 Frédérique Le Roux1,2*K. Mathias Wegner3Craig Baker-Austin4Luigi Vezzulli5Carlos R. Osorio6Carmen Amaro7Jennifer M. Ritchie8Tom Defoirdt9Delphine Destoumieux-Garzón10Melanie Blokesch11Didier Mazel12Annick Jacq13Felipe Cava14Lone Gram15Carolin C. Wendling16Eckhard Strauch17Alexander Kirschner18Stephan Huehn19
Frédérique Le Roux1,2*K. Mathias Wegner3Craig Baker-Austin4Luigi Vezzulli5Carlos R. Osorio6Carmen Amaro7Jennifer M. Ritchie8Tom Defoirdt9Delphine Destoumieux-Garzón10Melanie Blokesch11Didier Mazel12Annick Jacq13Felipe Cava14Lone Gram15Carolin C. Wendling16Eckhard Strauch17Alexander Kirschner18Stephan Huehn19- 1Unié Physiologie Fonctionnelle des Organismes Marins, Ifremer, Plouzané, France
- 2CNRS, UMR 8227, Integrative Biology of Marine Models, Station Biologique de Roscoff, Sorbonne Universités, UPMC Paris 06, Roscoff cedex, France
- 3Coastal Ecology, Alfred Wegener Institute Helmholtz Centre for Polar and Marine Research, List, Germany
- 4Cefas, Weymouth, UK
- 5Department of Earth, Environmental and Life Sciences, University of Genoa, Genoa, Italy
- 6Departamento de Microbioloxía e Parasitoloxía, Instituto de Acuicultura, Universidade de Santiago de Compostela, Santiago de Compostela, Spain
- 7Estructura de Investigación Interdisciplinar en Biotecnología y Biomedicina, Department of Microbiology and Ecology, University of Valencia, Valencia, Spain
- 8Faculty of Health and Medical Sciences, University of Surrey, Guildford, UK
- 9UGent Aquaculture R&D Consortium, Ghent University, Ghent, Belgium
- 10Interactions Hôtes-Pathogènes-Environnements, UMR 5244, CNRS, Ifremer, Université de Perpignan Via Domita, Université de Montpellier, Montpellier, France
- 11Laboratory of Molecular Microbiology, Global Health Institute, School of Life Sciences, Ecole Polytechnique Fédérale de Lausanne, Lausanne, Switzerland
- 12Département Génomes et Génétique, CNRS UMR3525, Unité Plasticité du Génome Bactérien, Institut Pasteur, Paris, France
- 13Institute for Integrative Biology of the Cell, CEA, CNRS, Université Paris-Sud, Orsay, France
- 14Laboratory for Molecular Infection Medicine Sweden, Department of Molecular Biology, Umeå Centre for Microbial Research, Umeå University, Umeå, Sweden
- 15Department of Systems Biology, Technical University of Denmark, Kongens Lyngby, Denmark
- 16Geomar, Helmholtz Centre for Ocean Research Kiel, Kiel, Germany
- 17Federal Institute for Risk Assessment, National Reference Laboratory for Monitoring Bacteriological Contamination of Bivalve Molluscs, Berlin, Germany
- 18Institute for Hygiene and Applied Immunology, Medical University of Vienna, Vienna, Austria
- 19Institute of Food Hygiene, Free University Berlin, Berlin, Germany
Global change has caused a worldwide increase in reports of Vibrio-associated diseases with ecosystem-wide impacts on humans and marine animals. In Europe, higher prevalence of human infections followed regional climatic trends with outbreaks occurring during episodes of unusually warm weather. Similar patterns were also observed in Vibrio-associated diseases affecting marine organisms such as fish, bivalves and corals. Basic knowledge is still lacking on the ecology and evolutionary biology of these bacteria as well as on their virulence mechanisms. Current limitations in experimental systems to study infection and the lack of diagnostic tools still prevent a better understanding of Vibrio emergence. A major challenge is to foster cooperation between fundamental and applied research in order to investigate the consequences of pathogen emergence in natural Vibrio populations and answer federative questions that meet societal needs. Here we report the proceedings of the first European workshop dedicated to these specific goals of the Vibrio research community by connecting current knowledge to societal issues related to ocean health and food security.
State of the Art and Perspectives of Vibrio Research in Europe
According to the European Environment Agency the rise of global sea surface temperature (SST) is one of the major physical impacts of climate change. However, SST in coastal European seas has increased 4–7 times faster over the past few decades than in the global oceans (Reid et al., 2011). This local increase in SST has been linked to outbreaks of Vibrio-associated human illness caused by Vibrio cholerae non O1-non-O139, V. parahaemolyticus, and V. vulnificus in several European countries (Table 1). However, the lack of mandatory notification systems for Vibrio-associated illnesses prevents accurate estimates of the number of Vibrio infections occurring in Europe. Also mass mortalities of marine animals increase in frequency (Table 1), particularly in heavily polluted coastal areas, suggesting human activities as a factor favoring disease epidemics. Prominent examples include several Vibrio species associated with the recent great devastation of oyster beds in France. The salmonid farming industry is constantly threatened by V. salmonicida and V. anguillarum. Moreover, different subspecies of Photobacterium damselae are associated with diseases in cultured fish species like sole, sea bass, sea bream and turbot, while V. vulnificus causes hemorrhagic septicaemia in eel, derbio, tilapia, trout and shrimps but can also cause septicemia in humans. Finally, evidence has accumulated linking Vibrio infections (e.g., V. coralliilyticus) to increasing mass mortalities of benthic corals (e.g., Paramuricea clavata) in the NW Mediterranean Sea.
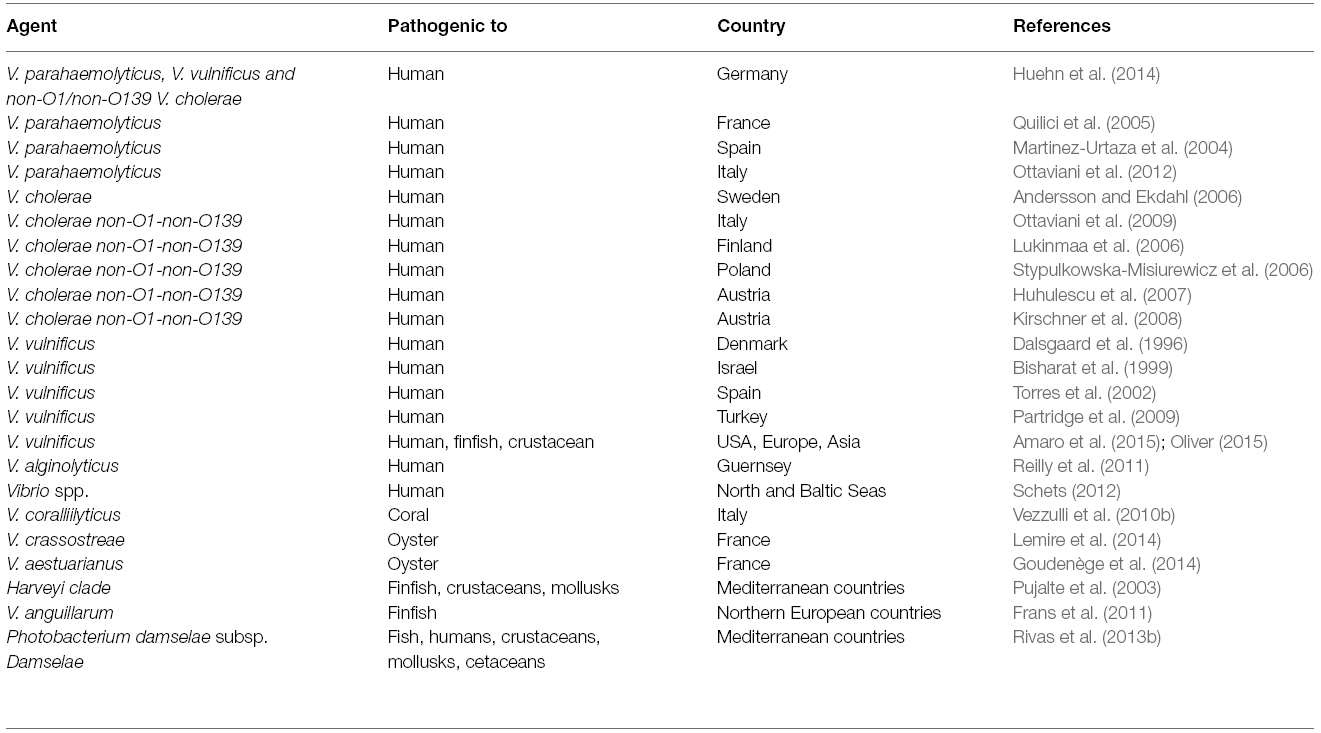
Table 1. Recent Vibrio-associated diseases caused by Vibrio in Europe.
To cover the large diversity of infectious vibrios, the development of operational tools to identify and detect emergent pathogens is essential to zoosanitary monitoring of cultivated species as well as on wild animal populations. Yet, compared to human pandemic strains, little is known about the virulence mechanisms of emergent environmental vibrios. This lack of knowledge may be attributed to the high genetic diversity of Vibrio isolates and the diversity/plurality of virulence mechanisms. To date pathogenic capacity cannot be inferred by taxonomic affiliation, because virulence factors (e.g., secretion systems, toxins) are rarely species-specific and are often shared between Vibrio species by lateral gene transfer. On top of that there are very few animal models to distinguish pathogenic strains and extend our understanding of the mechanisms involved in host-microbe interactions. Hence the elucidation of virulence for agent and target is a prerequisite to develop prophylactic methods to fight infectious diseases.
Due to the extent of the environmental, economical, and public health consequences resulting from Vibrio infections, a large scientific community is working on these bacteria in Europe. In order to join fundamental and applied research teams and to investigate the emergence of pathogens in natural Vibrio populations, we organized the first European workshop dedicated to the research on vibrios in Paris (11–12th March 2015), that provided a forum for experts in Vibrio ecology, evolution and pathogenesis to address societal issues involving ocean health and food security.
Vibrio Spread in Europe linked with climate change
Vibrios preferentially grow in warm (>15°C) saline aquatic environments. Warming of marine and saline inland waters is likely to support larger numbers of Vibrio populations and consequently an increased risk of Vibrio infections. An increase in the prevalence of human infections caused by V. parahaemolyticus, V. cholerae non-O1-non-O139 and V. vulnificus has been recorded in Europe even at high latitudes (Baker-Austin et al., 2013). In northern Europe, the increase in reported infections corresponds both in time and space with spikes in domestically-acquired Vibrio cases in “heatwave” years. Similarly, samples collected in the last 60 years by the continuous plankton recorder (CPR) survey (Vezzulli et al., 2012) showed that the genus Vibrio, including the human pathogen V. cholerae, has increased in prevalence in the last 44 years in the coastal North Sea, and that this increase is correlated with warming SST. Elevated water temperatures might also facilitate the successful invasion of pathogenic variants via food trade (Nair et al., 2007), ballast water (Dobbs et al., 2013), travelers (Fillion and Mileno, 2015) or natural animals. For example, migrating birds may act as vectors of intercontinental transport of V. cholerae (Vezzulli et al., 2010a). The direct comparison of the population structure of V. cholerae from a major bird sanctuary (Lake Neusiedl, Austria), with strains collected from six other European countries revealed that several strains in the lake shared the same alleles with other European strains, consistent with pan-European transport between distant ecosystems via birds (A. Kirschner, unpublished data). As a future challenge, macro-ecological studies on the impact of climate change on Vibrio persistence and spread in the aquatic environment combined with studies investigating climate change effects on epidemiologically relevant variables, such as host susceptibility and exposure are needed to significantly improve prediction and mitigation strategies against the future occurrence of Vibrio disease outbreaks.
Virulence as a Function of Biotic Interactions With Host and Microbiome
Virulence is a widespread phenomenon across the Vibrio phylogeny (Wendling et al., 2014). Its expression critically depends on biotic interactions with the host but also with other resident microbiota. On the host side, spatially-structured cross-infection experiments indicated that virulence of only distantly related Vibrio strains was lower when infecting oysters from the same geographic location. This suggests that oyster hosts are locally adapted and have evolved resistance to genetic factors shared within Vibrio populations (Wendling and Wegner, 2015). When considering interactions of Vibrio with the resident microbiome, the hemolymph microbiome modulates infections but is vulnerable to environmental disturbance (Lokmer and Wegner, 2015). Accordingly, Vibrio disease cannot be seen as an isolated event but needs to be considered in the context of the microbiome, which includes other non-virulent Vibrio. Indeed, the successive replacement of non-virulent with virulent strains during oyster infections occurs in the natural environment (Lemire et al., 2014) and the amplification of virulence in the presence of non-virulent strains suggests that also non-virulent strains contribute directly or indirectly to the development of disease. Future research on Vibrio disease should therefore focus on the higher order biotic interactions between the environment, the host and the pathogenic as well as the non-pathogenic fractions of microbial communities.
A key feature of the interaction between microbes within a community is the production of molecules that determine behavior like antagonism, competition or cooperation. Cell-to-cell communication in vibrios coordinates virulence gene expression based on the biotic and abiotic environment (Defoirdt, 2014). For example, the three-channel quorum sensing (QS) system of V. harveyi controls the pathogenicity of the bacterium toward different aquatic hosts (Defoirdt and Sorgeloos, 2012; Pande et al., 2013), and our most recent research revealed that another signaling molecule, indole, controls the virulence of V. anguillarum toward sea bass larvae (Li et al., 2014b). Another potential signaling mechanism has been described based on the production and release of high concentrations of D-amino acids into the extracellular milieu (Lam et al., 2009). First discovered in V. cholerae, these D-amino acids are different from those known to be part of the cell wall in bacteria (D-Ala and D-Glu) and were therefore called non-canonical D-amino acids (NCDAAs; Cava et al., 2011a). NCDAAs released into the media by producer strains can affect non-producer organisms beneficially or detrimentally in a particular niche (Cava et al., 2011b; Alvarez et al., 2014). The possible implications of NCDAAs in the biological processes of co-inhabitants still remains to be investigated but the enormous energy demand suggests that these molecules should have a great impact in poly-microbial communities. Finally several strains of Vibrio have been demonstrated to produce potent antibacterial agents (andrimid and holomycin) or agents that block QS regulated genes (solonamides, ngercheumicin) in human pathogens (Wietz et al., 2010; Mansson et al., 2011; Kjaerulff et al., 2013; Nielsen et al., 2014). Comparative and functional genomics using software like antiSMASH (Medema et al., 2011) could identify the genetic determinants of these secondary metabolites (polyketide synthases and non-ribosomal peptide synthetases). Hence further elucidation of virulence regulatory mechanisms will enable us to better understand Vibrio-host interactions and ecology, and to identify targets for the design of novel agents to control disease caused by vibrios.
Horizontal Gene Transfer, Genome Plasticity, and Chromosome Partitioning
Evolution of Vibrio species is often driven by mobile genetic elements via horizontal gene transfer (HGT). However, very little is known about HGT in environmental Vibrio isolates infecting marine organisms. In 75 marine Vibrio spp. isolated from the broad-nosed pipefish, Syngnathus typhle, associated prophages were characterized and the virulence of strains carrying different prophages was then assessed by comparing the relative expression of 44 immune genes during controlled infection experiments on juvenile pipefish. Preliminary results suggest that virulence is significantly influenced by the associated prophages, further supporting a role for bacteriophages in manipulating the virulence of environmental Vibrio isolates (C. Wendling unpublished data).
Virulence of V. vulnificus and P. damselae subsp. damselae in fish is determined by transferable plasmids (pVvbt2 in V. vulnificus and pPHDD1 in P. damselae). pVvbt2 contains two highly conserved virulence genes involved in serum resistance (vep07) and the ability to grow from eel transferrin (vep20) (Pajuelo et al., 2015). Interestingly, pPHDD1 also contains vep07 and vep20 homologs suggesting that both genes are involved in resistance to fish innate immunity. pVvbt2 also encodes RtxA13, a toxin belonging to MARTX (multifunctional, autoprocessive, repeat in toxin) family. RtxA13 is considered a host-non-specific virulence factor because it is involved in resistance to phagocytosis by murine and human phagocytes as well as in eel death (Lee et al., 2013). The other virulence plasmid, pPHDD1, encodes phospholipase-D damselysin (Dly) and the pore-forming toxin HlyApl (Rivas et al., 2011). A second HlyA (HlyAch) is encoded in chromosome I (Rivas et al., 2013a, 2014) and the three toxins contribute to hemolysis and virulence, and are secreted by a type-two secretion system (Rivas et al., 2015). While the two HlyA hemolysis produce an additive effect, Dly and any of the two HlyA interact in a synergistic manner, being responsible for maximal virulence for fish and for mice (Rivas et al., 2013a). Due to their host range and their duality as pathogens for both poikilotherm and homeotherm animals, P. damselae and V. vulnificus constitute valuable biological models to study the role of mobile genetic elements in the rise of novel pathogenic strategies.
Vibrios contain large chromosomal integrons (Cambray et al., 2010) and belong to the group of naturally competent bacteria, which allows them to absorb free DNA from their surrounding environment and recombine it into their genome (Seitz and Blokesch, 2013a). For V. cholerae, entry into competence is tightly regulated and requires growth to high cell densities on chitinous surfaces (Meibom et al., 2005; Lo Scrudato and Blokesch, 2012, 2013). Uptake of external DNA is accomplished by a sophisticated DNA-uptake machinery (Seitz and Blokesch, 2013b, 2014; Seitz et al., 2014). As the competence regulon also encompasses the type VI secretion system-encoding gene clusters, HGT is enhanced through deliberate killing of neighboring non-sibling cells followed by the transfer of their DNA (Borgeaud et al., 2015).
The presence of two chromosomes is another characteristic feature of vibrios. While distinctive localization patterns have been described for the two chromosomes, the selective advantages brought by this bipartite architecture are still under debate (Val et al., 2012, 2014). Replication of both chromosomes is tightly coupled so that replication termination is synchronized (Rasmussen et al., 2007). Moreover, the chromosomal position of genes determines the relative copy number during growth thereby impacting the bacteriums physiology (Soler-Bistue et al., 2015). Notably, mechanistic aspects of chromosome organization, architecture, and cell cycle-dependent dynamics are only starting to be deciphered (Yamaichi et al., 2012; Demarre et al., 2014). The elucidation of the mechanisms that coordinate the interplay between chromosomes, accessory replicons, mobile DNA and HGT mechanisms is essential to better apprehend the evolution and niche adaptation of Vibrio species.
Adaptation of Pathogenic Vibrios to Intracellular Life
The pathogenic V. tasmaniensis strain LGP32, a member of the V. splendidus clade (Gay et al., 2004) was found to be a facultative intracellular pathogen of oyster immune cells called hemocytes (Duperthuy et al., 2011). This is a rare example of Vibrio adapted to intracellular life. The virulence of LGP32 in oysters correlated with the ability to enter hemocytes (Duperthuy et al., 2010, 2011). Both cellular invasion and pathogenicity depend on the major outer membrane protein OmpU, which serves as an adhesin to invade host cells. Once inside the phagosome, LGP32 releases outer membrane vesicles (OMVs) that protect the organism against antimicrobial peptides and act as vehicles for the delivery of virulence factors (Destoumieux-Garzón et al., 2014; Vanhove et al., 2015). Moreover, entry into hemocytes and intracellular survival of LGP32 are required for expression of LGP32 cytotoxicity toward hemocytes. This capacity to survive intracellularly relies on potent antioxidant and copper tolerance responses, both of which are highly induced in the hostile environment of the phagosome.
Small regulatory RNAs have been shown to play important roles in regulating virulence gene expression in response to conditions encountered in the host. sRNAs present in multicopies such as Qrrs and CsrBs were found in several Vibrio spp. to mediate QS regulation of virulence gene expression (Nguyen and Jacq, 2014). One peculiarity of the Splendidus clade seems to be the presence of four highly expressed copies of the CsrB sRNAs in their genome, instead of 2–3 found in other vibrios (Lenz et al., 2005; Toffano-Nioche et al., 2012). CsrB sRNAs are highly transcribed inside oyster hemocytes suggesting a role in adaptation to the intracellular environment (Vanhove et al., submitted). The landscape and phylogeny of putative sRNAs encoded by LGP32 demonstrate rapid vertical evolution, with a vast majority of sRNAs being species/strain specific, and only a small number (28/250) conserved in all Vibrio sequenced so far (Toffano-Nioche et al., 2012). Thus, sRNAs contribute to a high diversity between species and provide opportunities for adaptation/colonization of new hosts and virulence emergence, a question that will be tackled by comparative functional studies of conserved Vibrio sRNAs.
Model Systems to Study Pathogenicity Mechanisms and Vibrio-host Interactions
Microbiologists are increasingly aware that how organisms behave in situ in the “real world” might be distinct from those that occur in laboratory monocultures grown under tightly controlled conditions (Smith, 2000). Thus, model systems, which replicate at least part of the natural processes of infection, are needed in order to examine the relevance and biological impact of in vitro findings. In vivo models that reproduce the main clinical and pathological signs of disease seen following the consumption of contaminated food or water, are available for toxigenic and non-toxigenic V. cholerae, and for V. parahaemolyticus (Ritchie et al., 2010, 2012; Shin et al., 2011). In these studies, a combination of microbiological, histological and genetic analysis was used to identify key virulence factors and the pathologic mechanisms associated with the respective strains (e.g., see Zhou et al., 2013, 2014). However, a growing number of Vibrio-associated illnesses are associated with a diverse group of strains, some of which lack known virulence factors (Garcia et al., 2009; Jones et al., 2012; Ottaviani et al., 2012). A future challenge will be to examine the pathogenesis of these strains and identify additional virulence markers, which should be used to improve risk assessment tools targeted to the different pathogens. Furthermore, a growing number of human Vibrio infections in Europe were not food-borne, but instead associated with the ability of non-O1-non-O139 V. cholerae, V. parahaemolyticus, or V. vulnificus to cause septicemia via wound infections (Table 1). Models to examine this aspect of their pathogenicity are currently lacking and should become a high priority given the poor prognosis of individuals acquiring this type of infection.
Next to models for human pathogens there is also an increasing need for aquatic animal models. However, studies aimed at investigating the pathogenicity mechanisms in aquatic hosts are often confounded by the presence of the natural microbiota (which usually contains Vibrio spp.). Gnotobiotic animals provide researchers with a means to examine host-microbe interactions without interference or influence from unknown microbiota (Gordon and Pesti, 1971). A model based on the use of gnotobiotic 1-day old larvae of brine shrimp (Artemia franciscana) has been recently developed to study V. campbellii, V. harveyi, or V. anguillarum pathogenesis (Defoirdt et al., 2005). An alternative model system for V. anguillarum involves the use of gnotobiotic European sea bass (Dicentrarchus labrax) larvae, where survival is monitored over 1 week (Li et al., 2014a). Finally, specific-pathogen-free (SPF) juveniles of C. gigas (Petton et al., 2013, 2015) have been developed to investigate the diversity and dynamics of microbial populations in an oceanic environment during disease. When combined with methods to monitor gene expression and activity of vibrios during infection (e.g., Ruwandeepika et al., 2011; Defoirdt and Sorgeloos, 2012), a better understanding of the infection process(es) will emerge.
Conclusion
This workshop clearly demonstrated the importance of vibrios to our understanding of emergent diseases in marine and inland aquatic ecosystems as well as their potential impact on society. The rising frequency of disease events not only affects humans directly but also indirectly by reducing food security and ecosystem health. The synergistic investigation of mechanistic and ecological processes contributing to disease is therefore paramount for our understanding of the larger scale consequences of changing Vibrio populations. A better understanding of Vibrio ecology is pivotal for the development of prevention and mitigation strategies. In addition, the mechanistic knowledge of virulence regulatory mechanisms could ultimately be used to inhibit disease. However, these tasks are complicated by the high diversity present within Vibrio populations, and the fact that biotic interactions within and between microbial communities, modify disease expression on different levels. Therefore, we have to consider Vibrio disease as an emergent, multi-faceted phenomenon that will require experimental model systems covering molecules to whole organisms. Expertise for most of these crucial challenges already exists and became united at the workshop under the European umbrella of Vibrio research thereby fostering a more productive combination of basic and applied research in the future (Figure 1).
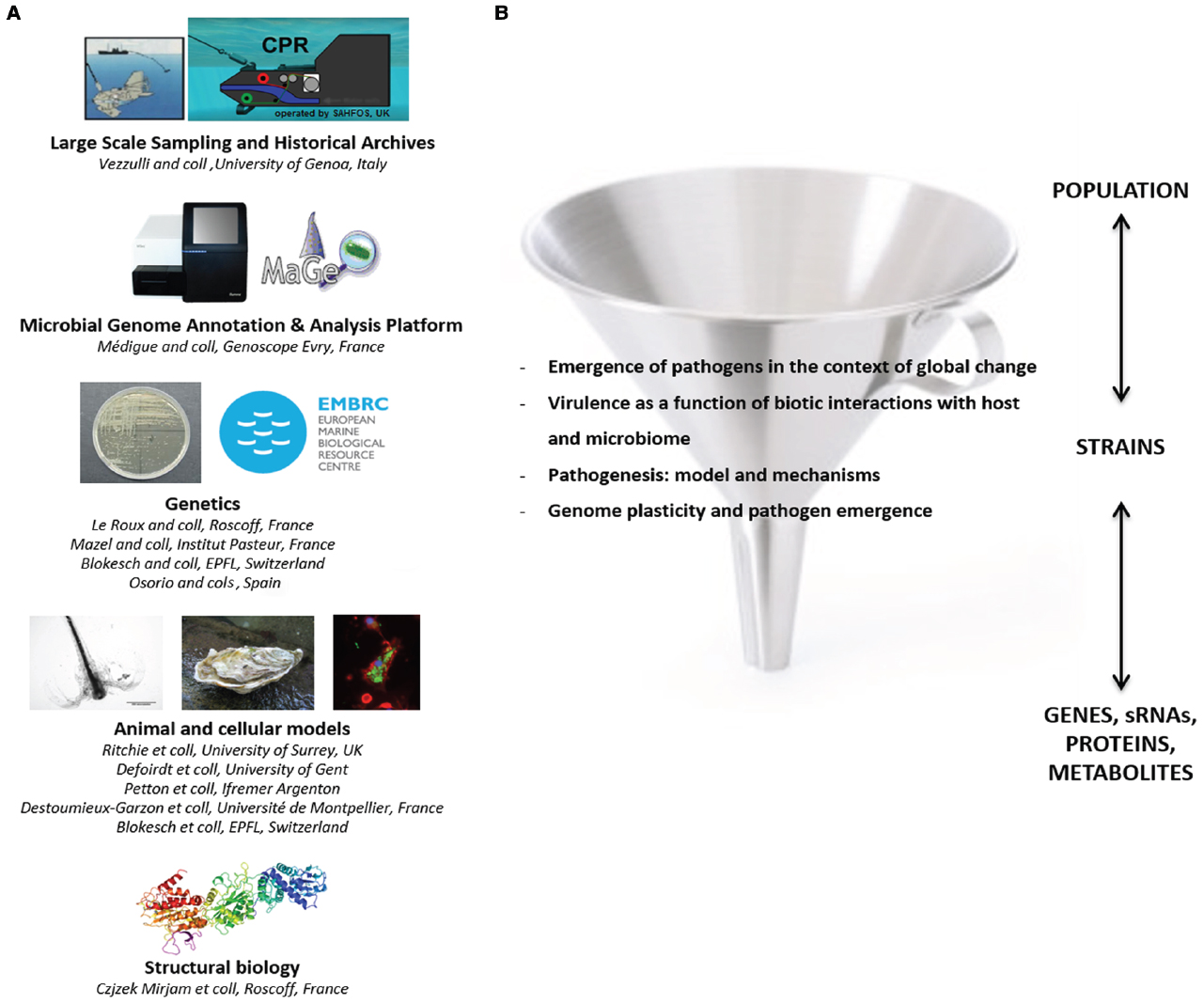
Figure 1. Perspectives of the European Vibrio network. (A) Sharing of common tools and databases. Large-scale sampling for Vibrio collection in the environment and retrospective analysis of Vibrio populations could be improved using the continuous plankton recorder (CPR) technology and the historical CPR archive. An “encyclopedia of Vibrio genome sequences” could be developed by the Genoscope (Evry, France) allowing access for the community to the Microbial genome annotation and analysis platform (MAGE). A genetic resource center, initially created under the scope of EMBRC France, could be improved thanks to other teams performing genetic development. Several teams developing in vivo and in vitro models to investigate host-Vibrio interactions as well as structural biology were also identified. Training sessions (such as summer schools) in the field of bioinformatics or microbial genetics could be organized. (B) Collaborations addressing specific questions have already been stimulated by the workshop.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We warmly thank EUROMARINE, EMBRC-France, the Agence Nationale de la Recherche (ANR) for their financial support and EFOR organization to this first workshop.
References
Alvarez, L., Espaillat, A., Hermoso, J. A., De Pedro, M. A., and Cava, F. (2014). Peptidoglycan remodeling by the coordinated action of multispecific enzymes. Microb. Drug Resist. 20, 190–198. doi: 10.1089/mdr.2014.0047
Amaro, C., Sanjuán, E., Fouz, B., Pajuelo, D., Lee, C., Hor, L., et al. (2015). The Fish Pathogen Vibrio vulnificus Biotype 2: Epidemiology, Phylogeny, and Virulence Factors Involved in Warm-Water Vibriosis. Microbiol Spect. 3.VE-0005-2014. doi: 10.1128/microbiolspec.VE-0005-2014
Andersson, Y., and Ekdahl, K. (2006). Wound infections due to Vibrio cholerae in Sweden after swimming in the Baltic Sea, summer 2006. Euro. Surveill. 11, E060803.2.
Baker-Austin, C., Trinanes, J. A., Taylor, N. G. H., Hartnell, R., Siitonen, A., and Martinez-Urtaza, J. (2013). Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim. Chang. 3, 73–77. doi: 10.1038/nclimate1628
Bisharat, N., Agmon, V., Finkelstein, R., Raz, R., Ben-Dror, G., Lerner, L., et al. (1999). Clinical, epidemiological, and microbiological features of Vibrio vulnificus biogroup 3 causing outbreaks of wound infection and bacteraemia in Israel. Israel Vibrio Study Group. Lancet 354, 1421–1424.
Borgeaud, S., Metzger, L. C., Scrignari, T., and Blokesch, M. (2015). The type VI secretion system of Vibrio cholerae fosters horizontal gene transfer. Science 347, 63–67. doi: 10.1126/science.1260064
Cambray, G., Guerout, A. M., and Mazel, D. (2010). Integrons. Annu. Rev. Genet. 44, 141–166. doi: 10.1146/annurev-genet-102209-163504
Cava, F., De Pedro, M. A., Lam, H., Davis, B. M., and Waldor, M. K. (2011a). Distinct pathways for modification of the bacterial cell wall by non-canonical D-amino acids. EMBO J. 30, 3442–3453. doi: 10.1038/emboj.2011.246
Cava, F., Lam, H., De Pedro, M. A., and Waldor, M. K. (2011b). Emerging knowledge of regulatory roles of D-amino acids in bacteria. Cell. Mol. Life Sci. 68, 817–831. doi: 10.1007/s00018-010-0571-8
Dalsgaard, A., Frimodt-Moller, N., Bruun, B., Hoi, L., and Larsen, J. L. (1996). Clinical manifestations and molecular epidemiology of Vibrio vulnificus infections in Denmark. Eur. J. Clin. Microbiol. Infect. Dis. 15, 227–232.
Defoirdt, T. (2014). Virulence mechanisms of bacterial aquaculture pathogens and antivirulence therapy for aquaculture. Rev. Aquacult. 6, 100–114. doi: 10.1111/raq.12030
Defoirdt, T., Bossier, P., Sorgeloos, P., and Verstraete, W. (2005). The impact of mutations in the quorum sensing systems of Aeromonas hydrophila, Vibrio anguillarum, and Vibrio harveyi on their virulence towards gnotobiotically cultured Artemia franciscana. Environ. Microbiol. 7, 1239–1247. doi: 10.1111/j.1462-2920.2005.00807.x
Defoirdt, T., and Sorgeloos, P. (2012). Monitoring of Vibrio harveyi quorum sensing activity in real time during infection of brine shrimp larvae. ISME J. 6, 2314–2319. doi: 10.1038/ismej.2012.58
Demarre, G., Galli, E., Muresan, L., Paly, E., David, A., Possoz, C., et al. (2014). Differential management of the replication terminus regions of the two Vibrio cholerae chromosomes during cell division. PLoS Genet. 10:e1004557. doi: 10.1371/journal.pgen.1004557
Destoumieux-Garzón, D., Duperthuy, M., Vanhove, A. S., Schmitt, P., and Wai, S. N. (2014). Resistance to Antimicrobial Peptides in vibrios. Antibiotics 3, 23. doi: 10.3390/antibiotics3040540
Dobbs, F. C., Goodrich, A. L., Thomson, F. K. III, and Hynes, W. (2013). Pandemic serotypes of Vibrio cholerae isolated from ship’s ballast tanks and coastal waters: assessment of antibiotic resistance and virulence genes (tcpA and ctxA). Microb. Ecol. 65, 969–974. doi: 10.1007/s00248-013-0182-7
Duperthuy, M., Binesse, J., Le Roux, F., Romestand, B., Caro, A., Got, P., et al. (2010). The major outer membrane protein OmpU of Vibrio splendidus contributes to host antimicrobial peptide resistance and is required for virulence in the oyster Crassostrea gigas. Environ. Microbiol. 12, 951–963. doi: 10.1111/j.1462-2920.2009.02138.x
Duperthuy, M., Schmitt, P., Garzon, E., Caro, A., Rosa, R. D., Le Roux, F., et al. (2011). Use of OmpU porins for attachment and invasion of Crassostrea gigas immune cells by the oyster pathogen Vibrio splendidus. Proc. Natl. Acad. Sci. U.S.A. 108, 2993–2998. doi: 10.1073/pnas.1015326108
Fillion, K., and Mileno, M. D. (2015). Cholera in travelers: shifting tides in epidemiology, management, and prevention. Curr. Infect. Dis. Rep. 17, 455. doi: 10.1007/s11908-014-0455-4
Frans, I., Michiels, C. W., Bossier, P., Willems, K. A., Lievens, B., and Rediers, H. (2011). Vibrio anguillarum as a fish pathogen: virulence factors, diagnosis and prevention. J. Fish Dis. 34, 643–661. doi: 10.1111/j.1365-2761.2011.01279.x
Garcia, K., Torres, R., Uribe, P., Hernandez, C., Rioseco, M. L., Romero, J., et al. (2009). Dynamics of clinical and environmental Vibrio parahaemolyticus strains during seafood-related summer diarrhea outbreaks in southern Chile. Appl. Environ. Microbiol. 75, 7482–7487. doi: 10.1128/AEM.01662-09
Gay, M., Berthe, F. C., and Le Roux, F. (2004). Screening of Vibrio isolates to develop an experimental infection model in the Pacific oyster Crassostrea gigas. Dis. Aquat. Organ 59, 49–56. doi: 10.3354/dao059049
Gordon, H. A., and Pesti, L. (1971). The gnotobiotic animal as a tool in the study of host microbial relationships. Bacteriol. Rev. 35, 390–429.
Goudenège, D., Travers, M. A., Lemire, A., Petton, B., Haffner, P., Labreuche, Y., et al. (2014). A single regulatory gene is sufficient to alter Vibrio aestuarianus pathogenicity in oysters. Environ. Microbiol. doi: 10.1111/1462-2920.12699 [Epub ahead of print].
Huehn, S., Eichhorn, C., Urmersbach, S., Breidenbach, J., Bechlars, S., Bier, N., et al. (2014). Pathogenic vibrios in environmental, seafood and clinical sources in Germany. Int. J. Med. Microbiol. 304, 843–850. doi: 10.1016/j.ijmm.2014.07.010
Huhulescu, S., Indra, A., Feierl, G., Stoeger, A., Ruppitsch, W., Sarkar, B., et al. (2007). Occurrence of Vibrio cholerae serogroups other than O1 and O139 in Austria. Wien Klin Wochenschr 119, 235–241. doi: 10.1007/s00508-006-0747-2
Jones, J. L., Ludeke, C. H., Bowers, J. C., Garrett, N., Fischer, M., Parsons, M. B., et al. (2012). Biochemical, serological, and virulence characterization of clinical and oyster Vibrio parahaemolyticus isolates. J. Clin. Microbiol. 50, 2343–2352. doi: 10.1128/JCM.00196-12
Kirschner, A. K., Schlesinger, J., Farnleitner, A. H., Hornek, R., Suss, B., Golda, B., et al. (2008). Rapid growth of planktonic Vibrio cholerae non-O1/non-O139 strains in a large alkaline lake in Austria: dependence on temperature and dissolved organic carbon quality. Appl. Environ. Microbiol. 74, 2004–2015. doi: 10.1128/AEM.01739-07
Kjaerulff, L., Nielsen, A., Mansson, M., Gram, L., Larsen, T. O., Ingmer, H., et al. (2013). Identification of four new agr quorum sensing-interfering cyclodepsipeptides from a marine Photobacterium. Mar. Drugs 11, 5051–5062. doi: 10.3390/md11125051
Lam, H., Oh, D. C., Cava, F., Takacs, C. N., Clardy, J., De Pedro, M. A., et al. (2009). D-amino acids govern stationary phase cell wall remodeling in bacteria. Science 325, 1552–1555. doi: 10.1126/science.1178123
Lee, C. T., Pajuelo, D., Llorens, A., Chen, Y. H., Leiro, J. M., Padrós, F., et al. (2013). MARTX of Vibrio vulnificus biotype 2 is a virulence and survival factor. Environ. Microbiol. 15, 419–432. doi: 10.1111/j.1462-2920.2012.02854.x
Lemire, A., Goudenege, D., Versigny, T., Petton, B., Calteau, A., Labreuche, Y., et al. (2014). Populations, not clones, are the unit of Vibrio pathogenesis in naturally infected oysters. ISME J. 9, 1523–1531. doi: 10.1038/ismej.2014.233
Lenz, D. H., Miller, M. B., Zhu, J., Kulkarni, R. V., and Bassler, B. L. (2005). CsrA and three redundant small RNAs regulate quorum sensing in Vibrio cholerae. Mol. Microbiol. 58, 1186–1202. doi: 10.1111/j.1365-2958.2005.04902.x
Li, X., Defoirdt, T., Yang, Q., Laureau, S., Bossier, P., and Dierckens, K. (2014a). Host-induced increase in larval sea bass mortality in a gnotobiotic challenge test with Vibrio anguillarum. Dis. Aquat. Organ 108, 211–216. doi: 10.3354/dao02722
Li, X., Yang, Q., Dierckens, K., Milton, D. L., and Defoirdt, T. (2014b). RpoS and indole signaling control the virulence of Vibrio anguillarum towards gnotobiotic sea bass (Dicentrarchus labrax) larvae. PLoS ONE 9:e111801. doi: 10.1371/journal.pone.0111801
Lo Scrudato, M., and Blokesch, M. (2012). The regulatory network of natural competence and transformation of Vibrio cholerae. PLoS Genet. 8:e1002778. doi: 10.1371/journal.pgen.1002778
Lo Scrudato, M., and Blokesch, M. (2013). A transcriptional regulator linking quorum sensing and chitin induction to render Vibrio cholerae naturally transformable. Nucleic Acids Res. 41, 3644–3658. doi: 10.1093/nar/gkt041
Lokmer, A., and Wegner, K. M. (2015). Hemolymph microbiome of Pacific oysters in response to temperature, temperature stress and infection. ISME J. 9, 670–682. doi: 10.1038/ismej.2014.160
Lukinmaa, S., Mattila, K., Lehtinen, V., Hakkinen, M., Koskela, M., and Siitonen, A. (2006). Territorial waters of the Baltic Sea as a source of infections caused by Vibrio cholerae non-O1, non-O139: report of 3 hospitalized cases. Diagn. Microbiol. Infect. Dis. 54, 1–6. doi: 10.1016/j.diagmicrobio.2005.06.020
Mansson, M., Nielsen, A., Kjaerulff, L., Gotfredsen, C. H., Wietz, M., Ingmer, H., et al. (2011). Inhibition of virulence gene expression in Staphylococcus aureus by novel depsipeptides from a marine Photobacterium. Mar. Drugs 9, 2537–2552. doi: 10.3390/md9122537
Martinez-Urtaza, J., Lozano-Leon, A., DePaola, A., Ishibashi, M., Shimada, K., Nishibuchi, M., et al. (2004). Characterization of pathogenic Vibrio parahaemolyticus isolates from clinical sources in Spain and comparison with asian and north American pandemic isolates. J. Clin. Microbiol. 42, 4672–4678. doi: 10.1128/JCM.42.10.4672-4678.2004
Medema, M. H., Blin, K., Cimermancic, P., De Jager, V., Zakrzewski, P., Fischbach, M. A., et al. (2011). antiSMASH: rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 39, W339–W346. doi: 10.1093/nar/gkr466
Meibom, K. L., Blokesch, M., Dolganov, N. A., Wu, C. Y., and Schoolnik, G. K. (2005). Chitin induces natural competence in Vibrio cholerae. Science 310, 1824–1827. doi: 10.1126/science.1120096
Nair, G. B., Ramamurthy, T., Bhattacharya, S. K., Dutta, B., Takeda, Y., and Sack, D. A. (2007). Global dissemination of Vibrio parahaemolyticus serotype O3:K6 and its serovariants. Clin. Microbiol. Rev 20, 39–48. doi: 10.1128/CMR.00025-06
Nguyen, A. N., and Jacq, A. (2014). Small RNAs in the Vibrionaceae: an ocean still to be explored. Wiley Interdiscip Rev. RNA 5, 381–392. doi: 10.1002/wrna.1218
Nielsen, A., Mansson, M., Bojer, M. S., Gram, L., Larsen, T. O., Novick, R. P., et al. (2014). Solonamide B inhibits quorum sensing and reduces Staphylococcus aureus mediated killing of human neutrophils. PLoS ONE 9:e84992. doi: 10.1371/journal.pone.0084992
Oliver, J. (2015). The biology of Vibrio vulnificus. Microbiol. Spect. 3. doi: 10.1128/microbiolspec.VE-0001-2014
Ottaviani, D., Leoni, F., Rocchegiani, E., Santarelli, S., Masini, L., Di Trani, V., et al. (2009). Prevalence and virulence properties of non-O1 non-O139 Vibrio cholerae strains from seafood and clinical samples collected in Italy. Int. J. Food Microbiol. 132, 47–53. doi: 10.1016/j.ijfoodmicro.2009.03.014
Ottaviani, D., Leoni, F., Serra, R., Serracca, L., Decastelli, L., Rocchegiani, E., et al. (2012). Non-toxigenic Vibrio parahaemolyticus strains causing acute gastroenteritis. J. Clin. Microbiol. 50, 4141–4143. doi: 10.1128/JCM.01993-12
Pajuelo, D., Lee, C. T., Roig, F. J., Hor, L. I., and Amaro, C. (2015). Novel host-specific iron acquisition system in the zoonotic pathogen Vibrio vulnificus. Environ. Microbiol. 17, 2076–2089. doi: 10.1111/1462-2920.12782
Pande, G. S., Natrah, F. M., Sorgeloos, P., Bossier, P., and Defoirdt, T. (2013). The Vibrio campbellii quorum sensing signals have a different impact on virulence of the bacterium towards different crustacean hosts. Vet. Microbiol. 167, 540–545. doi: 10.1016/j.vetmic.2013.08.021
Partridge, D. G., Townsend, R., Larkin, S., and Parsons, H. K. (2009). Vibrio vulnificus: an unusual mode of acquisition and novel use of rapid susceptibility testing. J. Clin. Pathol. 62, 370–372. doi: 10.1136/jcp.2008.059238
Petton, B., Marianne, A.-B., Boudry, P., and Pernet, F. (2015). Factors influencing disease-induced mortality of Pacific oysters Crassostrea gigas. Aquacult. Environ. Interact. 6, 205–222. doi: 10.3354/aei00125
Petton, B., Pernet, F., Robert, R., and Boudry, P. (2013). Temperature influence on pathogen transmission and subsequent mortalities in juvenile Pacific oysters Crassostrea gigas. Aquacult. Environ. Interact. 3, 257–273. doi: 10.3354/aei00070
Pujalte, M. J., Sitjà-Bobadilla, A., Macián, M. C., Belloch, C., Alvarez-Pellitero, P., Pérez-Sánchez, J., et al. (2003). Virulence and molecular typing of Vibrio harveyi strains isolated from cultured dentex, gilthead sea bream and European sea bass. Syst. Appl. Microbiol. 26, 284–292. doi: 10.1078/072320203322346146
Quilici, M. L., Robert-Pillot, A., Picart, J., and Fournier, J. M. (2005). Pandemic Vibrio parahaemolyticus O3:K6 spread, France. Emerg. Infect. Dis 11, 1148–1149. doi: 10.3201/eid1107.041008
Rasmussen, T., Jensen, R. B., and Skovgaard, O. (2007). The two chromosomes of Vibrio cholerae are initiated at different time points in the cell cycle. EMBO J. 26, 3124–3131. doi: 10.1038/sj.emboj.7601747
Reilly, G. D., Reilly, C. A., Smith, E. G., and Baker-Austin, C. (2011). Vibrio alginolyticus-associated wound infection acquired in British waters, Guernsey, July 2011. Euro. Surveill. 16, pii 19994.
Reid, P. C., Gorick, G., and Edwards, M. (2011). Climate change and European Marine Ecosystem Research. Plymouth: Sir Alister Hardy Foundation for Ocean Science, 53.
Ritchie, J. M., Rui, H., Bronson, R. T., and Waldor, M. K. (2010). Back to the future: studying cholera pathogenesis using infant rabbits. mBio 1, e00047-10. doi: 10.1128/mBio.00047-10
Ritchie, J. M., Rui, H., Zhou, X., Iida, T., Kodoma, T., Ito, S., et al. (2012). Inflammation and disintegration of intestinal villi in an experimental model for Vibrio parahaemolyticus-induced diarrhea. PLoS Pathog. 8:e1002593. doi: 10.1371/journal.ppat.1002593
Rivas, A. J., Balado, M., Lemos, M. L., and Osorio, C. R. (2011). The Photobacterium damselae subsp. damselae hemolysins damselysin and HlyA are encoded within a new virulence plasmid. Infect. Immun. 79, 4617–4627. doi: 10.1128/IAI.05436-11
Rivas, A. J., Balado, M., Lemos, M. L., and Osorio, C. R. (2013a). Synergistic and additive effects of chromosomal and plasmid-encoded hemolysins contribute to hemolysis and virulence in Photobacterium damselae subsp. damselae. Infect. Immun. 81, 3287–3299. doi: 10.1128/IAI.00155-13
Rivas, A. J., Lemos, M. L., and Osotio, C. R. (2013b). Photobacterium damselae subsp. damselae, a bacterium pathogenic for marine animals and humans. Front. Microbiol. 4:283. doi: 10.3389/fmicb.2013.00283
Rivas, A. J., Labella, A. M., Borrego, J. J., Lemos, M. L., and Osorio, C. R. (2014). Evidence for horizontal gene transfer, gene duplication and genetic variation as driving forces of the diversity of haemolytic phenotypes in Photobacterium damselae subsp. damselae. FEMS Microbiol. Lett. 355, 152–162. doi: 10.1111/1574-6968.12464
Rivas, A. J., Vences, A., Husmann, M., Lemos, M. L., and Osorio, C. R. (2015). Photobacterium damselae subsp. damselae major virulence factors Dly, plasmid-encoded HlyA, and chromosome-encoded HlyA are secreted via the type II secretion system. Infect. Immun. 83, 1246–1256. doi: 10.1128/IAI.02608-14
Ruwandeepika, H. A., Bhowmick, P. P., Karunasagar, I., Bossier, P., and Defoirdt, T. (2011). Quorum sensing regulation of virulence gene expression in Vibrio harveyi in vitro and in vivo during infection of gnotobiotic brine shrimp larvae. Environ. Microbiol. Rep. 3, 597–602. doi: 10.1111/j.1758-2229.2011.00268.x
Schets, C. (2012). “Pathogenic Vibrio spp. in Northern European Waters,” in International Symposium, 31 May—1 June 2012 in Koblenz, ed. B. F. Gewässerkunde (Koblenz: Bundesministerium für Verkehr, Bau und Stadtentwicklung), 18–21.
Seitz, P., and Blokesch, M. (2013a). Cues and regulatory pathways involved in natural competence and transformation in pathogenic and environmental Gram-negative bacteria. FEMS Microbiol. Rev. 37, 336–363. doi: 10.1111/j.1574-6976.2012.00353.x
Seitz, P., and Blokesch, M. (2013b). DNA-uptake machinery of naturally competent Vibrio cholerae. Proc. Natl. Acad. Sci. U.S.A. 110, 17987–17992. doi: 10.1073/pnas.1315647110
Seitz, P., and Blokesch, M. (2014). DNA transport across the outer and inner membranes of naturally transformable Vibrio cholerae is spatially but not temporally coupled. mBio 5, e01409-14. doi: 10.1128/mBio.01409-14
Seitz, P., Pezeshgi Modarres, H., Borgeaud, S., Bulushev, R. D., Steinbock, L. J., Radenovic, A., et al. (2014). ComEA is essential for the transfer of external DNA into the periplasm in naturally transformable Vibrio cholerae cells. PLoS Genet. 10:e1004066. doi: 10.1371/journal.pgen.1004066
Shin, O. S., Tam, V. C., Suzuki, M., Ritchie, J. M., Bronson, R. T., Waldor, M. K., et al. (2011). Type III secretion is essential for the rapidly fatal diarrheal disease caused by non-O1, non-O139 Vibrio cholerae. mBio 2, e00106–e00111. doi: 10.1128/mBio.00106-11
Smith, H. (2000). Questions about the behaviour of bacterial pathogens in vivo. Philos. Trans. R. Soc. Lond. B Biol. Sci. 355, 551–564. doi: 10.1098/rstb.2000.0597
Soler-Bistue, A., Mondotte, J. A., Bland, M. J., Val, M. E., Saleh, M. C., and Mazel, D. (2015). Genomic location of the major ribosomal protein gene locus determines Vibrio cholerae global growth and infectivity. PLoS Genet. 11:e1005156. doi: 10.1371/journal.pgen.1005156
Stypulkowska-Misiurewicz, H., Pancer, K., and Roszkowiak, A. (2006). Two unrelated cases of septicaemia due to Vibrio cholerae non-O1, non-O139 in Poland, July and August 2006. Eur. Surveill. 11, E061130.2.
Toffano-Nioche, C., Nguyen, A. N., Kuchly, C., Ott, A., Gautheret, D., Bouloc, P., et al. (2012). Transcriptomic profiling of the oyster pathogen Vibrio splendidus opens a window on the evolutionary dynamics of the small RNA repertoire in the Vibrio genus. RNA 18, 2201–2219. doi: 10.1261/rna.033324.112
Torres, L., Escobar, S., Lopez, A. I., Marco, M. L., and Pobo, V. (2002). Wound Infection due to Vibrio vulnificus in Spain. Eur. J. Clin. Microbiol. Infect. Dis. 21, 537–538. doi: 10.1007/s10096-002-0767-4
Val, M. E., Skovgaard, O., Ducos-Galand, M., Bland, M. J., and Mazel, D. (2012). Genome engineering in Vibrio cholerae: a feasible approach to address biological issues. PLoS Genet. 8:e1002472. doi: 10.1371/journal.pgen.1002472
Val, M. E., Soler-Bistue, A., Bland, M. J., and Mazel, D. (2014). Management of multipartite genomes: the Vibrio cholerae model. Curr. Opin. Microbiol. 22, 120–126. doi: 10.1016/j.mib.2014.10.003
Vanhove, A. S., Duperthuy, M., Charriere, G. M., Le Roux, F., Goudenege, D., Gourbal, B., et al. (2015). Outer membrane vesicles are vehicles for the delivery of Vibrio tasmaniensis virulence factors to oyster immune cells. Environ. Microbiol. 17, 1152–1165. doi: 10.1111/1462-2920.12535.
Vezzulli, L., Brettar, I., Pezzati, E., Reid, P. C., Colwell, R. R., Höfle, M. G., et al. (2012). Long-term effects of ocean warming on the prokaryotic community: evidence from the vibrios. ISME J. 6, 21–30. doi: 10.1038/ismej.2011.89
Vezzulli, L., Pruzzo, C., Huq, A., and Colwell, R. R. (2010a). Environmental reservoirs of Vibrio cholerae and their role in cholera. Environ. Microbiol. Rep. 2, 27–33. doi: 10.1111/j.1758-2229.2009.00128.x
Vezzulli, L., Previati, C., Pruzzo, C., Marchese, A., Bourne, D. G., Cerrano, C., et al. (2010b). Vibrio infections triggering mass mortality events in a warming Mediterranean Sea. Environ. Microbiol. 12, 2007–2019. doi: 10.1111/j.1462-2920.2010.02209.x
Wendling, C. C., Batista, F. M., and Wegner, K. M. (2014). Persistence, seasonal dynamics and pathogenic potential of Vibrio communities from Pacific oyster hemolymph. PLoS ONE 9:e94256. doi: 10.1371/journal.pone.0094256
Wendling, C. C., and Wegner, K. M. (2015). Adaptation to enemy shifts: rapid resistance evolution to local Vibrio spp. in invasive Pacific oysters. Proc. Biol. Sci. 282, 20142244. doi: 10.1098/rspb.2014.2244
Wietz, M., Mansson, M., Gotfredsen, C. H., Larsen, T. O., and Gram, L. (2010). Antibacterial compounds from marine Vibrionaceae isolated on a global expedition. Mar. Drugs 8, 2946–2960. doi: 10.3390/md8122946
Yamaichi, Y., Bruckner, R., Ringgaard, S., Moll, A., Cameron, D. E., Briegel, A., et al. (2012). A multidomain hub anchors the chromosome segregation and chemotactic machinery to the bacterial pole. Genes Dev. 26, 2348–2360. doi: 10.1101/gad.199869.112
Zhou, X., Gewurz, B. E., Ritchie, J. M., Takasaki, K., Greenfield, H., Kieff, E., et al. (2013). A Vibrio parahaemolyticus T3SS effector mediatates pathogenesis by independently enabling intestinal colonisation and inhibiting TAK1 activation. Cell Rep. 3, 1690–1702. doi: 10.1016/j.celrep.2013.03.039
Keywords: global warming, human health, aquaculture, interactions, animal model, bacterial disease, genome plasticity, european network
Citation: Le Roux F, Wegner KM, Baker-Austin C, Vezzulli L, Osorio CR, Amaro C, Ritchie JM, Defoirdt T, Destoumieux-Garzón D, Blokesch M, Mazel D, Jacq A, Cava F, Gram L, Wendling CC, Strauch E, Kirschner A and Huehn S (2015) The emergence of Vibrio pathogens in Europe: ecology, evolution, and pathogenesis (Paris, 11–12th March 2015). Front. Microbiol. 6:830. doi: 10.3389/fmicb.2015.00830
Received: 15 May 2015; Accepted: 28 July 2015;
Published: 13 August 2015.
Edited by:
Maurizio Labbate, University of Technology, Sydney, AustraliaReviewed by:
Daniela Ceccarelli, University of Maryland, USAYan Boucher, University of Alberta, Canada
Fabiano Thompson, Federal University of Rio de Janeiro, Brazil
Copyright © 2015 Le Roux, Wegner, Baker-Austin, Vezzulli, Osorio, Amaro, Ritchie, Defoirdt, Destoumieux-Garzón, Blokesch, Mazel, Jacq, Cava, Gram, Wendling, Strauch, Kirschner and Huehn. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frédérique Le Roux, Sorbonne Université, UPMC Paris 06, CNRS, UMR 8227, Integrative Biology of Marine Models, Station Biologique de Roscoff, CS 90074, F-29688 Roscoff Cedex, France,Zmxlcm91eEBzYi1yb3Njb2ZmLmZy