 Xiaodong Liu
Xiaodong Liu Yantao Luo†
Yantao Luo† Zhefei Li
Zhefei Li- State Key Laboratory of Crop Stress Biology in Arid Areas, Shaanxi Key Laboratory of Agricultural and Environmental Microbiology, College of Life Sciences, Northwest Agriculture & Forestry University, Yangling, China
Mesorhizobium alhagi CCNWXJ12-2T, isolated from root nodules of the desert plant Alhagi sparsifolia, contains two type III secretion systems (T3SSs). T3SSs are specialized machinery with wide distribution in bacteria that inject effector proteins into target cells. Our previous study showed that the expression of M. alhagi T3SS1 is upregulated in high-salt conditions. Here, phylogenetic analysis of T3SS1 using the core protein RhcU suggested that T3SS1 belongs to the α-Rhc II subgroup of the Rhc T3SS family. To elaborate the function of M. alhagi CCNWXJ12-2T T3SS1 in stress adaption, two T3SS1 mutants (ΔrhcQ and ΔMA29250) were constructed and analyzed. β-galactosidase transcriptional fusion assays showed that activity of the promoter of T3SS1 was induced by salts. Mutant ΔrhcQ was more sensitive to NaCl and LiCl than the wild-type, but ΔMA29250 was not. Both mutants were more sensitive to KCl than the wild-type. The intracellular Na+ concentration in ΔrhcQ in high-NaCl conditions (0.4 M) increased by 37% compared to that of the wild-type strain, while the Na+ concentration in ΔMA29250 increased by 13%. The K+ concentration in both mutants increased by 16% compared to the wild-type in high-KCl conditions (0.3 M). Strain ΔrhcQ showed decreased survival compared to the wild-type after treatment with H2O2, while the survival rate of ΔMA29250 was almost the same as that of the wild-type. Antioxidant enzyme activities in ΔrhcQ were lower than those in the wild-type strain, but this was not the case for ΔMA29250. Our data elucidate the beneficial effects of T3SS1 in the adaption of M. alhagi CCNWXJ12-2T to stress.
Introduction
The type III secretion systems (T3SSs) are widespread in Gram-negative bacteria; their main function is to translocate effector proteins from bacteria into host cells. Many reports have shown that T3SSs play an important role in bacterial virulence (Galán and Collmer, 1999; Rolhion et al., 2016; Wang et al., 2016) and in symbiotic interactions between microorganisms and their hosts (Silver et al., 2007; Buttner and He, 2009; Correa et al., 2012). Because of the important roles that T3SSs play in bacteria–host cell interactions, the T3SSs are arguably one of the best characterized protein injection systems (Galán et al., 2014). There are also some reports on T3SS expression in high-salt conditions. In Yersinia enterocolitica, the expression of a chromosomally encoded T3SS was found to be triggered in high-salt conditions and at low temperature (Haller et al., 2000). Microarray analysis of Pseudomonas aeruginosa showed that expression of T3SS and associated cytotoxins was upregulated in hyperosmotic stress conditions (Aspedon et al., 2006). Similar results were also found for Salmonella enterica serovar Typhimurium (Mizusaki et al., 2008). Most research has focused on changes in pathogenicity and protein secretion between T3SS-related mutants and wild-type strains (Haller et al., 2000; Galán et al., 2014). However, little is known about the functions of T3SSs in stress resistance.
In rhizobial T3SSs, the core genes are called the rhc genes (Rhizobium conserved) to distinguish them from phytopathogenic bacterial T3SSs (Viprey et al., 1998). Phylogenetic analysis of T3SS core genes suggested that the rhizobial T3SSs could constitute an individual family (the Rhc family); further analysis showed that the Rhc family could be divided into four subgroups: α-Rhc I, α-Rhc II, α-Rhc III, and β-Rhc (Tampakaki, 2014). Most T3SSs in the α-Rhc I group play an important role in symbiosis formation, for example, in Bradyrhizobium japonicum (Krause et al., 2002), Sinorhizobium fredii (Lorio et al., 2004), and Mesorhizobium loti (Sanchez et al., 2012). However, there is little knowledge regarding the functions of T3SSs in group α-Rhc II (Tampakaki, 2014).
Mesorhizobium alhagi CCNWXJ12-2T (=ACCC 14561T = HAMBI 3019T) is a soil-dwelling α-proteobacterium isolated from root nodules of the desert plant Alhagi sparsifolia and possessing high salt resistance (up to 0.8 M NaCl) (Chen et al., 2010). Two T3SSs, designated as T3SS1 and T3SS2, were found in the draft genome sequence of M. alhagi CCNWXJ12-2T (GenBank accession number AHAM00000000) (Zhou et al., 2012). To investigate the transcriptional changes in M. alhagi CCNWXJ12-2T in high salt conditions, a transcriptome analysis was conducted previously and the results revealed that the expression of T3SS1 was upregulated significantly in high-salt conditions, while the expression of T3SS2 was downregulated (GEO accession number GSE57306) (Liu et al., 2014). Phylogenetic analysis of RhcU (or SctU), a core protein of T3SS, suggested that M. alhagi T3SS1 belongs to the α-Rhc II group and T3SS2 belongs to the α-Rhc I group.
The upregulation of M. alhagi T3SS1 in high-salt conditions and little knowledge of T3SS functions in α-Rhc II group prompted us to explore here the role of T3SS1 in salt resistance and stress adaption by M. alhagi CCNWXJ12-2T. The promoter activity of T3SS1 was studied under salt stresses and the phenotypes of two T3SS1 gene knockout strains were compared to the wild-type strain. Our results showed that T3SS1 plays an important role in stress adaption in M. alhagi CCNWXJ12-2T. We propose a new function of the α-Rhc II group T3SSs in rhizobia.
Materials and Methods
Bacterial Strains and Growth Conditions
Table 1 lists the bacterial strains and plasmids used in this work. The wild-type strain of M. alhagi CCNWXJ12-2T, ΔrhcQ and ΔMA29250 mutants, and their complemented strains were typically grown in tryptone–yeast extract (TY) broth (5 g tryptone, 3 g yeast extract, and 0.7 g CaCl2⋅2H2O per liter) at 28°C. Salt mannitol (SM) medium (0.2 g MgSO4⋅7H2O, 0.1 g CaCl2, 0.5g KNO3, 0.5 g K2HPO4, 0.1 g NaCl, 10 g mannitol, 75 mg pantothenic acid, 75 mg biotin, and 75 mg thiamine per liter) was used to isolate gene knockout mutants and complementation strains. Escherichia coli was grown in Luria-Bertani (LB) broth (10 g tryptone, 5 g yeast extract, and 10 g NaCl per liter) at 37°C. All bacteria were incubated in aerobic conditions at 180 rpm. Where necessary, antibiotics were added at the following concentrations: kanamycin 100 μg/ml; gentamicin 50 μg/ml.

TABLE 1. Bacterial strains and plasmids used in this study.
Phylogenetic Analysis of T3SSs
Phylogenetic analysis was performed using the protein sequences of RhcU, one of the core T3SS proteins, including RhcU1 (locus_tag = “MAXJ12_29220,” from T3SS1) and RhcU2 (locus_tag = “MAXJ12_24267,” from T3SS2). Other closely related RhcU sequences of rhizobia were retrieved from the NCBI1 database. MEGA 5.1 software was used to align the protein sequences with the multiple alignment method ClustalW and then to analyze the phylogenetic relationships between different species using the neighbor-joining method with 1000 bootstrap replicates (Tamura et al., 2011). Evolutionary distances were computed using Poisson correction and are expressed in terms of the number of amino acid substitutions per site (Zuckercandl and Pauling, 1965).
Construction of Promoter Reporter Strain
The promoter reporter plasmid pBML was derived from pBBR1MCS-5. The lacZ promoter in pBBR1MCS-5 was deleted and a fragment of lacZY cloned from E. coli K-12 was used to replace lacZα in pBBR1MCS-5; the new plasmid was called pBML. A DNA fragment (named TP13) containing rhcC and the putative promoter of T3SS1 (the upstream 200 bp of rhcC) was amplified from M. alhagi CCNWXJ12-2T genomic DNA using primers TP1 and TP3 (Supplementary Table S1). The DNA fragment was digested with HindIII and XbaI and then cloned into pBML digested with the same restriction enzymes. Finally, the promoter reporter plasmid, designated pBMLTP13, was transformed into E. coli DH5α and the strain was named DH-TP13.
To generate promoter reporter strain M. alhagi XJTP13, a triparental mating procedure was conducted as described previously to transform the plasmids from E. coli into CCNWXJ12-2T (Liu et al., 2014). Briefly, strain DH-TP13, strain E. coli MM249 containing helper plasmid pRK2013, and M. alhagi CCNWXJ12-2T were mixed together and incubated on a TY plate for 4 days. Then, SM agar plates containing gentamicin were used to isolate M. alhagi containing pBMLTP13. Strain XJTP13 was verified by PCR and sequencing.
β-Galactosidase Transcriptional Fusion Assay
Strain XJTP13 was first grown to OD600 ≈0.8 in 20 ml TY broth containing gentamicin. Then, 20 μl suspensions were added to 20 ml TY broth containing gentamicin and different concentrations of additional salts (0, 0.1 M NaCl, 0.2 M NaCl, 0.3 M NaCl, 0.4 M NaCl, 0.1 M LiCl, 0.2 M LiCl, 0.1 M KCl, 0.2 M KCl, or 0.3 M KCl) and grown to OD600 ≈0.8. Then, 100 μl of each culture was subjected to β-galactosidase activity assay using o-nitrophenyl-β-D-galactopyranoside as the substrate (Zhang and Bremer, 1995). Three independent biological repetitions were conducted.
Construction of Gene Knockout Mutants and Complementation Strains
Supplementary Table S1 lists primers used in construction of gene knockout mutants and complemented strains. A rhcQ (locus_tag = “MAXJ12_29275”) in-frame deletion mutant of M. alhagi CCNWXJ12-2T was constructed using plasmid pK18rhcQ. Primer pairs RhcQ-US/RhcQ-UA and RhcQ-DS/RhcQ-DA were used to amplify the 522-bp upstream DNA fragment and 568-bp downstream DNA fragment of rhcQ, respectively. The upstream and downstream DNA fragments were digested with EcoRI/XbaI and XbaI/HindIII, respectively, then the digested DNA fragments were cloned into linearized plasmid pK18mobsacB digested with EcoRI and HindIII, to generate pK18rhcQ.
An in-frame deletion mutant of gene MA29250 (locus_tag = “MAXJ12_29250”) was constructed using plasmid pK18MA29250. The procedure used to produce pK18MA29250 was similar to the procedure for pK18rhcQ construction. Briefly, the upstream and downstream DNA fragments of MA29250 were amplified using primer pairs MA29250-US/MA29250-UA and MA29250-DS/MA29250-DA and then digested using the same restriction enzymes as used for pK18rhcQ construction. The digested DNA fragments were then cloned into plasmid pK18mobsacB digested with EcoRI and HindIII to generate pK18MA29250.
To complement the ΔrhcQ and ΔMA29250 mutants, primer pairs CRhcQA/CRhcQB and CMA29250A/CMA29250B were used to amplify rhcQ and MA29250 gene fragments from genomic DNA of M. alhagi CCNWXJ12-2T. The gene fragments were then cloned into plasmid pBML digested with XbaI using the ClonExpress MultiS One Step Cloning Kit (Vazyme Biotech, China), to generate complementation plasmids pBMLrhcQ and pBMLMA29250.
For construction of in-frame deletion mutants and complementation strains, a triparental mating procedure was conducted as described previously (Liu et al., 2014). Briefly, the recipient strains (the wild-type strain or the mutants), the donor strains (E. coli DH5α containing the constructed plasmids), and the helper strain (E. coli MM249 containing helper plasmid pRK2013) were grown to log phase and mixed together in the ratio 4:1:1. The mixtures were cultured on TY plates for 4 days. For in-frame deletion mutant isolation, SM plates containing kanamycin were used to isolate single exchange mutants (gene knockout plasmids integrated into the genome of M. alhagi CCNWXJ12-2T), then TY plates containing sucrose (5 g/100 ml) were used to isolate the double exchange mutants (in-frame gene deletion mutants). SM agar plates containing gentamicin were used initially to isolate the complementation strains; the single exchange gene knockout mutants isolated from SM plates were then smeared onto TY plates containing sucrose (5 g/100 ml) to isolate the double exchange mutants. The gene knockout mutants and complemented strains were verified by PCR.
Salt Resistance Assay
Salt resistance assays were conducted as described previously (Liu et al., 2016). Wild-type M. alhagi CCNWXJ12-2T, ΔrhcQ and ΔMA29250, and the complemented strains were grown to log phase in 20 ml TY broth with shaking at 28°C. Next, 20 μl of the cultures were inoculated into 20 ml fresh TY broth and grown to OD600 ≈0.8. The cultures were diluted fourfold with sterile H2O, then 5 μl of each were spotted onto TY agar plates with or without salts (NaCl 0.4 M, LiCl 0.3 M, or KCl 0.3 M). Plates were incubated at 28°C for 4 days.
Measurement of Total Cellular Na+ and K+ Concentrations
Inocula were prepared as described in the section on salt resistance assays. Each inoculum (500 μl, OD600 ≈0.2) was smeared onto TY agar plates containing different salts (no added salt, 0.4 M NaCl or 0.3 M KCl) and incubated at 28°C for 4 days (plates containing 0 and 0.4 M NaCl) or 7 days (plates containing 0.3 M KCl). The bacteria were then collected in 1.5-ml sterile tubes using sterile stainless steel spoons and dried at 60°C for 10 h using an air dryer. The total cellular Na+ and K+ concentrations were measured using an atomic absorption spectrophotometer (Hitachi, Tokyo, Japan).
Hydrogen Peroxide Sensitivity Assay
H2O2 resistance assays were conducted as described previously (Liu et al., 2016). Wild-type M. alhagi CCNWXJ12-2T, ΔrhcQ, ΔMA29250, and the complementation strains (CΔrhcQ and CΔMA29250) were cultured to OD600 ≈0.6 at 28°C in TY broth containing antibiotics and then diluted with sterile H2O to OD600 ≈0.1. After treatment with 10 mM H2O2 for 30 min, the survival rates of all the strains were measured by colony counts compared to cells without peroxide treatment, calculated as follows: [(CFU per ml after treatment with H2O2)/(CFU per ml before treatment with H2O2)] × 100.
Antioxidant Enzyme Activity Assay
A previous report showed that peroxidase activity was not detectable in M. alhagi CCNWXJ12-2T (Liu et al., 2016), so here, we measured catalase (CAT) and superoxide dismutase (SOD) activities in normal and stressed conditions. Wild-type M. alhagi CCNWXJ12-2T, ΔrhcQ, ΔMA29250, CΔrhcQ, and CΔMA29250 were grown in TY broth containing antibiotics to OD600 ≈0.8. Each strain (20 μl suspension) was inoculated into two bottles containing 20 ml TY broth and one bottle containing 20 ml TY broth plus 0.4 M NaCl, and grown to OD600 ≈0.8. For the two bottles without additional NaCl, one was treated with 0.1 mM H2O2 for 30 min and the other without. All cells were collected in 50-ml tubes by centrifugation, resuspended in phosphate-buffered saline, and lysed by ultrasonication. The antioxidant enzyme activities were measured using a CAT Assay Kit and a SOD Assay Kit (Suzhou Comin Biotechnology, China). Total soluble protein concentrations were determined using a BCA Protein Assay Kit (CoWin Biosciences, China).
Statistical Analysis
Statistical differences between the wild-type strain and the mutants in different conditions were assessed by t-test using SPSS version 15.0 (SPSS Inc., Chicago, IL, United States). Differences were considered significant at P < 0.05.
Results
Phylogenetic Analysis of T3SSs in M. alhagi CCNWXJ12-2T
Protein sequences of RhcU, one of the core T3SS proteins, were used to build an evolutionary tree of different rhizobia. Three subgroups of Rhc family T3SSs were apparent in the phylogenetic analysis. M. alhagi CCNWXJ12-2T T3SS1 was in subgroup α-Rhc II and T3SS2 was in subgroup α-Rhc I (Figure 1).
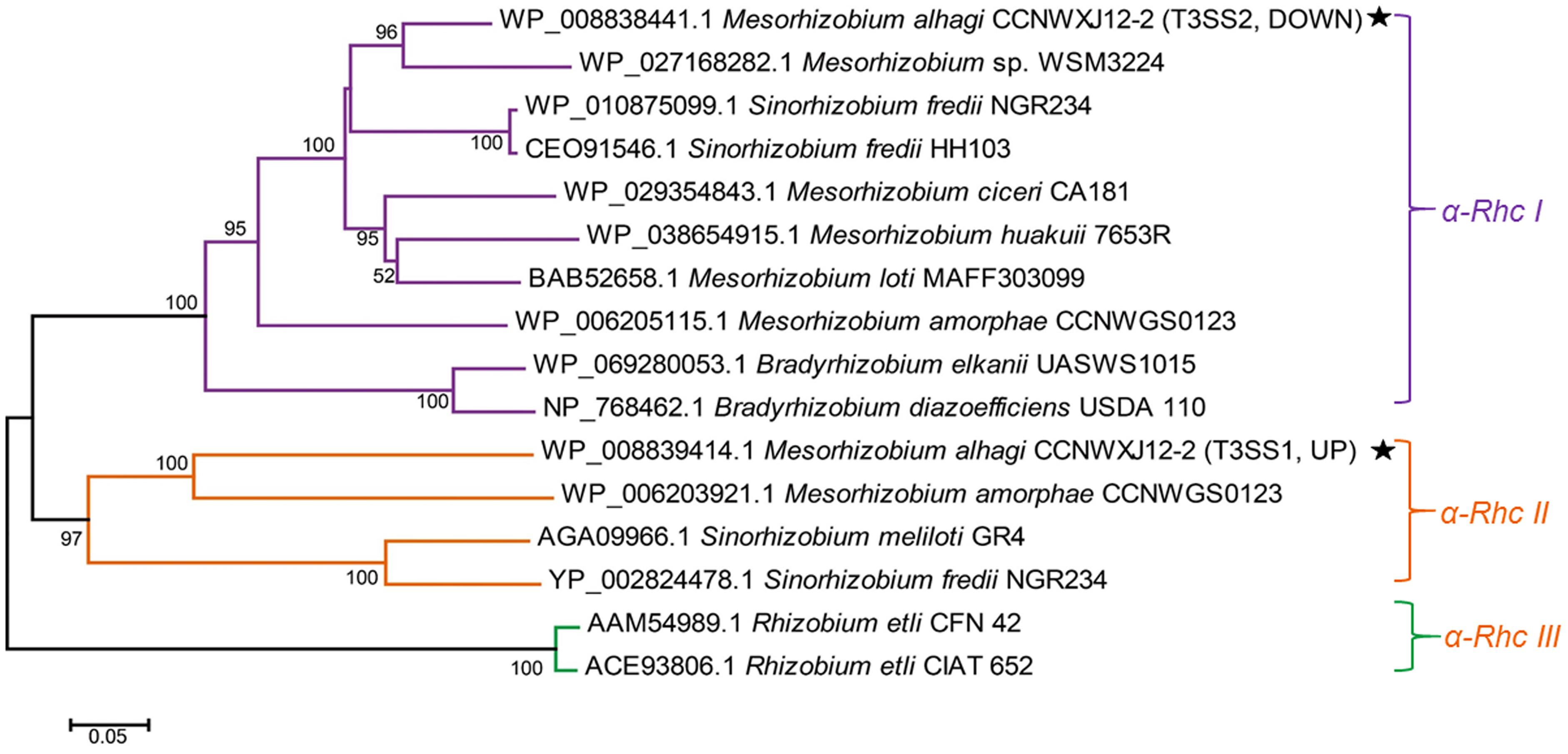
FIGURE 1. Neighbor-joining tree of RhcU sequences from two type III secretion systems (T3SSs) in the draft genome of M. alhagi CCNWXJ12-2T. The black stars indicate T3SS1 and T3SS2. UP and DOWN indicate the expression pattern of the T3SSs in high-salt conditions (3). Evolutionary distances were estimated using the p-distance method and are in units of the number of amino acid differences per site. Numbers to the left of the branches are bootstrap percentages for 1000 replications. Bootstrap values > 50% are shown, and the scale bar represents the number of substitutions per site. Evolutionary analysis was conducted using MEGA 5.1 (Tamura et al., 2011).
T3SS1 Promoter Activity Is Induced by Salts
The T3SS1 genes are in an operon in the genome of M. alhagi CCNWXJ12-2T (Figure 2). The putative promoter (200 bp upstream of rhcC) (Figure 2) was used with the whole gene of rhcC to construct a promoter reporter plasmid. The promoter activities were measured in different conditions using β-galactosidase tests. The β-galactosidase activities increased with increasing salt concentration (Figure 3). At the same concentration of different salts (i.e., NaCl, LiCl, and KCl), the β-galactosidase activities were almost the same. The β-galactosidase activities at the lowest salt concentration tested (0.1 M) were almost three times higher than that in the control group (no added salt), while at salt concentrations of 0.3 M they were about six times higher than that in the control group.
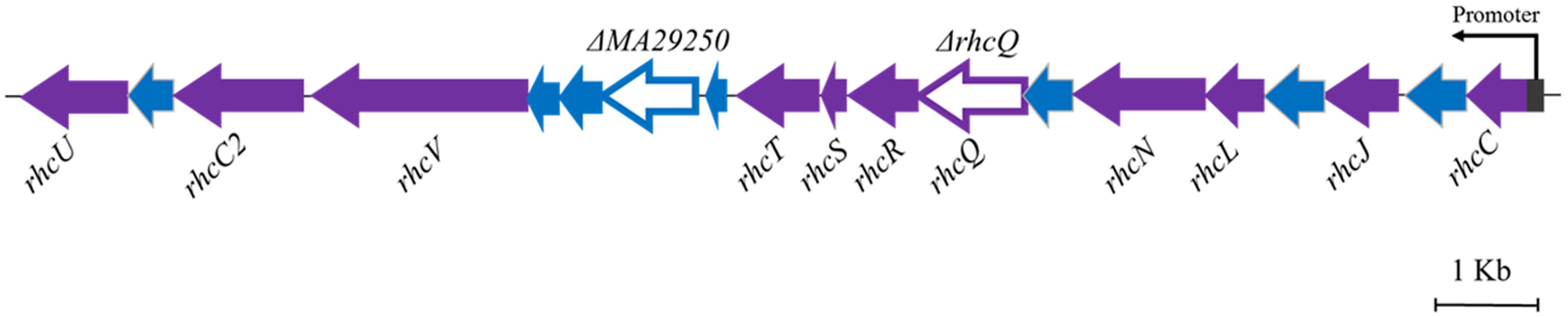
FIGURE 2. Genetic organization of M. alhagi CCNWXJ12-2T T3SS1. Purple arrows indicate conserved T3SS structural component genes. Blue arrows indicate hypothetical protein encoding genes. The genes chosen to construct in-frame deletion mutants are shown as open triangles and the mutant names are above the genes. The putative promoter of T3SS1 is shown as a dark gray rectangle and the arrow indicates the direction of transcription.
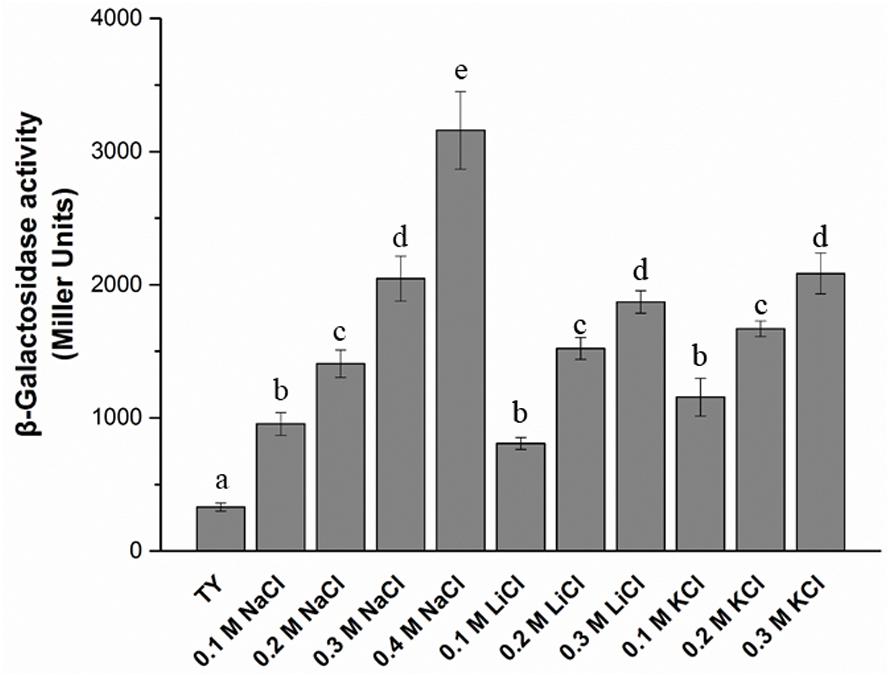
FIGURE 3. β-galactosidase activity of promoter reporter strain M. alhagi XJTP13 on exposure to different salts and salt concentrations. Strain XJTP13 was grown to OD600 ≈0.8 in TY broth containing added salts (0–0.4 M NaCl, 0.1–0.2 M LiCl, or 0.1–0.3 M KCl). Then, 100 μl of each culture was taken for β-galactosidase activity assay. Values given are means ± SDs of three replicates. The significance of differences is shown at the P < 0.05 level (t-test). Different lowercase letters mean significant differences between two columns.
Decrease in Salt Resistance of T3SS1-Related Mutants
Two gene knockout mutants, ΔrhcQ and ΔMA29250, were constructed to study the role T3SS1 plays in salt resistance. One of the genes knocked-out (Figure 2) is rhcQ (locus_tag = “MAXJ12_29275”), encoding a component of the C-ring of T3SS1 which plays an important role in T3SS assembly, and the other is MA29250, encoding a hypothetical protein. MA29250 (locus_tag = “MAXJ12_29250”) was randomly chosen to study its function. The salt resistance of strain ΔrhcQ was decreased compared to the wild-type strain (Figure 4). However, the salt resistance of ΔMA29250 was different from that of ΔrhcQ. The NaCl and LiCl tolerances of ΔMA29250 were similar to those of the wild-type, while the KCl tolerance of ΔMA29250 was decreased dramatically (Figure 4). Both mutants showed more sensitivity to KCl than to NaCl and LiCl, indicating that T3SS1 plays a more important role in resistance to KCl than to the other salts. All the complemented strains had similar salt resistance to the wild-type (Figure 4).

FIGURE 4. Sensitivity of M. alhagi CCNWXJ12-2T to different salts. Wild-type (WT), mutants (ΔrhcQ and ΔMA29250), and the complementation strains (CΔrhcQ and CΔMA29250) were grown to OD600 ≈0.8 in TY broth containing appropriate antibiotics, and then adjusted to OD600 ≈0.2. The inocula were serially diluted and spotted onto TY agar plates containing no additional salt or containing 0.4 M NaCl, 0.3 M LiCl, or 0.3 M KCl. The 10-fold serial dilutions are shown. All plates were incubated at 28°C for 4 days.
Total Cellular Na+ and K+ Concentration Measurement
To explore the reasons for the decrease in salt resistance of the mutants ΔrhcQ and ΔMA29250, total cellular Na+ and K+ were measured after growth in 0.4 M NaCl and 0.3 M KCl, respectively. The Na+ and K+ concentrations in both mutants increased compared to the wild-type strain (Figure 5). In 0.4 M NaCl, the Na+ concentration in ΔrhcQ increased significantly (by about 37%) compared to the wild-type strain, while the Na+ concentration of ΔMA29250 increased non-significantly (about 13%) (Figure 5A). In 0.3 M KCl, the increases in K+ concentration in strains ΔrhcQ and ΔMA29250 were significant, both around 16% compared to the wild-type (Figure 5B). Although the increments of K+ concentrations in the mutants were lower than that of the Na+ concentration in strain ΔrhcQ, the overall K+ concentrations in all strains grown in 0.3 M KCl were about twice the Na+ concentrations in cells grown in 0.4 M NaCl (Figure 5). The higher K+ concentrations may explain the more serious growth repression caused by KCl than by NaCl shown in Figure 2. The Na+ and K+ concentrations in the complemented strains were similar to those in the wild-type.
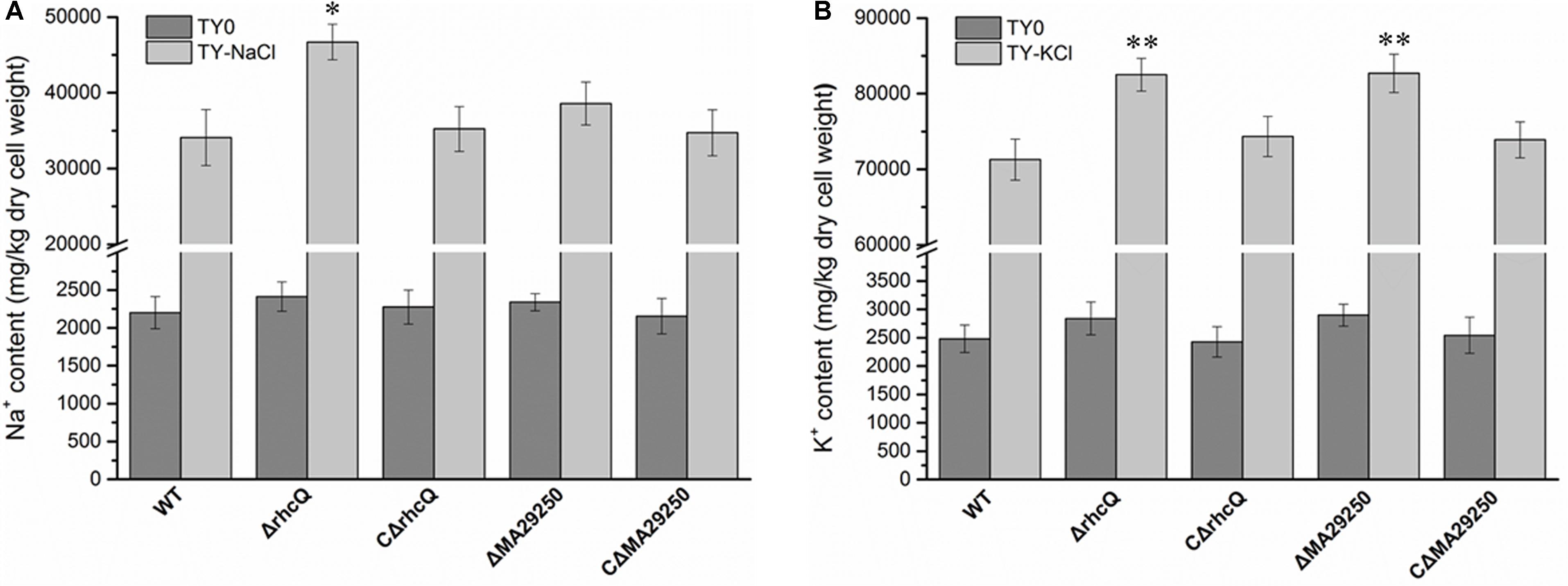
FIGURE 5. Total cellular Na+ and K+ concentrations of M. alhagi CCNWXJ12-2T. Wild-type (WT), mutants (ΔrhcQ and ΔMA29250), and the complementation strains (CΔrhcQ and CΔMA29250) were grown to OD600 ≈0.8 in TY broth containing appropriate antibiotics. A total of 500 microliters of each inoculum were plated on TY agar plates, TY plates containing 0.4 M NaCl, and TY plates containing 0.3 M KCl (all containing appropriate antibiotics) and inoculated at 28°C for 5 days. Then, the bacterial cells were collected for Na+ and K+ concentration measurement; those from TY plates containing 0.4 M NaCl were collected for Na+ concentration measurement, and those from TY plates containing 0.3 M KCl were collected for K+ concentration measurement. (A) Na+ concentrations in different strains; (B) K+ concentrations in different strains. Three independent experiments were conducted and the error bars indicate SDs. Significant differences between the WT strain and other strains are indicated (∗P < 0.05; ∗∗P < 0.01).
Effect of T3SS1 on Antioxidant Capacity in M. alhagi CCNWXJ12-2T
To test whether T3SS1 has a role in oxidative stress resistance, we tested the antioxidant capacity of the mutants. The survival rate of ΔrhcQ after treatment with 10 mM H2O2 for 30 min was significantly decreased (by 50%) compared to the wild-type strain, while the survival rate of ΔMA29250 was almost the same as that of the wild-type (Figure 6). The survival rates of the complemented strains after H2O2 treatment were also almost the same as that of the wild-type strain (Figure 6). The antioxidant enzyme activities of all the strains were measured in different conditions. In normal conditions (with no stresses), the SOD activity of ΔrhcQ showed a significant decrease, while the CAT activities of both mutants and the SOD activity of ΔMA29250 showed no significant difference compared to the wild-type. The CAT and SOD activities of all strains were increased to varying degrees under oxidative stress (25–89%) and salt stress (110–160%) compared to normal conditions. The CAT and SOD activities of ΔrhcQ under salt and oxidative stresses were significantly decreased compared to the wild-type (68–78% of the level for the wild-type strain), while in ΔMA29250, the CAT and SOD activities showed no significant differences compared to the wild-type (Figure 7).
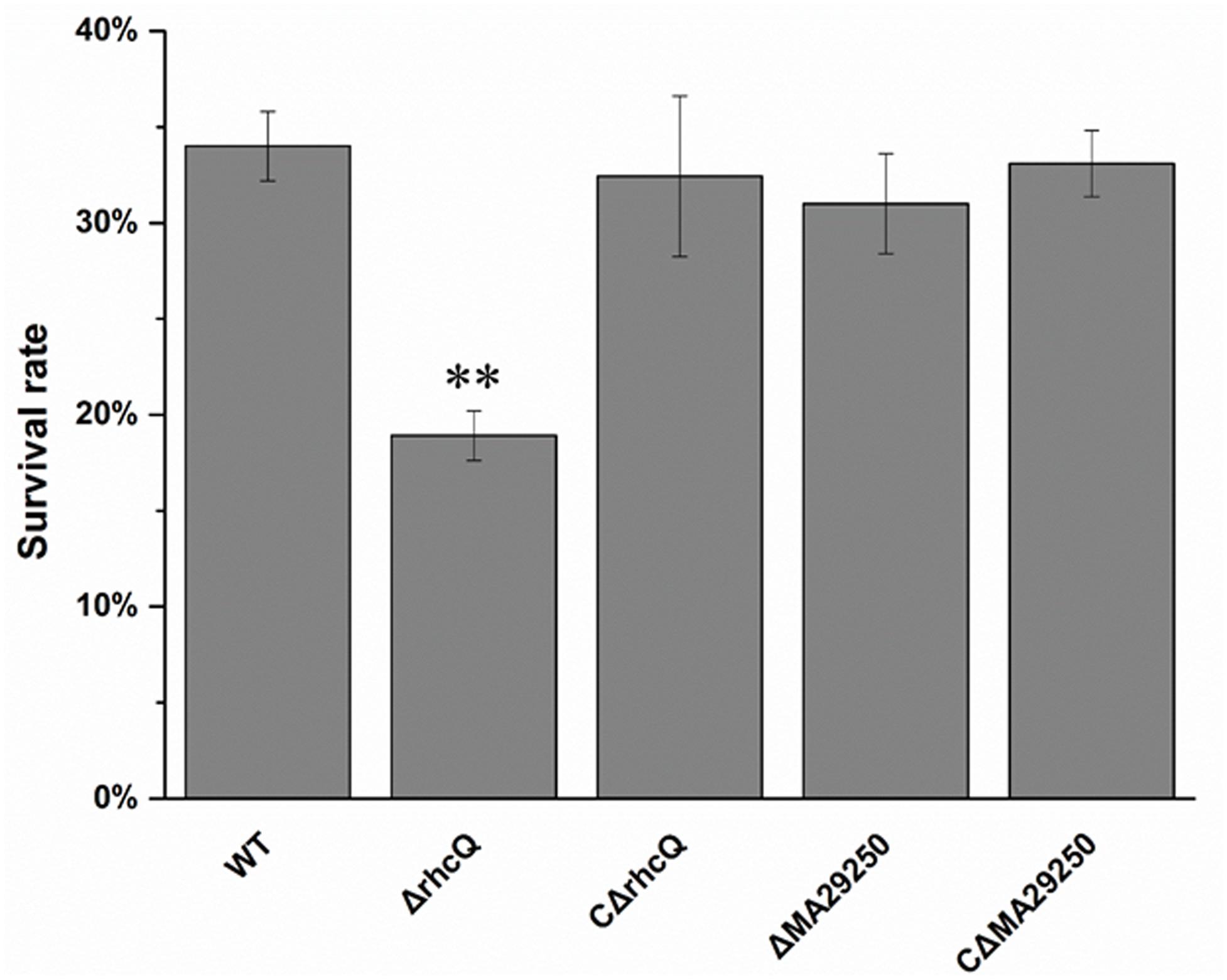
FIGURE 6. Survival rate of M. alhagi CCNWXJ12-2T under oxidative stress. Wild-type (WT), mutants (ΔrhcQ and ΔMA29250), and the complemented strains (CΔrhcQ and CΔMA29250) were grown to OD600 ≈0.8 in TY broth containing appropriate antibiotics, and then adjusted to OD600 ≈0.2. The cultures were treated with 10 mM H2O2 for 30 min or incubated without H2O2 as a control. Cell survival rates were then counted and compared to untreated cells. The mean values of three independent experiments are shown and the error bars indicate SDs. Significant differences between the WT strain and other strains are indicated (∗∗P < 0.01).
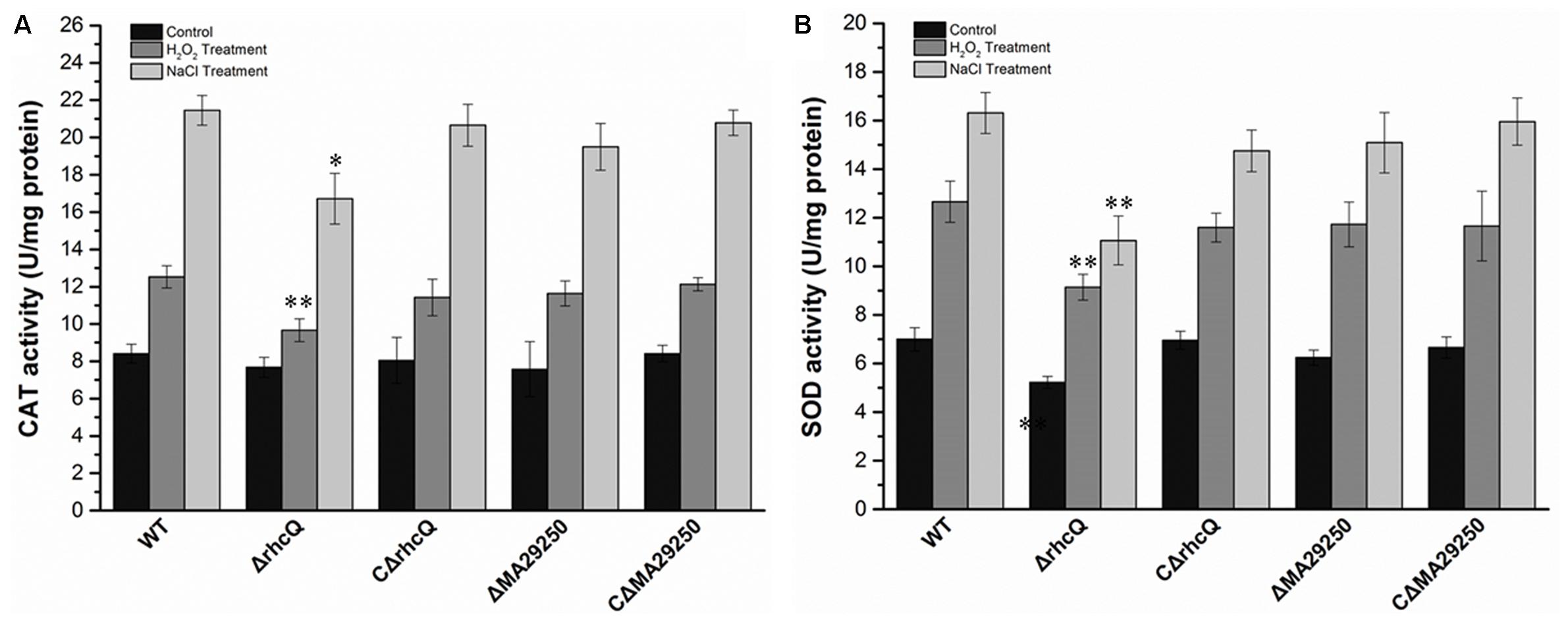
FIGURE 7. Antioxidant enzyme activities of M. alhagi CCNWXJ12-2T under oxidative and salt stress. Catalase (CAT) and superoxide dismutase (SOD) activities of the wild-type (WT), mutants (ΔrhcQ and ΔMA29250), and complemented strains (CΔrhcQ and CΔMA29250) were measured in three independent biological experiments. (A) CAT activities; (B) SOD activities. Three biological replicates were conducted and the error bars indicate SDs. Significant differences of the enzyme activities in different conditions between the WT strain and other strains are indicated (∗P < 0.05; ∗∗P < 0.01).
Discussion
Our previous transcriptome analysis of M. alhagi CCNWXJ12-2T in salt-free and high-salt conditions revealed different expression patterns of the T3SSs: T3SS1 was upregulated in high-salt conditions, while the expression of T3SS2 was downregulated (Liu et al., 2014). In the present study, phylogenetic analysis of the two T3SS core proteins, showed that T3SS1 and T3SS2 were members of subgroups α-Rhc II and α-Rhc I, respectively (Figure 1). Most T3SSs in subgroup α-Rhc I play important roles in symbiosis between the rhizobia and leguminous plants, while studies of the functions of T3SSs in subgroup α-Rhc II remain limited (Tampakaki, 2014). A former study showed that the expression of T3SS-II (belonging to α-Rhc II subgroup) in Sinorhizobium sp. NGR234 was upregulated in nodules formed with Vigna unguiculata and Leucaena leucocephala while the T3SS-II deletion mutant did not show any effect on nodule formation (Li et al., 2013). And another study of P. aeruginosa found that the osmaotic stress could induce the expression of T3SS with associated cytotoxins (Aspedon et al., 2006). In a study of S. enterica Serovar Typhimurium found that some simple salts, such as NaCl, KCl, NH4Cl, etc., could induce the assembly of SPI1-encoded T3SS and proteins secreted by T3SS (Mizusaki et al., 2008). Although the upregulation of the T3SS expression by simple salts was confirmed in these strains, the effects of T3SS on the salt tolerance were not studied. Here, we focused on the function of T3SS1, which is in subgroup α-Rhc II.
To explore the functions of T3SS1 in stress adaption by M. alhagi CCNWXJ12-2T, we first studied the promoter activities of T3SS1 on exposure to different salts and salt concentrations. β-galactosidase activities were used to reflect the promoter activities and increased with increasing concentrations of salts (Figure 3). This result encouraged us to study further the function of T3SS1 in M. alhagi CCNWXJ12-2T. Two genes were chosen to construct T3SS1-related mutants: rhcQ and MA29250. The loss of rhcQ could completely inhibit the assembly of T3SS in other bacteria (Biemans-Oldehinkel et al., 2011; Bzymek et al., 2012). Here, we hypothesized that the function of T3SS1 was completely inhibited in the ΔrhcQ mutant. The hypothetical protein encoding gene MA29250 was randomly chosen from among the hypothetical proteins in the T3SS1 gene cluster for primary tests of its function. Our results showed a difference in stress resistance of the two mutants. Strain ΔrhcQ showed more sensitivity to NaCl, LiCl, KCl, and H2O2; ΔMA29250 showed similar stress resistance to the wild-type strain, except toward KCl (Figures 4, 6). The results confirmed our hypothesis that the deletion of ΔrhcQ could completely disrupt the function of T3SS1; however, the deletion of MA29250 only partially influenced the function of T3SS1. This phenomenon promoted us to study the role of T3SS1 in resistance to KCl.
The K+ concentration in both mutants was increased compared to the wild-type (Figure 5B), indicating that T3SS1 plays a role in adjusting the intracellular K+ concentration. Meanwhile, the Na+ concentration in both mutants was also higher than that in the wild-type strain, but there was a smaller increase in mutant ΔMA29250 compared to ΔrhcQ (Figure 5A). These findings suggested that rhcQ plays a more important role in T3SS1 than MA29250, and that T3SS1 plays an important role in salt stress-resistance.
Environmental stresses such as hyperosmolality, acid, and high-salt concentrations can trigger secondary oxidative stress in bacteria (Mols and Abee, 2011). Levels of reactive oxygen species can be increased significantly when cells are grown in high-salt conditions (Koh et al., 2010; Calderon-Torres et al., 2011). Therefore, the oxidative stress resistance of the two mutants was measured. ΔrhcQ showed lower oxidative stress resistance than ΔMA29250 and the wild-type (Figure 6). This result again suggested that rhcQ plays a more important role in T3SS1 than MA29250. CAT and SOD activities in ΔrhcQ were lower than those in the wild-type and ΔMA29250 (Figure 7). Our results also showed that T3SS1 plays an important role in resistance to both salt and oxidative stress.
Author Contributions
XL, ZL, and GW made conceived and designed of the study. XL and YL conducted the laboratory work. XL and ZL carried out the data analysis and manuscript writing. All the authors read and approved the final manuscript.
Funding
This research was financially supported by the National Natural Science Foundation of China (313701427 and 31270012).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00263/full#supplementary-material
Footnotes
References
Aspedon, A., Palmer, K., and Whiteley, M. (2006). Microarray analysis of the osmotic stress response in Pseudomonas aeruginosa. J. Bacteriol. 188, 2721–2725. doi: 10.1128/JB.188.7.2721-2725.2006
Biemans-Oldehinkel, E., Sal-Man, N., Deng, W., Foster, L. J., and Finlay, B. B. (2011). Quantitative proteomic analysis reveals formation of an EscL-EscQ-EscN Type III complex in enteropathogenic Escherichia coli. J. Bacteriol. 193, 5514–5519. doi: 10.1128/JB.05235-11
Buttner, D., and He, S. Y. (2009). Type III protein secretion in plant pathogenic bacteria. Plant Physiol. 150, 1656–1664. doi: 10.1104/pp.109.139089
Bzymek, K. P., Hamaoka, B. Y., and Ghosh, P. (2012). Two translation products of Yersinia yscQ assemble to form a complex essential to Type III secretion. Biochemistry 51, 1669–1677. doi: 10.1021/bi201792p
Calderon-Torres, M., Castro, D. E., Montero, P., and Pena, A. (2011). DhARO4 induction and tyrosine nitration in response to reactive radicals generated by salt stress in Debaryomyces hansenii. Yeast 28, 733–746. doi: 10.1002/yea.1903
Chen, W. M., Zhu, W. F., Bontemps, C., Young, J. P., and Wei, G. H. (2010). Mesorhizobium alhagi sp. nov, isolated from wild Alhagi sparsifolia in north-western China. Int. J. Syst. Evol. Microbiol. 60(Pt 4), 958–962. doi: 10.1099/ijs.0.014043-0
Correa, V. R., Majerczak, D. R., Ammar, E. D., Merighi, M., Pratt, R. C., Hogenhout, S. A., et al. (2012). The bacterium Pantoea stewartii uses two different Type III secretion systems to colonize its plant host and insect vector. Appl. Environ. Microbiol. 78, 6327–6336. doi: 10.1128/Aem.00892-12
Ditta, G., Stanfield, S., Corbin, D., and Helinski, D. R. (1980). Broad host range DNA cloning system for gram-negative bacteria: construction of a gene bank of Rhizobium meliloti. Proc. Natl. Acad. Sci. U.S.A. 77, 7347–7351. doi: 10.1073/pnas.77.12.7347
Galán, J. E., and Collmer, A. (1999). Type III secretion machines: bacterial devices for protein delivery into host cells. Science 284, 1322–1328. doi: 10.1126/science.284.5418.1322
Galán, J. E., Lara-Tejero, M., Marlovits, T. C., and Wagner, S. (2014). Bacterial Type III secretion systems: specialized nanomachines for protein delivery into target cells. Annu. Rev. Microbiol. 68, 415–438. doi: 10.1146/annurev-micro-092412-155725
Haller, J. C., Carlson, S., Pederson, K. J., and Pierson, D. E. (2000). A chromosomally encoded Type III secretion pathway in Yersinia enterocolitica is important in virulence. Mol. Microbiol. 36, 1436–1446. doi: 10.1046/j.1365-2958.2000.01964.x
Hanahan, D. (1983). Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 166, 557–580. doi: 10.1016/S0022-2836(83)80284-8
Koh, M. J., Lee, H. S., Rhee, J. E., and Choi, S. H. (2010). Evidence that Vibrio vulnificus AhpC2 is essential for survival under high salinity by modulating intracellular level of ROS. J. Microbiol. 48, 129–133. doi: 10.1007/s12275-009-0227-2
Kovach, M. E., Elzer, P. H., Hill, D. S., Robertson, G. T., Farris, M. A., Roop, R. M., et al. (1995). 4 new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 166, 175–176. doi: 10.1016/0378-1119(95)00584-1
Krause, A., Doerfel, A., and Gottfert, M. (2002). Mutational and transcriptional analysis of the Type III secretion system of Bradyrhizobium japonicum. Mol. Plant Microbe Interact. 15, 1228–1235. doi: 10.1094/Mpmi.2002.15.12.1228
Li, Y., Tian, C. F., Chen, W. F., Wang, L., Sui, X. H., and Chen, W. X. (2013). High-resolution transcriptomic analyses of Sinorhizobium sp. NGR234 bacteroids in determinate nodules of Vigna unguiculata and indeterminate nodules of Leucaena leucocephala. PLoS One 8:e70531. doi: 10.1371/journal.pone.0070531
Liu, X., Luo, Y., Li, Z., and Wei, G. (2016). Functional analysis of PrkA - a putative serine protein kinase from Mesorhizobium alhagi CCNWXJ12-2 - in stress resistance. BMC Microbiol. 16:227. doi: 10.1186/s12866-016-0849-6
Liu, X. D., Luo, Y. T., Mohamed, O. A., Liu, D. Y., and Wei, G. H. (2014). Global transcriptome analysis of Mesorhizobium alhagi CCNWXJ12-2 under salt stress. BMC Microbiol. 14:319. doi: 10.1186/s12866-014-0319-y.
Lorio, J. C., Kim, W. S., and Krishnan, H. B. (2004). NopB, a soybean cultivar-specificity protein from Sinorhizobium fredii USDA257, is a Type III secreted protein. Mol. Plant Microbe Interact. 17, 1259–1268. doi: 10.1094/Mpmi.2004.17.11.1259
Mizusaki, H., Takaya, A., Yamamoto, T., and Aizawa, S. (2008). Signal pathway in salt-activated expression of the Salmonella pathogenicity island 1 Type III secretion system in Salmonella enterica serovar Typhimurium. J. Bacteriol. 190, 4624–4631. doi: 10.1128/JB.01957-07
Mols, M., and Abee, T. (2011). Primary and secondary oxidative stress in Bacillus. Environ. Microbiol. 13, 1387–1394. doi: 10.1111/j.1462-2920.2011.02433.x
Rolhion, N., Furniss, R. C. D., Grabe, G., Ryan, A., Liu, M., Matthews, S. A., et al. (2016). Inhibition of nuclear transport of NF-kappa B p65 by the Salmonella Type III secretion system effector SpvD. PLoS Pathog. 12:e1005653. doi: 10.1371/journal.ppat.1005653
Sanchez, C., Mercante, V., Babuin, M. F., and Lepek, V. C. (2012). Dual effect of Mesorhizobium loti T3SS functionality on the symbiotic process. FEMS Microbiol. Lett. 330, 148–156. doi: 10.1111/j.1574-6968.2012.02545.x
Schafer, A., Tauch, A., Jager, W., Kalinowski, J., Thierbach, G., and Puhler, A. (1994). Small mobilizable multipurpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19 - selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 145, 69–73. doi: 10.1016/0378-1119(94)90324-7
Silver, A. C., Kikuchi, Y., Fadl, A. A., Sha, J., Chopra, A. K., and Graf, J. (2007). Interaction between innate immune cells and a bacterial Type III secretion system in mutualistic and pathogenic associations. Proc. Natl. Acad. Sci. U.S.A. 104, 9481–9486. doi: 10.1073/pnas.0700286104
Tampakaki, A. P. (2014). Commonalities and differences of T3SSs in rhizobia and plant pathogenic bacteria. Front. Plant Sci. 5:114. doi: 10.3389/fpls.2014.00114
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739. doi: 10.1093/molbev/msr121
Viprey, V., Del Greco, A., Golinowski, W., Broughton, W. J., and Perret, X. (1998). Symbiotic implications of Type III protein secretion machinery in Rhizobium. Mol. Microbiol. 28, 1381–1389. doi: 10.1046/j.1365-2958.1998.00920.x
Wang, S. H., Liu, X., Xu, X., Yang, D. H., Wang, D., Han, X. G., et al. (2016). Escherichia coli Type III secretion system 2 ATPase EivC is involved in the motility and virulence of avian pathogenic Escherichia coli. Front. Microbiol. 7:1387. doi: 10.3389/fmicb.2016.01387
Watanabe, T., Seno, M., Sasada, R., and Igarashi, K. (1990). Molecular characterization of recombinant human acidic fibroblast growth factor produced in E. coli: comparative studies with human basic fibroblast growth factor. Mol. Endocrinol. 4, 869–879. doi: 10.1210/mend-4-6-869
Zhang, X., and Bremer, H. (1995). Control of the Escherichia coli rrnB P1 promoter strength by ppGpp. J. Biol. Chem. 270, 11181–11189. doi: 10.1074/jbc.270.19.11181
Zhou, M. L., Chen, W. M., Chen, H. Y., and Wei, G. H. (2012). Draft genome sequence of Mesorhizobium alhagi CCNWXJ12-2(Tau), a novel salt-resistant species isolated from the Desert of Northwestern China. J. Bacteriol. 194, 1261–1262. doi: 10.1128/Jb.06635-11
Keywords: Mesorhizobium alhagi, type III secretion system, stress adaption, β-galactosidase assay, Na+ content measurement
Citation: Liu X, Luo Y, Li Z and Wei G (2018) Functional Analysis of a Putative Type III Secretion System in Stress Adaption by Mesorhizobium alhagi CCNWXJ12-2T. Front. Microbiol. 9:263. doi: 10.3389/fmicb.2018.00263
Received: 22 November 2017; Accepted: 02 February 2018;
Published: 20 February 2018.
Edited by:
Manuel Kleiner, North Carolina State University, United StatesReviewed by:
Michael Göttfert, Technische Universität Dresden, GermanyMuhammad Yahya Khan, University of Agriculture Faisalabad, Pakistan
Copyright © 2018 Liu, Luo, Li and Wei. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gehong Wei, d2VpZ2Vob25nQG53c3VhZi5lZHUuY24= Zhefei Li, bGl6aGVmZWlAaG90bWFpbC5jb20=
†These authors have contributed equally to this work.