 Alexandra Irrgang1
Alexandra Irrgang1 Linda Falgenhauer2
Linda Falgenhauer2 Jennie Fischer1Hiren Ghosh2Elisabet Guiral1,3
Jennie Fischer1Hiren Ghosh2Elisabet Guiral1,3 Beatriz Guerra1,4Silvia Schmoger1Can Imirzalioglu2
Beatriz Guerra1,4Silvia Schmoger1Can Imirzalioglu2 Trinad Chakraborty2
Trinad Chakraborty2 Jens A. Hammerl1*Annemarie Käsbohrer1,5
Jens A. Hammerl1*Annemarie Käsbohrer1,5- 1Department Biological Safety, German Federal Institute for Risk Assessment, Berlin, Germany
- 2Institute of Medical Microbiology, German Center for Infection Research, Partner Site Giessen-Marburg-Langen, Justus Liebig University, Giessen, Germany
- 3Barcelona Institute for Global Health (ISGlobal), Hospital Clínic-Universitat de Barcelona, Barcelona, Spain
- 4European Food Safety Authority, Parma, Italy
- 5Institute of Veterinary Public Health, Department for Farm Animals and Veterinary Public Health, University of Veterinary Medicine, Vienna, Austria
Extended-spectrum beta-lactamases (ESBL) mediating resistance to 3rd generation cephalosporins are a major public health issue. As food may be a vehicle in the spread of ESLB-producing bacteria, a study on the occurrence of cephalosporin-resistantu Escherichia coli in food was initiated. A total of 404 ESBL-producing isolates were obtained from animal-derived food samples (e.g., poultry products, pork, beef and raw milk) between 2011 and 2013. As CTX-M-15 is the most abundant enzyme in ESBL-producing E. coli causing human infections, this study focusses on E. coli isolates from food samples harboring the blaCTX-M-15 gene. The blaCTX-M-15 gene was detected in 5.2% (n = 21) of all isolates. Molecular analyses revealed a phylogenetic group A ST167 clone that was repeatedly isolated from raw milk and beef samples over a period of 6 months. The analyses indicate that spread of CTX-M-15-producing E. coli in German food samples were associated with a multireplicon IncF (FIA FIB FII) plasmid and additional antimicrobial resistance genes such as aac(6)-Ib-cr, blaOXA−1, catB3, different tet-variants as well as a class 1 integron with an aadA5/dfrA17 gene cassette. In addition, four phylogenetic group A ST410 isolates were detected. Three of them carried a chromosomal copy of the blaCTX-M-15 gene and a single isolate with the gene on a 90 kb IncF plasmid. The blaCTX-M-15 gene was always associated with the ISEcp1 element. In conclusion, CTX-M-15-producing E. coli were detected in German food samples. Among isolates of different matrices, two prominent clonal lineages, namely A-ST167 and A-ST410, were identified. These lineages may be important for the foodborne dissemination of CTX-M-15-producing E. coli in Germany. Interestingly, these clonal lineages were reported to be widely distributed and especially prevalent in isolates from humans and livestock. Transmission of CTX-M-15-harboring isolates from food-producing animals to food appears probable, as isolates obtained from livestock and food samples within the same time period exhibit comparable characteristics as compared to isolates detected from human. However, the routes and direction of transmission need further investigation.
Introduction
Resistance to 3rd generation cephalosporins in bacterial pathogens is of great concern in human medicine, since treatment options become increasingly limited in infections caused by multidrug-resistant Enterobacteriaceae. The most common resistance mechanisms in 3rd generation cephalosporin-resistant Enterobacteriaceae is the production of beta-lactamases (ESBL, AmpC and carbapenemases). The emergence and dissemination of ESBL-producing Enterobacteriaceae is mainly driven by horizontal gene transfer, especially conjugation/mobilization, as the enzymes are usually encoded on plasmids (Bonnet, 2004; Carattoli, 2013). Epidemic plasmids, which are detected amongst farm and companion animals, food and humans, belong to the incompatibility groups (Inc.,) F, A/C, N, HI2, I1 and K (EFSA Panel on Biological Hazards, 2011). However, increasing reports of chromosomal localization of antibiotic resistance genes, indicates that spread of the cephalosporin resistance might also be mediated via clonal spread (Hirai et al., 2013; Price et al., 2013; Rodriguez et al., 2014). The ESBL/AmpC genes are sometimes flanked by mobile genetic elements (e.g., transposons, IS elements or class 1 integrons), which are also responsible for successful transmission, and in case of ISEcp1 and ISCR1, also involved in the expression of the genes (Poirel et al., 2008).
ESBL-producing isolates are frequently reported from samples of livestock origin. Spread and persistence has been demonstrated in different studies (Carattoli, 2009; Liebana et al., 2013). Transmission of ESBL/AmpC-producing Escherichia coli from animal to humans is assumed. Contaminated food as a transmission vehicle is often discussed, but direct evidence to support this hypothesis is rare (Leverstein-van Hall et al., 2011). Often, transmission is suggested by indirect evidence through the detection of similar clones, plasmids or sequence types in different populations (EFSA Panel on Biological Hazards, 2011). In Germany, infections with ESBL-producing E. coli in humans are most commonly associated with CTX-M-15 enzymes, followed by CTX-M-1, -14 and -27 (Ewers et al., 2012; Valenza et al., 2014; Falgenhauer et al., 2016a; Pietsch et al., 2017). In contrast, the most common ESBL-type in animals is CTX-M-1, whereas CTX-M-15 is underrepresented in samples from animal livestock in European countries (EFSA Panel on Biological Hazards, 2011; Day et al., 2016). Similar observations were also made in food. Studies from Germany on chicken meat revealed that the most detected ESBL enzymes belonged to the CTX-M-1 type or SHV (Kola et al., 2012; Campos et al., 2014). Neither in these studies nor in a comprehensive study on ESBL in food from the UK, CTX-M-15 enzymes could be detected (Randall et al., 2017). Nevertheless, blaCTX-M-15-encoding E. coli from animal sources in Europe have been described (Lopez-Cerero et al., 2011; Valentin et al., 2014). The risk of contaminated food for the consumers was clearly shown within the German EHEC-outbreak in 2011 caused by the consumption of fenugreek sprouts contaminated with CTX-M-15-producing E. coli O104:H4 clone (Beutin and Martin, 2012; Weiser et al., 2013).
One of the aims of the German national research consortium RESET (2011–2016) was to reveal possible transmission pathways for ESBL/AmpC-producing Enterobacteriaceae. Harmonized protocols were established for the isolation of phenotypically cephalosporin-resistant bacteria from livestock, environment, food, companion animals, and humans to generate a comparable set of data. A previous study on CTX-M-15-producing E. coli of livestock origin (Fischer et al., 2014) found a frequent occurrence of isolates belonging to the clonal complex 10 (CC10), as well as clonal spread of ST410 isolates. Supporting this, a phylogenetic analysis based on whole genome data of CTX-M-15-producing isolates obtained from German livestock, companion animals, humans and environment was carried out, revealing interspecies dissemination of ST410 clones (Falgenhauer et al., 2016a). In the present study, CTX-M-15-producing E. coli isolated from animal food samples of different matrices were taken in the same period (2011–2013) as the livestock samples and were comprehensively characterized.
Materials and Methods
Bacterial Isolates and Cultivation
More than 2,500 food samples of different origins (poultry, cattle, swine, vegetables) and matrices (meat and meat preparations, raw milk, cheese, vegetables) were taken by official food inspectors and investigated by German state laboratories (Saxony, Lower Saxony, Hesse, Bavaria). Food samples from processing plants, retail and raw milk samples were collected at the farm level. From each sample, 25 g were investigated by a non-selective pre-enrichment step for 18–24 h at 37°C in lysogeny broth (LB) following selective cultivation of 10 μl aliquots on MacConkey agar supplemented with 1 mg/L cefotaxime (CTX, Sigma-Aldrich, Munich, Germany) for 18–24 h at 37°C. The identification of E. coli was confirmed by MALDI-TOF (Biotyper, Bruker). From each sample one E. coli isolate phenotypically resistant to CTX was sent to the German National Institute for Risk Assessment (BfR). Positive samples were obtained from all analyzed matrices, even though only raw milk cheese was burdened and only one vegetable sample was tested positive. The ESBL genotype of 437 isolates was verified by PCR and Sanger sequencing as previously described (Rodriguez et al., 2009). Isolates positive for blaCTX-M-15 were included in this study and further characterized. Phylogenetic groups were classified as previously described (Doumith et al., 2012). The antimicrobial resistance pattern was determined by microbroth dilution according to CLSI guidelines (CLSI M07-A9) at the National Reference Laboratory for Antimicrobial Resistance (NRL-AR, BfR). The used antimicrobial panel was in concordance to the decision 2013/652/EU of the European Union commission and was carried out with microtiter plates from TREK Diagnostic Systems (Thermo Fisher Scientific, Schwerte, Germany).
Molecular Typing and Characterization
Molecular characterization was performed using pulsed-field gel-electrophoresis (PFGE) and multi-locus sequence typing (MLST). Phylogenetic relationship of the isolates was determined using XbaI-PFGE analysis according to the PulseNet protocol (https://www.cdc.gov/pulsenet/pathogens/protocols.html). MLST was performed using the Achtman scheme (adk, fumC, gyrB, icd, mdh, purA, recA; http://mlst.warwick.ac.uk/mlst/dbs/Ecoli).
The location of the blaCTX-M-15 gene was determined by S1-nuclease PFGE (1–25 s 17 h, 120°, 6 V/cm) with subsequent southern blot hybridization using a blaCTX-M-15 PCR-probe (Rodriguez et al., 2009). A chromosomal location of the blaCTX-M-15 gene was assumed for isolates in which no positive signal had been detected. Plasmids harboring the blaCTX-M-15 gene were isolated by alkaline lysis and transformed into E. coli DH10B™ competent cells (Invitrogen™, Thermo Fisher Scientific, Schwerte, Germany) (Birnboim and Doly, 1979; Rodriguez et al., 2009). Selection of transformed cells was carried out on LB agar supplemented with 1 mg/L CTX. Transformation of plasmids was confirmed by PCR. Incompatibility groups of the transferred plasmids were determined by PCR using the PBRT kit (Diatheva, Cartoceto PU, Italy). When transformation experiments were inconclusive, incompatibility group of the blaCTX-M-15 harboring plasmid was determined using PFGE/southern blot hybridization with probes specific for IncF and IncI1. The ISEcp1 element was detected by using modified ALA3/ALA4 Primer (5′-TTTGCGCATACAGCGGCACAC-3′/5′-CTATCCGTACAAGGGAG-3′) (Rodriguez et al., 2014).
Next Generation Sequencing (NGS) and in silico Analyses
Additionally, whole genome sequencing of the isolates was performed. Therefore, genomic DNA was isolated from overnight cultures using the PureLink®; Genomic DNA Mini Kit (Thermo Fisher Scientific, Schwerte, Germany). A NexteraXT library was generated and sequenced on a MiSeq benchtop sequencer (Illumina, CA, USA) with 2 × 300 bp paired-end reads. Raw reads were assembled using SPAdes (v 3.5.0) (Bankevich et al., 2012). Whole-genome-based phylogenetic analysis was performed using HarvestSuite (ParSNP) (Treangen et al., 2014).
Resistance genes, virulence genes, serotype and pMLST were predicted using the web-based tools of the Center for Genomic Epidemiology (Zankari et al., 2012; Carattoli et al., 2014; Joensen et al., 2014, 2015).
Accession Numbers
Whole genome sequences of the isolates have been deposited in the European Nucleotide Archive (ENA). Accession numbers of isolates RL16, RL25, RL36, RL40, RL63, RL162, RL195, RL212, RL224, RL230, RL239, RL330, RL331, RL345, RL346, RL364, RL379, RL406-0, RL452,RL464, and RL465 are summarized in the Table S2.
Results and Discussion
Persistence of the ST167 Clone amongst German CTX-M-15 Food Isolates
From 437 E. coli isolates phenotypically resistant to 3rd generation cephalosporins obtained from animal derived food, 404 isolates were confirmed as ESBL/AmpC-producing bacteria. Of these, 21 (5.2%) isolates harbored the blaCTX-M-15 gene. This is in agreement with the observations, that while resistance to 3rd generation cephalosporins in Germany and other European countries is frequently mediated by CTX-M-15 enzymes in isolates from human origin, they are of low prevalence in bacteria from livestock (Pfeiffer et al., 2013; Brolund, 2014; Valentin et al., 2014). A comparable study from the UK even found no CTX-M-15-producing E. coli in food samples from animals and non-animal sources while there was an overall prevalence of ESBL-producing E. coli of 27.5% of the meat samples (Randall et al., 2017)
The main characteristics of the isolates is given in Table 1. There are distinct similarities regarding detected STs, pMLST of IncF plasmids, class 1 integrons or virulence between isolates obtained from food and animal origin. These results suggest a transmission from animal to food (Fischer et al., 2014).
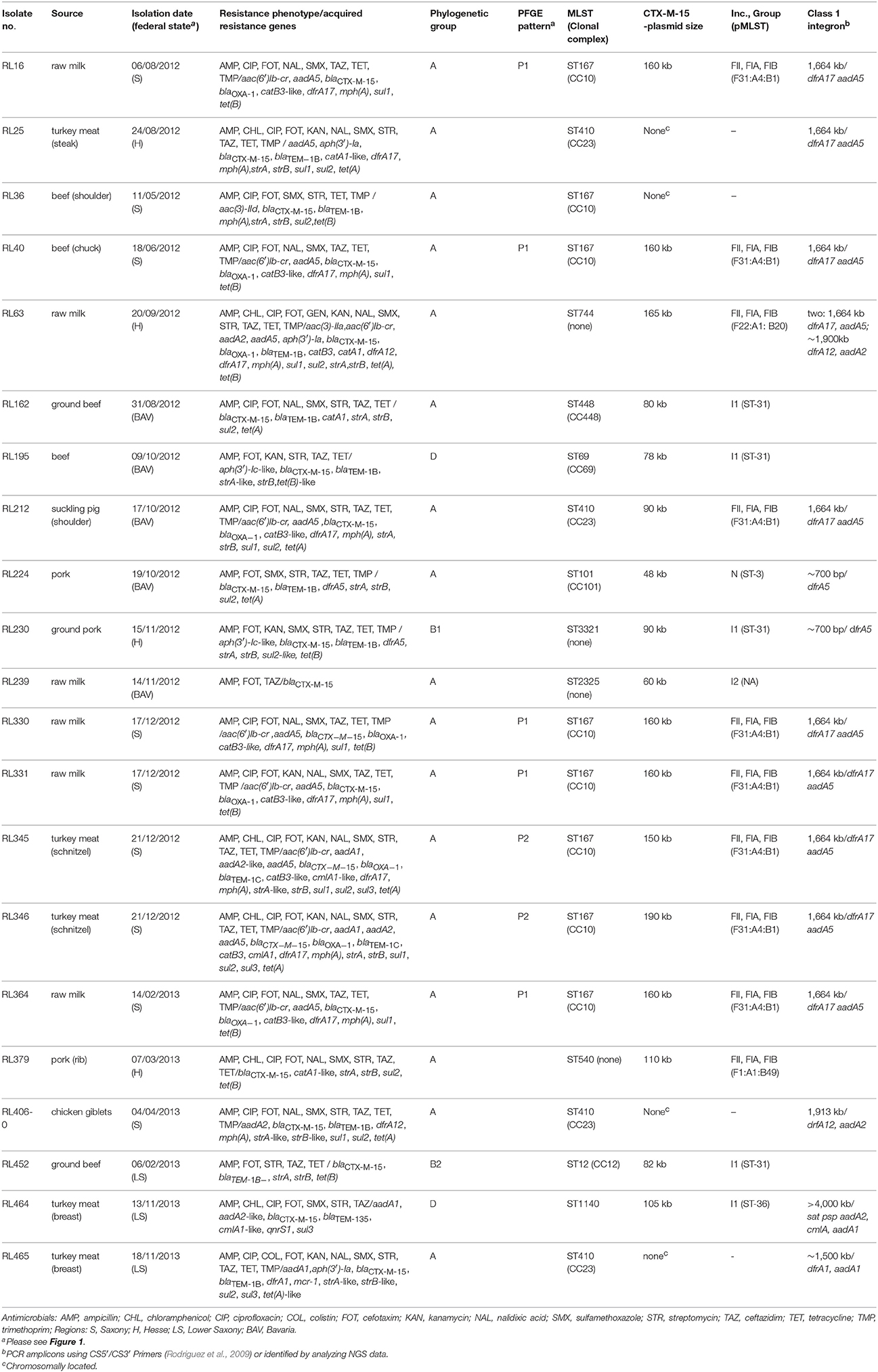
Table 1. Overview of the characteristics of CTX-M-15-producing E. coli isolates obtained from food samples.
There is a predominance of isolates belonging to clonal complex (CC) 10 (n = 8) and CC23 (n = 4) of the phylogenetic group A. Although the sequence types (ST) 38 and ST131 are typically observed in humans (Rodriguez et al., 2014),the frequent presence of ST167/ST617 (CC10) and ST410 (CC23) isolates from food samples in this study concurs with previous reports of isolates from animal samples within the same time period (2011–2013) and from human stool samples. In particular, those reported isolates harbored blaCTX-M-15 and were members of the same phylogenetic group (Fischer et al., 2014; Ben Sallem et al., 2015). This underlines a possible transmission from animals to humans via contaminated food.
All CC10 isolates belong to ST167. Five of them showed an almost identical XbaI PFGE pattern (P1; Figure 1). These strains have been isolated over a period of 6 months from samples of raw milk (n = 4) and beef (n = 1) in Saxony (Eastern Germany). Isolates were obtained from four different samples taken at different time points from three different postal code locations. Milk samples were obtained from farms that were nearby, whereas the beef sample was taken about 200 km away. Therefore, a geographical spread of the clone might have occurred. In general, the phylogenetic group A clonal complex 10 (ST10/167/617) represents a successful clonal lineage, which can be found in humans, livestock, as well as in companion animals (Ewers et al., 2012). In this study two additional ST167 isolates obtained from turkey meat showed similar PFGE restriction patterns (P2; Figure 1) which are distinguishable from those of the raw milk isolates. Nevertheless, all eight ST167 isolates cluster when performing phylogenetic analysis based on whole genome sequences (Figure 2) and all isolates of this clade belong to the same serotype as shown by NGS data (Table S1).
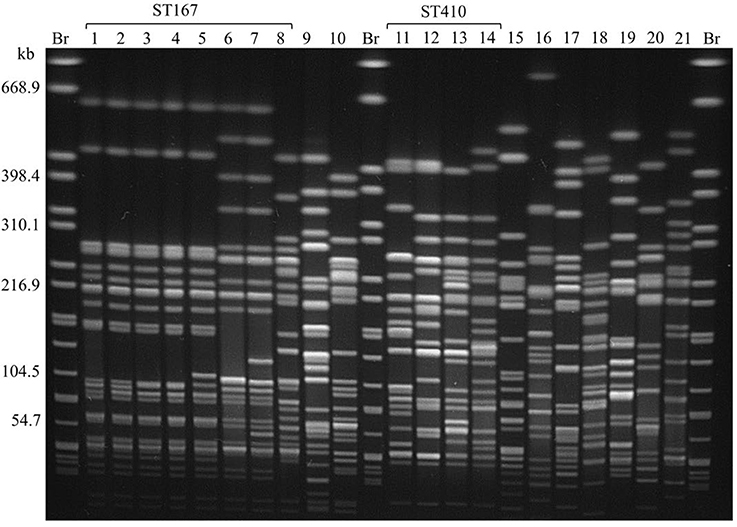
Figure 1. XbaI PFGE of E. coli isolates obtained from food harboring the blaCTX−M−15 gene. Br = S. Braenderup H9812, used as size standard. Lanes:
1: RL16 2: RL40 3: RL330 4: RL331 5: RL364 6: RL345 7: RL346
8: RL36 9: RL162 10: RL63 11: RL25 12: RL212 13: RL406-0 14: RL465
15: RL195 16: RL452 17: RL239 18: RL224 19: RL230 20: RL2379 21: RL464
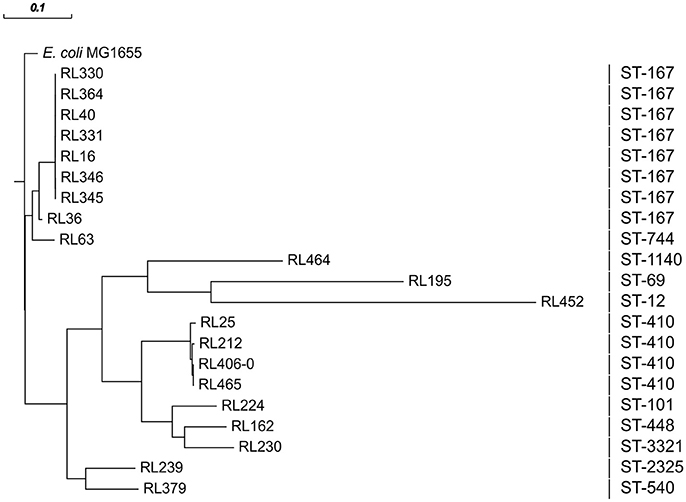
Figure 2. Phylogenetic analysis of all 21 sequenced CTX-M-15-producing isolates obtained from food performed by HarvestSuite (ParSNP), using E. coli MG1655 (NC_000913) as a reference. The scale defines the degree of the phylogenetic relationship of the isolates.
Detection of a Circulating ST410 Clone and the Impact of the Chromosomal Localization of the Beta-Lactamase Gene
There are several isolates (n = 4) belonging to ST410 harboring mainly a chromosomal location of the blaCTX-M-15 gene. Only in one isolate (RL212) the blaCTX-M-15 was located on a 90 kb multireplicon IncF plasmid as described above. However, NGS data revealed a close phylogenetic relationship of all isolates (Figure 2). In the Supplementary Material, a comparison based on whole genome sequences with ST410 isolates from German livestock, companion animals, humans and environment is shown (Supplementary Figure 1). The food-related isolates can be found in three of the five clades (B, C, D), which otherwise comprise of isolates from farm or farm environment-related samples as well as samples of human origin. In concordance with the other strains of clade B, blaCTX-M-15 of RL25 integrated at a distinct location in the rhsE cassette, which is known as a hotspot for insertion sequences and recombination in E. coli (Saier, 2008). The blaCTX-M-15 of RL465 (Falgenhauer et al., 2016b) and RL406-0 also integrated at the same location known for the other members of clade C, at a defective lambdoid prophage region. The results of the current study further extend previous findings of interspecies circulation of ST410 clones to include food and point out the potential risk of contaminated food as transmission vehicles for consumers (Falgenhauer et al., 2016a).
Apart from the three ST410 isolates, a chromosomal localization is also likely for RL63 (ST176). The stable integration of the blaCTX-M-15 genes into the chromosome is reported for different MLST variants (Falgenhauer et al., 2016a). These findings demonstrate that the chromosomal integration of the blaCTX-M-15 gene occurred in several independent events and emphasize that a chromosomal location of this gene might be more common than anticipated (Rodriguez et al., 2014).
blaCTX-M-15 Is Mainly Located on Plasmids of the Incompatibility Group IncF
Apart from strain RL36, where the blaCTX-M-15 seems to be located on the chromosome, the remaining seven ST167 isolates (PFGE pattern P1 and P2) harbor the blaCTX-M-15 gene on 150–190 kb multireplicon IncFIA/FIB/FII plasmids. These large IncF plasmids (>150 kb), as well as the ST410 IncF plasmid, additionally harbored an aac(6)-1b-cr gene (plasmid mediated quinolone resistance gene). The further correlation of blaCTX-M-15-encoding IncF plasmids with the detection of blaOXA-1, catB3 and tet genes is also described by Lopez-Cerero et al. (2011). IncFII plasmids carrying blaCTX-M-15 are known to be highly transferable (Carattoli, 2009). Except for plasmids from RL63 and RL379, all multireplicon IncF plasmids of this study belong to the pMLST F31:A4:B1, indicating plasmid-related spread of blaCTX-M-15 carrying E. coli within different food production chains.
The blaCTX-M-15 - aac(6)-1b-cr- harboring IncF plasmids were also associated with an 1,664 kb large class 1 integron containing a dfrA17/aadA5 gene cassette (Table 1) encoding for trimethoprim and aminoglycoside resistance. Similar class 1 integrons associated with blaCTX-M-15 of phylogenetic group A E. coli were detected in isolates of livestock and companion origin as well as in samples of healthy humans worldwide (Dureja et al., 2014; Fischer et al., 2014).
Association of blaCTX-M-15 with Mobile Genetic Elements
The blaCTX-M-15 gene was associated with an upstream located ISEcp1 element in all isolates, and has been frequently reported for blaCTX-M-15 positive isolates (Lartigue et al., 2004; Smet et al., 2010). The association with ISEcp1 was even detected for chromosomally encoded blaCTX-M-15 genes. This suggests that the resistance gene can be easily mobilized. Transposition of blaCTX−M genes associated with the ISEcp1 element was demonstrated in vitro (Lartigue et al., 2004). The PCR for ISEcp1 was positive in all isolates except for one. For the isolate RL25 (ST410, chromosomal blaCTX-M-15) insertion event of an IS1-element into the tnpA gene (transposase encoding) was detected at identical position to those found in ST410 isolates of different origin in the same clade (Fischer et al., 2014; Falgenhauer et al., 2016a). ISEcp1 elements, which are truncated by different IS elements, are occasionally reported and their effects on mobilization and expression of bla genes as well as their role in plasmid evolution have been discussed (Smet et al., 2010; Alonso et al., 2017).
Virulence Associated Genes amongst CTX-M-15 Isolates
In addition, the occurrence of virulence genes in the isolates was examined (Table S1). Most of the food isolates contained relatively few virulence genes. These included bacteriocins, glutamate decarboxylase, capsule synthesizing enzymes and serum survival genes. These virulence associated genes were also recognized in CTX-M-15-producing isolates from animals (Fischer et al., 2014). One isolate (RL346) harbored senB, which encodes an enterotoxin, that is responsible for enterotoxic activity of enteroinvasive E. coli (EIEC) and Shigella spp. (Nataro et al., 1995). Another isolate (RL452) carried two toxin genes (ncf1–cytotoxic necrotizing factor, involved in urinary tract infection (Mills et al., 2000), and vat–vacuolating autotransporter toxin, known to mediate increased fitness of uropathogenic E. coli (UPEC) during systemic infections (Nichols et al., 2016). This isolate belonged to phylogenetic group B2. These findings support the general assumption of low pathogenic potential in isolates of phylogenetic group A and B1 (major phylogenetic groups detected in this study) as compared to the higher virulence properties in isolates of the phylogenetic group B2 and D.
Conclusion
In conclusion, blaCTX-M-15 positive E. coli have been detected in ESBL-producing isolates obtained from food, albeit with a low prevalence. There are two major findings regarding the spread of these resistance genes in these isolates: (1) the blaCTX-M-15 can either be spread by successful IncF plasmids (pMLST: F31:A4:B1) or (2) it can be transmitted by clonal spread of ST410 isolates harboring a chromosomally encoded gene. This clone was also found in samples of animal and human origin within the same sampling period. Their virtual identity to animal-derived isolates indicates an animal origin of the isolates found in food samples, although cross-contamination cannot be ruled out. Independently, there is a risk for consumers related to exposure to ESBL genes by contaminated food, although a quantification of this issue is not possible. In future, the distribution of CTX-M-types should be closely monitored in a one-health approach, in particular by whole genome analysis of isolates, to detect actual trends and delineate dissemination pathways of the beta-lactamases.
Author Contributions
AK, TC, and BG designed the study. AI, JF, EG, and SS performed the experiments. AI, SS, LF, HG, and CI performed WGS-sequencing and bioinformatics. AI, LF, SS, and JAH analyzed the data. AI, JAH, and LF wrote the manuscript and prepared the tables and figures. All authors edited the manuscript.
Funding
This work was funded by grants from the BMBF, German Federal Ministry for Education and Research in the course of RESET (FKZ01Kl1013B, FKZ01Kl13B, 01KI1313G) and German Center for Infection Research, DZIF/grant number 8000 701-3 [HZI] to TC and CI and TI06.001/8032808811 to TC.
Disclaimer
BG is employed with the European Food Safety Authority (EFSA) in its BIOCONTAM Unit that provides scientific and administrative support to EFSA's scientific activities. The positions and opinions presented in this article are those of the authors alone and are not intended to represent the views or scientific works of EFSA. The other authors have nothing to proclaim.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors gratefully acknowledge the support of the regional laboratories and authorities by collecting the samples and providing the isolates in the framework of the study. We thank Christina Gerstmann, Nathalia Lest, and Mateo San Jose for excellent technical assistance. We thank Dr. Nancy Mah for critically reading the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02318/full#supplementary-material
References
Alonso, C. A., Zarazaga, M., Ben Sallem, R., Jouini, A., Ben Slama, K., and Torres, C. (2017). Antibiotic resistance in Escherichia coli in husbandry animals: the African perspective. Lett. Appl. Microbiol. 64, 318–334. doi: 10.1111/lam.12724
Bankevich, A., Nurk, S., Antipov, D., Gurevich, A. A., Dvorkin, M., Kulikov, A. S., et al. (2012). SPAdes a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477. doi: 10.1089/cmb.2012.0021
Ben Sallem, R., Ben Slama, K., Estepa, V., Cheikhna, E. O., Mohamed, A. M., Chairat, S., et al. (2015). Detection of CTX-M-15-producing Escherichia coli isolates of lineages ST410-A, ST617-A and ST354-D in faecal samples of hospitalized patients in a Mauritanian hospital. J. Chemother. 27, 114–116. doi: 10.1179/1973947814Y.0000000172
Beutin, L., and Martin, A. (2012). Outbreak of Shiga toxin-producing Escherichia coli (STEC) O104H4 infection in Germany causes a paradigm shift with regard to human pathogenicity of STEC strains. J. Food Prot. 75, 408–418. doi: 10.4315/0362-028X.JFP-11-452
Birnboim, H. C., and Doly, J. (1979). A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res. 7, 1513–1523. doi: 10.1093/nar/7.6.1513
Bonnet, R. (2004). Growing group of extended-spectrum-lactamases the CTX-M enzymes. Antimicrob. Agents Chemother. 48, 1–14. doi: 10.1128/AAC.48.1.1-14.2004
Brolund, A. (2014). Overview of ESBL-producing Enterobacteriaceae from a Nordic perspective. Infect. Ecol. Epidemiol. 4:24555. doi: 10.3402/iee.v4.24555
Campos, C. B., Fenner, I., Wiese, N., Lensing, C., Christner, M., Rohde, H., Aepfelbacher, M., et al. (2014). Prevalence and genotypes of extended spectrum beta-lactamases in Enterobacteriaceae isolated from human stool and chicken meat in Hamburg, Germany. Int. J. Med. Microbiol. 304, 678–684. doi: 10.1016/j.ijmm.2014.04.012
Carattoli, A. (2009). Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 53, 2227–2238. doi: 10.1128/AAC.01707-08
Carattoli, A. (2013). Plasmids and the spread of resistance. Int. J. Med. Microbiol. 303, 298–304. doi: 10.1016/j.ijmm.2013.02.001
Carattoli, A., Zankari, E., Garcia-Fernandez, A., Voldby, L. M., Lund, O., Villa, L., et al. (2014). In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 58, 3895–3903. doi: 10.1128/AAC.02412-14
Day, M. J., Rodriguez, I., van Essen-Zandbergen, A., Dierikx, C., Kadlec, K., Schink, A.-K., et al. (2016). Diversity of STs, plasmids and ESBL genes among Escherichia coli from humans, animals and food in Germany, the Netherlands and the UK. J. Antimicrob. Chemother. 71, 1178–1182. doi: 10.1093/jac/dkv485
Doumith, M., Day, M. J., Hope, R., Wain, J., and Woodford, N. (2012). Improved multiplex PCR strategy for rapid assignment of the four major Escherichia coli phylogenetic groups. J. Clin. Microbiol. 50, 3108–3110. doi: 10.1128/JCM.01468-12
Dureja, C., Mahajan, S., and Raychaudhuri, S. (2014). Phylogenetic distribution and prevalence of genes encoding class I Integrons and CTX-M-15 extended-spectrum beta-lactamases in Escherichia coli isolates from healthy humans in Chandigarh, India. PLoS ONE 9:e112551. doi: 10.1371/journal.pone.0112551
EFSA Panel on Biological Hazards (2011). Scientific opinion on the public health risks of bacterial strains producing extended-spectrum β-lactamases and/or AmpC β-lactamases in food and food-producing animals. EFSA J. 9, 2322.
Ewers, C., Bethe, A., Semmler, T., Guenther, S., and Wieler, L. H. (2012). Extended-spectrum beta-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health a global perspective. Clin. Microbiol. Infect. 18, 646–655. doi: 10.1111/j.1469-0691.2012.03850.x
Falgenhauer, L., Imirzalioglu, C., Ghosh, H., Gwozdzinski, K., Schmiedel, J., Gentil, K., et al. (2016a). Circulation of clonal populations of fluoroquinolone-resistant CTX-M-15-producing Escherichia coli ST410 in humans and animals in Germany. Int. J. Antimicrob. Agents 47, 457–465. doi: 10.1016/j.ijantimicag.2016.03.019
Falgenhauer, L., Waezsada, S.-E., Gwozdzinski, K., Ghosh, H., Doijad, S., Bunk, B., et al. (2016b). Chromosomal locations of mcr-1 and blaCTX-M-15 in fluoroquinolone-resistant Escherichia coli ST410. Emerg. Infect. Dis. 22, 1689–1691. doi: 10.3201/eid2209.160692
Fischer, J., Rodríguez, I., Baumann, B., Guiral, E., Beutin, L., Schroeter, A., et al. (2014). blaCTX-M-15-carrying Escherichia coli and Salmonella isolates from livestock and food in Germany. J. Antimicrob. Chemother. 69, 2951–2958. doi: 10.1093/jac/dku270
Hirai, I., Fukui, N., Taguchi, M., Yamauchi, K., Nakamura, T., Okano, S., et al. (2013). Detection of chromosomal blaCTX-M-15in Escherichia coli O25b-B2-ST131 isolates from the Kinki region of Japan. Int. J. Antimicrob. Agents 42, 500–506. doi: 10.1016/j.ijantimicag.2013.08.005
Joensen, K. G., Scheutz, F., Lund, O., Hasman, H., Kaas, R. S., Nielsen, E. M., et al. (2014). Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Microbiol. 52, 1501–1510. doi: 10.1128/JCM.03617-13
Joensen, K. G., Tetzschner, A. M. M., Iguchi, A., Aarestrup, F. M., and Scheutz, F. (2015). Rapid and easy in silico serotyping of Escherichia coli isolates by use of whole-genome sequencing data. J. Clin. Microbiol. 53, 2410–2426. doi: 10.1128/JCM.00008-15
Kola, A., Kohler, C., Pfeifer, Y., Schwab, F., Kühn, K., Schulz, K., et al. (2012). High prevalence of extended-spectrum-β-lactamase-producing Enterobacteriaceae in organic and conventional retail chicken meat, Germany. J. Antimicrob. Chemother. 67, 2631–2634. doi: 10.1093/jac/dks295
Lartigue, M.-F., Poirel, L., and Nordmann, P. (2004). Diversity of genetic environment of blaCTX-M genes. FEMS Microbiol. Lett. 234, 201–207. doi: 10.1111/j.1574-6968.2004.tb09534.x
Leverstein-van Hall, M. A., Dierikx, C. M., Cohen Stuart, J., Voets, G. M., van den Munckhof, M., P van Essen-Zandbergen, A., et al. (2011). Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin. Microbiol. Infect. 17, 873–880. doi: 10.1111/j.1469-0691.2011.03497.x
Liebana, E., Carattoli, A., Coque, T. M., Hasman, H., Magiorakos, A.-P., Mevius, D., et al. (2013). Public health risks of enterobacterial isolates producing extended-spectrum beta-lactamases or AmpC beta-lactamases in food and food-producing animals an EU perspective of epidemiology, analytical methods, risk factors, and control options. Clin. Infect. Dis. 56, 1030–1037. doi: 10.1093/cid/cis1043
Lopez-Cerero, L., Egea, P., Serrano, L., Navarro, D., Mora, A., Blanco, J., et al. (2011). Characterisation of clinical and food animal Escherichia coli isolates producing CTX-M-15 extended-spectrum beta-lactamase belonging to ST410 phylogroup A. Int. J. Antimicrob. Agents 37, 365–367. doi: 10.1016/j.ijantimicag.2011.01.001
Mills, M., Meysick, K. C., and O'Brien, A. D. (2000). Cytotoxic necrotizing factor type 1 of uropathogenic Escherichia coli kills cultured human uroepithelial 5637 cells by an apoptotic mechanism. Infect. Immun. 68, 5869–5880. doi: 10.1128/IAI.68.10.5869-5880.2000
Nataro, J. P., Seriwatana, J., Fasano, A., Maneval, D. R., Guers, L. D., Noriega, F., et al. (1995). Identification and cloning of a novel plasmid-encoded enterotoxin of enteroinvasive Escherichia coli and Shigella strains. Infect. Immun. 63, 4721–4728.
Nichols, K. B., Totsika, M., Moriel, D. G., Lo, A. W., Yang, J., Wurpel, D. J., et al. (2016). Molecular characterization of the vacuolating autotransporter toxin in uropathogenic Escherichia coli. J. Bacteriol. 198, 1487–1498. doi: 10.1128/JB.00791-15
Pfeiffer, Y., Eller, C., Leistner, R., Valenza, G., Nickel, S., Guerra, B., et al. (2013). ESBL producer as human pathogens and the zoonotic reservoir. Hyg. Med. 38, 294–299. Available online at: http://edoc.rki.de/oa/articles/reU6BzKbhvAcI/PDF/29PNIiHL2fRac.pdf
Pietsch, M., Eller, C., Wendt, C., Holfelder, M., Falgenhauer, L., Fruth, A., et al. (2017). Molecular characterisation of extended-spectrum beta-lactamase (ESBL)-producingEscherichia coli isolates from hospital and ambulatory patients in Germany. Vet. Microbiol. 200, 130–137. doi: 10.1016/j.vetmic.2015.11.028
Poirel, L., Naas, T., and Nordmann, P. (2008). Genetic support of extended-spectrum beta-lactamases. Clin. Microbiol. Infect. 14(Suppl. 1), 75–81. doi: 10.1111/j.1469-0691.2007.01865.x
Price, L. B., Johnson, J. R., Aziz, M., Clabots, C., Johnston, B., Tchesnokova, V., et al. (2013). The epidemic of extended-spectrum-beta-lactamase-producing Escherichia coli ST131 is driven by a single highly pathogenic subclone, H30-Rx. mBio 4:13. doi: 10.1128/mBio.00377-13
Randall, L. P., Lodge, M. P., Elviss, N. C., Lemma, F. L., Hopkins, K. L., Teale, C. J., et al. (2017). Evaluation of meat, fruit and vegetables from retail stores in five United Kingdom regions as sources of extended-spectrum beta-lactamase (ESBL)-producing and carbapenem-resistant Escherichia coli. Int. J. Food Microbiol. 241, 283–290. doi: 10.1016/j.ijfoodmicro.2016.10.036
Rodriguez, I., Barownick, W., Helmuth, R., Mendoza, M. C., Rodicio, M. R., Schroeter, A., et al. (2009). Extended-spectrum beta-lactamases and AmpC beta-lactamases in ceftiofur-resistant Salmonella enterica isolates from food and livestock obtained in Germany during 2003-07. J. Antimicrob. Chemother. 64, 301–309. doi: 10.1093/jac/dkp195
Rodriguez, I., Thomas, K., van Essen, A., Schink, A.-,K Day, M., Chattaway, M., et al. (2014). Chromosomal location of blaCTX-M genes in clinical isolates of Escherichia coli from Germany, the Netherlands and the UK. Int. J. Antimicrob. Agents 43, 553–557. doi: 10.1016/j.ijantimicag.2014.02.019
Saier, M. H. Jr. (2008). The bacterial chromosome. Crit. Rev. Biochem. Mol. Biol. 43, 89–134. doi: 10.1080/10409230801921262
Smet, A., van Nieuwerburgh, F., Vandekerckhove, T. T. M., Martel, A., Deforce, D., Butaye, P., et al. (2010). Complete nucleotide sequence of CTX-M-15-plasmids from clinical Escherichia coli isolates insertional events of transposons and insertion sequences. PLoS ONE 5:e11202. doi: 10.1371/journal.pone.0011202
Treangen, T. J., Ondov, B. D., Koren, S., and Phillippy, A. M. (2014). The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 15:571. doi: 10.1186/s13059-014-0524-x
Valentin, L., Sharp, H., Hille, K., Seibt, U., Fischer, J., Pfeifer, Y., et al. (2014). Subgrouping of ESBL-producing Escherichia coli from animal and human sources an approach to quantify the distribution of ESBL types between different reservoirs. Int. J. Med. Microbiol. 304, 805–816. doi: 10.1016/j.ijmm.2014.07.015
Valenza, G., Nickel, S., Pfeifer, Y., Eller, C., Krupa, E., Lehner-Reindl, V., et al. (2014). Extended-spectrum-beta-lactamase-producing Escherichia coli as intestinal colonizers in the German community. Antimicrob. Agents Chemother. 58, 1228–1230. doi: 10.1128/AAC.01993-13
Weiser, A. A., Gross, S., Schielke, A., Wigger, J.-F., Ernert, A., Adolphs, J., et al. (2013). Trace-back and trace-forward tools developed ad hoc and used during the STEC O104H4 outbreak 2011 in Germany and generic concepts for future outbreak situations. Foodborne Pathog. Dis. 10, 263–269. doi: 10.1089/fpd.2012.1296
Keywords: antimicrobial resistance, CTX-M-15, livestock, genome, plasmid, distribution, ESBL
Citation: Irrgang A, Falgenhauer L, Fischer J, Ghosh H, Guiral E, Guerra B, Schmoger S, Imirzalioglu C, Chakraborty T, Hammerl JA and Käsbohrer A (2017) CTX-M-15-Producing E. coli Isolates from Food Products in Germany Are Mainly Associated with an IncF-Type Plasmid and Belong to Two Predominant Clonal E. coli Lineages. Front. Microbiol. 8:2318. doi: 10.3389/fmicb.2017.02318
Received: 03 June 2017; Accepted: 09 November 2017;
Published: 21 November 2017.
Edited by:
Alberto Quesada, Universidad de Extremadura, SpainReviewed by:
Campos Jorge Campos, ESTM-IPLeiria, PortugalLuisa Peixe, Universidade do Porto, Portugal
Copyright © 2017 Irrgang, Falgenhauer, Fischer, Ghosh, Guiral, Guerra, Schmoger, Imirzalioglu, Chakraborty, Hammerl and Käsbohrer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jens A. Hammerl, amVucy1hbmRyZS5oYW1tZXJsQGJmci5idW5kLmRl