 Floriane Gaucher1,2
Floriane Gaucher1,2 Koffigan Kponouglo1
Koffigan Kponouglo1 Houem Rabah1,3
Houem Rabah1,3 Sylvie Bonnassie1,4
Sylvie Bonnassie1,4 Jordane Ossemond1
Jordane Ossemond1 Sandrine Pottier5
Sandrine Pottier5 Julien Jardin1Valérie Briard-Bion1Pierre Marchand2
Julien Jardin1Valérie Briard-Bion1Pierre Marchand2 Philippe Blanc2
Philippe Blanc2 Romain Jeantet1†
Romain Jeantet1† Gwénaël Jan1*†
Gwénaël Jan1*†- 1UMR STLO, Agrocampus Ouest, INRA, Rennes, France
- 2Bioprox, Levallois-Perret, France
- 3Bba, Pôle Agronomique Ouest, Régions Bretagne et Pays de la Loire, Rennes, France
- 4Université de Rennes I, Rennes, France
- 5CNRS, ISCR – UMR 6226, PRISM, BIOSIT – UMS 3480 Université de Rennes I, Rennes, France
Propionibacterium freudenreichii is a beneficial bacterium with documented effects on the gut microbiota and on inflammation. Its presence within the animal and human intestinal microbiota was correlated with immunomodulatory effects, mediated by both propionibacterial surface components and by secreted metabolites. It is widely implemented, both in the manufacture of fermented dairy products such as Swiss-type cheeses, and in the production of probiotic food complements, under the form of freeze-dried powders. The bottleneck of this drying process consists in the limited survival of bacteria during drying and storage. Protective pre-treatments have been applied to other bacteria and may, in a strain-dependent manner, confer enhanced resistance. However, very little information was yet published on P. freudenreichii adaptation to freeze-drying. In this report, an immunomodulatory strain of this probiotic bacterium was cultured under hyperosmotic constraint in order to trigger osmoadaptation. This adaptation was then combined with acid or thermal pre-treatment. Such combination led to accumulation of key stress proteins, of intracellular compatible solute glycine betaine, to modulation of the propionibacterial membrane composition, and to enhanced survival upon freeze-drying. This work opens new perspectives for efficient production of live and active probiotic propionibacteria.
Introduction
Incidence of inflammatory bowel disease (IBD), of irritable bowel syndrome (IBS) and of antibiotic-associated diarrhea (ADD) is increasing in developed countries in relation with modifications of the life style. Indeed, the Western diet contains high amount of fat and proteins. The diet has an important impact on the modulation of the microbiota. A dysbiosis of the microbiota is involved in many disorders like inflammation, allergy and atopy (Moco et al., 2014; Skonieczna-Żydecka et al., 2018; Soularue et al., 2018). Ingestion of a mixture of probiotic bacteria induced or prolonged a remission in patients suffering from ulcerative colitis (Ganji-Arjenaki and Rafieian-Kopaei, 2018). In addition, the strain Lactobacillus plantarum 299V and Bifidobacterium infantis 35624 exerted healing effect on IBS (Ducrotté, 2012; Yuan et al., 2017; Sniffen et al., 2018). Probiotic are defined as “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” (FAO/WHO, 2001). Early exposition to key immunomodulatory symbiotic bacteria and or/probiotic prevent allergy and atopy (Fujimura et al., 2016; Butel et al., 2018). Bifidobacteria and propionibacteria constitute protective and early colonizing probiotic symbionts (Chang et al., 2017; Colliou et al., 2017). Furthermore, allergic diseases in Caesarean-delivered children, at 13-year follow-up, was prevented by an early exposition to a mixture of lactic acid bacteria an propionibacteria (Kallio et al., 2018).
Propionibacterium freudenreichii possesses the QPS (qualified Presumption of Safety EFS, European Union) and the GRAS (Generally Recognized As Safe, United States, FDA) status (Cousin et al., 2010; Rabah et al., 2018). It is consumed in high amounts due to its presence in Swiss-type cheeses, and also in probiotic food supplements. During the manufacturing, P. freudenreichii is responsible of the production of the beneficial short chain fatty acids acetate and propionate, the B9 (folate) and B12 (cobalamin) vitamins, and the bifidogenic compounds DHNA (1,4-dihydroxy-2-naphtoic acid) and ACNQ (2-amino-3-carboxy-1,4-naphthoquinone) (Rabah et al., 2017). Select strains of P. freudenreichii can modulate the gut microbiota in the colitis context and in healthy conditions (Bouglé et al., 1999; Hojo et al., 2002; Seki et al., 2004; Mitsuyama et al., 2007). This modulation depends of the production of the DHNA and ACNQ molecules which favor bifidobacteria at the expense of pathobiont Bacteroides (Isawa et al., 2002). Consumption of P. freudenreichii protected animals from induced intestinal disease (Foligne et al., 2010; Plé et al., 2015, 2016) and revealed healing effect on ulcerative colitis in humans (Okada, 2006; Mitsuyama et al., 2007). To optimize its effects mediated by beneficial metabolites, P. freudenreichii should be consumed alive. It can be consumed within Swiss-type cheese or under a powder form in tablets or capsules. The International Dairy Federation (IDF) recommends a minimum of 107 live probiotic bacterial cells per gram or milliliter of production at the time of consumption (Corona-Hernandez et al., 2013).
Most of the time, probiotic bacteria are stored, transported and consumed under a powder form. Probiotics microorganisms like bacteria and yeasts are dried to produce easy-to-use preparations, which can be implemented in food, feed or pharmaceutical industry (Huang et al., 2017). Two drying processes can be used to dry bacteria: freeze-drying and spray drying. Freeze-drying is the most used process, keeps alive the largest percentage of bacteria, but it has the disadvantage of being an expensive discontinuous process (Santivarangkna et al., 2007). However, freeze-drying imposes cold and osmotic stresses (Gaucher et al., 2019b), and causes the appearance of holes in the membrane, which can lead to cell death (Carvalho et al., 2004; Giulio et al., 2005). After drying, bacteria used as probiotic will further be exposed to harsh conditions during the digestion process. Some of the probiotic actions, however, need live bacteria in the colon. Tolerance toward stomach acidity and intestinal bile salts can constitute a limit of the probiotic potential.
Bacteria adaptation increase bacteria resistance to different stresses like acid stress (Jan et al., 2002; Broadbent et al., 2010), bile salt stress (Leverrier et al., 2003, 2004), and drying (Desmond et al., 2001; Carvalho et al., 2004; Silva et al., 2005; Li et al., 2009; Paéz et al., 2012). Cross-protections provided by adaptation are generally strain-dependent (Gaucher et al., 2019b). In a previous study, osmoadaptation exerted a positive impact on P. freudenreichii CIRM-BIA 129 viability during freeze-drying, but not during different challenges such as heat, acid or bile salts stresses (Gaucher et al., 2019a). Concerning P. freudenreichii freeze-drying, almost nothing has been published. Acid adaptation and/or heat adaptation have, however, been reported to increase Lactobacillus casei (Paéz et al., 2012), L. plantarum (Paéz et al., 2012), Lactobacillus bulgaricus (Li et al., 2009), and Lactobacillus reuteri (Palmfeldt and Hahn-Hägerdal, 2000; Koch et al., 2008) viability during freeze-drying as well as Lactobacillus paracasei (Desmond et al., 2001), and Lactobacillus delbrueckii (Silva et al., 2005) viability during spray drying. During these adaptations, the membrane fluidity of the bacteria will be adjusted by the modulation of the composition of fatty acids, in particularly the amount of saturated (SFA) and unsaturated (UFA) fatty acids (Lanciotti et al., 2001; Machado et al., 2004; Streit et al., 2008; Broadbent et al., 2010). General stress protein like Clps, DnaK, GroES or GroEL are overproduced during these three adaptations (Jan et al., 2001; Leverrier et al., 2004; Savijoki et al., 2005; Dalmasso et al., 2012; Huang et al., 2016a). During osmoadaptation, P. freudenreichii is able to accumulate glutamate, trehalose and glycine betaine (Gaucher et al., 2019a). In hyperosmotic media, bacteria need to accumulate compatible solutes in order to preserve their turgescent pressure and to enable cell growth and division. Compatible solutes can be imported into the cell from the growth medium, or synthesized de novo (Csonka and Hanson, 1991). Glycine betaine is a compatible solute accumulated by different bacteria, including Staphylococcus aureus, Escherichia coli, Enterococcus faecalis, and P. freudenreichii, during osmoadaptation (Kunin and Rudy, 1991; Graham and Wilkinson, 1992; Gaucher et al., 2019a). Glutamate has already been reported as a compatible solutes accumulated during osmoadaptation (Empadinhas and Viete-Vallejo, 2008). Trehalose is a compatible solutes accumulated during acid adaptation and osmoadaptation by P. freudenreichii (Cardoso et al., 2007; Huang et al., 2016a; Gaucher et al., 2019a). In the present study, we combined osmoadaptation with acid or heat adaptation, in order to improve P. freudenreichii CIRM-BIA 129 resistance. We investigate the impact of the modulation of the fatty acid composition, compatible solutes accumulation and expression of general stress proteins, on P. freudenreichii tolerance toward different stresses and during freeze-drying. Combining two protective pretreatments led to enhance survival of P. freudenreichii upon freeze-drying.
Materials and Methods
Strains and Pre-culture
Propionibacterium freudenreichii CIRM-BIA 129 (equivalent ITG P20) was provided, stored and maintained by the CIRM-BIA Biological Resource Center (Centre International de Ressources Microbiennes-Bactéries d’Intérêt Alimentaire, INRA, Rennes, France). P. freudenreichii CIRM-BIA 129 was routinely cultivated in yeast extract lactate (YEL) broth (Malik et al., 1968). YEL medium contained, per liter, 21.4 g DL-lactate (60%), 10 g Yeast extract, 10 g tryptone, 0.328 g K2HPO4, and 0.056 g MnSO4. This leads to a non-protein nitrogen content of 14.2 g. P. freudenreichii was grown at 30°C without agitation under microaerophilic condition.
Bacterial Adaptation
Propionibacterium freudenreichii CIRM-BIA 129 was grown under different condition: in YEL medium (0.429 osmol), or in YEL medium with 0.9 M NaCl (YEL+NaCl, 1.958 osmol) to induce osmoadaptation. Heat or acid adaptation where realized at the beginning of the stationary phase. Heat adaptation was performed by placing 25 mL (in a 50 mL Falcon tube) of P. freudenreichii cultures in a water bath at 42°C for 1 h (Anastasiou et al., 2006), and acid adaptation was performed by adjusting the culture at pH = 5 and then incubated during 1 h at 30°C (Jan et al., 2001).
Stress Challenges
Heat, oxidative, bile salts and acid challenge were applied to cultures at the beginning of stationary-phase (when maximal OD was reached), or after adaptations. A strong heat challenge was performed by placing 2 mL (in a 15 mL Falcon tube) of P. freudenreichii culture in a water bath at 60°C for 10 min (Leverrier et al., 2004). A lower heat stress was performed by placing 2 mL (in a 15 mL Falcon tube) of P. freudenreichii culture in a water bath at 55°C for 30 min (Leverrier et al., 2004). Oxidative challenge was applied by adding 1.25 mM of hydrogen peroxide (Labogros, France) to 2 mL of P. freudenreichii culture during 1 h at 30°C (Serata et al., 2016). Acid challenge was applied by re-suspending P. freudenreichii in MMO medium adjusted to pH 2.0 by using HCl at 30°C followed by a 1 h incubation (Jan et al., 2000). Bile salts challenge was performed by adding 1 g.L–1 of a bile salts mixture (an equimolar mixture of cholate and deoxycholate; Sigma Chemical, St. Louis, MO, United States) in the culture during 1 h at 37°C (Leverrier et al., 2003). CFU counting was performed after challenge. In order to calculated survival percentage, a CFU counting was made, with untreated culture left for the same time at 30°C as a control.
Freeze-Drying
Propionibacterium freudenreichii CIRM-BIA 129 were grown in the YEL and YEL+NaCl medium and with or without acid or heat adaptation. At the beginning of the stationary phase, cultures were harvested (8000 g, 10 min, 30°C). Pellets were then homogenized in a maltodextrin solution (100 g.L–1) (Roquette, France). The bacterial solutions were then freeze-dried (2253-04, Serail, France).
Fatty Acids Analysis
Propionibacterium freudenreichii CIRM-BIA 129 was grown in YEL and YEL+NaCl medium with or without acid or heat adaptation at the beginning of the stationary-phase. Cells were then washed with sterile distilled water. Cells were harvested by centrifugation (8000 g, 15 min). Saponification was performed by adding 3 mL of sodium methoxide (3.75 M in methanol) (Sigma Chemical, St. Louis, MO, United States) and shaking vigorously for 10 s and incubated during 25 min at 100°C. Samples were cooled to performed methylation, a solution of HCL and methanol were added (HCl 3.5 M and methanol 42% final concentration), samples were vortexed 10 s and incubated during 10 min at 80°C. Before the extraction of BAME (bacterial acid methyl ester) samples were cooled on ice and a solution of hexane and diethyl ether (hexane 50% and diethyl ether 50%) was added. Samples were agitated during 10 min and then decanted, aqueous phases were removed before organic phases washing with NaOH 3 M solution. Samples were then agitated and decanted. Organic phases were then collected for analysis.
The analyses were performed on an Agilent gas chromatograph (7890A) equipped with a BPX70 capillary column (120 m × 0.25 mm × 0.25 μm, SGE, Victoria, Australia) and coupled to a flame ionization detector (Agilent Technologies, Les Ulis, France). Hydrogen was used as carrier gas, and the injection volume was 0.5 μL. Injection was done by a cool on column injector. Detection temperature was 250°C. Agilent MSD ChemStation software was used for data acquisition. Components were identified from the retention time measured from BAME (Bacterial Acid Methyl Ester CP) standards (Merck, France).
Results were expressed as relative percentages of each fatty acid, which were calculated as the ratio of the surface area of the considered peak to the total area of all peaks. The ratio of unsaturated to saturated fatty acids (U/S) was determined. Analyses were made in triplicate.
Identification and Quantification of Osmoprotectants Accumulated by P. freudenreichii CIRM-BIA 129
Extraction of Accumulated Osmoprotectants
Propionibacterium freudenreichii CIRM-BIA 129 was grown in YEL and YEL+NaCl medium with or without acid adaptation at the beginning of the stationary-phase. During exponential phase (OD = 0.8), cells were harvested by centrifugation (8000 g 10 min). Cells were washed twice in a NaCl solution with the same osmolarity than the culture medium. Cells were then re-suspended in 2 mL of distilled water, then 8 mL of absolute ethanol were added. The suspension was homogenized and centrifuged (8000 g, 10 min) in order to remove cell fragments. The supernatant extract was evaporated during 7 h with a rotary evaporator. Dried extracts were then solubilized in deuterium oxide (Sigma-Aldrich, United States).
NMR (Nuclear Magnetic Resonance) Analyses
All 1H and 13C NMR spectra were recorded at 298 K on a Bruker Avance 500 spectrometer equipped with a 5 mm TCI triple-resonance cryoprobe (PRISM core facility, Rennes). 1H spectra were acquired with a 6 kHz spectral width, 32 K data points and a total repetition time of 6.73 s. 13C spectra were acquired using a proton power-gated decoupling sequence with a 30° flip angle, a 30 kHz spectral width, 64 K data points and a total repetition time of 3.08 s. The data were processed with Topspin software (Bruker Biospin). Before applying the Fourier transform, free induction decays of 1H spectra were treated with an exponential broadening of 0.3 Hz.
Samples were solubilized in D2O. 3-(Trimethylsilyl) propionic-2,2,3,3-d4 acid sodium salt (TSP-d4) (Sigma-Aldrich, United States) served as an internal reference for 1H and 13C chemical shifts. The relative concentration of trehalose, glutamate and glycine betaine in the samples was determined by the integration of the peaks’ areas of their 1H signals relative to the internal standard TMSP. Results are expressed as NMR relative units (RU).
Label Free Proteomics
Whole-Cell Protein Extraction and Protein Tryptic Digestion
The label free proteomics has been conducted as described by Gaucher et al. (2019a). At the beginning of stationary phase or after acid adaptation, P. freudenreichii cells were harvested by centrifugation and washed twice with PBS buffer (NaCl 8 g.L–1, KCl 2 g.L–1 KH2PO4 2 g.L–1, Na2HPO4 12H2O 35,8 g.L–1). Briefly, cells pellets were washed, and cells were resuspended in a lysis solution. The solution was sonicated and cells were broken using zirconium beads in a homogenizer. Proteins were harvested, cleaned and quantified before digestion and protein were extracted before digestion. Tryptic digestion was performed on 100 μg of whole-cell proteins from each sample during 15 h at 37°C using Sequencing Grade Modified Trypsin (Promega, Madison, WI, United States) according to the manufacturer’s instructions and as described previously (Huang et al., 2018). Spectrophotometric-grade trifluoroacetic acid (TFA) (Sigma-Aldrich, United States) was added in order to stop the digestion.
Nano-LC-MS/MS
Experiments were performed as previously described (Huang et al., 2018). Briefly, experiments were performed using a nano RSLC Dionex U3000 system fitted to a Q-Exactive mass spectrometer (Thermo Fisher Scientific, San Jose, CA, United States) equipped with a nano-electrospray ion source. Peptides separation was performed on a reversed-phase column (PepMap 100 C18, 75 μm i.d., 250 mm length, 3 μm particle size, 100 Å pore size; Dionex, Amsterdam, Netherlands) and The spectra of eluted peptides were recorded in full MS mode and selected in a mass range 250–2000 m/z for MS spectra with a resolution of 70,000 at m/z 200. For each scan, the ten most intense ions were selected for fragmentation. MS/MS spectra were recorded with a resolution of 17,500 at m/z 200 and the parent ion was subsequently excluded from MS/MS fragmentation for 20 s. The instrument was externally calibrated according to the supplier’s instructions.
Protein Identification
Protein identification was performed as previously described (Huang et al., 2018). Peptides were identified from the MS/MS spectra using X!Tandem pipeline software (Langella et al., 2017). The search was performed against the proteome of strain P. freudenreichii CIRM-BIA 129 (ITG P20) (downloaded from NCBI.nlm.nih.gov on the 23rd of August 2018). The strain is registered as “P. freudenreichii subsp. freudenreichii ITG P20” (GenBank: CCBE000000000.1, BioProject PRJEB4826). Database search parameters were specified as follows: trypsin cleavage was used and the peptide mass tolerance was set to 10 ppm for MS and 0.05 Da for MS/MS. Oxidation of methionine and phosphorylation of threonine, serine and tryptophan were selected as a variable modification. For each peptide identified, a minimum score corresponding to an e-value below 0.05 was considered as a prerequisite for peptide validation.
Protein Quantification
Protein quantification was performed as previously described (Huang et al., 2018). Every peptide identified by tandem mass spectrometry was quantified using the free MassChroQ software (Valot et al., 2011) before data treatment and statistical analysis within the R software (R 3.2.2, Project for statistical computing). A specific R package called “MassChroqR” was used to automatically filter dubious peptides for which standard deviation of retention time was superior to 30 s and to regroup peptide quantification data into proteins. For peak counting analysis, variance analysis was performed on proteins with a minimum peak ratio of 1.5 between both culture conditions. Proteins with an adjusted p-value < 0.05 were considered significantly different.
For XIC based quantification, normalization was performed to take into account possible global quantitative variations between LC-MS runs. Peptides shared between different proteins were automatically excluded from the data set as well as peptides present in less than 85% of samples. Missing data were then imputed from a linear regression based on other peptide intensities for the same protein (Blein-Nicolas et al., 2015). Analysis of variance was used to determine proteins with significantly different abundance between our two culture conditions.
SDS-PAGE and Western-Blotting
The extraction, the SDS-Page and the Western blot have been conducted as Rabah et al. (2018). Briefly, P. freudenreichii was grown in YEL or YEL+NaCl with or without acid or heat adaptation. Cells were harvested by centrifugation, washed two times with PBS. After centrifugation, supernatants were removed and lysis solution (Tris–HCl 50 mM pH 7.5, SDS 0.3%, DTT200 mM) was added to bacterial pellets and samples were frozen prior to sonication and cell lysis using Precellys® Evolution homogenizer. Resulting SDS extracts were recovered by centrifugation (21,000 × g, 4°C, 20 min) and analyzed by one-dimensional electrophoresis SDS-PAGE. The protein extracts from triplicate experiments were pooled and the protein amount was determined using 2-D Quant Kit (GE Healthcare). Protein extracts were separated by 12% SDS–PAGE and then transferred to PVDF membranes (GE Healthcare). A solution with 3% non-fat dry milk diluted in TBS (Tris 10 mM, NaCl 0.15 M, 0.3% tween 20) was added on the PVDF membranes to perform locking. Membranes were washed and then incubated over-night at 4°C with anti-SlpB primary antibodies purified from rabbit sera (AGRO-BIO, France) at the dilution 1:10,000. After washing, membranes were incubated with secondary antibodies: anti-rabbit IgG conjugated with horseradish peroxidase (1:15,000, AGRO-BIO, France) for 2 h at room temperature. Bound antibodies were visualized with ECL Plus system (GE Healthcare, Vélizy, France) and blots were scanned using the Syngene GBox (Ozyme, Saint-Quentin-en-Yvelines, France).
Statistical Analysis
The data were from triplicate samples. All the results are presented as mean value with standard deviation. Statistical analyses using one-way ANOVA with Tukey post hoc analyses for multiple comparison. Calculations were performed using GraphPad Prism Software (Prism 7 for Windows).
Results
P. freudenreichii Viability During Challenges and Freeze-Drying
We subjected P. freudenreichii to freeze-drying, heat, acid, and bile salts stress challenges, in order to monitor its stress tolerance. P. freudenreichii CIRM-BIA 129 viability was determined by numeration (CFU counting) before and after these challenges, which were selected as being relevant to freeze-drying process, Swiss-type cheese making, and to digestive constraints. As shown in Figure 1A, addition of salt (0.9 M NaCl) to the YEL growth medium increased P. freudenreichii CIRM-BIA 129 viability during freeze-drying (43 to 74.4%), yet showing no effect on its viability during heat lethal challenge (55°C, 30 min) (Figure 1B). This hyperosmotic constraint even had a negative impact on P. freudenreichii CIRM-BIA 129 survival during acid (pH 2, 1 h, 36.4 to 1.8%) and bile salt (1 g.L–1, 1 h, 38.3 to 0%) challenges (Figures 1C,D). Osmoadaptation, acid adaptation, and the combination thereof led to a viability below the detection threshold during bile salts challenge. A sublethal heat pretreatment (42°C, 1 h) had per se a positive impact on survival during a subsequent heat lethal challenge (Figure 1B, 32.5 to 87.6%), yet a negative impact on survival during acid challenges (Figure 1C, 36.4 to 10%, respectively). Acid adaptation (pH 5, 1 h) per se did not enhance tolerance, neither toward these lethal challenges, nor toward freeze-drying. Heat adaptation, or acid adaptation, combined to osmoadaptation, had a negative or no impact on P. freudenreichii CIRM-BIA 129 tolerance toward stress challenges drying (Figures 1B–D). However, such a combination enhanced P. freudenreichii CIRM-BIA 129 survival during freeze-drying (Figure 1A, 43 to 90.5 and 96.7%, respectively). Best viability during freeze-drying was obtained for P. freudenreichii CIRM-BIA 129 grown in YEL+NaCl, with an acid-adaptation at the beginning of the stationary phase. Further analyses were then performed on P. freudenreichii CIRM-BIA 129, grown in YEL or in YEL+NaCl, with or without acid or heat adaptation, in order to elucidate mechanism responsible for enhanced tolerance toward freeze-drying.
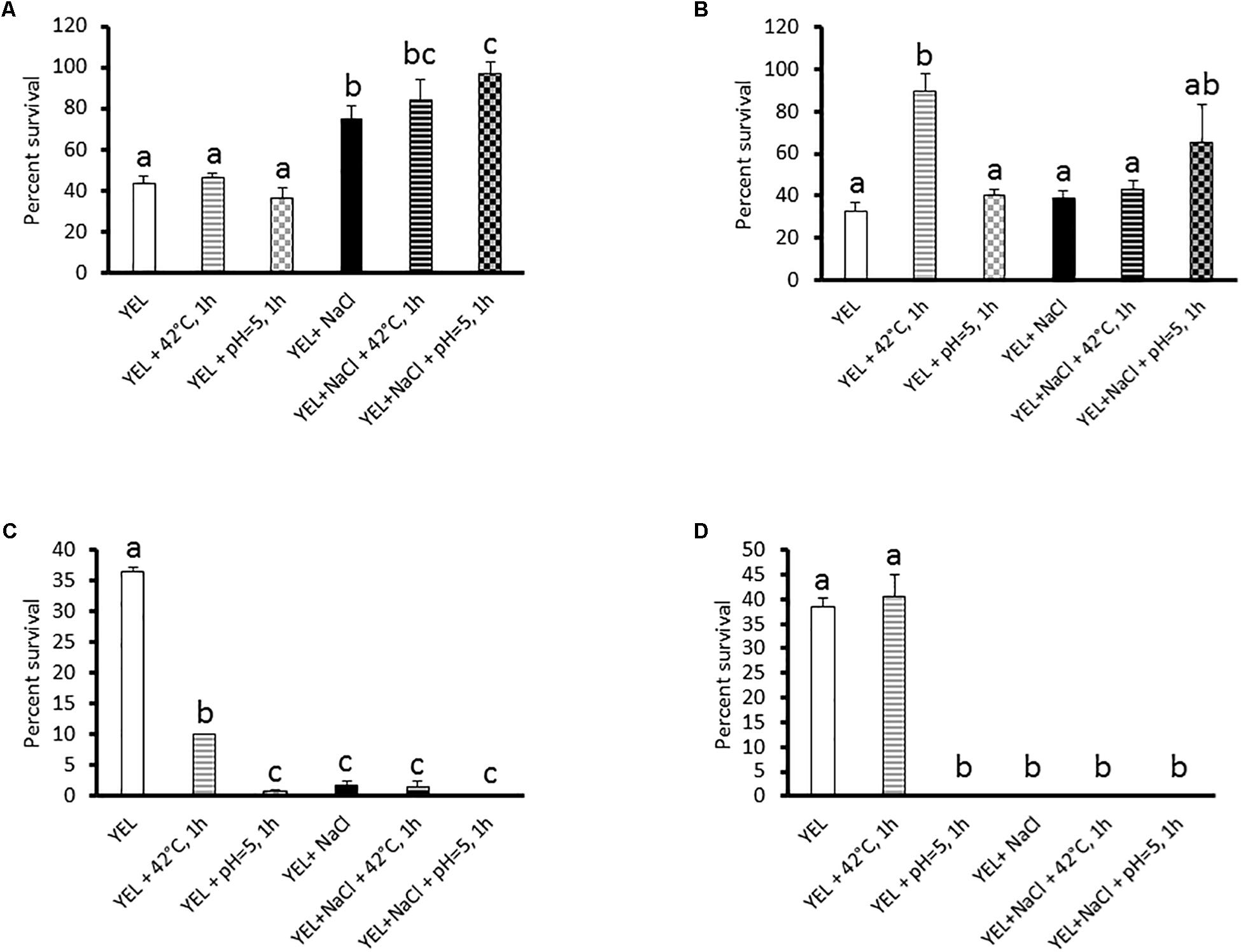
Figure 1. Viability of Propionibacterium freudenreichii CIRM-BIA 129 after freeze-drying (A), heat (B), acid (C), and bile salts (D) challenges. The strain was previously grown in YEL and YEL+NaCl broths, and subjected to heat and acid adaptation. Errors bars represent the standard deviation for triplicate experiments. Significances differences are reported with different letters above columns (p > 0.05). YEL: Yeast Extract Lactate. 42°C, 1 h: thermo-adaptation at 42°C during 1 h. pH = 5, 1 h: acid-adaptation at pH = 5 during 1 h.
Membrane Fatty Acids Composition Is Modulated by Acid, Heat and Osmoadaptation
The fatty acid composition of membrane lipids was analyzed to better understand P. freudenreichii CIRM-BIA 129 adaptation to high osmotic pressure, to thermal stress or to acidification. As shown in Figure 2, a total of 23 fatty acids were detected in P. freudenreichii CIRM-BIA 129 membrane, using gas chromatography, reported in Supplementary Table S1. This includes saturated, unsaturated and branched fatty acids. P. freudenreichii CIRM-BIA 129 contains a high proportion of branched fatty acids (52.37–62.15%), and lower amounts of saturated (13.54–18.30%), and unsaturated ones (4.28–16.45%). The anteiso C15:0 was detected as the major fatty acid, which proportion was furthermore highly modulated during P. freudenreichii CIRM-BIA 129 adaptations (Figure 1A). Osmoadaptation increased the amount of long fatty acids (anteiso C17:0, C17:0, and C19:0) and decreased the amount of short fatty acid (C12:0 and C13:0). Accordingly, the C15-C22/C10-C14 ratio was enhanced as a result of osmoadaptation and of acid adaptation (Figure 2B). The amount of C16;1n7 and C18:1n9c decrease sharply for the bacteria grown in salty medium, by contrast the saturated fatty acid C16:0 and C17:0 are increasing during osmoadaptation (Supplementary Table S1). Such variations led to enhanced saturated/unsaturated ratio upon osmoadaptation (Figure 2C). Heat or acid-adaptation, applied after osmoadaptation can modulate this ratio. Saturated and branched fatty acid proportions are modulated in the different conditions, but the ratio saturated/branched did not show significant variations (Figure 2D).
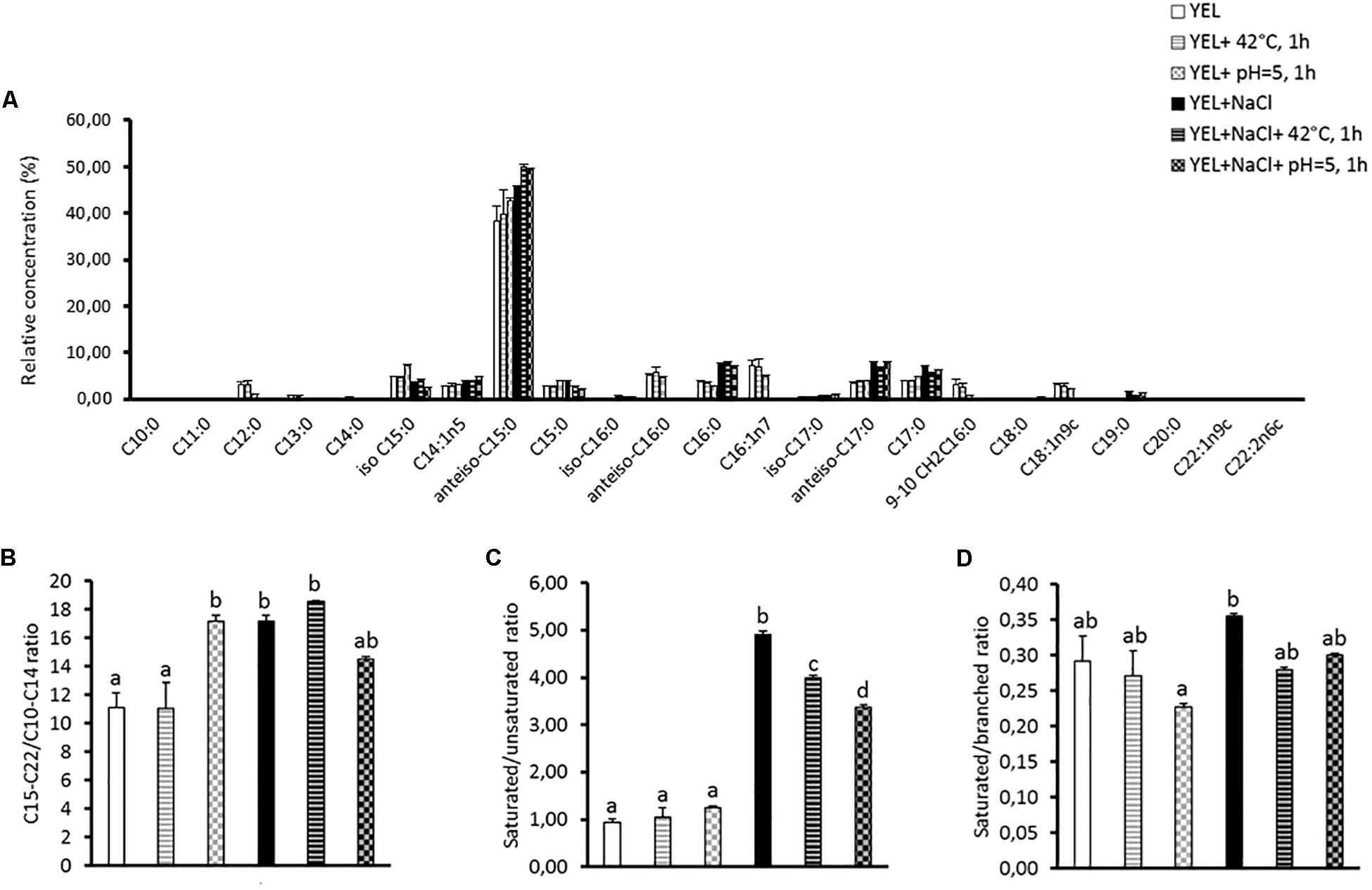
Figure 2. Membrane fatty acids (% of total membrane fatty acids) of Propionibacterium freudenreichii CIRM-BIA 129 in different conditions. The fatty acids composition (A), the saturated/unsaturated fatty acids ratios (SFA/UFA) (B), the saturated/branched fatty acids ratio (SFA/BFA) (C) and the C15-C22/C10-C14 ratio (D) are shown for the strain grown in YEL and YEL+NaCl broths and subjected to heat and acid adaptation. Error bars represent the standard deviation for triplicate experiments. Significant differences are reported with different letters above the columns (p > 0.05). YEL: Yeast Extract Lactate. 42°C, 1 h: thermo-adaptation at 42°C during 1 h. pH = 5, 1 h: acid-adaptation at pH = 5 during 1 h.
Osmoprotectants Accumulation Is Modulated by the Acid and Osmoadaptation
When cultivated in the isotonic and rich YEL growth medium in the absence of NaCl and of acid or heat stress, no trehalose, no glutamate and no glycine betaine was detected in the cytoplasm of P. freudenreichii CIRM-BIA 129 (Figure 3). However, acid adaptation, at the end of growth, at the beginning of stationary phase, triggered glutamate accumulation in the same medium. During growth in rich YEL medium in the presence of NaCl, P. freudenreichii CIRM-BIA 129 mainly accumulated glycine betaine, as well as lower amounts of glutamate and of trehalose. In these osmoadapted cultures, acid adaptation at the beginning of the stationary phase further increased accumulation of glycine and of trehalose, at the expense of glutamate.
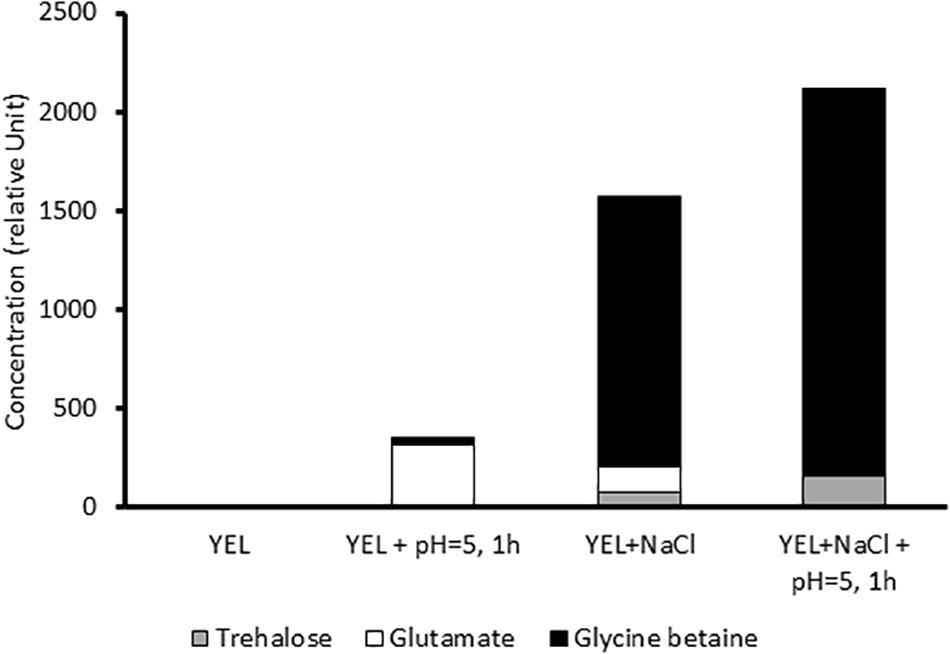
Figure 3. Compatible solutes accumulation (relative units) after growth in YEL and YEL+NaCl broths, with and without acid adaptation.
Modulation of Protein Expression During Osmoadaptation and Acid Adaptation
To understand the respective impact of P. freudenreichii CIRM-BIA 129 osmoadaptation and of osmoadaptation coupled to acid adaptation, we realized a proteomic differential analysis. Theoretically, 2176 proteins are encoded by P. freudenreichii CIRM-BIA 129 genome (GCA_000723545.1). In this study, a total of 857 proteins were detected using mass spectrometry. This represents a coverage of 39%. Proteomics data have been deposit into INRA data base (Jardin, 2019). We focused on proteins that were differentially abundant for at least one condition between YEL, YEL+NaCl, and YEL+NaCl+pH = 5, 1 h, with a minimum ratio of 1.5 or less than 0.66. A total of 216 proteins were significantly modulated, 208 between YEL and YEL+NaCl (see Supplementary Table S2) and eight proteins between YEL+NACl and YEL+NaCl+pH = 5, 1 h (Table 1). As shown in Figure 4, the heat map indicated that the two proteomes corresponding to, respectively, YEL and YEL+NaCl, were very different from the control one corresponding to YEL. By contrast, the proteome corresponding to YEL+NACl+pH = 5, 1 h is very closed to that of bacteria grown in YEL+NaCl. Salt addition strongly affected P. freudenreichii CIRM-BIA 129 proteome, while acid adaptation after osmoadaptation modulated the expression of few proteins. More precisely, 20 proteins modulated during osmoadaptation were involved in amino acids metabolism and transport, 17 proteins in carbohydrate transport and metabolism, five proteins in lipid transport and metabolism, six proteins in cell wall-membrane-envelope biogenesis. Another set of 17 proteins belonged to the “post-translational modification, protein turnover and chaperones” category (Table 2). All the other modulated proteins are presented in Supplementary Table S2.
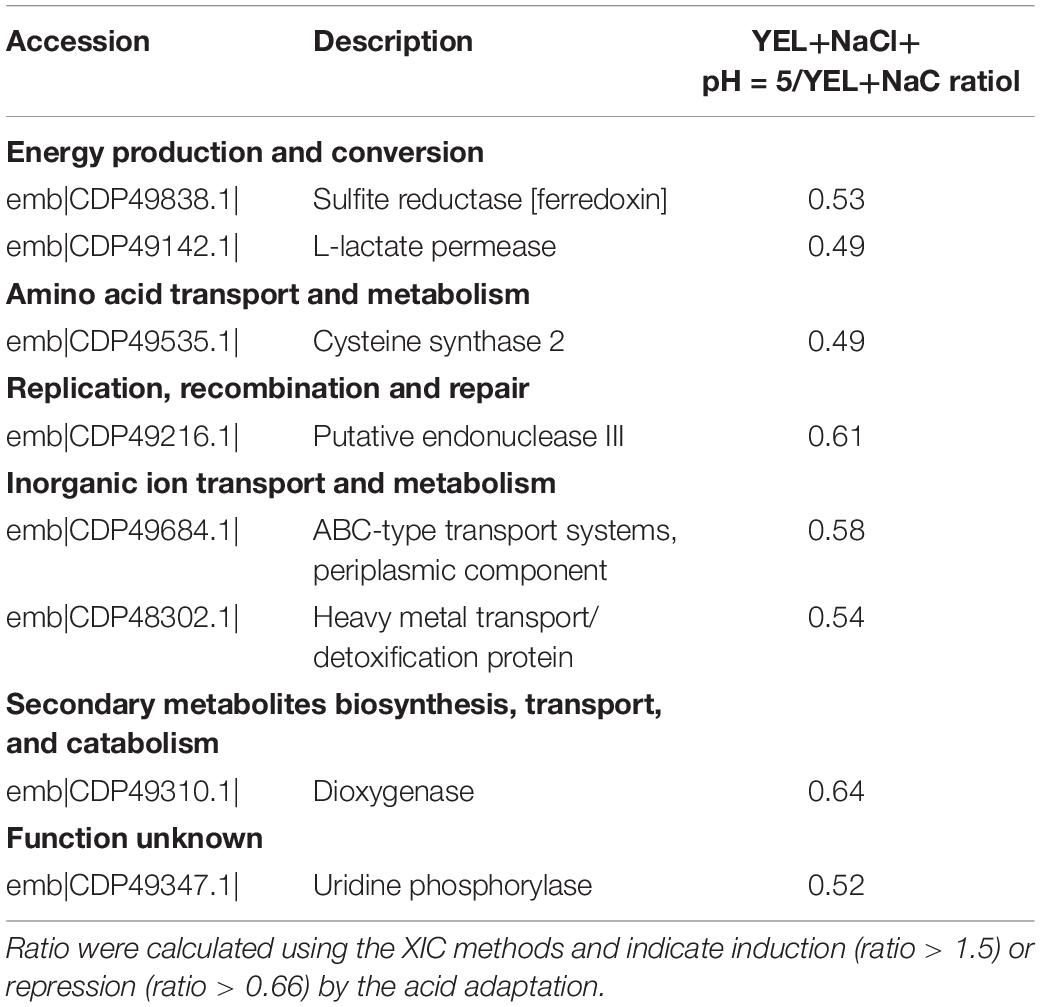
Table 1. Proteins modulated during acid-adaptation after osmoadaptation. Proteins were determined by using a database composed of proteome of P. freudenreichii CIRM-BIA 129 (downloaded from NCBI.nlm.nih.gov 2018).
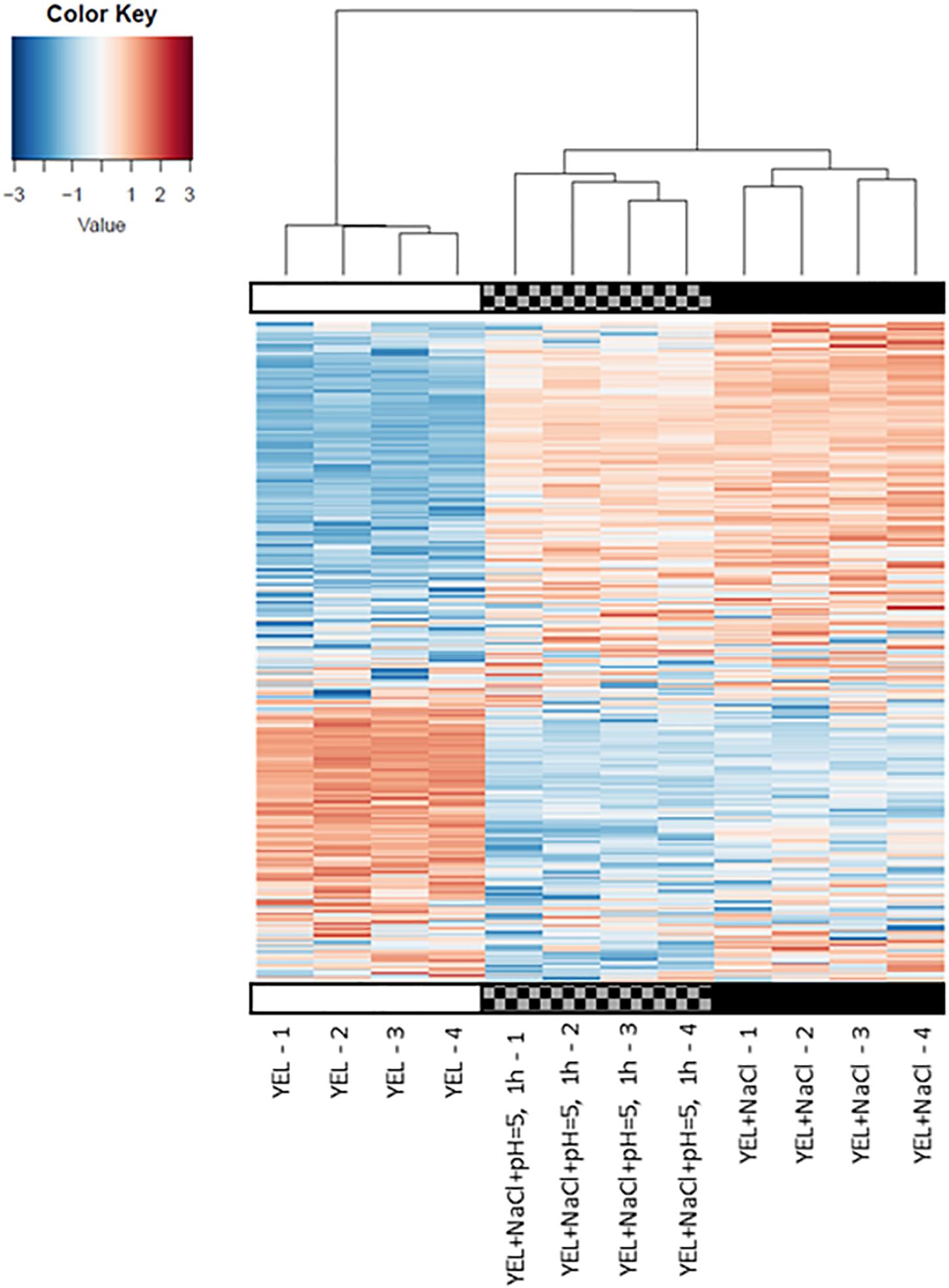
Figure 4. Effects of osmoadaptation and acid-adaptation on P. freudenreichii CIRM-BIA 129 proteome.
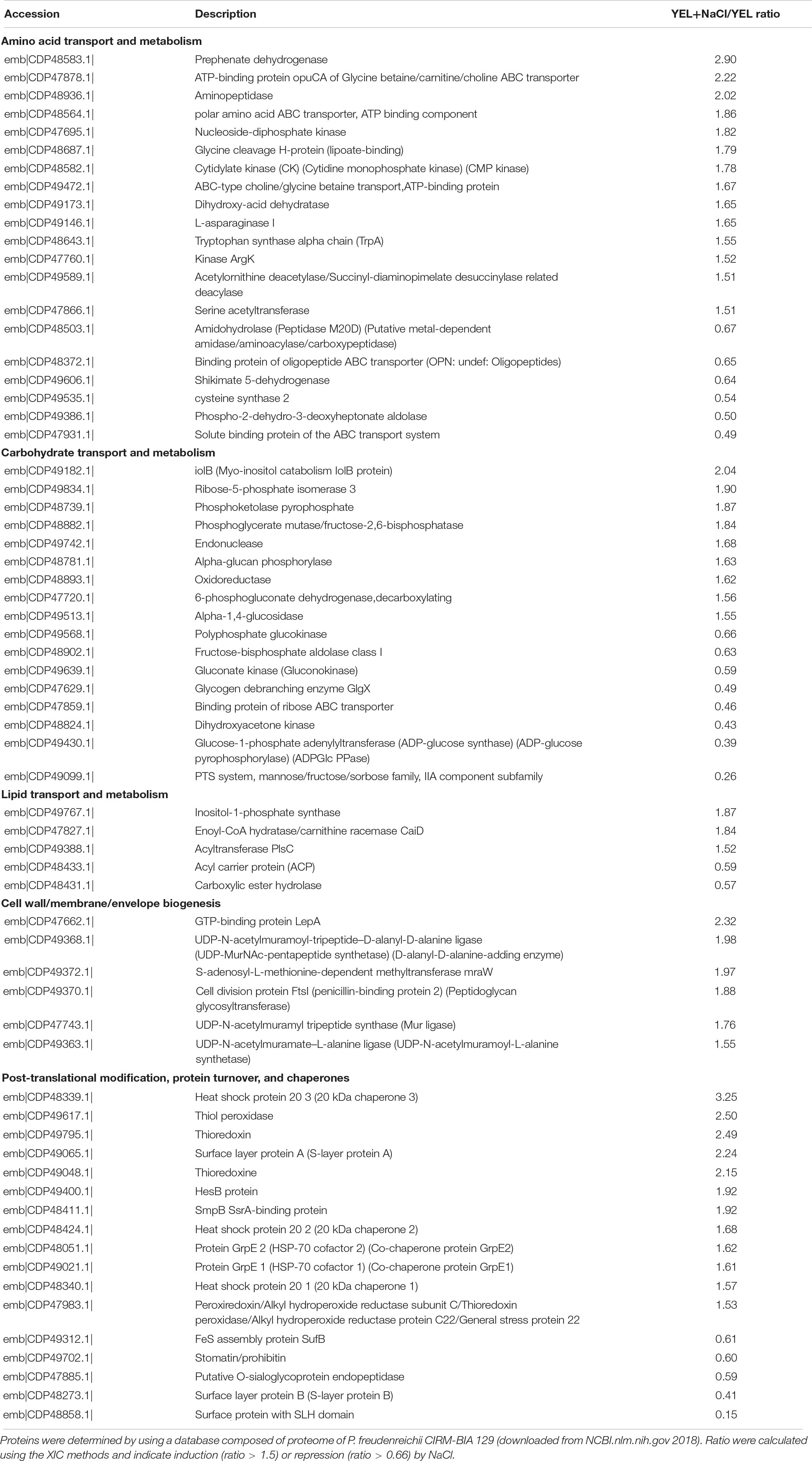
Table 2. Proteins modulated during osmoadaptation belonging to the COG category “amino acid metabolism and transport,” “carbohydrate transport and metabolism,” “lipid transport and metabolism” and “cell wall-membrane-envelope biogenesis.”
Modulation of S-Layer Protein B on P. freudenreichii CIRM-BIA 129 Cell Wall
Western blotting, using antibodies directed against P. freudenreichii CIRM-BIA 129 SlpB protein, allowed detection of this protein in all P. freudenreichii whole-cell protein extracts, whatever the growth conditions, but to different extents (Figure 5A). Indeed, the amount of detected SlpB was lower for all the cultures performed in YEL with added NaCl. Concerning the culture supernatants, no SlpB was detected in control conditions, i.e., cultures in the rich YEL medium. However, heat and acid adaptation led to the release of a part of SlpB into the growth supernatant. Less SlpB protein was detected in whole cell extracts when P. freudenreichii CIRM-BIA 129 was cultivated in the presence of NaCl, while traces of this protein were detected in the corresponding supernatants.
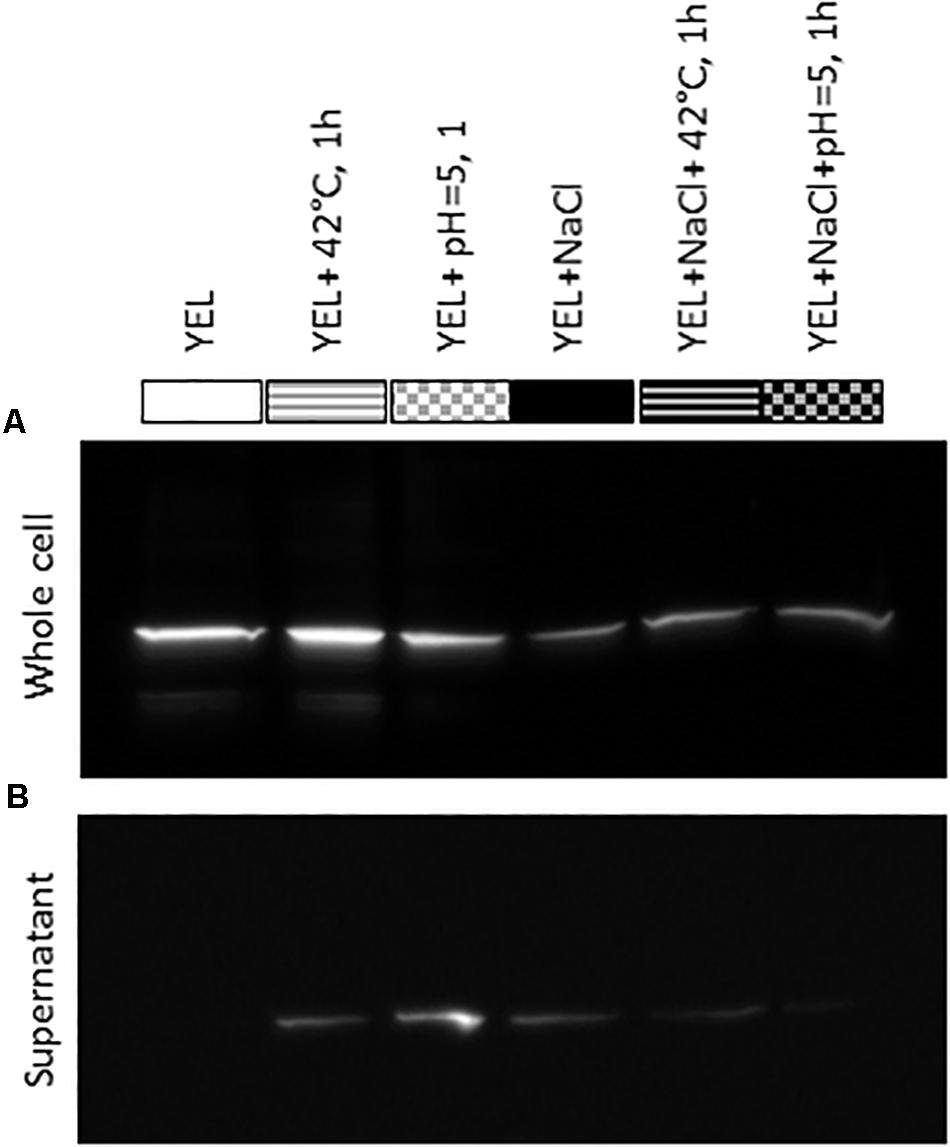
Figure 5. Detection of SlpB protein in Propionibacterium freudenreichii CIRM-BIA 129 whole-cell protein extracts (A) and supernatants (B) for cultures performed in YEL and YEL+NaCl broths, and after heat and acid adaptations. YEL: Yeast Extract Lactate. 42°C, 1 h: thermo-adaptation at 42°C during 1 h. pH = 5, 1 h: acid-adaptation at pH = 5 during 1 h.
Discussion
Heat and Acid-Adaptations, Coupled to Osmoadaptation, Trigger Cross-Protection to Lethal Challenges
Osmoadaptation, i.e., growth in the rich YEL medium in the presence of 0.9 M NaCl, triggered enhanced viability of P. freudenreichii CIRM-BIA 129 during freeze drying, as reported previously for P. freudenreichii and for L. bulgaricus (Carvalho et al., 2003; Gaucher et al., 2019a). Remarkably, a mild acid adaptation, coupled to such an osmoadaptation, further enhanced P. freudenreichii CIRM-BIA 129 tolerance toward freeze-drying. This opens new perspectives to improve industrial production of this probiotic. However, osmoadaptation, coupled or not to heat or acid-adaptation, dramatically reduced P. freudenreichii CIRM-BIA 129 tolerance toward the digestive stresses mimicked by acid and bile salt challenges. By contrast, acid adaptation reportedly increases acid tolerance in another strain of P. freudenreichii and in L. casei (Jan et al., 2000; Broadbent et al., 2010). Moreover, heat adaptation increases bile salts and acid tolerance in P. freudenreichii (Jan et al., 2000, 2002; Leverrier et al., 2004). In the present study, heat adaptation alone increased P. freudenreichii CIRM-BIA 129 viability during heat challenge, in accordance with the increased thermotolerance of heat-adapted P. freudenreichii (Anastasiou et al., 2006), L. paracasei and Lactobacillus kefiranofaciens to heat shock (Desmond et al., 2001). To our knowledge, the present report is the first describing optimal tolerance toward freeze-drying, as a result of osmoadaptation combined with mild acid pretreatment.
Modulation of the Membrane Fatty Acid Composition Increases the Viability During Freeze-Drying
Osmoadaptation strongly increased the saturated/unsaturated fatty acids ratio in the membrane of P. freudenreichii CIRM-BIA 129, as reported for L. casei (Machado et al., 2004). Heat adaptation reportedly decreases this ratio in the Swiss-type cheese starter Lactobacillus helveticus (Lanciotti et al., 2001), while not in P. freudenreichii CIRM-BIA 129 in the present study. Heat or acid-adaptation modulated the increase of the ratio saturated/unsaturated triggered by osmoadaptation. The ratio SFA/BFA (BFA: Branched fatty acid) was not significantly altered whatever the growth and treatment conditions. Osmoadaptation led to enhanced fatty acid chain length. Indeed, salt addition and acid adaptation, but not thermal adaptation, increased the C15-C22/C10-C14 ratio. Thus, P. freudenreichii CIRM-BIA 129 responded to environmental stresses by modulating its membrane fluidity via the saturated/unsaturated ratio, with a profound increase of anteiso-C15:0, the main membrane fatty acid in P. freudenreichii (Thierry et al., 2011) and known to play a key role in bacterial stress adaptation (Annous et al., 1997). Anteiso-C15, a fatty acid abundant in halophilic bacteria (Guan et al., 2018; Jang et al., 2018; Ding et al., 2019), is accordingly increased under hyperosmotic conditions in halotolerant Gram-positive bacteria such as Planococcus sp. (Miller, 1985), Staphylococcus epidermidis (Komaratat and Kates, 1975), and S. aureus (Kanemasa et al., 1972).
Acid Adaptation, Coupled to Osmoadaptation, Increases Compatible Solutes Accumulation
Propionibacterium freudenreichii CIRM-BIA 129 accumulated in this study high amounts of glycine betaine during osmoadaptation, as already described (Gaucher et al., 2019a). Glycine betaine is an important compatible solute accumulated by different bacteria such as L. plantarum and Lactococcus lactis during osmoadaptation (Glaasker et al., 1996; Romeo et al., 2003). Only limited amounts of glutamate and trehalose were accumulated by P. freudenreichii CIRM-BIA 129 during osmoadaptation. Glutamate is a key molecule of the primary osmoregulation response (Csonka and Hanson, 1991) and is accumulated by L. plantarum (Glaasker et al., 1996; Kets et al., 1996). It should be noted that the rich YEL medium, composed of yeast extract, lactate and peptone, contains non-protein nitrogen (12.4 g), including glutamate and glycine betaine. The P. freudenreichii genome allows prediction of at least two glycine betaine import systems, i.e., OpuCA and an ABC-type choline/glycine betaine transporter (see below). Accordingly, P. freudenreichii was previously shown to accumulate glycine betaine in hyperosmotic conditions (Boyaval et al., 1999). Trehalose is accumulated in many different conditions such as osmotic, acid, oxidative and cold stresses (Cardoso et al., 2004, 2007; Thierry et al., 2011; Huang et al., 2016b). Trehalose can be synthetized by the OtsA-Otsb and TreS pathway previously described in P. freudenreichii (Cardoso et al., 2007). Accumulation of these three osmoprotectants, in response to osmotic constraint, is thus a general adaptation mechanism (Csonka and Hanson, 1991). Interestingly, acid adaptation further enhanced osmoprotectants intracellular accumulation and particularly that of glycine betaine. This enhanced accumulation is in accordance with the enhanced tolerance toward the freeze-drying challenge. To better address mechanisms responsible for such an adaptation, a whole-cell proteomic differential analysis was thus undertaken.
NaCl Induces Major Changes in P. freudenreichii CIRM-BIA 129 Proteome, While Acid Adaptation Down-Regulated Part of the NaCl Stress Proteome
Osmoadaptation triggered major modifications in the whole-cell protein content of P. freudenreichii CIRM-BIA 129, as illustrated by the heatmap in Figure 4. The patterns corresponding to YEL and to YEL+NaCl exhibit numerous differences, while acid adaptation had little impact on the proteome. The NaCl stress proteome included many proteins involved in amino acid transport and metabolism. In particular, the ABC-type choline/glycine betaine transporter (emb|CDP49472.1|) and the ATP-binding protein OpuCA of glycine betaine/carnitine/choline ABC transporter (emb|CDP47878.1|) were in higher abundance, in accordance with the observed glycine betaine accumulation in the presence of salt (0.9 M NaCl). Different proteins such as a polar amino acid ABC transporter, an aminopeptidase and enzymes involved in amino acid biosynthesis were also enhanced and this shall lead to an enhanced intracellular pool of amino acids. Prephenate dehydrogenase (emb|CDP48583.1|) and L-asparaginase (emb|CDP49146.1|), which liberate ammonia/ammonium from amino acids, take part in intracellular pH homeostasis. L-asparaginase has already been reported as a stress protein in Salmonella typhimurium (Sonck et al., 2009).
About proteins involved in the transport and metabolism of carbohydrate, proteins corresponding to the utilization of alternative carbon sources are induced, including Myo-inositol catabolism IolB protein (emb|CDP49182.1|), as it happens during unfavorable conditions in Corynebacterium glutamicum (Chen et al., 2019). The pentose phosphate pathway, including the phosphoketolase pyrophosphate (emb|CDP48739.1|) and the ribose-5-phosphate isomerase 3 (emb|CDP49834.1|), is also enhanced. Stress-induction of this pathway, involved in intracellular homeostasis, has already been described for propionibacteria (Gaucher et al., 2019a), for bifidobacteria (Sanchez et al., 2007) and for bacilli (Goswami et al., 2018). By contrast, PTS (PhosphoTransferase System, emb|CDP49099.1|) is reduced by NaCl as in Listeria monocytogenes (Bae et al., 2012), and so is glycolysis (fructose-bisphosphate aldoase 1, emb|CDP48902.1|) and glycogen utilization (glycogen debranching enzyme GlgX, emb|CDP47629.1|).
Concerning lipid and transport metabolism, the induction of inositol-1-phosphate synthase (emb|CDP49767.1|) and of acyltransferase PslC (emb|CDP49388.1|) indicates enhanced biosynthesis of phospholipids, in accordance with the observed effect of salt on membrane fatty acids. These proteins are also involved in stress response in Serratia plymuthica and C. glutamicum (Nurlinawati et al., 2015; Chen et al., 2019).
Concerning cell wall, enzymes involved in peptidoglycan synthesis, such as UDP-N-acetylmuramate-L-alanine ligase (emb|CDP49363.1|), which adds short polypeptides to UDP-acetylmuramic acid, are enhanced. So is the cell division protein FtsI (emb|CDP49370.1|). By contrast, the S-layer-type proteins anchored into the cell wall via SLH domains, i.e., SlpB (emb|CDP48273.1|) was reduced in the presence of salt, in accordance with the western blot analysis (Figure 5) showing reduce SlpB. SlpB is known to be involved in immunomodulation by P. freudenreichii, with an anti-inflammatory effect (Deutsch et al., 2017). By contrast, P. freudenreichii CIRM-BIA 129 surface protein SlpA (emb|CDP49065.1|) was enhanced in the presence of NaCl, in accordance with a report showing an impact of osmotic stress on the expression of S-layer proteins in L. acidophilus (Palomino et al., 2016).
Adaptation to the osmotic constraint clearly enhanced the amount of general stress proteins involved in protein repair or turnover. This includes chaperones such as the three paralogs of Hsp 20 in P. freudenreichii CIRM-BIA 129, which protect other proteins from denaturations, as well as the two paralogs of GrpE, which prevent the aggregation of stress-denatured proteins. NaCl adaptation also enhanced proteins involved in oxidative remediation like thioredoxin (emb|CDP49795.1| and emb|CDP49048.1|), thiol peroxidase (emb|CDP49617.1|) and peroxiredoxin (emb|CDP47983.1|). In addition, the enhanced SmpB protein belongs to the superfamily of SsrA-binding proteins, which are RNA-binding proteins involved in translational surveillance (Moore and Sauer, 2007).
Acid adaptation, after osmoadaptation, did not induce important additional changes in P. freudenreichii CIRM-BIA 129 proteome, compared to osmoadaptation per se. Only eight proteins were repressed during the acid adaptation after the osmoadaptation.
Conclusion
As a conclusion, this work highlights the positive impact of coupling osmoadaptation with a mild acid pretreatment, prior to freeze-drying. It moreover allows better understanding of adaptation mechanisms. The high accumulation of glycine betaine and the modulation of membrane fatty acid composition increased P. freudenreichii CIRM-BIA 129 viability during freeze-drying, opening new perspectives for efficient production of live and active probiotic propionibacteria.
Data Availability Statement
All datasets generated for this study are included in the manuscript/Supplementary Files.
Author Contributions
FG, KK, HR, SB, JO, SP, JJ, and VB-B performed the experiments. PM, PB, RJ, and GJ supervised the work. All the authors participated in the writing of the manuscript.
Funding
The authors declare that this study received funding from Bioprox and ANRT. The funders financed the thesis project Viabilité Améliorée de Bactéries BENEfiques.
Conflict of Interest
FG, PM, and PB was employed by company Bioprox and HR was employed by company Bba.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
FG is the recipient of a joint doctoral fellowship from Bioprox and from the French ANRT (Association Nationale de la Recherche et de la Technologie). The authors thank Ella Guiole and Eva Cherin for technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.02324/full#supplementary-material
TABLE S1 | Membrane fatty acids composition after different adaptations. The strain was previously grown in YEL and YEL+NaCl broths, and subjected to heat and acid-adaptation. Relative membrane fatty acids concentration are expressed in percentage.
TABLE S2 | Proteins modulated during osmoadaptation. Proteins were determined by using a database composed of proteome of P. freudenreichii CIRM-BIA 129 (downloaded from NCBI.nlm.nih.gov 2018). Ratio were calculated using the XIC methods and indicate induction (ratio > 1.5) or repression (ratio > 0.66) by NaCl.
References
Anastasiou, R., Leverrier, P., Krestas, I., Rouault, A., Kalantzopoulos, G., Boyaval, P., et al. (2006). Changes in protein synthesis during thermal adaptation of Propionibacterium freudenreichii subsp. shermanii. Int. J. Food Microbiol. 108, 301–314. doi: 10.1016/j.ijfoodmicro.2005.11.015
Annous, B. A., Becker, L. A., Bayles, D. O., Labeda, D. P., and Wilkinson, B. J. (1997). Critical role of anteiso-C15:0 fatty acid in the growth of Listeria monocytogenes at low temperatures. Appl. Environ. Microbiol. 63, 3887–3894.
Bae, D., Liu, C., Zhang, T., Jones, M., Peterson, S. N., and Wang, C. (2012). Global gene expression of Listeria monocytogenes to salt stress. J. Food Protect. 75, 906–912. doi: 10.4315/0362-028X.JFP-11-282
Blein-Nicolas, M., Albertin, W., da Silva, T., Valot, B., Balliau, T., Masneuf-Pomarède, I., et al. (2015). A systems approach to elucidate heterosis of protein abundances in yeast. Mol. Cell. Proteomics 14, 2056–2071. doi: 10.1074/mcp.M115.048058
Bouglé, D., Roland, N., and Lebeurrier, F. (1999). Effect of Propionibacteria supplementation on fecal Bifidobacteria and segmental colonic transit time in healthy human subjects. Scand. J. Gastroenterol. 34, 144–148. doi: 10.1080/00365529950172998
Boyaval, P., Deborde, C., Corre, C., Blanco, C., and Bégué, É (1999). Stress and osmoprotection in propionibacteria. Dairy Sci. Technol. 79, 59–69.
Broadbent, J. R., Larsen, R. L., Deibel, V., and Steele, J. L. (2010). Physiological and transcriptional response of Lactobacillus casei ATCC 334 to acid stress. J. Bacteriol. 192, 2445–2458. doi: 10.1128/JB.01618-9
Butel, M.-J., Waligora-Dupriet, A.-J., and Wydau-Dematteis, S. (2018). The developing gut microbiota and its consequences for health. J. Dev. Orig. Health Dis. 9, 590–597. doi: 10.1017/S2040174418000119
Cardoso, F. S., Castro, R. F., Borges, N., and Santos, H. (2007). Biochemical and genetic characterization of the pathways for trehalose metabolism in Propionibacterium freudenreichii, and their role in stress response. Microbiology 153, 270–280. doi: 10.1099/mic.0.29262-0
Cardoso, F. S., Gaspar, P., Hugenholtz, J., Ramos, A., and Santos, H. (2004). Enhancement of trehalose production in dairy propionibacteria through manipulation of environmental conditions. Int. J. Food Microbiol. 91, 195–204. doi: 10.1016/S0168-1605(03)00387-8
Carvalho, A. S., Silva, J., Ho, P., Teixeira, P., Malcata, F. X., and Gibbs, P. (2003). Effects of addition of sucrose and salt, and of starvation upon thermotolerance and survival during storage of freeze-dried Lactobacillus delbrueckii ssp bulgaricus. J. Food Sci. 68, 2538–2541. doi: 10.1111/j.1365-2621.2003.tb07057.x
Carvalho, A. S., Silva, J., Ho, P., Teixeira, P., Malcata, F. X., and Gibbs, P. (2004). Relevant factors for the preparation of freeze-dried lactic acid bacteria. Int. Dairy J. 14, 835–847. doi: 10.1016/j.idairyj.2004.02.001
Chang, H.-Y., Chen, J.-H., Chang, J.-H., Lin, H.-C., Lin, C.-Y., and Peng, C.-C. (2017). Multiple strains probiotics appear to be the most effective probiotics in the prevention of necrotizing enterocolitis and mortality: an updated meta-analysis. PLoS One 12:e0171579. doi: 10.1371/journal.pone.0171579
Chen, C., Chen, K., Su, T., Zhang, B., Li, G., Pan, J., et al. (2019). Myo-inositol-1-phosphate synthase (Ino-1) functions as a protection mechanism in Corynebacterium glutamicum under oxidative stress. Microbiol. Open 8:e00721. doi: 10.1002/mbo3.721
Colliou, N., Ge, Y., Sahay, B., Gong, M., Zadeh, M., Owen, J. L., et al. (2017). Commensal Propionibacterium strain UF1 mitigates intestinal inflammation via Th17 cell regulation. J. Clin. Invest. 127, 3970–3986. doi: 10.1172/JCI95376
Corona-Hernandez, R. I., Álvarez-Parrilla, E., Lizardi-Mendoza, J., Islas-Rubio, A. R., de la Rosa, L. A., and Wall-Medrano, A. (2013). Structural stability and viability of microencapsulated probiotic bacteria: a review. Comp. Rev. Food Sci. Food Safety 12, 614–628. doi: 10.1111/1541-4337.12030
Cousin, F. J., Mater, D. D. G., Foligne, B., and Jan, G. (2010). Dairy propionibacteria as human probiotics: a review of recent evidence. Dairy Sci. Technol. 91, 1–26. doi: 10.1051/dst/2010032
Csonka, L. N., and Hanson, A. D. (1991). Prokaryotic osmoregulation: genetics and physiology. Annu. Rev. Microbiol. 45, 569–606. doi: 10.1146/annurev.mi.45.100191.003033
Dalmasso, M., Aubert, J., Briard-Bion, V., Chuat, V., Deutsch, S.-M., Even, S., et al. (2012). A Temporal -omic Study of Propionibacterium freudenreichii CIRM-BIA1T adaptation strategies in conditions mimicking cheese ripening in the cold. PLoS One 7:e29083. doi: 10.1371/journal.pone.0029083
Desmond, C., Stanton, C., Fitzgerald, G. F., Collins, K., and Paul Ross, R. (2001). Environmental adaptation of probiotic lactobacilli towards improvement of performance during spray drying. Int. Dairy J. 11, 801–808. doi: 10.1016/S0958-6946(01)00121-2
Deutsch, S.-M., Mariadassou, M., Nicolas, P., Parayre, S., Le Guellec, R., Chuat, V., et al. (2017). Identification of proteins involved in the anti-inflammatory properties of Propionibacterium freudenreichii by means of a multi-strain study. Sci. Rep. 7:46409. doi: 10.1038/srep46409
Ding, Z.-G., Ji, Y., Yin, M., Zhao, Y.-R., Feng, Y.-Z., Chunyu, W.-X., et al. (2019). Phytoactinopolyspora halophila sp. nov., a halophilic actinomycete isolated from a saline soil. Int. J. Syst. Evol Microbio l 69, 384–389. doi: 10.1099/ijsem.0.003153
Ducrotté, P. (2012). Clinical trial: Lactobacillus plantarum 299v (DSM 9843) improves symptoms of irritable bowel syndrome. World J. Gastroenterol. 18, 4012–4018. doi: 10.3748/wjg.v18.i30.4012
Empadinhas, N., and Viete-Vallejo, O. (2008). Osmoadaptation mechanisms in prokaryotes: distribution of compatible solutes. Int. Microbiol. 1, 151–161. doi: 10.2436/20.1501.01.55
FAO/WHO (2001). Evaluation of Health and Nutritional Properties of Powder Milk and Live Lactic Acid Bacteria. Food and Agriculture Organization of the United Nations and World Health Organization expert Consultation Report. Rome: FAO.
Foligne, B., Deutsch, S.-M., Breton, J., Cousin, F. J., Dewulf, J., Samson, M., et al. (2010). promising immunomodulatory effects of selected strains of dairy propionibacteria as evidenced in vitro and in vivo. Appl. Environ. Microbiol. 76, 8259–8264. doi: 10.1128/AEM.01976-10
Fujimura, K. E., Sitarik, A. R., Havstad, S., Lin, D. L., Levan, S., Fadrosh, D., et al. (2016). Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat. Med. 22, 1187–1191. doi: 10.1038/nm.4176
Ganji-Arjenaki, M., and Rafieian-Kopaei, M. (2018). Probiotics are a good choice in remission of inflammatory bowel diseases: a meta analysis and systematic review. J. Cell. Physiol. 233, 2091–2103. doi: 10.1002/jcp.25911
Gaucher, F., Bonnassie, S., Rabah, H., Leverrier, P., Pottier, S., Jardin, J., et al. (2019a). Benefits and drawbacks of osmotic adjustment in Propionibacterium freudenreichii. J. Proteomics 204:103400. doi: 10.1016/j.jprot.2019.103400
Gaucher, F., Bonnassie, S., Rabah, H., Marchand, P., Blanc, P., Jeantet, R., et al. (2019b). Review: adaptation of beneficial propionibacteria, lactobacilli, and bifidobacteria improves tolerance toward technological and digestive stresses. Front. Microbiol. 10:841. doi: 10.3389/fmicb.2019.00841
Giulio, B. D., Orlando, P., Barba, G., Coppola, R., Rosa, M. D., Sada, A., et al. (2005). Use of alginate and cryo-protective sugars to improve the viability of lactic acid bacteria after freezing and freeze-drying. World J. Microbiol. Biotechnol. 21, 739–746. doi: 10.1007/s11274-004-4735-2
Glaasker, E., Konings, W. N., and Poolman, B. (1996). Osmotic regulation of intracellular solute pools in Lactobacillus plantarum. J. Bacteriol. 178, 575–582.
Goswami, G., Panda, D., Samanta, R., Boro, R. C., Modi, M. K., Bujarbaruah, K. M., et al. (2018). Bacillus megaterium adapts to acid stress condition through a network of genes: insight from a genome-wide transcriptome analysis. Sci. Rep. 8:16105. doi: 10.1038/s41598-018-34221-0
Graham, J. E., and Wilkinson, B. J. (1992). Staphylococcus aureus osmoregulation: roles for choline, glycine betaine, proline, and taurine. J. Bacteriol. 174, 2711–2716. doi: 10.1128/jb.174.8.2711-2716.1992
Guan, H.-L., Zhang, Y.-J., Lu, X.-J., Jia, M., Zhang, Z.-Y., Gao, X.-H., et al. (2018). Gracilibacillus eburneus sp.nov., a moderately halophilic bacterium isolated from Xinjiang province, China. Arch. Microbiol. 200, 423–429. doi: 10.1007/s00203-017-1450-6
Hojo, K., Yoda, N., Tsuchita, H., Ohtsu, T., Seki, K., Taketomo, N., et al. (2002). Effect of ingested culture of Propionibacterium freudenreichii ET-3 on fecal microflora and stool frequency in healthy females. Biosci. Microflora 21, 115–120. doi: 10.12938/bifidus1996.21.115
Huang, S., Cauty, C., Dolivet, A., Le Loir, Y., Chen, X. D., Schuck, P., et al. (2016a). Double use of highly concentrated sweet whey to improve the biomass production and viability of spray-dried probiotic bacteria. J. Funct. Foods 23, 453–463. doi: 10.1016/j.jff.2016.02.050
Huang, S., Rabah, H., Jardin, J., Briard-Bion, V., Parayre, S., Maillard, M.-B., et al. (2016b). Hyperconcentrated sweet whey, a new culture medium that enhances Propionibacterium freudenreichii stress tolerance. Appl. Environ. Microbiol. 82, 4641–4651. doi: 10.1128/AEM.00748-16
Huang, S., Gaucher, F., Cauty, C., Jardin, J., Le Loir, Y., Jeantet, R., et al. (2018). Growth in hyper-concentrated sweet whey triggers multi stress tolerance and spray drying survival in Lactobacillus casei BL23: from the molecular basis to new perspectives for sustainable probiotic production. Front. Microbiol. 9:2548. doi: 10.3389/fmicb.2018.02548
Huang, S., Vignolles, M.-L., Chen, X. D., Le Loir, Y., Jan, G., Schuck, P., et al. (2017). Spray drying of probiotics and other food-grade bacteria: a review. Trends Food Sci. Tech. 63, 1–17. doi: 10.1016/j.tifs.2017.02.007
Isawa, K., Hojo, K., Yoda, N., Kamiyama, T., Makino, S., Saito, M., et al. (2002). Isolation and identification of a new bifidogenic growth stimulator produced by Propionibacterium freudenreichii ET-3. Biosci. Biotechnol. Biochem. 66, 679–681. doi: 10.1271/bbb.66.679
Jan, G., Leverrier, P., Pichereau, V., and Boyaval, P. (2001). Changes in protein synthesis and morphology during acid adaptation of Propionibacterium freudenreichii. Appl. Environ. Microbiol. 67, 2029–2036. doi: 10.1128/AEM.67.5.2029-2036.2001
Jan, G., Leverrier, P., Proudy, I., and Roland, N. (2002). Survival and beneficial effects of propionibacteria in the human gut: in vivo and in vitro investigations. Le Lait 82, 131–144. doi: 10.1051/lait:2001012
Jan, G., Rouault, A., and Maubois, J.-L. (2000). Acid stress susceptibility and acid adaptation of Propionibacterium freudenreichii subsp. shermanii. Int. Dairy J. 80, 325–336. doi: 10.1051/lait:2000128
Jang, J.-Y., Oh, Y. J., Lim, S. K., Park, H. K., Lee, C., Kim, J. Y., et al. (2018). Salicibibacter kimchii gen. nov., sp. nov., a moderately halophilic and alkalitolerant bacterium in the family Bacillaceae, isolated from kimchi. J. Microbiol. 56, 880–885. doi: 10.1007/s12275-018-8518-0
Jardin, J. (2019). Data from Propionibacterium freudenreichii CIRM-BIA 129 osmoadaptation coupled to acid-adaptation increases its viability during freeze-drying. Portail Data Inra 1. doi: 10.15454/5cwcll
Kallio, S., Kukkonen, A. K., Savilahti, E., and Kuitunen, M. (2018). Perinatal probiotic intervention prevented allergic disease in a Caesarean-delivered subgroup at 13-year follow-up. Clin. Exp. Allergy 49, 506–515. doi: 10.1111/cea.13321
Kanemasa, Y., Yoshioka, T., and Hayashi, H. (1972). Alteration of the phospholipid composition of Staphylococcus aureus cultured in medium containing NaCl. Biochim. Biophys. Acta 280, 444–450.
Kets, E., Teunissen, P., and de Bont, J. (1996). Effect of compatible solutes on survival of lactic Acid bacteria subjected to drying. Appl. Environ. Microbiol. 62, 259–261.
Koch, S., Eugster-Meier, E., Oberson, G., Meile, L., and Lacroix, C. (2008). Effects of strains and growth conditions on autolytic activity and survival to freezing and lyophilization of Lactobacillus delbrueckii ssp. lactis isolated from cheese. Int. Dairy J. 18, 187–196. doi: 10.1016/j.idairyj.2007.07.009
Komaratat, P., and Kates, M. (1975). The lipid composition of a halotolerant species of staphylococcus epidermidis. Biochim. Biophys. Acta 398, 464–484. doi: 10.1016/0005-2760(75)90197-6
Kunin, C. M., and Rudy, J. (1991). Effect of NaCl-induced osmotic stress on intracellular concentrations of glycine betaine and potasssium in Escherichia coli, Enterococcus faecalis, and Staphylococci. Transl. Res. 118, 217–224. doi: 10.5555/uri:pii:002221439190065F
Lanciotti, R., Guerzoni, M. E., and Cocconcelli, P. S. (2001). Alteration in cellular fatty acid composition as a response to salt, acid, oxidative and thermal stresses in Lactobacillus helveticus. Microbiology 147, 2255–2264. doi: 10.1099/00221287-147-8-2255
Langella, O., Valot, B., Balliau, T., Blein-Nicolas, M., Bonhomme, L., and Zivy, M. (2017). X!TandemPipeline: a tool to manage sequence redundancy for protein inference and phosphosite identification. J. Proteome Res. 16, 494–503. doi: 10.1021/acs.jproteome.6b00632
Leverrier, P., Dimova, D., Pichereau, V., Auffray, Y., Boyaval, P., and Jan, G. (2003). Susceptibility and adaptive response to bile salts in Propionibacterium freudenreichii: physiological and proteomic analysis. Appl. Environ. Microbiol. 69, 3809–3818. doi: 10.1128/AEM.69.7.3809-3818.2003
Leverrier, P., Vissers, J. P. C., Rouault, A., Boyaval, P., and Jan, G. (2004). Mass spectrometry proteomic analysis of stress adaptation reveals both common and distinct response pathways in Propionibacterium freudenreichii. Arch. Microbiol. 181, 215–230. doi: 10.1007/s00203-003-0646-0
Li, C., Zhao, J.-L., Wang, Y.-T., Han, X., and Liu, N. (2009). Synthesis of cyclopropane fatty acid and its effect on freeze-drying survival of Lactobacillus bulgaricus L2 at different growth conditions. World J. Microbiol. Biotechnol. 25, 1659–1665. doi: 10.1007/s11274-009-0060-0
Machado, M. C., López, C. S., Heras, H., and Rivas, E. A. (2004). Osmotic response in Lactobacillus casei ATCC 393: biochemical and biophysical characteristics of membrane. Arch. Bioch. Biophys. 422, 61–70. doi: 10.1016/j.abb.2003.11.001
Malik, A. C., Reinbold, G. W., and Vedamuthu, E. R. (1968). An evaluation of the taxonomy of Propionibacterium. Can. J. Microbiol. 14, 1185–1191.
Miller, K. J. (1985). Effects of temperature and sodium chloride concentration on the phospholipid and fatty acid compositions of a halotolerant Planococcus sp. J. Bacteriol. 162, 263–270.
Mitsuyama, K., Masuda, J., Yamasaki, H., Kuwaki, K., Kitazaki, S., Koga, H., et al. (2007). Treatment of ulcerative colitis with milk whey culture with Propionibacterium freudenreichii. J. Intest Microbiol. 21, 143–147.
Moco, S., Candela, M., Chuang, E., Draper, C., Cominetti, O., Montoliu, I., et al. (2014). systems biology approaches for inflammatory bowel disease: emphasis on gut microbial metabolism. Inflamm. Bowel Dis. 20, 2104–2114. doi: 10.1097/MIB.0000000000000116
Moore, S. D., and Sauer, R. T. (2007). The tmRNA system for translational surveillance and ribosome rescue. Annu. Rev. Biochem. 76, 101–124. doi: 10.1146/annurev.biochem.75.103004.142733
Nurlinawati, K., Aertsen, A., and Michiels, C. W. (2015). Role of 1-acyl-sn-glycerol-3-phosphate acyltransferase in psychrotrophy and stress tolerance of Serratia plymuthica RVH1. Res. Microbiol. 166, 28–37. doi: 10.1016/j.resmic.2014.11.001
Okada, Y. (2006). Propionibacterium freudenreichii component 1.4-dihydroxy-2-naphthoic acid (DHNA) attenuates dextran sodium sulphate induced colitis by modulation of bacterial flora and lymphocyte homing. Gut 55, 681–688. doi: 10.1136/gut.2005.070490
Paéz, R., Lavari, L., Vinderola, G., Audero, G., Cuatrin, A., Zaritzky, N., et al. (2012). Effect of heat treatment and spray drying on lactobacilli viability and resistance to simulated gastrointestinal digestion. Food Res. Int. 48, 748–754. doi: 10.1016/j.foodres.2012.06.018
Palmfeldt, J., and Hahn-Hägerdal, B. (2000). Influence of culture pH on survival of Lactobacillus reuteri subjected to freeze-drying. Int. J. Food Microbiol. 55, 235–238. doi: 10.1016/S0168-1605(00)00176-8
Palomino, M. M., Waehner, P. M., Fina Martin, J., Ojeda, P., Malone, L., Sánchez Rivas, C., et al. (2016). Influence of osmotic stress on the profile and gene expression of surface layer proteins in Lactobacillus acidophilus ATCC 4356. Appl. Microbiol. Biotechnol. 100, 8475–8484. doi: 10.1007/s00253-016-7698-y
Plé, C., Breton, J., Richoux, R., Nurdin, M., Deutsch, S.-M., Falentin, H., et al. (2016). Combining selected immunomodulatory Propionibacterium freudenreichii and Lactobacillus delbrueckii strains: reverse engineering development of an anti-inflammatory cheese. Mol. Nutr. Food Res. 60, 935–948. doi: 10.1002/mnfr.201500580
Plé, C., Richoux, R., Jardin, J., Nurdin, M., Briard-Bion, V., Parayre, S., et al. (2015). Single-strain starter experimental cheese reveals anti-inflammatory effect of Propionibacterium freudenreichii CIRM BIA 129 in TNBS-colitis model. J. Funct. Foods 18, 575–585. doi: 10.1016/j.jff.2015.08.015
Rabah, H., Ménard, O., Gaucher, F., do Carmo, F. L. R., Dupont, D., and Jan, G. (2018). Cheese matrix protects the immunomodulatory surface protein SlpB of Propionibacterium freudenreichii during in vitro digestion. Food Res. Int. 106, 712–721. doi: 10.1016/j.foodres.2018.01.035
Rabah, H., Rosa do Carmo, F., and Jan, G. (2017). Dairy Propionibacteria: versatile probiotics. Microorganisms 5:24. doi: 10.3390/microorganisms5020024
Romeo, Y., Obis, D., Bouvier, J., Guillot, A., Fourçans, A., Bouvier, I., et al. (2003). Osmoregulation in Lactococcus lactis: BusR, a transcriptional repressor of the glycine betaine uptake system BusA. Mol. Microbiol. 47, 1135–1147.
Sanchez, B., Champomier-Verges, M.-C., Collado, M. D. C., Anglade, P., Baraige, F., Sanz, Y., et al. (2007). Low-pH adaptation and the acid tolerance response of Bifidobacterium longum biotype longum. Appl. Environ. Microbiol. 73, 6450–6459. doi: 10.1128/AEM.00886-7
Santivarangkna, C., Kulozik, U., and Foerst, P. (2007). Alternative drying processes for the industrial preservation of lactic acid starter cultures. Biotechnol. Progr. 23, 302–315. doi: 10.1021/bp060268f
Savijoki, K., Suokko, A., Palva, A., Valmu, L., Kalkkinen, N., and Varmanen, P. (2005). Effect of heat-shock and bile salts on protein synthesis of Bifidobacterium longum revealed by [35S]methionine labelling and two-dimensional gel electrophoresis. FEMS Microbiol. Lett. 248, 207–215. doi: 10.1016/j.femsle.2005.05.032
Seki, K., Nakao, H., Umino, H., Isshiki, H., Yoda, N., and Tachihara, R. (2004). Effects of fermented milk whey containing novel bifidogenic growth stimulator produced by Propionibacterium on fecal bacteria, putrefactive metabolite, defecation frequency and fecal properties in senile volunteers needed serious nursing-care taking enteral nutrition by tube feeding. J. Intstinal Microbiol. 18, 107–115.
Serata, M., Kiwaki, M., and Iino, T. (2016). Functional analysis of a novel hydrogen peroxide resistance gene in Lactobacillus casei strain Shirota. Microbiology 162, 1885–1894. doi: 10.1099/mic.0.000379
Silva, J., Carvalho, A. S., Ferreira, R., Vitorino, R., Amado, F., Domingues, P., et al. (2005). Effect of the pH of growth on the survival of Lactobacillus delbrueckii subsp. bulgaricus to stress conditions during spray-drying. J. Appl. Microbiol. 98, 775–782. doi: 10.1111/j.1365-2672.2004.02516.x
Skonieczna-Żydecka, K., Marlicz, W., Misera, A., Koulaouzidis, A., and Żoniewski, I. (2018). Microbiome—the missing link in the gut-brain axis: focus on its role in gastrointestinal and mental health. J. Clin. Med. 7:521. doi: 10.3390/jcm7120521
Sniffen, J. C., McFarland, L. V., Evans, C. T., and Goldstein, E. J. C. (2018). Choosing an appropriate probiotic product for your patient: an evidence-based practical guide. PLoS One 13:e0209205. doi: 10.1371/journal.pone.0209205
Sonck, K. A. J., Kint, G., Schoofs, G., Vander Wauven, C., Vanderleyden, J., and De Keersmaecker, S. C. J. (2009). The proteome of Salmonella typhimurium grown under in vivo-mimicking conditions. Proteomics 9, 565–579. doi: 10.1002/pmic.200700476
Soularue, E., Lepage, P., Colombel, J. F., Coutzac, C., Faleck, D., Marthey, L., et al. (2018). Enterocolitis due to immune checkpoint inhibitors: a systematic review. Gut 67, 2056–2067. doi: 10.1136/gutjnl-2018-316948
Streit, F., Delettre, J., Corrieu, G., and Béal, C. (2008). Acid adaptation of Lactobacillus delbrueckii subsp. bulgaricus induces physiological responses at membrane and cytosolic levels that improves cryotolerance. J. Appl. Microbiol. 105, 1071–1080. doi: 10.1111/j.1365-2672.2008.03848.x
Thierry, A., Deutsch, S.-M., Falentin, H., Dalmasso, M., Cousin, F. J., and Jan, G. (2011). New insights into physiology and metabolism of Propionibacterium freudenreichii. Int. J. Food Microbiol. 149, 19–27. doi: 10.1016/j.ijfoodmicro.2011.04.026
Valot, B., Langella, O., Nano, E., and Zivy, M. (2011). MassChroQ: a versatile tool for mass spectrometry quantification. Proteomics 11, 3572–3577. doi: 10.1002/pmic.201100120
Keywords: bacteria adaptation, freeze-drying, probiotic, starters, osmoadaptation, stress
Citation: Gaucher F, Kponouglo K, Rabah H, Bonnassie S, Ossemond J, Pottier S, Jardin J, Briard-Bion V, Marchand P, Blanc P, Jeantet R and Jan G (2019) Propionibacterium freudenreichii CIRM-BIA 129 Osmoadaptation Coupled to Acid-Adaptation Increases Its Viability During Freeze-Drying. Front. Microbiol. 10:2324. doi: 10.3389/fmicb.2019.02324
Received: 16 July 2019; Accepted: 23 September 2019;
Published: 09 October 2019.
Edited by:
Eugenia Bezirtzoglou, Democritus University of Thrace, GreeceReviewed by:
Erhard Bremer, University of Marburg, GermanyVincent Peton, University of Greifswald, Germany
Jorge Reinheimer, National University of the Littoral, Argentina
Copyright © 2019 Gaucher, Kponouglo, Rabah, Bonnassie, Ossemond, Pottier, Jardin, Briard-Bion, Marchand, Blanc, Jeantet and Jan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gwénaël Jan, Z3dlbmFlbC5qYW5AaW5yYS5mcg==
†These authors share senior authorship