capacity of DCs to elicit a Th1 response. Interestingly, curcumin-treated DCs exhibited increased endocytic capacity, suggesting that curcumin maintains DCs in a resting state. These effects were associated with curcumin-mediated inhibition of central signaling pathways such as the nuclear factor-kappa B (NF-κB) and mitogen activated protein kinase (MAPK) cascades in DCs. In particular, pre-treatment with curcumin prevented the LPS-triggered nuclear translocation of NF-κB and the phosphorylation of the p38, c-Jun N-terminal Kinase (JNK) and extracellular signal-regulated kinase (ERK) (20). Similar results were observed in human moDCs, where pre-treatment with curcumin inhibited the LPS and polyinosinic:polycytidylic acid (polyI:C)-induced upregulation of different activation markers (CD83, CD86, and HLA-DR) and the secretion of various cytokines including IL-12, TNF and IL-10 (21). Furthermore, curcumin reduced endocytosis and the ability of DCs to prime T cell proliferation. Additionally, curcumin significantly reduced the production of the chemokine CXCL10 and prevented the migration of DCs in response to CCL19 and CCL21, further suggesting its immunosuppressive potential (21). Further studies confirmed that curcumin promotes the development of tolerogenic DCs. Curcumin-treated murine BM-DCs generated from B6 mice show increased IL-10 production, and support the differentiation of Tregs from naive CD4+ T cells (22). Additionally, transforming growth factor-β (TGF-β) and retinoic acid produced by curcumin-treated DCs were also required to induce Treg differentiation.
Mechanistically, curcumin induces an anti-inflammatory phenotype in DCs by activating nuclear factor-erythroid 2-related factor 2 (NRF2) and promoting the expression of heme oxygenase 1 (HO-1) (23, 24). Brück et al. showed that curcumin induced HO-1 expression and signal transducer and activator of transcription 3 (STAT3) phosphorylation in LPS-activated mouse BM-DCs thereby inhibiting their ability to promote Th1 and Th17 polarization through repression of IL-12b and IL-23a transcription. Similarly, curcumin was also able to inhibit the functionality of LPS-stimulated human moDCs through upregulating the immunomodulatory enzyme, HO-1 (24). In a subsequent study the authors also demonstrated that upregulation of HO-1 is dependent on the activation of AMP-activated protein kinase (AMPK). Generally, AMPK negatively regulates the mammalian target of rapamycin (mTOR) signaling cascade, thus inhibiting the glycolytic reprogramming required for full DC activation (25). Furthermore, curcumin inhibited Th17 differentiation by suppressing the production of IL-6 and IL-23 in mouse BM-DCs (26). This effect might be associated with the upregulation of metabotropic glutamate receptor-4 (mGluR4), a molecule known to favor Treg development, and thus inhibit autoimmunity (26).
It was also demonstrated that curcumin-treated human moDCs and murine CD11c+ DCs promote Treg differentiation and expansion both in vitro and in vivo (27). These findings were corroborated by in vivo experiments demonstrating that Tregs generated in the presence of curcumin-treated DCs mitigated Th1-mediated colitis in mice (22). In a mouse model of inflammatory bowel disease (IBD), curcumin alleviated disease symptoms as evidenced by reduced colonic damage and decreased inflammatory cell infiltration to the colonic mucosa (28). Moreover, curcumin not only decreased the number of CD11c+/MHC II+ DCs in the Peyer patches, but also reduced the level of different costimulatory molecules on their surface (28). Additionally, tetramethylcurcumin, an analog of curcumin, was shown to stimulate the release of immunosuppressive extracellular vesicles (EVs) from BM-DCs cultured with ovalbumin (29). Intranasal administration of these EVs induced Treg differentiation in a mouse model of allergic rhinitis thereby alleviating disease symptoms and inflammation (29). In patients with psoriasis, curcumin reduced the expression of IL-17, GM-CSF and IFN-γ in PBMCs further supporting its therapeutic relevance (30).
In silico studies have elucidated potential molecular targets that may underlie the anti-inflammatory and antioxidant effects of curcumin. Strong binding to both epidermal growth factor receptor (EGFR) and NF-κB suggests a synergistic blockade of upstream receptor-mediated and downstream transcriptional inflammatory signaling (31). Inhibition of EGFR signaling can attenuate pro-inflammatory cellular responses by reducing activation of MAPK and STAT pathways, while suppression of NF-κB activity decreases transcription of inflammatory cytokines. Molecular docking results indicate that curcumin binds to dihydrofolate reductase (DHFR) with an affinity comparable to methotrexate, a clinically established immunosuppressive agent for RA (32). Curcumin also shows binding potential to matrix metalloproteinase 3 (MMP-3), a protease whose expression and activity are elevated in chronic inflammatory diseases, including RA (33). MMP3 inhibition may prevent extracellular matrix degradation and inflammatory tissue damage. Docking studies have revealed that curcumin can inhibit the NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3) inflammasome, potentially reducing caspase-1 activation and subsequent IL-1β maturation (34). In silico analyses of 15 curcumin analogues show binding to multiple subdomains of cyclooxygenase-2 (COX-2), a pivotal enzyme in prostaglandin synthesis during inflammation (35). Molecular docking and dynamics simulations suggest that curcumin can embed into the hydrophobic pocket of myeloid differentiation protein 2 (MD-2), a co-receptor for TLR4 that mediates lipopolysaccharide (LPS)-induced signaling, thereby dampening TLR4-mediated activation (36). Further, in silico investigations suggest that curcumin along with quercetin, EGCG, and, resveratrol can inhibit TLR4-mediated signaling through directly interacting with TLR4 and IL-1 receptor-associated kinase 1 (IRAK1), a kinase downstream of TLR-4 (37). These compounds were further predicted to interact with caspase-3, which is primarily known for its role in apoptosis but also plays a significant role in regulating inflammation (37).
3.2 Gingerols and shogaols
Ginger (Zingiber officinale), like turmeric, belongs to the Zingiberaceae family and has a long history of use as both a spice and medicinal plant. Its rhizome is rich in biologically active compounds, such as gingerols and shogaols, which are primarily responsible for its therapeutic effects. 6-gingerol is the principal active compound in fresh ginger, whereas 6-shogaol predominates in dried preparations (38). While numerous studies have shown that gingerols and shogaols influence the function of various immune cell types, only two reports have explored their effects on DCs.
Our group has recently demonstrated that both 6-gingerol and 6-shogaol modulate the functionality of human moDCs (39). Both phenolic compounds decreased the expression of CD40, CD83, CD86 and HLA-DQ, as well as reduced the secretion of TNF, IL-6 and IL-10 cytokines by TLR-stimulated moDCs. Furthermore, both compounds could significantly reduce the inflammatory cytokine production in moDCs triggered by Escherichia coli, and thus their capacity to promote Th1 differentiation from naive CD4+ T cells. Our mechanistic studies revealed that 6-gingerol and 6-shogaol interfered with the TLR-mediated activation of NF-κB, MAPK and mTOR signaling cascades in moDCs. Further, we demonstrated that 6-shogaol, but not 6-gingerol, enhanced AMPK phosphorylation, and activated the NRF2/HO-1 axis suggesting its superior anti-inflammatory potential compared to 6-gingerol (39).
Another study demonstrated that 6-gingerol induced a tolerogenic phenotype in LPS-stimulated mouse BM-DCs as indicated by low expression of MHC II and costimulatory molecules, reduced cytokine secretion, and impaired ability to prime Th17 cells. In agreement with our results, it was also demonstrated that 6-gingerol suppressed DC functionality by modulating the NF-κB and MAPK signaling pathways in LPS-stimulated mouse BM-DCs. In vivo, 6-gingerol significantly ameliorated the severity of experimental autoimmune encephalomyelitis (EAE), a murine model of multiple sclerosis (MS), by inhibiting DC activation and Th17 polarization. Furthermore, 6-gingerol significantly inhibited inflammatory cell infiltration and demyelination in the central nervous system, and lowered the frequencies of CD11c+CD80+ activated DCs and Th17 cells in the spleen (40).
In silico studies have identified several molecular targets that may contribute to the anti-inflammatory and antioxidant properties of 6-gingerol and 6-shogaol. Both compounds can interact with 5-lipoxygenase (5-LOX) (41), an enzyme responsible for leukotriene biosynthesis, suggesting a role in suppressing lipid mediator–driven inflammation. Thiophene derivatives of 6-shogaol have been predicted to activate the NRF2 antioxidant pathway by binding to key regulators, including Kelch-like ECH-associated protein 1 (KEAP1) and glycogen synthase kinase 3-β (GSK-3β) that might lead to the upregulation of cytoprotective and anti-inflammatory genes (42). Additionally, 6-gingerol derivatives showed binding ability to cyclooxygenase-1 (COX-1) (43), indicating potential to reduce prostaglandin-mediated inflammatory responses.
3.3 Resveratrol
Resveratrol is a polyphenol that has various sources such as berries, grapes, red wine and peanuts. When sold as a food supplement, resveratrol is predominantly extracted from Japanese knotweed (Polygonum cuspidatum), a plant traditionally used in East Asian medicine (44). Accumulating evidence suggests its anti-inflammatory and immunomodulatory effects, including its protective role in respiratory system diseases such as asthma and chronic obstructive pulmonary disease (COPD), partly through its action on DCs (45).
In 2004, Kim et al. demonstrated that resveratrol inhibited LPS-mediated activation of BM-DCs in vitro. Resveratrol significantly reduced the production of IL-12 and the surface expression of CD80, CD86, MHC I and MHC II molecules, while preserving the endocytic capacity of DCs. Consequently, these DCs also exhibited decreased T cell stimulatory capacity (46). Similar results were obtained when resveratrol was added to human moDCs (47, 48). Specifically, resveratrol decreased the nuclear translocation of NF-κB p65, the production of IL-12 as well as the expression of CD40, CD80 and CD86 costimulatory molecules in LPS-stimulated human moDCs (47). In contrast, resveratrol markedly upregulated immunoglobulin-like transcript (ILT) 3 and ILT4, which deliver inhibitory signals and promote a tolerogenic phenotype in DCs (47). Therefore, resveratrol-treated moDCs were poor stimulators of CD4+ T cell proliferation and migration. Interestingly, resveratrol-treated moDCs promoted the proliferation of IL-10 secreting T cells but failed to induce forkhead box P3 (FOXP3) expression. Comparable findings were reported in TNF-stimulated moDCs (48), where resveratrol reduced NF-κB p65 nuclear translocation, CD83 and CD86 expression, and IL-12 and IL-23 production, while increasing IL-10 levels. Furthermore, resveratrol also suppressed the capacity of TNF-stimulated DCs to initiate CD3+ T cell proliferation. Resveratrol also blocked moDC activation in response to advanced glycation end products (AGEs) (49), known to contribute to the pathogenesis of autoimmune diseases via DC activation (50). In particular, resveratrol inhibited the expression of various activation markers, costimulatory molecules, and the receptor for AGE (RAGE) and suppressed the production of inflammatory cytokines in moDCs stimulated with glycated albumin. It also reduced the activation of the MAPK and NF-κB pathways, and impaired the allostimulatory potential of moDCs (49).
A recent study demonstrated that the immunomodulatory effect of resveratrol depends highly on its structural features and mode of delivery. Hydroxylated and methylated derivatives are characterized by different biological activities compared to the parent compound (51). Resveratrol and its hydroxylated derivative, piceatannol, potently inhibited the LPS-induced production of reactive oxygen species (ROS), whereas the methylated monomers of resveratrol showed a reduced antioxidant capacity in mouse BM-DCs (51). These compounds also differ in their potential to elicit cytokine production by BM-DCs in response to LPS and bacteria implying the importance of structure with regards to the immunomodulatory potential of resveratrol. Another study demonstrated that resveratrol encapsulated in nanostructured lipid carriers (NLC) more effectively suppressed TNF-induced CD83 expression, IL-12 and IL-23 production, and NF-κB p65 phosphorylation than free resveratrol in moDCs (52).
Subsequent studies further demonstrated that resveratrol can alleviate lung inflammation by modulating DC functions. In vivo, resveratrol ameliorated acute lung injury (ALI) in mice challenged intratracheally with LPS (53). Resveratrol pre-treatment greatly reduced the levels of pro-inflammatory cytokines, including, TNF, IL-6, IL-12, whereas increased the levels of anti-inflammatory cytokines, including TGF-β, IL-10, IL-13 and IL-33 in the bronchoalveolar lavage fluid (BALF) of ALI mice. Resveratrol dramatically decreased the expression of CD80, CD86 and MHC II, the production of IL-12, while increased ILT3 expression and IL-10 secretion in pulmonary and splenic cDCs of ALI mice. In vitro, resveratrol pretreatment of BM-DCs inhibited LPS-induced activation, cytokine production, and T cell stimulatory capacity of DCs as well (53).
Human moDCs differentiated from monocytes of chronic obstructive pulmonary disease (COPD) patients expressed increased levels of CD80, CD86 and IFN-α as compared with those of healthy individuals (54). Resveratrol pre-treatment reduced the level of these molecules, possibly via downregulation microRNA-34, an important regulator of inflammatory responses (55).
A subsequent study investigated how the stability, bioavailability and thus the bioactivity of resveratrol could be enhanced by irradiation. Intriguingly, γ-irradiated resveratrol has a lower toxicity compared to its intact form, and has strong immunosuppressive properties as it can significantly inhibit inflammatory cytokine production, costimulatory molecule expression, and antigen-presentation by LPS-activated mouse BM-DCs (56). It also promoted IL-10 production and Treg production when added to differentiating BM-DCs.
Oral administration of γ-irradiated resveratrol attenuated the clinical signs of colitis in DSS-treated mice, suggesting the therapeutic potential of γ-irradiated resveratrol in IBD (56). However, the lack of direct functional comparison of γ-irradiated resveratrol to its unmodified form limits conclusions regarding its superior ability to induce tolerogenic DCs.
In silico studies have revealed that TLR4, IRAK1, and caspase-3 are molecular targets of resveratrol (37). In addition, MMP-2 and MMP-9 have been reported as targets, whose inhibition may reduce extracellular matrix degradation and attenuate inflammation (57).
3.4 Epigallocatechin-3-gallate
Green tea (Camellia sinensis) is a popular beverage that is consumed worldwide for its health benefits. EGCG is a flavonoid that represents the major bioactive compound in green tea and is known for its strong antioxidant capacity (58). Moreover, it has gained a great attention for its anti-inflammatory and immunomodulatory potential.
The immunomodulatory effect of EGCG has been first investigated on mouse BM-DCs (59). This study showed that EGCG effectively inhibits DC functions, as it was able to inhibit IL-12 production and downregulate the expression of CD80, CD86, MHC I and MHC II molecules in LPS-stimulated murine BM-DCs. EGCG-treated DCs were poor inducers of T cell proliferation and activation. The authors suggested that EGCG antagonized the LPS-mediated functionality of DCs by suppressing MAPK and NF-κB activation. Another study demonstrated that EGCG can inhibit IL-12 production while increasing TNF secretion in BM-derived DCs stimulated with LPS, muramyl-dipeptide or Legionella pneumophila (60). However, the divergent effects of EGCG on IL-12 and TNF production have not been elucidated yet. In the same year, it was also shown that EGCG pretreatment suppressed COX-2 expression, prostaglandin E2 (PGE2) and indoleamine 2,3-dioxygenase (IDO) production in BM-DCs in response to IFN-γ, likely through STAT1 inhibition (61). In human moDCs, EGCG exhibited similar anti-inflammatory effects by attenuating TLR4-mediated signaling. In LPS-stimulated moDCs, EGCG reduced the expression of HLA-DR, CD80 and CD83, and impaired their ability to promote T cell proliferation (62). Conversely, EGCG increased IL-10 production and the endocytic ability of LPS-stimulated DCs suggesting that EGCG maintains DCs in their resting state (62). Experiments performed with murine BM-DCs further demonstrated that the 67 kDa laminin receptor (67LR) is essential for mediating the anti-inflammatory actions of EGCG in DCs (63). EGCG reduced the LPS-induced production of IL-1β, IL-6 and TNF, the expression of CD80, CD86, MHC I and MHC II molecules, and NF-κB and MAPK activation in murine BM-DCs. Interestingly, the inhibitory effect of EGCG was abrogated upon pre-treatment with anti-67LR antibodies, suggesting that the inhibitory actions of EGCG are mediated through 67LR. In addition, EGCG elevated the expression of Toll-interacting protein (Tollip), a negative regulator of TLR signaling through 67LR suggesting that the anti-inflammatory actions of EGCG might be partially mediated by Tollip upregulation in DCs.
Previously it was reported that EGCG”Me, a 3-O-methylated derivative of EGCG, significantly reduced TLR4 expression and thereby exerted anti-inflammatory effects in mouse peritoneal macrophages (64). On the contrary, EGCG3”Me supplementation increased TLR5 expression on lamina propria DCs and macrophages, and enhanced vaccine-induced immune response in mice immunized with a split influenza vaccine (65). Although, EGCG treatment showed overall beneficial effects, the contradiction between its immunosuppressive and immunostimulatory effects warrants further investigation.
Molecular docking and modeling revealed that EGCG can directly bind to TLR4, IRAK1 and caspase-3 (37). In silico studies further predicted NF-κB as a principal target of EGCG, suggesting that its anti-inflammatory effects may be mediated through suppression of NF-κB activation and downstream cytokine production (66). Furthermore, EGCG was found to interact with IκB kinase β (IKKβ), a key regulator of NF-κB signaling, indicating that EGCG may inhibit the phosphorylation and degradation of IκB, thereby preventing NF-κB nuclear translocation and inflammatory gene transcription (67).
3.5 Quercetin
Quercetin is a flavonoid found in various fruits, vegetables, and medicinal plants. It exhibits a broad range of bioactivities, including antioxidant and anti-inflammatory effects, and has been suggested to alleviate allergy symptoms (68). Several studies indicate that quercetin decreases inflammation primarily by inhibiting DC activation. It also acts as a natural, albeit indirect, ligand for the aryl hydrocarbon receptor (Ahr), which plays a key role in regulating immune responses and promoting tolerogenic properties in DCs (69).
In vitro studies demonstrated that quercetin significantly reduced the expression of MHC II and different costimulatory molecules (CD40, CD80, CD86) as well as the production of various cytokines (IL-1 α/β, IL-6, IL-10, IL-12) and chemokines (MCP-1, MIP-1α/β, RANTES) in LPS-stimulated mouse BM-DCs (70). Quercetin also blocked endocytosis by resting DCs and suppressed LPS-stimulated migration of DCs both in vitro and in vivo. Furthermore, quercetin abrogated T cell activation and proliferation induced by LPS-stimulated BM-DCs. The study also demonstrated that quercetin blocked the LPS-triggered activation of ERK, JNK, Akt and the degradation of IκB indicating that quercetin suppresses DC activation via interfering with the MAPK, Akt and NF-κB signaling pathways (70).
In human moDCs, quercetin impaired the LPS-mediated production of IL-12 and the upregulation of CD83, CD86, HLA-DR and CCR7 (71). It was revealed that quercetin downregulated CD83 through facilitating direct binding of Ahr to the CD83 promoter region. In addition, quercetin decreased the ability of DCs to activate T cells, whereas increased their potential to initiate Treg differentiation in coculture with naive T cells. The data showed that quercetin induced a tolerogenic phenotype in DCs via upregulating various immunomodulatory molecules, including Disabled-2 (Dab2), ILT3, ILT4 and ILT5 inhibitory receptors, and the ATP-degrading ectoenzymes CD39 and CD73 (71). Blockade of Dab2, a negative regulator of intracellular signaling reversed the inhibitory effects of quercetin on BMDC activation suggesting that the regulatory effects of quercetin might be mediated via Dab2 upregulation (72).
Co-administration with piperine, a known bioenhancer, was shown to augment the anti-inflammatory potential of quercetin (73). Genome-wide transcriptome analysis revealed that quercetin and piperine delivered via reconstituted oil bodies (ROBs-QP) significantly downregulated various inflammatory mediators, and decreased the expression of molecules associated with antigen presentation and activation in mouse BM-DCs (74). DCs exposed to ROBs-QP failed to upregulated CCR7, migrate to lymph nodes and efficiently present antigens to naive T cells (74). Intraperitoneal delivery of ROBs-QP also ameliorated DSS-induced colitis symptoms in mice highlighting its potential for treating inflammatory diseases (73). Another report showed that quercetin administration reduced atherosclerosis progression in apolipoprotein E knock out mice by suppressing DC activation (72). Immunohistochemical analysis revealed that quercetin reduced DC and macrophage accumulation in atherosclerotic lesions, and decreased serum IL-6 and IL-12, while increased IL-10 levels. In vivo, quercetin also alleviated contact hypersensitivity response elicited via injection of 2,4-dinitrofluorobenzene (DNFB)-pulsed DCs to mice, indicating that quercetin could be used to prevent delayed-type hypersensitivity (70). Furthermore, a recent study demonstrated that quercetin is also able to suppress neuroinflammation in EAE mice by inhibiting DC activation and Th1/Th17 cell differentiation (75). The experimental results suggest that this effect was mediated through inhibition of STAT4 (75), a transcription factor central to DC activation and implicated in autoimmune disease pathogenesis (76).
These findings demonstrate that quercetin efficiently inhibits LPS-induced DC activation, and attenuates different types of inflammatory reactions in vivo, suggesting its potential as a therapeutic agent to treat various DC-mediated inflammatory conditions.
Molecular docking analysis revealed that quercetin has considerable binding affinity to IKKβ, a core component of the NF-κB signaling pathway, as well as to the antioxidant enzyme superoxide dismutase (SOD) (77), suggesting that its anti-inflammatory effects are mediated largely through suppression of NF-κB signaling and modulation of oxidative stress. Furthermore, in silico studies have indicated Death-Associated Protein Kinase 1 (DAPK1) as a potential molecular target for quercetin and its analogs, implicating possible neuroprotective and anti-inflammatory roles through modulation of DAPK1 activity (78).
3.6 Apigenin
Apigenin is a ubiquitous flavonoid synthetized by many different types of plants. One of the richest natural sources of apigenin is chamomile (Matricaria recutita), which has been traditionally consumed as an herbal tea to reduce anxiety and treat gastrointestinal complaints (79). Like many flavonoids, apigenin has also been reported to reduce inflammation and oxidative stress (80). Apigenin also has an impact on the immune system by modulating the biological activities of various immune cells, including DCs.
In mouse BM-DCs, apigenin significantly suppressed the LPS-induced production of IL-12 and the expression of CD80, CD86, MHC I, and MHC II molecules (81). In addition, apigenin-treated DCs showed reduced ability to initiate allogeneic T cell proliferation and Th1 differentiation, while displayed high endocytic capacity suggesting that apigenin maintained DCs in their resting state. Apigenin also blocked the NF-κB and MAPK signaling pathways in mouse BM-DCs (81). It also suppressed TLR7- and TLR9-mediated IL-6 and IFN-α production by spleen DCs of lupus mice (82). In human blood derived DCs, apigenin reduced the LPS-triggered expression of CD40, CD83, CD86, CCR7, and MHC I and MHC II molecules, as well as production of IL-1β, IL-6, IL-12 and IL-23 while inducing secretion of IL-10 and TGF-β (83). Consequently, apigenin decreased the ability of LPS-triggered DCs to induce Th1 and Th17 polarization while increased the number of Tregs. In LPS-stimulated DCs, apigenin prevented the nuclear translocation of RelB, a transcription factor, which is involved in the non-canonical activation of the NF-κB pathway and thus in the control of DC activation (83).
In vivo, apigenin suppressed splenic DC activation and IFN-γ production of splenic CD4+ T cells in LPS-challenged mice and decreased the ability of trinitrobenzene sulfonic acid (TNBS)-pulsed murine BM-DCs to induce contact hypersensitivity (81). In SNF1 mice, a spontaneous lupus mouse model, apigenin reduced COX-2 expression in immune cells including DCs (82). Apigenin also reduced nucleosome-induced IFN-γ and IL-17 response by splenic T cells and autoantibody production of splenic B cells of SNF1 mice. Intraperitoneal injection of apigenin also decreased serum autoantibody levels and delayed the development of glomerulonephritis. In collagen induced arthritis (CIA) mice, an animal model of RA, apigenin reduced joint inflammation, swelling and destruction, most likely via inhibition of DC functions (84). In CIA mice, apigenin reduced levels of TNF, IL-1β and IL-6 in the serum and supernatants from the lymph nodes, and blocked DC activation as shown by reduced expression of costimulatory molecules and MHC II in CD11c+ cells. Apigenin also reduced DC migration to the draining lymph nodes of CIA mice (84). These data indicate that apigenin exerts its immunosuppressive effect in arthritis by inhibiting DC activation and migration. Apigenin also reduced the severity of EAE in vivo both in progressive (C57BL/6) and relapse-remitting (SJL/J) mouse models of MS. Apigenin decreased immune cell infiltration and reduced demyelination in the CNS of EAE mice while retaining immune cells in the periphery including the blood, spleen and lymph nodes. Apigenin increased the number of CD11c+ DCs in all 3 peripheral compartments, and downmodulated the levels of MHC II and CD86 on splenocyte-derived DCs (83). It also reduced the surface expression of α4 integrin on the surface of splenic DCs and CD4+ T cell from EAE mice, thereby decreasing their ability to cross the blood-brain barrier (85).
A recent in silico study have further identified the molecular targets of apigenin involved in inflammatory pathways, including IKK, the NF-κB p50–p65 heterodimer, p38 MAPK, and COX-2 (86). By modulating these key signaling molecules, apigenin might exert broad anti-inflammatory effects through inhibition of NF-κB activation, MAPK signaling, and prostaglandin synthesis, thereby contributing to the regulation of DC functions.
3.7 Capsaicin
Chili pepper (Capsicum annuum) is a commonly used spice that has also various health-promoting attributes such as anti-inflammatory, antioxidant, and antimicrobial properties (87). Capsaicin is a naturally occurring alkaloid in chili pepper responsible for its pungent taste, as well as many of its health benefits. Although, several studies have reported beneficial effects of capsaicin in various autoimmune diseases (reviewed in (88)), its effects on DCs remained controversial and its immunomodulatory mechanisms are not completely elucidated.
Initial studies demonstrated that mouse BM-DCs express the capsaicin receptor transient receptor potential channel vanilloid type 1 (TRPV1), and engagement of this receptor by capsaicin promotes DC activation (89). Capsaicin increased surface expression of MHC II and CD86 that could be inhibited by the TRPV1 antagonist capsazepine. Intradermal injection of capsaicin led to the migration of DCs to the draining lymph nodes in TRPV1 expressing mice, but not in TRPV1 deficient animals (89). Studies in human DCs also confirmed TRPV1 expression; however, unlike in mouse BM-DCs, capsaicin exerted anti-inflammatory effects in them (90). In particular, capsaicin dose-dependently decreased the expression of CD83 and CCR7, endocytosis of Escherichia coli, and the production of IL-6 and IL-12 by moDCs stimulated with pro-inflammatory cytokines. The divergent response between species might arise from dosage differences. The dose of capsaicin was 150 µM in the mouse study, while it was only 1 µM in the study performed with human DCs. A subsequent study showed that TRPV1 functions as a calcium channel and induces the release of calcitonin-gene related peptide (CGRP) upon capsaicin exposure in mouse splenic DCs (91). CGRP attenuated LPS-stimulated CD11c+ splenic DC responses, as it was shown by their reduced expression of CD80/CD86, decreased production of TNF and increased release of IL-10. Capsaicin also significantly reduced IFN-γ secretion in whole spleen cell cultures under Th1 polarizing conditions. These data suggest that capsaicin might contribute to immune regulation via CGRP-mediated suppression of DC activation (91).
Recent preclinical studies indicate that capsaicin treatment is able to ameliorate autoimmune disease symptoms. In an experimental model of autoimmune neuropathy, orally administered capsaicin reduced sciatic nerve demyelination and inflammatory cell infiltration when given prophylactically (92). Similarly, 0.075% capsaicin skin cream alleviated DSS-induced colitis symptoms including colon shortening, diarrhea and weight loss in mice while improved their epithelial barrier integrity and gut microbiota composition (93).
It has been known for many years that capsaicin binds to TRPV1, specifically within a ligand-binding pocket formed by transmembrane segments (94). Experimental evidence also indicates that capsaicin might interact with the ATP binding site of molecular chaperone heat-shock protein 90 (Hsp90), a key protein that stabilizes and activates signaling molecules involved in inflammation, such as NF-κB and MAPK pathways. By inhibiting Hsp90, capsaicin may disrupt these pro-inflammatory signaling cascades, thereby reducing inflammatory responses (95). Molecular docking studies suggest that capsaicin may also interact with other signaling proteins, including Abelson tyrosine-protein kinase (c-Abl), c-Src kinase, p38 MAP kinase, and VEGF receptor, that all play key roles in propagating inflammatory signals (96). In silico studies further predict that capsaicin can bind to and potentially modulate the activity of key inflammatory mediators such as COX-2, IL-6, and TGF-β (97), supporting its immunomodulatory properties. These interactions ranging from well-established TRPV1 binding to computationally predicted protein targets underpin the complex role of capsaicin in immune regulation and inflammation.
3.8 Berberine
Berberine is an isoquinoline alkaloid found in some plants like goldenseal, goldthread and Oregon grape. Its most abundant natural source is barberry (Berberis species), traditionally used in Asian medicine to treat fever, infections, digestive disorders, and other pathologies (98). Both in vitro and in vivo studies show that berberine possesses strong anti-inflammatory and immunomodulatory activities, and thus propose the use of berberine as a therapeutic agent for the treatment of inflammatory disorders (99).
In LPS-treated human moDCs, berberine reduced CD40, CD80 and CD86 expression, and lowered IL-1β, IL-6 and TNF production, impairing their capacity to prime Th17 responses (100) In LPS-stimulated mouse BM-DCs, berberine inhibited the secretion of TNF and IL-12, as well as the production of IL-6 and TGF-β that supposedly contributed to the inhibition of Th1 and Th17 polarization, respectively. The authors further suggest that berberine exerts its effect through inhibiting dopamine receptor-mediated signaling pathways (100). In DC2.4 cells, berberine greatly increased the secretion of IDO and TGF-β that might contribute to the ability of DCs to induce Treg differentiation, while decreasing Th17 proliferation (101). Additional data indicate that berberine is able to inhibit Th17 responses both directly and indirectly through repressing DC functions (102).
Berberine-mediated suppression of Th1 and Th17 responses has been further substantiated in animal models of type I diabetes (103) and MS (104). In addition, berberine was also shown to ameliorate DSS-induced colitis symptoms as well as Th1 and Th17 responses in mice (100). In the context of DSS-induced murine ulcerative colitis (UC), berberine inhibited colon damage and restored mucosal barrier homeostasis by inhibiting the infiltration and activation of immune cells (105). Berberine treatment decreased the percentage of inflammatory cells including DCs, Th1 and Th17 cells in the mesenteric lymph nodes and lamina propria of DSS-treated mice. In addition, berberine reduced serum levels of inflammatory cytokines including TNF, IL-1β, IL-6 and IFN-γ, and increased enteric glial cell functions (105). In streptozotocin-induced diabetic retinopathy (DR) berberine lowered serum levels of glucose, TNF, IL-1β, IL-6 and IL-17 (101). In the spleen and lymph nodes of DR mice, berberine reduced the frequency and activation of DCs. In addition, berberine lowered the ratio of Th17/Tregs indicating that berberine can suppress DC activation and influence T cell differentiation as well. These results are in line with a previous report (102), showing that berberine might affect T cell responses both directly and indirectly through DCs.
Recent in silico studies have indicated IKKα, as a primary molecular target of berberine (106). Additionally, berberine demonstrated strong binding affinity to the allosteric sites of AMP-activated protein kinase (AMPK) α- and β-subunits, which could contribute to the modulation of inflammatory responses as well (107). Together, these molecular interactions highlight the potential of berberine to suppress pro-inflammatory signaling and regulate DC functions.
3.9 Ginsenoside
Ginsenosides are a class of steroid glycosides, and triterpene saponins found exclusively in the roots of Panax species such as ginseng (Panax ginseng), traditional used in East Asian to enhance physical and mental performances (108). In the last few decades, the major bioactive compounds of ginseng have been shown to modulate various immune cells including DCs (109).
More than 200 structurally diverse ginsenosides exist, and most of them are categorized in the groups of protopanaxadiol (e.g. Rb1, Rc, Rd, Re, Rg3, Rg5) and protopanaxatriol (e.g. Rg1, F4, Rg6) types of glycosides (110). An early study evaluated 21 different ginsenosides for their ability to modulate LPS-triggered IL-12 production of BM-DCs, finding ginsenosides Rg6 and F4 the most effective (111). Another study demonstrated that ginsenoside Rg1 more effectively reduced LPS-triggered IL-6 and TNF production by murine DC2.4 cells than ginsenoside Rb1 (112). Interestingly, the inhibitory effects of ginsenosides Rg1 and Rb1 were diminished when the two compounds were combined (112). A fraction of ginsenosides containing predominantly Rc, Rg3, Rd and Rb1, decreased CD40, CD80, CD86, and MHC II expression by human LPS-stimulated moDCs (113). These ginsenosides also suppressed the ability of Staphylococcus aureus-primed moDCs to induce naïve CD4+ T cell proliferation and IFN-γ production. Ginsenoside metabolite compound K (CK), the main deglycosylated metabolite of ginsenosides, decreased CD80, CD86 and MHC II expression on mouse BM-DCs as well as their capacity to prime T cell proliferation in vitro (114). Ginsenoside Rg5 could greatly increase the efferocytotic capacity, the clearance of apoptotic cells by BM-DCs from db/db mice, via inhibiting Solute Carrier Family 7 Member 11 (SLC7A11), a negative regulator of efferocytosis (115). The authors further found that ginsenoside Rg5 inhibited SLC7A11 activity via direct binding.
In vivo, ginsenoside Rg5 promoted wound healing in the skin of diabetic (db/db) mice by increasing efferocytosis by DCs (115). Furthermore, CK decreased the proportion of DCs in the lymph nodes of collagen induced arthritis mice by lowering CCL21 levels in the lymph nodes and CCR7 expression on the surface of DCs (114). However, various ginsenosides can alleviate disease symptoms in mouse (116) and rat models (117) of MS; their impact on DCs has remained unexplored.
Molecular docking analyses demonstrated that ginsenoside Rb1 can bind to TLR4 (118), while ginsenoside Rf was identified as a potential ligand for peroxisome proliferator-activated receptor gamma (PPARγ), a nuclear receptor that plays an important role in regulating inflammation by controlling the expression of COX-2 (119). Additionally, molecular docking and thermal shift assays confirmed the interaction between CK and Annexin A2. This interaction prevented Annexin A2 from binding to the NF-κB p50 subunit and their nuclear co-localization, thereby attenuating NF-κB activation and downstream gene transcription (120).
4 Discussion
Since time immemorial, plants have been extensively used to alleviate pain and treat different type of illnesses around the world (121). Traditionally, different parts of plants including roots, leaves, seeds and fruits have been used in herbal remedy preparation. In the last few decades, it has been revealed that the bioactive compounds found in plants, also called phytochemicals, are responsible for their pharmacological actions. Research has also shown that a multitude of plant-derived bioactive compounds have significant promise in preventing and curing chronic illnesses. In addition, many phytochemicals have been recognized for their immunomodulatory activity and significant contribution to the maintenance of the body’s homeostasis (122).
Despite the fact that phytochemicals are widely distributed in fruits, vegetables and herbs, diet is insufficient to reach therapeutic levels, mainly due to their fast metabolism in the gut and liver. Moreover, phytochemicals have limited bioavailability due to their poor solubility and stability. Besides, the dietary intake form may also affect the bioavailability of phytochemicals (123). Therefore, new delivery systems and formulation techniques are under development to increase the stability, the bioavailability, and thus the therapeutic efficacy of plant-derived bioactive compounds (124). Nevertheless, the application of plant-derived compounds for therapeutic purposes is still in its infancy and faces many different challenges. In particular, in vitro studies show that mostly large doses (in the µM range) are required to elicit the beneficial effects of some phytochemicals that is hard to achieve in vivo. Moreover, high concentrations of certain phytochemicals might already have adverse and cytotoxic effects (125). For instance, in murine models, relatively high concentrations of capsaicin promoted DC activation (89), in contrast to the anti-inflammatory effects in human moDCs at markedly lower doses (90). These divergent outcomes likely arise from species-specific differences in DC subset sensitivity and the dose-dependent immune-modulatory properties of capsaicin. Moreover, berberine was shown to induce apoptosis in murine BM-DCs and splenic DCs in a dose-dependent manner (from 2 to 50 µM) while not affecting other immune cell types such as macrophages, B and T cells (126). Actually, this feature of berberine could be exploited for therapeutic purposes. Intraperitoneal injection of berberine to CIA mice alleviated disease symptoms, most probably due to its ability to selectively induce apoptosis of DCs, and thus significantly reduce their proportion in the spleen and lymph nodes of CIA mice (126). These studies underscore the current lack of consensus on the optimal dose of phytochemicals for therapeutic application; therefore, further studies are needed to assess their clinically relevant levels.
The administration route is another factor that needs to be optimized to ensure the effectiveness and safety of plant bioactive compounds. In general, orally delivered phytochemicals are poorly absorbed, since many plant-derived compounds are conjugated with glucuronide, sulfate or glutathione moieties in the gut epithelium or liver, and then excreted in urine and bile (123). Therefore, intraperitoneal administration is preferred over the oral route to avoid the potential degradation of biological agents in in vivo animal experiments (127). In relation to plant-derived compounds, a study showed that intraperitoneal injection of berberin led to a better anti-arthritic effect than its oral administration (126). Nevertheless, intraperitoneal injection is rarely used in the clinics, thus the efficacy of other forms of administration routes such as intravenous or subcutaneous needs to be explored for plant-derived compounds. It is more likely, though, that improving the delivery efficiency into the target tissue by nanoformulation might be the key to overcome these limitations (128). For instance, a recent study shows that gingerol encapsulation within lipid nanoparticles enhanced its stability and thus promoted its osteogenic, chemopreventive, and antibacterial properties in 3D-printed bone scaffolds (129). Another study demonstrated that β-cyclodextrin inclusion technology could enhance the stability and bioavailability of 6-shogaol (130) and curcumin (131) as well. Furthermore, encapsulation of resveratrol in nanostructured lipid carriers not only increased its stability and maintained its activity, but also decreased the necessary dose for inhibiting DC functions (52). The potency of nanocarries in DC-based immunotherapy has been extensively reviewed elsewhere (132, 133).
Another promising strategy to improve the bioavailability and thereby enhance the efficacy of phytochemicals is their combined use. Piperine acts as a broad-spectrum bioenhancer by increasing the intestinal absorption of several phytochemicals, which consequently amplifies their anti-inflammatory effects. For instance, co-administration with piperine has been shown to augment the anti-inflammatory potential of quercetin (73). Specifically, the combined administration of quercetin and piperine in the form of reconstituted oil bodies significantly improved the suppression of inflammatory cytokine production by DCs, even at low doses. Piperin is also documented to increase the bioavailability of curcumin (134). It must also be noted that enhancing the intestinal absorption of phytochemicals such as curcumin is particularly important from the perspective of DCs, as they are abundant in the gut mucosa and actively sample antigens from the intestinal lumen. Consequently, increased local bioavailability of phytochemicals in the gut allows them to directly modulate the function and phenotype of intestinal DCs, potentially shaping mucosal immune responses more effectively. Further studies in murine RAW264.7 cells suggest that ginsenoside Rg1 can synergize with glucocorticoids to enhance their anti-inflammatory effect (135). Additionally, immunomodulatory synergy has been reported between curcumin and capsaicin (97), curcumin and resveratrol (136), as well as EGCG and quercetin (137). Furthermore, curcumin showed synergistic anti-cancer effects when combined with apigenin (138). However, to date, no specific in vitro studies have directly evaluated the effects of these combinations on DCs.
Another challenge associated with phytochemicals is that plant-derived compounds might affect several cell types at the same time when administrated systemically. The phytochemicals introduced in this review exert significant inhibitory effects on DC functions; however, several studies showed that various phytochemicals are able to directly affect other immune cell types such as T cells and macrophages. These data suggest that plant-derived compounds probably target ubiquitous cellular signaling pathways that might lead to unexpected or undesired pharmacological effects. Although, in vivo animal studies showed that several phytochemicals might be effective in the treatment of autoimmune diseases associated with aberrant DC activation, the precise molecular mechanism underlying their activity needs to be explored.
In silico analyses have already predicted multiple molecular targets for these phytochemicals through approaches such as molecular docking and direct binding simulations. Notably, molecular targets include several components of the NF-κB signaling cascade, as well as key pro-inflammatory enzymes such as LOX and COX, whose activity can be attenuated through direct ligand binding. In addition, several of these phytochemicals play a crucial role in activating the antioxidant system through direct binding to NRF2, SOD and inhibiting the Keap1. Such interactions offer a plausible mechanistic basis for the anti-inflammatory and anti-oxidant phenotypes documented in vitro and in vivo. Furthermore, these observations underscore the complex interplay between phytochemicals and multiple signaling pathways, highlighting the necessity of further studies to precisely define their molecular targets and mechanisms of action. Understanding these multifaceted interactions will be crucial for harnessing phytochemicals as targeted therapeutics for immune-mediated diseases while minimizing their off-target effects.
Accumulating evidence indicates that multiple plant-derived compounds can modulate DC activity, including cytokine secretion and their T cell-priming capacity (Figure 1). Therefore, phytochemicals could be utilized to the generation of tolerogenic DCs for DC-based therapies that holds significant potential for the treatment of autoimmune diseases. Although, numerous protocols have been developed to manipulate the functionality of DCs prior to re-introduction into patients, to date, clinical outcomes have not been satisfactory. There are still many challenges in DC therapy that needs to be resolved such as the delivery root, optimal cell number and type of DC to be used. Additionally, a prerequisite for tolerogenic DCs is to maintain their suppressive phenotype in inflammatory environments. Therefore, the protocols for generating tolerogenic DCs also include an activation step using LPS or a cocktail of inflammatory cytokines. This also contributes to the upregulation of CCR7, which is necessary to the migration of DCs to the lymph nodes and thus to the modulation of T cell responses. Nevertheless, tolerogenic DCs generated with the immunosuppressive agents such as vitamin D3, dexamethasone or rapamycin have reduced CCR7 levels and thus impaired capacity to migrate to lymph nodes (16). Although, migration is a significant attribute of DC functionality, relatively low number of studies examined the effects of plant-derived compounds on DC migration. Given the variability and abundance of biologically active compounds in plants, it is also conceivable, that some phytochemicals are able to induce tolerogenic characteristics in DC without inhibiting their migratory potential. It is also important to mention that similar to most DC clinical trials, studies examining the effect of phytochemicals on human DCs have been carried out with moDCs, which have an inherent low migratory ability and thus rather orchestrate local immune responses (9, 16, 139). In comparison to moDCs, cDCs have a superior capacity to migrate to lymph nodes and present antigens to naive T cells (140). Thus, in future experimental studies it would be desirable to investigate the effects of plant-derived compounds on circulating DC types, especially on conventional DCs, which might serve as a more potent alternative to moDCs for DC vaccination (140).
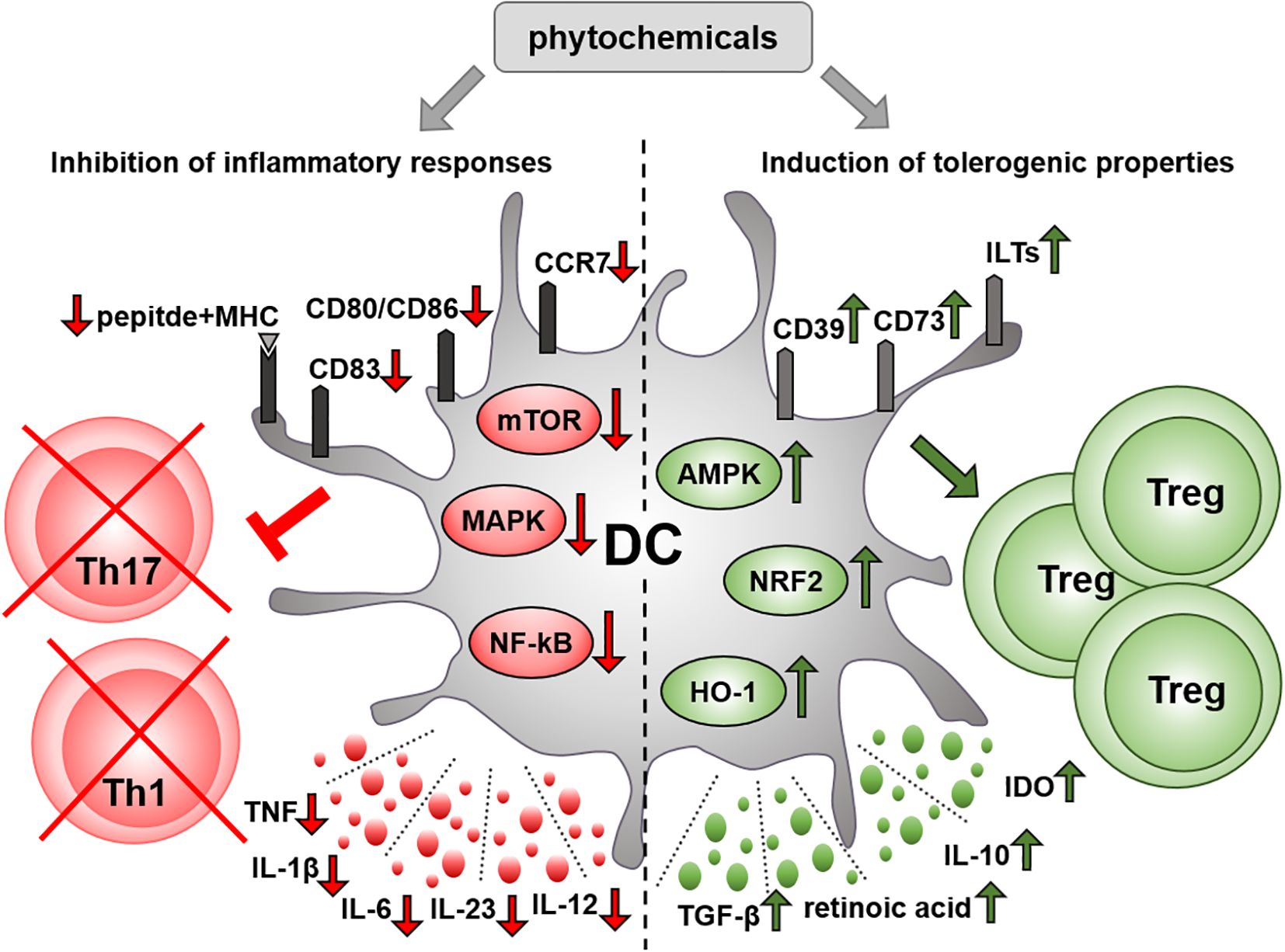
Figure 1. Immunomodulatory effect of phytochemicals on DCs. Various phytochemicals can inhibit the immunostimulatory function of DCs through multiple mechanisms. On the one hand, phytochemicals may impair the ability of DCs to respond to inflammatory stimuli. In particular, they can suppress the production of inflammatory mediators, and downregulate the expression of costimulatory and antigen-presenting molecules by interfering with key inflammatory signaling pathways in DCs. On the other hand, plant-derived bioactive compounds may promote a tolerogenic phenotype in DCs, characterized by upregulated expression of tolerogenic cell surface markers and increased production of anti-inflammatory mediators. Several phytochemicals suppress the capacity of DCs to induce inflammatory T helper cell subsets, such as Th1 and Th17, while increasing their ability to drive the differentiation of regulatory T cells. AMPK, AMP-activated protein kinase; CD, cluster of differentiation; DC, dendritic cell; HO-1, heme oxygenase 1; IDO, indoleamine 2,3-dioxygenase, IL, interleukin; ILT, immunoglobulin-like transcript; MAPK, mitogen activated protein kinase; MHC, major histocompatibility complex; mTOR, mammalian target of rapamycin; NF-κB, nuclear factor-kappa B; NRF2, nuclear factor-erythroid 2-related factor 2; TGF, transforming growth factor; Th, T helper cell; TNF, tumor necrosis factor; Treg, regulatory T cell.
In conclusion, plant-derived bioactive compounds might open up new avenues in the treatment of autoimmune diseases. Several clinical trials of phytochemicals including curcumin, EGCG, resveratrol, apigenin and berberine for the treatment of various autoimmune diseases have been recently completed or are still ongoing that indicates the potential clinical benefits of these compounds (Table 3.). Alternatively, several plant-derived bioactive compounds might serve as potential tools for the generation of DC-based vaccines (Figure 2). In addition to the phytochemicals introduced in this review, many others might have the ability to modulate DC responses that could be exploited for therapeutic purposes. For instance, a recent study demonstrated that even a rose flavor compound, namely β-damascone, was also able to suppress DC-mediated immune responses, and thus to ameliorate contact hypersensitivity in mice (141). All these data imply that the plant kingdom is one of the richest sources of bioactive compounds with pharmaceutical activity, and holds a great potential for the discovery of new therapeutic agents.
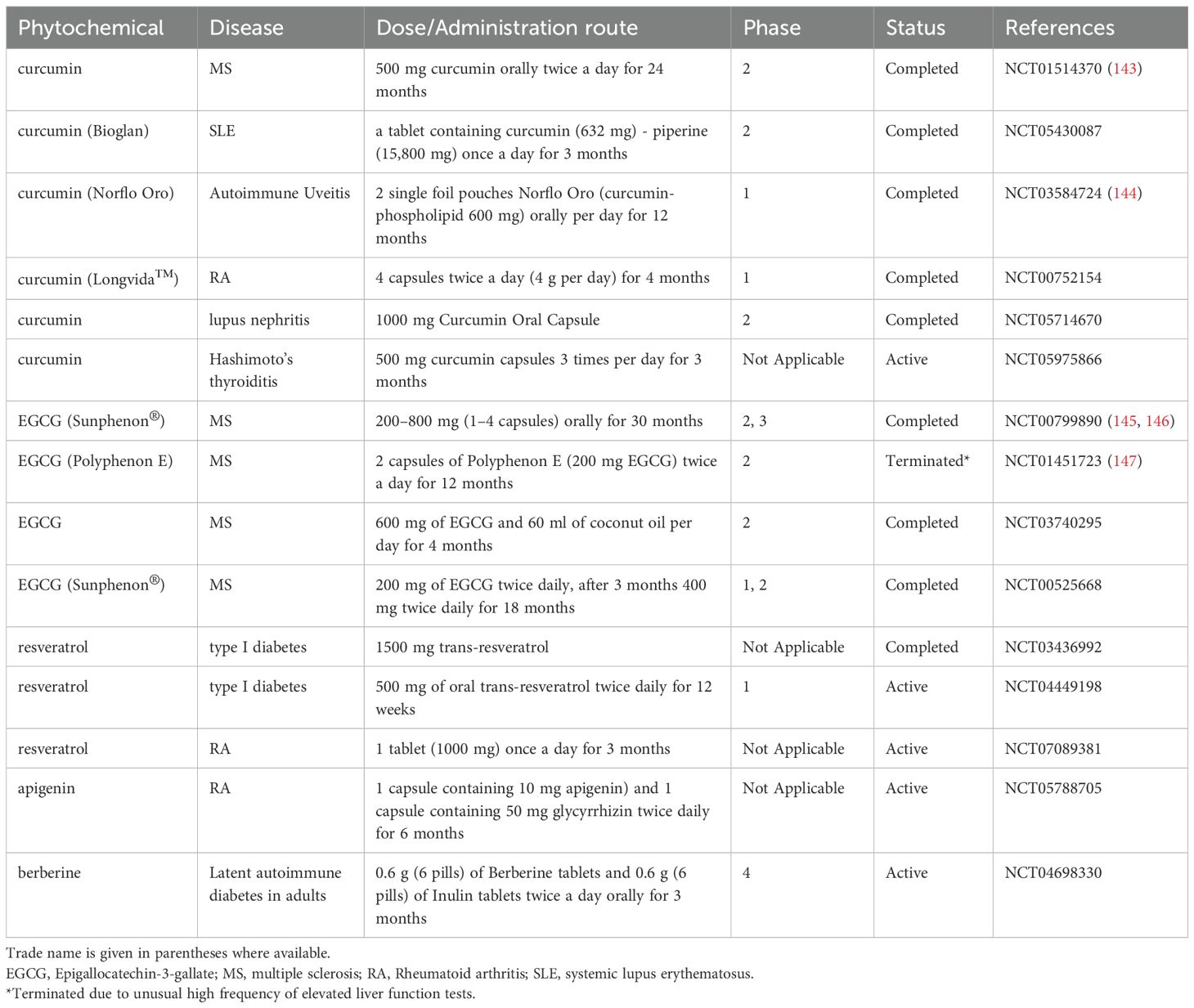
Table 3. Phytochemicals for autoimmune diseases in clinical trials.
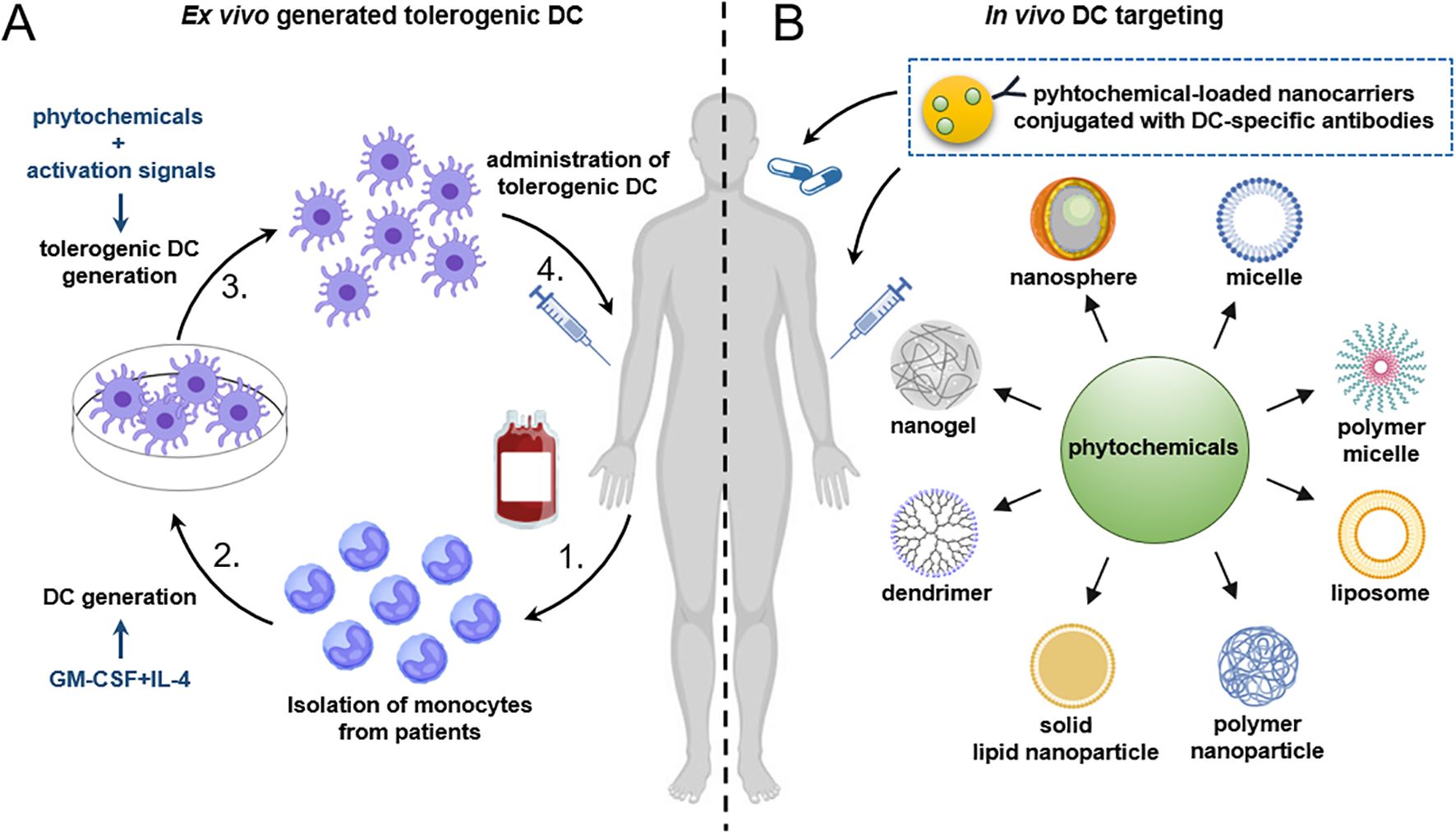
Figure 2. Future strategies for the generation of tolerogenic DCs using plant-derived compounds. (A) For the ex vivo generation of tolerogenic DCs, CD14+ monocytes are isolated from peripheral blood of patients with autoimmune diseases and then differentiated into DCs in the presence of GM-CSF and IL-4. A tolerogenic state in DCs can be achieved by the addition of plant-derived bioactive compounds. To ensure the stability of tolerogenic DCs under inflammatory conditions, activation stimuli may also be applied. Alternatively, DCs can be pulsed with disease-relevant antigens as well. Finally, the tolerogenic DCs are reintroduced into the patient. (B) A promising future strategy involves the in vivo targeting of DCs using nanoparticles loaded with phytochemicals alone, or in combination with autoantigen-associated peptides. Alternatively, nanoparticles might be loaded with antibodies against specific surface antigens expressed on DCs to enhance their targeting specificity. Finally, nanoparticles loaded with immunomodulatory agents such as phytochemicals can be administered intravenously, subcutaneously, or via non-invasive routes such as oral delivery. DC, dendritic cell; GM-CSF, granulocyte-macrophage colony-stimulating factor; IL-4, interleukin-4.
Author contributions
TF: Conceptualization, Writing – original draft, Writing – review & editing. KP: Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by the National Research, Development and Innovation Office (NKFIH PD 135193 to TF and FK 142782 to KP). Supported by the University of Debrecen Program for Scientific Publication.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Cabeza-Cabrerizo M, Cardoso A, Minutti CM, Pereira da Costa M, and Reis e Sousa C. Dendritic cells revisited. Annu Rev Immunol. (2021) 39:131–66. doi: 10.1146/annurev-immunol-061020-053707
2. Bergonzi MC, Heard CM, and Garcia-Pardo J. Bioactive molecules from plants: discovery and pharmaceutical applications. Pharmaceutics. (2022) 14. doi: 10.3390/pharmaceutics14102116
3. Al-Khayri JM, Rashmi R, Toppo V, Chole PB, Banadka A, Sudheer WN, et al. Plant secondary metabolites: the weapons for biotic stress management. Metabolites. (2023) 13. doi: 10.3390/metabo13060716
4. Sorrenti V, Buro I, Consoli V, and Vanella L. Recent advances in health benefits of bioactive compounds from food wastes and by-products: biochemical aspects. Int J Mol Sci. (2023) 24. doi: 10.3390/ijms24032019
5. Steinman RM and Cohn ZA. Identification of a novel cell type in peripheral lymphoid organs of mice. II. Functional properties in vitro. J Exp Med. (1974) 139:380–97. doi: 10.1084/jem.139.2.380
6. Steinman RM and Cohn ZA. Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J Exp Med. (1973) 137:1142–62. doi: 10.1084/jem.137.5.1142
7. Collin M and Bigley V. Human dendritic cell subsets: an update. Immunology. (2018) 154:3–20. doi: 10.1111/imm.12888
8. Bencze D, Fekete T, and Pazmandi K. Type I interferon production of plasmacytoid dendritic cells under control. Int J Mol Sci. (2021) 22. doi: 10.3390/ijms22084190
9. Backer RA, Probst HC, and Clausen BE. Classical DC2 subsets and monocyte-derived DC: Delineating the developmental and functional relationship. Eur J Immunol. (2023) 53:e2149548. doi: 10.1002/eji.202149548
10. Sallusto F and Lanzavecchia A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J Exp Med. (1994) 179:1109–18. doi: 10.1084/jem.179.4.1109
11. Doebel T, Voisin B, and Nagao K. Langerhans cells - the macrophage in dendritic cell clothing. Trends Immunol. (2017) 38:817–28. doi: 10.1016/j.it.2017.06.008
12. Crozat K, Guiton R, Guilliams M, Henri S, Baranek T, Schwartz-Cornil I, et al. Comparative genomics as a tool to reveal functional equivalences between human and mouse dendritic cell subsets. Immunol Rev. (2010) 234:177–98. doi: 10.1111/j.0105-2896.2009.00868.x
13. Dalod M and Scheu S. Dendritic cell functions in vivo: A user's guide to current and next- generation mutant mouse models. Eur J Immunol. (2022) 52:1712–49. doi: 10.1002/eji.202149513
14. Wilson NS, El-Sukkari D, and Villadangos JA. Dendritic cells constitutively present self antigens in their immature state in vivo and regulate antigen presentation by controlling the rates of MHC class II synthesis and endocytosis. Blood. (2004) 103:2187–95. doi: 10.1182/blood-2003-08-2729
15. Silva Z, Ferro T, Almeida D, Soares H, Ferreira JA, Deschepper FM, et al. MHC class I stability is modulated by cell surface sialylation in human dendritic cells. Pharmaceutics. (2020) 12. doi: 10.3390/pharmaceutics12030249
16. Mansilla MJ, Hilkens CMU, and Martinez-Caceres EM. Challenges in tolerogenic dendritic cell therapy for autoimmune diseases: the route of administration. Immunother Adv. (2023) 3:ltad012. doi: 10.1093/immadv/ltad012
17. Ganguly D, Haak S, Sisirak V, and Reizis B. The role of dendritic cells in autoimmunity. Nat Rev Immunol. (2013) 13:566–77. doi: 10.1038/nri3477
18. Hewlings SJ and Kalman DS. Curcumin: A review of its effects on human health. Foods. (2017) 6. doi: 10.3390/foods6100092
19. Cozmin M, Lungu II, Gutu C, Stefanache A, Duceac LD, Soltuzu BD, et al. Turmeric: from spice to cure. A review of the anti-cancer, radioprotective and anti-inflammatory effects of turmeric sourced compounds. Front Nutr. (2024) 11:1399888. doi: 10.3389/fnut.2024.1399888
20. Kim GY, Kim KH, Lee SH, Yoon MS, Lee HJ, Moon DO, et al. Curcumin inhibits immunostimulatory function of dendritic cells: MAPKs and translocation of NF-kappa B as potential targets. J Immunol. (2005) 174:8116–24. doi: 10.4049/jimmunol.174.12.8116
21. Shirley SA, Montpetit AJ, Lockey RF, and Mohapatra SS. Curcumin prevents human dendritic cell response to immune stimulants. Biochem Biophys Res Commun. (2008) 374:431–6. doi: 10.1016/j.bbrc.2008.07.051
22. Cong Y, Wang L, Konrad A, Schoeb T, and Elson CO. Curcumin induces the tolerogenic dendritic cell that promotes differentiation of intestine-protective regulatory T cells. Eur J Immunol. (2009) 39:3134–46. doi: 10.1002/eji.200939052
23. Bruck J, Holstein J, Glocova I, Seidel U, Geisel J, Kanno T, et al. Nutritional control of IL-23/Th17-mediated autoimmune disease through HO-1/STAT3 activation. Sci Rep. (2017) 7:44482. doi: 10.1038/srep44482
24. Campbell NK, Fitzgerald HK, Malara A, Hambly R, Sweeney CM, Kirby B, et al. Naturally derived Heme-Oxygenase 1 inducers attenuate inflammatory responses in human dendritic cells and T cells: relevance for psoriasis treatment. Sci Rep. (2018) 8:10287. doi: 10.1038/s41598-018-28488-6
25. Everts B and Pearce EJ. Metabolic control of dendritic cell activation and function: recent advances and clinical implications. Front Immunol. (2014) 5:203. doi: 10.3389/fimmu.2014.00203
26. Zhao G, Liu Y, Yi X, Wang Y, Qiao S, Li Z, et al. Curcumin inhibiting Th17 cell differentiation by regulating the metabotropic glutamate receptor-4 expression on dendritic cells. Int Immunopharmacol. (2017) 46:80–6. doi: 10.1016/j.intimp.2017.02.017
27. Rogers NM, Kireta S, and Coates PT. Curcumin induces maturation-arrested dendritic cells that expand regulatory T cells in vitro and in vivo. Clin Exp Immunol. (2010) 162:460–73. doi: 10.1111/j.1365-2249.2010.04232.x
28. Zhao HM, Xu R, Huang XY, Cheng SM, Huang MF, Yue HY, et al. Curcumin suppressed activation of dendritic cells via JAK/STAT/SOCS signal in mice with experimental colitis. Front Pharmacol. (2016) 7:455. doi: 10.3389/fphar.2016.00455
29. Mo LH, Han HY, Jin QR, Song YN, Wu GH, Zhang Y, et al. T cell activator-carrying extracellular vesicles induce antigen-specific regulatory T cells. Clin Exp Immunol. (2021) 206:129–40. doi: 10.1111/cei.13655
30. Snyder JP and Amiel E. Regulation of dendritic cell immune function and metabolism by cellular nutrient sensor mammalian target of rapamycin (mTOR). Front Immunol. (2018) 9:3145. doi: 10.3389/fimmu.2018.03145
31. Saeed MEM, Yucer R, Dawood M, Hegazy MF, Drif A, Ooko E, et al. In silico and in vitro screening of 50 curcumin compounds as EGFR and NF-kappaB inhibitors. Int J Mol Sci. (2022) 23. doi: 10.3390/ijms23073966
32. Hobani Y, Jerah A, and Bidwai A. A comparative molecular docking study of curcumin and methotrexate to dihydrofolate reductase. Bioinformation. (2017) 13:63–6. doi: 10.6026/97320630013063
33. Jerah A, Hobani Y, Kumar BV, and Bidwai A. Curcumin binds in silico to anti-cancer drug target enzyme MMP-3 (human stromelysin-1) with affinity comparable to two known inhibitors of the enzyme. Bioinformation. (2015) 11:387–92. doi: 10.6026/97320630011387
34. Jena AB, Dash UC, and Duttaroy AK. An in silico investigation on the interactions of curcumin and epigallocatechin-3-gallate with NLRP3 Inflammasome complex. Biomedicine pharmacotherapy = Biomedecine pharmacotherapie. (2022) 156:113890. doi: 10.1016/j.biopha.2022.113890
35. Sohilait MR, Pranowo HD, and Haryadi W. Molecular docking analysis of curcumin analogues with COX-2. Bioinformation. (2017) 13:356–9. doi: 10.6026/97320630013356
36. Wang Z, Chen G, Chen L, Liu X, Fu W, Zhang Y, et al. Insights into the binding mode of curcumin to MD-2: studies from molecular docking, molecular dynamics simulations and experimental assessments. Mol bioSystems. (2015) 11:1933–8. doi: 10.1039/C5MB00085H
37. Andres CMC, Munguira EB, Juan CA, Lobo F, and Perez-Lebena E. Perez de la Lastra JM. In Silico Exploration of Natural Antioxidants for Sepsis Drug Discovery. Molecules. (2025) 30. doi: 10.3390/molecules30112288
38. Pazmandi K, Szollosi AG, and Fekete T. The "root" causes behind the anti-inflammatory actions of ginger compounds in immune cells. Front Immunol. (2024) 15:1400956. doi: 10.3389/fimmu.2024.1400956
39. Pazmandi K, Agics B, Szollosi AG, Bacsi A, and Fekete T. Ginger-derived bioactive compounds attenuate the Toll-like receptor mediated responses of human dendritic cells. Eur J Pharmacol. (2024) 967:176399. doi: 10.1016/j.ejphar.2024.176399
40. Han JJ, Li X, Ye ZQ, Lu XY, Yang T, Tian J, et al. Treatment with 6-gingerol regulates dendritic cell activity and ameliorates the severity of experimental autoimmune encephalomyelitis. Mol Nutr Food Res. (2019) 63:e1801356. doi: 10.1002/mnfr.201801356
41. Ley-Martinez JS, Ortega-Valencia JE, Garcia-Barradas O, Jimenez-Fernandez M, Uribe-Lam E, Vencedor-Meraz CI, et al. Active compounds in zingiber officinale as possible redox inhibitors of 5-lipoxygenase using an in silico approach. Int J Mol Sci. (2022) 23. doi: 10.3390/ijms23116093
42. Mak KK, Shiming Z, Sakirolla R, Balijepalli MK, Dinkova-Kostova AT, Epemolu O, et al. Synthesis of new shogaol analogues as NRF2 activators and evaluation of their anti-inflammatory activity, modes of action and metabolic stability. Antioxidants. (2023) 12. doi: 10.3390/antiox12020475
43. Ahmed SHH, Gonda T, Agbadua OG, Girst G, Berkecz R, Kusz N, et al. Preparation and evaluation of 6-gingerol derivatives as novel antioxidants and antiplatelet agents. Antioxidants (Basel). (2023) 12. doi: 10.3390/antiox12030744
44. Bensa M, Vovk I, and Glavnik V. Resveratrol food supplement products and the challenges of accurate label information to ensure food safety for consumers. Nutrients. (2023) 15. doi: 10.3390/nu15020474
45. de Sa Coutinho D, Pacheco MT, Frozza RL, and Bernardi A. Anti-inflammatory effects of resveratrol: mechanistic insights. Int J Mol Sci. (2018) 19. doi: 10.3390/ijms19061812
46. Kim GY, Cho H, Ahn SC, Oh YH, Lee CM, and Park YM. Resveratrol inhibits phenotypic and functional maturation of murine bone marrow-derived dendritic cells. Int Immunopharmacol. (2004) 4:245–53. doi: 10.1016/j.intimp.2003.12.009
47. Svajger U, Obermajer N, and Jeras M. Dendritic cells treated with resveratrol during differentiation from monocytes gain substantial tolerogenic properties upon activation. Immunology. (2010) 129:525–35. doi: 10.1111/j.1365-2567.2009.03205.x
48. Silva AM, Oliveira MI, Sette L, Almeida CR, Oliveira MJ, Barbosa MA, et al. Resveratrol as a natural anti-tumor necrosis factor-alpha molecule: implications to dendritic cells and their crosstalk with mesenchymal stromal cells. PloS One. (2014) 9:e91406. doi: 10.1371/journal.pone.0091406
49. Buttari B, Profumo E, Facchiano F, Ozturk EI, Segoni L, Saso L, et al. Resveratrol prevents dendritic cell maturation in response to advanced glycation end products. Oxid Med Cell longevity. (2013) 2013:574029. doi: 10.1155/2013/574029
50. Delrue C, Speeckaert R, Delanghe JR, and Speeckaert MM. The potential influence of advanced glycation end products and (s)RAGE in rheumatic diseases. Int J Mol Sci. (2023) 24. doi: 10.3390/ijms24032894
51. Johnsen PR, Pinna C, Mattio L, Strube MB, Di Nunzio M, Iametti S, et al. Investigation of the effects of monomeric and dimeric stilbenoids on bacteria-induced cytokines and LPS-induced ROS formation in bone marrow-derived dendritic cells. Int J Mol Sci. (2023) 24. doi: 10.3390/ijms24032731
52. Barbosa JP, Neves AR, Silva AM, Barbosa MA, Reis MS, and Santos SG. Nanostructured lipid carriers loaded with resveratrol modulate human dendritic cells. Int J Nanomedicine. (2016) 11:3501–16. doi: 10.2147/IJN.S108694
53. Guo B, Peng Y, Gu Y, Zhong Y, Su C, Liu L, et al. Resveratrol pretreatment mitigates LPS-induced acute lung injury by regulating conventional dendritic cells' maturation and function. Open Life Sci. (2021) 16:1064–81. doi: 10.1515/biol-2021-0110
54. Wang X, Zhang C, Huang G, Han D, Guo Y, Meng X, et al. Resveratrol inhibits dysfunction of dendritic cells from chronic obstructive pulmonary disease patients through promoting miR-34. Int J Clin Exp Pathol. (2015) 8:5145–52.
55. Das K and Rao LVM. The role of microRNAs in inflammation. Int J Mol Sci. (2022) 23. doi: 10.3390/ijms232415479
56. Kim WS, Song HY, Mushtaq S, Kim JM, Byun EH, Yuk JM, et al. Therapeutic potential of gamma-irradiated resveratrol in ulcerative colitis via the anti-inflammatory activity and differentiation of tolerogenic dendritic cells. Cell Physiol Biochem. (2019) 52:1117–38. doi: 10.33594/000000076
57. Pandey AK, Bhattacharya P, Shukla SC, Paul S, and Patnaik R. Resveratrol inhibits matrix metalloproteinases to attenuate neuronal damage in cerebral ischemia: a molecular docking study exploring possible neuroprotection. Neural regeneration Res. (2015) 10:568–75. doi: 10.4103/1673-5374.155429
58. Musial C, Kuban-Jankowska A, and Gorska-Ponikowska M. Beneficial properties of green tea catechins. Int J Mol Sci. (2020) 21. doi: 10.3390/ijms21051744
59. Ahn SC, Kim GY, Kim JH, Baik SW, Han MK, Lee HJ, et al. Epigallocatechin-3-gallate, constituent of green tea, suppresses the LPS-induced phenotypic and functional maturation of murine dendritic cells through inhibition of mitogen-activated protein kinases and NF-kappaB. Biochem Biophys Res Commun. (2004) 313:148–55. doi: 10.1016/j.bbrc.2003.11.108
60. Rogers J, Perkins I, van Olphen A, Burdash N, Klein TW, and Friedman H. Epigallocatechin gallate modulates cytokine production by bone marrow-derived dendritic cells stimulated with lipopolysaccharide or muramyldipeptide, or infected with Legionella pneumophila. Exp Biol Med (Maywood). (2005) 230:645–51. doi: 10.1177/153537020523000906
61. Jeong YI, Jung ID, Lee JS, Lee CM, Lee JD, and Park YM. (-)-Epigallocatechin gallate suppresses indoleamine 2,3-dioxygenase expression in murine dendritic cells: evidences for the COX-2 and STAT1 as potential targets. Biochem Biophys Res Commun. (2007) 354:1004–9. doi: 10.1016/j.bbrc.2007.01.076
62. Yoneyama S, Kawai K, Tsuno NH, Okaji Y, Asakage M, Tsuchiya T, et al. Epigallocatechin gallate affects human dendritic cell differentiation and maturation. J Allergy Clin Immunol. (2008) 121:209–14. doi: 10.1016/j.jaci.2007.08.026
63. Byun EB, Choi HG, Sung NY, and Byun EH. Green tea polyphenol epigallocatechin-3-gallate inhibits TLR4 signaling through the 67-kDa laminin receptor on lipopolysaccharide-stimulated dendritic cells. Biochem Biophys Res Commun. (2012) 426:480–5. doi: 10.1016/j.bbrc.2012.08.096
64. Kumazoe M, Nakamura Y, Yamashita M, Suzuki T, Takamatsu K, Huang Y, et al. Green tea polyphenol epigallocatechin-3-gallate suppresses toll-like receptor 4 expression via up-regulation of E3 ubiquitin-protein ligase RNF216. J Biol Chem. (2017) 292:4077–88. doi: 10.1074/jbc.M116.755959
65. Kumazoe M, Takamatsu K, Horie F, Yoshitomi R, Hamagami H, Tanaka H, et al. Methylated (-)-epigallocatechin 3-O-gallate potentiates the effect of split vaccine accompanied with upregulation of Toll-like receptor 5. Sci Rep. (2021) 11:23101. doi: 10.1038/s41598-021-02346-4
66. Suhail M, Rehan M, Tarique M, Tabrez S, Husain A, and Zughaibi TA. Targeting a transcription factor NF-kappaB by green tea catechins using in silico and in vitro studies in pancreatic cancer. Front Nutr. (2022) 9:1078642. doi: 10.3389/fnut.2022.1078642
67. Wang W, Xiong X, Li X, Zhang Q, Yang W, and Du L. In silico investigation of the anti-tumor mechanisms of epigallocatechin-3-gallate. Molecules. (2019) 24. doi: 10.3390/molecules24071445
68. Mlcek J, Jurikova T, Skrovankova S, and Sochor J. Quercetin and its anti-allergic immune response. Molecules. (2016) 21. doi: 10.3390/molecules21050623
69. Mohammadi-Bardbori A, Bengtsson J, Rannug U, Rannug A, and Wincent E. Quercetin, resveratrol, and curcumin are indirect activators of the aryl hydrocarbon receptor (AHR). Chem Res Toxicol. (2012) 25:1878–84. doi: 10.1021/tx300169e
70. Huang RY, Yu YL, Cheng WC, OuYang CN, Fu E, and Chu CL. Immunosuppressive effect of quercetin on dendritic cell activation and function. J Immunol. (2010) 184:6815–21. doi: 10.4049/jimmunol.0903991
71. Michalski J, Deinzer A, Stich L, Zinser E, Steinkasserer A, and Knippertz I. Quercetin induces an immunoregulatory phenotype in maturing human dendritic cells. Immunobiology. (2020) 225:151929. doi: 10.1016/j.imbio.2020.151929
72. Lin W, Wang W, Wang D, and Ling W. Quercetin protects against atherosclerosis by inhibiting dendritic cell activation. Mol Nutr Food Res. (2017) 61. doi: 10.1002/mnfr.201700031
73. Cavalcanti E, Vadrucci E, Delvecchio FR, Addabbo F, Bettini S, Liou R, et al. Administration of reconstituted polyphenol oil bodies efficiently suppresses dendritic cell inflammatory pathways and acute intestinal inflammation. PloS One. (2014) 9:e88898. doi: 10.1371/journal.pone.0088898
74. Delvecchio FR, Vadrucci E, Cavalcanti E, De Santis S, Kunde D, Vacca M, et al. Polyphenol administration impairs T-cell proliferation by imprinting a distinct dendritic cell maturational profile. Eur J Immunol. (2015) 45:2638–49. doi: 10.1002/eji.201545679
75. Zhou F, Guo YX, Gao R, Ji XY, Tang YX, Wang LB, et al. Quercetin regulates dendritic cell activation by targeting STAT4 in the treatment of experimental autoimmune encephalomyelitis. Toxicol Appl Pharmacol. (2024) 488:116980. doi: 10.1016/j.taap.2024.116980
76. Fukao T, Frucht DM, Yap G, Gadina M, O'Shea JJ, and Koyasu S. Inducible expression of Stat4 in dendritic cells and macrophages and its critical role in innate and adaptive immune responses. J Immunol. (2001) 166:4446–55. doi: 10.4049/jimmunol.166.7.4446
77. Bastin A, Teimouri M, Faramarz S, Shabani M, Doustimotlagh AH, and Sadeghi A. In vitro and molecular docking analysis of quercetin as an anti-inflammatory and antioxidant. Curr Pharm design. (2023) 29:883–91. doi: 10.2174/1381612829666230330084043
78. Sun Y, Zhao J, Lu Y, Ngo FY, Shuai B, Zhang ZJ, et al. In silico prediction of quercetin analogs for targeting death-associated protein kinase 1 (DAPK1) against alzheimer's disease. Curr neuropharmacology. (2024) 22:2353–67. doi: 10.2174/1570159X22666240515090434
79. Dai YL, Li Y, Wang Q, Niu FJ, Li KW, Wang YY, et al. Chamomile: A review of its traditional uses, chemical constituents, pharmacological activities and quality control studies. Molecules. (2022) 28. doi: 10.3390/molecules28010133
80. Singh A, Singh J, Parween G, Khator R, and Monga V. A comprehensive review of apigenin a dietary flavonoid: biological sources, nutraceutical prospects, chemistry and pharmacological insights and health benefits. Crit Rev Food Sci Nutr. (2025) 65:4529–65. doi: 10.1080/10408398.2024.2390550
81. Yoon MS, Lee JS, Choi BM, Jeong YI, Lee CM, Park JH, et al. Apigenin inhibits immunostimulatory function of dendritic cells: Implication of immunotherapeutic adjuvant. Mol Pharmacol. (2006) 70:1033–44. doi: 10.1124/mol.106.024547
82. Kang HK, Ecklund D, Liu M, and Datta SK. Apigenin, a non-mutagenic dietary flavonoid, suppresses lupus by inhibiting autoantigen presentation for expansion of autoreactive Th1 and Th17 cells. Arthritis Res Ther. (2009) 11:R59. doi: 10.1186/ar2682
83. Ginwala R, Bhavsar R, Moore P, Bernui M, Singh N, Bearoff F, et al. Apigenin modulates dendritic cell activities and curbs inflammation via relB inhibition in the context of neuroinflammatory diseases. J Neuroimmune Pharmacol. (2021) 16:403–24. doi: 10.1007/s11481-020-09933-8
84. Li X, Han Y, Zhou Q, Jie H, He Y, Han J, et al. Apigenin, a potent suppressor of dendritic cell maturation and migration, protects against collagen-induced arthritis. J Cell Mol Med. (2016) 20:170–80. doi: 10.1111/jcmm.12717
85. Ginwala R, McTish E, Raman C, Singh N, Nagarkatti M, Nagarkatti P, et al. Apigenin, a natural flavonoid, attenuates EAE severity through the modulation of dendritic cell and other immune cell functions. J Neuroimmune Pharmacol. (2016) 11:36–47. doi: 10.1007/s11481-015-9617-x
86. Peanlikhit T, Aryal U, Welsh JS, Shroyer KR, and Rithidech KN. Evaluation of the inhibitory potential of apigenin and related flavonoids on various proteins associated with human diseases using autoDock. Int J Mol Sci. (2025) 26. doi: 10.3390/ijms26062548
87. Azlan A, Sultana S, Huei CS, and Razman MR. Antioxidant, anti-obesity, nutritional and other beneficial effects of different chili pepper: A review. Molecules. (2022) 27. doi: 10.3390/molecules27030898
88. Deng Y, Huang X, Wu H, Zhao M, Lu Q, Israeli E, et al. Some like it hot: The emerging role of spicy food (capsaicin) in autoimmune diseases. Autoimmun Rev. (2016) 15:451–6. doi: 10.1016/j.autrev.2016.01.009
89. Basu S and Srivastava P. Immunological role of neuronal receptor vanilloid receptor 1 expressed on dendritic cells. Proc Natl Acad Sci United States America. (2005) 102:5120–5. doi: 10.1073/pnas.0407780102
90. Toth BI, Benko S, Szollosi AG, Kovacs L, Rajnavolgyi E, and Biro T. Transient receptor potential vanilloid-1 signaling inhibits differentiation and activation of human dendritic cells. FEBS letters. (2009) 583:1619–24. doi: 10.1016/j.febslet.2009.04.031
91. Assas MB, Wakid MH, Zakai HA, Miyan JA, and Pennock JL. Transient receptor potential vanilloid 1 expression and function in splenic dendritic cells: a potential role in immune homeostasis. Immunology. (2016) 147:292–304. doi: 10.1111/imm.12562
92. Motte J, Ambrosius B, Gruter T, Bachir H, Sgodzai M, Pedreiturria X, et al. Capsaicin-enriched diet ameliorates autoimmune neuritis in rats. J Neuroinflammation. (2018) 15:122. doi: 10.1186/s12974-018-1165-x
93. Santos EA, Silva JL, Leocadio PCL, Andrade MER, Queiroz-Junior CM, Oliveira NSS, et al. Cutaneous application of capsaicin cream reduces clinical signs of experimental colitis and repairs intestinal barrier integrity by modulating the gut microbiota and tight junction proteins. ACS Pharmacol Transl Sci. (2024) 7:2143–53. doi: 10.1021/acsptsci.4c00207
94. Yang F, Xiao X, Cheng W, Yang W, Yu P, Song Z, et al. Structural mechanism underlying capsaicin binding and activation of the TRPV1 ion channel. Nat Chem Biol. (2015) 11:518–24. doi: 10.1038/nchembio.1835
95. Patwardhan CA, Kommalapati VK, Llbiyi T, Singh D, Alfa E, Horuzsko A, et al. Capsaicin binds the N-terminus of Hsp90, induces lysosomal degradation of Hsp70, and enhances the anti-tumor effects of 17-AAG (Tanespimycin). Sci Rep. (2023) 13:13790. doi: 10.1038/s41598-023-40933-9
96. Kunjiappan S, Sankaranarayanan M, Karan Kumar B, Pavadai P, Babkiewicz E, Maszczyk P, et al. Capsaicin-loaded solid lipid nanoparticles: design, biodistribution, in silico modeling and in vitro cytotoxicity evaluation. Nanotechnology. (2021) 32:095101. doi: 10.1088/1361-6528/abc57e
97. Vasanthkumar T, Hanumanthappa M, and Lakshminarayana R. Curcumin and capsaicin modulates LPS induced expression of COX-2, IL-6 and TGF-beta in human peripheral blood mononuclear cells. Cytotechnology. (2019) 71:963–76. doi: 10.1007/s10616-019-00338-x
98. Neag MA, Mocan A, Echeverria J, Pop RM, Bocsan CI, Crisan G, et al. Berberine: botanical occurrence, traditional uses, extraction methods, and relevance in cardiovascular, metabolic, hepatic, and renal disorders. Front Pharmacol. (2018) 9:557. doi: 10.3389/fphar.2018.00557
99. Kalmarzi RN, Naleini SN, Ashtary-Larky D, Peluso I, Jouybari L, Rafi A, et al. Anti-inflammatory and immunomodulatory effects of barberry (Berberis vulgaris) and its main compounds. Oxid Med Cell Longev. (2019) 2019:6183965. doi: 10.1155/2019/6183965
100. Kawano M, Takagi R, Kaneko A, and Matsushita S. Berberine is a dopamine D1- and D2-like receptor antagonist and ameliorates experimentally induced colitis by suppressing innate and adaptive immune responses. J Neuroimmunol. (2015) 289:43–55. doi: 10.1016/j.jneuroim.2015.10.001
101. Yang Y, Wen Z, Zhang Y, Li P, Zhao J, Sun Y, et al. Berberine alleviates diabetic retinopathy by regulating the Th17/Treg ratio. Immunol letters. (2024) 267:106862. doi: 10.1016/j.imlet.2024.106862
102. Yang Y, Qi J, Wang Q, Du L, Zhou Y, Yu H, et al. Berberine suppresses Th17 and dendritic cell responses. Invest Ophthalmol Visual science. (2013) 54:2516–22. doi: 10.1167/iovs.12-11217
103. Cui G, Qin X, Zhang Y, Gong Z, Ge B, and Zang YQ. Berberine differentially modulates the activities of ERK, p38 MAPK, and JNK to suppress Th17 and Th1 T cell differentiation in type 1 diabetic mice. J Biol Chem. (2009) 284:28420–9. doi: 10.1074/jbc.M109.012674
104. Qin X, Guo BT, Wan B, Fang L, Lu L, Wu L, et al. Regulation of Th1 and Th17 cell differentiation and amelioration of experimental autoimmune encephalomyelitis by natural product compound berberine. J Immunol. (2010) 185:1855–63. doi: 10.4049/jimmunol.0903853
105. Li H, Fan C, Lu H, Feng C, He P, Yang X, et al. Protective role of berberine on ulcerative colitis through modulating enteric glial cells-intestinal epithelial cells-immune cells interactions. Acta Pharm Sin B. (2020) 10:447–61. doi: 10.1016/j.apsb.2019.08.006
106. Reddi KK, Li H, Li W, and Tetali SD. Berberine, A phytoalkaloid, inhibits inflammatory response induced by LPS through NF-kappabeta pathway: possible involvement of the IKKalpha. Molecules. (2021) 26. doi: 10.3390/molecules26164733
107. Kakoti BB, Zothantluanga JH, Deka K, Halder RK, and Roy D. In silico design and computational screening of berberine derivatives for potential antidiabetic activity through allosteric activation of the AMPK pathway. In silico Pharmacol. (2025) 13:12. doi: 10.1007/s40203-024-00295-0
108. Ratan ZA, Haidere MF, Hong YH, Park SH, Lee JO, Lee J, et al. Pharmacological potential of ginseng and its major component ginsenosides. J Ginseng Res. (2021) 45:199–210. doi: 10.1016/j.jgr.2020.02.004
109. Kang S and Min H. Ginseng, the 'Immunity boost': the effects of panax ginseng on immune system. J Ginseng Res. (2012) 36:354–68. doi: 10.5142/jgr.2012.36.4.354
110. Chen W, Balan P, and Popovich DG. Comparison of ginsenoside components of various tissues of New Zealand forest-grown asian ginseng (Panax ginseng) and american ginseng (Panax quinquefolium L.). Biomolecules. (2020) 10. doi: 10.3390/biom10030372
111. Tung NH, Quang TH, Son JH, Koo JE, Hong HJ, Koh YS, et al. Inhibitory effect of ginsenosides from steamed ginseng-leaves and flowers on the LPS-stimulated IL-12 production in bone marrow-derived dendritic cells. Arch Pharm Res. (2011) 34:681–5. doi: 10.1007/s12272-011-0419-2
112. Rhule A, Rase B, Smith JR, and Shepherd DM. Toll-like receptor ligand-induced activation of murine DC2.4 cells is attenuated by Panax notoginseng. J ethnopharmacology. (2008) 116:179–86. doi: 10.1016/j.jep.2007.11.019
113. Lee YJ, Son YM, Gu MJ, Song KD, Park SM, Song HJ, et al. Ginsenoside fractions regulate the action of monocytes and their differentiation into dendritic cells. J Ginseng Res. (2015) 39:29–37. doi: 10.1016/j.jgr.2014.07.003
114. Chen J, Wu H, Wang Q, Chang Y, Liu K, and Wei W. Ginsenoside metabolite compound K suppresses T-cell priming via modulation of dendritic cell trafficking and costimulatory signals, resulting in alleviation of collagen-induced arthritis. J Pharmacol Exp Ther. (2015) 353:71–9. doi: 10.1124/jpet.114.220665
115. Xia W, Zhu Z, Xiang S, and Yang Y. Ginsenoside Rg5 promotes wound healing in diabetes by reducing the negative regulation of SLC7A11 on the efferocytosis of dendritic cells. J Ginseng Res. (2023) 47:784–94. doi: 10.1016/j.jgr.2023.06.006
116. Oh J, Kwon TW, Choi JH, Kim Y, Moon SK, Nah SY, et al. Ginsenoside-Re inhibits experimental autoimmune encephalomyelitis as a mouse model of multiple sclerosis by downregulating TLR4/MyD88/NF-kappaB signaling pathways. Phytomedicine: Int J phytotherapy phytopharmacology. (2024) 122:155065.
117. Lee MJ, Jang M, Choi J, Chang BS, Kim DY, Kim SH, et al. Korean red ginseng and ginsenoside-rb1/-rg1 alleviate experimental autoimmune encephalomyelitis by suppressing th1 and th17 cells and upregulating regulatory T cells. Mol Neurobiol. (2016) 53:1977–2002. doi: 10.1007/s12035-015-9131-4
118. Gao H, Kang N, Hu C, Zhang Z, Xu Q, Liu Y, et al. Ginsenoside Rb1 exerts anti-inflammatory effects in vitro and in vivo by modulating toll-like receptor 4 dimerization and NF-kB/MAPKs signaling pathways. Phytomedicine: Int J phytotherapy phytopharmacology. (2020) 69:153197. doi: 10.1016/j.phymed.2020.153197
119. Song H, Park J, Choi K, Lee J, Chen J, Park HJ, et al. Ginsenoside Rf inhibits cyclooxygenase-2 induction via peroxisome proliferator-activated receptor gamma in A549 cells. J ginseng Res. (2019) 43:319–25. doi: 10.1016/j.jgr.2018.11.007
120. Wang YS, Zhu H, Li H, Li Y, Zhao B, and Jin YH. Ginsenoside compound K inhibits nuclear factor-kappa B by targeting Annexin A2. J ginseng Res. (2019) 43:452–9. doi: 10.1016/j.jgr.2018.04.002
121. Petrovska BB. Historical review of medicinal plants' usage. Pharmacogn Rev. (2012) 6:1–5. doi: 10.4103/0973-7847.95849
122. Vieira SF, Reis RL, Ferreira H, and Neves NM. Plant-derived bioactive compounds as key players in the modulation of immune-related conditions. Phytochem Rev. (2025) 24:343–460. doi: 10.1007/s11101-024-09955-7
123. Selby-Pham SNB, Miller RB, Howell K, Dunshea F, and Bennett LE. Physicochemical properties of dietary phytochemicals can predict their passive absorption in the human small intestine. Sci Rep. (2017) 7:1931. doi: 10.1038/s41598-017-01888-w
124. Kyriakoudi A, Spanidi E, Mourtzinos I, and Gardikis K. Innovative delivery systems loaded with plant bioactive ingredients: formulation approaches and applications. Plants (Basel). (2021) 10. doi: 10.3390/plants10061238
125. Chan MM, Chen R, and Fong D. Targeting cancer stem cells with dietary phytochemical - Repositioned drug combinations. Cancer Lett. (2018) 433:53–64. doi: 10.1016/j.canlet.2018.06.034
126. Hu Z, Jiao Q, Ding J, Liu F, Liu R, Shan L, et al. Berberine induces dendritic cell apoptosis and has therapeutic potential for rheumatoid arthritis. Arthritis Rheumatol. (2011) 63:949–59. doi: 10.1002/art.30202
127. Al Shoyaib A, Archie SR, and Karamyan VT. Intraperitoneal route of drug administration: should it be used in experimental animal studies? Pharm Res. (2019) 37:12. doi: 10.1007/s11095-019-2745-x
128. Han HS, Koo SY, and Choi KY. Emerging nanoformulation strategies for phytocompounds and applications from drug delivery to phototherapy to imaging. Bioact Mater. (2022) 14:182–205. doi: 10.1016/j.bioactmat.2021.11.027
129. Bose S, White B, and Chaudhari VS. Gingerol encapsulated lipid nanoparticles on 3D-printed scaffolds for orthopedic and dental applications. Biomaterials advances. (2025) 172:214194. doi: 10.1016/j.bioadv.2025.214194
130. Li R, Bao R, Yang QX, Wang QL, Adu-Frimpong M, Wei QY, et al. 6]-Shogaol/beta-CDs inclusion complex: preparation, characterisation, in vivo pharmacokinetics, and in situ intestinal perfusion study. J microencapsulation. (2019) 36:500–12. doi: 10.1080/02652048.2019.1649480
131. Longwei J, Xia N, Wang F, Xie C, Ye R, Tang H, et al. Preparation and characterization of curcumin/β-cyclodextrin nanoparticles by nanoprecipitation to improve the stability and bioavailability of curcumin. LWT. (2022) 171. doi: 10.1016/j.lwt.2022.114149
132. Li H, Yang YG, and Sun T. Nanoparticle-based drug delivery systems for induction of tolerance and treatment of autoimmune diseases. Front Bioeng Biotechnol. (2022) 10:889291. doi: 10.3389/fbioe.2022.889291
133. Hashemi V, Farhadi S, Ghasemi Chaleshtari M, Seashore-Ludlow B, Masjedi A, Hojjat-Farsangi M, et al. Nanomedicine for improvement of dendritic cell-based cancer immunotherapy. Int Immunopharmacol. (2020) 83:106446. doi: 10.1016/j.intimp.2020.106446
134. Zeng X, Cai D, Zeng Q, Chen Z, Zhong G, Zhuo J, et al. Selective reduction in the expression of UGTs and SULTs, a novel mechanism by which piperine enhances the bioavailability of curcumin in rat. Biopharmaceutics Drug disposition. (2017) 38:3–19. doi: 10.1002/bdd.2049
135. Song Y, Zhao F, Zhang L, Du Y, Wang T, and Fu F. Ginsenoside Rg1 exerts synergistic anti-inflammatory effects with low doses of glucocorticoids in vitro. Fitoterapia. (2013) 91:173–9. doi: 10.1016/j.fitote.2013.09.001
136. Focaccetti C, Palumbo C, Benvenuto M, Carrano R, Melaiu O, Nardozi D, et al. The combination of bioavailable concentrations of curcumin and resveratrol shapes immune responses while retaining the ability to reduce cancer cell survival. Int J Mol Sci. (2023) 25. doi: 10.3390/ijms25010232
137. Liu H, Wang L, Li F, Jiang Y, Guan H, Wang D, et al. The synergistic protection of EGCG and quercetin against streptozotocin (STZ)-induced NIT-1 pancreatic beta cell damage via upregulation of BCL-2 expression by miR-16-5p. J Nutr Biochem. (2021) 96:108748. doi: 10.1016/j.jnutbio.2021.108748
138. Choudhury D, Ganguli A, Dastidar DG, Acharya BR, Das A, and Chakrabarti G. Apigenin shows synergistic anticancer activity with curcumin by binding at different sites of tubulin. Biochimie. (2013) 95:1297–309. doi: 10.1016/j.biochi.2013.02.010
139. Kikete S, Luo L, Jia B, Wang L, Ondieki G, and Bian Y. Plant-derived polysaccharides activate dendritic cell-based anti-cancer immunity. Cytotechnology. (2018) 70:1097–110. doi: 10.1007/s10616-018-0202-z
140. Bol KF, Schreibelt G, Rabold K, Wculek SK, Schwarze JK, Dzionek A, et al. The clinical application of cancer immunotherapy based on naturally circulating dendritic cells. J Immunother Cancer. (2019) 7:109. doi: 10.1186/s40425-019-0580-6
141. Kodama N, Okada H, Hachisu M, Ando M, Ito N, Nagata K, et al. A rose flavor compound activating the NRF2 pathway in dendritic cells ameliorates contact hypersensitivity in mice. Front Nutr. (2023) 10:1081263. doi: 10.3389/fnut.2023.1081263
142. Carrillo-Martinez EJ, Flores-Hernandez FY, Salazar-Montes AM, Nario-Chaidez HF, and Hernandez-Ortega LD. Quercetin, a flavonoid with great pharmacological capacity. Molecules. (2024) 29. doi: 10.3390/molecules29051000
143. Petracca M, Quarantelli M, Moccia M, Vacca G, Satelliti B, D'Ambrosio G, et al. ProspeCtive study to evaluate efficacy, safety and tOlerability of dietary supplemeNT of Curcumin (BCM95) in subjects with Active relapsing MultIple Sclerosis treated with subcutaNeous Interferon beta 1a 44 mcg TIW (CONTAIN): A randomized, controlled trial. Mult Scler Relat Disord. (2021) 56:103274. doi: 10.1016/j.msard.2021.103274
144. Allegri P, Cimino L, Davis JL, and Tugal-Tutkun I. Norflo Oro study g. Assessment of the Anti-inflammatory Effects of NORFLO(R) ORO in Acute Relapses of HLA-B27-associated Autoimmune Uveitis: A Multicenter, Randomized, Placebo-controlled, Double-blind Clinical Study. Ocul Immunol Inflamm. (2023) 31:526–35. doi: 10.1080/09273948.2022.2039210
145. Klumbies K, Rust R, Dorr J, Konietschke F, Paul F, Bellmann-Strobl J, et al. Retinal thickness analysis in progressive multiple sclerosis patients treated with epigallocatechin gallate: optical coherence tomography results from the SUPREMES study. Front Neurol. (2021) 12:615790. doi: 10.3389/fneur.2021.615790
146. Rust R, Chien C, Scheel M, Brandt AU, Dorr J, Wuerfel J, et al. Epigallocatechin gallate in progressive MS: A randomized, placebo-controlled trial. Neurol Neuroimmunol Neuroinflamm. (2021) 8. doi: 10.1212/NXI.0000000000000964
147. Lovera J, Ramos A, Devier D, Garrison V, Kovner B, Reza T, et al. Polyphenon E, non-futile at neuroprotection in multiple sclerosis but unpredictably hepatotoxic: Phase I single group and phase II randomized placebo-controlled studies. J Neurol Sci. (2015) 358:46–52. doi: 10.1016/j.jns.2015.08.006
Keywords: phytochemicals, dendritic cells, bioactive compounds, inflammation, Autoimmune disease, therapy, tolerogenic
Citation: Fekete T and Pázmándi K (2025) Phytochemicals as modulators of dendritic cell functions: implications for tolerogenic cell-based therapy. Front. Immunol. 16:1653803. doi: 10.3389/fimmu.2025.1653803
Received: 25 June 2025; Accepted: 29 October 2025;
Published: 11 November 2025.
Edited by:
Oscar Herrera-Calderon, Universidad Nacional Mayor de San Marcos, PeruReviewed by:
Cristopher Anel Boya, Instituto de Investigaciones Científicas y Servicios de Alta Tecnología, PanamaSalyoc Tapia-Rojas, Universisdad Cientifica del Sur, Peru
Copyright © 2025 Fekete and Pázmándi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tünde Fekete, ZmVrZXRlLnR1bmRlQG1lZC51bmlkZWIuaHU=