 María-Efigenia Álvarez-Cao
María-Efigenia Álvarez-Cao María-Esperanza Cerdán
María-Esperanza Cerdán María-Isabel González-Siso
María-Isabel González-Siso Manuel Becerra
Manuel Becerra- Grupo EXPRELA, Centro de Investigacións Científicas Avanzadas (CICA), Facultade de Ciencias, Universidade da Coruña, A Coruña, Spain
Molasses are sub-products of the sugar industry, rich in sucrose and containing other sugars like raffinose, glucose, and fructose. Alpha-galactosidases (EC. 3.2.1.22) catalyze the hydrolysis of alpha-(1,6) bonds of galactose residues in galacto-oligosaccharides (melibiose, raffinose, and stachyose) and complex galactomannans. Alpha-galactosidases have important applications, mainly in the food industry but also in the pharmaceutical and bioenergy sectors. However, the cost of the enzyme limits the profitability of most of these applications. The use of cheap sub-products, such as molasses, as substrates for production of alpha-galactosidases, reduces the cost of the enzymes and contributes to the circular economy. Alpha-galactosidase is a specially indicated bioproduct since, at the same time, it allows to use the raffinose present in molasses. This work describes the development of a two-step system for the valuation of beet molasses, based on their use as substrate for alpha-galactosidase and bioethanol production by Saccharomyces cerevisiae. Since this yeast secretes high amounts of invertase, to avoid congest the secretory route and to facilitate alpha-galactosidase purification from the culture medium, a mutant in the SUC2 gene (encoding invertase) was constructed. After a statistical optimization of culture conditions, this mutant yielded a very high rate of molasses bioconversion to alpha-galactosidase. In the second step, the SUC2 wild type yeast strain fermented the remaining sucrose to ethanol. A procedure to recycle the yeast biomass, by using it as nitrogen source to supplement molasses, was also developed.
Introduction
Molasses are viscous dark sub-products resulting from the sugar-making industry; they are rich in sucrose and contain lower amounts of other sugars like raffinose, glucose, and fructose. Due to the high sucrose content (about 50% by dry weight), at present molasses serve mostly as substrate for the industrial production of bioethanol by the yeast Saccharomyces cerevisiae (Akbas and Stark, 2016). The use of food industry wastes as sustainable substrates for the microbial synthesis of other biotechnological products besides ethanol, such as enzymes and other active ingredients, is increasingly gaining field in the context of the circular economy (da Silva, 2016). From this point of view, molasses provides a good carbon source for microbial cell growth (Santos et al., 2010), and the yeast biomass itself, generated in the bioprocess, may be used after a simple treatment (Ferreira et al., 2010) as source of nitrogen and B-group vitamins to complement molasses.
Saccharomyces cerevisiae is undoubtedly the best performing microorganism for molasses fermentation to bioethanol (Akbas et al., 2014). This yeast is also a widely employed host for heterologous protein production and secretion (Nielsen, 2013). However, there are no reports hitherto on the use of molasses as substrate with the aim of heterologous protein secretion by S. cerevisiae.
Alpha-galactosidases (EC. 3.2.1.22) catalyze the hydrolysis of alpha-(1,6) bonds of galactose residues in galacto-oligosaccharides (melibiose, raffinose, and stachyose) and complex galactomannans. Some alpha-galactosidases can also synthesize oligosaccharides by transglycosylation reactions under substrate supersaturation conditions (Spangenberg et al., 2000). Indeed alpha-galactosidases show a great variety of uses, mainly in the food industry such us improvement of separation of sucrose from beet (Linden, 1982), reduction of the content of non-digestible oligosaccharides of legume-derived food products (Katrolia et al., 2012), obtention of the low-calorie sweetener tagatose (Kim et al., 2009), improvement of rheological properties of galactomannans (Dey et al., 1993), and synthesis of prebiotics (Dai et al., 2018), but also in other sectors like pharmaceutical (Arends et al., 2017) and bioenergy (Rodrigues-Dutra et al., 2017). S. cerevisiae alpha-galactosidase (ScAGal), encoded by the MEL1 gene (GeneBank X03102), is a highly glycosylated 471-amino acid extracellular protein, and the crystal structures of the complexes with melibiose and raffinose have been reported (Fernández-Leiro et al., 2010).
The cost of the enzyme actually limits the profitability of most of the above cited applications. The use of cheap sub-products, such as molasses and whey, as substrates for production of ScAGal, was previously reported and might favor the economy of the processes (Álvarez-Cao et al., 2018). The aim of this work is to develop a system for the valuation of beet molasses, based on their use as substrate for ScAGal and bioethanol production by S. cerevisiae in two-steps. A first step using an invertase-deficient mutant (transformed with a ScAGal over-expressing plasmid), which not only favors the secretion-purification of the enzyme due to the absence of invertase (Liljeström et al., 1991), but also increases the yield of ScAGal production, because the mutant uses the sugars present at low concentration in the molasses but not sucrose, and then the metabolism is preferentially respiratory under aerobic conditions. In the second step, the invertase wild-type strain is used to convert the sucrose to ethanol under yet established fermentation conditions.
With the engineered strains and two-step system here developed, a high rate of molasses bioconversion to alpha-galactosidase was obtained (culture conditions were statistically optimized), and sucrose was efficiently converted to ethanol. A procedure to recycle the yeast biomass, by using it as nitrogen source to supplement molasses, was also developed.
Materials and Methods
Microorganisms, Expression Vectors, and Culture Media
Saccharomyces cerevisiae BJ3505 [pep4::HIS3, prb-Δ1.6R HIS3, lys2-208, trp1-Δ101, ura 3-52, gal2, can1] (Eastman Kodak Company) was selected to construct a mutant depleted of invertase activity to be used as host for heterologous protein expression. Escherichia coli XL1-Blue [recA1 endA1 gyrA96 thi-1 hsdR17 supE44 relA1 lac [F'proAB lacIqZDM15 Tn10 (Tetr)]] (Stratagene Cloning Systems) was employed for standard DNA recombinant techniques (Ausubel et al., 2003).
The plasmids YEpMEL1 [ampr ori 2 μ MEL1 TRP1] and YEpMEL1His [ampr ori 2 μ MEL1His TRP1] (Fernández-Leiro, 2011), bearing the MEL1 gene that encodes ScAGal, were chosen as expression vectors. The vector YEpFLAG-1 (Eastman Kodak Company) was used as control of the expression system.
LB [1% (w/v) tryptone, 0.5% (w/v) yeast extract, 0.5% (w/v) NaCl], and YPD [1% (w/v) yeast extract, 2% (w/v) peptone, 2% (w/v) glucose] were used as culture media for growth and maintenance of bacteria and yeasts, respectively. LB was supplemented with 100 mg/L ampicillin (Sigma Aldrich) for the propagation of plasmids in bacteria. A complete medium without uracil (CM-Ura) or tryptophan (CM-Trp) (Zitomer and Hall, 1976) was used for selection in S. cerevisiae. Two percent (w/v) bacteriological agar was added to solid media. A modified YPHSM medium and beet molasses (provided by AB Azucarera Iberia, Spain) based media were used for ScAGal production. Molasses were diluted in distilled water ratio 1:1 (v/v), to facilitate handling, and centrifuged at 10,000 rpm for 15 min to remove solid impurities. Ampicillin was filtered through 0.22 μm membrane (Sartorius AG) and the rest of media components were sterilized by autoclave at 121°C for 20 min. The composition of the media used in this work is shown in Table 1.

Table 1. Summary of the strategy of study of media and culture conditions on the production of ScAGal.
Construction of an Invertase Defective Mutant
The SUC2 gene was deleted in the S. cerevisiae strain BJ3505 using the integrative cassette suc2Δ(266-388)URA3 that contains the suc2Δ(266-388) deletion and the URA3 gene as selection marker (Figure 1). All oligonucleotides used as primers in this work are showed in Table S1.
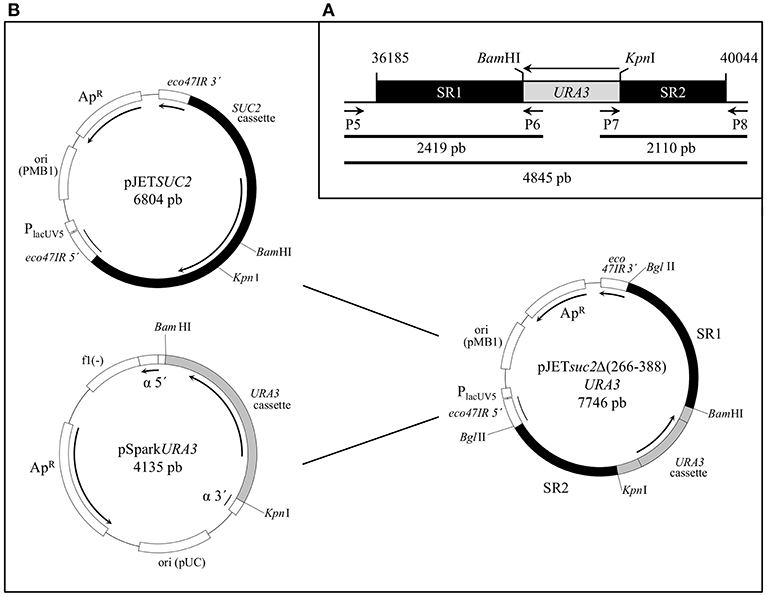
Figure 1. Construction of S. cerevisiae strain BJ3505Δsuc2. (A) The sequence [36,185–40,044] bp of chromosome 9 of S. cerevisiae (Gene ID: 854644; NC_001141.2) containing the SUC2 ORF was chosen to carry out the suc2Δ(266–388) deletion by replacement with the URA3 cassette between sites BamHI and KpnI. SR1 and SR2 are the chromosomal recombination sequences and the primers pairs P5-P6, P7-P8, and P5-P8 are used to obtain the fragments 2,419, 2,110, and 4,845 bp, respectively to check the chromosomal integration. (B) Physical maps of the plasmids constructed to generate strain BJ3505Δsuc2: the URA3 cassette from pSparkURA3 was cloned between the BamHI-KpnI sites of pJETSUC2 to obtain pJETsuc2Δ(266–388)URA3, which was digested with BglII to release the suc2Δ(266–388)URA3 cassette that was integrated through SR1 and SR2 sequences into genome of the origin strain.
Construction of SUC2 and URA3 Cassettes
The ORF of the SUC2 gene was identified by means of the tool BLAST at GeneBank (http://www.ncbi.nlm.nih.gov/) and the homology among the different S. cerevisiae strains in the database. The nucleotide sequence [36185–40044] in the chromosome 9 of S. cerevisiae S288c (Gene ID: 854644; NC_001141.2) was used and the design of the integrative cassette is showed in Figure 1A. A region longer than 1 Kb upstream and downstream of the SUC2 ORF was used to compete with the marker ura3-52 of the strain BJ3505. The fragment of 3,831 bp length corresponding to the SUC2 cassette was amplified with the primer pairs P1–P2 (Table S1) from genomic DNA of the strain BJ3505 as template, by the protocol described in Hoffman (2001). A fragment of 1,308 bp flanked by the restriction sites BamHI and KpnI was amplified from the vector YEplac195 (Gietz and Sugino, 1988) using the primer pairs P3–P4 (Table S1) to obtain the cassette URA3 with its promoter and terminator regions. PCRs were performed using Phusion High-Fidelity DNA Polymerase (Thermo Fisher Scientific).
Construction of Plasmids pJETSUC2, pSparkURA3, and pJETsuc2Δ(266–388)URA3
The PCR products obtained from amplification of SUC2 and URA3 cassettes were cloned into vectors pJET1.2/blunt (CloneJET PCR Cloning Kit, Thermo Fisher Scientific) and pSpark IV (pSpark DNA Cloning System, Canvax Biotech) respectively, following the protocol recommended by each supplier, to create the plasmids pJETSUC2 and pSparkURA3. The BamHI-KpnI fragment of 1,308 bp from pSparkURA3 was cloned between the BamHI-KpnI sites of pJETSUC2 into chemically competent XL1-Blue cells to construct the plasmid pJETsuc2Δ(266–388)URA3 with the recombination sequences SR1 (1,962 bp) and SR2 (1,503 bp) that allowed the chromosomal integration (Figure 1B). All the plasmids generated in this work were propagated and extracted from the bacterial cells using GeneJET Plasmid Miniprep Kit (Thermo Fisher Scientific). The inserts were identified by PCR of transformant colonies using DreamTaq polymerase (Thermo Fisher Scientific) and restriction analysis, and correct sequences confirmed by sequencing (Servizos de Apoio á Investigación, Universidade da Coruña).
Integration of the suc2Δ(266–388)URA3 Cassette in the BJ3505 Genome
The plasmid pJETsuc2Δ(266–388)URA3 was digested with BglII to separate the cassette suc2Δ(266–388)URA3 and further cloning in BJ3505 competent cells transformed by the lithium acetate method (Ito et al., 1983). Chromosomal integration succeeded by two homologous recombination points provided by SR1 and SR2 sequences. Recombinants were selected by growth in CM-Ura and the invertase-deficient strain, named BJ3505ΔSuc2, was verified by functional and PCR analysis using DreamTaq polymerase (Thermo Fisher Scientific) (Figure 2).
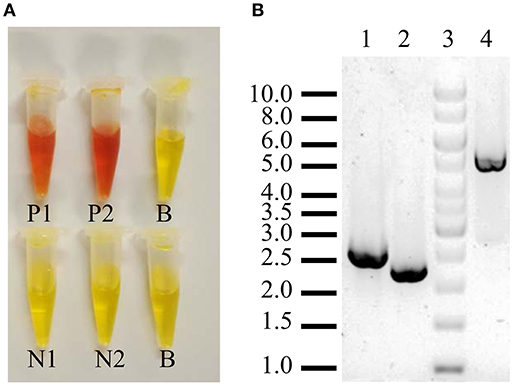
Figure 2. Verification of S. cerevisiae strain BJ3505Δsuc2. (A) Negative (yellow color) and positive (orange color) functional assay of extracellular (1) and intracellular invertase activity (2) as described in section Analytical Methods. N, strain BJ3505Δsuc2; P, strain BJ3505 (parental); B, blank. (B) PCR analysis of the “upstream” (2,419 bp, lane 1), “downstream” (2,110 bp, lane2), and between them (4,845 bp, lane 4) of the SUC2 gene that codes for the invertase. Lane 3, GeneRuler 1Kb DNA Ladder (Thermo Fisher Scientific).
Transformation and Inoculum Preparation for ScAGal Production Cultures
The strains BJ3505ΔSuc2 and BJ3505 were transformed with plasmids YEpFLAG-1, YEpMEL1, and YEpMEL1His, by a modification of the lithium acetate method (Chen et al., 1992). Transformants were selected on solid CM-Trp after 48 h at 30°C. A single colony was taken to prepare a liquid CM-Trp pre-culture up to an optical density at 600 nm (OD600) of 5 (stationary phase) at 30°C and 250 rpm for 72 h, which was used to inoculate up to units OD600 corresponding to each production culture tested, as described above.
Influence of Media and Culture Conditions on ScAGal Production
Preliminary studies on cell growth determined by optical density at 600 nm (Ausubel et al., 2003) and ScAGal production from strains BJ3505Δsuc2 and BJ3505 were performed with PR, YR, and YRaut media (Table 1). Beet molasses were added at the desired concentration, determined as total sugars percentage, and supplemented with yeast extract or peptone to provide nitrogen source.
Yeast extract coming from biomass of ended-cultures was prepared according to a modification of the autolysis method described by Belem et al. (1997). Yeast cells were centrifuged (5,000 rpm, 15 min, 4°C), resuspended 1:2 (w/v) in 0.1 M KH2PO4 (pH 6.5), and autolyzed at 50°C and 150 rpm for 30 h. Supernatant was recovered by centrifugation (10,000 rpm, 15 min), cell debris was resuspended 1:10 in the same buffer and disintegrated by sonication (Sonics Vibra cell) for 10 min at 7 s intervals to facilitate the release of the remaining soluble cellular material in a second supernatant recovered by centrifugation. Both supernatants were mixed and added at 0.5% (v/v) to supplement the culture media. The RNA released from the final autolysate was analyzed by 1.5% (w/v) agarose gel electrophoresis using 1% (w/v) commercial yeast extract as control (Figure S1A).
Cultures were inoculated, from the pre-cultures of transformed strains described above, by triplicate in flasks containing 20% volume of medium, and incubated at 30°C and 250 rpm. A modified YPHSM medium (Table 1) inoculated to obtain an initial OD600 of 0.5 was used to statistically compare the expression systems YEpMEL1 and YEpMEL1His by analysis of variance (ANOVA) with 95% confidence intervals. Profiles of sugar consumption and products formation in the molasses-based media YR and PR were initially determined from cultures inoculated at OD600 of 2. Cultures in YR and YRaut media inoculated at OD600 of 4 were used to compare the assimilation of the carbon source with commercial yeast extract and autolyzed biomass, respectively. Growth kinetics and substrate to products conversion was evaluated from YR cultures inoculated at OD600 of 4. In this case, when the consumption of sugar was negligible, half of each culture volume was separated while the other half was maintained in the same conditions. Under sterile conditions, the volume separated of each culture was centrifuged (5,000 rpm, 10 min, 4°C), the supernatant was refreshed with 1% (w/v) yeast extract and inoculated at the same initial OD600 with cells coming from the yeast pellet. Cultures were stopped after 84 h and the last step was repeated but only with the BJ3505Δsuc2 cultures and using as inoculum the BJ3505 at the same initial OD600. Samples were taken at different incubation times to determine biomass, residual sugars, ethanol, and enzyme activities (invertase and alpha-galactosidase). A summary of the strategy of study of media and culture conditions is shown in Table 1.
Microscopy
An optical microscope (Nikon Eclipse 50) was used to observe directly, without fixation, the cells from the collected culture samples. For observation under transmission electron microscopy (TEM) (Servizos de Apoio á Investigación, Universidade da Coruña), the cells were fixed in 100 mM sodium cacodylate buffer pH 7.2 as described in Bauer et al. (2001).
Optimization of ScAGal Production by the Surface Response Methodology
Statistical optimization of beet molasses based cultures for ScAGal production was performed using the response surface methodology (RSM) with the strain BJ3505Δsuc2/YEpMEL1. The experimental factors selected as independent variables were 4: concentration of molasses and yeast extract, inoculum size, and culture time. The dependent variable or response was extracellular alpha-galactosidase activity. A central composite design (CCD) was applied to study the effects and interactions among the four factors at five different levels. Their coded values were –α, −1, 0, +1, +α, being α = 2k/4, k the number of independent variables and 0 the central point. Table 2 shows the levels and real values of the factors, calculated according to the equation described by de Faria et al. (2013). The RS was adjusted to a polynomic second order equation that correlates the measured response with the independent variables (Dilipkumar et al., 2014), and the optimum value of the CCD was obtained solving the regression equation and analyzing the surface response contour plots. An ANOVA with 95% confidence intervals was performed to determine the significance of the effects.
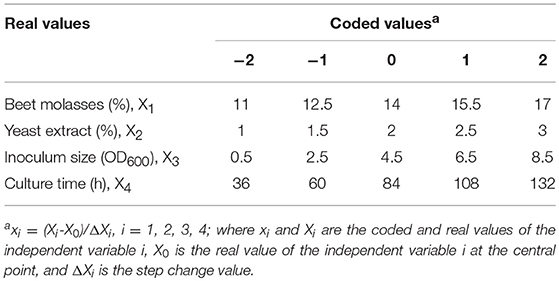
Table 2. Experimental domain and codification of the independent variables in the CCD to the optimization of the ScAGal production by BJ3505Δsuc2/YEpMEL1.
Bioreactor Operation
Bioreactor cultures were run in a Biostat-MD (Braun-Biotech) 2 L fermentor, 1 L working volume, with pH control to evaluate its effect on ScAGal production. For this, the fermentations were carried out without and with adjustment to pH 6 during the course of the culture. Medium composition and inoculum size were selected from the results of the CCD described above. Bioreactor was sterilized at 121°C for 30 min, and then aseptically inoculated and supplemented with 300 mg/L ampicillin. Culture conditions were 30°C, 2 L/min air flow and 300 rpm, for 120 h (Ausubel et al., 2003). Sterilized 1 M NaOH or 1 M HCl were added as needed to adjust pH. Samples were taken at regular time intervals to determine yeast biomass, extracellular alpha-galactosidase activity, and conversion of substrates into secondary metabolites. Plasmid stability was determined as percentage of viable Trp+ colonies, after seeding diluted samples of the culture on Petri dishes with CM and CM-Trp and counting colony forming units by the following equation,
Analytical Methods
Molasses Sugar and Protein Content
The sugars present in the diluted molasses used to prepare the culture media were identified and quantified by High-Performance Liquid Chromatography (HPLC) using an external standard formed by known concentrations of raffinose, sucrose, glucose, and fructose ranging 4–0.06 mg/mL. An estimate of the nitrogen fraction in the diluted molasses according to the protein content was measured by the Bradford's method using DC Protein Assay Kit (Bio Rad) and bovine serum albumin as standard.
Biomass, Residual Sugars, and Ethanol Determination
Cell growth in shaken flasks was measured as units of OD600 with an UV-Visible espectrophotometer (Biospectrometer Kinetic Eppendorf) to express the biomass generated. Cell-free medium was used to measure ethanol, with the enzymatic assay Ethanol UV method (NZYTtech), and residual sugar. The latter was quantified indirectly using the DNS method (Miller, 1959) by means of the analysis of reducing sugars content in the samples before and after hydrolysis at 95°C for 5 min with 18.5% (w/v) HCl followed by neutralization with 25% (w/v) NaOH. For cultures in bioreactor, biomass was quantified as dry weight from 5 mL samples, after being centrifuged at 5,000 rpm for 5 min, washed with distilled water, centrifuged again, and dried at 105°C until constant weight. The supernatant was used to measure sugar consumption, ethanol, and other metabolites formation by HPLC using an external standard composed by known concentrations of raffinose, sucrose, galactose, glucose, fructose, glycerol, and ethanol ranging 4–0.06 mg/mL.
HPLC Analysis
Sugar Pack Waters (6.5 × 300 mm) column and Refractive Index Detector (Cienytech) were used. Samples were previously clarified with cartridges HyperSep Silica SPE Column (Fisher Scientific) to avoid interferences due to pigments and other impurities coming from molasses. Column temperature 80°C, detector temperature 37°C, and sensitivity 32 were the running conditions using as mobile phase 100 μM EDTA-Calcium (Sigma Aldrich) at flow rate of 0.5 mL/min. Eluted compounds were identified and quantified using sorbitol (1 mg/mL) as internal standard, since it does not interfere with the retention times of the compounds under study (Xu et al., 2015).
Enzyme Activity Assay
Extracellular alpha-galactosidase activity was measured by the method of Ryan et al. (1998), incubating the samples with a 10 mM solution of the synthetic substrate p-nitrophenyl-α-galactose (Sigma Aldrich) in McIlvaine buffer (pH 4) at 40°C. At two consecutive time intervals, the reaction was stopped with 0.5 M Na2CO3 and released p-nitrophenol was measured at 400 nm (molar extinction coefficient: 18.20 mmol−1.cm−1). Invertase activity, extracellular, and intracellular was measured from supernatant and sediment of culture samples, respectively, after permeabilization with 15% (v/v) chloroform and 0.01% (w/v) SDS. The samples were incubated with 100 mM sucrose in 50 mM acetate buffer (pH 5) for 10 min at 40°C, and reducing sugars were determined by the DNS method (Miller, 1959). One enzyme unit (U) is defined as the amount of enzyme that releases one μmol of product per minute under assay conditions.
Statistical Analysis
Statistical data treatment was done using the StatGraphics Centurion XVI package and graphics construction was performed with the program SigmaPlot 12.0. Significance of data was evaluated with Student's t-test and results were considered significant for p ≤ 0.05.
Results
Construction of an Invertase Defective Mutant
The strain BJ3505Δsuc2 was constructed by the procedure described in the Materials and Methods section. The correctness of the mutation was verified by functional measurement of invertase activity that was fully depleted in the mutant (Figure 2A) and by PCR analysis (Figure 2B, see also Figure 1A) that showed bands of the expected size in function of the chromosomal integration.
Approach to a Medium for ScAGal Production Based on Beet Molasses
Cultures in YPHSM of the strains BJ3505Δsuc2 and corresponding wild type BJ3505, transformed with the plasmids YEpMEL1His or YEpMEL1, with and without affinity purification tag, respectively, were performed to select the best ScAGal expression system. YPHSM is the recommended medium for heterologous protein expression with the YEpFLAG-1 system (Eastman Kodak Company) that was used to construct the plasmids (Fernández-Leiro, 2011). Growth was similar in the four cases (OD600 = 90 ± 10; data not shown), and maximum extracellular alpha-galactosidase activity was similar for the two strains but dependent on the plasmids, 43 and 25 U/mL for YEpMEL1 and YEpMEL1His, respectively, after 216 h of incubation (Figure 3). Although the statistical analysis shows that there is not a direct correlation between ScAGal secretion and lack of invertase in glucose (YPHSM) medium (Table S2), the advantage of obtaining alpha-galactosidase preparations not contaminated with invertase is clear. Therefore, BJ3505Δsuc2/YEpMEL1 was selected as an improved ScAGal expression system for the following experiments.
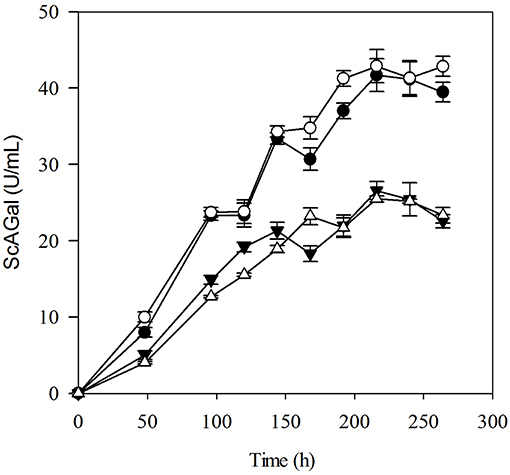
Figure 3. Time course of extracellular alpha-galactosidase activity using a conventional culture medium. Cultures in the YPHSM medium were inoculated to an OD600 of 0.5 from pre-cultures of strains BJ3505Δsuc2 (empty symbol) and BJ3505 (full symbol) transformed with the plasmids YEpMEL1 (circle), and YEpMEL1His (triangle).
Raw molasses used in this work contained 59.7% sucrose, 2.9% raffinose, 2.4% fructose, 1.2% glucose, and 0.3% nitrogen fraction. Beet molasses are limited in biotin (vitamin B7) and other sources of nitrogen, and yeast biomass can be valuable for using as nitrogen and vitamins supplement for microbial growth (Ferreira et al., 2010). Then, the effects on sugar consumption and ScAGal extracellular production of molasses supplementation with yeast extract and peptone using the YR and PR media, respectively, were assayed and are shown in Figure 4. Results prove that, as expected due to the Δsuc2 mutation, the strain BJ3505Δsuc2 uses only 10–20% of the sugars provided by the media, while the strain BJ3505 (with intact SUC2 gene) uses the 90% at the same culture time. An increase of sugar consumption rate was observed for strains transformed with YEpMEL1 due to alpha-galactosidase activity (Figure 4A). From Figure 4B, it is inferred that in YR and PR media, BJ3505Δsuc2/YEpMEL1 produces more extracellular ScAGal than BJ3505/YEpMEL1 does, i.e., indeed invertase depletion favors ScAGal secretion in sucrose-rich medium, and that yeast extract can be used as supplement instead of peptone, since ScAGal production by BJ3505Δsuc2/YEpMEL1 is quite similar with both supplements. Furthermore, the recycling of yeast biomass as molasses supplement was analyzed and it was found that the autolysis treatment provides a source rich in low molecular weight nucleotides, as is well-documented (Belem et al., 1997). Although the results also confirmed very close values of sugar consumption and extracellular ScAGal production (Figure S1B), we continued to use commercial yeast extract for the following experiments, as it is more standard and reproducible among laboratories.
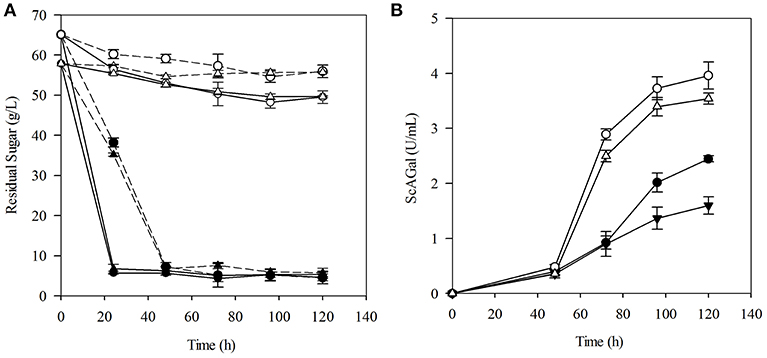
Figure 4. Evaluation of culture media based on beet molasses for ScAGal production. Time course of residual sugar (A) and extracellular alpha-galactosidase activity (B) by BJ3505Δsuc2 (empty symbol) and BJ3505 (full symbol) in culture media YR (triangle) and PR (circle). The solid and dashed lines correspond to the strains transformed with the plasmid YEpMEL1 and YEpFLAG-1, respectively. The media YR and PR were prepared with 8% beet molasses supplemented with 1% yeast extract or 2% peptone and the cultures were inoculated to OD600 = 2.
Figure 5 shows culture profiles of BJ3505Δsuc2/YEpMEL1 and the corresponding wild type BJ3505/YEpMEL1 in 8% beet molasses supplemented with 1% yeast extract. It is observed that both strains reach similar growth levels but while BJ3505/YEpMEL1 consumed 88% sugars (native invertase hydrolyzed sucrose) and produced 13 g/L of ethanol, BJ3505Δsuc2/YEpMEL1 consumed 12% sugars producing only 1.8 g/L ethanol that was later metabolized as carbon source (Phase I). After 40 h of incubation, all available sugar was consumed by both strains and a part of the cultures was supplemented again with 1% yeast extract, which caused that BJ3505/YEpMEL1 shifted to use ethanol as carbon source and to secrete ScAGal, reaching a biomass four-fold higher than BJ3505Δsuc2/YEpMEL1 that continued secreting ScAGal (Phase II). Finally, the cell free medium at the end of the BJ3505Δsuc2/YEpMEL1 culture was inoculated with BJ3505/YEpMEL1 that converted the sucrose into ethanol (Phase III). So, the metabolism of BJ3505/YEpMEL1 is fermentative and produces ethanol from sucrose in 8% molasses, even under aerated conditions, known as Crabtree effect (Marques et al., 2015). However, BJ3505Δsuc2/YEpMEL1 shows a respiratory metabolism, probably due to the lower amount of sugars available since this strain cannot use sucrose, and metabolizes the rest of sugars present in molasses producing alpha-galactosidase more efficiently.
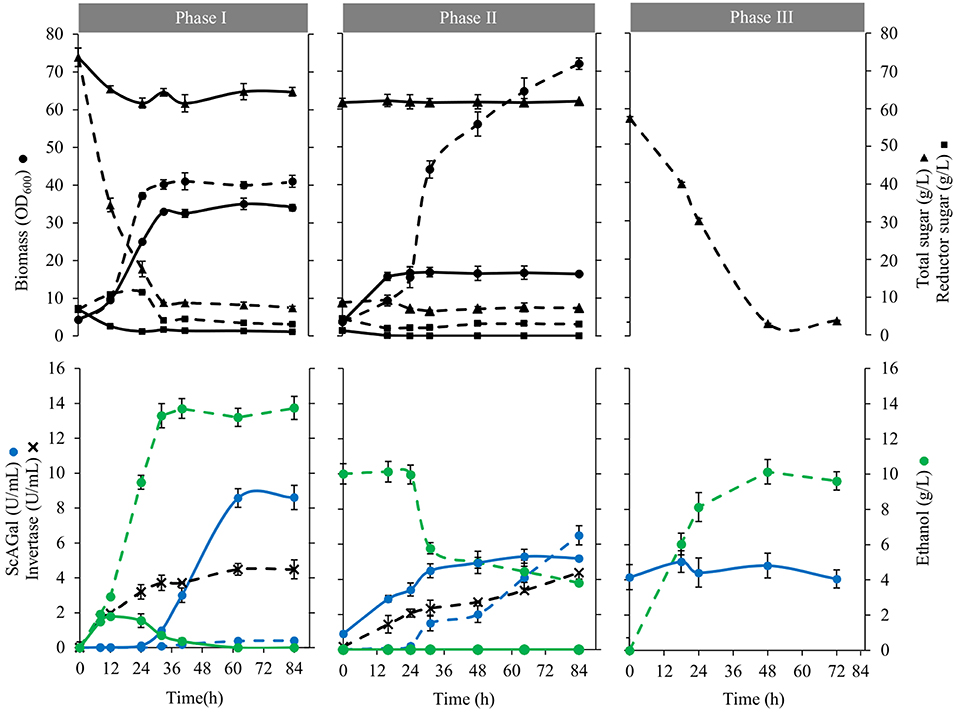
Figure 5. Growth kinetic and conversion of substrates to products of strains BJ3505Δsuc2/YEpMEL1 and BJ3505/YEpMEL1. Biomass (circles), total sugar (triangles), reducing sugar (squares), ethanol (green circles), extracellular alpha-galactosidase activity (blue circles), and extracellular invertase activity (blades) of cultures BJ3505Δsuc2/YEpMEL1 (solid line) and BJ3505/YEpMEL1 (dashed line). The cultures were inoculated in the YR production medium (8% beet molasses, 1% yeast extract) (Phase I). After a period of growth of 40 h, part of each of the cultures was separated and cooled with 1% yeast extract (Phase II) while the rest remained in the same conditions of phase I. The free medium of cells recovered from final cultures of BJ3505Δsuc2/YEpMEL1 was reused as a production medium using strain BJ3505/YEpMEL1 (Phase III). In each phase the same initial cell density was maintained (OD600 = 4).
Moreover, both strains BJ3505Δsuc2/YEpMEL1 and BJ3505/YEpMEL1 show morphological differences depending on the culture phase (Figure 6). By optical microscopy, the presence of particles around the vacuole is observed in BJ3505/YEpMEL1, while no vacuole is visible in BJ3505Δsuc2/YEpMEL1 and there are fewer particles in the cytoplasm (Phase I, Figure 6A). However, when BJ3505/YEpMEL1 starts to use ethanol as carbon source, the morphology of both strains is similar (Phase II, Figure 6A). The TEM images show in more detail the intense cytoplasmic activity in BJ3505Δsuc2/YEpMEL1 possibly as a result of the production of heterologous protein compared to the strain of origin (Phase I, Figure 6B).
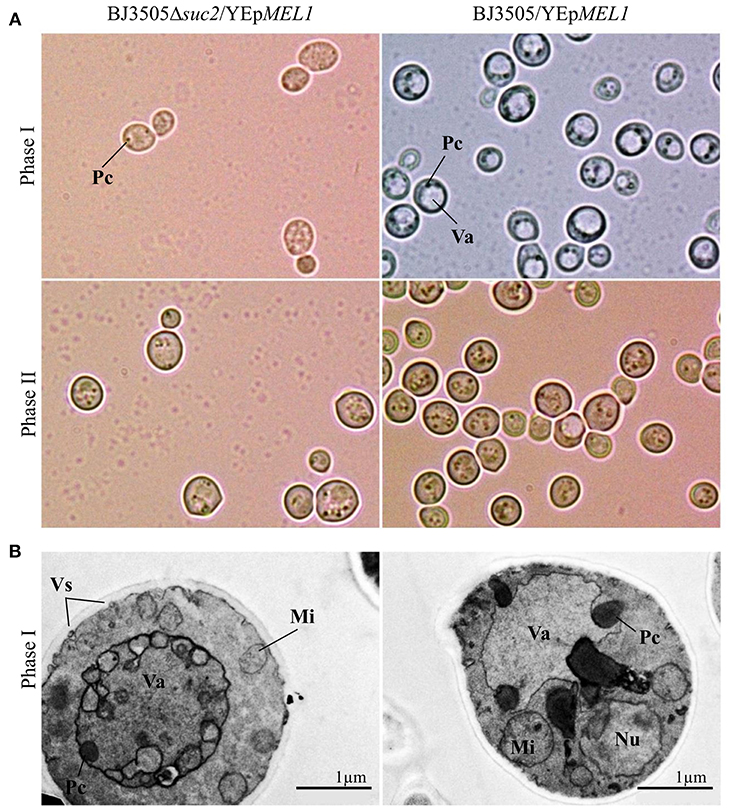
Figure 6. Visualization of cultures of BJ3505Δsuc2/YEpMEL1 and BJ3505/YEpMEL1 by optical microscopy (A) and TEM (B). Va, vacuole; Pc, Cytoplasmic particles; Nu, nucleus; Mi, mitochondria; Vs, secretion vesicles.
Optimization of ScAGal Production by BJ3505Δsuc2/YEpMEL1 From Beet Molasses
The matrix of CCD shown in Table 3 defined a set of 30 experiments, including six replicas of the central point to estimate the experimental error of the production technique. The corresponding results are also shown. Following, the surface response methodology (SRM) was applied to the experimental observed data in order to adjust the response function (extracellular alpha-galactosidase activity) and reveal the simultaneous influence of the four studied variables (molasses concentration, yeast extract concentration, inoculum size, and incubation time) on it. The best condition for ScAGal production corresponded to experiment number 13 that reached an extracellular alpha-galactosidase activity of 17 U/mL. In contrast, the worst condition (experiment 23) showed an activity value as low as 0.001 U/mL.
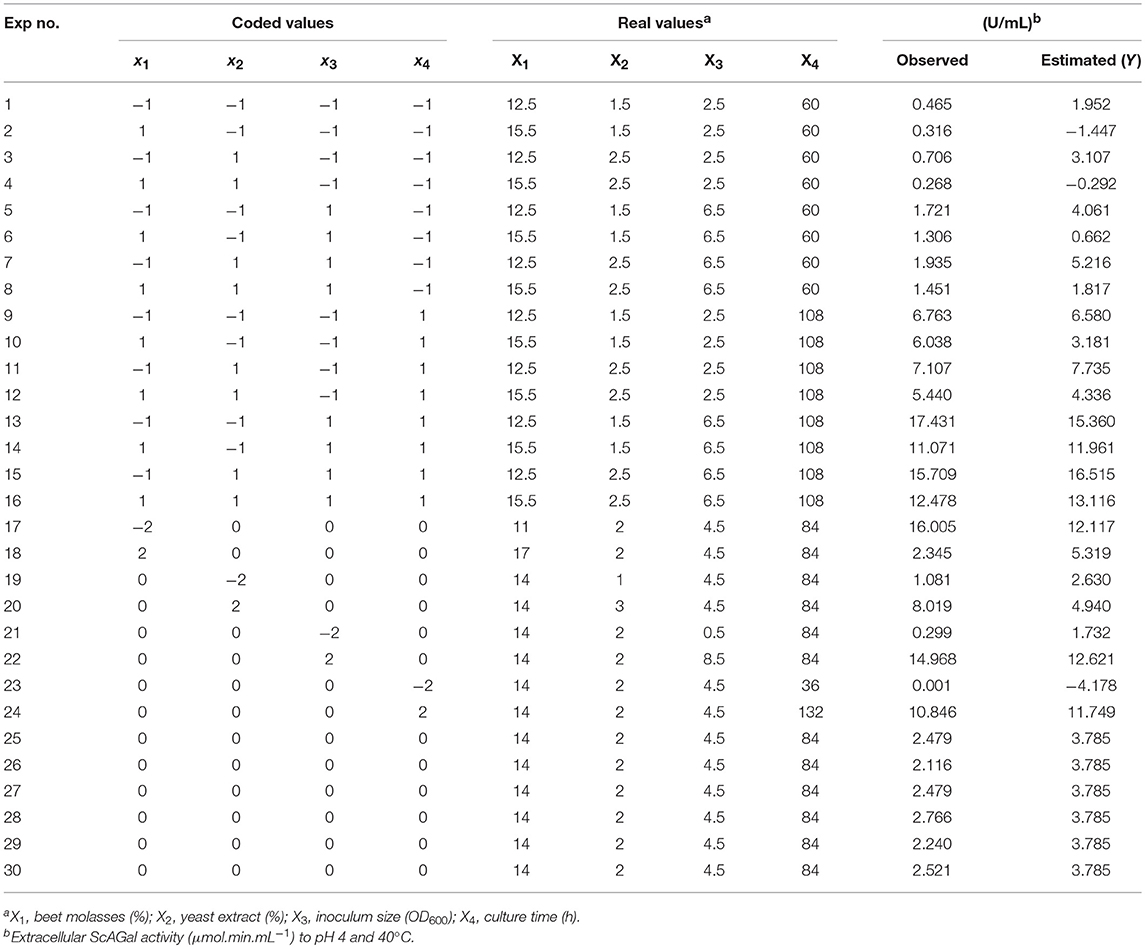
Table 3. Experimental matrix according to the CCD and results observed and predicted by the MSR to the optimization of the ScAGal production by BJ3505Δsuc2/YEpMEL1.
To validate the significance of the model obtained, the ANOVA presented in Table 4 was performed. The Pareto graphic analysis of the model shows the significance of the influence of the variables on the response (Figure 7A). The effects statistically not significant at the 95% confidence level were excluded from the model (and do not appear in Table 4), except yeast extract concentration that was the closest to the significance limit. Inoculum size and incubation time showed a positive effect on the response, whereas molasses concentration effect was negative. The effect of yeast extract concentration was smaller than the other three variables as corresponds to its lower significance level. Moreover, the representation of the single interaction that was significant, between inoculum size and incubation time (Figure 7B), shows that the increase of the inoculum size provokes a higher increase of the response if the incubation time also increases. The correlation coefficient (R2) that explains the 86.8% of the variation of the response and the p-value > 0.05 for the lack of fit test prove that the regression model is significant and can be used to describe the production of ScAGal into the defined experimental domain. Therefore, the following equation describes the relationship between response and variables: Extracellular alpha-galactosidase activity (Y) = 3.78–1.70x1+ 0.58x2 + 2.72x3 + 3.98x4 + 1.23x1 x1+ 0.85x3 x3 + 1.67Cx3x4.
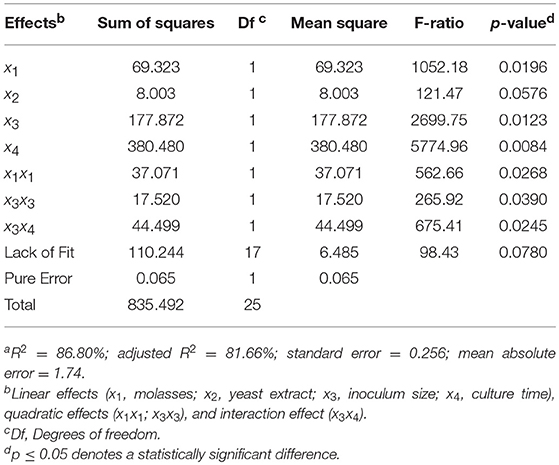
Table 4. ANOVA for the response surface quadratic model to the optimization of the ScAGal production by BJ3505Δsuc2/YEpMEL1a.
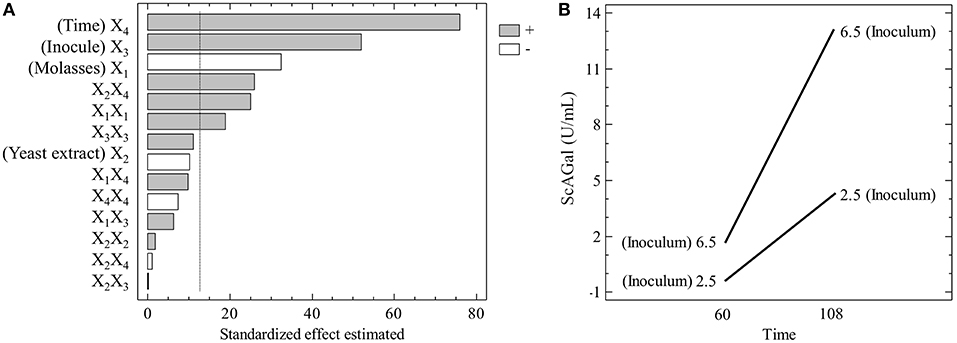
Figure 7. Pareto graphic and main interactions on the production of ScAGal. (A) Independent variables on the response at the 95% level of significance (vertical line) before remove the statistically insignificant effects. (B) Interaction Time/Inoculum effect, where the time varies from −1 to +1 while the inoculum remains constant at the value +1 (up line) and −1 (down line).
SRM and contour plot analysis reinforce that the effect of increasing incubation time (T) is more positive as inoculum size (I) increases (Figure 8A), and also when molasses concentration (M) decreases (Figure 8B). The combination of maximum I and minimum M also drives to an increase in ScAGal production (Figure 8C), while the effect of yeast extract (YE) is scarcely relevant in the experimental domain studied (Figures 8D–F).
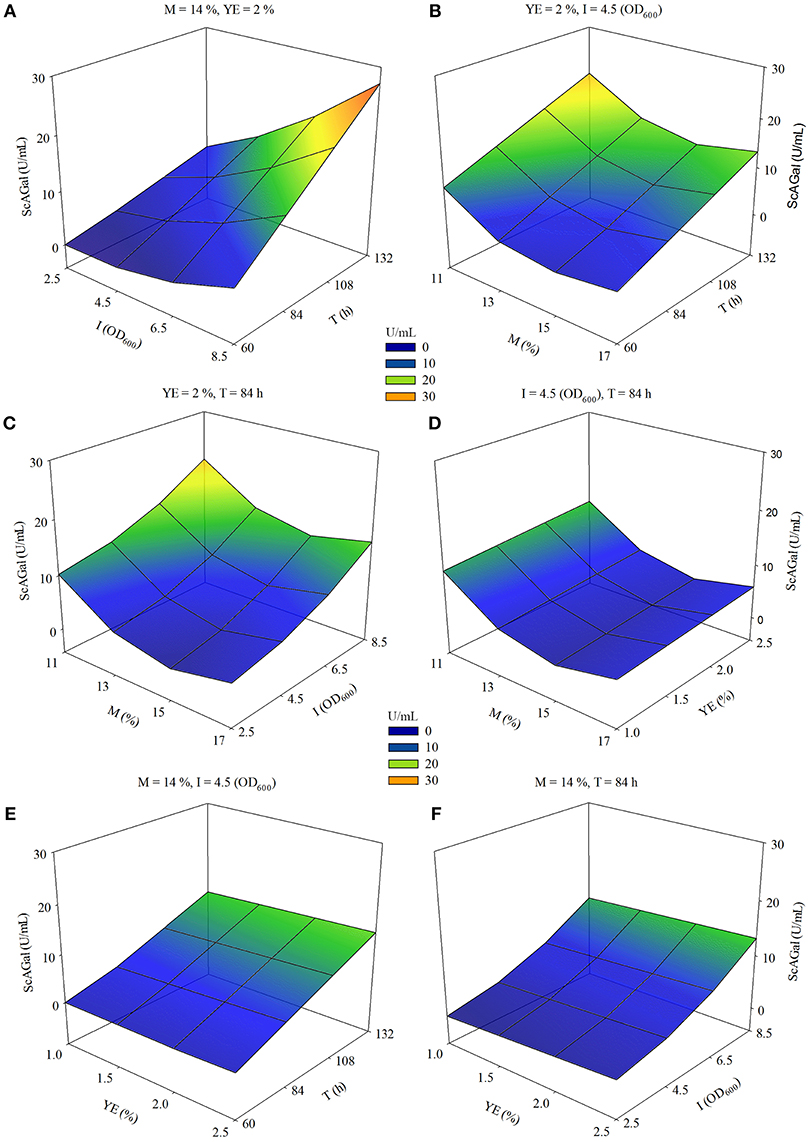
Figure 8. Response surface-contour plots for extracellular ScAGal production by the strain BJ3505Δsuc2/YEpMEL1 on beet molasses based medium using RSM. Estimated extracellular alpha-galactosidase activity (ScAGal) as function of (A) inoculum size (I) and time (T); (B) beet molasses (M) and time (T); (C) beet molasses (M) and inoculum size (I); (D) beet molasses (M) and yeast extract (YE); (E) yeast extract (YE) and time (T); (F) yeast extract (YE) and inoculum size (I). The values of the third and fourth variables were remained constant at level 0.
The maximum ScAGal production value is obtained in the experimental domain conditions x1 = −2, x2 = 0, x3 = +2, x4 = +2, which correspond to the real values 11% M, 2% YE, 8.5OD600 I, and 132 h T, respectively. According to the estimated model, an increase in YE (x2 = 0, X2 = 2%) would slightly increase the ScAGal production (above 4%) compared to the value x2 = −2 (X2 = 1%) of the same variable, keeping the rest of the conditions optimized. Therefore, we decided to use 1% YE to economize the productive process and since it is also the concentration usually used in commercial culture media. Under these conditions, the strain BJ3505Δsuc2/YEpMEL1 yielded the maximum extracellular alpha-galactosidase activity of 24 U/mL (Figure S2). This is a value close to predicted by the model (34 U/mL) and represents an improvement of the 72% with respect to the ScAGal production observed in the first studies performed in this work.
Therefore, in addition to being sustainable and low-cost, we have proved that molasses are also promising substrates for ScAGal production with the BJ3505Δsuc2/YEpMEL1 strain engineered in this work.
Scale-Up to 2-L Bioreactor
The strain BJ3505Δsuc2/YEpMEL1 was cultured in 2-L bioreactors with 1-L working volume of optimized YR (beet molasses-yeast extract) medium under controlled pH conditions. S. cerevisiae strains grow well between pH 4.5 and 6.5 (Walker and Stewart, 2016), and we decided to maintain bioreactors at pH 6 since it slightly improved the ScAGal production according to our previously observed experiments (Álvarez-Cao, 2017). Figure 9A shows the monitoring of bioreactors using optimized YR without (Bioreactor 1) and with (Bioreactor 2) adjustments at pH 6 during the culture time, where we found unexpected results that had not been considered before. First, cellular growth stopped at 72 h in both cases, but biomass and extracellular alpha-galactosidase activity were approximately double when the culture was maintain at pH 6, reaching about 20 g/L and 30 U/L, respectively, at 120 h of culture. Second, the pH value suddenly increased to pH 9 when it was not adjusted during the course of the culture. Apart from ScAGal, other metabolites were produced depending on the culture phase, shown in Table 5. Thus, during the first 24 h, the strain used all the glucose and only 73% (Bioreactor 1) or 82% (Bioreactor 1) of the fructose to produce ethanol and glycerol as co-metabolites. At the same time, galactose appears in the medium coming from 5% (Bioreactor 1) to 25% (Bioreactor 2) of the hydrolysis of raffinose by the action of ScAGal. Galactose concentration was maintained constant thereafter in both cultures, meaning that the strain prefers to use other monosaccharides (Marques et al., 2015). On the other hand, in the bioreactor without adjustment at pH 6 (Bioreactor 1; Figure 9A), from 24 to 48 h the strain metabolism was preferetially fermentative producing 8 g/L ethanol and 1.6 g/L glycerol while ScAGal activity was kept constant. During the following 48–96 h, strain's metabolism shifted to respiratory using ethanol and glycerol as carbon sources, and ScAGal expression sharply increased. The next 24 h, the production of ethanol and glycerol re-started and ScAGal activity reached 20 U/mL at 120 h of culture. However, when the bioreactor pH was maintained at 6 (Bioreactor 2; Figure 9A), the strain metabolism was respiro-fermentative during all culture, producing ethanol, glycerol, and ScAGal until 48 h, from 48 to 72 h consumed all the previously produced ethanol and a part of the glycerol while ScAGal increased up to 20 U/mL, and from 72 to 120 h, ethanol was newly produced and levels of ScAGal reached the highest values obtained (30 U/mL). If both cultures were extended, ethanol consumption and ScAGal production could be increased. These results are superior to those obtained in a similar characteristics bioreactor using the rich and much more expensive medium YPHSM with 5% (w/v) glucose (a level of assimilable sugars comparable to YR medium) that yielded more biomass (25 g/L) but less ScAGal (25 U/mL) (Bioreactor 3; Figure 9A). So, scale-up to 1-L culture in bioreactor with pH control allowed to increase a 21% the levels of ScAGal production compared with the best results obtained using shaken flasks (Figure S2).
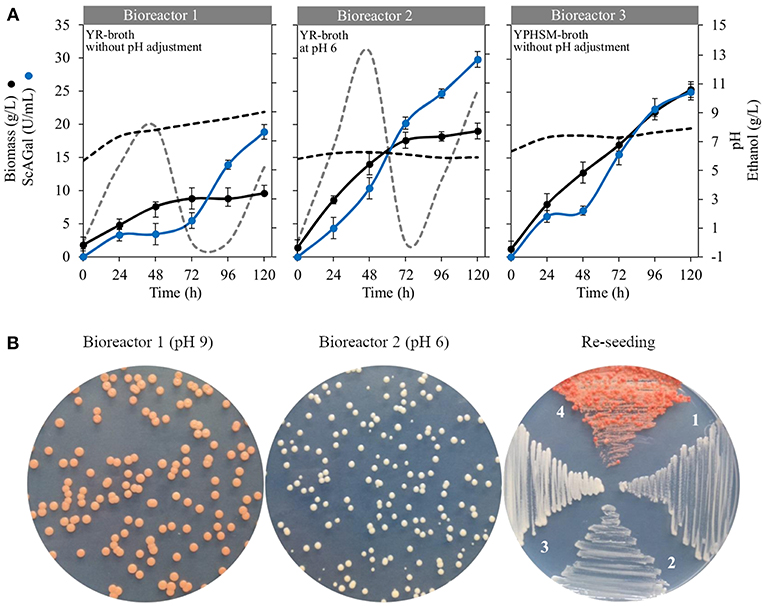
Figure 9. Cultivation of strain BJ3505Δsuc2/YEpMEL1 in bioreactors under controlled pH conditions. (A) Time course of cellular biomass (black circles), extracellular alpha-galactosidase activity (blue circles), and pH (black dashed line) in optimized YR (Bioreactor 1 and 2) and YPHSM (Bioreactor 3). The production of ethanol (gray dashed line) is shown in bioreactors 1 and 2. (B) Macroscopic visualization of cell colonies from ended-bioreactors, at pH 9 and pH 6, and re-seeding on selective medium CM-Trp. Re-seeding: 1, BJ3505/YEpMEL1 in YPHSM (pH 7); 2, BJ3505ΔSuc2/YEpMEL1 in YPHSM (pH 7); 3, BJ3505ΔSuc2/YEpMEL1 in YR (pH 6); 4, BJ3505ΔSuc2/YEpMEL1 in YR (pH 9).
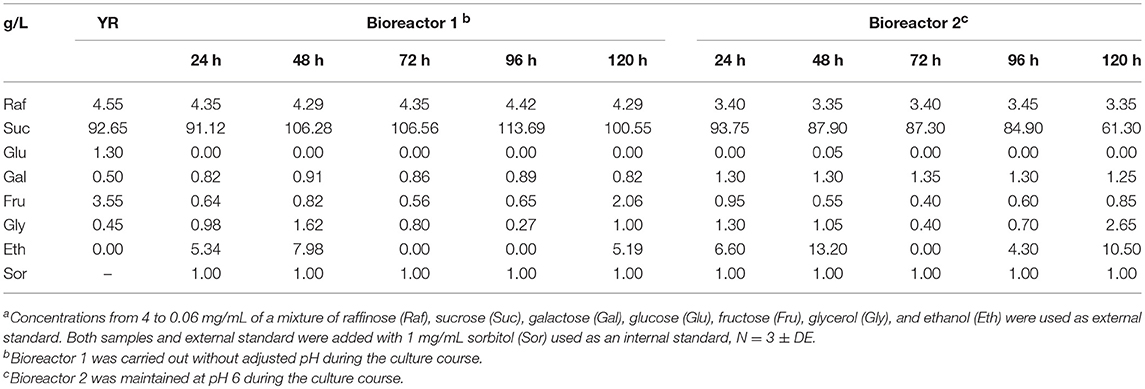
Table 5. HPLC analysis to quantify the conversion of substrates to products during the cultivation of bioreactors with strain BJ3505Δsuc2/YEpMEL1a.
Strikingly, when cells coming from ended-cultures of both bioreactors, at pH 9 and pH 6, were seeded in solid CM-Trp medium, only those coming from pH 9 acquired pink color (Bioreactor 1; Figure 9B) while the second ones maintained the normal phenotype (Bioreactor 2; Figure 9B). This characteristic was maintained during successive re-seedings for 10 months (Re-seeding; Figure 9B). Moreover, pink colonies grew faster than white ones and with a peculiar morphology. On the other hand, plasmid stability resulted to be about 98% in both bioreactors, independently of pH value, after 120 h of culture.
Discussion
Molasses are highly abundant sub-products of the food industry that are hitherto being fermented to ethanol at industrial level due to their richness in sucrose. The yeasts employed are adapted strains of S. cerevisiae, that produces high amounts of extracellular invertase, a native enzyme that hydrolyses sucrose into glucose and fructose, and is a Crabtree positive yeast, which means that preferentially ferments high concentrations of sugars to ethanol even under aerated conditions (Marques et al., 2015).
In this work, we started from the hypothesis that a mutant strain not-producing invertase (Δsuc2), i.e., unable to use sucrose, would metabolize the sugars present in molasses at lower concentration (raffinose, glucose, fructose) that could be used to produce a recombinant protein. In our case, the selected recombinant protein was ScAGal, an enzyme widely used in several industries and with large market, that would also simultaneously allow to use the raffinose present in molasses since ScAGal shows a high affinity for this sugar (Fernández-Leiro et al., 2010). The remaining sucrose could be fermented by wild type S. cerevisae in a coupled second-step classical industrial process.
Figure 10 shows the scheme of a proposed productive process of ScAGal and bioethanol in the conditions optimized in this work, using beet molasses as substrate and the constructed engineered yeast strain. In the first step, ScAGal would be obtained from cultures of the strain BJ3505Δsuc2/YEpMEL1 in molasses medium. In the second step, bioethanol would be obtained from cultures of any wild type yeast strain, in our case the strain of origin BJ3505, in post-incubate cell-free sucrose-rich medium resulting from the first step after ScAGal recovery by tangential flow filtration (TFF). The ethanol produced in step 2 could be extracted for biofuel use, and also recycled as carbon source for ScAGal production in the step 1. Moreover, one part of the yeast biomass obtained (of each of the two strains) would serve as inoculum of the following culture, and the rest would be autolyzed and then used to supplement molasses instead of the more expensive commercial yeast extract.
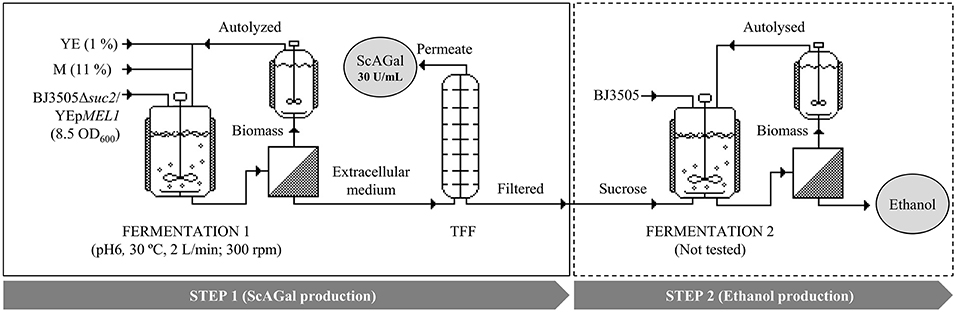
Figure 10. Simulation of a two-step bioprocess to obtain ScAGaL and ethanol from beet molasses-based medium. The ended-culture of the fermentation 1 carried out by the invertase-deficient strain BJ3505Δsuc2/YEpMEL1 is centrifuged, and the extracellular medium filtered by TTF to obtain a permeate with the recombinant protein, ScAGal (Step 1). The sucrose recovered in the filtrate is used in the fermentation 2 to produce ethanol by a strain of S. cerevisiae with invertase activity (Step 2). The biomass recovered from both fermentations can be used as yeast extract (YE) supplement after an autolysis treatment. M, beet molasses; YE, yeast extract; TFF, tangential flow filtration. Modificated imagen from SuperPro Designer.
During the research conducted to develop the first step of this proposed coupled process, we found several interesting observations. On the one hand, the Δsuc2 strain produced more extracellular ScAGal from molasses than the corresponding wild type strain. We attributed this result to the relief of the secretory route due to the absence of invertase (Liljeström et al., 1991) and to the shift to a respiratory metabolism due to the inability to use sucrose (Finn et al., 2006). In addition, ScAGal became easier to purify from the culture medium without contaminating invertase. On the other hand, the Δsuc2 strain presented some curious morphological changes. In the first place, the absence of vacuolar particles that are observed in the wild type strain growing in molasses, and that in the case of the mutant are cytosolic. Under stress conditions, the vacuole is prone to flow between non-fragmented and fragmented forms due in part to its association with other cellular organelles (endoplasmic reticulum, Golgi complex, secretory, and endocytic vesicles) as components of interconnected branched systems (Gibson et al., 2007). This change of morphology could be attributed to a stress response due to environmental factors, such as glucose depletion, tolerance to ethanol (Gibson et al., 2007; Kato et al., 2011; Grousl et al., 2013), or to the switch from fermentative to respiratory metabolism since the wild type strain growing in ethanol does not present such vacuolar particles. In the second place, the Δsuc2 strain gains a pink color in a pH-dependent manner that could be related to the synthesis of carotenoid pigments by yeast as has also been reported in the literature for other microorganisms (Querijero-Palacpac et al., 1990; Schmidt et al., 2011). However, the molecular mechanism of these morphological changes merits to be the subject of further research.
It is well-known that high concentrations of sucrose exert hyperosmotic stress on S. cerevisiae which synthesizes antioxidant and osmoregulatory molecules to avoid lethal injuries, such as trehalose and glycerol, among others (Gibson et al., 2007). The accumulation of trehalose, as osmotic protector, with similar retention time than sucrose or the induction of the synthesis of other hydrolases involved in sucrose metabolism (Marques et al., 2015) could explain the increase and decrease in sucrose levels observed during cultures without and with pH control, respectively (Table 5).
Finally, we reported for the first time the statistical optimization of the production of ScAGal from molasses (using CCD) with high yields of enzyme production, while the fermentation to ethanol of the remaining sucrose was just verified but not optimized since this procedure is nowadays ongoing at industrial scale. In addition, the stability of the plasmid expressing ScAGal is not a problem since 98% of the cells growing in molasses retained the plasmid at the end of the cultures. In fact, we obtained a production of 30 U/mL of ScAGal which is 1.6-fold higher than the recently reported with a Kluyveromyces lactis engineered strain growing in a mixture of cheese whey and molasses (Álvarez-Cao et al., 2018) and higher than the 10.4 U/mL obtained for Aspergillus fumigatus alpha-galactosidase expressed in A. sojae in an optimized synthetic medium in the presence of 10.5% (w/v) molasses (Gurkok et al., 2011). The alpha-galactosidase yield reached is among the highest of those obtained in submerged fermentation for the alpha-galactosidase enzyme of different origins on various carbon sources (Anisha, 2017).
Conclusion
A procedure for valuation of beet molasses has been developed that consists in the high-yield production of ScAGal, an enzyme with wide market, by an engineered yeast strain from the less abundant sugars, followed by ethanol fermentation of sucrose, the most abundant sugar, by a wild type strain. The engineered strain is a Δsuc2 mutant transformed with a plasmid that drives secretion of ScAGal to the culture medium. The mutation causes a shift to a more respiratory metabolism in molasses and facilitates extracellular ScAGal purification by avoiding contamination with invertase.
Data Availability
All datasets generated for this study are included in the manuscript and/or the supplementary files.
Author Contributions
M-EC, M-IG-S, and MB contributed conception and design of the study. M-EA-C performed the experimental work (data acquisition); M-EA-C, M-IG-S, and MB contributed to analysis or interpretation of data. M-EA-C and M-IG-S wrote the first draft of the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version.
Funding
This work was supported by the Fundación Barrié, Fondo de Investigación en Ciencia [grant Bio-alfa-gal]; and the Xunta de Galicia, Consolidación Grupos Referencia Competitiva, co-financed by FEDER [grant ED431C 2016–012].
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
AB Azucarera Iberia provided molasses.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.00405/full#supplementary-material
References
Akbas, M. Y., Sar, T., and Ozcelik, B. (2014). Improved ethanol production from cheese whey, whey powder, and sugar beet molasses by “Vitreoscilla hemoglobin expressing” Escherichia coli. Biosci. Biotechnol. Biochem. 78, 687–694. doi: 10.1080/09168451.2014.896734
Akbas, M. Y., and Stark, B. C. (2016). Recent trends in bioethanol production from food processing byproducts. J. Ind. Microbiol. Biotechnol. 43, 1593–1609. doi: 10.1007/s10295-016-1821-z
Álvarez-Cao, M.-E. (2017). Optimización de la Producción Heteróloga de la Enzima α-Galactosidasa de Saccharomyces cerevisiae a Partir de Residuos Agroindustriales. Ph.D. thesis, Universidade da Coruña. Available online at: https://ruc.udc.es/dspace/handle/2183/19936
Álvarez-Cao, M.-E., Rico-Díaz, A., Cerdán, M.-E., Becerra, M., and González-Siso, M.-I. (2018). Valuation of agro-industrial wastes as substrates for heterologous production of α-galactosidase. Microb. Cell Fact. 17, 1–13. doi: 10.1186/s12934-018-0988-6
Anisha, G. S. (2017). “α-Galactosidases,” in Current Developments in Biotechnology and Bioengineering: Production, Isolation and Purification of Industrial Products, eds A. Pandey, S. Negi, and C. Soccol (Amsterdam: Elsevier B.V.), 369–394. doi: 10.1016/B978-0-444-63662-1.00016-6
Arends, M., Biegstraaten, M., Hughes, D. A., Mehta, A., Elliott, P. M., Oder, D., et al. (2017). Retrospective study of long-term outcomes of enzyme replacement therapy in Fabry disease: analysis of prognostic factors. PLoS ONE 12:e0182379. doi: 10.1371/journal.pone.0182379
Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A., et al. (2003). Current Protocols in Molecular Biology. New York, NY: John Wiley and Sons, Inc. doi: 10.1002/mrd.1080010210
Bauer, C., Herzog, V., and Bauer, M. F. (2001). Improved technique for electron microscope visualization of yeast membrane structure. Microsc. Microanal. 7, 530–534. doi: 10.1007/s10005-001-0020-4
Belem, M. A. F., Gibbs, B. F., and Lee, B. H. (1997). Enzymatic production of ribonucleotides from autolysates of Kluyveromyces marxianus grown on whey. J. Food Sci. 62, 851–857. doi: 10.1111/j.1365-2621.1997.tb15470.x
Chen, D.-C., Yang, B.-C., and Kuo, T.-T. (1992). One-step transformation of yeast in stationary phase. Curr. Genet. 21, 83–84. doi: 10.1007/BF00318659
da Silva, L. L. (2016). Adding value to agro-industrial wastes. Ind. Chem. 2, 1–2. doi: 10.4172/2469-9764.1000e103
Dai, Z., Lyu, W., Xiang, X., Tang, Y., Hu, B., Ou, S., et al. (2018). Immunomodulatory effects of enzymatic-synthesized α-galactooligosaccharides and evaluation of the structure–activity relationship. J. Agric. Food Chem. 66, 9070–9079. doi: 10.1021/acs.jafc.8b01939
de Faria, J. T., Rocha, P. F., Converti, A., Passos, F. M. L., Minim, L. A., and Sampaio, F. C. (2013). Statistical investigation of Kluyveromyces lactis cells permeabilization with ethanol by response surface methodology. Braz. J. Microbiol. 44, 1067–1074. doi: 10.1590/S1517-83822013000400007
Dey, P. M., Patel, S., and Brownleader, M. D. (1993). Induction of alpha-galactosidase in Penicillium ochrochloron by guar (Cyamopsis tetragonobola) gum. Biotechnol. Appl. Biochem. 17, 361–371. doi: 10.1111/j.1470-8744.1993.tb00250.x
Dilipkumar, M., Rajasimman, M., and Rajamohan, N. (2014). Optimization, kinetics, and modeling of inulinase production by K. marxianus var. marxianus. Prep. Biochem. Biotechnol. 44, 291–309. doi: 10.1080/10826068.2013.812567
Fernández-Leiro, R. (2011). Structural Characterization and Directed Evolution of an α-Galactosidase From Saccharomyces cerevisiae. Ph.D. thesis, Universidade da Coruña.
Fernández-Leiro, R., Pereira-Rodriguez, A., Cerdan, M. E., Becerra, M., and Sanz-Aparicio, J. (2010). Structural analysis of Saccharomyces cerevisiae α-galactosidase and its complexes with natural substrates reveals new insights into substrate specificity of GH27 glycosidases. J. Biol. Chem. 285, 28020–28033. doi: 10.1074/jbc.M110.144584
Ferreira, I. M. P. L. V. O., Pinho, O., Vieira, E., and Tavarela, J. G. (2010). Brewer's Saccharomyces yeast biomass: characteristics and potential applications. Trends Food Sci. Technol. 21, 77–84. doi: 10.1016/j.tifs.2009.10.008
Finn, B., Harvey, L. M., and McNeil, B. (2006). Near-infrared spectroscopic monitoring of biomass, glucose, ethanol and protein content in a high cell density baker's yeast fed-batch bioprocess. Yeast 23, 507–517. doi: 10.1002/yea.1371
Gibson, B. R., Lawrence, S. J., Leclaire, J. P. R., Powell, C. D., and Smart, K. A. (2007). Yeast responses to stresses associated with industrial brewery handling. FEMS Microbiol. Rev. 31, 535–569. doi: 10.1111/j.1574-6976.2007.00076.x
Gietz, R. D., and Sugino, A. (1988). New yeast-Escherichia coli shuttle vectors constructed with in vitro mutagenized yeast genes lacking six-base pair restriction site. Gene 74, 527–534. doi: 10.1016/0378-1119(88)90185-0
Grousl, T., Ivanov, P., Malcova, I., Pompach, P., Frydlova, I., Slaba, R., et al. (2013). Heat shock-induced accumulation of translation elongation and termination factors precedes assembly of stress granules in S. cerevisiae. PLoS ONE 8, 1–17. doi: 10.1371/journal.pone.0057083
Gurkok, S., Cekmecelioglu, D., and Ogel, Z. B. (2011). Optimization of culture conditions for Aspergillus sojae expressing an Aspergillus fumigatus α-galactosidase. Bioresour. Technol. 102, 4925–4929. doi: 10.1016/j.biortech.2011.01.036
Hoffman, C. S. (2001). “Preparation of yeast DNA,” in Current Protocols in Molecular Biology, eds F. M. Ausubel, R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and k. Struhl (New York, NY: John Wiley and Sons, Inc.), 13.11.1–13.11.4. doi: 10.1002/0471142727.mb1311s39
Ito, H., Fukuda, Y., Murata, K., and Kimura, A. (1983). Transformation of intact yeast cells treated with alkali cations. J. Bacteriol. 153, 163–168.
Kato, K., Yamamoto, Y., and Izawa, S. (2011). Severe ethanol stress induces assembly of stress granules in Saccharomyces cerevisiae. Yeast 28, 339–347. doi: 10.1002/yea.1842
Katrolia, P., Jia, H., Yan, Q., Song, S., Jiang, Z., and Xu, H. (2012). Characterization of a protease-resistant α-galactosidase from the thermophilic fungus Rhizomucor miehei and its application in removal of raffinose family oligosaccharides. Bioresour. Technol. 110, 578–586. doi: 10.1016/j.biortech.2012.01.144
Kim, S.-B., Kim, J.-H., Lee, Y.-M., Kim, J.-H., Park, S.-W., and Lee, K.-P. (2009). Process for Manufacturing Tagatose Using Soy Oligosaccharide. WO2009/096693
Liljeström, P. L., Tubb, R. S., and Korhola, M. P. (1991). Construction of New α-Galactosidase Producing Yeast Strains and the Industrial Application of These Strains. US5055401A
Linden, J. C. (1982). Immobilized α-d-galactosidase in the sugar beet industry. Enzyme Microb. Technol. 1982:103. doi: 10.1016/0141-0229(82)90103-X
Marques, W. L., Raghavendran, V., Stambuk, B. U., and Gombert, A. K. (2015). Sucrose and Saccharomyces cerevisiae: a relationship most sweet. FEMS Yeast Res. 16, 1–48. doi: 10.1093/femsyr/fov107
Miller, G. L. (1959). Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Biochem. 31, 426–428. doi: 10.1021/ac60147a030
Nielsen, J. (2013). Production of biopharmaceutical proteins by yeast: advances through metabolic engineering. Bioengineered 4, 207–211. doi: 10.4161/bioe.22856
Querijero-Palacpac, N. M., Martinez, M. R., and Boussiba, S. (1990). Mass cultivation of the nitrogen-fixing cyanobacterium Gloeotrichia natans, indigenous to rice-fields. J. Appl. Phycol. 2, 319–325. doi: 10.1007/BF02180921
Rodrigues-Dutra, T., Monteze-Guimarães, V., Mascarenhas-Varela, E., da Silva-Fialho, L., Ferreira-Milagres, A. M., Falkoski, D. L., et al. (2017). A Chrysoporthe cubensis enzyme cocktail produced from a low-cost carbon source with high biomass hydrolysis efficiency. Sci. Rep. 7:3893. doi: 10.1038/s41598-017-04262-y
Ryan, M. P., Jones, R., and Morse, R. H. (1998). SWI-SNF complex participation in transcriptional activation at a step subsequent to activator binding. Mol. Cell. Biol. 18, 1774–1782. doi: 10.1128/MCB.18.4.1774
Santos, A. C., Silva Bezerra, M., dos Santos Pereira, H., dos Santos, E. S., and de Macedo, G. R. (2010). Production and recovery of rhamnolipids using sugar cane molasses as carbon source. J. Chem. Chem. Eng. 4, 27–33. doi: 10.17265/1934-7375/2010.11.005
Schmidt, I., Schewe, H., Gassel, S., Jin, C., Buckingham, J., Hümbelin, M., et al. (2011). Biotechnological production of astaxanthin with Phaffia rhodozyma/Xanthophyllomyces dendrorhous. Appl. Microbiol. Biotechnol. 89, 555–571. doi: 10.1007/s00253-010-2976-6
Spangenberg, P., André, C., Dion, M., Rabiller, C., and Mattes, R. (2000). Comparative study of new alpha-galactosidases in transglycosylation reactions. Carbohydr. Res. 329, 65–73. doi: 10.1016/S0008-6215(00)00170-1
Walker, G. M., and Stewart, G. G. (2016). Saccharomyces cerevisiae in the production of fermented beverages. Beverages 2, 1–12. doi: 10.3390/beverages2040030
Xu, W., Liang, L., and Zhu, M. (2015). Determination of sugars in molasses by HPLC following solid-phase extraction. Int. J. Food Prop. 18, 547–557. doi: 10.1080/10942912.2013.837064
Keywords: alpha-galactosidase, invertase, Saccharomyces cerevisiae, beet molasses, bioconversion, ethanol
Citation: Álvarez-Cao M-E, Cerdán M-E, González-Siso M-I and Becerra M (2019) Bioconversion of Beet Molasses to Alpha-Galactosidase and Ethanol. Front. Microbiol. 10:405. doi: 10.3389/fmicb.2019.00405
Received: 11 October 2018; Accepted: 15 February 2019;
Published: 07 March 2019.
Edited by:
Laurent Dufossé, Université de la Réunion, FranceReviewed by:
Anna Irini Koukkou, University of Ioannina, GreeceMarta Wilk, Wroclaw University of Economics, Poland
Copyright © 2019 Álvarez-Cao, Cerdán, González-Siso and Becerra. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María-Isabel González-Siso, bWlnc0B1ZGMuZXM=