 Rosalia Mamani1†
Rosalia Mamani1† Saskia Camille Flament-Simon1†
Saskia Camille Flament-Simon1† Vanesa García1
Vanesa García1 Azucena Mora1
Azucena Mora1 María Pilar Alonso2
María Pilar Alonso2 Cecilia López1
Cecilia López1 Isidro García-Meniño1
Isidro García-Meniño1 Dafne Díaz-Jiménez1
Dafne Díaz-Jiménez1 Jesús E. Blanco1
Jesús E. Blanco1 Miguel Blanco1
Miguel Blanco1 Jorge Blanco1*
Jorge Blanco1*- 1Laboratorio de Referencia de E. coli, Departamento de Microbioloxía e Parasitoloxía, Facultade de Veterinaria, Universidade de Santiago de Compostela, Lugo, Spain
- 2Unidade de Microbioloxía Clínica, Hospital Universitario Lucus Augusti, Lugo, Spain
The aim of the present study was to examine the prevalence and determine the molecular characteristics of extended-spectrum β-lactamase-producing Escherichia coli (ESBL-EC) causing bacteraemia in a Spanish Hospital over a 12-year period (2000 to 2011). As far as we know, this is the first study which has investigated and compared the serotypes, phylogroups, clonotypes, virotypes, and PFGE profiles of ST131 and non-ST131 clones of bacteraemia ESBL-EC isolates. Of the 2,427 E. coli bloodstream isolates, 96 (4.0%) were positive for ESBL production: 40 for CTX-M-15, 36 for CTX-M-14, eight for CTX-M-1, four for CTX-M-9, CTX-M-32, and SHV-12. The number of ESBL-EC increased from 1.0% during 2000 to 2005 to 5.5% during 2006–2011 (P < 0.001). The 96 ESBL-EC isolates belonged to 36 different STs. The commonest was ST131 (41 isolates), followed by ST58, ST354, ST393 and ST405 (four isolates each). Most CTX-M-15 isolates (87.5%, 35/40) were ST131, whereas the 36 CTX-M-14 isolates belonged to 23 different STs and only 3 (8.3%) of them were ST131. The 35 ST131 CTX-M-15-producing isolates belonged to the H30Rx subclone and 29 of them showed the virotype A. A drastic change in ST131 virotypes happened in 2011 due to the emergence of the virotypes E (sat, papGII, cnf1, hlyA, and kpsMII-K5) and F (sat, papGII, and kpsMII-K5) which displaced virotype A (afa/draBC, afa operon FM955459, sat, and kpsMII-K2). Although the 96 ESBL-EC isolates showed 21 O serogroups and 17 H flagellar antigens, 39 belonged to serotype O25b:H4 (ST131 isolates). The second most prevalent serotype (O15:H1) was found to be associated with another important high-risk clone (ST393). In conclusion, the ST131 was the most frequent sequence type, being the H30Rx subclone responsible for the significant increase of ESBL-EC isolates since 2006. Here, we report two new virotypes (E and F) of the H30Rx subclone emerged in 2011. Future molecular studies are needed to understand the dynamics of expansion of this successful high-risk subclone in order to prevent its spread and establish the importance of the two new virotypes.
Introduction
Escherichia coli is the main cause of bloodstream infections (BSIs) and in recent years there has been a very significant increase in the number of infections caused by multidrug-resistant isolates, and especially by extended-spectrum β-lactamase-producing E. coli (ESBL-EC) (Tumbarello et al., 2010; de Kraker et al., 2013; Vihta et al., 2018). ESBL production in E. coli has increased due mainly to the spread of CTX-M enzymes. The high-risk clones have played a very important role in the current pandemic spread, especially the ST131 clone associated with the CTX-M-15 enzyme (Nicolas-Chanoine et al., 2008; Chung et al., 2012; Peirano et al., 2012; Adams-Sapper et al., 2013; Banerjee et al., 2013; Colpan et al., 2013; Matsumura et al., 2013; Johnson et al., 2017; Kallonen et al., 2017; Roer et al., 2017; Mendis et al., 2018).
The majority of ST131 isolates belong to the O25b:H4 serotype, subclone H30, clonotype CH40-30 and clade C. The H30Rx (clade C2) subset often carries blaCTX–M–15, whereas H30R-M27 (clade C1-M27) subset is positive for blaCTX–M–27 and H30R-nM27 (clade C1-nM27) subset is often positive for blaCTX–M–14. However, there are other two ST131 subclones less frequently expanded: H22 (clonotype CH40-22, clade B) and H41 (clonotype CH40-41, clade A). Isolates of subclone H41 usually belong to serotype O16:H5 (Price et al., 2013; Olesen et al., 2014; Peirano et al., 2014; Matsumura et al., 2017a,b; Harris et al., 2018). Generally, clades A and B show fluoroquinolone (FQ) susceptibility and rarely carry ESBL plasmids, while most isolates of clade C are FQ-resistant. Clade B evolved into clade C by acquisition of several prophages, genomic islands, the fimH30 allele and mutations within gyrA and parC genes, mainly during the late 1980s (Ben Zakour et al., 2016; McNally et al., 2016; Stoesser et al., 2016; Pitout and DeVinney, 2017). In addition, considering the content of virulence genes, ST131 isolates can be classified into 12 virotypes (A to F) (Blanco et al., 2013; Dahbi et al., 2014; Mora et al., 2014).
Currently, few data are available on molecular epidemiology of bacteraemia caused by ST131 and non-ST131 clones of ESBL-EC, especially in Europe (Rodríguez-Baño et al., 2012; Horner et al., 2014; Day et al., 2016; Merino et al., 2016; Roer et al., 2017). Thus, the aim of the present study was to examine the prevalence and determine the molecular characteristics of ESBL-EC causing bacteraemia in a Spanish Hospital over a 12-year period (2000–2011).
Materials and Methods
E. coli Bloodstream Isolates
The present study included 2,427 non-duplicate, clinically relevant E. coli isolates recovered from blood of patients at Hospital Universitario Lucus Augusti (HULA) (formely Complejo Hospitalario Xeral-Calde) between January 2000 and December 2011. All available isolates from that time period were analyzed. HULA is a university hospital with 740 beds, which serves an urban and rural population of approximately 265,000 inhabitants in Lugo, Spain. Ethics approval was not required according to national and institutional guidelines.
Antibiotic Susceptibility Testing and ESBL Typing
ESBL production was detected by the double disk synergy test (Jarlier et al., 1988). The type of ESBL was established by PCR and sequencing using the TEM-, SHV-, CTX-M-1, and CTX-M-9 group-specific primers (Leflon-Guibout et al., 2004; Blanco et al., 2009).
Phylogenetic Grouping, MLST, CH Typing, and Identification of H30Rx Subclone
The determination of the phylogenetic groups (A, B1, B2, C, D, E, F) was carried out by the protocol of Clermont et al. (2013). The sequence types (STs) were established following the multilocus sequence typing (MLST) scheme of Achtman1 (Wirth et al., 2006). Clonotype identification was determined by fumC and fimH (CH) sequencing (Weissman et al., 2012; Tchesnokova et al., 2013). The H30Rx subclone was established by PCR detection of a specific SNP (G723A) of ybbW gene (Banerjee et al., 2013).
O and H Typing
The serotyping was carried out using all available O (O1 to O181) and H (H1 to H56) antisera (Guinée et al., 1981). Isolates that did not react with any antisera were designated as ONT or HNT (NT, non-typeable) and those non motile were denoted as HNM. The O25b subtype was determined by PCR (Clermont et al., 2008).
Virulence Genotyping
Virulence genes were screened by PCR as documented previously (Dahbi et al., 2014), using specific primers (Supplementary Table S1). The virulence gene score was the number of extraintestinal virulence-associated genes detected. Isolates were defined presumptively as extraintestinal pathogenic E. coli (ExPEC) (Johnson et al., 2015) if positive for ≥2 of 5 markers, including papAH and/or papC, sfa/focDE, afa/draBC, kpsM II, and iutA, and as uropathogenic E. coli (UPEC) (Spurbeck et al., 2012) if positive for ≥3 of 4 markers, including chuA, fyuA, vat, and yfcV. The virotypes A to F of the ST131 isolates was assigned according to the scheme developed by Dahbi et al. (2014).
Pulsed Field Gel Electrophoresis (PFGE)
XbaI-PFGE profiles were determined as previously described (Mora et al., 2009, 2010) and imported into BioNumerics (Applied Maths, Sint-Martens-Latem, Belgium) and clustered using the UPGMA algorithm and applying Dice coefficient and 1% tolerance.
Statistical Analysis
All the P-values were calculated using the Fisher’s exact test, except for the comparison of the means that was performed using the test one-way ANOVA test. P-values <0.05 were considered statistically significant.
Results
Prevalence of ESBL-Producing E. coli (ESBL-EC)
During 2000–2011, 2,427 E. coli bloodstream isolates were analyzed, of which 96 (4.0%) exhibited ESBL production. The number of ESBL-EC isolates increased from 1.0% (8/827) during 2000–2005 to 5.5% (88/1,600) during 2006–2011 (P < 0.001) (Figure 1, Table 1, and Supplementary Table S2).
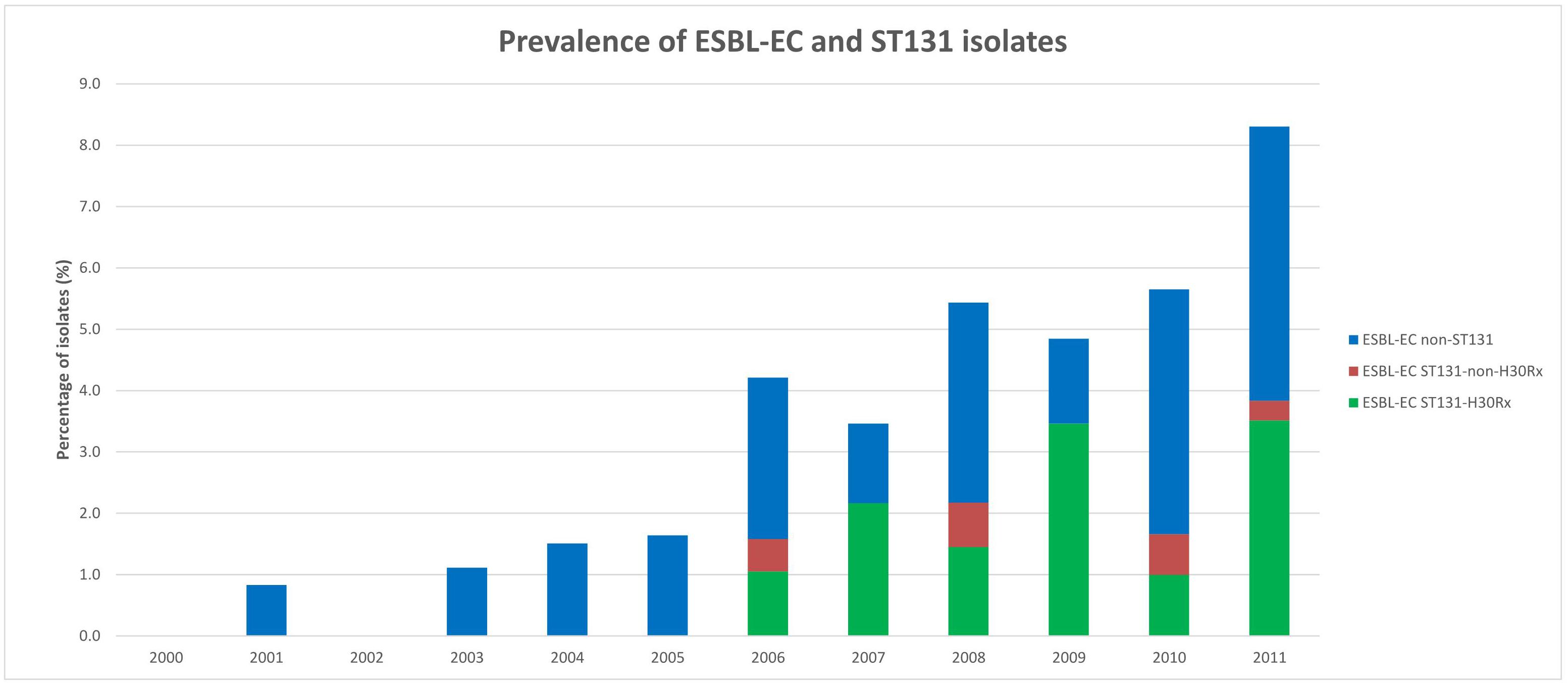
Figure 1. Prevalence of extended-spectrum β-lactamase-producing E. coli (ESBL-EC) and ESBL-EC ST131 isolates. A total 2,427 E. coli bloodstream isolates were analyzed, of which 96 (4.0%) exhibited ESBL production. A total of 41 ESBL-EC ST131 isolates were identified and 35 of them belonged to the H30Rx subclone.
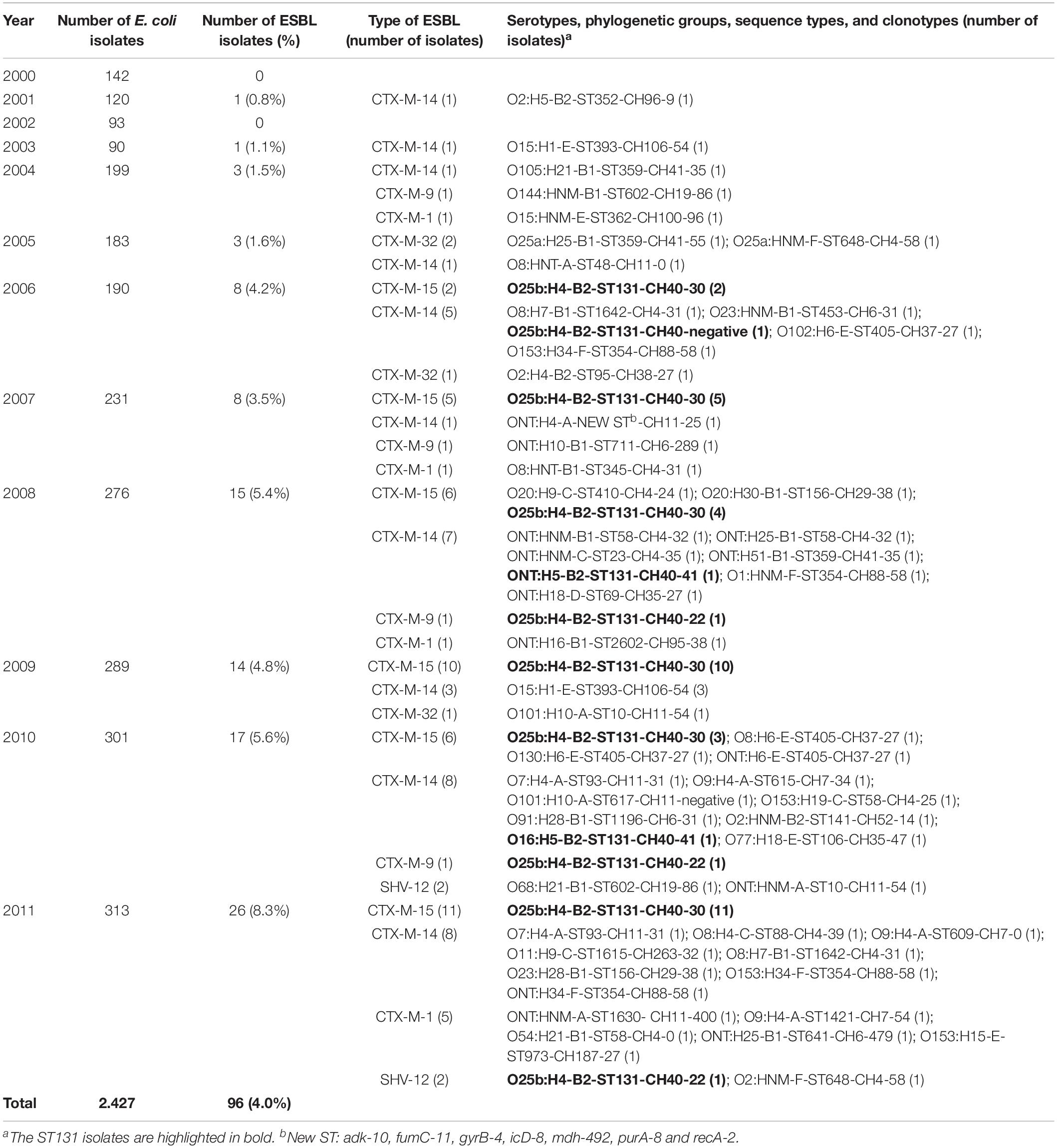
Table 1. Prevalence, serotypes, phylogenetic groups, STs, and clonotypes of extended-spectrum β-lactamase-producing E. coli (ESBL-EC) bloodstream isolates.
ESBL Enzymes
Of the 96 ESBL-EC isolates, 40 were positive for CTX-M-15, 36 for CTX-M-14, eight for CTX-M-1, and four for CTX-M-9, CTX-M-32, and SHV-12 each (Figure 2 and Table 1).
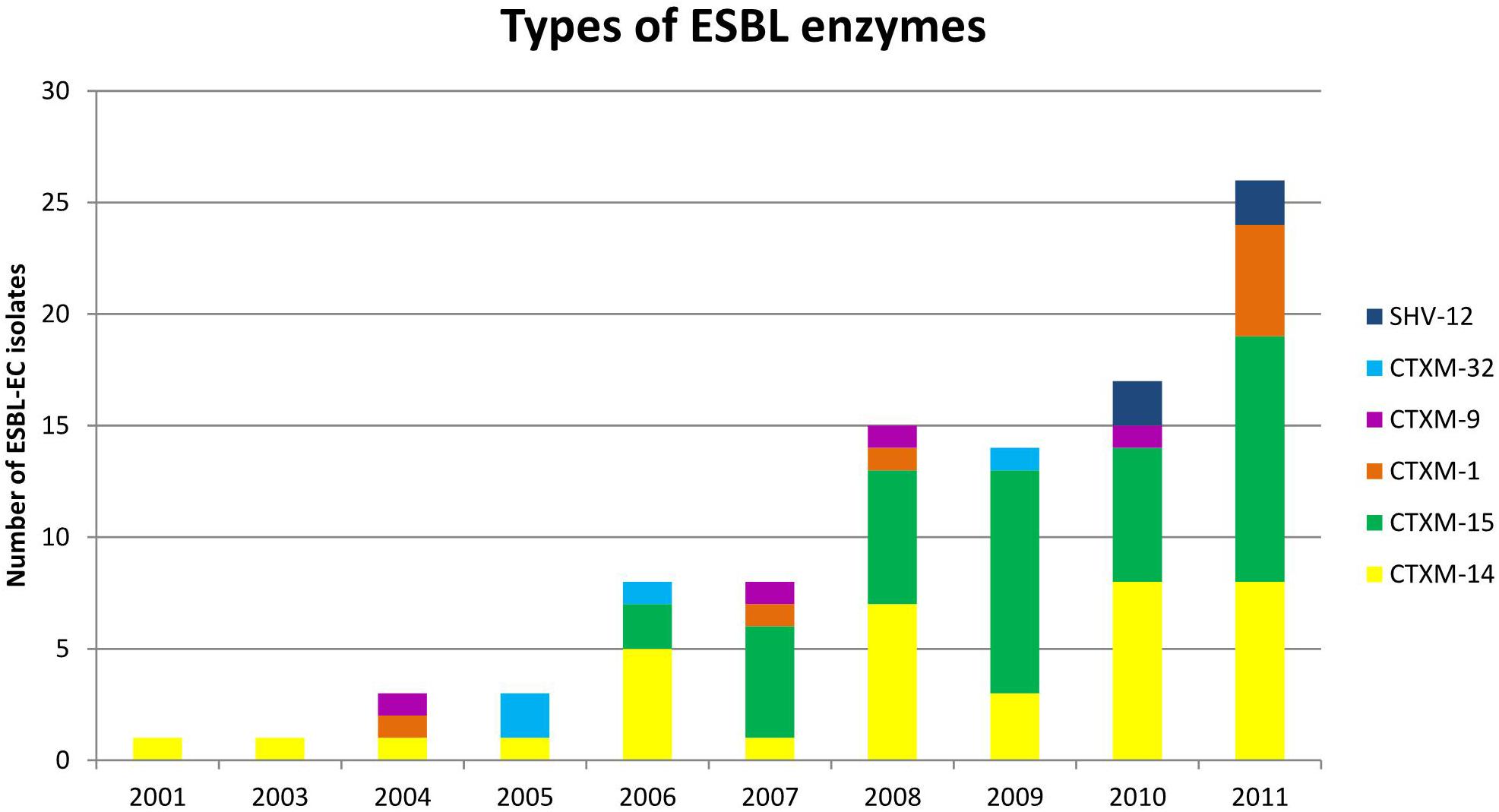
Figure 2. Distribution of extended-spectrum β-lactamase (ESBL) enzymes. Of the 96 ESBL-EC bloodstream isolates, 40 were positive for CTX-M-15, 36 for CTX-M-14, eight for CTX-M-1, and four for CTX-M-9, CTX-M-32, and SHV-12 each.
Phylogenetic Groups
The commonest phylogenetic group was B2 (44 isolates), followed by B1 (18 isolates), A and E (11 isolates from each), F (six isolates), C (five isolates), and D (one isolate).
The majority of CTX-M-15-producing (87.5%, 35/40) isolates belonged to phylogenetic group B2. In contrast, only five (14%) of 36 CTX-M-14-producing isolates were assigned to this phylogenetic group (P < 0.001) (Table 1).
Multilocus Sequence Types
The 96 ESBL-EC isolates belonged to 36 different STs (Table 1). The commonest was ST131 (41 isolates), followed by ST58 (four isolates), ST354 (four isolates), ST393 (four isolates), ST405 (four isolates), and ST359 (three isolates). Most CTX-M-15 isolates (87.5%, 35/40) were ST131, whereas the CTX-M-14 isolates belonged to 23 different STs, none of which accounted for >12% and only 3 (8.3%) of 36 isolates were ST131 (Table 1).
Clonotypes
CH typing identified 40 clonotypes, with CH40-30 being the most prevalent, which was present in 35 ST131 CTX-M-15-producing isolates. Other prevalent clonotypes were: CH41-35 (three ST359 isolates), CH40-22 (three ST131 isolates), CH106-54 (four ST393 isolates), CH37-27 (four ST405 isolates), and CH88-58 (four ST354 isolates) (Table 1).
Serotypes
The 96 ESBL-EC isolates belonged to 21 O serogroups and expressed 17 H flagellar antigens. However, 39 ST131 isolates belonged to serotype O25b:H4. Other prevalent serotypes were: O15:H1 (four ST393 isolates), O9:H4 (three CC46 isolates), O7:H4 (two ST93 isolates), O8:H7 (two ST1642 isolates), O101:H10 (two CC10 isolates), and O153:H34 (two ST354 isolates) (Table 1).
Virulence Genes
The ST131 isolates showed a higher virulence score (range 11 to 21; mean 13.49) compared with the non-ST131 isolates (range 1 to 18; mean 8.31) (P < 0.001).
Fourteen virulence genes (afa/draBC, afaFM955459, yfcV, sat, cnf1, iucD, iutA, fyuA, chuA, kpsM II, kpsM II-K2, malX, usp, and ompT) were significantly associated with ST131 isolates, whereas only five virulence genes (fimAvMT78, hlyF, vat, iroN, cvaC, iss, and traT) were significantly associated with non-ST131 isolates.
Of the 96 ESBL-EC isolates, 65.6% were classified as ExPEC and 44.8% classified as UPEC. The prevalence of ExPEC (97.6 vs. 41.8%) (P < 0.001) and UPEC (90.2 vs. 10.9%) (P < 0.001) status were higher within ST131 isolates than within non-ST131 isolates (Table 2).
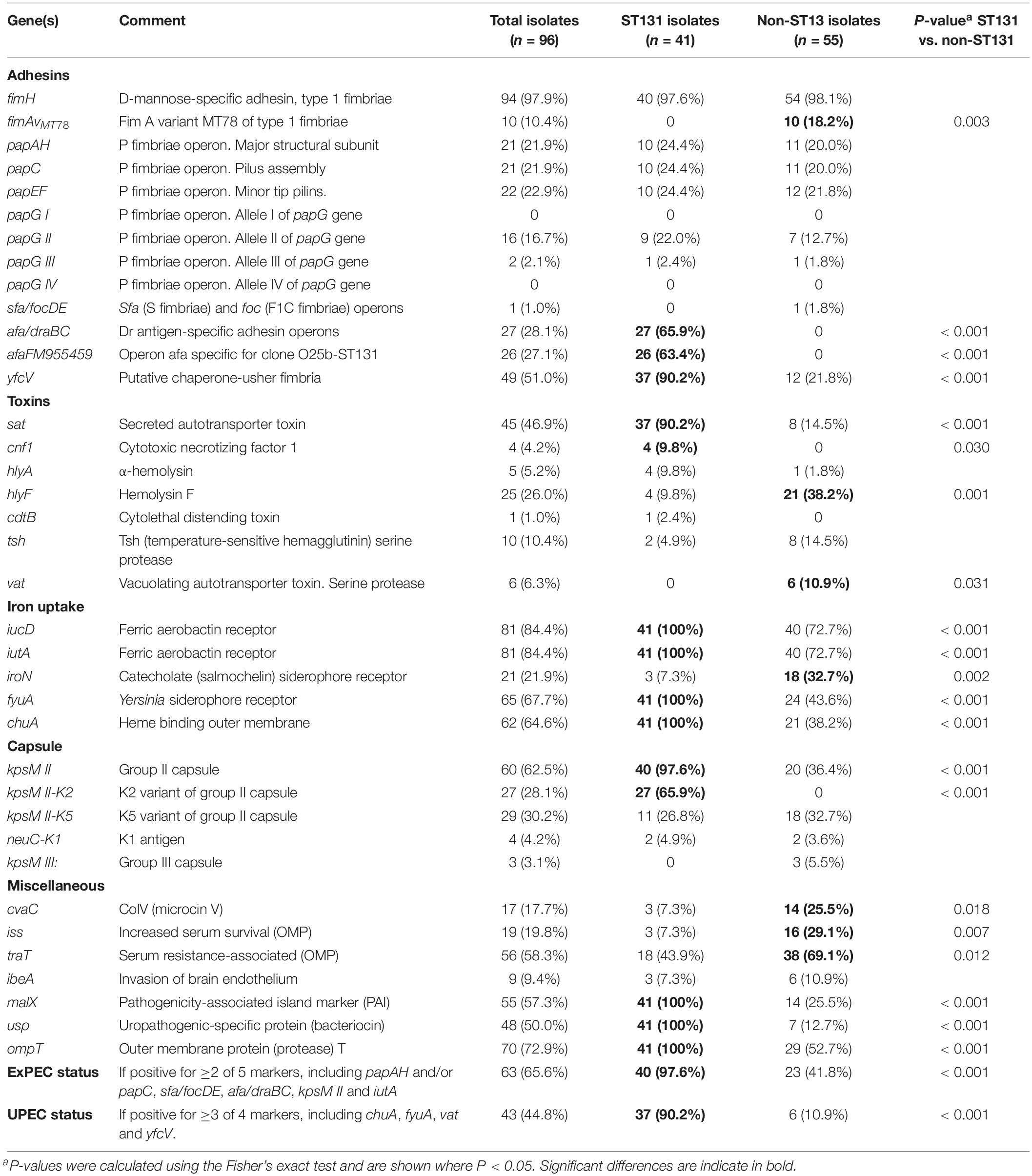
Table 2. Virulence genes in ST131 and non-ST131 bloodstream isolates.
H30Rx Subclone and Virotypes of ST131 Isolates
The 35 ST131 O25b:H4 CTX-M-15-producing isolates belonged to the H30Rx subclone. Twenty-six of the 35 H30Rx isolates showed the virotype A, four isolates the virotype E and five isolates the virotype F. In contrast, the three isolates of H22 subclone showed the virotypes D2 (one isolate) and D4 (two isolates). Besides, one of the two isolates belonging to H41 subclone showed the virotype C3, while the other was non-typeable (NT).
In 2011, there was a very drastic change in the virotypes, due to the emergence of the virotypes E (sat, papGII, cnf1, hlyA, and kpsMII-K5) and F (sat, papGII, and kpsMII-K5) which have displaced the virotype A (afa/draBC, afa operon FM955459, sat, and kpsMII-K2). Thus 8 (66.7%) of the 12 ST131 isolates obtained in 2011 belonged to virotypes E and F vs. 1 (3.4%) of 29 ST131 isolates from 2006 to 2010 (P < 0.001). By contrast, while in the first period 23 (79.3%) of 29 ST131 isolates belonged to virotype A in the last year represented only 3 (25%) of 12 isolates (P = 0.002) (Figure 3).
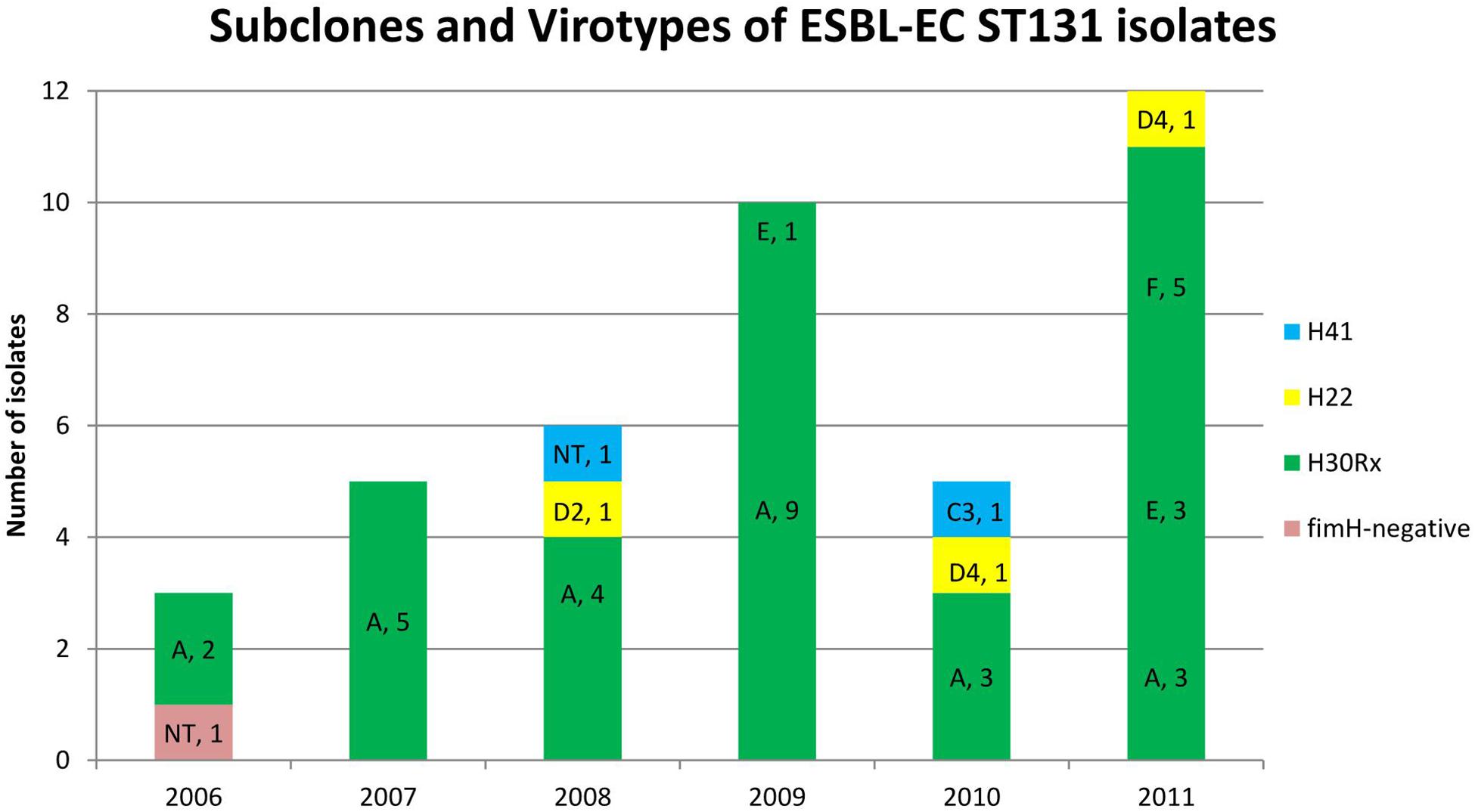
Figure 3. Distribution of subclones and virotypes of extended-spectrum β-lactamase-producing E. coli (ESBL-EC) ST131 bloodstream isolates. A drastic change in ST131 virotypes happened in 2011 due to the emergence of the virotypes E and F, which displaced virotype A.
Macrorestriction Profiles by PFGE
Supplementary Figures S1, S2 show a dendrogram with the XbaI macrorestriction profiles of the ST131 and non-ST131 which showed a similarity of 61.3 and 50.2%, respectively. A high genetic diversity was detected among the 40 ST131 isolates, showing 31 different PFGE profiles. However, the isolates remained distributed within three virotype-specific clusters of 75% (four virotype E isolates), 84.2% (25 virotype A isolates) and 94.6% identity (five virotype F isolates). As expected, the 55 non-ST131 isolates showed even more heterogeneity, with 55 different profiles. However, the majority of isolates belonging to the same STs, clonotypes and O:H serotypes grouped together in the dendrogram.
Discussion
The management of BSIs due to E. coli has been complicated due to antimicrobial resistance emergence, especially since 2000. A low proportion of the bacteraemia E. coli isolates recovered in our hospital between 1989 and 1992 were resistant to extended-spectrum cephalosporins (ESC) (0.5%) and fluoroquinolones (FQs) (1%), and none of the 213 E. coli isolates was multidrug resistant (MDR). However, a significant number turned out to be resistant (ESC 8%, FQs 31%, and MDR 16%) within 614 bacteraemia E. coli isolates obtained between 2010 and 2011 (unpublished data).
Escherichia coli BSIs are increasing in Europe and worldwide mainly due to the increased resistance to antibiotics, and especially to the expansion of high-risk clones such as ST131 (Tumbarello et al., 2010; de Kraker et al., 2013; Vihta et al., 2018). Since 2006, it is evident that the prevalence of ESBL-EC has raised in our hospital. This increase has been due to the spread of the multidrug-resistant H30Rx subclone associated with the production of CTX-M-15. Thus, the number of ESBL-EC isolates increased from 1.0% during 2000–2005 to 5.5% during 2006–2011. While during the first period 0% of the ESBL-EC isolates belonged to H30Rx subclone, in the second period this subclone represented 39.8%. A similar situation has been reported in other hospitals worldwide. Specifically, in a centralized Canadian region (Calgary), Peirano et al. (2012) investigated the prevalence and molecular characteristics of ESBL-EC bloodstream isolates obtained from the year 2000 to 2010 and found that these isolates increased significantly in the last years of the study (2% during 2000–2006 vs. 8% during 2007–2010). ST131 was first described in 2001, and its prevalence remained stable until 2006. However, since 2007, the prevalence of ST131 isolates increased significantly. Like in our study, most isolates from Canada produced either CTX-M-15 or CTX-M-14 (Peirano et al., 2012).
In the present study, while most of the 40 CTX-M-15-producing isolates belonged to ST131, the 36 CTX-M-14 positive isolates were distributed within 23 different STs. Similar results were observed by Merino et al. (2016) who reported a prevalence of 9.2% (39/425) of ESBL-EC among isolates causing bacteraemia of urinary origin in eight Spanish hospitals during 2010 and 2011. Of the 39 ESBL-EC isolates, 21 produced CTX-M-15 and 11 CTX-M-14. Fifteen STs were identified, but as in our study, the ST131 H30Rx subclone was predominant among CTX-M-15 isolates (20 of 21 isolates). Another seven STs (ST69, ST156, ST359, ST405, ST410, ST453, and ST609) reported by Merino et al. (2016) were also identified in the present study. However, the phylogroup distribution of our study differed from the one showed by Merino et al. (2016): A (11.5 vs. 7.7%), B1 (18.8 vs. 23.1%), B2 (45.8 vs. 53.8%), C (5.2 vs. 5.1%), D (1.0 vs. 10.3%), E (11.5 vs. 0%), and F (6.3 vs. 0%).
In our investigation, the CH typing identified 40 clonotypes, subdiving ST131 isolates in three subclones: CH40-30 (35 H30-Rx isolates of subclade C2), CH40-22 (three H22 isolates of clade B), and CH40-41 (two H41 isolates of clade A). Recently, Roer et al. (2018) presented a new Web tool for CH typing. To determine the resolution of it, 243 E. coli isolates were analyzed. Those isolates were resistant to third-generation cephalosporins and obtained from Danish patients with BSIs. A total of 48 different STs were identified, with ST131 the most common (50.2% of isolates; 95 CH40-30, 14 CH40-27, 11 CH40-41, one CH40-22, and one CH40-35). In addition to ST131, other 15 STs (ST23, ST58, ST69, ST88, ST93, ST95, ST141, ST345, ST354, ST393, ST405, ST410, ST453, ST617, and ST648) were identified in both studies. Thus, 72.9% of the Spanish isolates presented the same STs as 72.8% of the Danish isolates. Besides, the majority of the Spanish and Danish bloodstream isolates of the same ST displayed as well the same clonotype. Also, many of the STs found among our isolates were identified within the ESBL-EC isolates collected from BSIs in Japan (ST10, ST23, ST58, ST69, ST93, ST95, ST131, ST156, ST354, ST362, ST393, ST405, ST602, and ST648) (Matsumura et al., 2013), in Korea (ST10, ST69, ST95, ST131, ST354, ST393, ST405, ST410, ST453, ST617, ST648, and ST1642) (Kang et al., 2013), and in China (ST10, ST23, ST58, ST69, ST95, ST131, ST393, ST405, ST410, ST602, ST617, ST648, and ST1642) (Wang et al., 2016).
In Japan and other Asian countries, the CTX-M-27-producing C1/H30R-M27 and CTX-M-14-producing C1/H30R-nM27 subclades are more frequently isolated than the CTX-M-15-producing C2/H30Rx subclade (Matsumura et al., 2017b). We have not detected these two subclades within the collection of the study reported here (2000-2011). However, the epidemiologic situation seems to be changing in Spain since we have detected the subclade C1/H30R-M27 (five cases) within 92 ESBL-EC obtained from patients with urinary tract infections (84 cases) and other extraintestinal infections of our hospital in 2015, being the most frequent after the C2/H30Rx subclade (27 cases) (Flament-Simon et al., 2019).
The ST131 bloodstream isolates showed a higher virulence score than non-ST131 isolates. Furthermore, the prevalence of the ExPEC and UPEC status was also higher among the ST131 isolates than in the non-ST131 isolates, supporting the idea of a greater virulence potential of ST131, confirming the findings of previous studies (Blanco et al., 2011, 2013; Dahbi et al., 2014; Merino et al., 2016). However, the success of ST131 isolates cannot be explained exclusively by its high number of virulence genes. ST131 isolates are carriers of both virulence and resistance genes, which is very rare in classical extraintestinal pathogenic E. coli isolates (Nicolas-Chanoine et al., 2014). In addition, most ESBL-producing ST131 isolates are resistant to both extended-spectrum cephalosporins and fluoroquinolones (Colpan et al., 2013; Johnson et al., 2013). Diverse fitness cost associated with high-level resistance to fluoroquinolones was reported to contribute to the selection of the international clones of methicillin-resistant Staphylococcus aureus, Clostridium difficile ESBL-producing Klebsiella pneumoniae and E. coli (Fuzi et al., 2017). It is also important to highlight that the major reservoir of human extraintestinal pathogenic E. coli is the human digestive tract and that ST131 is the most competitive of the phylogenetic group B2 clones known to colonize the human digestive tract (Blanc et al., 2014; Nicolas-Chanoine et al., 2014). Recently, McNally et al. (2019) after analyzing the genome of 1,094 systematically sampled bacteremia ST131 isolates from the British Society for Antimicrobial Chemotherapy (BSAC) collection, found that clade C had accumulated a significantly elevated allelic diversity, particularly enriched for genes involved in anaerobic metabolism as well as other loci important for colonization of the human host by ExPEC.
In the present study, most of the isolates ST131 showed the virotype A. The international pulsotype L, which was responsible for the significant increase in BSIs in Calgary since 2007, also belonged to the virotype A and subclone H30Rx (Peirano et al., 2014). However, in many countries, the virotype C is the most frequent among ESBL-EC isolates belonging to H30Rx subclone (Olesen et al., 2014; Peirano et al., 2014). The isolates of virotypes E and F obtained during the last year (2011) of our study may possibly represent new sublineages of the ST131 clonal group.
A high genetic diversity was detected by PFGE. However, the ST131 isolates grouped in three specific clusters of virotypes A, E, and F suggesting a clonal basis for the virotypes (Blanco et al., 2013). In future studies it would be very interesting to determine the whole genome sequence of the ST131 isolates belonging to virotypes A, E and F to know if they really belong to different branches within the H30Rx C2 subclade.
In the present study we have not detected any carbapenem-resistant E. coli, but O25b:H4-ST131- H30-Rx isolates of virotype E and co-producing CTX-M-15 and OXA-48 were recently reported in another Spanish hospital (HUCA, Oviedo) near our region (de Toro et al., 2017).
The present study has certain limitations such as it was performed in a single hospital, non-ESBL ST131 isolates were not included, lack of whole genome sequencing (WGS) analysis or the fact that the last isolates were obtained during 2011.
Conclusion
ST131 was the sequence type most frequently detected; moreover, the H30Rx subclone was responsible for the significant increase of ESBL-EC bloodstream isolates since 2006. In the year 2011 two new virotypes (E and F) of H30Rx subclone have emerged. Future molecular studies are needed to understand the dynamics of expansion of this successful high-risk subclone in order to prevent its spread and establish the importance of the two new virotypes.
Data Availability
The raw data supporting the conclusions of this manuscript will be made available by the authors, without undue reservation, to any qualified researcher (Supplementary Table S2).
Author Contributions
RM, SF-S, VG, MA, AM, CL, IG-M, DD-J, JEB, and MB undertook the laboratory work. JB conceived the concept for the research and designed the experiments. All authors provided critical input and contributed to the manuscript writing and approved its final version.
Funding
This study was supported by the following projects: PI16/01477 from the Plan Estatal de I+D+I 2013-2016, Instituto de Salud Carlos III (ISCIII), Subdirección General de Evaluación y Fomento de la Investigación, and Fondo Europeo de Desarrollo Regional (FEDER); AGL2013-47852-R from the Spanish Ministerio de Economía y Competitividad (MINECO) and FEDER; AGL2016-79343-R from the Spanish Agencia Estatal de Investigación (AEI) and FEDER; and CN2012/303 and ED431C2017/57 from the Consellería de Cultura, Educación e Ordenación Universitaria, (Xunta de Galicia) and FEDER.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
RM acknowledges the grant of the Agencia Española de Cooperación Internacional (AECI) (Ministerio de Asuntos Exteriores y de Cooperación). SF-S acknowledges the FPU programme for the grant (FPU15/02644) from the Secretaría General de Universidades, Spanish Ministerio de Educación, Cultura y Deporte. IG-M and VG acknowledge the Consellería de Cultura, Educación e Ordenación Universitaria, Xunta de Galicia for his predoctoral grant (ED481A-2015/149) and her postdoctoral grant (ED481B2018/018), respectively.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.01530/full#supplementary-material
Footnotes
References
Adams-Sapper, S., Diep, B. A., Perdreau-Remington, F., and Riley, L. W. (2013). Clonal composition and community clustering of drug-susceptible and -resistant Escherichia coli isolates from bloodstream infections. Antimicrob. Agents Chemother. 57, 490–497. doi: 10.1128/AAC.01025-12
Banerjee, R., Robicsek, A., Kuskowski, M. A., Porter, S., Johnston, B. D., Sokurenko, E., et al. (2013). Molecular epidemiology of Escherichia coli sequence type 131 and Its H30 and H30-Rx subclones among extended-spectrum-β-lactamase-positive and -negative E. coli clinical isolates from the Chicago Region, 2007 to 2010. Antimicrob. Agents Chemother. 57, 6385–6388. doi: 10.1128/AAC.01604-13
Ben Zakour, N. L., Alsheikh-Hussain, A. S., Ashcroft, M. M., Khanh Nhu, N. T., Roberts, L. W., Stanton-Cook, M., et al. (2016). Sequential acquisition of virulence and fluoroquinolone resistance has shaped the evolution of Escherichia coli ST131. mBio 7:e00347-16. doi: 10.1128/mBio.00347-16
Blanc, V., Leflon-Guibout, V., Blanco, J., Haenni, M., Madec, J. Y., Rafignon, G., et al. (2014). Prevalence of day-care centre children (France) with faecal CTX-M-producing Escherichia coli comprising O25b:H4 and O16:H5 ST131 strains. J. Antimicrob. Chemother. 69, 1231–1237. doi: 10.1093/jac/dkt519
Blanco, J., Mora, A., Mamani, R., López, C., Blanco, M., Dahbi, G., et al. (2011). National survey of Escherichia coli causing extraintestinal infections reveals the spread of drug-resistant clonal groups O25b:H4-B2-ST131, O15:H1-D-ST393 and CGA-D-ST69 with high virulence gene content in Spain. J. Antimicrob. Chemother. 66, 2011–2021. doi: 10.1093/jac/dkr235
Blanco, J., Mora, A., Mamani, R., López, C., Blanco, M., Dahbi, G., et al. (2013). Four main virotypes among extended-spectrum-β-lactamase-producing isolates of Escherichia coli O25b:H4-B2-ST131: bacterial, epidemiological, and clinical characteristics. J. Clin. Microbiol. 51, 3358–3367. doi: 10.1128/JCM.01555-13
Blanco, M., Alonso, M. P., Nicolas-Chanoine, M. H., Dahbi, G., Mora, A., Blanco, J. E., et al. (2009). Molecular epidemiology of Escherichia coli producing extended-spectrum β-lactamases in Lugo (Spain): dissemination of clone O25b:H4-ST131 producing CTX-M-15. J. Antimicrob. Chemother. 63, 1135–1141. doi: 10.1093/jac/dkp122
Chung, H. C., Lai, C. H., Lin, J. N., Huang, C. K., Liang, S. H., Chen, W. F., et al. (2012). Bacteremia caused by extended-spectrum-β-lactamase-producing Escherichia coli sequence type ST131 and non-ST131 clones: comparison of demographic data, clinical features, and mortality. Antimicrob. Agents Chemother. 56, 618–622. doi: 10.1128/AAC.05753-11
Clermont, O., Christenson, J. K., Denamur, E., and Gordon, D. M. (2013). The Clermont Escherichia coli phylo-typing method revisited: improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 5, 58–65. doi: 10.1111/1758-2229.12019
Clermont, O., Lavollay, M., Vimont, S., Deschamps, C., Forestier, C., Branger, C., et al. (2008). The CTX-M-15-producing Escherichia coli diffusing clone belongs to a highly virulent B2 phylogenetic subgroup. J. Antimicrob. Chemother. 61, 1024–1028. doi: 10.1093/jac/dkn084
Colpan, A., Johnston, B., Porter, S., Clabots, C., Anway, R., and Thao, L. (2013). Escherichia coli sequence type 131 (ST131) subclone H30 as an emergent multidrug-resistant pathogen among US veterans. Clin. Infect. Dis. 57, 1256–1265. doi: 10.1093/cid/cit503
Dahbi, G., Mora, A., Mamani, R., López, C., Alonso, M. P., Marzoa, J., et al. (2014). Molecular epidemiology and virulence of Escherichia coli O16:H5-ST131: comparison with H30 and H30-Rx subclones of O25b:H4-ST131. Int. J. Med. Microbiol. 304, 1247–1257. doi: 10.1016/j.ijmm.2014.10.002
Day, M. J., Doumith, M., Abernethy, J., Hope, R., Reynolds, R., Wain, J., et al. (2016). Population structure of Escherichia coli causing bacteraemia in the UK and Ireland between 2001 and 2010. J. Antimicrob. Chemother. 71, 2139–2142. doi: 10.1093/jac/dkw145
de Kraker, M. E., Jarlier, V., Monen, J. C., Heuer, O. E., van de Sande, N., and Grundmann, H. (2013). The changing epidemiology of bacteraemias in Europe: trends from the European antimicrobial resistance surveillance system. Clin. Microbiol. Infect. 19, 860–868. doi: 10.1111/1469-0691.12028
de Toro, M., Fernández, J., García, V., Mora, A., Blanco, J., de la Cruz, F., et al. (2017). Whole genome sequencing, molecular typing and in vivo virulence of OXA-48-producing Escherichia coli isolates including ST131 H30-Rx, H22 and H41 subclones. Sci. Rep. 21:2103. doi: 10.1038/s41598-017-12015-0
Flament-Simon, S. C., García, V., Duprilot, M., Mayer, N., Alonso, M. P., García-Meniño, I., et al. (2019). “High prevalence of ST131 subclades C2 and C1-M27 producing CTX-M-15 and CTX-M-27, respectively, among ESBL-producing Escherichia coli causing extraintestinal infections in Spain and France during 2015,” in Proceedings of the 29th European Congress of Clinical Microbiology and Infectious Diseases, (Amsterdam),
Fuzi, M., Szabo, D., and Csercsik, R. (2017). Double-serine fluoroquinolone resistance mutations advance major international clones and lineages of various multi-drug resistant bacteria. Front. Microbiol. 8:2261. doi: 10.3389/fmicb.2017.02261
Guinée, P. A. M., Jansen, W. H., Wadström, T., and Sellwood, R. (1981). “Escherichia coli associated with neonatal diarrhoea in piglets and calves,” in Laboratory Diagnosis in Neoanatal Calf and Pig Diarrhoea. Current Topics in Veterinary and Animal Science, Vol. 13, eds P. W. de Leeuw and P. A. M. Guinée (Leiden: Martinus Nijhoff Publishers), 126–162. doi: 10.1007/978-94-009-8328-1_18
Harris, P. N. A., Ben Zakour, N. L., Roberts, L. W., Wailan, A. M., Zowawi, H. M., Tambyah, P. A., et al. (2018). Whole genome analysis of cephalosporin-resistant Escherichia coli from bloodstream infections in Australia, New Zealand and Singapore: high prevalence of CMY-2 producers and ST131 carrying blaCTX-M-15 and blaCTX-M-27. J. Antimicrob. Chemother. 73, 634–642. doi: 10.1093/jac/dkx466
Horner, C., Fawley, W., Morris, K., Parnell, P., Denton, M., and Wilcox, M. (2014). Escherichia coli bacteraemia: 2 years of prospective regional surveillance (2010-12). J. Antimicrob. Chemother. 69, 91–100. doi: 10.1093/jac/dkt333
Jarlier, V., Nicolas, M. H., Fournier, G., and Philippon, A. (1988). Extended broad-spectrum β-lactamases conferring transferable resistance to newer β-lactam agents in Enterobacteriaceae: hospital prevalence and susceptibility patterns. Rev. Infect. Dis. 10, 867–878. doi: 10.1093/clinids/10.4.867
Johnson, J. R., Porter, S., Johnston, B., Kuskowski, M. A., Spurbeck, R. R., Mobley, H. L., et al. (2015). Host characteristics and bacterial traits predict experimental virulence for Escherichia coli bloodstream isolates from patients with urosepsis. Open Forum Infect. Dis. 2:ofv083. doi: 10.1093/ofid/ofv083
Johnson, J. R., Porter, S., Thuras, P., and Castanheira, M. (2017). The pandemic H30 subclone of sequence type 131 (ST131) as the leading cause of multidrug-resistant Escherichia coli infections in the United States (2011-2012). Open Forum Infect. Dis. 4:ofx089. doi: 10.1093/ofid/ofx089
Johnson, J. R., Tchesnokova, V., Johnston, B., Clabots, C., Roberts, P. L., Billig, M., et al. (2013). Abrupt emergence of a single dominant multidrug-resistant strain of Escherichia coli. J. Infect. Dis. 207, 919–928. doi: 10.1093/infdis/jis933
Kallonen, T., Brodrick, H. J., Harris, S. R., Corander, J., Brown, N. M., Martin, V., et al. (2017). Systematic longitudinal survey of invasive Escherichia coli in England demonstrates a stable population structure only transiently disturbed by the emergence of ST131. Genome Res. doi: 10.1101/gr.216606.116 [Epub ahead of print].
Kang, C. I., Cha, M. K., Kim, S. H., Ko, K. S., Wi, Y. M., Chung, D. R., et al. (2013). Clinical and molecular epidemiology of community-onset bacteremia caused by extended-spectrum β-lactamase-producing Escherichia coli over a 6-year period. J. Korean Med. Sci. 28, 998–1004. doi: 10.3346/jkms.2013.28.7.998
Leflon-Guibout, V., Jurand, C., Bonacorsi, S., Espinasse, F., Guelfi, M. C., Duportail, F., et al. (2004). Emergence and spread of three clonally related virulent isolates of CTX-M-15-producing Escherichia coli with variable resistance to aminoglycosides and tetracycline in a French geriatric hospital. Antimicrob. Agents Chemother. 48, 3736–3742. doi: 10.1128/aac.48.10.3736-3742.2004
Matsumura, Y., Nagao, M., Iguchi, M., Yagi, T., Komori, T., Fujita, N., et al. (2013). Molecular and clinical characterization of plasmid-mediated AmpC β-lactamase-producing Escherichia coli bacteraemia: a comparison with extended-spectrum β-lactamase-producing and non-resistant E. coli bacteraemia. Clin. Microbiol. Infect. 19, 161–168. doi: 10.1111/j.1469-0691.2012.03762.x
Matsumura, Y., Pitout, J. D. D., Peirano, G., DeVinney, R., Noguchi, T., Yamamoto, M., et al. (2017b). Rapid identification of different Escherichia coli sequence type 131 clades. Antimicrob. Agents Chemother. 61:e00179-17. doi: 10.1128/AAC.00179-17
Matsumura, Y., Noguchi, T., Tanaka, M., Kanahashi, T., Yamamoto, M., Nagao, M., et al. (2017a). Population structure of Japanese extraintestinal pathogenic Escherichia coli and its relationship with antimicrobial resistance. J. Antimicrob. Chemother. 72, 1040–1049. doi: 10.1093/jac/dkw530
McNally, A., Kallonen, T., Connor, C., Abudahab, K., Aanensen, D. M., Horner, C., et al. (2019). Diversification of colonization factors in a multidrug-resistant Escherichia coli lineage evolving under negative frequency-dependent selection. mBio 10:e00644-19. doi: 10.1128/mBio.00644-19
McNally, A., Oren, Y., Kelly, D., Pascoe, B., Dunn, S., Sreecharan, T., et al. (2016). Combined analysis of variation in core, accessory and regulatory genome regions provides a super-resolution view into the evolution of bacterial populations. PLoS Genet. 12:e1006280. doi: 10.1371/journal.pgen.1006280
Mendis, S. M., Vasoo, S., Johnston, B. D., Porter, S. B., Cunningham, S. A., Menon, S. R., et al. (2018). Clinical and molecular correlates of Escherichia coli bloodstream infection from two geographically diverse centers in Rochester, Minnesota, and Singapore. Antimicrob. Agents Chemother. 62, e937–e918. doi: 10.1128/AAC.00937-18
Merino, I., Shaw, E., Horcajada, J. P., Cercenado, E., Mirelis, B., Pallarés, M. A., et al. (2016). CTX-M-15-H30Rx-ST131 subclone is one of the main causes of healthcare-associated ESBL-producing Escherichia coli bacteraemia of urinary origin in Spain. J. Antimicrob. Chemother. 71, 2125–2130. doi: 10.1093/jac/dkw133
Mora, A., Dahbi, G., López, C., Mamani, R., Marzoa, J., Dion, S., et al. (2014). Virulence patterns in a murine sepsis model of ST131 Escherichia coli clinical isolates belonging to serotypes O25b:H4 and O16:H5 are associated to specific virotypes. PLoS One 9:e87025. doi: 10.1371/journal.pone.0087025
Mora, A., Herrera, A., Mamani, R., López, C., Alonso, M. P., Blanco, J. E., et al. (2010). Recent emergence of clonal group O25b:K1:H4-B2-ST131 ibeA strains among Escherichia coli poultry isolates, including CTX-M-9-producing strains, and comparison with clinical human isolates. Appl. Environ. Microbiol. 76, 6991–6997. doi: 10.1128/AEM.01112-10
Mora, A., López, C., Dabhi, G., Blanco, M., Blanco, J. E., Alonso, M. P., et al. (2009). Extraintestinal pathogenic Escherichia coli O1:K1:H7/NM from human and avian origin: detection of clonal groups B2 ST95 and D ST59 with different host distribution. BMC Microbiol. 9:132. doi: 10.1186/1471-2180-9-132
Nicolas-Chanoine, M. H., Bertrand, X., and Madec, J. Y. (2014). Escherichia coli ST131, an intriguing clonal group. Clin. Microbiol. Rev. 27, 543–574. doi: 10.1128/CMR.00125-13
Nicolas-Chanoine, M. H., Blanco, J., Leflon-Guibout, V., Demarty, R., Alonso, M. P., Caniça, M. M., et al. (2008). Intercontinental emergence of Escherichia coli clone O25:H4-ST131 producing CTX-M-15. J. Antimicrob. Chemother. 61, 273–281. doi: 10.1093/jac/dkm464
Olesen, B., Frimodt-Møller, J., Leihof, R. F., Struve, C., Johnston, B., Hansen, D. S., et al. (2014). Temporal trends in antimicrobial resistance and virulence-associated traits within the Escherichia coli sequence type 131 clonal group and its H30 and H30-Rx subclones, 1968 to 2012. Antimicrob. Agents Chemother. 58, 6886–6895. doi: 10.1128/AAC.03679-14
Peirano, G., van der Bij, A. K., Freeman, J. L., Poirel, L., Nordmann, P., Costello, M., et al. (2014). Characteristics of Escherichia coli sequence type 131 isolates that produce extended-spectrum β-lactamases: global distribution of the H30-Rx sublineage. Antimicrob. Agents Chemother. 58, 3762–3767. doi: 10.1128/AAC.02428-14
Peirano, G., van der Bij, A. K., Gregson, D. B., and Pitout, J. D. (2012). Molecular epidemiology over an 11-year period (2000 to 2010) of extended-spectrum β-lactamase-producing Escherichia coli causing bacteremia in a centralized Canadian region. J. Clin. Microbiol. 50, 294–299. doi: 10.1128/JCM.06025-11
Pitout, J. D., and DeVinney, R. (2017). Escherichia coli ST131: a multidrug-resistant clone primed for global domination. F1000Res. 6:F1000FacultyRev–195. doi: 10.12688/f1000research.10609.1
Price, L. B., Johnson, J. R., Aziz, M., Clabots, C., Johnston, B., Tchesnokova, V., et al. (2013). The epidemic of extended-spectrum-β-lactamase-producing Escherichia coli ST131 is driven by a single highly pathogenic subclone, H30-Rx. mBio 4, e00377-13. doi: 10.1128/mBio.00377-13
Rodríguez-Baño, J., Mingorance, J., Fernández-Romero, N., Serrano, L., López-Cerero, L., Pascual, A., et al. (2012). Virulence profiles of bacteremic extended-spectrum β-lactamase-producing Escherichia coli: association with epidemiological and clinical features. PLoS One 7:e44238. doi: 10.1371/journal.pone.0044238
Roer, L., Hansen, F., Thomsen, M. C. F., Knudsen, J. D., Hansen, D. S., Wang, M., et al. (2017). WGS-based surveillance of third-generation cephalosporin-resistant Escherichia coli from bloodstream infections in Denmark. J. Antimicrob. Chemother. 72, 1922–1929. doi: 10.1093/jac/dkx092
Roer, L., Johannesen, T. B., Hansen, F., Stegger, M., Tchesnokova, V., Sokurenko, E., et al. (2018). CHTyper, a Web tool for subtyping of extraintestinal pathogenic Escherichia coli based on the fumC and fimH alleles. J. Clin. Microbiol. 56:e00063-18. doi: 10.1128/JCM.00063-18
Spurbeck, R. R., Dinh, P. C. Jr., Walk, S. T., Stapleton, A. E., Hooton, T. M., Nolan, L. K., et al. (2012). Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract. Infect. Immun. 80, 4115–4122. doi: 10.1128/IAI.00752-12
Stoesser, N., Sheppard, A. E., Pankhurst, L., De Maio, N., Moore, C. E., Sebra, R., et al. (2016). Evolutionary history of the global emergence of the Escherichia coli epidemic clone ST131. mBio 7:e02162. doi: 10.1128/mBio.02162-15
Tchesnokova, V., Billig, M., Chattopadhyay, S., Linardopoulou, E., Aprikian, P., Roberts, P. L., et al. (2013). Predictive diagnostics for Escherichia coli infections based on the clonal association of antimicrobial resistance and clinical outcome. J. Clin. Microbiol. 51, 2991–2999. doi: 10.1128/JCM.00984-13
Tumbarello, M., Spanu, T., Di Bidino, R., Marchetti, M., Ruggeri, M., Trecarichi, E. M., et al. (2010). Costs of bloodstream infections caused by Escherichia coli and influence of extended-spectrum-beta-lactamase production and inadequate initial antibiotic therapy. Antimicrob. Agents Chemother. 54, 4085–4091. doi: 10.1128/AAC.00143-10
Vihta, K. D., Stoesser, N., Llewelyn, M. J., Quan, T. P., Davies, T., Fawcett, N. J., et al. (2018). Trends over time in Escherichia coli bloodstream infections, urinary tract infections, and antibiotic susceptibilities in Oxfordshire, UK, 1998-2016: a study of electronic health records. Lancet Infect. Dis. 18, 1138–1149. doi: 10.1016/S1473-3099(18)30353-0
Wang, S., Zhao, S. Y., Xiao, S. Z., Gu, F. F., Liu, Q. Z., Tang, J., et al. (2016). Antimicrobial resistance and molecular epidemiology of Escherichia coli causing bloodstream infections in three hospitals in Shanghai, China. PLoS One 11:e0147740. doi: 10.1371/journal.pone.0147740
Weissman, S. J., Johnson, J. R., Tchesnokova, V., Billig, M., Dykhuizen, D., Riddell, K., et al. (2012). High-resolution two-locus clonal typing of extraintestinal pathogenic Escherichia coli. Appl. Environ. Microbiol. 78, 1353–1360. doi: 10.1128/AEM.06663-11
Keywords: clonotypes, E. coli, ESBL, ExPEC, H30Rx subclone, ST131 clonal group
Citation: Mamani R, Flament-Simon SC, García V, Mora A, Alonso MP, López C, García-Meniño I, Díaz-Jiménez D, Blanco JE, Blanco M and Blanco J (2019) Sequence Types, Clonotypes, Serotypes, and Virotypes of Extended-Spectrum β-Lactamase-Producing Escherichia coli Causing Bacteraemia in a Spanish Hospital Over a 12-Year Period (2000 to 2011). Front. Microbiol. 10:1530. doi: 10.3389/fmicb.2019.01530
Received: 19 February 2019; Accepted: 18 June 2019;
Published: 16 July 2019.
Edited by:
Miklos Fuzi, Semmelweis University, HungaryReviewed by:
Sebastian Guenther, University of Greifswald, GermanyXavier Bertrand, Centre Hospitalier Universitaire de Besançon, France
Rowland Cobbold, The University of Queensland, Australia
Copyright © 2019 Mamani, Flament-Simon, García, Mora, Alonso, López, García-Meniño, Díaz-Jiménez, Blanco, Blanco and Blanco. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jorge Blanco, am9yZ2UuYmxhbmNvQHVzYy5lcw==
†These authors have contributed equally to this work