 Zhiyang Liu
Zhiyang Liu Xuejuan Li
Xuejuan Li Feng Sun
Feng Sun Tong Zhou
Tong Zhou Yijun Zhou
Yijun Zhou- 1Institute of Plant Protection, Jiangsu Academy of Agricultural Sciences, Jiangsu Technical Service Center of Diagnosis and Detection for Plant Virus Diseases, Nanjing, China
- 2Scientific Observation and Experimental Station of Crop Pests in Nanjing, Ministry of Agriculture, Nanjing, China
Rice stripe virus (RSV) causes a severe disease in Oryza sativa (rice) in many Eastern Asian countries. The NS3 protein of RSV is a viral suppressor of RNA silencing, but plant host factors interacting with NS3 have not been reported yet. Here, we present evidence that expression of RSV NS3 in Arabidopsis thaliana causes developmental abnormalities. Through yeast two-hybrid screening and a luciferase complementation imaging assay, we demonstrate that RSV NS3 interacted with OsCIPK30, a CBL (calcineurin B-like proteins)-interaction protein kinase protein. Furthermore, OsCIPK30 was overexpressed to investigate the function of OsCIPK30 in rice. Our investigation showed that overexpression of OsCIPK30 in rice could delay the RSV symptoms and show milder RSV symptoms. In addition, the expression of pathogenesis-related genes was increased in OsCIPK30 transgenic rice. These results suggest that overexpression of OsCIPK30 positively regulates pathogenesis-related genes to enhance the tolerance to RSV in rice. Our findings provide new insight into the molecular mechanism underlying resistance to RSV disease.
Introduction
Rice stripe disease, caused by Rice stripe virus (RSV), is one of the most destructive diseases in rice (Wei et al., 2009). RSV is transmitted transovarially in a circulative manner by vector insects, primarily the small brown planthopper (SBPH; Laodelphax striatellus), exhibiting chlorotic and necrotic stripes in rice plant leaves followed by plant wilting (Hibino, 1996; Falk and Tsai, 1998). RSV was first reported in Japan in 1897 (Kisimoto, 1965) and is currently present in many East Asian countries, including China, where it was first reported in 1963. It is now widely spread in at least 16 different provinces of China (Wei et al., 2009).
Rice stripe virus belongs to the Tenuivirus genus, and the viral genome consists of four single-stranded RNA segments (Hibino, 1996; Falk and Tsai, 1998), which encode seven ORFs using a negative or ambisense coding strategy (Ramirez and Haenni, 1994). RNA1 is the largest RNA segment and has a single ORF in the viral-complementary sense, encoding an RNA-dependent RNA polymerase (RdRp) (Barbier et al., 1992; Toriyama et al., 1994). Each of the other three segments (RNA2, 3, and 4) has two have two open reading frames (ORFs), with one on the viral RNA (vRNA) and the other on the viral complementary strand (vcRNA) (Zhu et al., 1991, 1992; Takahashi et al., 1993). RSV vRNA2 encodes a membrane-associated protein (also named NS2) that has been reported to be an RNA silencing suppressor (Du et al., 2011). The vcRNA2 encodes a glycoprotein (also named NSvc2), which moves from the endoplasmic reticulum to the Golgi bodies via the coat protein complexes I – and coat protein complexes II-dependent secretion pathway (Zhao et al., 2012; Yao et al., 2014). vRNA3 encodes an RNA silencing suppressor (also named NS3), suppressing both local and systemic RNA silencing in the plant and vcRNA3 encodes a nucleocapsid protein (also named NP) from the vcRNA (Xiong et al., 2009; Shen et al., 2010). RSV vRNA4 encodes a disease-specific protein (also named SP) that accumulates in both infected plant and insect cells (Toriyama, 1986). The protein encoded by vcRNA4 (also named MP) has been identified as a cell-to-cell movement protein (Xiong et al., 2008).
Thus far, few host proteins have been reported to interact with RSV proteins. For example, DnaJ protein and a small heat-shock protein (HSP) of rice can interact with RSV MP, OsSGS3 (suppressor of gene silencing 3) can interact with NS2, and PsbP, a 23-kDa oxygen-evolving complex protein of plants, has been reported to interact with SP, resulting in the enhancement of virus symptoms (Lu et al., 2009; Du et al., 2011; Kong et al., 2014). Recently, host HSP70 and HSP20 have been shown to interact with RSV RdRp, the former of which is necessary for RSV infection and the latter of which can be altered by RSV for its expression pattern (Jiang et al., 2014; Li et al., 2015). Although NS3 was reported as a suppressor of RNA silencing, the host factors that may interact with NS3 are still unknown.
Plant CIPKs can be activated by interacting with calcineurin B-like proteins (CBLs) to transduce calcium signals by phosphorylating downstream signaling components, containing a Ser/Thr protein kinase domain in their N terminal region and an NAF domain [a novel protein interaction domain (first called by Albrecht et al., 2001)] in their C-terminal region (Luan, 2009). A systematic genome analysis revealed the existence of over 30 OsCIPK genes in rice (Kolukisaoglu et al., 2004; Xiang et al., 2007). Thus far, several studies have postulated the possible functions of OsCIPKs. OsCIPK19 is involved in the responses to light, nutrients, and low temperature (Ohba et al., 2000). OsCIPK23 is suggested to function in pollination and drought stress responses (Yang et al., 2008). The expression of OsCIPK31 is involved in diverse signals, such as cold, salt, light, cytokinins, and sugars (Kim et al., 2003). The mutant of OsCIPK31 exhibited hypersensitive phenotypes to abscisic acid, salt, mannitol, and glucose (Piao et al., 2010). Overexpression of OsCIPK3, OsCIPK12, and OsCIPK15 enhances tolerance to cold, drought, and salt stress, respectively (Xiang et al., 2007). OsCIPK15 has also been shown to play an important role in O2-deficiency tolerance in rice (Lee et al., 2009). To date, many OsCIPKs genes have been reported to be involved in response to abiotic stresses, but only few have been reported to be involved in responses to biotic stresses.
Here, we provide evidence showing that host OsCIPK30 interacts with RSV NS3, and that the overexpression of OsCIPK30 in rice (Oryza sativa L. japonica. cv. Nipponbare) positively regulates pathogenesis-related genes to enhance tolerance to RSV.
Materials and Methods
Sources of Virus, Vectors, and Plant Materials
Rice plants infected with RSV were collected from Jiangsu province in China. Young instar nymphs of SBPHs were fed on the RSV-infected rice plants for 2 days to acquire the virus and were maintained on “Wuyujing No. 3” rice plants grown in an insect-rearing room at a temperature of 25 ± 3°C, 55 ± 5% relative humidity and under a light intensity of 200 μmol m-2 s-1 (16 h photoperiod). The proportions of viruliferous SBPHs were confirmed by dot-ELISA (Sun et al., 2011).
Arabidopsis thaliana (ecotype Columbia-0, Col-0) seeds were grown in potting soil in a growth chamber at 24°C under 200 μmol m-2 s-1 illumination. Rice plants used in this study were grown inside a growth chamber at 26°C, 80% relative humidity. Additionally, all plants were kept under 16-hlight/8-h dark photoperiod conditions.
Inoculation of RSV
Rice plants (Oryza sativa L. spp. japonica. cv. Nipponbare) were inoculated with three viruliferous SBPHs per plant and were kept in a growth chamber. After inoculation for 3 days, planthoppers were removed back to rice field. And the inoculation does not hurt SBPHs at all, we just let them feed on the tested rice plants. Plants were maintained in a growth chamber for symptom development, and RSV-free SBPHs were used for mock inoculation.
Generation of Transgenic Plants
Total RNA was extracted from RSV infected rice. cDNA synthesis and PCR were conducted as described previously (Sun et al., 2016) to get the full length of NS3. The full-length NS3 DNA was integrated into the pENTR/D-TOPO vector (Invitrogen, Carlsbad, CA, United States) and was then cloned into the pBA-Flag-6Myc-DC binary vector (Zhang et al., 2005) using the LR clonase reaction mixture to induce constitutive expression of the NS3 gene in Arabidopsis. The resultant 35S:NS3 construct was introduced into Agrobacterium tumefaciens strain GV3101. Agrobacteria-mediated transformation of A. thaliana ecotype Col-0 with the NS3 gene was conducted using the floral dip method (Clough and Bent, 1998; Zhang et al., 2006). The transgenic seeds were selected on standard MS medium (Murashige and Skoog, 1962) containing the appropriate antibiotics: 100 mg/L carbenicillin (Sangon Biotech, Shanghai, China).
Total RNA was extracted from Oryza sativa L. spp. japonica. cv. Nipponbare and the full-length OsCIPK30 cDNA was cloned into the pUN1301 vector (Biovector, Co., Ltd, Beijing, China) to generate the Ubi-OsCIPK30 plasmid. The construct was introduced into A. tumefaciens strain GV3101. Agrobacteria-mediated transformation protocols for rice were published previously (Wu et al., 2007). Regenerated transgenic plants (T0) were transplanted into soil; and the T1 progeny germinated from seeds were grown in soil inside a greenhouse. The T1 generations were confirmed by RT-PCR for testing hygromycin gene. Empty pUN1301 vector-transformed plants were used as the wild type (WT) controls.
Alignment and Phylogenetic Analysis
Sequence alignment of 33 members of the OsCIPK family in rice was performed using ClustalW. Phylogenetic analysis of the 33 OsCIPKs in rice based on amino acid sequences carried out using a neighbor-joining (NJ) method with MEGA version 4.0.
Yeast Two-Hybrid Assay
To construct the bait vector for the assay, the coding sequence of NS3 was cloned into the yeast GAL4 binding domain vector pGBKT7, as instructed (Clontech, Mountain View, CA, United States). Construction and screening of a rice cDNA library (Oryza sativa L. spp. japonica. cv. Nipponbare) were performed as described in the BD Matchmaker Library Construction and Screening Kits User Manual (Clontech, Mountain View, CA, United States). Briefly, the rice cDNA library was screened using pGBKT7-NS3 as the bait in the Saccharomyces cerevisiae strain Gold (Clontech, Mountain View, CA, United States). The positive clones were selected on a histidine-deficient medium and then confirmed by β-gal assays. The coding sequence of OsCIPK5 (Os01g0206700), OsCIPK25 (Os06g0543400), OsCIPK26 (Os02g0161000), OsCIPK29 (Os07g0678300), OsCIPK30 (Os01g0759200), and OsCIPK32 (Os12g0132200) was PCR-amplified from rice leaf cDNA and then inserted into the yeast GAL4 activation domain vector, pGADT7 (Clontech, Mountain View, CA, United States).
Combinations of the plasmids BD-NS3 and AD-OsCIPK5, BD-NS3 and AD-OsCIPK25, BD-NS3 and AD-OsCIPK26, BD-NS3 and AD-OsCIPK29, BD-NS3 and AD-OsCIPK30, or BD-NS3 and AD-OsCIPK32 were co-transformed into the S. cerevisiae strain Gold. BD-53 and AD-T were also co-transformed into S. cerevisiae strain Gold to serve as a positive control. S. cerevisiae co-transformed with BD-Lam and AD-T, BD-NS3 and AD, or BD and AD-OsCIPK30 were as used as negative controls. All transformants were grown at 30°C for 72 h on synthetic medium lacking Leu and Trp and then transferred to medium lacking adenine, His, Leu, and Trp and supplemented with 0.5 mg ml-1 Aureobasidin A (AbA).
Luciferase Complementation Imaging (LCI) Assay
The LCI was performed on 4-week-old Nicotiana benthamiana (N. benthamiana) leaves infiltrated with various combinations of A. tumefaciens GV3101 harboring pCambia-Myc-NS3-nLUC and pCambia-cLUC-3HA-candidate proteins. The agrobacteria containing the plasmids were incubated with 25 μM beta-estradiol for 3 h prior to infiltration, and all cultures were adjusted to OD600 = 0.8. The transfected leaves were assayed 2 days after agroinfiltration by adding the substrate (10 mM luciferin). The sprayed leaves were incubated in total darkness for 5 min and photographed using the Tanon 5200 Luminescent Imaging Workstation with an EMCCD camera. The images were processed with TanonImage software (Tanon Science & Technology, Shanghai, China).
Western Blot Assay
Extraction of total protein from leaf tissues (100 mg per sample) for western blotting was done as described previously (Sun et al., 2016). The membranes were probed with anti-Myc antibodies (Sigma-Aldrich, St. Louis, MO, United States). The secondary antibody used in the study was a goat anti-rabbit IgG conjugated with alkaline phosphatase (Sigma-Aldrich, St. Louis, MO, United States).
Real-Time Quantitative RT-PCR (qRT-PCR)
Equal amounts of the RNA samples from five independent plants of the same treatment were pooled. Total RNA was extracted from each treatment using RNAiso Plus reagent (Takara, Dalian, China), and cDNA synthesis and PCR were conducted as described previously (Sun et al., 2016). Actin1 (Os03g0718100) was used as an internal standard. qRT-PCR was performed using the Bio-Rad iQ5 Real-time PCR System and SYBR Premix Ex Taq and the primers used in this assay are listed in Supplementary Data Sheet 1 (Takara, Dalian, China). The comparative cycle threshold method was used to calculate the relative mRNA levels (Applied Biosystems, 1997; Johnson et al., 2000). All qRT-PCR experiments were performed at least three times.
Statistical Analysis
Data were obtained from three replicates and analyzed by unpaired two-tailed Student’s t-test with GraphPad Prism software (GraphPad Software, Inc., La Jolla, CA, United States). Values of P < 0.05 or P < 0.01 were considered to indicate different levels of statistical significance.
Results
RSV NS3 Causes Developmental Abnormalities in Arabidopsis
To study the function of NS3, we generated transgenic Arabidopsis lines overexpressing full-length RSV NS3 with Flag-6Myc epitopes. These transgenic plants were confirmed by western blot assays (Figure 1B). Importantly, the majority of the transgenic lines exhibited developmental abnormalities with serrate true leaves (Figure 1A). The phenotypes of transgenic plants indicated that NS3 might impact growth and development in Arabidopsis and might also influence rice.

FIGURE 1. NS3 causes developmental abnormalities in Arabidopsis. (A) Morphological defects of Arabidopsis transgenic plants expressing RSV NS3. Photographs were taken 30 days after sowing. (B) Western blot analysis of 35S-Flag-6Myc-NS3 transgenic lines. The hyphen indicates the locations of the tagged NS3 proteins (23.87 KDa NS3 plus 7.2 KDa 6Myc). +: 35S-Flag-6Myc-SERRATE; –: Col-0; 1–8: individual NS3 transgenic lines.
RSV NS3 Interacts with OsCIPK30
To identify plant host factor(s) that interact with NS3 during RSV infection, we screened a rice cDNA library using RSV NS3 as the bait through a yeast two-hybrid assay. Twenty-one positive cDNA clones were identified. One of them was a fragment of the OsCIPK30 gene. To test the interaction between the full-length OsCIPK30 and NS3, the ORFs of OsCIPK30 and NS3 were amplified and integrated into pGADT7 and pGBKT7, respectively. The recombinant plasmid (AD-30) was co-transformed into yeast cells with BD-NS3. Figures 2A,B show the positive interaction between the full-length OsCIPK30 and NS3 through yeast two-hybrid assays.
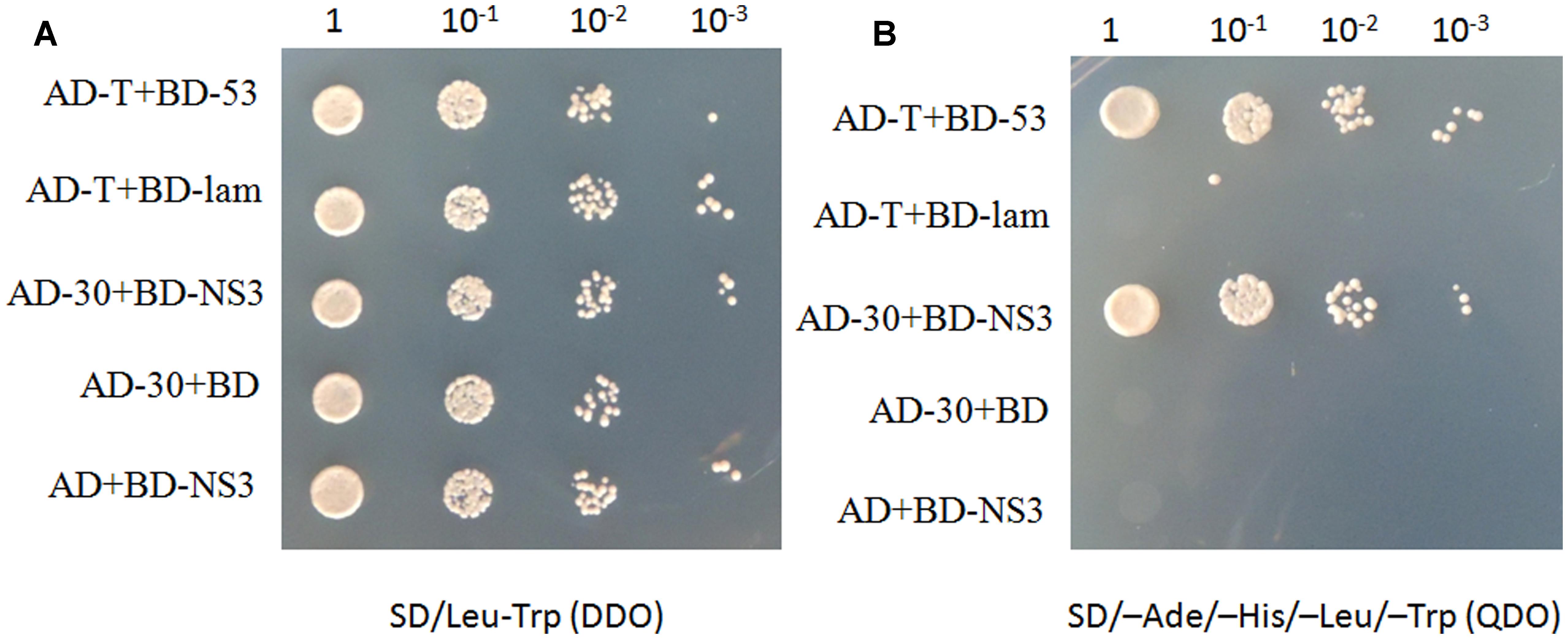
FIGURE 2. NS3 and OsCIPK30 interactions are detected in the yeast two-hybrid assay. Yeast strain Gold was co-transformed with the indicated plasmids shown on the left side of the panels. (A) The co-transformants were spread onto the SD/Leu-Trp (DDO) selection medium and (B) the SD/-Ade/-His/-Leu/-Trp (QDO) selection medium. AD-T+BD-53 were used as positive control, AD-T+BD-lam were used as negative control.
To demonstrate whether NS3 and OsCIPK30 also interacted in plant cells, a luciferase complementation imaging (LCI) assay was used. In the LCI experiments, the N- and C-terminal regions of firefly luciferase (NLuc and CLuc) were fused to NS3 and OsCIPK30, respectively, and were transiently expressed in N. benthamiana. When NLuc and CLuc are close to each other through the interaction of the two proteins, catalytic activity is restored and recorded through the CCD camera. In our LCI screening, NS3 displayed LUC complementation with OsCIPK30, suggesting that NS3 is physically close to OsCIPK30 in vivo (Figure 3C). Together, these assays clearly indicated that NS3 and OsCIPK30 interact in vivo.
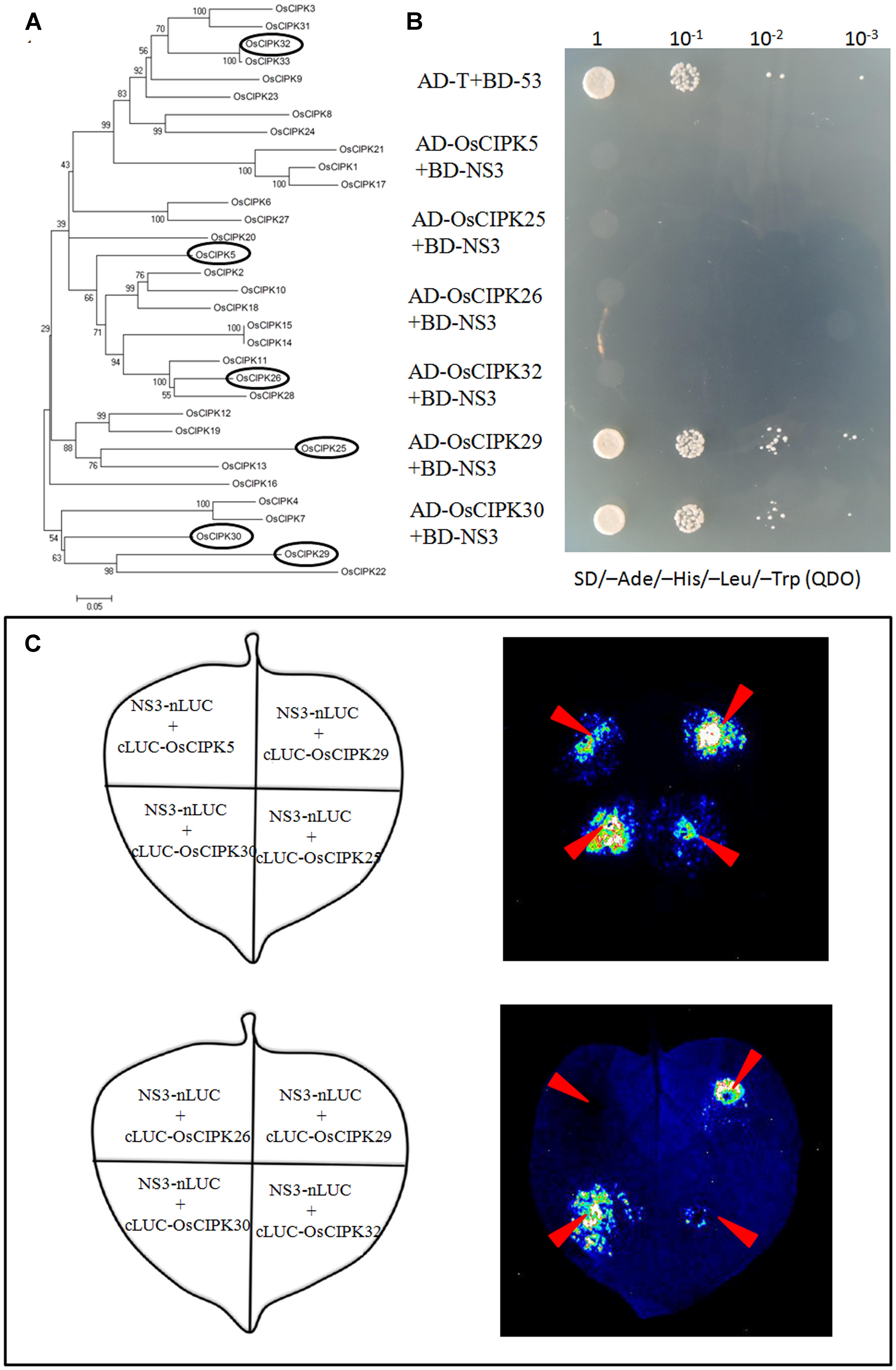
FIGURE 3. Phylogenetic analysis of OsCIPK proteins and the interaction between NS3 and OsCIPK5, OsCIPK25, OsCIPK26, OsCIPK29, OsCIPK30, and OsCIPK32 in yeast cells and plant cells. (A) Phylogenetic analysis of all 33 OsCIPK proteins. The analysis was performed using MEGA 4 software. The ellipses indicate the proteins chosen for the interaction assay. (B) The interaction between NS3 and OsCIPK5, OsCIPK25, OsCIPK26, OsCIPK29, OsCIPK30, and OsCIPK32 by yeast two-hybrid assay. The co-transformants were spread onto the SD/-Ade/-His/-Leu/-Trp (QDO) selection medium. (C) The interaction between NS3 and OsCIPK5, OsCIPK25, OsCIPK26, OsCIPK29, OsCIPK30, and OsCIPK32 by a luciferase complementation imaging (LCI) assay. Schematic representation of the LCI assay shows the different combinations of infiltrated constructs that were fused either to the N-terminal (NLuc) or C-terminal (CLuc) regions of luciferase. The infiltration positions of the constructs (red arrows) and luminescence signals resulting from the protein–protein interaction in a leaf are shown.
As mentioned above, OsCIPK30 belongs to the CIPK family in rice. According to the Institute for Genomic Research [TIGR] database and NCBI database, we collected all 33 CIPK proteins in rice named from OsCIPK1 to OsCIPK33. Phylogenetic analysis of OsCIPKs showed that the proteins could be classified into several groups (Figure 3A); we chose the other five OsCIPK proteins from different groups and performed Y2H and LCI assays to identify whether they also interacted with NS3. The results showed that only OsCIPK29 was able to interact with NS3, whereas OsCIPK5, 25, 26, and 32 could not (Figures 3B,C).
OsCIPK30 Is Induced by RSV Infection
To investigate whether RSV can induce the expression of OsCIPKs (OsCIPK5, OsCIPK25, OsCIPK26, OsCIPK29, OsCIPK30, and OsCIPK32), we examined the relative transcript level of these six OsCIPK genes in RSV-infected rice at 7 days post-inoculation (dpi). Real-time PCR showed that only three OsCIPKs were up-regulated, and two of them exhibited highly significant differences compared to the mock rice, suggesting that OsCIPK29 and OsCIPK30 might be involved in the host response to RSV infection (Figure 4).
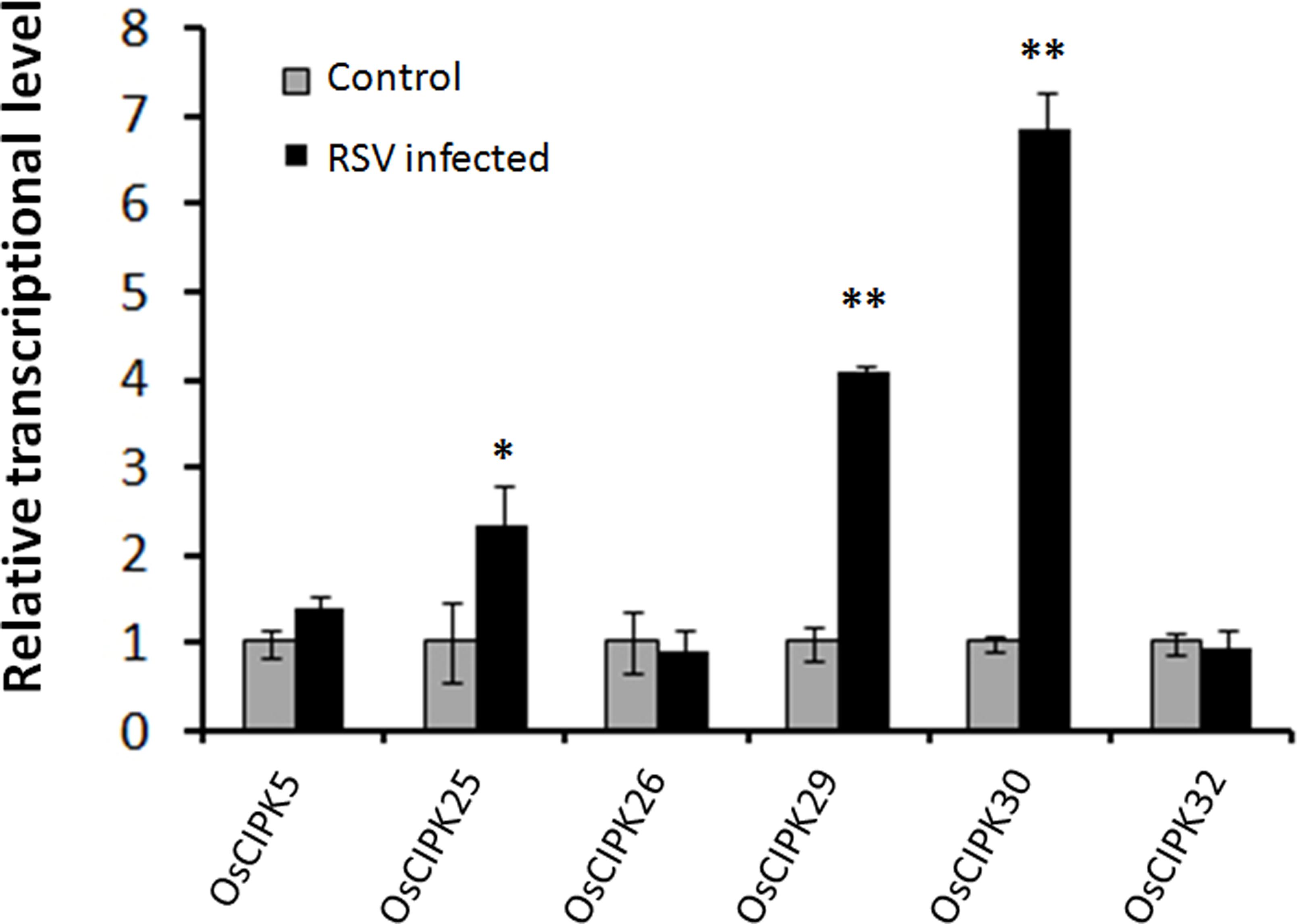
FIGURE 4. Expression of OsCIPK genes infected by RSV. The assay was performed by real-time RT-PCR. RNA was isolated from RSV-infected rice plants at 7 days post-inoculation. The relative mRNA levels were calculated with respect to the expression level of the corresponding transcript in the wild type (WT) rice plants without RSV infection. All the experiments were repeated three times, and similar results were obtained. The data represent the means ± SD of triplicate measurements. Asterisks above the columns represent significance based on an unpaired, two-tailed Student’s t-test relative to the WT. ∗∗P < 0.01; ∗P < 0.05.
Overexpression of OsCIPK30 Enhances the Tolerance to RSV
Given that the transcription level of OsCIPK30 is the highest in six OsCIPKs when RSV infects the host, we generated the overexpression transgenic rice of OsCIPK30 to further investigate its function. Under greenhouse conditions, the Ubi-OsCIPK30 rice plants showed normal growth compared with WT (Figure 5A). We then inoculated WT and Ubi-OsCIPK30 transgenic rice plants with RSV and observed the symptom development of the plants over a time course.
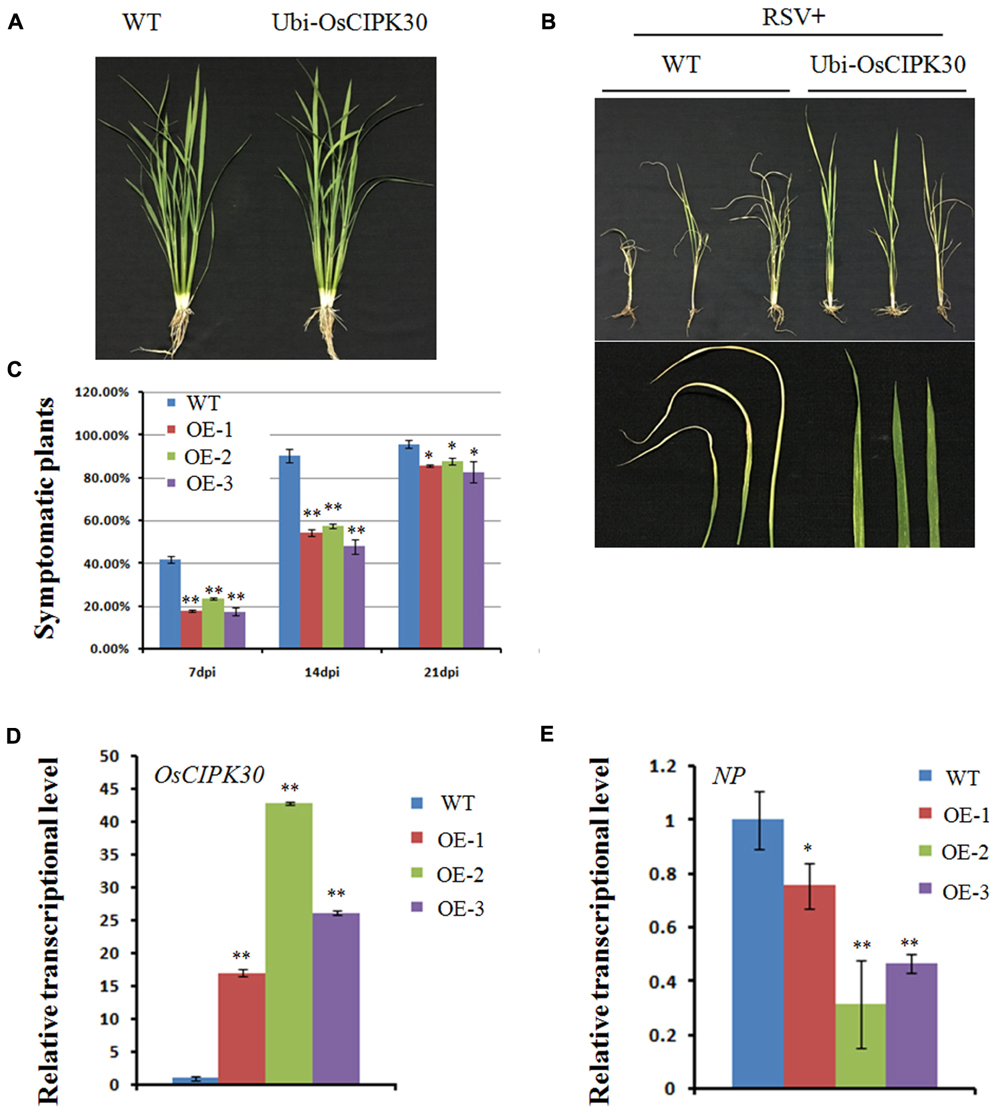
FIGURE 5. Effect of OsCIPK30 overexpression on RSV infection in rice. (A) Growth of the WT and Ubi-OsCIPK30 transgenic rice (T1 generation). (B) WT and Ubi-OsCIPK30 T1 transgenic rice were inoculated with RSV and photographed at 21 dpi. The lower panel of (B) shows the symptomatic leaves of the representative plants. (C) Temporal changes of RSV symptom development in WT and Ubi-OsCIPK30 T1 transgenic rice (OE-1, OE-2, OE-3). Thirty plants were used for each treatment in the experiment. (D) The expression level of OsCIPK30 in T1 transgenic rice (OE-1, OE-2, OE-3) by real-time RT-PCR at 21 dpi with RSV. The relative mRNA levels were calculated with respect to the expression level of the corresponding transcript in the WT rice plant. (E) The expression level of NP in T1 transgenic rice (OE-1, OE-2, OE-3) by real-time RT-PCR at 21 dpi with RSV. The relative mRNA levels were calculated with respect to the expression level of the corresponding transcript in the WT rice plant. All the experiments were repeated three times, and similar results were obtained. The data represent the means ± SD of triplicate measurements. The asterisks above the columns represent significance based on an unpaired, two-tailed Student’s t-test relative to the WT. ∗∗P < 0.01; ∗P < 0.05.
Fewer transgenic rice plants showed symptoms than WT rice at 7 and 14 dpi; moreover, the Ubi-OsCIPK30 rice showed milder RSV symptoms than those in the WT rice at 21 dpi (Figures 5B,C). These results suggested that Ubi-OsCIPK30 rice could delay the RSV symptoms and might influence the RSV infection. The real-time PCR results demonstrated that the expression levels of OsCIPK30 in transgenic rice plants were significantly increased, while the transcriptional level of the RSV NP gene was decreased (Figures 5D,E), indicating that the enhanced tolerance was associated with increased accumulation of OsCIPK30 transcripts.
Pathogenesis-Related Genes Are Up-regulated in OsCIPK30-Overexpressing Plants after RSV Infection
A previous study showed several pathogenesis-related genes involved in RSV infection (Satoh et al., 2010), and our results also showed that OsCIPK30 might influence RSV infection. To determine whether OsCIPK30 is involved in pathogenesis-related defense systems, we analyzed the expression patterns of several pathogenesis-related genes. The second youngest leaves were collected from five independent plants of Ubi-OsCIPK30 rice and WT rice at 7 dpi. The real-time PCR showed that RSV infection strongly induced the expression of PR1, PR10, PRB1-2, and PRB1-3 in OsCIPK30-overexpressing plants, showing significant differences between transgenic plants and WT plants, while the expression levels of the PR genes were not changed in un-inoculated OsCIPK30-overexpressing plants compared with WT rice (Figures 6A–D). We also took an experiment showing the effect of the PR1, PR10, PRB1-2, and PRB1-3 expression in WT plants by RSV infection (Figure 7). The results showed that the expression levels of PRB1-3 increased after RSV infection in WT rice, but PR1, PR10, PRB1-2 not. Together with the results showing in Figure 6, we conclude that the pathogenesis-related genes are triggered in OsCIPK30-overexpressing plants after RSV infection. Taken together, these results suggest that OsCIPK30 positively regulates the pathogenesis-related genes to enhance tolerance to RSV.
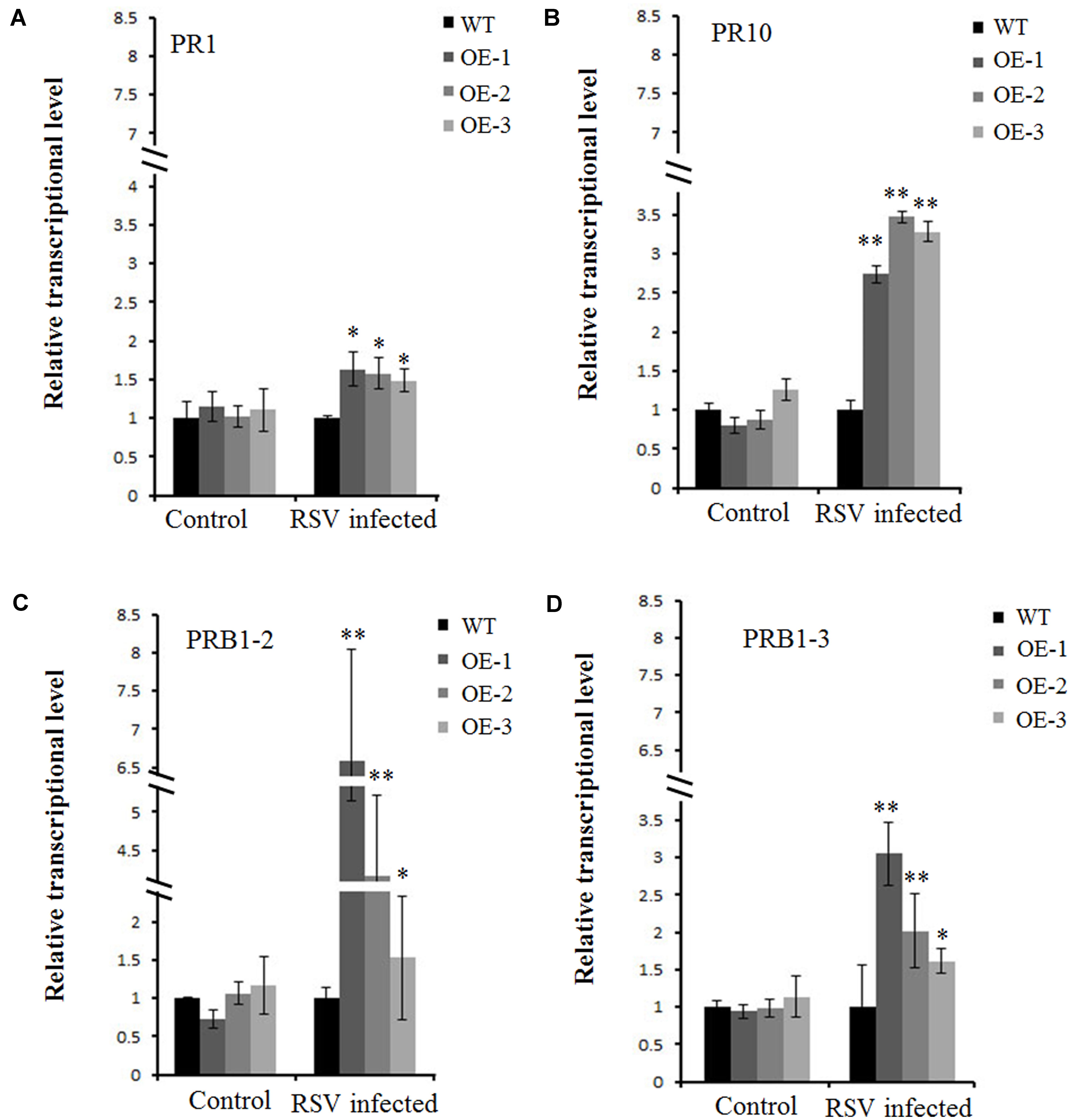
FIGURE 6. Transcriptionally modulated OsCIPK30 influenced the expression of a set of pathogenesis-related genes. (A) The expression levels of PR10 were significantly higher in OsCIPK30-overexpressing rice plants (OE-1, OE-2, OE-3) than WT plants after RSV infection. (B) The expression levels of PR1 were higher in OsCIPK30-overexpressing rice plants (OE-1, OE-2, OE-3) than WT plants after RSV infection. (C) The expression levels of PRB1-2 were significantly higher in OsCIPK30-overexpressing rice plants (OE-1, OE-2, OE-3) than WT plants after RSV infection. (D) The expression levels of PRB1-3 were significantly higher in OsCIPK30-overexpressing rice plants (OE-1, OE-2, OE-3) than WT plants after RSV infection. The un-inoculated plants were used as control. All experiments were repeated three times, and similar results were obtained. The data represent the means ± SD of triplicate measurements. Asterisks above the columns represent significance based on an unpaired, two-tailed Student’s t-test relative to the WT plants. ∗∗P < 0.01; ∗P < 0.05.
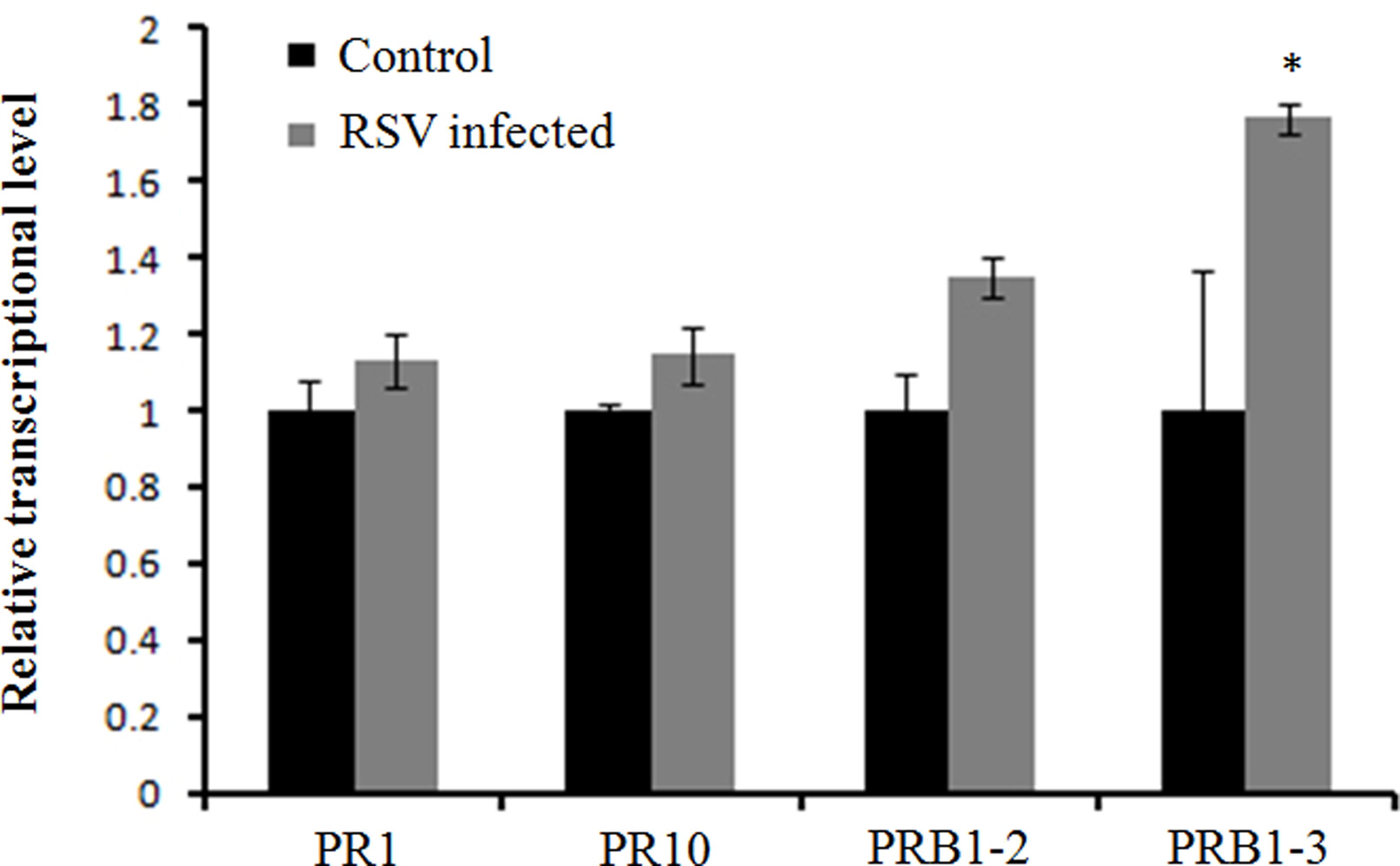
FIGURE 7. Expression of pathogenesis-related genes infected by RSV. The assay was performed by real-time RT-PCR. RNA was isolated from RSV-infected rice plants at 7 days post-inoculation. The relative mRNA levels were calculated with respect to the expression level of the corresponding transcript in the WT rice plants without RSV infection. All the experiments were repeated three times, and similar results were obtained. The data represent the means ± SD of triplicate measurements. Asterisks above the columns represent significance based on an unpaired, two-tailed Student’s t-test relative to the WT. ∗∗P < 0.01; ∗P < 0.05.
Discussion
We detected and confirmed an interaction between RSV NS3 and OsCIPK29 and OsCIPK30 (Figures 2, 3). To the best of our knowledge, this is the first report of plant host factors interacting with RSV NS3. NS3 is a viral suppressor of RNA silencing, and transgenic rice expressing NS3 could accumulate more RSV than in WT plants at the early stage of infection (Wu et al., 2014). In this study, we provided the transgenic Arabidopsis expressing NS3, which caused developmental abnormalities in plants (Figure 1A). All the results indicate that NS3 may be a pathogenic factor. Our results showed that NS3 could interact with OsCIPK29 and OsCIPK30 but failed to interact with other OsCIPK proteins. Given that OsCIPK genes have similar functional domains (the complete protein kinase domain and the NAF domain) (Xiang et al., 2007), the reason for the specific interaction between NS3 and OsCIPK29, 30 remains unclear.
As mentioned above, OsCIPK genes play some important roles in the response to abiotic stresses, such as drought, cold, salt, light, and sugars (Yu et al., 2014). In this study, OsCIPK30-overexpressing plants caused mild symptoms and induced high expression of pathogenesis-related genes (PR1, PR10, PRB1-2, and PRB1-3) upon RSV infection (Figures 5, 6), suggesting that OsCIPK30 may be involved in biotic stress. This provides further evidence that OsCIPK genes are activated in response to both abiotic and biotic stresses.
CIPK proteins can activate kinase activity though interacting with calcineurin-B-like (CBL) proteins and form plant-specific Ca2+ sensor-effector modules (Yang and Poovaiah, 2003; Luan, 2009; Weinl and Kudla, 2009). There is some evidence showing that some components of this module are endogenous RNA silencing regulators that can be recruited by some plant viruses, such as the Tobacco etch virus (TEV), Tomato golden mosaic virus (TGMV), and Tomato yellow leaf curl China virus (TYLCCNV) (Anandalakshmi et al., 2000; Chung and Sunter, 2014; Li et al., 2014). Since NS3 has been reported as a viral suppressor of RNA silencing (VSR), we hypothesize that NS3 could recruit OsCIPK30 to perform its VSR function. However, this hypothesis remains highly speculative until the mechanism of the interaction between these two proteins is identified. We tried to use a co-localization assay to investigate the relationship between NS3 and OsCIPK30 (Supplementary Figure 1). The assays showed that NS3, OsCIPK30 localized in both the cytoplasm and nucleus when they were expressed in tobacco leaf epidermal cells alone. We also observed that the localization of OsCIPK30 was not obviously different from that when co-expressed with NS3, which indicated that the localization of OsCIPK30 and NS3 was not affected by each other.
Compared with WT, OsCIPK30-overexpressing rice induced high expression of pathogenesis-related genes in infected rice plants (Figure 6). The production and accumulation of pathogenesis-related proteins in plants in response to biotic stresses is well-known and is considered a crucial mechanism for plant defense (van Loon et al., 2006; Sels et al., 2008). For example, the accumulation of OsPR1a was enhanced with the treatment of protein phosphatase inhibitor cantharidin; the expression of PR-4b from Theobroma cacao is increased after infection in resistant genotypes, while in the susceptible genotypes the expression was only activated at the final stages of infection; OgPR1a from Oryza grandiglumis conferred disease resistance on Arabidopsis to bacterial and fungal infections (Agrawal et al., 2000; Pereira Menezes et al., 2014; Shin et al., 2014). Our results support and enrich the above opinions and indicate that the overexpression of OsCIPK30 induces the high expression of pathogenesis-related genes to enhance tolerance to RSV.
Conclusion
Our results showed that the elevation of the pathogenesis-related genes occurs upon OsCIPK30 sensing RSV infection. The findings therefore provide new insight into the molecular mechanism underlying resistance to RSV disease.
Author Contributions
YZ and TZ conceived and designed the experiments. ZL, XL, and FS performed the experiments. ZL and XL analyzed the data. ZL wrote and revised the paper.
Funding
This work was supported by the National Natural Science Foundation of China (31601704), the China Postdoctoral Science Foundation (2016M591797), and Jiangsu Postdoctoral research funding program.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
The authors are grateful to Dr. Zhonghui Zhang (South China Normal University, China) for providing the plasmids for the luciferase complementation imaging assay.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02322/full#supplementary-material
DATA SHEET 1 | Primers used in this study.
References
Agrawal, G. K., Jwa, N. S., and Rakwal, R. (2000). A novel rice (Oryza sativa L.) acidic PR1 gene highly responsive to cut, phytohormones, and protein phosphatase inhibitors. Biochem. Biophys. Res. Commun. 274, 157–165. doi: 10.1006/bbrc.2000.3114
Albrecht, V., Ritz, O., Linder, S., Harter, K., and Kudla, J. (2001). The NAF domain defines a novel protein-protein interaction module conserved in Ca2+-regulated kinases. EMBO J. 20, 1051–1063. doi: 10.1093/emboj/20.5.1051
Anandalakshmi, R., Marathe, R., Ge, X., Herr, J. M. Jr., Mau, C., Mallory, A., et al. (2000). A calmodulin-related protein that suppresses posttranscriptional gene silencing in plants. Science 290, 142–144. doi: 10.1126/science.290.5489.142
Applied Biosystems (1997). “Relative quantitation of gene expression” in User Bulletin #2: ABI Prism 7700 Sequence Detection System, ed. K. J. Livak (Foster City, CA: Applied Biosystems).
Barbier, P., Takahashi, M., Nakamura, I., Toriyama, S., and Ishihama, A. (1992). Solubilization and promoter analysis of RNA polymerase from rice stripe virus. J. Virol. 66, 6171–6174.
Chung, H. Y., and Sunter, G. (2014). Interaction between the transcription factor AtTIFY4B and begomovirus AL2 protein impacts pathogenicity. Plant Mol. Biol. 86, 185–200. doi: 10.1007/s11103-014-0222-9
Clough, S. J., and Bent, A.F. (1998). Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743. doi: 10.1046/j.1365-313x.1998.00343.x
Du, Z., Xiao, D., Wu, J., Jia, D., Yuan, Z., Liu, Y., et al. (2011). p2 of Rice stripe virus (RSV) interacts with OsSGS3 and is a silencing suppressor. Mol. Plant Pathol. 12, 808–814. doi: 10.1111/j.1364-3703.2011.00716.x
Falk, B. W., and Tsai, J. H. (1998). Biology and molecular biology of viruses in the genus Tenuivirus. Annu. Rev. Phytopathol. 36, 139–163. doi: 10.1146/annurev.phyto.36.1.139
Hibino, H. (1996). Biology and epidemiology of rice viruses. Annu. Rev. Phytopathol. 34, 249–274. doi: 10.1146/annurev.phyto.34.1.249
Jiang, S., Lu, Y., Li, K., Lin, L., Zheng, H., Yan, F., et al. (2014). Heat shock protein 70 is necessary for Rice stripe virus infection in plants. Mol. Plant Pathol. 15, 907–917. doi: 10.1111/mpp.12153
Johnson, M. R., Wang, K., Smith, J. B., Heslin, M. J., and Diasio, R. B. (2000). Quantitation of dihydropyrimidine dehydrogenase expression by real-time reverse transcription polymerase chain reaction. Anal. Biochem. 278, 175–184. doi: 10.1006/abio.1999.4461
Kim, K. N., Lee, J. S., Han, H., Choi, S. A., Go, S. J., and Yoon, I. S. (2003). Isolation and characterization of a novel rice Ca2+-regulated protein kinase gene involved in responses to diverse signals including cold, light, cytokinins, sugars and salts. Plant Mol. Biol. 52, 1191–1202. doi: 10.1023/B:PLAN.0000004330.62660.a2
Kisimoto, R. (1965). “On the transovarial passage of the Rice stripe virus through the small brown planthopper, Laodelphax striatellus Fallen,” in Proceedings of the Conference on Relationships between Arthropods and Plant Pathogenic Viruses, Tokyo, 73–90.
Kolukisaoglu, U., Weinl, S., Blazevic, D., Batistic, O., and Kudla, J. (2004). Calcium sensors and their interacting protein kinases: genomics of the Arabidopsis and rice CBL-CIPK signaling networks. Plant Physiol. 134, 43–58. doi: 10.1104/pp.103.033068
Kong, L., Wu, J., Lu, L., Xu, Y., and Zhou, X. (2014). Interaction between Rice stripe virus disease-specific protein and host PsbP enhances virus symptoms. Mol. Plant 7, 691–708. doi: 10.1093/mp/sst158
Lee, K. W., Chen, P. W., Lu, C. A., Chen, S., Ho, T. H., and Yu, S. M. (2009). Coordinated responses to oxygen and sugar deficiency allow rice seedlings to tolerate flooding. Sci. Signal. 2, r61. doi: 10.1126/scisignal.2000333
Li, F., Huang, C., Li, Z., and Zhou, X. (2014). Suppression of RNA silencing by a plant DNA virus satellite requires a host calmodulin-like protein to repress RDR6 expression. PLOS Pathog. 10:e1003921. doi: 10.1371/journal.ppat.1003921
Li, J., Xiang, C. Y., Yang, J., Chen, J. P., and Zhang, H. M. (2015). Interaction of HSP20 with a viral RdRp changes its sub-cellular localization and distribution pattern in plants. Sci. Rep. 5:14016. doi: 10.1038/srep14016
Lu, L., Du, Z., Qin, M., Wang, P., Lan, H., Niu, X., et al. (2009). Pc4, a putative movement protein of Rice stripe virus, interacts with a type I DnaJ protein and a small Hsp of rice. Virus Genes 38, 320–327. doi: 10.1007/s11262-008-0324-z
Luan, S. (2009). The CBL-CIPK network in plant calcium signaling. Trends Plant Sci. 14, 37–42. doi: 10.1016/j.tplants.2008.10.005
Murashige, T., and Skoog, F. (1962). A revised medium for rapid growth and Bio assays with tobacco tissue cultures. Physiol. Plant. 15, 473–497. doi: 10.1111/j.1399-3054.1962.tb08052.x
Ohba, H., Steward, N., Kawasaki, S., Berberich, T., Ikeda, Y., Koizumi, N., et al. (2000). Diverse response of rice and maize genes encoding homologs of WPK4, an SNF1-related protein kinase from wheat, to light, nutrients, low temperature and cytokinins. Mol. Gen. Genet. 263, 359–366. doi: 10.1007/s004380051179
Pereira Menezes, S., de Andrade Silva, E. M., Matos Lima, E., Oliveira de Sousa, A., Silva Andrade, B., Santos Lima Lemos, L., et al. (2014). The pathogenesis-related protein PR-4b from Theobroma cacao presents RNase activity, Ca2+ and Mg2+ dependent-DNase activity and antifungal action on Moniliophthora perniciosa. BMC Plant Biol. 14:161. doi: 10.1186/1471-2229-14-161
Piao, H. L., Xuan, Y. H., Park, S. H., Je, B. I., Park, S. J., Kim, C. M., et al. (2010). OsCIPK31, a CBL-interacting protein kinase is involved in germination and seedling growth under abiotic stress conditions in rice plants. Mol. Cells 30, 19–27. doi: 10.1007/s10059-010-0084-1
Ramirez, B. C., and Haenni, A. L. (1994). Molecular biology of tenuiviruses, a remarkable group of plant viruses. J. Gen. Virol. 75, 467–475. doi: 10.1099/0022-1317-75-3-467
Satoh, K., Kondoh, H., Sasaya, T., Shimizu, T., Choi, I. R., Omura, T., et al. (2010). Selective modification of rice (Oryza sativa) gene expression by rice stripe virus infection. J. Gen. Virol. 91, 294–305. doi: 10.1099/vir.0.015990-0
Sels, J., Mathys, J., De Coninck, B. M., Cammue, B. P., and De Bolle, M. F. (2008). Plant pathogenesis-related (PR) proteins: a focus on PR peptides. Plant Physiol. Biochem. 46, 941–950. doi: 10.1016/j.plaphy.2008.06.011
Shen, M., Xu, Y., Jia, R., Zhou, X., and Ye, K. (2010). Size-independent and noncooperative recognition of dsRNA by the rice stripe virus RNA silencing suppressor NS3. J. Mol. Biol. 404, 665–679. doi: 10.1016/j.jmb.2010.10.007
Shin, S. H., Pak, J. H., Kim, M. J., Kim, H. J., Oh, J. S., Choi, H. K., et al. (2014). An Acidic PATHOGENESIS-RELATED1 Gene of Oryza grandiglumis is involved in disease resistance response against bacterial infection. Plant Pathol. J. 30, 208–214. doi: 10.5423/PPJ.NT.11.2013.0112
Sun, F., Fang, P., Li, J., Du, L., Lan, Y., Zhou, T., et al. (2016). RNA-seq-based digital gene expression analysis reveals modification of host defense responses by rice stripe virus during disease symptom development in Arabidopsis. Virol. J. 13, 202. doi: 10.1186/s12985-016-0663-7
Sun, F., Yuan, X., Zhou, T., Fan, Y., and Zhou, Y. (2011). Arabidopsis is susceptible to Rice stripe virus infections. J. Phytopathol. 159, 767–772. doi: 10.1111/j.1439-0434.2011.01840.x
Takahashi, M., Toriyama, S., Hamamatsu, C., and Ishihama, A. (1993). Nucleotide sequence and possible ambisense coding strategy of rice stripe virus RNA segment 2. J. Gen. Virol. 74, 769–773. doi: 10.1099/0022-1317-74-4-769
Toriyama, S. (1986). Rice stripe virus: prototype of a new group of viruses that replicate in plants and insects. Microbiol. Sci. 3, 347–351.
Toriyama, S., Takahashi, M., Sano, Y., Shimizu, T., and Ishihama, A. (1994). Nucleotide sequence of RNA 1, the largest genomic segment of rice stripe virus, the prototype of the tenuiviruses. J. Gen. Virol. 75, 3569–3579. doi: 10.1099/0022-1317-75-12-3569
van Loon, L. C., Rep, M., and Pieterse, C. M. (2006). Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 44, 135–162. doi: 10.1146/annurev.phyto.44.070505.143425
Wei, T. Y., Yang, J. G., Liao, F. R., Gao, F. L., Lu, L. M., Zhang, X. T., et al. (2009). Genetic diversity and population structure of rice stripe virus in China. J. Gen. Virol. 90, 1025–1034. doi: 10.1099/vir.0.006858-0
Weinl, S., and Kudla, J. (2009). The CBL-CIPK Ca2+-decoding signaling network: function and perspectives. New Phytol. 184, 517–528. doi: 10.1111/j.1469-8137.2009.02938.x
Wu, G., Wang, J., Yang, Y., Dong, B., Wang, Y., Sun, G., et al. (2014). Transgenic rice expressing rice stripe virus NS3 protein, a suppressor of RNA silencing, shows resistance to rice blast disease. Virus Genes 48, 566–569. doi: 10.1007/s11262-014-1051-2
Wu, J., Yu, L., Li, L., Hu, J., Zhou, J., and Zhou, X. (2007). Oral immunization with transgenic rice seeds expressing VP2 protein of infectious bursal disease virus induces protective immune responses in chickens. Plant Biotechnol. J. 5, 570–578. doi: 10.1111/j.1467-7652.2007.00270.x
Xiang, Y., Huang, Y., and Xiong, L. (2007). Characterization of stress-responsive CIPK genes in rice for stress tolerance improvement. Plant Physiol. 144, 1416–1428. doi: 10.1104/pp.107.101295
Xiong, R., Wu, J., Zhou, Y., and Zhou, X. (2008). Identification of a movement protein of the Tenuivirus rice stripe virus. J. Virol. 82, 12304–12311. doi: 10.1128/JVI.01696-08
Xiong, R., Wu, J., Zhou, Y., and Zhou, X. (2009). Characterization and subcellular localization of an RNA silencing suppressor encoded by Rice stripe tenuivirus. Virology 387, 29–40. doi: 10.1016/j.virol.2009.01.045
Yang, T., and Poovaiah, B. W. (2003). Calcium/calmodulin-mediated signal network in plants. Trends Plant Sci. 8, 505–512. doi: 10.1016/j.tplants.2003.09.004
Yang, W., Kong, Z., Omo-Ikerodah, E., Xu, W., Li, Q., and Xue, Y. (2008). Calcineurin B-like interacting protein kinase OsCIPK23 functions in pollination and drought stress responses in rice (Oryza sativa L.). J. Genet. Genomics 35, 531–543, S1–S2. doi: 10.1016/S1673-8527(08)60073-9
Yao, M., Liu, X., Li, S., Xu, Y., Zhou, Y., Zhou, X., et al. (2014). Rice stripe tenuivirus NSvc2 glycoproteins targeted to the golgi body by the N-terminal transmembrane domain and adjacent cytosolic 24 amino acids via the COP I- and COP II-dependent secretion pathway. J. Virol. 88, 3223–3234. doi: 10.1128/JVI.03006-13
Yu, Q., An, L., and Li, W. (2014). The CBL-CIPK network mediates different signaling pathways in plants. Plant Cell Rep. 33, 203–214. doi: 10.1007/s00299-013-1507-1
Zhang, X., Garreton, V., and Chua, N. H. (2005). The AIP2 E3 ligase acts as a novel negative regulator of ABA signaling by promoting ABI3 degradation. Genes Dev. 19, 1532–1543. doi: 10.1101/gad.1318705
Zhang, X., Henriques, R., Lin, S. S., Niu, Q. W., and Chua, N. H. (2006). Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 1, 641–646. doi: 10.1038/nprot.2006.97
Zhao, S. L., Dai, X. J., Liang, J. S., and Liang, C. Y. (2012). Surface display of rice stripe virus NSvc2 and analysis of its membrane fusion activity. Virol. Sin. 27, 100–108. doi: 10.1007/s12250-012-3237-x
Zhu, Y., Hayakawa, T., and Toriyama, S. (1992). Complete nucleotide sequence of RNA 4 of rice stripe virus isolate T, and comparison with another isolate and with maize stripe virus. J. Gen. Virol. 73, 1309–1312. doi: 10.1099/0022-1317-73-5-1309
Keywords: Rice stripe virus, NS3, interaction, OsCIPK30, pathogenesis-related genes
Citation: Liu Z, Li X, Sun F, Zhou T and Zhou Y (2017) Overexpression of OsCIPK30 Enhances Plant Tolerance to Rice stripe virus. Front. Microbiol. 8:2322. doi: 10.3389/fmicb.2017.02322
Received: 20 August 2017; Accepted: 10 November 2017;
Published: 24 November 2017.
Edited by:
Jean-François Laliberté, Institut National de la Recherche Scientifique, CanadaReviewed by:
Svetlana Folimonova, University of Florida, United StatesIl-Ryong Choi, International Rice Research Institute, Philippines
Copyright © 2017 Liu, Li, Sun, Zhou and Zhou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yijun Zhou, eWp6aG91QGphYXMuYWMuY24= Tong Zhou, emhvdXRvbmdAamFhcy5hYy5jbg==