 Mayte Rueda-Munguía1,2
Mayte Rueda-Munguía1,2 Luis Alberto Luévano-Martínez1,2
Luis Alberto Luévano-Martínez1,2 Gerardo García-Rivas1,2
Gerardo García-Rivas1,2 Elena Cristina Castillo1,2*
Elena Cristina Castillo1,2* Omar Lozano1,2*
Omar Lozano1,2*- 1Escuela de Medicina y Ciencias de la Salud Tec Salud, Tecnologico de Monterrey, Monterrey, Mexico
- 2Institute for Obesity Research, Tecnologico de Monterrey, Monterrey, Mexico
The intricate interplay between macrophage biology and lipid metabolism has emerged as a critical determinant of metabolic homeostasis, disease progression and pathogenesis. This comprehensive review explores the molecular mechanisms through which fatty acids activate macrophage function, emphasizing their selective engagement of pattern recognition receptors such as Toll-like receptors (TLRs), CD36, and GPR120. Notably, saturated fatty acids (SFAs) like lauric acid (C12:0) and palmitic acid (C16:0) activate TLR2 and TLR4 signaling pathways. Palmitic acid triggers mitochondrial dysfunction and lysosomal destabilization, leading to NLRP3 inflammasome activation and chronic low-grade inflammation. In contrast, ω-3 polyunsaturated fatty acids (PUFAs), such as docosahexaenoic acid, help resolve inflammation through GPR120-mediated signaling and the production of specialized pro-resolving mediators (SPMs) like resolvins, protectins, and maresins. This review establishes a paradigm for understanding the complex relationship between dietary lipids, innate immunity, and metabolic health, with broad implications for immunometabolic interventions.
Introduction
The immune system, an intricate network of cells including macrophages, neutrophils, dendritic cells, and natural killer (NK) cells (1, 2) relies significantly on the multifaceted functions of their cells to maintain systemic homeostasis (3). Of particular interest are macrophages, which constitute approximately 10% of immune cells (4), exhibit remarkable heterogeneity and plasticity (5), enabling them to play pivotal roles in both promoting and resolving inflammation.
Macrophages can be broadly classified into two categories: monocyte-derived macrophages, which originate from the bone marrow, circulate in the bloodstream, and subsequently extravasate into tissues, where they differentiate into macrophages; and tissue-resident macrophages, such as alveolar macrophages in the lungs and Kupffer cells in the liver, which emerge during fetal development from precursors in the yolk sac and fetal liver precursors (6). Unlike monocyte-derived macrophages, tissue-resident macrophages sustain their population through local proliferation, allowing them to adapt and respond specifically to their microenvironment (7).
Depending on external stimuli, macrophages polarize into distinct phenotypes, each performing specialized functions within tissues (8). The two primary phenotypes are pro-inflammatory macrophages (M1), which are activated in response to stimuli such as lipopolysaccharides (LPS) and cytokines like INF-γ (9), and the anti-inflammatory macrophages (M2), which are activated in the presence of interleukin (IL)-4 and IL-13, that play key roles in inflammation resolution, tissue repair, and homeostasis maintenance (10).
Beyond their traditional immune functions, macrophages are increasingly recognized as key regulators in metabolic diseases. In this context, fatty acids, long known for their roles as energy substrates and structural components, exert significant influence on macrophage polarization and function by activating or suppressing key signaling pathways (11). In metabolic disorders, macrophages often exhibit a mixed phenotype, distinct from classical M1 and M2 polarization, referred to as metabolically activated (MA) macrophages (12).
In addition to the classical M1 and M2 polarization spectrum, single-cell transcriptomic studies have identified a metabolically specialized subset of macrophages termed lipid-associated macrophages (LAMs). These cells mostly arise in adipose tissue during obesity and are characterized by the expression of genes associated with lipid metabolism, including TREM2, CD9 and lipoprotein lipase (LPL) (13). LAMs localize around hypertrophic or necrotic adipocytes, where they phagocytose lipids and cellular debris, hence fulfilling a protective and homeostatic function during initial metabolic stress. However, under conditions of chronic obesity, their sustained activation leads to the production of pro-inflammatory cytokines and maladaptive tissue remodeling, ultimately exacerbating metabolic dysfunction (14). Collectively, LAMs connect metabolism and immunology by merging lipid handling with inflammatory signaling, positioning them as a central component of the immunometabolic framework of adipose tissue.
In the context of a diet rich in animal fats and processed foods, oversupply of saturated fatty acids (SFAs) within tissues leads to their accumulation, particularly in adipose tissue, where macrophages accumulate excess fatty acids (15). This accumulation of SFAs influences macrophages’ function beyond their metabolic contributions as energy resources, leading to a MA phenotype (Figure 1). MA macrophages activate M1-like inflammatory pathways, promoting low-grade chronic inflammation and contributing to the pathogenesis of metabolic disorders (16, 17). The disruption in the function of macrophages occurs not only in the adipose tissue but also in tissue-resident macrophages, such as Kupffer cells in the liver and microglia in the brain (6). This systemic impact may drive chronic inflammation and increase susceptibility to a range of metabolic disorders, including obesity, diabetes type II, and cardiovascular diseases (18, 19).
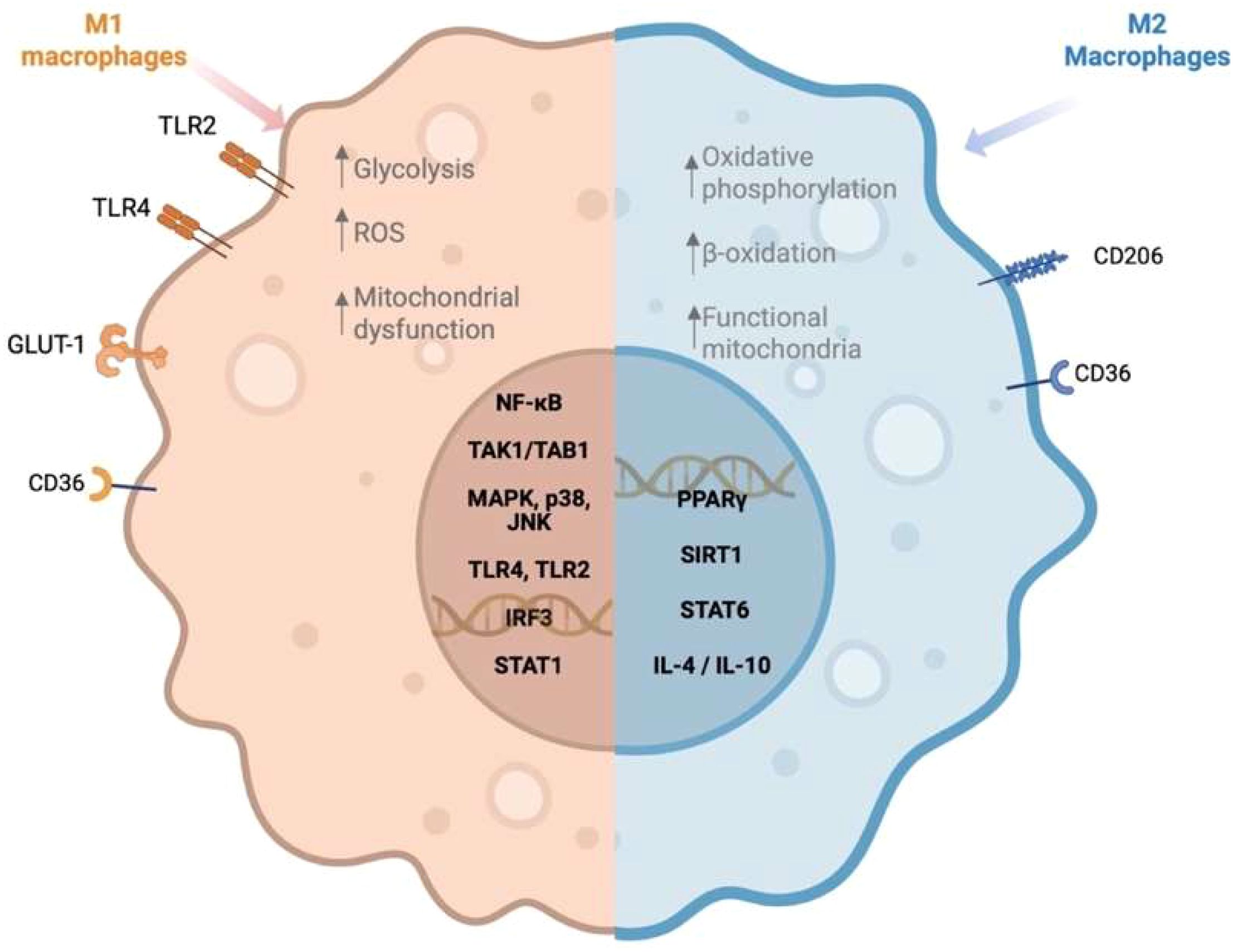
Figure 1. Macrophage polarization in response to saturated and unsaturated fatty acids. Created in https://BioRender.com.
Dietary SFAs, such as palmitic acid and lauric acid (C:12), activate Toll-like receptor 4 (TLR4) signaling pathways, leading to the activation of nuclear factor-kappa B (NF-κB) and the subsequent release of pro-inflammatory cytokines, including tumor necrosis factor-alpha (TNF-α), IL-1β, and IL-6, a pleiotropic cytokine associated with insulin resistance (20, 21). In addition, palmitic acid induces mitochondrial dysfunction and lysosomal destabilization, culminating in the activation of the NLRP3 inflammasome, a multiprotein complex strongly associated with obesity and type II diabetes (22). Caspase-1, a key component of the NLRP3 inflammasome, facilitates the maturation and activation of IL-1β, thereby amplifying the inflammasome response (23). However, evidence suggests that other inflammasomes, such as NLRC4, though less well characterized in the context of obesity, also contribute to IL-1β production and the exacerbation of inflammatory states (23).
Conversely, M2 macrophage polarization, driven by cytokines like IL-4, IL-13, IL-10, and transforming growth factor beta (TGF-β), serves a compensatory role in controlling inflammation, promoting tissue repair, and maintaining metabolic homeostasis (9). Certain unsaturated fatty acids (UFAs), such as ω-3 fatty acids like eicosapentaenoic acid and docosahexaenoic acid, have been shown to support M2 polarization and exert anti-inflammatory responses, counteracting the pro-inflammatory effects of SFAs (24). Notably, IL-10 released from M2 macrophages suppresses mechanistic target of rapamycin complex 1 (mTORC1) signaling, a pathway known to regulate glycolysis and mitochondrial respiration in immune cells, thereby reducing inflammation and oxidative stress (25). Additionally, TGF-β plays a crucial role in regulating adipogenesis, T cell differentiation, and mitochondrial metabolism in adipose tissue, further supporting metabolic homeostasis (26). These cytokines collectively counteract immune and metabolic dysfunction in obesity and insulin resistance, partly by mitigating the mitochondrial dysfunction and lysosomal destabilization induced by SFAs like palmitic acid. This shift toward M2 polarization and the anti-inflammatory phenotype supports metabolic health, particularly in counteracting chronic inflammation induced by diet-related factors.
In the context of metabolic disorders, the widespread dietary shift towards ultra-processed foods (UPFs), characterized by high SFA and deficiency in essential nutrients, exacerbates metabolic dysregulation (27). UPFs activate inflammatory pathways, including TLR4-NF-κB signaling, which directly impacts immune cell function, particularly macrophages (28). This signaling cascade skews macrophage polarization toward a MA phenotype, contributing to chronic low-grade inflammation and insulin resistance (29).
The excess SFAs present in UPFs also promote mitochondrial dysfunction and lysosomal destabilization in macrophages, further amplifying the inflammatory response by activating NLRP3 inflammasomes (30). These alterations in macrophage function, driven by the dietary intake of UPFs, are closely linked to the increased risk of obesity, type II diabetes, and cardiovascular diseases. Furthermore, epidemiological studies show that each 20-gram increase in UPF consumption is associated with a significant increase in serum cholesterol (TC) (β = 1.214; 95% CI:.159-2.269) and a reduction in high-density lipoprotein (HDL) (β = -0.371; 95% CI: -0.675 to -0.067), both of which contribute to dyslipidemia and promote a pro-inflammatory environment, exacerbating metabolic imbalance (27). These findings underscore the detrimental impact of UPFs on macrophage function and their role in driving immune dysfunction and metabolic disorders.
This review aims to elucidate and analyze the multifaceted interactions between macrophages and fatty acids and their contributions to the pathogenesis of metabolic and cardiovascular diseases. By bridging cellular mechanisms with dietary influences, we provide a comprehensive framework for understanding the immuno-metabolic dysregulation underpinning these conditions while identifying potential therapeutic targets.
For the compilation of the presented information in this review, keyword searches employing Boolean operators, such as “macrophages AND metabolic diseases,” “macrophages AND diabetes OR obesity,” “macrophages AND fatty acids,” and “fatty acids AND inflammation,” were used. A literature search was conducted across databases, including PubMed and ScienceDirect.
Impact of SFA on the function of macrophages
A defining component of the modern Western diet is SFAs, which have been implicated in the rising global prevalence of metabolic diseases (31). SFAs are a class of lipids characterized by their fully saturated molecular structure, lacking double bonds between carbon atoms. These fatty acids are abundant in animal-derived products such as meat and dairy, as well as in certain tropical oils, including coconut and palm oil (32).
Fatty acids serve as crucial substrates for triglyceride (TG) storage and membrane formation, with their metabolism intricately linked to cellular energy balance. While TG serves as a physiological adaptation to excess energy intake, chronic overconsumption of SFAs disrupts lipid homeostasis, leading to ectopic TG accumulation in non-adipose tissues, a phenomenon known as lipotoxicity (30). Lipotoxic stress induces cellular dysfunction, particularly in hepatocytes, cardiomyocytes, and macrophages, where excessive SFAs are metabolized into bioactive lipids such as ceramides and diacylglycerols (DAGs) (33). These lipid intermediates disrupt insulin signaling and predispose macrophages to an inflammatory phenotype (11).
Exposure to SFAs drives macrophage polarization toward a pro-inflammatory M1-like phenotype. Specifically, palmitic acid (C16:0) triggers TLR signaling, activating the NF-κB and c-Jun N-terminal kinase (JNK) pathways (19). This signaling cascade primes macrophages to secrete cytokines such as TNF-α and IL-1β, contributing to the chronic low-grade inflammation characteristic of metabolic diseases (19). In addition to palmitic acid, other SFAs, including myristic acid (C14:0), lauric acid, and stearic acid (C18:0), have also been shown to induce mitochondrial dysfunction and impair oxidative phosphorylation, leading to a disruption of normal cellular energy metabolism (16). These alterations lead to increased production of reactive oxygen species (ROS), contributing to metabolic stress and promoting insulin resistance (34).
Additionally, both saturated and unsaturated fatty acids can be taken up through the scavenger receptor CD36, which also mediates the uptake of oxidized-low density lipoproteins (oxLDL) (35, 36). In the context of SFAs, CD36-mediated lipid accumulation impairs cholesterol efflux and promotes foam cell formation, thereby amplifying pro-inflammatory signaling (37, 38). Collectively, these SFA-induced changes in macrophage function accelerate metabolic dysfunction, reinforcing the link between dietary composition and the pathogenesis of metabolic disorders, including type II diabetes and cardiovascular diseases (34).
Findings summarized in Table 1 highlight the interactions between various fatty acids and key metabolic pathways, demonstrating their impact on macrophage inflammation and metabolism. Palmitic acid activates mitogen-activated protein kinase (MAPK) and TLR4 pathways, leading to increased secretion of monocyte chemoattractant protein-1 (MCP-1) and promoting inflammatory responses via extracellular signal-regulated kinases 1/2 (ERK1/2) and TLR2/4 activation (19, 39). This pro-inflammatory effect is further amplified when palmitic acid is combined with palmitoleic acid (C16:1) (40). Notably, palmitoleic acid, frequently exerts opposing, anti-inflammatory actions. Experimental evidence shows that palmitoleic acid attenuates high-fat diet-induced proinflammatory macrophage polarization via AMPK activation (40) and inhibits palmitic acid-induced macrophage activation, thereby mitigating skeletal muscle insulin resistance in co-culture models of J774 macrophages and C2CL12 myotubes (41). Nonetheless, its protective effects are context-dependent: they are more pronounced at physiological concentrations (≤50 μM) and in macrophage or adipose tissue models, whereas in microglial cells, palmitoleic acid provides limited protection against palmitate-induced inflammation (42).
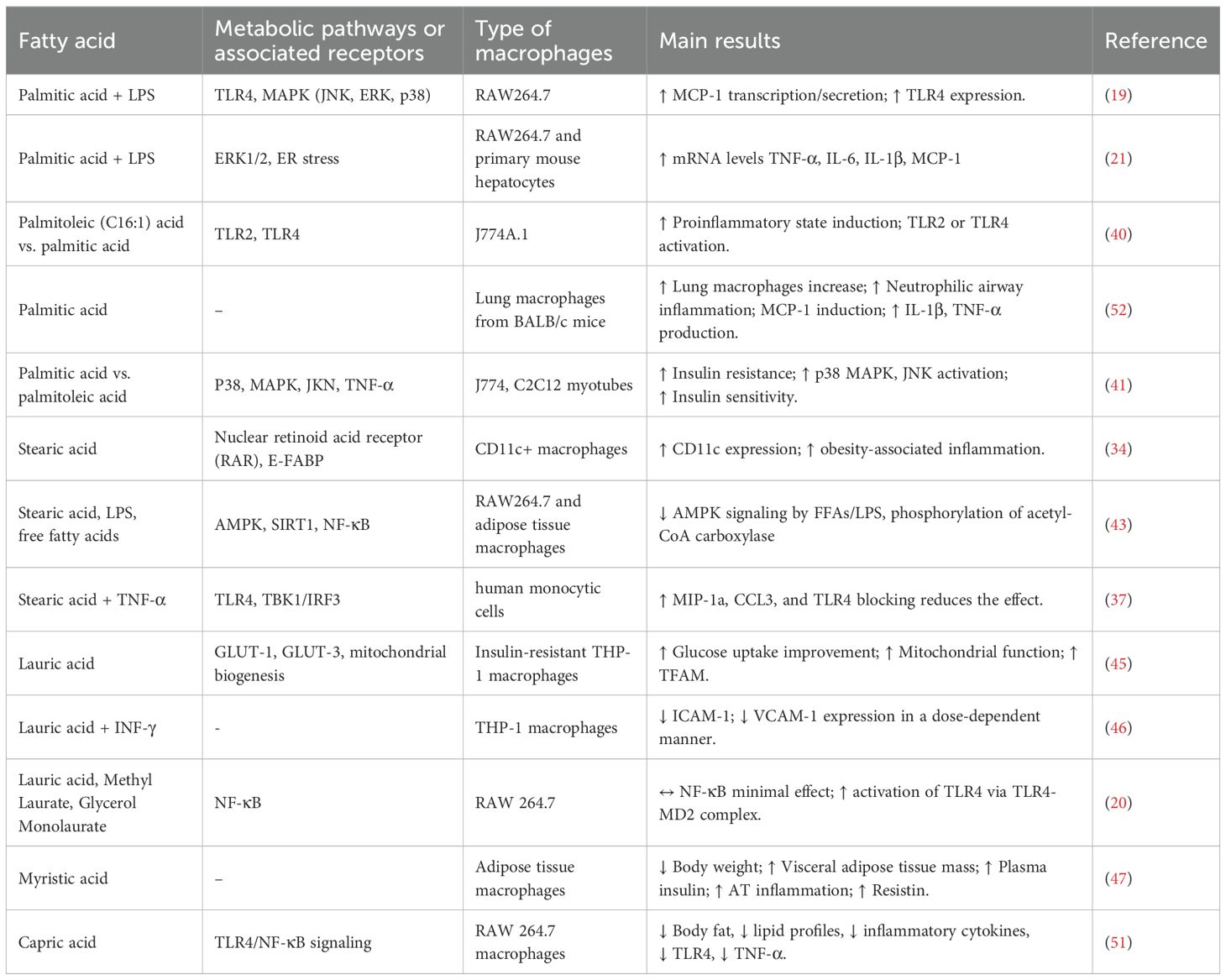
Table 1. Impact of SFAs on macrophages’ metabolic pathways and their effects.
Stearic acid influences CD11c expression through nuclear retinoid acid receptors and epidermal fatty acid-binding protein (E-FABP), both of which are critical in driving macrophage differentiation towards a pro-inflammatory CD11+ phenotype (34). Furthermore, stearic acid influences key metabolic pathways by inhibiting the AMP-activated protein kinase (AMPK) phosphorylation and sirtuin 1 (SIRT1) dependent signaling, two central regulators of cellular energy homeostasis (43). The activation of MAPK and SIRT1 has been implicated in metabolic programming, promoting mitochondrial biogenesis, lipid oxidation, and anti-inflammatory responses, thereby playing a protective role against obesity-induced insulin resistance and chronic inflammation (44).
There are instances where SFAs exhibit context-dependent or even contradictory effects, as illustrated by lauric acid. In insulin-resistant THP-1 macrophages, lauric acid enhanced mitochondrial biogenesis and glucose uptake at concentrations of 5-50 μM (24 h), thereby increasing the expression of PGC-1α, TFAM, GLUT1, and GLUT-3, thereby attenuating insulin resistance and promoting metabolic health (45). In line with these findings, lauric acid has been reported to suppress the expression of adhesion molecules ICAM-1 and VCAM-1 in human macrophages, further supporting its anti-inflammatory potential (46). In contrast, lauric acid activated pro-inflammatory signaling in RAW264.7 macrophages by inducing TLR4 dimerization and its recruitment into lipid rafts with MD-2, a proximal step that initiates NF-κB activation in a manner similar to LPS (20). Available studies do not report explicit cell-viability thresholds for lauric acid. However, in insulin-resistant THP-1 macrophages, micromolar exposures (5-50 μM, 24h) were well tolerated and improved mitochondrial function (45), whereas in RAW264.7 cells, higher doses (~150 μM sodium laurate) rapidly triggered TLR4/MD-2-mediated NF-κB activation (20). These findings suggest that the inflammatory outcome depends on the context due to variations in dosage conditions.
By contrast, myristic acid is associated with increased body weight, adipose tissue inflammation, and insulin resistance, contributing to the metabolic disease progression. This indicates that dietary supplementation with myristic acid exacerbates adipose tissue inflammation and systemic insulin resistance in mice subjected to a high-fat diet (47). The study observed increased macrophage infiltration and elevated expression of pro-inflammatory markers in adipose tissue, alongside higher plasma insulin levels and impaired glucose tolerance (48).
On the other hand, capric acid (10:0) has demonstrated potential metabolic benefits, reducing body fat, improving lipid profiles, and suppressing inflammatory responses through TLR4 receptor and NF-κB signaling (49–51).
These findings underscore the complex and nuanced roles of fatty acids in regulating macrophage function and inflammation. While certain SFAs, such as lauric acid and capric acid, may confer metabolic or anti-inflammatory benefits under defined conditions, others like myristic promote inflammation and metabolic dysfunction. Understanding the specific effects of different fatty acids on macrophage function could inform targeted therapeutic strategies for managing metabolic disorders and inflammation-related conditions.
Activation of macrophages by SFAs, particularly palmitic acid, induces a pro-inflammatory phenotype characterized by elevated production of cytokines such as TNF-α, IL-6, and IL-1β, alongside activation of the NF-κB pathway (17). This state is associated with endoplasmic reticulum stress and generation of reactive oxygen species (ROS), contributing to chronic inflammation and insulin resistance in metabolic diseases (28). SFAs activate receptors such as TLR4, TLR2, and the scavenger receptor CD36, which together trigger downstream signaling pathways including NF-κB, JNK, and p38 MAPK, as well as the NLRP3 inflammasome (22). Additionally, SFA disrupt lipid metabolism by promoting lipid accumulation and enhancing fatty acid oxidation, resulting in low-grade, sustained inflammation when exposure is prolonged. In contrast, macrophage activation by lipopolysaccharide (LPS) occurs exclusively through TLR4, which engages the MyD88 and TRIF signaling pathways, leading to the activation of NF-κB and production of cytokines through a typically acute and transient inflammatory response (23, 28). These differences highlight not only the specific signaling pathways involved but also how the persistence and resolution of the stimulus shape the temporal dynamics of the immune response, influencing whether inflammation becomes chronic (17, 53).
Impact of unsaturated fatty acids on the function of macrophages
UFAs, distinguished by the presence of one or more double bonds in their hydrocarbon chain, exist either in cis or trans configurations based on spatial arrangements around the double bond. The presence of cis double bonds introduces a structural bend, conferring flexibility to the hydrocarbon chain. This property prevents tight packing within membrane phospholipids, thereby increasing the fluidity of the membrane (54). Dietary inclusion of UFAs has been associated with numerous health benefits, including enhanced cardiovascular health, anti-inflammatory effects, and improved cognitive function (2, 55). Among UFAs, polyunsaturated (PUFAs) and monounsaturated (MUFAs) are particularly recognized for their roles in supporting metabolic and immune functions, with each type offering distinct but complementary health benefits; enhancing tissue repair, and contributing to cardiovascular health by reducing triglycerides and improving endothelial function (56). PUFAs, which contain two or more double bonds, are primarily classified into ω-3 and ω-6 families, whereas MUFAs possess a single double bond, conferring them structural and functional properties distinct from PUFAs. Both play a critical role by activating or suppressing distinct signaling pathways that regulate polarization and inflammatory responses function.
Like SFAs, PUFAs and MUFAs exert varying effects on inflammation and metabolism, with some demonstrating anti-inflammatory benefits while others promote inflammatory responses. These effects depend on the type of fatty acid and its dietary context. For example, ω-3 PUFAs such as eicosapentaenoic acid and docosahexaenoic acid are generally anti-inflammatory (57, 58), whereas ω-6 PUFAs, like arachidonic acid (C20:4) (59), can promote inflammation when consumed excessively. Similarly, MUFAs such as oleic acid (C:18:1) (60) have anti-inflammatory properties, but oxidized MUFAs may contribute to inflammatory responses. Overall, while UFAs are broadly associated with beneficial effects on inflammation and metabolic health, certain PUFAs and MUFAs may exert pro-inflammatory effects under specific conditions.
Polyunsaturated fatty acids
PUFAs play a critical role in regulating metabolic pathways and are integral components of phospholipid bilayers in cell membranes, where their unsaturated hydrocarbon chains contribute to fluidity and flexibility (54). Functionally, PUFAs serve as precursors for bioactive lipid mediators known as oxylipins, which are generated via enzymatic pathways (e.g., cyclooxygenase [COX], lipoxygenase [LOX], and cytochrome P450 systems) as well as non-enzymatic processes involving free radicals (61). Oxylipins regulate key biological processes, including inflammation, tissue homeostasis, and intercellular communication, by influencing intracellular signal pathways. Importantly, their effects are context dependent. Pro-inflammatory oxylipins such as prostaglandins and leukotrienes, derived from arachidonic acid, amplify inflammation, while resolvins, maresins, and protectins form eicosapentaenoic acid and promote resolution and tissue repair (62).
One key pathway in macrophages is that mediated by the peroxisome proliferator-activated receptor gamma (PPAR-γ), through which PUFAs promote an M2 phenotype, reprogramming immune cells into a reparative state and fostering an anti-inflammatory environment (59). Additionally, PUFAs enhance the cholesterol efflux pathways in macrophages, reducing foam cell formation and exerting anti-atherogenic effects (63, 64).
In macrophages, ω-3 PUFAs facilitate the transition from inflammation to resolution, supporting tissue repair by reinforcing the M2 macrophage state. This effect is mediated through the production of specialized pro-resolving mediators (SPMs), which include resolvins, maresins, and protectins (65). D-series resolvins, derived from eicosapentaenoic acid and docosahexaenoic acid, such as resolvin D1 (RvD1), are potent anti-inflammatory mediators, suppressing the NF-κB signaling pathway, enhancing macrophage phagocytosis, and promoting the clearance of inflammatory cells and debris (66). Maresins and protectins, both synthesized from docosahexaenoic acid, further contribute to inflammation resolution by promoting tissue repair and macrophage-mediated efferocytosis (the clearance of apoptotic cells) (67, 68). Mechanistically RvD1 signals through G-protein-coupled receptors (GPCRs) such as ALX/FPR2 and GPR32, both expressed on macrophages, initiating a coordinated anti-inflammatory signaling cascade that reduces pro-inflammatory cytokine secretion and promote efferocytosis (67, 68).
In addition to the D-series, E-series resolvins derived from eicosapentaenoic acid, most notably resolvin E1 (RvE1), exert complementary effects in resolution. RvE1 was originally characterized in murine peritonitis models and human peripheral blood neutrophils and monocytes, where it inhibits leukotriene-driven inflammation by antagonizing BLT1 while simultaneously engaging ChemR23 (ERV1) to promote efferocytosis and limit neutrophil infiltration (69). In addition to resolvins, several families of specialized pro-resolving mediators (SPMs) demonstrate receptor specificity. Maresin-1 was shown in human macrophages and mouse peritonitis models to engage LGR6 and orchestrate regeneration processes, while protectin PD1 has been associated with GPR37, imparting neuroprotective and pro-resolving properties (70, 71). These pathways collectively illustrate the integration of several PUFA-derived mediators and their corresponding receptors to promote homeostasis, tissue repair, and resolution.
In addition, α-linolenic acid (C18:3), an essential ω-3 PUFA, exhibits antioxidative properties that regulate cellular redox balance (72). By upregulating key enzymes such as superoxide dismutase (SOD) and catalase, α-linolenic acid helps mitigate ROS production, thereby limiting oxidative stress and reducing ROS-induced cellular damage in macrophages (61). This protective mechanism may play a crucial role in preventing chronic inflammation. Moreover, α-linolenic acid influences immune responses by inhibiting the production of pro-inflammatory cytokines (48) and enhancing oxylipin-mediated inflammation resolution (61).
On the other hand, ω-6 PUFAs exhibit distinct pro-inflammatory and anti-inflammatory properties, with their effects on macrophages playing a crucial role in inflammatory processes. For instance, linolenic acid, an essential ω-6 PUFA, enhances macrophage activation and remodels lipid raft composition, thereby promoting key signaling pathways (73). In contrast, conjugated linoleic acid, a group of geometric and positional isomers of linoleic acid (C18:2), impacts adipose tissue inflammation by altering adipocyte differentiation. Conjugated linoleic acid supplementation has been shown to reduce inflammation in obese rats by decreasing pro-inflammatory M1 macrophages and increasing anti-inflammatory M2 macrophages (74). Specifically, the 10,12-conjugated linoleic acid isomer promotes weight loss and increases anti-inflammatory macrophages in adipose tissue, underscoring its therapeutic potential in mitigating obesity-induced inflammation (75).
Similarly, γ-linolenic acid (C18:3), another ω-6 PUFA, exerts context-dependent effects on macrophage responses by exhibiting both pro-inflammatory and anti-inflammatory effects. Evidence shows that γ-linolenic acid can suppress NF-κB and AP-1 activation, thereby inhibiting the expression of inflammatory mediators (76). However, other studies indicate that γ-linolenic acid does not consistently affect cytokines such as IL-6, suggesting that its role may vary depending on the experimental context (77). This dual function highlights the complex interplay of linoleic acid, conjugated linoleic acid, and γ-linolenic acid within inflammatory processes, revealing a dynamic regulation of macrophage activity. γ-linolenic acid illustrates the context-dependent effects of PUFA: in RAW264.7 macrophages, it increases IL-6 secretion and activates NF-κB/AP-1 signaling, hence promoting chronic inflammation (76). Conversely, γ-Linoleic acid can stimulate PPAR-γ in human immune cells, increasing IL-10 production and thereby suppressing pro-inflammatory mediators (15). This highlights that the impact of certain fatty acids on macrophage activity is profoundly influenced by cellular environment, dose, and metabolic state.
The interaction between PUFAs and macrophages is important because it influences inflammatory responses, which has implications for the development of therapeutic strategies targeting inflammatory diseases. Understanding the nuanced effects of different PUFAs on macrophage function may offer important lessons for future PUFA-based therapies aimed at mitigating metabolic and immune disorders. Table 2 provides an overview of the various types of PUFAs, their dietary sources, and their physiological effects.
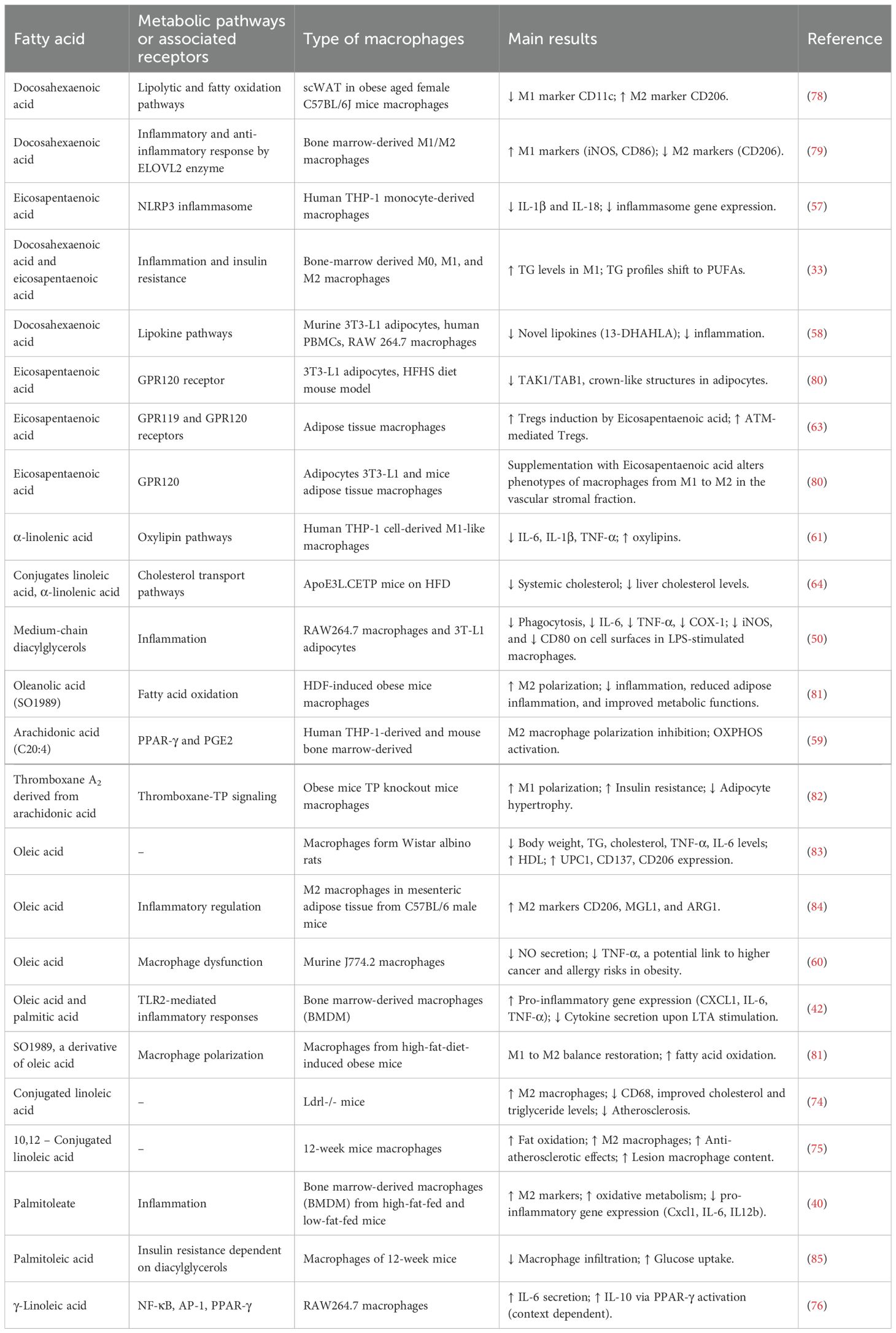
Table 2. Metabolic pathways of PUFAs in macrophages and their effects.
Impact of monounsaturated fatty acids on macrophages’ function
MUFAs, characterized by a single double bond, typically in the cis configuration, are abundant in plant-based oils such as olive, canola, and avocado oil. The Mediterranean diet, rich in MUFAs, has been associated with health benefits, including cardiovascular protection, anti-inflammatory effects, and improved lipid profiles. In this diet pattern, approximately 60% of total fat intake is derived from MUFAs, which have been shown to lower blood pressure, enhance glucose metabolism, and reshape gut microbiome, thereby reducing the risk of coronary artery disease (56).
Among MUFAs, oleic acid —a major component of olive oil—exhibits notable anti-inflammatory effects and improves insulin sensitivity by suppressing TLR4 signaling in macrophages (60, 86). This suppression downregulates downstream inflammatory pathways, particularly those mediated by NF-κB and MAPK kinases, leading to a reduction in pro-inflammatory cytokines such as IL-6 (42). Consequently, this suppression contributes to an overall anti-inflammatory response. Furthermore, oleic acid promotes macrophage polarization toward the M2 phenotype by activating PPAR-γ, favoring a reparative and anti-inflammatory state (83, 84). This activation not only enhances the expression of genes involved in the differentiation of insulin-sensitive adipocytes but also supports inflammation resolution by reducing macrophage infiltration into adipose tissue, a key driver of obesity-associated inflammation.
In addition to its effects on macrophage polarization, oleic acid (C18:1 cis-9) and other MUFAs also mitigate oxidative stress by enhancing the activity of antioxidant enzymes such as superoxide dismutase (SOD), glutathione peroxidase (GPx), and catalase (72). This mechanism protects cells from oxidative damage, further facilitating inflammation resolution. Importantly, PPAR-γ activation by MUFAs not only reduced NF-κB signaling but also synergistically enhanced antioxidant enzyme activity, creating a dual mechanism that simultaneously combats oxidative stress while promoting anti-inflammatory macrophage function (36). This combined action helps reduce macrophage accumulation in adipose tissue, thus alleviating inflammation associated with obesity and metabolic disorders (87).
However, not all MUFAs exhibit the same effects. Erucic acid (C22:1), found in yellow mustard, has been reported to act as a natural inhibitor of PPAR-γ transcriptional activity, improving insulin sensitivity and reducing macrophage infiltration into adipose tissue (88). This effect contrasts with the typical benefits of PPAR-γ activation, seen with oleic acid and ω-3 PUFAs, where PPAR-γ activation promotes an anti-inflammatory M2 macrophage phenotype and contributes to inflammation resolution (89). These findings underscore that MUFAs are a heterogeneous group of fatty acids, with effects that depend on the specific fatty acid, its interaction with PPAR-γ, and its metabolic context. The differing impact of MUFAs on macrophage function illustrates the importance of a more comprehensive understanding of how various MUFAs may either upregulate or downregulate specific inflammation pathways.
A key consideration in the dual role of PPAR-γ in macrophage function is that while PPAR-γ activation reduces inflammatory cytokine production in M1 macrophages and potentiates M2 activation, it may also impair full M2c polarization, a macrophage subtype critical for achieving an inflammatory-resolution state (90). This suggests that chronic PPAR-γ activation, as observed in obesity or metabolic syndrome, may hinder complete inflammation resolution. In this context, PPAR-γ antagonism during monocyte-to-macrophage differentiation has been proposed as a strategy to promote M2c-like polarization, helping restore anti-inflammatory conditions in chronic inflammatory settings (89, 90). These findings emphasize the complex-dependent effects of PPAR-γ on macrophage function, which points to the therapeutic benefits of selectively targeting specific PPAR-γ isoforms to either enhance or inhibit macrophage polarization and metabolic pathways depending on context.
While many MUFAs (such as oleic acid) are generally anti-inflammatory and promote M2 polarization, nervonic acid (C24:1) exhibits a more complex role in inflammation. Nervonic acid has been studied for its effects on immune function in RAW 264.7 macrophages, where it has been found to activate NF-κB signaling, leading to the increased production of pro-inflammatory cytokines (e.g., TNF-α and IL-6) as well as enhanced nitric oxide (NO) and ROS generation in concentrations of 12.5 μM or higher (91). These findings suggest that nervonic acid exerts dose-dependent effects on macrophage function, reinforcing that not all MUFAs follow the same pattern regarding macrophage polarization and inflammation. Further research is needed to fully elucidate the mechanisms of nervonic acid in different physiological contexts and explore its potential role in macrophage activity (72).
In animal models, such as in JCR rats (a model of obesity), vaccenic acid (C18:1 trans-11), a precursor to trans fats derived from ruminants, has been shown to suppress pro-inflammatory lymphocyte activity and improve the inflammatory profile of mesenteric lymphocytes (92). These findings point out the importance of MUFAs in regulating macrophage activity and inflammation, particularly in obesity-related metabolic disorders (see Table 3). Additionally, these results highlight the complexity and diversity of MUFA effects on immune function, demonstrating that MUFAs cannot be generalized into one anti-inflammatory category.
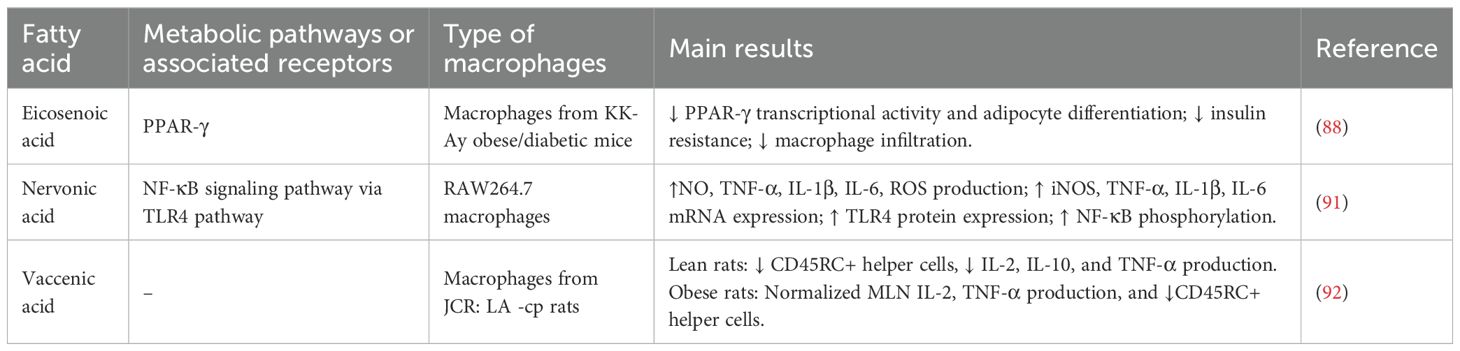
Table 3. Summary of MUFAs effects on metabolic and immunological pathways.
Concluding remarks
The intricate interplay between fatty acid metabolism and immune cell function unveils a complex regulatory network that impacts both physiological and pathological processes. As illustrated in Figure 2, the metabolic activation of macrophages in response to different types of fatty acids reveals divergent functional outcomes. Fatty acid oxidation plays a critical role in promoting alternative and regulatory macrophage phenotypes, particularly the M2 phenotype, which is involved in tissue remodeling, wound healing, and anti-inflammatory cytokine production. Key metabolic regulators, such as PPAR-γ and AMPK, enhance fatty acid uptake and oxidation in M2 macrophages, supporting their anti-inflammatory and reparative functions (43, 59).
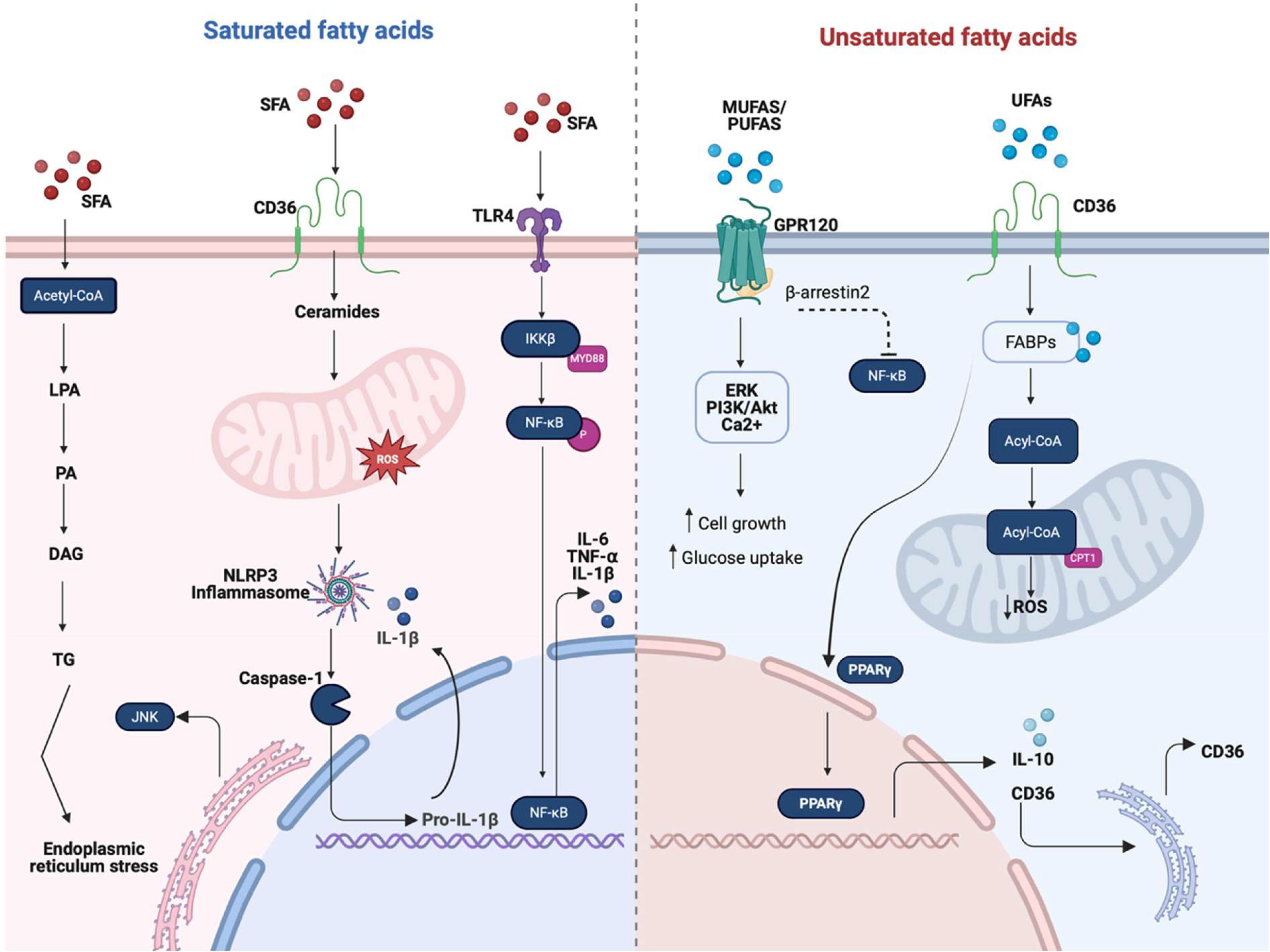
Figure 2. Overview of the main macrophage activation pathways induced by saturated (SFAs) and unsaturated fatty acids (UFAs, including MUFAs and PUFAs). Fatty acids regulate macrophage function by acting as both metabolic substrates and signaling molecules that shape inflammatory responses. SFAs are metabolized into triglycerides (TGs), contributing to endoplasmic reticulum (ER) stress and the activation of pro-inflammatory pathways involving CD36, ceramides, and reactive oxygen species (ROS), which trigger the NLRP3 inflammasome. This cascade promotes inflammation by disrupting membrane fluidity and enhancing TLR4-mediated NF-κB signaling, leading to the production of proinflammatory cytokines such as IL-6, TNF-α, and IL-1β. In contrast, UFAs exert anti-inflammatory and pro-resolving effects. They activate GPR120, which engages multiple downstream pathways, including ERK, PI3K/AKT, and Ca2+ signaling, that collectively enhance glucose uptake and cell growth. UFAs also signal through CD36 and fatty acid-binding proteins (FABPs), which transport fatty acids to mitochondria for conversion to acetyl-CoA. Acetyl-CoA enters β-oxidation, facilitated by carnitine palmitoyl transferase 1 (CPT1), thereby enhancing mitochondrial respiration and reducing ROS levels. This metabolic shift supports the activation of PPAR-γ and the production of IL-10, reinforcing an M2 phenotype through feedback to CD36. Together, these pathways highlight the dual immunometabolic roles of fatty acids in macrophage polarization and suggest potential targets for therapeutic intervention in metabolic inflammatory diseases. Created in https://BioRender.com.
In contrast, de novo lipogenesis and fatty acid synthesis are closely associated with the pro-inflammatory M1 macrophage phenotype, which is related to rapid clonal expansion and cytokine production. This metabolic pathway supplies essential lipids for membrane biogenesis and biosynthesis of signaling molecules, such as inflammatory eicosanoids derived from arachidonic acid. The metabolic shift toward glycolysis in M1 macrophages is further regulated by sterol regulatory element-binding protein 1 (SREBP1), which ensures a continuous supply of substrates for fatty acid synthesis to sustain the heightened metabolic demands of inflammation (17).
Our revision highlights the distinct roles of SFAs, MUFAs, and PUFAs in shaping macrophage behavior and function. SFAs activate pattern recognition receptors, particularly TLR4, leading to NF-κB activation and subsequent production of inflammatory cytokines like TNF-α and IL-6. Chronic exposure to SFAs has been implicated in the development of metabolic inflammation and insulin resistance (19). In contrast, PUFAs serve as precursors for SPMs and inhibit NF-κB, thereby reducing pro-inflammatory cytokines production (58). It is important to note that exceptions within each fatty acid class have been reported in the literature, and readers are referred to the previous sections for further details. MUFAs, such as oleic acid, generally exert anti-inflammatory effects by increasing membrane fluidity, attenuating TLR signaling, and promoting oxidative metabolism (84). Moreover, MUFAs have been shown to counteract some of the proinflammatory actions of SFAs, thereby improving insulin sensitivity and reducing macrophage activation.
It is critical to recognize that a significant portion of the current evidence establishing a connection between macrophage polarization and fatty acids derived from in vitro studies that employ primary or immortalized cell culture systems. Despite the fact that these models offer mechanistic insights into lipid-immune interactions, they are unable to completely approximate the in vivo environment, which is characterized by the critical roles of tissue microenvironment, systemic metabolism, and immune crosstalk. Consequently, it is critical to ascertain the translational relevance of these findings by conducting studies in animal models. Ultimately, these findings need to be validated in human models. For instance, research conducted in mouse models with high-fat diets have shown that macrophage phenotypes in adipose tissue and liver are adjusted by dietary fatty acids (6, 85). Likewise, the in vivo validation of fatty acid-immune interactions was underscored by the alteration of mesenteric lymphocyte function in obese JCR rats by vaccenic acid supplementation (92). The integration of these models will be essential for bridging the knowledge gap between cellular discoveries and clinical applications.
In conclusion, the multifaceted interplay between fatty acids and macrophage function represents a compelling nexus with significant implications for human health. As research continues to unravel the complexities of these interactions, the potential to harness fatty acids as therapeutic agents for managing inflammatory and metabolic diseases emerges as a promising avenue for future research and clinical interventions. This review offers a comprehensive synthesis of current knowledge and underscores the need for further investigation into the intersection of fatty acid metabolism and macrophage biology, paving the way for innovative therapeutic strategies and translational applications.
Author contributions
MR-M: Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft. LL-M: Conceptualization, Methodology, Validation, Writing – original draft. GG-R: Conceptualization, Funding acquisition, Supervision, Validation, Writing – original draft. EC: Methodology, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. OL: Conceptualization, Funding acquisition, Methodology, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was supported in part by Tecnologico de Monterrey through the Challenge-based research projects IJXT070-22TE55001 and IJXT070-22EG57001; and the SECIHTI project CBF-2025-I-2520.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Essandoh K, Li Y, Huo J, and Fan GC. MiRNA-mediated macrophage polarization and its potential role in the regulation of inflammatory response. Shock. (2016) 46:122–31. doi: 10.1097/SHK.0000000000000604
2. Childs CE, Calder PC, and Miles EA. Diet and immune function. Nutrients. (2019) 11:1933. doi: 10.3390/nu11081933
3. Heieis GA, Patente TA, Almeida L, Vrieling F, Tak T, Perona-Wright G, et al. Metabolic heterogeneity of tissue-resident macrophages in homeostasis and during helminth infection. Nat Commun. (2023) 14:5627. doi: 10.1038/s41467-023-41353-z
4. Sender R, Weiss Y, Navon Y, Milo I, Azulay N, Keren L, et al. The total mass, number, and distribution of immune cells in the human body. Proc Natl Acad Sci. (2023) 120:e2308511120. doi: 10.1073/pnas.2308511120
5. Specht H, Emmott E, Petelski AA, Huffman RG, Perlman DH, Serra M, et al. Single-cell proteomic and transcriptomic analysis of macrophage heterogeneity using SCoPE2. Genome Biol. (2021) 22:50. doi: 10.1186/s13059-021-02267-5
6. Diehl KL, Vorac J, Hofmann K, Meiser P, Unterweger I, Kuerschner L, et al. Kupffer cells sense free fatty acids and regulate hepatic lipid metabolism in high-fat diet and inflammation. Cells. (2020) 9:2258. doi: 10.3390/cells9102258
7. Lendeckel U, Venz S, and Wolke C. Macrophages: shapes and functions. ChemTexts. (2022) 8:12. doi: 10.1007/s40828-022-00163-4
8. Redka DS, Gütschow M, Grinstein S, and Canton J. Differential ability of proinflammatory and anti-inflammatory macrophages to perform macropinocytosis. Mol Biol Cell. (2018) 29:53–65. Gruenberg JE. doi: 10.1091/mbc.E17-06-0419
9. Huang C, Du W, Ni Y, Lan G, and Shi G. The effect of short-chain fatty acids on M2 macrophages polarization in vitro and in vivo. Clin Exp Immunol. (2022) 207:53–64. doi: 10.1093/cei/uxab028
10. Hou C, Yan L, Sun K, Zhou T, Zou Y, Xiong W, et al. Nuclear receptor corepressor 1 deficiency exacerbates asthma by modulating macrophage polarization. Cell Death Discov. (2023) 9:429. doi: 10.1038/s41420-023-01724-3
11. Worthmann A, Ridder J, Piel SYL, Evangelakos I, Musfeldt M, Voß H, et al. Fatty acid synthesis suppresses dietary polyunsaturated fatty acid use. Nat Commun. (2024) 15:45. doi: 10.1038/s41467-023-44364-y
12. Luévano-Martínez LA, Méndez-Fernández A, Rueda-Munguía M, Bonilla-Ruelas D, Brunck MEG, and García-Rivas G. Glycerol metabolism is activated in both palmitic acid-stimulated and adipose tissue macrophages from a murine model of cardiometabolic heart failure. J Physiol. (2025) 20:JP287791. doi: 10.1113/JP287791
13. Jaitin DA, Adlung L, Thaiss CA, Weiner A, Li B, Descamps H, et al. Lipid-associated macrophages control metabolic homeostasis in a trem2-dependent manner. Cell. (2019) 178:686–98. doi: 10.1016/j.cell.2019.05.054
14. Wculek SK, Dunphy G, Heras-Murillo I, Mastrangelo A, and Sancho D. Metabolism of tissue macrophages in homeostasis and pathology. Cell Mol Immunol. (2022) 19:384–408. doi: 10.1038/s41423-021-00791-9
15. Li P, Hao Z, Wu J, Ma C, Xu Y, Li J, et al. Comparative proteomic analysis of polarized human THP-1 and mouse RAW264.7 macrophages. Front Immunol. (2021) 12:700009. doi: 10.3389/fimmu.2021.700009
16. Huynh K, Pernes G, Mellett NA, Meikle PJ, Murphy AJ, and Lancaster GI. Lipidomic profiling of murine macrophages treated with fatty acids of varying chain length and saturation status. Metabolites. (2018) 8:29. doi: 10.3390/metabo8020029
17. Oishi Y, Spann NJ, Link VM, Muse ED, Strid T, Edillor C, et al. SREBP1 contributes to resolution of pro-inflammatory TLR4 signaling by reprogramming fatty acid metabolism. Cell Metab. (2017) 25:412–27. doi: 10.1016/j.cmet.2016.11.009
18. Das UN. Saturated fatty acids, MUFAs and PUFAs regulate ferroptosis. Cell Chem Biol. (2019) 26:309–11. doi: 10.1016/j.chembiol.2019.03.001
19. Wang Y, Qian Y, Fang Q, Zhong P, Li W, Wang L, et al. Saturated palmitic acid induces myocardial inflammatory injuries through direct binding to TLR4 accessory protein MD2. Nat Commun. (2017) 8:13997. doi: 10.1038/ncomms13997
20. Wong SW, Kwon MJ, Choi AMK, Kim HP, Nakahira K, and Hwang DH. Fatty Acids Modulate Toll-like Receptor 4 Activation through Regulation of Receptor Dimerization and Recruitment into Lipid Rafts in a Reactive Oxygen Species-dependent Manner. J Biol Chem. (2009) 284:27384–92. doi: 10.1074/jbc.M109.044065
21. Zhang Y, Hao J, Sun Y, and Li B. Saturated fatty acids induce ceramide-associated macrophage cell death. J Vis Exp. (2017) 128):56535. doi: 10.3791/56535
22. Zhang Y, Luo L, Xu X, Wu J, Wang F, Lu Y, et al. Acetylation is required for full activation of the NLRP3 inflammasome. Nat Commun. (2023) 14:8396. doi: 10.1038/s41467-023-44203-0
23. Wu J, Fernandes-Alnemri T, and Alnemri ES. Involvement of the AIM2, NLRC4, and NLRP3 inflammasomes in caspase-1 activation by listeria monocytogenes. J Clin Immunol. (2010) 30:693–702. doi: 10.1007/s10875-010-9425-2
24. L’homme L, Esser N, Riva L, Scheen A, Paquot N, Piette J, et al. Unsaturated fatty acids prevent activation of NLRP3 inflammasome in human monocytes/macrophages. J Lipid Res. (2013) 54:2998–3008. doi: 10.1194/jlr.M037861
25. Man K, Kallies A, and Vasanthakumar A. Resident and migratory adipose immune cells control systemic metabolism and thermogenesis. Cell Mol Immunol. (2022) 19:421–31. doi: 10.1038/s41423-021-00804-7
26. Zhao M, Okunishi K, Bu Y, Kikuchi O, Wang H, Kitamura T, et al. Targeting activin receptor–like kinase 7 ameliorates adiposity and associated metabolic disorders. JCI Insight. (2023) 8:e161229. doi: 10.1172/jci.insight.161229
27. Heidari Seyedmahalleh M, Nasli-Esfahani E, Zeinalabedini M, and Azadbakht L. Association of ultra-processed food consumption with cardiovascular risk factors among patients with type-2 diabetes mellitus. Nutr Diabetes. (2024) 14:89. doi: 10.1038/s41387-024-00337-8
28. Lee SM, Son KN, Shah D, Ali M, Balasubramaniam A, Shukla D, et al. Histatin-1 attenuates LPS-induced inflammatory signaling in RAW264.7 macrophages. Int J Mol Sci. (2021) 22:7856. doi: 10.3390/ijms22157856
29. Haghikia A, Jörg S, Duscha A, Berg J, Manzel A, Waschbisch A, et al. Dietary fatty acids directly impact central nervous system autoimmunity via the small intestine. Immunity. (2015) 43:817–29. doi: 10.1016/j.immuni.2015.09.007
30. Deng L, Kersten S, and Stienstra R. Triacylglycerol uptake and handling by macrophages: From fatty acids to lipoproteins. Prog Lipid Res. (2023) 92:101250. doi: 10.1016/j.plipres.2023.101250
31. Burr SD, Chen Y, Hartley CP, Zhao X, and Liu J. Replacement of saturated fatty acids with linoleic acid in western diet attenuates atherosclerosis in a mouse model with inducible ablation of hepatic LDL receptor. Sci Rep. (2023) 13:16832. doi: 10.1038/s41598-023-44030-9
32. Roopashree PG, Shetty SS, Shetty VV, and Nalilu SK. Medium-chain fatty acids and breast cancer risk by receptor and pathological subtypes. Nutrients. (2022) 14:5351. doi: 10.3390/nu14245351
33. Morgan PK, Huynh K, Pernes G, Miotto PM, Mellett NA, Giles C, et al. Macrophage polarization state affects lipid composition and the channeling of exogenous fatty acids into endogenous lipid pools. J Biol Chem. (2021) 297:101341. doi: 10.1016/j.jbc.2021.101341
34. Zeng J, Zhang Y, Hao J, Sun Y, Liu S, Bernlohr DA, et al. Stearic acid induces CD11c expression in proinflammatory macrophages via epidermal fatty acid binding protein. J Immunol. (2018) 200:3407–19. doi: 10.4049/jimmunol.1701416
35. Yang P, Qin H, Li Y, Xiao A, Zheng E, Zeng H, et al. CD36-mediated metabolic crosstalk between tumor cells and macrophages affects liver metastasis. Nat Commun. (2022) 13:5782. doi: 10.1038/s41467-022-33349-y
36. Vazquez MM, Gutierrez MV, Salvatore SR, Puiatti M, Dato VA, Chiabrando GA, et al. Nitro-oleic acid, a ligand of CD36, reduces cholesterol accumulation by modulating oxidized-LDL uptake and cholesterol efflux in RAW264. 7 macrophages. Redox Biol. (2020) 36:101591. doi: 10.1016/j.redox.2020.101591
37. Kochumon S, Arefanian H, Azim R, Shenouda S, Jacob T, Abu Khalaf N, et al. Stearic acid and TNF-α Co-operatively potentiate MIP-1α Production in monocytic cells via myD88 independent TLR4/TBK/IRF3 signaling pathway. Biomedicines. (2020) 8:403. doi: 10.3390/biomedicines8100403
38. Wang S, Lin Y, Gao L, Yang Z, Lin J, Ren S, et al. PPAR-γ integrates obesity and adipocyte clock through epigenetic regulation of Bmal1. Theranostics. (2022) 12:1589–606. doi: 10.7150/thno.69054
39. Wang X, Jiang X, Deng B, Xiao J, Jin J, and Huang Z. Lipopolysaccharide and palmitic acid synergistically induced MCP-1 production via MAPK-meditated TLR4 signaling pathway in RAW264.7 cells. Lipids Health Dis. (2019) 18:71. doi: 10.1186/s12944-019-1017-4
40. Chan KL, Pillon NJ, Sivaloganathan DM, Costford SR, Liu Z, Théret M, et al. Palmitoleate reverses high fat-induced proinflammatory macrophage polarization via AMP-activated protein kinase (AMPK). J Biol Chem. (2015) 290:16979–88. doi: 10.1074/jbc.M115.646992
41. Talbot NA, Wheeler-Jones CP, and Cleasby ME. Palmitoleic acid prevents palmitic acid-induced macrophage activation and consequent p38 MAPK-mediated skeletal muscle insulin resistance. Mol Cell Endocrinol. (2014) 393:129–42. doi: 10.1016/j.mce.2014.06.010
42. Howe AM, Burke S, O’Reilly ME, McGillicuddy FC, and Costello DA. Palmitic acid and oleic acid differently modulate TLR2-mediated inflammatory responses in microglia and macrophages. Mol Neurobiol. (2022) 59:2348–62. doi: 10.1007/s12035-022-02756-z
43. Yang Z, Kahn BB, Shi H, and Zhong XB. Macrophage α1 AMP-activated protein kinase (α1AMPK) antagonizes fatty acid-induced inflammation through SIRT1. J Biol Chem. (2010) 285:19051–9. doi: 10.1074/jbc.M110.123620
44. Shao D, Lian Z, Di Y, Zhang L, Rajoka MSR, Zhang Y, et al. Dietary compounds have potential in controlling atherosclerosis by modulating macrophage cholesterol metabolism and inflammation via miRNA. NPJ Sci Food. (2018) 2:13. doi: 10.1038/s41538-018-0022-8
45. Tham YY, Choo QC, Muhammad TST, and Chew CH. Lauric acid alleviates insulin resistance by improving mitochondrial biogenesis in THP-1 macrophages. Mol Biol Rep. (2020) 47:9595–607. doi: 10.1007/s11033-020-06019-9
46. Lim WS, Gan MSY, Ong MHL, and Chew CH. Lauric acid abolishes interferon-gamma (IFN-γ)-induction of intercellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1) expression in human macrophages. Asian Pac J Reprod. (2015) 4:217–21. doi: 10.1016/j.apjr.2015.06.005
47. Saraswathi V, Kumar N, Ai W, Gopal T, Bhatt S, Harris EN, et al. Myristic acid supplementation aggravates high fat diet-induced adipose inflammation and systemic insulin resistance in mice. Biomolecules. (2022) 12:739. doi: 10.3390/biom12060739
48. Hung HC, Tsai SF, Chou HW, Tsai MJ, Hsu PL, and Kuo YM. Dietary fatty acids differentially affect secretion of pro-inflammatory cytokines in human THP-1 monocytes. Sci Rep. (2023) 13:5511. doi: 10.1038/s41598-023-32710-5
49. Zhang X, Xue C, Xu Q, Zhang Y, Li H, Li F, et al. Caprylic acid suppresses inflammation via TLR4/NF-κB signaling and improves atherosclerosis in ApoE-deficient mice. Nutr Metab. (2019) 16:40. doi: 10.1186/s12986-019-0359-2
50. Yu S, Choi JH, Kim HJ, Park SH, woong G, and Kim W. In vitro evidence of anti-inflammatory and anti-obesity effects of medium-chain fatty acid-diacylglycerols. J Microbiol Biotechnol. (2017) 27:1617–27. doi: 10.4014/jmb.1703.03071
51. Gaete PV, Nieves-Barreto LD, Guatibonza-García V, Losada-Barragán M, Vargas-Sánchez K, and Mendivil CO. Medium-chain fatty acids modify macrophage expression of metabolic and inflammatory genes in a PPAR β/δ-dependent manner. Sci Rep. (2023) 13:11573. doi: 10.1038/s41598-023-38700-x
52. Tashiro H, Takahashi K, Sadamatsu H, Kato G, Kurata K, Kimura S, et al. Saturated fatty acid increases lung macrophages and augments house dust mite-induced airway inflammation in mice fed with high-fat diet. Inflammation. (2017) 40:1072–86. doi: 10.1007/s10753-017-0550-4
53. Fernandes HB, De Oliveira IM, Postler TS, Lima SQ, Santos CAC, Oliveira MS, et al. Transcriptomic analysis reveals that RasGEF1b deletion alters basal and LPS-induced expression of genes involved in chemotaxis and cytokine responses in macrophages. Sci Rep. (2023) 13:19614. doi: 10.1038/s41598-023-47040-9
54. Fidalgo Rodríguez JL, Dynarowicz-Latka P, and Minones Conde J. Interactions of cholesterol and 7−ketocholesterol with unsaturated fatty acids of different unsaturation degree – The monolayer study. Biochim Biophys Acta BBA - Biomembr. (2019) 1861:1428–36. doi: 10.1016/j.bbamem.2019.06.002
55. Yamada H, Umemoto T, Kakei M, Momomura SI, Kawakami M, Ishikawa SE, et al. The effect of omega-3 fatty acid supplementation on weight loss and cognitive function in overweight or obese individuals on weight-loss diet. Nutr Hosp. (2022) 39:803–813. Available online at: https://www.nutricionhospitalaria.org/articles/03992/show.
56. Kim KH, Kim Y, and Seo KW. Efficacy of monounsaturated fatty acids in reducing risk of the cardiovascular diseases, cancer, inflammation, and insulin resistance: a narrative review. Ann Clin Nutr Metab. (2023) 15:2–7. doi: 10.15747/ACNM.2023.15.1.2
57. Lee KR, Midgette Y, and Shah R. Fish oil derived omega 3 fatty acids suppress adipose NLRP3 inflammasome signaling in human obesity. J Endocr Soc. (2019) 3:504–15. doi: 10.1210/js.2018-00220
58. Kuda O, Brezinova M, Rombaldova M, Slavikova B, Posta M, Beier P, et al. Docosahexaenoic acid–derived fatty acid esters of hydroxy fatty acids (FAHFAs) with anti-inflammatory properties. Diabetes. (2016) 65:2580–90. doi: 10.2337/db16-0385
59. Xu M, Wang X, Li Y, Geng X, Jia X, Zhang L, et al. Arachidonic acid metabolism controls macrophage alternative activation through regulating oxidative phosphorylation in PPARγ Dependent manner. Front Immunol. (2021) 12:618501. doi: 10.3389/fimmu.2021.618501
60. Shiomi N and Watanabe K. Effects of oleic acid on murine macrophage dysfunction. J BioMed Sci Eng. (2013) 06:654–60. doi: 10.4236/jbise.2013.66080
61. Pauls SD, Rodway LA, Winter T, Taylor CG, Zahradka P, and Aukema HM. Anti-inflammatory effects of α-linolenic acid in M1-like macrophages are associated with enhanced production of oxylipins from α-linolenic and linoleic acid. J Nutr Biochem. (2018) 57:121–9. doi: 10.1016/j.jnutbio.2018.03.020
62. Von Gerichten J, West AL, Irvine NA, Miles EA, Calder PC, Lillycrop KA, et al. Oxylipin secretion by human CD3+ T lymphocytes in vitro is modified by the exogenous essential fatty acid ratio and life stage. Front Immunol. (2023) 14:1206733. doi: 10.3389/fimmu.2023.1206733
63. Onodera T, Fukuhara A, Shin J, Hayakawa T, Otsuki M, and Shimomura I. Eicosapentaenoic acid and 5-HEPE enhance macrophage-mediated Treg induction in mice. Sci Rep. (2017) 7:4560. doi: 10.1038/s41598-017-04474-2
64. O’Reilly ME, Lenighan YM, Dillon E, Kajani S, Curley S, Bruen R, et al. Conjugated linoleic acid and alpha linolenic acid improve cholesterol homeostasis in obesity by modulating distinct hepatic protein pathways. Mol Nutr Food Res. (2020) 64:1900599. doi: 10.1002/mnfr.201900599
65. Serhan CN. Pro-resolving lipid mediators are leads for resolution physiology. Nature. (2014) 510:92–101. doi: 10.1038/nature13479
66. jie KS and Garg NJ. Metabolic programming of macrophage functions and pathogens control. Redox Biol. (2019) 24:101198. doi: 10.1016/j.redox.2019.101198
67. Herová M, Schmid M, Gemperle C, and Hersberger M. ChemR23, the receptor for chemerin and resolvin E1, is expressed and functional on M1 but not on M2 macrophages. J Immunol. (2015) 194:2330–7. doi: 10.4049/jimmunol.1402166
68. Schmid M, Gemperle C, Rimann N, and Hersberger M. Resolvin D1 Polarizes Primary Human Macrophages toward a Proresolution Phenotype through GPR32. J Immunol. (2016) 196:3429–37. doi: 10.4049/jimmunol.1501701
69. Arita M, Yoshida M, Hong S, Tjonahen E, Glickman JN, Petasis NA, et al. Resolvin E1, an endogenous lipid mediator derived from omega-3 eicosapentaenoic acid, protects against 2,4,6-trinitrobenzene sulfonic acid-induced colitis. Proc Natl Acad Sci. (2005) 102:7671–6. doi: 10.1073/pnas.0409271102
70. Krishnamoorthy N, Walker KH, Brüggemann TR, Tavares LP, Smith EW, Nijmeh J, et al. The Maresin 1–LGR6 axis decreases respiratory syncytial virus-induced lung inflammation. Proc Natl Acad Sci. (2023) 120:e2206480120. doi: 10.1073/pnas.2206480120
71. Liu S, Bai T, Liu X, Zhao W, Li X, Sui Y, et al. Role and regulatory mechanism of GPR37 in neurological diseases. Front Cell Neurosci. (2025) 19:1617682. doi: 10.3389/fncel.2025.1617682
72. Shramko VS, Striukova EV, Polonskaya YV, Stakhneva EM, Volkova MV, Kurguzov AV, et al. Associations of antioxidant enzymes with the concentration of fatty acids in the blood of men with coronary artery atherosclerosis. J Pers Med. (2021) 11:1281. doi: 10.3390/jpm11121281
73. Hennig B, Lei W, Arzuaga X, Ghosh DD, Saraswathi V, and Toborek M. Linoleic acid induces proinflammatory events in vascular endothelial cells via activation of PI3K/Akt and ERK1/2 signaling. J Nutr Biochem. (2006) 17:766–72. doi: 10.1016/j.jnutbio.2006.01.005
74. Abdou AG, Bendary MA, Abdou SE, and Amer GS. Impact of conjugated linoleic acid on obesity and its association with macrophage recruitment: experimental and immunohistochemical study. J Microsc Ultrastruct. (2024) 12:142–7. doi: 10.4103/jmau.jmau_25_22
75. Kanter JE, Goodspeed L, Wang S, Kramer F, Wietecha T, Gomes-Kjerulf D, et al. 10,12 conjugated linoleic acid-driven weight loss is protective against atherosclerosis in mice and is associated with alternative macrophage enrichment in perivascular adipose tissue. Nutrients. (2018) 10:1416. doi: 10.3390/nu10101416
76. Chang CS, Sun HL, Lii CK, Chen HW, Chen PY, and Liu KL. Gamma-linolenic acid inhibits inflammatory responses by regulating NF-κB and AP-1 activation in lipopolysaccharide-induced RAW 264.7 macrophages. Inflammation. (2010) 33:46–57. doi: 10.1007/s10753-009-9157-8
77. Baker EJ, Valenzuela CA, Van Dooremalen WTM, Martínez-Fernández L, Yaqoob P, Miles EA, et al. Gamma-linolenic and pinolenic acids exert anti-inflammatory effects in cultured human endothelial cells through their elongation products. Mol Nutr Food Res. (2020) 64:2000382. doi: 10.1002/mnfr.202000382
78. Félix-Soriano E, Sáinz N, Fernández-Galilea M, Gil-Iturbe E, Celay J, Martínez-Climent JA, et al. Chronic docosahexaenoic acid supplementation improves metabolic plasticity in subcutaneous adipose tissue of aged obese female mice. J Nutr Biochem. (2023) 111:109153. doi: 10.1016/j.jnutbio.2022.109153
79. Talamonti E, Pauter AM, Asadi A, Fischer AW, Chiurchiù V, and Jacobsson A. Impairment of systemic DHA synthesis affects macrophage plasticity and polarization: implications for DHA supplementation during inflammation. Cell Mol Life Sci. (2017) 74:2815–26. doi: 10.1007/s00018-017-2498-9
80. Yamada H, Umemoto T, Kakei M, Momomura SI, Kawakami M, Ishikawa SE, et al. Eicosapentaenoic acid shows anti-inflammatory effect via GPR120 in 3T3-L1 adipocytes and attenuates adipose tissue inflammation in diet-induced obese mice. Nutr Metab. (2017) 14:33. doi: 10.1186/s12986-017-0188-0
81. Yang N, Tang Q, Qin W, Li Z, Wang D, Zhang W, et al. Treatment of obesity-related inflammation with a novel synthetic pentacyclic oleanane triterpenoids via modulation of macrophage polarization. EBioMedicine. (2019) 45:473–86. doi: 10.1016/j.ebiom.2019.06.053
82. Xu R, Dai Y, Zheng X, Yan Y, He Z, Zhang H, et al. Thromboxane A2-TP axis promotes adipose tissue macrophages M1 polarization leading to insulin resistance in obesity. Biochem Pharmacol. (2023) 210:115465. doi: 10.1016/j.bcp.2023.115465
83. Salama A, Amin MM, and Hassan A. Effects of oleic acid and/or exercise on diet-induced thermogenesis and obesity in rats: involvement of beige adipocyte differentiation and macrophage M1 inhibition. Res Pharm Sci. (2023) 18:219–30. doi: 10.4103/1735-5362.367800
84. Camell C and Smith CW. Dietary oleic acid increases M2 macrophages in the mesenteric adipose tissue. Bassaganya-Riera J editor. PloS One. (2013) 8:e75147. doi: 10.1371/journal.pone.0075147
85. Liang Q, Zheng Y, Meng F, Jiang X, Zhen Q, Lu Z, et al. Palmitoleic acid on top of HFD ameliorates insulin resistance independent of diacylglycerols and alters gut microbiota in C57BL/6J mice. Food Sci Hum Wellness. (2024) 13:856–68. doi: 10.26599/FSHW.2022.9250073
86. Lee C, Barnett J, and Reaven PD. Liposomes enriched in oleic acid are less susceptible to oxidation and have less proinflammatory activity when exposed to oxidizing conditions. J Lipid Res. (1998) 39:1239–47. doi: 10.1016/S0022-2275(20)32548-7
87. Sun Y, Wang J, Guo X, Zhu N, Niu L, Ding X, et al. Oleic acid and eicosapentaenoic acid reverse palmitic acid-induced insulin resistance in human hepG2 cells via the reactive oxygen species/JUN pathway. Genomics Proteomics Bioinf. (2021) 19:754–71. doi: 10.1016/j.gpb.2019.06.005
88. Takahashi A, Ishizaki M, Kimira Y, Egashira Y, and Hirai S. Erucic acid-rich yellow mustard oil improves insulin resistance in KK-ay mice. Molecules. (2021) 26:546. doi: 10.3390/molecules26030546
89. Zuo S, Wang Y, Bao H, Zhang Z, Yang N, Jia M, et al. Lipid synthesis, triggered by PPARγ T166 dephosphorylation, sustains reparative function of macrophages during tissue repair. Nat Commun. (2024) 15:7269. doi: 10.1038/s41467-024-51736-5
90. Zizzo G and Cohen PL. The PPAR-γ antagonist GW9662 elicits differentiation of M2c-like cells and upregulation of the MerTK/Gas6 axis: a key role for PPAR-γ in human macrophage polarization. J Inflamm. (2015) 12:36. doi: 10.1186/s12950-015-0081-4
91. Yuan SN, Feng CY, Rong Y, Guo X, Chen YY, Fraga-Corral M, et al. Immunomodulatory effects of nervonic acid in RAW264.7 macrophages. J Food Biochem. (2023) 992311. doi: 10.1155/2023/9923111
Keywords: macrophages, fatty acids, inflammation, metabolic diseases, immunometabolism
Citation: Rueda-Munguía M, Luévano-Martínez LA, García-Rivas G, Castillo EC and Lozano O (2025) Macrophages: their role in immunity and their relationship with fatty acids in health and disease. Front. Immunol. 16:1694892. doi: 10.3389/fimmu.2025.1694892
Received: 29 August 2025; Accepted: 14 November 2025; Revised: 10 October 2025;
Published: 02 December 2025.
Edited by:
Fabrizia Bonacina, University of Milan, ItalyReviewed by:
Jin Young Huh, Sogang University, Republic of KoreaCopyright © 2025 Rueda-Munguía, Luévano-Martínez, García-Rivas, Castillo and Lozano. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Omar Lozano, b21hci5sb3phbm9AdGVjLm14; Elena Cristina Castillo, ZWNnb256YWxlekB0ZWMubXg=