 Kathryn Jane Turnbull1,2,3Ievgen Dzhygyr2,3Søren Lindemose1
Kathryn Jane Turnbull1,2,3Ievgen Dzhygyr2,3Søren Lindemose1 Vasili Hauryliuk2,3,4
Vasili Hauryliuk2,3,4 Mohammad Roghanian1,2,3*
Mohammad Roghanian1,2,3*- 1Centre for Bacterial Stress Response and Persistence, Department of Biology, University of Copenhagen, Copenhagen, Denmark
- 2Department of Molecular Biology, Umeå University, Umeå, Sweden
- 3Laboratory for Molecular Infection Medicine Sweden (MIMS), Umeå University, Umeå, Sweden
- 4Institute of Technology, University of Tartu, Tartu, Estonia
Amino acid starvation in Escherichia coli activates the enzymatic activity of the stringent factor RelA, leading to accumulation of the alarmone nucleotide (p)ppGpp. The alarmone acts as an intercellular messenger to regulate transcription, translation and metabolism to mediate bacterial stress adaptation. The enzymatic activity of RelA is subject to multi-layered allosteric control executed both by ligands – such as “starved” ribosomal complexes, deacylated tRNA and pppGpp – and by individual RelA domains. The auto-regulation of RelA is proposed to act either in cis (inhibition of the enzymatic activity of the N-terminal region, NTD, by regulatory C-terminal region, CTD) or in trans (CTD-mediated dimerization leading to enzyme inhibition). In this report, we probed the regulatory roles of the individual domains of E. coli RelA and our results are not indicative of RelA dimerization being the key regulatory mechanism. First, at growth-permitting levels, ectopic expression of RelA CTD does not interfere with activation of native RelA, indicating lack of regulation via inhibitory complex formation in the cell. Second, in our biochemical assays, increasing RelA concentration does not decrease the enzyme activity, as would be expected in the case of efficient auto-inhibition via dimerization. Third, while high-level CTD expression efficiently inhibits the growth, the effect is independent of native RelA and is mediated by direct inhibition of protein synthesis, likely via direct interaction with the ribosomal A-site. Finally, deletion of the RRM domain of the CTD region leads to growth inhibition mediated by accumulation of (p)ppGpp, suggesting de-regulation of the synthetic activity in this mutant.
Introduction
Bacteria have numerous mechanisms for sensing and adapting to stressful conditions, such as nutrient limitations. The stringent response is a near-ubiquitous bacterial stress response which is mediated by accumulation of two hyper-phosphorylated derivatives of GTP and GDP: guanosine pentaphosphate (pppGpp) and guanosine tetraphosphate (ppGpp), collectively referred to as (p)ppGpp (Hauryliuk et al., 2015). The stringent response and (p)ppGpp have important roles in the regulation of bacterial virulence, survival during host invasion, and antibiotic resistance (Dalebroux et al., 2010; Dalebroux and Swanson, 2012; Hauryliuk et al., 2015).
RelA/SpoT homolog (RSH) are ubiquitous bacterial enzymes responsible for synthesis and degradation of (p)ppGpp (Atkinson et al., 2011; Jimmy et al., 2019). In the majority of beta- and gamma-proteobacteria, including Escherichia coli, the RSHs are represented by two multi-domain enzymes, the namesakes of the protein family – RelA and SpoT. The “long” multi-domain RSHs are comprised of two functional regions: the catalytic N-terminal (NTD) and the regulatory C-terminal (CTD) (Figure 1A; Atkinson et al., 2011). The NTD encompasses the (p)ppGpp hydrolase domain (HD; enzymatically inactive in RelA) as well as a (p)ppGpp synthetase domain (SYNTH). The CTD is comprised of four domains: Thr-tRNA synthetase, GTPase and SpoT domain (TGS domain); helical domain; zing finger domain (ZFD); and the RNA recognition motif (RRM). RelA is the best studied of all of the RSH representatives and its regulation is the focus of this study.
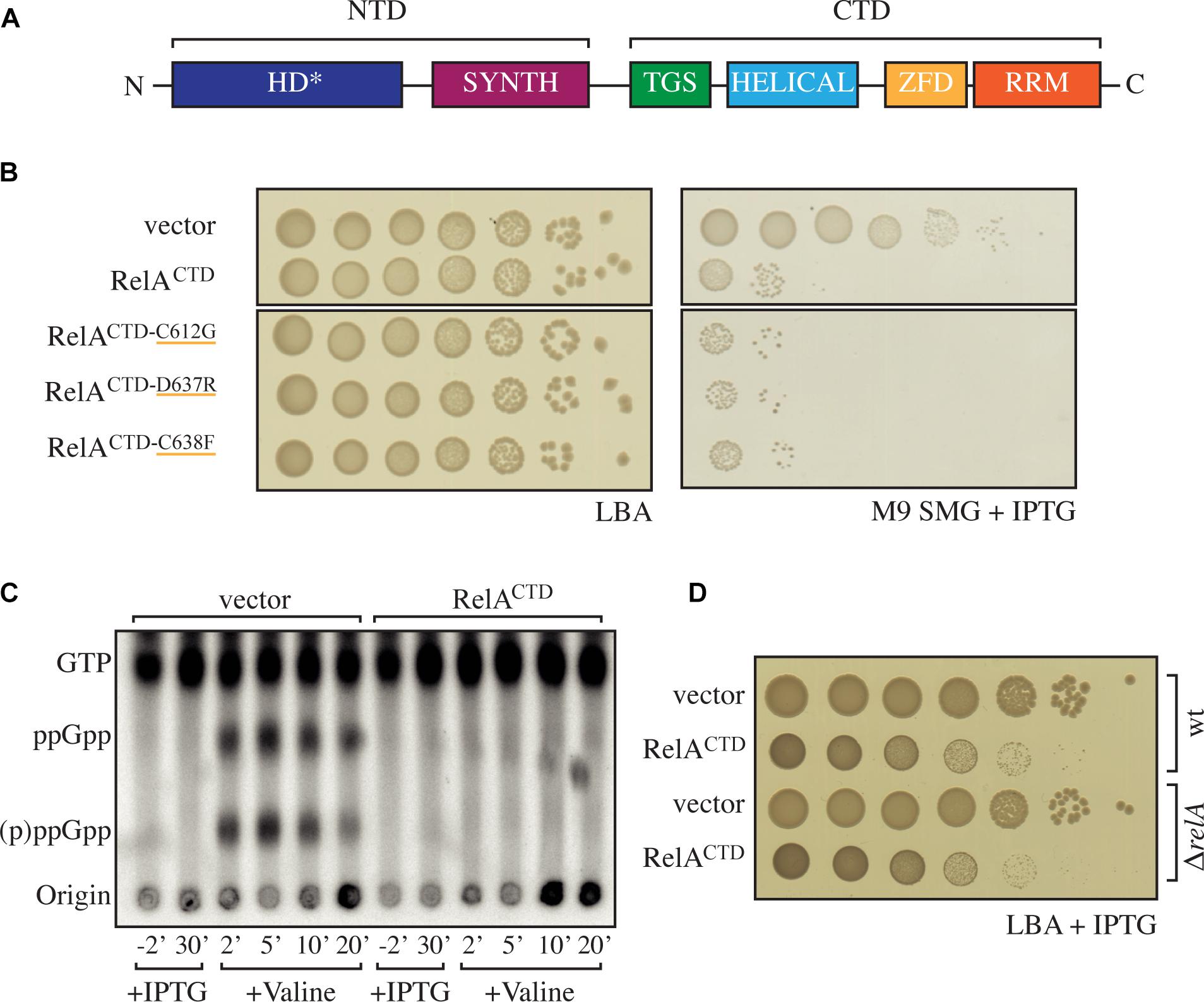
Figure 1. High-level ectopic expression of RelACTD is inhibitory to growth independently of native RelA’s activity. (A) Domain structure of RelA, the enzymatically inactive (p)ppGpp hydrolysis (HD∗) and a functional (p)ppGpp synthesis (SYNTH) NTD domains. TGS (ThrRS, GTPase, and SpoT), Helical, ZFD (Zinc Finger Domain) and RRM (RNA recognition motif) domains comprise the regulatory CTD region. (B) Escherichia coli MG1655 (wt) cells were transformed with high copy IPTG inducible vector, pMG25 (vector), pMG25:relACTD, pMG25:RelACTD–C612G, pMG25:RelACTD–D637R, and pMG25:RelACTD–C638F. The transformed cells were grown at 37°C overnight in M9 minimal medium with 100 μg/ml ampicillin. Ten-fold serial dilutions were made and spotted onto LB agar (LBA) supplemented with 100 μg/ml ampicillin, as a loading control, and onto SMG plates supplemented with 100 μg/ml ampicillin and 1 mM IPTG to induce relA variant expression. (C) Representative audioradiogram of a PEI Cellulose TLC plate showing (p)ppGpp accumulation of E. coli MG1655 carrying pMG25 (vector) or pMG25:relACTD upon valine-induced isoleucine starvation. See section “Materials and Methods” for more details. (D) Overnight cultures of E. coli MG1655 (wt) and MG1655ΔrelA (ΔrelA) transformed with pMG25 (vector) or pMG25:relACTD, grown in LB with 100 μg/ml ampicillin. Ten-fold serial dilutions of were made and spotted onto LB agar supplemented with 1 mM IPTG.
While RelA has no detectable hydrolytic activity (Shyp et al., 2012), it possesses a strong ribosome-dependent (p)ppGpp synthetic activity that is activated by deacylated tRNA in the ribosomal A-site – ribosomal “starved” complexes (Haseltine and Block, 1973). In the absence of the starved ribosomal complexes, RelA displays a very low (p)ppGpp synthetic activity that is not stimulated by deacylated tRNA (Wendrich et al., 2002; Knutsson Jenvert and Holmberg Schiavone, 2005; Kudrin et al., 2018). Deletion of the CTD results in (p)ppGpp accumulation in the cell (Schreiber et al., 1991; Svitil et al., 1993; Gropp et al., 2001), suggesting that the CTD is essential for regulation via repression of the synthetic activity of RelA.
Two opposing models have been presented for the regulatory role of the CTD. The first was proposed by three independent groups (Gropp et al., 2001; Yang and Ishiguro, 2001; Jain et al., 2006) suggesting that under optimal growth conditions, RelA exists as a synthetically inactive dimer and upon amino acid starvation the enzyme is activated in the monomeric form. The dimerization was proposed to be mediated by the Zinc Finger Domain [ZDF; alternatively referred to as CC, conserved cysteine as per (Atkinson et al., 2011)], and mutations C612G, D637R, and C638F were proposed to abrogate dimerization (Gropp et al., 2001; Jain et al., 2006). Importantly, the ZDF is crucial for RelA interaction with ribosome, forming specific contacts with the A-site Finger rRNA element that acts as an anchoring point for RelA:ribosome complex formation (Arenz et al., 2016; Brown et al., 2016; Loveland et al., 2016; Kudrin et al., 2018). The second model was proposed by a study of a bi-functional “long” RSH enzyme Rel from Streptococcus equisimilis (RelSeq) which suggested that the CTD of Rel represses the (p)ppGpp synthesis by NTD via intramolecular contacts in cis (Mechold et al., 2002).
Here, we provide evidence that E. coli RelA is predominantly regulated through intramolecular (autoinhibition of the NTD by the CTD in cis) rather than intermolecular (dimerization, autoinhibition in trans) interactions.
Materials and Methods
Bacterial Strains, Plasmids and Growth Conditions
The strains and plasmids used in this study are listed in Table 1. Oligonucleotides used in this study are provided in Supplementary Table S1. For growth assays in Luria–Bertani (LB, BD Difco – Thermo Fisher Scientific) medium or on LB agar, the relevant strains were grown overnight in LB medium supplemented with antibiotic selection. For growth assays on SMG (Serine, Methionine, and Glycine) plates (Uzan and Danchin, 1978), the strains were grown overnight in M9-glucose minimal medium (1 × M9 salts, 1 μg μl–1 thiamine, 1 mM MgCl2, 0.1 mM CaCl, pH 7.4; 10 × M9 salts: 80 g Na2HPO4, 30 g KH2PO4, 5 g NaCl per 1 L) supplemented with antibiotics for plasmid selection. The SMG plates were made by supplementing M9-glucose minimal medium with 0.7% agarose, 100 μg/ml of Serine, Methionine, and Glycine and appropriate antibiotic selection. Expression from LacI-regulated promoters on solid or in liquid media was induced by the addition of 1 mM of IPTG. Bacterial cultures were routinely grown overnight at 37°C, and plates were incubated at 37°C for 18 h. 30 μg/ml ampicillin was used to select for low copy number plasmid pNDM220 and its derivatives, and 100 μg/ml ampicillin was used to select for the high copy number plasmid pMG25 and its derivatives. Growth assays present in figure panels originate from a single plate that were brought together, where applicable, this is indicated by a gap. All growth curves were performed in flasks, grown at 37°C, with shaking at 160 rpm, unless otherwise indicated.

Table 1. Bacterial strains and plasmids used in this study.
(p)ppGpp Measurements by Thin Layer Chromatography (TLC)
To determine the (p)ppGpp levels in cells we modified the method from Cashel (1994) and Tian et al. (2016). In brief, overnight cultures were diluted 100-fold in 5 ml of MOPS glucose minimal medium supplemented with all five nucleobases (10 μg/ml of each, Sigma-Alderich) and incubated at 37°C with shaking. All of the strains had similar growth rates in this medium. At an optical density at 600 nm (OD600) of ≈0.5, cells were diluted 10-fold to an OD600 of ≈0.05 and were left to grow with shaking at 37°C with H332PO4 (100 μCi/ml). When looking at the effect of RelACTD expressed from the low copy number plasmid (Figure 2C and Supplementary Figure S4), 1 mM IPTG was also added at this point. The cells were then grown for ≈2 generations (OD600 ≈0.2) before amino acid starvation was induced by the addition of either 500 μg/ml valine (Sigma-Aldrich) or 400 μg/ml serine hydroxamate (SHX) (Sigma-Aldrich). Whereas, due to its inhibitory effect on growth, RelACTD was expressed from the high copy number plasmid for 30 min after the cells were grown for ≈2 generations (OD600 ≈0.2). Subsequently, amino acid starvation was induced by the addition of valine (Figure 1C). Fifty-microliter samples were withdrawn before and at various times (see figures) after the induction of amino acid starvation. With the induction of RelA and RelAΔRRM (Figure 4D), after labeling of cells for ≈2 generations, 1 mM IPTG was added and 50 μl samples were taken at −2, 10, 30, and 60 min. The reactions were stopped by the addition of 10 μl of ice-cold 2 M formic acid. A 10 μl aliquot of each reaction mixture was loaded on polyethyleneimine (PEI) cellulose TLC plates (GE Healthcare) and separated by chromatography in 1.5 M potassium phosphate at pH 3.4. The TLC plates were revealed by phosphorimaging (GE Healthcare) and analyzed using ImageQuant software (GE Healthcare). The increase in the level of (p)ppGpp was normalized to the basal level (time zero) for each strain.
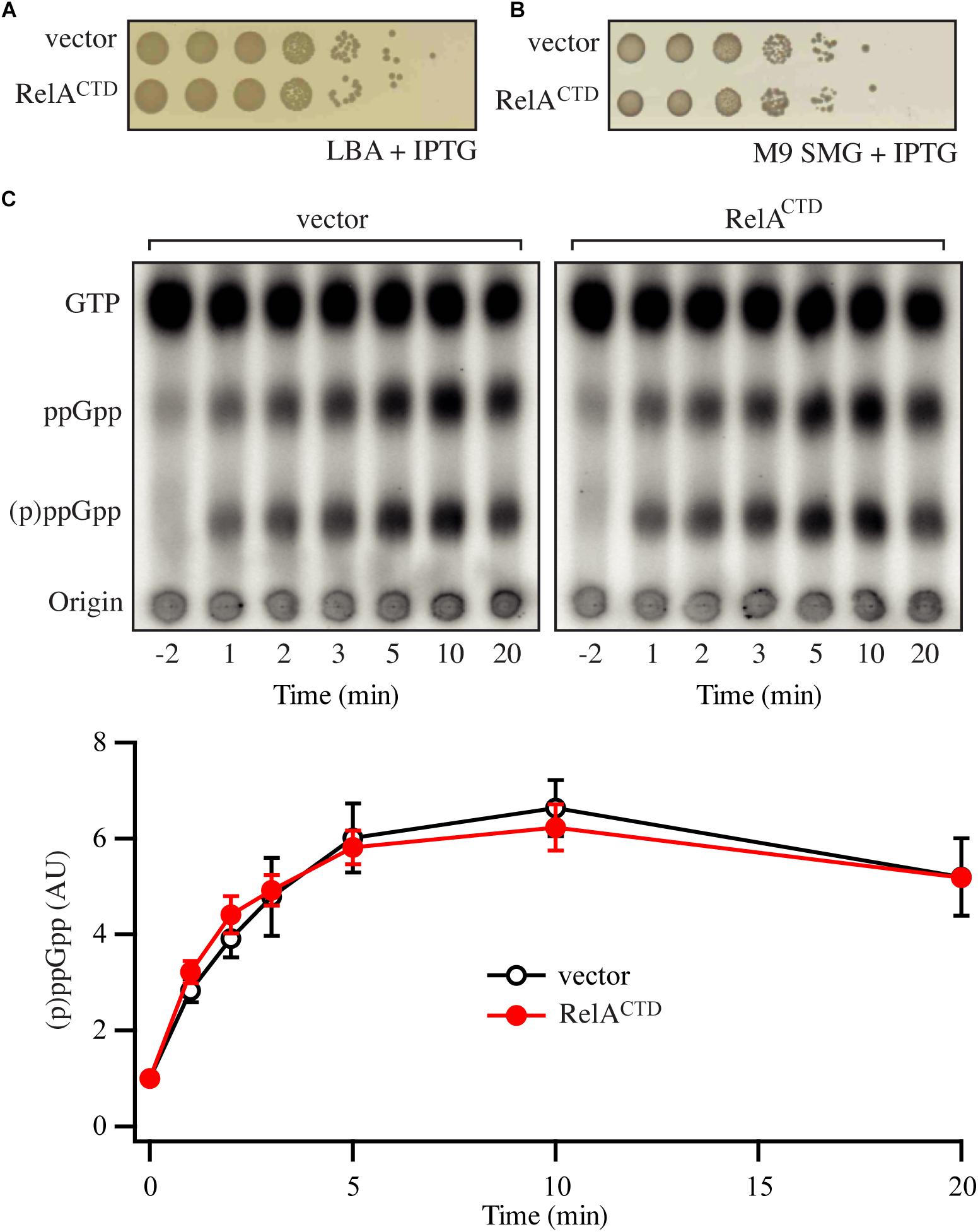
Figure 2. Low-level ectopic expression of RelACTD does not interfere with the functionality of endogenous RelA. (A,B) Overnight cultures of E. coli MG1655, transformed with low copy IPTG inducible vector pNDM220 (vector) or pNDM220:relACTD, were grown at 37°C overnight in M9 minimal medium with 30 μg/ml ampicillin. Ten-fold serial dilutions were made and spotted onto LB agar (LBA) and SMG plates, both supplemented with 30 μg/ml ampicillin and 1 mM IPTG. (C) Representative audioradiogram of a PEI Cellulose TLC plate showing (p)ppGpp accumulation of E. coli MG1655 carrying pNDM220 (vector) or pNDM220:relACTD upon valine-induced isoleucine starvation. The curves represent the average fold increase for three independent measurements, and the error bars represent standard errors. The levels of (p)ppGpp were normalized to the pre-starved level for each strain at –2 min. See experimental procedures for more details.
Monitoring Translation Efficiency
Overnight E. coli MG1655 cells harboring the CII-YFP plasmid pSEM3034UR2 kindly donated by Szabolcs Semsey and pMG25 (−ve), pMG25:relA, pMG25:relAG251E, pMG25:relACTD were grown into balanced growth in MOPS glucose minimal medium, before back-dilution to 0.05 OD600 into flat-bottom 96-well microtiter plate (Sterilin, Thermo Fisher Scientific) and placed in Synergy H1 (BioTek) microplate reader. Subsequently, the cultures were grown at 37°C with continuous orbital shaking. Every 15 min growth measured by taking the optical density (OD600), and the fluorescence was read from the bottom at 485 nm excitation and the 520 nm emission wavelengths (Figure 3).
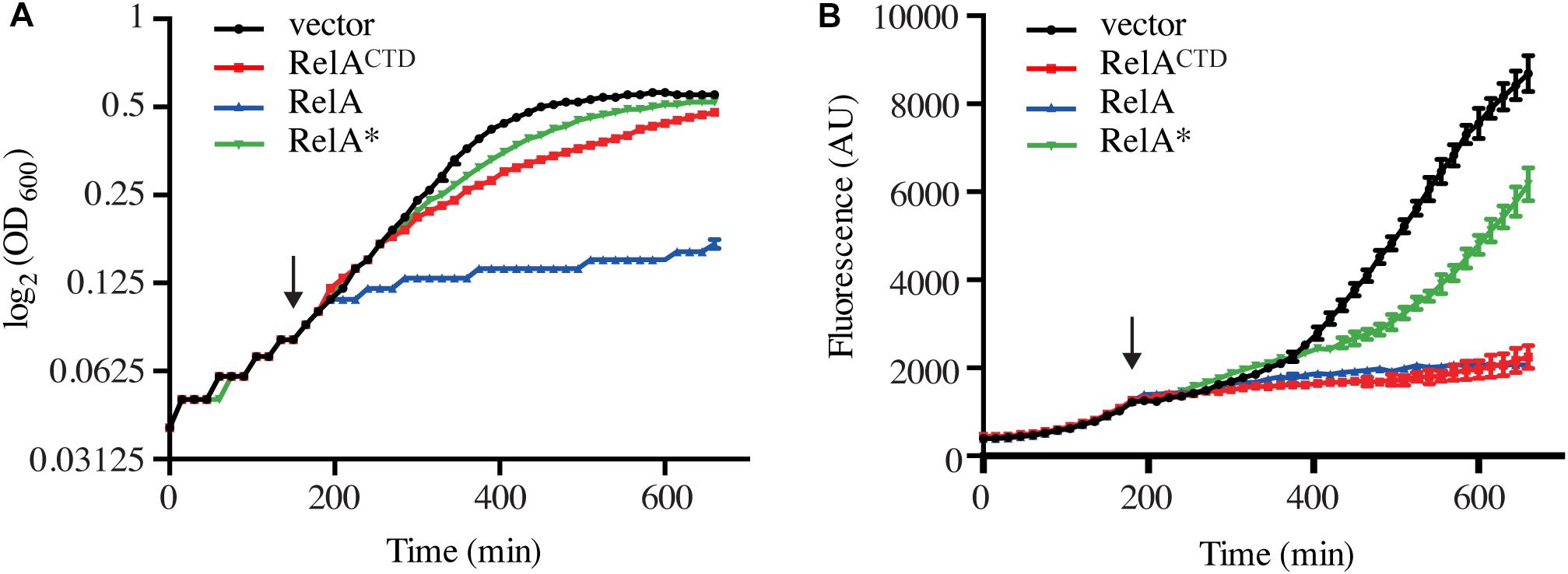
Figure 3. High-level ectopic expression of RelACTD impairs translation. Exponentially growing E. coli MG1655 cells harboring the CII-YFP plasmid were back-diluted to OD600 0.05, before the expression of relA, catalytically inactive G251E mutant relA∗, or relACTD from high copy vector pMG25 was induced by addition of 1 mM IPTG (indicated by the arrow). (A) Growth, measured at 600 nm (OD600) and (B) the fluorescence of YFP, measured at 520 nm was monitored every 15 min for 11 h. See section “Materials and Methods” for more details.
Western Blotting
For the detection of overproduced RelA and RelAΔRRM the appropriate plasmid was induced as described in the text and samples taken at the indicated time points (Figure 4). At each time point, 1 ml samples were taken and the cells were harvested by centrifugation at 4°C. Subsequently, the cell pellet was resuspended in an appropriate volume of 1× LDS loading buffer (Life Technologies) and boiled for 5 min. 20 μl of the samples was then resolved on a 4–12% NuPAGE gel (Life Technologies). Subsequently, the proteins were transferred using a semi-dry blotting apparatus (Hoefer Scientific Instruments) to a PVDF membrane (Amersham) and incubated with the primary antibody in PBS with 0.1% Tween-20 and 5% milk powder. The primary antibodies used in this study were raised in rabbit against a peptide sequence from the ZFD of RelA (Eurogentec). Next, membranes were incubated with HR conjugated anti-rabbit IgG antibody (Sigma) in PBS with 0.1% Tween-20 and 5% milk powder. Finally, the protein bands were visualized using Pierce ECL chemiluminescence substrate (Thermo Fisher Scientific) according to manufacturer’s instructions and the signal was detected in an Imagequant LAS4100.
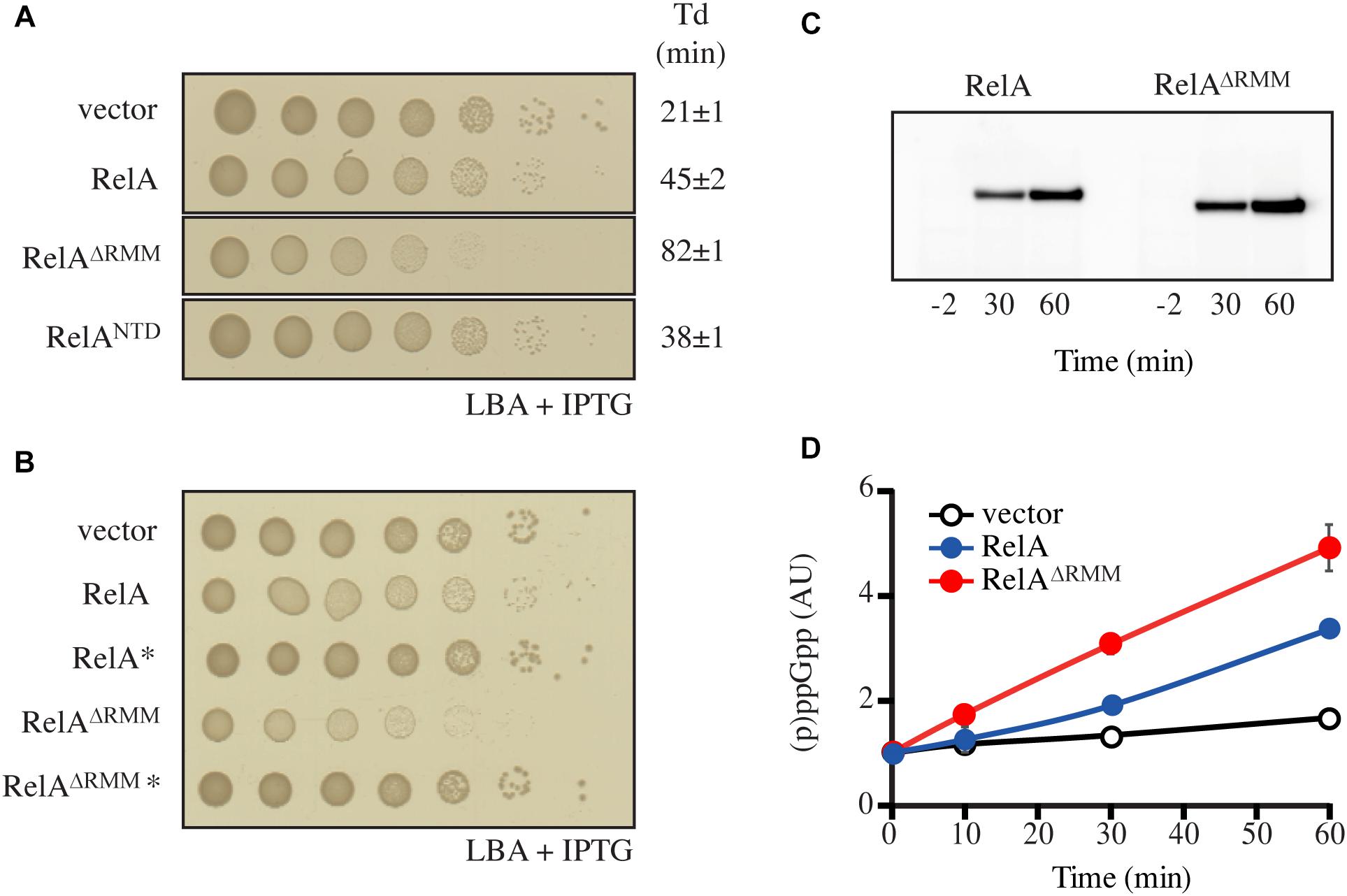
Figure 4. Truncation of RelA leads to accumulation of (p)ppGpp. (A,B) E. coli MG1655 cells were transformed with low copy IPTG inducible vector, pNDM220 (vector), pNDM220:relA, pNDM220:relA∗, pNDM220:relAΔRRM, pNDM220:relAΔRRM*, or pNDM220:relANTD. Ten-fold serial dilutions of overnight LB cultures were made and spotted onto LB agar (LBA) supplemented with 30 μg/ml ampicillin and 1 mM IPTG. Loading controls are presented in Supplementary Figure S6B. (C) E. coli MG1655ΔrelA transformed with pNDM220:relA or pNDM220:relAΔRRM were grown at 37°C to OD600 0.5, before 1 mM IPTG was added for plasmid induction. Samples were withdrawn at the mentioned times (minutes after induction) and western blotting was performed as described in the experimental procedures. (D) E. coli MG1655 carrying pNDM220 (vector), pNDM220:relA, or pNDM220:relAΔRRM were grown exponentially in MOPS minimal medium with 30 μg/ml ampicillin before 1 mM IPTG was added, for plasmid induction at time 0 min. The curves represent the average fold increase of (p)ppGpp for two independent measurements and the error bars represent standard errors. The levels of (p)ppGpp were normalized to the pre-starved level (–2 min) for each strain.
Purification of E. coli 70S Ribosomes and Untagged Native RelA
Escherichia coli 70S ribosomes were prepared from RNase I-deficient E. coli strain MRE600 (Kurylo et al., 2016). Bacteria were grown in 2YT medium (Sigma-Alderich) to OD600 ≈0.5, collected by centrifugation, and the ribosomes were purified by sucrose gradient centrifugation as described for preparation of Enterococcus faecalis 70S earlier (Murina et al., 2018).
For purification of RelA, E. coli BL21 DE3 harboring pET24d:his10-SUMO-relA expression construct were grown, induced, harvested and lysed as previously described (Kudrin et al., 2018). All liquid chromatography steps were performed using ÄKTA Avant 25 system and chromatographic columns from GE Healthcare were used. In order to exclude a possibility of substitution of Zn2+ ions in RelA’s ZFD for Ni2+ during purification on metal affinity chromatography column (Block et al., 2009), HisTrap 5 HP column was stripped from Ni2+ (according to manufacturer recommendations) and loaded with 10 ml of 100 mM Zn(OAc)2, pH 5.0. The column was then washed with four column volumes of deionized water and pre-equilibrated with four column volumes of binding buffer (25 mM Hepes pH 7.6, 320 mM NaCl, 10 mM imidazole, 5 mM MgCl2, 4 mM BME, 10% glycerol). Clarified cell lysate (≈50 ml) was applied on the column at the flow rate 5 ml/minute. Then the column was washed with binding buffer (2.5 column volumes) and the protein was eluted with six column volumes of 0–100% gradient of elution buffer (binding buffer with 500 mM imidazole) and 2 ml fractions were collected into 96 deep well plates (Omega Bio-tek). The collected fractions were run on SDS–PAGE gel and the fractions corresponding to His10-SUMO-RelA with the least nucleic acid contamination was collected (≈5 ml, highlighted in Figure 5A; corresponding SDS–PAGE analysis is shown on Figure 5D). These fractions were applied to Hiprep 10/26 desalting column pre-equilibrated with the storage buffer (25 mM Hepes pH 7.6, 720 mM KCl, 50 mM arginine, 50 mM glutamic acid, 10 mM imidazole, 5 mM MgCl2, 4 mM BME, 10% glycerol) (Figure 5B). The peak was collected and concentrated to the volume not bigger than 5 ml using Amicon centrifugal filters with 50 kDa cut-off. His6-UlpI SUMO protease [Protein Expertise Platform (PEP), Umeå University] was added to the protein solution to a final concentration of 1.3 μM and incubated for 15 min at room temperature in order to cleave off His10-SUMO tag and the protein solution was applied on HisTrap 1 HP (GE Healthcare) loaded with Zn2+ and pre-equilibrated with storage buffer. The flow-through was collected and concentrated using Amicon concentrators with 50 kDa cut-off (Figure 5C). The concentrated protein was aliquoted per 20–30 μl, flash frozen in liquid nitrogen and stored at −80°C. The purity of RelA preparations was assessed by SDS–PAGE (Figure 5D) and spectrophotometrically [OD260/OD280 ratio below 0.8 corresponding to less than 5% RNA contamination (Layne, 1957)].
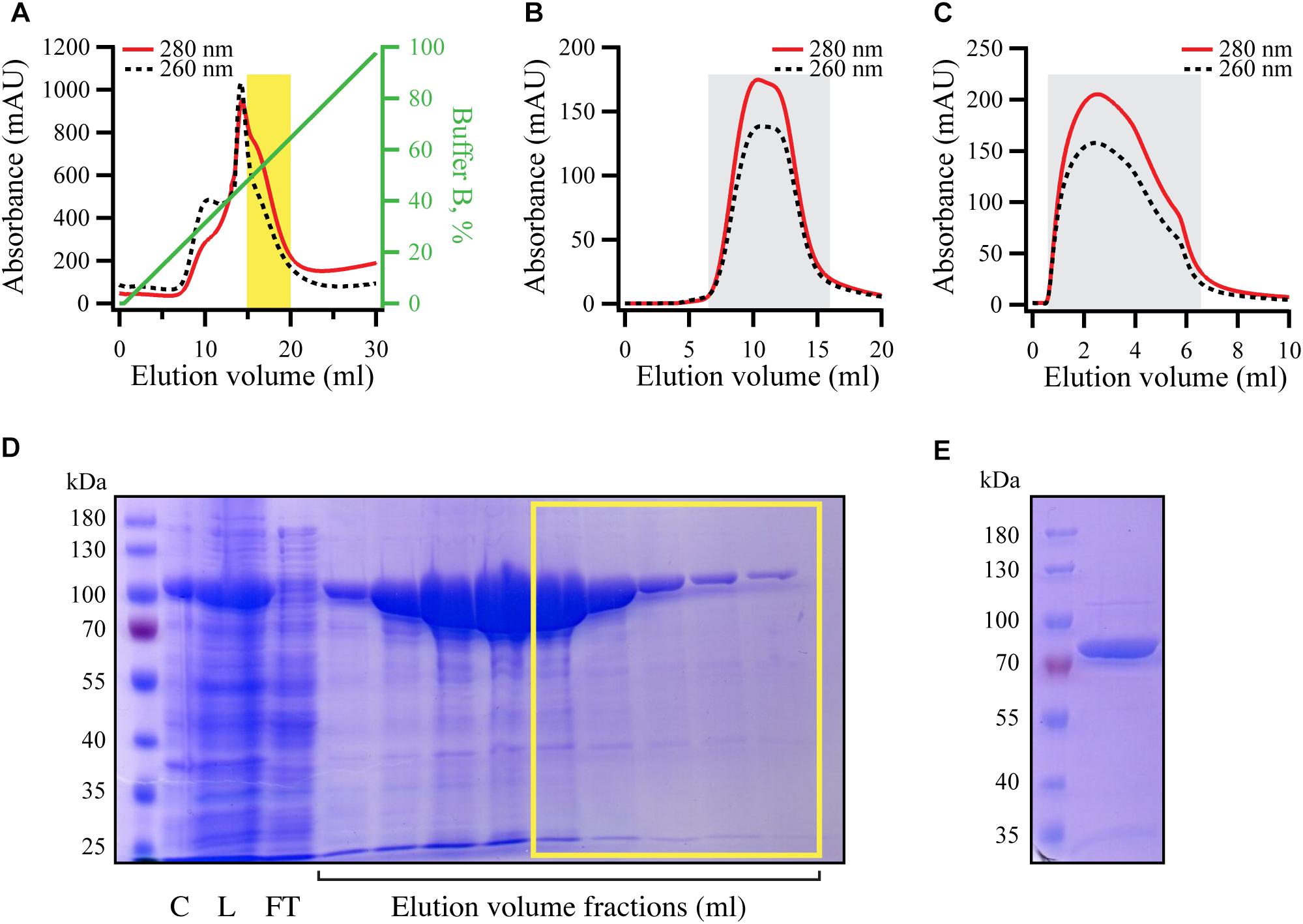
Figure 5. Purification of RNA-free untagged full-length E. coli RelA. N-terminally His10-SUMO tagged RelA was overexpressed and purified as described in detail in the section “Materials and Methods.” (A) Cells were lysed and subjected to immobilized metal affinity chromatography (IMAC). The fraction corresponding the RelA with the lowest contamination of nucleic acids (highlighted in yellow) was carried forward. (B) Following the buffer exchange, the grayed-out fractions were pooled and the His10-SUMO tag was cleaved off by the addition of His6-Ulp1. (C) His6-Ulp1 and the His10-SUMO tag was then purified from RelA by a second IMAC round. The grayed-out fractions were pulled, concentrated, aliquoted and stored –80°C. (D) SDS-PAGE showing the eluted fractions from the first IMAC round (A), fractions carried forward are highlighted with the yellow border. (E) SDS-PAGE analysis of purified native untagged RelA (≈84 kDa).
ppGpp Synthesis Assay
All experiments were performed at 37°C in HEPES:Polymix buffer, pH 7.6, with 5 mM Mg2+ (Antoun et al., 2004). Since increased concentration of KCl inhibits RelA activity, RelA stock solution containing 720 mM KCl was first diluted 7.2 times with HEPES:Polymix buffer without KCl to final KCl concentration of 100 mM and then added to the reaction mixture. The reaction mixture containing 1 mM [3H]GDP (PerkinElmer), 100 μM pppGpp as well as RelA at increasing concentrations (25, 50, 100, 200, and 400 nM), either supplemented or not by 0.6 μM 70S ribosomes, was preincubated for 3 min at 37°C and then the reaction was started by adding pre-warmed ATP to a final concentration of 1.5 mM. During the time course of the reaction 5 μl aliquots of the reaction mixture was quenched with 4 μl of 70% formic acid supplemented with 4 mM GTP and GDP (for UV shadowing). Time points were centrifuged for 5 min at 14,000 rpm, the supernatant was resolved on PEI-TLC (Macherey-Nagel) in 0.5 KH2PO4 buffer (pH 3.5) and the spots corresponding to [3H]GDP and [3H]ppGpp were cut out and quantified by scintillation in 5 ml of ScintiSafe scintillation cocktail (Thermo Fisher Scientific).
Results
High-Level Ectopic Expression of RelCTD Is Inhibitory to Growth Independently of Endogenous RelA
To test the functionality of RelA-mediated stringent response we utilized the so-called SMG test (minimal medium plates supplemented with serine, methionine, and glycine plates). Growth of E. coli on SMG is strictly dependent on the functionality of RelA (Uzan and Danchin, 1978). To investigate the effect of high-level ectopic expression of RelACTD (pUC derivative pMG25:RelACTD, PA1/O4/O3 promoter) on RelA activity, we assayed the growth of wild-type E. coli MG1655 (wt) cells growing on SMG. In agreement with the previous studies (Gropp et al., 2001; Yang and Ishiguro, 2001), high expression levels of RelACTD severely inhibit growth on SMG as compared to the empty vector control (Figure 1B). When E. coli is challenged with amino acid starvation, the levels of (p)ppGpp increase (Figure 1C, left side). At the same time, expression of RelACTD from the high-copy vector abrogates the response of native RelA to amino acid starvation and the levels of (p)ppGpp in the cells do not increase beyond the basal levels (Figure 1C, right side), i.e., native RelA fails to be activated in cells expressing high levels of RelACTD. The lack of activation of native RelA by amino acid starvation upon high-level expression of RelACTD could be explained either via direct inhibition of full-length RelA by RelACTD (autoregulation in trans) or interference with the activation of the full-length RelA by starved ribosomal complexes.
Next, we tested the ZFD domain mutations C612G, D637R, and C638F that were previously shown to render RelACTD unable to suppress the activity of native RelA (Gropp et al., 2001). We individually introduced these three mutations into the high copy plasmid encoding RelACTD to see whether the negative dominance of the CTD is abolished. The mentioned amino acid changes do not affect the expression or the stability of C-terminal fragments (Gropp et al., 2001). To our surprise, in our hands, overexpression of mutant variants of RelACTD still inhibits the growth of wt cells on SMG (Figure 1B). Therefore, high levels of RelACTD, with or without the mutated conserved residues, impair the functionality of RelA-mediated stringent response.
To test whether the observed effect of RelACTD overexpression on the stringent response is solely mediated by inactivation of endogenous RelA or via other mechanism(s) of growth inhibition, we investigated the effect of RelACTD expression in wt as well as ΔrelA backgrounds in both rich liquid and solid medium under unstarved conditions where functional RelA is not required. Surprisingly, upon induction of RelACTD, the growth rate is reduced identically in the wt and ΔrelA backgrounds, both in liquid and solid media indicative of the RelA-independent mechanism of growth inhibition (Figure 1D and Supplementary Figure S1A). Additionally, high expression levels of RelACTD bearing C612G, D637R, and C638F mutations are also inhibitory to growth in unstarved conditions (Supplementary Figure S2A), further reinforcing the idea growth inhibition being independent of the functionality of the endogenous RelA and the stringent response.
Low-Level Ectopic Expression of RelCTD Does Not Affect the Activation of Endogenous Full-Length RelA
Since high-level ectopic expression of RelACTD was inhibitory to growth independently of endogenous RelA (Figure 1D), no conclusions could be drawn with regard to its effect on the regulation of RelA. Ectopic expression of RelACTD from a high copy number plasmid results in accumulation of high levels of the recombinant protein in the cell (Supplementary Figure S1B). Given that there are only a few hundred molecules of RelA in the cell (Pedersen and Kjeldgaard, 1977; Justesen et al., 1986; Li et al., 2014; Schmidt et al., 2016), we switched to a very low copy number mini R1 plasmid (pNDM220, 1–2 copy per chromosome (Molin et al., 1979); PA1/O4/O3 promoter). We tested whether expression of the full-length RelA from this plasmid (pNDM220:relA) can restore the growth of a strain lacking relA (E. coli MG1655ΔrelA) on SMG. As seen from Supplementary Figure S3, expression of RelA in a ΔrelA background from this low copy number plasmid supports growth similar to wt cells. Notably, RelA bearing C612G, D637R, and C638F mutations in the ZFD domain also show full complementation (Supplementary Figure S3). As expected, the empty plasmid control and the catalytically inactive RelA mutant G251E [commonly designated as RelA∗ (Gropp et al., 2001)] fails to grow on SMG (Supplementary Figure S3). This result indicates that the expression from this plasmid resulted in more physiologically relevant levels of RelA.
Next, we repeated the experiments presented in the section “High-Level Ectopic Expression of RelCTD Is Inhibitory to Growth Independently of Endogenous RelA” using ectopic expression of RelACTD from the low copy number R1 plasmid (pNDM220:relACTD, PA1/O4/O3 promoter). As seen from Figure 2A, lower levels of RelACTD are not inhibitory to growth under unstarved conditions on LB agar. Likewise, at such expression levels, RelACTD does not affect the growth of starved wt cells on SMG plates (Figure 2B) or the out growth in SMG media (Supplementary Figure S5B), suggesting that native RelA in these conditions is still active. We then tested the profile of the of RelA mediated response upon amino acid starvation by measuring (p)ppGpp produced in the cell. As evident from the SMG plate assays (Figure 2B), low levels of RelACTD do not abrogate the functionality of endogenous RelA upon induction of the stringent response by addition of either valine that causes amino acid starvation in K12-based E. coli strains (Leavitt and Umbarger, 1962; Figure 2C) or SHX, the competitive inhibitor of seryl-tRNA synthetase (Tosa and Pizer, 1971; Supplementary Figure S4). Furthermore, in case additional domains other than the CTD are required, we wanted to explore the possibility of dimerization of full-length RelA enzyme. We hypothesized that if RelA is capable of dimerization, that endogenous expression of a catalytically inactive RelA mutant would bind and titrate out the native, catalytically active RelA, shifting the equilibrium away from free and active RelA in the cell, thus impairing the response to amino acid starvation. In order to test this theory, we repeated the above experiments with the inactive RelA G251E mutant, RelA∗ (pNDM220:relA∗). As seen from Supplementary Figure S5, low-level expression of RelA∗ does not affect growth on SMG plate or media, and does not affect the activation of endogenous RelA upon induction of amino acid starvation. Taken together, our data indicate that the activation of RelA is not predominantly regulated through intermolecular interactions.
High-Level Ectopic Expression of RelACTD Impairs Translation
The above data obtained using low expression vector suggests that RelACTD does not inhibit RelA activation by forming dimers. However, the question remained as to why high levels of RelACTD are inhibitory to growth (Figure 1D). All recent studies have unanimously showed that the CTD is critical for RelA binding to the ribosome (Agirrezabala et al., 2013; Arenz et al., 2016; Brown et al., 2016; Loveland et al., 2016; Winther et al., 2018). Furthermore, biochemical studies have shown that upon overexpression of RelACTD, the bulk of the protein is ribosome bound (Yang and Ishiguro, 2001). Therefore, we hypothesized that the reduced growth rate associated with high levels of RelACTD could be due to impairment of translation. To monitor the efficiency of translation upon expression of RelA variants (RelA, RelA∗ and RelACTD), we transformed wt cells with a low copy number plasmid encoding bacteriophage λ CII protein fused to Venus yellow fluorescence protein (CII-YFP) which allows the indirect monitoring of translation via measuring of fluorescence (Svenningsen et al., 2019).
As expected, wt cells with the empty vector exhibited increased levels of fluorescence correlated with growth, demonstrating active translation (Figures 3A,B). Wild-type cells harboring CII-YFP were grown exponentially before RelA and its variants were induced. High-level expression of full-length RelA results in accumulation of (p)ppGpp to levels at least as high as those seen during the stringent response (Schreiber et al., 1991; Svitil et al., 1993; Gropp et al., 2001). The alarmone is a potent inhibitor of bacterial growth targeting both transcription (Sorensen et al., 1994; Molodtsov et al., 2018), translation (Milon et al., 2006; Mitkevich et al., 2010) and ribosome assembly (Corrigan et al., 2016). Thus, as expected, upon over-expression of RelA, growth is immediately inhibited (Figure 3A). Consequently, the levels of fluorescence fail to increase, indicating arrest of translation (Figure 3B). Ectopic expression of RelACTD in mid-exponentially growing cells results in slower rather than total arrest of growth (Figure 3A), but the levels of fluorescence, as in the case of expression of the full-length RelA, do not increase suggesting translation is impaired (Figure 3B). Next, to test whether the presence of the NTD could regulate the inhibitory effect of the CTD, we checked the effect of expression of full-length catalytically inactive G251E mutant, RelA∗ (Supplementary Figure S3). As seen from Figure 3A, high-level expression of RelA∗ is inhibitory to growth, but to a lesser extent than RelACTD. Moreover, the levels of fluorescence gradually increased upon expression of RelA∗ indicating continuous decrease of translational efficiency in these cells. Notably, the inhibitory effect of RelACTD and RelA∗ in the cell is more pronounced when overnight cultures were back diluted to a lower optical density and allowed more time to accumulate the expressed product, suggesting that the inhibition of growth is dose-dependent (Supplementary Figure S2B). Taken together, these results suggest that RelACTD is inhibitory to growth as translation is impaired upon expression. RelA∗ is less toxic to growth and the presence of the NTD appears to moderate the toxic effect of the CTD.
The RRM Domain Is Crucial for Regulation of RelA’s Enzymatic Activity
The data presented above are suggestive of RelA being primely regulated, not via dimerization (in trans), but intramolecularly (in cis). To probe the autoregulatory roles of the individual domains, we systematically removed domains of the regulatory CTD region and tested the effects of their expression on growth (Figure 4A and Supplementary Figure S6A). Low-level expression of full-length RelA from pNDM220 in wt cells, was slightly inhibitory to growth (Figure 4A). Compared to the empty vector control, expression of RelA yielded smaller colonies on solid media and a reduced growth rate in LB liquid cultures of 21 to 45 min (Figure 4A and Supplementary Figure 6C), presumably due to elevated levels of (p)ppGpp. The expression of the constitutively active, but unstable, RelANTD (Schreiber et al., 1991; Svitil et al., 1993; Gropp et al., 2001) from the same plasmid resulted in similar growth inhibition (Figure 4A and Supplementary Figure S6C). Surprisingly, expression of RelA lacking the RRM domain (RelAΔRRM, Figure 1A), resulted in a more severe reduction of the growth rate than wild type RelA or the constitutively active RelANTD, as observed by a comparatively smaller colony size and an increased doubling time of 86 min (Figure 4A and Supplementary Figure S6C). Western blot analysis showed that upon expression, RelAΔRRM has similar abundance as RelA in the cells (Figure 4C). Thus, showing that the RRM domain is essential for the regulation of the (p)ppGpp synthetic activity of RelA.
To test if the growth inhibition by RelAΔRRM is mediated via over-production of (p)ppGpp, we repeated the plate assay described above using RelA and RelAΔRRM variants harboring the G251E mutation that abrogates RelA’s catalytic activity (Gropp et al., 2001). As seen in Figure 4B, the inhibition of growth is indeed dependent on the synthetic activity of RelA and RelAΔRRM. To further substantiate this hypothesis, we directly measured the levels of (p)ppGpp upon expression of RelA and RelAΔRRM in exponentially growing wt cells. Expression of RelA resulted in a gradual increase in the (p)ppGpp levels over time, and, consistent with a stronger growth inhibition (compare Figures 4A,B and Supplementary Figure S6C) the accumulation of the alarmone was more pronounced in the case of RelAΔRRM (Figure 4D). Taken together, our results collectively suggest that the RRM domain is crucial for preventing uncontrolled (p)ppGpp production by RelA.
Synthetic Activity of Native Untagged RelA Is Not Inhibited by Increasing Enzyme Concentrations
The concentration of RelA in E. coli is in the range of 50–100 nM (Pedersen and Kjeldgaard, 1977; Justesen et al., 1986; Li et al., 2014; Schmidt et al., 2016). We hypothesized that if there is regulation via dimerization is physiologically relevant, the dimerization affinity should be characterized by the equilibrium affinity constant (KD) in this concentration range, and, therefore, at the concentrations above the KD RelA would dimerize and become catalytically inhibited.
To test this hypothesis in the test tube, we purified native untagged E. coli RelA (Figure 5). Using untagged version of the protein for enzymatic studies is important since even addition of a C-terminal His6 tag can interfere with RelA’s regulation (Kudrin et al., 2018). To do so, we first subjected N-terminally His10-SUMO tagged RelA to immobilized metal affinity chromatography (IMAC). Since RelA makes extensive contacts with tRNA and ribosomal RNA which regulate its enzymatic activity (Arenz et al., 2016; Brown et al., 2016; Loveland et al., 2016; Kudrin et al., 2018; Winther et al., 2018), it was essential to purify the protein from nucleic acid contamination that is observed by monitoring both 260 nm and 280 nm UV traces during the IMAC purification step (Figure 5A). Therefore, we pooled only the fractions containing RelA with the lowest amounts of nucleic acid contamination, indicated by a yellow box in Figures 5A,D, rather than collecting the factions that contained the highest concentration of the protein as judged by absorbance at 280 nm and SDS PAGE analysis. Following a buffer exchange (Figure 5B), the His10-SUMO tag was cleaved by the addition of Ulp1, resulting in native untagged RelA (≈84 kDa), which was separated from the uncut version as well as the His10-SUMO tag by IMAC (Figure 5C). The purity of the protein preparation was checked by SDS–PAGE (Figure 5E), and importantly spectrophotometrically, giving the OD260/OD280 ratio of 0.8 corresponding to less than 5% RNA contamination (Layne, 1957).
We tested the synthetic activity of RelA in the range of concentrations from 25 to 400 nM, in the presence or absence of 0.6 μM 70S ribosomes (Figure 6). The enzymatic activity is largely insensitive to RelA concentration, suggesting that RelA is not regulated via through intermolecular interactions (dimerization).
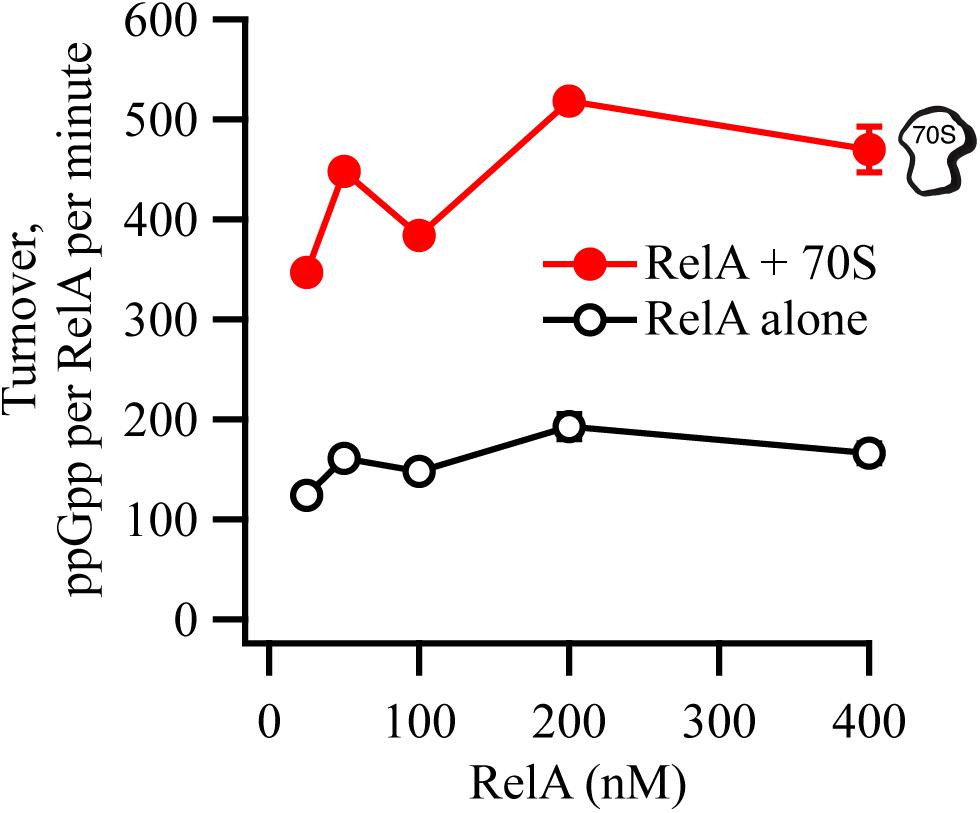
Figure 6. RelA’s ppGpp-synthetic activity is not inhibited by increasing concentrations of the enzyme. ppGpp synthetic activity of increasing concentrations of untagged native full-length RelA in the presence or absence of 0.6 μM E. coli 70S ribosomes, with 1 mM 3H-labeled GDP and 1.5 mM ATP. Enzymatic assays were performed at 37°C in HEPES:Polymix buffer, pH 7.5, in the presence of 5 mM Mg2+. Error bars represent SDs of the turnover estimates by linear regression.
Discussion
In this report, we provide evidence that the catalytic activity of the E. coli stringent factor RelA, is unlikely to be regulated through oligomerization in the cell, as it was previously suggested (Gropp et al., 2001; Yang and Ishiguro, 2001; Jain et al., 2006). However, our results are consistent with an alternative model for RelA regulation via intradomain regulation in cis, proposed earlier for “long” bifunctional RSH Rel from S. equisimilis (Mechold et al., 2002). This conclusion is substantiated by three lines of evidence.
The original “inhibition-via-dimerization” model of RelA regulation, was motivated by the observation of high-level expression of RelACTD leading to inhibition of both growth and (p)ppGpp accumulation upon amino acid starvation (Gropp et al., 2001; Yang and Ishiguro, 2001). By performing additional control experiments overlooked in the original reports, we demonstrate that high-level expression of RelACTD is inhibitory to growth in a RelA-independent manner, likely via direct inhibition of translation. Recent studies provide a possible structural explanation of the microbiological results. The CTD region is responsible for anchoring RelA to the ribosome by forming multiple contacts with rRNA and A-site tRNA (Agirrezabala et al., 2013; Arenz et al., 2016; Brown et al., 2016; Loveland et al., 2016; Winther et al., 2018). Once bound to the ribosome, RelACTD would sterically clash with other ribosomal ligands, such as elongation factors EF-Tu and ET-G (thus inhibiting translation), and endogenous full-length RelA [thus inhibiting (p)ppGpp accumulation upon amino acid starvation, resulting in a relaxed phenotype (Figures 1B,C)]. This interpretation is further substantiated by an earlier report demonstrating that RelACTD is ribosome-bound (Yang and Ishiguro, 2001). Conversely, at low expression levels, that are more similar to that of the endogenous full-length RelA, expression of neither RelACTD nor catalytically inactive G251E mutant RelA∗ compromises the response of wt MG1655 E. coli cells to acute amino acid starvation (Figure 2C and Supplementary Figure S4). We interpret this more physiologically relevant experiment to mean that neither of the constructs can drive the formation of catalytically inactive hetero-dimers with native endogenous full-length RelA. Finally, our biochemical experiments using untagged full-length RelA fail to detect reduction in the ppGpp synthetic activity upon increase in enzyme’s concentration that is expected to drive the formation of inactive dimers (Figure 6). Our biochemical results are in good agreement with gel-filtration analysis of full-length Thermus thermophilus Rel that failed to detect protein dimerization (Van Nerom et al., 2019). Importantly, the said gel-filtration experiments was performed at unphysiologically high ionic strength that is necessary to keep relatively (in comparison to E. coli RelA) soluble T. thermophilus Rel in solution, and, therefore, should not be over-interpreted. Finally, Mycobacterium tuberculosis Rel ectopically expressed in E. coli does not form oligomers, further supporting the monomeric nature of RelA/Rel enzymes (Jain et al., 2006).
In addition to characterizing the intact RelA regulatory CTD region, we have examined a C-terminally truncated RelA variant lacking the RRM domain (RelAΔRRM). Surprisingly, in the cell, the (p)ppGpp synthetic activity of the enzyme is unregulated in the absence of the RRM domain (Figure 4). Importantly, ectopic expression of RelAΔRRM, is more inhibitory to growth than that of well-characterized constitutively active RelANTD (Schreiber et al., 1991; Svitil et al., 1993; Gropp et al., 2001). A possible explanation is that while RelANTD is highly unstable and decays with a half-life of only a few minutes (Schreiber et al., 1991), the stability of RelAΔRRM is similar to full-length RelA, i.e., half-life of more than 2 h (Schreiber et al., 1991). Therefore, more efficient inhibition of growth by RelAΔRRM is likely a compound effect of, first, de-regulation of the enzyme leading to uncontrolled (p)ppGpp production similarly to RelANTD, and second, relatively high stability of the protein product. In agreement with our findings, deletion of the RRM domain of Caulobacter crescentus bifunctional ribosome-associated RSH, Rel, also leads to de-regulation of the protein and (p)ppGpp-mediated growth inhibition (Ronneau et al., 2019). Curiously, while Francisella tularensis RelA naturally lacks the RRM domain (Atkinson et al., 2011), the enzyme displays normal, wt-like enzymatic functionality in biochemical assays (Wilkinson et al., 2015). Further work is required to determine the exact regulatory role of the RRM domain that is near-universally conserved in “long” RSHs Rel, RelA and SpoT (Atkinson et al., 2011).
Data Availability
All datasets generated for this study are included in the manuscript and/or the Supplementary Files.
Author Contributions
MR conceived the study. MR and VH designed the experiments. KT, ID, SL, and MR performed the experiments, contributed to the data analysis, and drafted and revised the manuscript with contributions from VH. All authors read and approved the final version of the manuscript.
Funding
This work was carried out at both the Centre for Bacterial Stress Response and Persistence (BASP) at the University of Copenhagen, and the Department of Molecular Biology at Umeå University. This work was supported by a grant from the Danish National Research Foundation (DNFR120), the MIMS Excellence by Choice Postdoctoral Fellowship Programme to MR and the Swedish Research Council (grant 2017-03783 to VH).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Professor Kenn Gerdes, Dr. Anurag Sinha, and Dr. Farshid Jalalvand for helpful discussions and comments on the manuscript, Dr. Szabolcs Semsey for kindly providing plasmid pSEM3034UR2, and the Protein Expertise Platform (PEP) at Umeå University and Mikael Lindberg for constructing pET24d:His10-SUMO and purifying His6-Ulp1. This manuscript was deposited as a preprint to bioRxiv (Turnbull et al., 2019).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.01966/full#supplementary-material
References
Agirrezabala, X., Fernandez, I. S., Kelley, A. C., Carton, D. G., Ramakrishnan, V., and Valle, M. (2013). The ribosome triggers the stringent response by RelA via a highly distorted tRNA. EMBO Rep. 14, 811–816. doi: 10.1038/embor.2013.106
Antoun, A., Pavlov, M. Y., Tenson, T., and Ehrenberg, M. M. (2004). Ribosome formation from subunits studied by stopped-flow and Rayleigh light scattering. Biol. Proced. Online 6, 35–54. doi: 10.1251/bpo71
Arenz, S., Abdelshahid, M., Sohmen, D., Payoe, R., Starosta, A. L., Berninghausen, O., et al. (2016). The stringent factor RelA adopts an open conformation on the ribosome to stimulate ppGpp synthesis. Nucleic Acids Res. 44, 6471–6481. doi: 10.1093/nar/gkw470
Atkinson, G. C., Tenson, T., and Hauryliuk, V. (2011). The RelA/SpoT homolog (RSH) superfamily: distribution and functional evolution of ppGpp synthetases and hydrolases across the tree of life. PLoS One 6:e23479. doi: 10.1371/journal.pone.0023479
Block, H., Maertens, B., Spriestersbach, A., Brinker, N., Kubicek, J., Fabis, R., et al. (2009). Immobilized-metal affinity chromatography (IMAC): a review. Methods Enzymol. 463, 439–473. doi: 10.1016/S0076-6879(09)63027-5
Brown, A., Fernandez, I. S., Gordiyenko, Y., and Ramakrishnan, V. (2016). Ribosome-dependent activation of stringent control. Nature 534, 277–280. doi: 10.1038/nature17675
Cashel, M. (1994). “Detection of (p)ppGpp accumulation patterns in Escherichia coli mutants,” in Methods in Molecular Genetics, ed. K. W. Adolph (New York, N.Y: Academic Press), 341–356.
Cherepanov, P. P., and Wackernagel, W. (1995). Gene disruption in Escherichia coli: TcR and KmR cassettes with the option of flp-catalyzed excision of the antibiotic-resistance determinant. Gene 158, 9–14. doi: 10.1016/0378-1119(95)00193-a
Corrigan, R. M., Bellows, L. E., Wood, A., and Grundling, A. (2016). ppGpp negatively impacts ribosome assembly affecting growth and antimicrobial tolerance in Gram-positive bacteria. Proc. Natl. Acad. Sci. U.S.A. 113, E1710–E1719. doi: 10.1073/pnas.1522179113
Dalebroux, Z. D., Svensson, S. L., Gaynor, E. C., and Swanson, M. S. (2010). ppGpp conjures bacterial virulence. Microbiol. Mol. Biol. Rev. 74, 171–199. doi: 10.1128/MMBR.00046-09
Dalebroux, Z. D., and Swanson, M. S. (2012). ppGpp: magic beyond RNA polymerase. Nat. Rev. Microbiol. 10, 203–212. doi: 10.1038/nrmicro2720
Gotfredsen, M., and Gerdes, K. (1998). The Escherichia coli relBE genes belong to a new toxin-antitoxin gene family. Mol. Microbiol. 29, 1065–1076. doi: 10.1046/j.1365-2958.1998.00993.x
Gropp, M., Strausz, Y., Gross, M., and Glaser, G. (2001). Regulation of Escherichia coli RelA requires oligomerization of the C-terminal domain. J. Bacteriol. 183, 570–579. doi: 10.1128/jb.183.2.570-579.2001
Haseltine, W. A., and Block, R. (1973). Synthesis of guanosine tetra- and pentaphosphate requires the presence of a codon-specific, uncharged transfer ribonucleic acid in the acceptor site of ribosomes. Proc. Natl. Acad. Sci. U.S.A. 70, 1564–1568. doi: 10.1073/pnas.70.5.1564
Hauryliuk, V., Atkinson, G. C., Murakami, K. S., Tenson, T., and Gerdes, K. (2015). Recent functional insights into the role of (p)ppGpp in bacterial physiology. Nat. Rev. Microbiol. 13, 298–309. doi: 10.1038/nrmicro3448
Jain, V., Saleem-Batcha, R., China, A., and Chatterji, D. (2006). Molecular dissection of the mycobacterial stringent response protein Rel. Protein Sci. 15, 1449–1464. doi: 10.1110/ps.062117006
Jaskolska, M., and Gerdes, K. (2015). CRP-dependent positive autoregulation and proteolytic degradation regulate competence activator Sxy of Escherichia coli. Mol. Microbiol. 95, 833–845. doi: 10.1111/mmi.12901
Jimmy, S., Saha, C. K., Stavropoulos, C., Garcia-Pino, A., Hauryliuk, V., and Atkinson, G. C. (2019). Discovery of small alarmone synthetases and their inhibitors as toxin-antitoxin loci. bioRxiv [Preprint]. doi: 10.1101/575399
Justesen, J., Lund, T., Skou Pedersen, F., and Kjeldgaard, N. O. (1986). The physiology of stringent factor (ATP:GTP 3′-diphosphotransferase) in Escherichia coli. Biochimie 68, 715–722. doi: 10.1016/s0300-9084(86)80165-1
Knutsson Jenvert, R. M., and Holmberg Schiavone, L. (2005). Characterization of the tRNA and ribosome-dependent pppGpp-synthesis by recombinant stringent factor from Escherichia coli. FEBS J. 272, 685–695. doi: 10.1111/j.1742-4658.2004.04502.x
Kudrin, P., Dzhygyr, I., Ishiguro, K., Beljantseva, J., Maksimova, E., Oliveira, S. R. A., et al. (2018). The ribosomal A-site finger is crucial for binding and activation of the stringent factor RelA. Nucleic Acids Res. 46, 1973–1983. doi: 10.1093/nar/gky023
Kurylo, C. M., Alexander, N., Dass, R. A., Parks, M. M., Altman, R. A., Vincent, C. T., et al. (2016). Genome sequence and analysis of Escherichia coli MRE600, a colicinogenic, nonmotile strain that lacks RNase I and the type I methyltransferase, EcoKI. Genome Biol. Evol. 8, 742–752. doi: 10.1093/gbe/evw008
Layne, E. (1957). Spectrophotometric and turbidimetric methods for measuring proteins. Methods Enzymol. 3, 447–454. doi: 10.1016/s0076-6879(57)03413-8
Leavitt, R. I., and Umbarger, H. E. (1962). Isoleucine and valine metabolism in Escherichia coli. XI. valine inhibition of the growth of Escherichia coli strain K-12. J. Bacteriol. 83, 624–630.
Li, G. W., Burkhardt, D., Gross, C., and Weissman, J. S. (2014). Quantifying absolute protein synthesis rates reveals principles underlying allocation of cellular resources. Cell 157, 624–635. doi: 10.1016/j.cell.2014.02.033
Loveland, A. B., Bah, E., Madireddy, R., Zhang, Y., Brilot, A. F., Grigorieff, N., et al. (2016). Ribosome∗RelA structures reveal the mechanism of stringent response activation. eLife 5:e17029. doi: 10.7554/eLife.17029
Mechold, U., Murphy, H., Brown, L., and Cashel, M. (2002). Intramolecular regulation of the opposing (p)ppGpp catalytic activities of Rel(Seq), the Rel/Spo enzyme from Streptococcus equisimilis. J. Bacteriol. 184, 2878–2888. doi: 10.1128/jb.184.11.2878-2888.2002
Milon, P., Tischenko, E., Tomsic, J., Caserta, E., Folkers, G., La Teana, A., et al. (2006). The nucleotide-binding site of bacterial translation initiation factor 2 (IF2) as a metabolic sensor. Proc. Natl. Acad. Sci. U.S.A. 103, 13962–13967. doi: 10.1073/pnas.0606384103
Mitkevich, V. A., Ermakov, A., Kulikova, A. A., Tankov, S., Shyp, V., Soosaar, A., et al. (2010). Thermodynamic characterization of ppGpp binding to EF-G or IF2 and of initiator tRNA binding to free IF2 in the presence of GDP, GTP, or ppGpp. J. Mol. Biol. 402, 838–846. doi: 10.1016/j.jmb.2010.08.016
Molin, S., Stougaard, P., Uhlin, B. E., Gustafsson, P., and Nordstrom, K. (1979). Clustering of genes involved in replication, copy number control, incompatibility, and stable maintenance of the resistance plasmid R1drd-19. J. Bacteriol. 138, 70–79.
Molodtsov, V., Sineva, E., Zhang, L., Huang, X., Cashel, M., Ades, S. E., et al. (2018). Allosteric effector ppGpp potentiates the inhibition of transcript initiation by DksA. Mol. Cell 69, 828–839.e5. doi: 10.1016/j.molcel.2018.01.035
Murina, V., Kasari, M., Hauryliuk, V., and Atkinson, G. C. (2018). Antibiotic resistance ABCF proteins reset the peptidyl transferase centre of the ribosome to counter translational arrest. Nucleic Acids Res. 46, 3753–3763. doi: 10.1093/nar/gky050
Pedersen, F. S., and Kjeldgaard, N. O. (1977). Analysis of the relA gene product of Escherichia coli. Eur. J. Biochem. 76, 91–97. doi: 10.1111/j.1432-1033.1977.tb11573.x
Ronneau, S., Caballero-Montes, J., Coppine, J., Mayard, A., Garcia-Pino, A., and Hallez, R. (2019). Regulation of (p)ppGpp hydrolysis by a conserved archetypal regulatory domain. Nucleic Acids Res. 47, 843–854. doi: 10.1093/nar/gky1201
Schmidt, A., Kochanowski, K., Vedelaar, S., Ahrne, E., Volkmer, B., Callipo, L., et al. (2016). The quantitative and condition-dependent Escherichia coli proteome. Nat. Biotechnol. 34, 104–110. doi: 10.1038/nbt.3418
Schreiber, G., Metzger, S., Aizenman, E., Roza, S., Cashel, M., and Glaser, G. (1991). Overexpression of the relA gene in Escherichia coli. J. Biol. Chem. 266, 3760–3767.
Shyp, V., Tankov, S., Ermakov, A., Kudrin, P., English, B. P., Ehrenberg, M., et al. (2012). Positive allosteric feedback regulation of the stringent response enzyme RelA by its product. EMBO Rep. 13, 835–839. doi: 10.1038/embor.2012.106
Sorensen, M. A., Jensen, K. F., and Pedersen, S. (1994). High concentrations of ppGpp decrease the RNA chain growth rate. Implications for protein synthesis and translational fidelity during amino acid starvation in Escherichia coli. J. Mol. Biol. 236, 441–454. doi: 10.1006/jmbi.1994.1156
Svenningsen, M. S., Veress, A., Harms, A., Mitarai, N., and Semsey, S. (2019). Birth and resuscitation of (p)ppGpp induced antibiotic tolerant persister cells. Sci. Rep. 9:6056. doi: 10.1038/s41598-019-42403-7
Svitil, A. L., Cashel, M., and Zyskind, J. W. (1993). Guanosine tetraphosphate inhibits protein synthesis in vivo. A possible protective mechanism for starvation stress in Escherichia coli. J. Biol. Chem. 268, 2307–2311.
Tian, C., Roghanian, M., Jorgensen, M. G., Sneppen, K., Sorensen, M. A., Gerdes, K., et al. (2016). Rapid curtailing of the stringent response by toxin-antitoxin module-encoded mRNases. J. Bacteriol. 198, 1918–1926. doi: 10.1128/JB.00062-16
Tosa, T., and Pizer, L. I. (1971). Biochemical bases for the antimetabolite action of L-serine hydroxamate. J. Bacteriol. 106, 972–982.
Turnbull, K. J., Dzhygyr, I., Lindemose, S., Hauryliuk, V., and Roghanian, M. (2019). Intramolecular interactions dominate the autoregulation of Escherichia coli stringent factor RelA. bioRxiv [Preprint]. doi: 10.1101/680231
Uzan, M., and Danchin, A. (1978). Correlation between the serine sensitivity and the derepressibility of the ilv genes in Escherichia coli relA- mutants. Mol. Gen. Genet. 165, 21–30. doi: 10.1007/bf00270372
Van Nerom, K., Tamman, H., Takada, H., Hauryliuk, V., and Garcia-Pino, A. (2019). The Rel stringent factor from Thermus thermophilus: crystallization and X-ray analysis. Acta Cryst. F 75, 561–569. doi: 10.1107/S2053230X19010628
Wendrich, T. M., Blaha, G., Wilson, D. N., Marahiel, M. A., and Nierhaus, K. H. (2002). Dissection of the mechanism for the stringent factor RelA. Mol. Cell 10, 779–788. doi: 10.1016/s1097-2765(02)00656-1
Wilkinson, R. C., Batten, L. E., Wells, N. J., Oyston, P. C., and Roach, P. L. (2015). Biochemical studies on Francisella tularensis RelA in (p)ppGpp biosynthesis. Biosci. Rep. 35:e00268. doi: 10.1042/BSR20150229
Winther, K. S., Roghanian, M., and Gerdes, K. (2018). Activation of the stringent response by loading of RelA-tRNA complexes at the ribosomal A-Site. Mol. Cell 70, 95–105.e4. doi: 10.1016/j.molcel.2018.02.033
Keywords: (p)ppGpp, RelA, RSH, stringent response, ribosome, translation
Citation: Turnbull KJ, Dzhygyr I, Lindemose S, Hauryliuk V and Roghanian M (2019) Intramolecular Interactions Dominate the Autoregulation of Escherichia coli Stringent Factor RelA. Front. Microbiol. 10:1966. doi: 10.3389/fmicb.2019.01966
Received: 22 June 2019; Accepted: 09 August 2019;
Published: 27 August 2019.
Edited by:
Haike Antelmann, Freie Universität Berlin, GermanyReviewed by:
Gert Bange, University of Marburg, GermanyMichael Cashel, Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD), United States
Copyright © 2019 Turnbull, Dzhygyr, Lindemose, Hauryliuk and Roghanian. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mohammad Roghanian, bW9oYW1tYWQucm9naGFuaWFuQHVtdS5zZQ==