 Mónica Gandía
Mónica Gandía Sandra Garrigues
Sandra Garrigues Begoña Bolós
Begoña Bolós Paloma Manzanares
Paloma Manzanares Jose F. Marcos
Jose F. Marcos- Department of Biotechnology, Instituto de Agroquímica y Tecnología de Alimentos, Consejo Superior de Investigaciones Científicas, Valencia, Spain
Penicillium digitatum is the main postharvest pathogen of citrus fruit and is responsible for important economic losses in spite of the massive use of fungicides. The fungal cell wall (CW) and its specific component chitin are potential targets for the development of new antifungal molecules. Among these are the antifungal peptides and proteins that specifically interact with fungal CW. Chitin is synthesized by a complex family of chitin synthases (Chs), classified into up to eight classes within three divisions. Previously, we obtained and characterized a mutant of P. digitatum in the class VII gene (ΔchsVII), which contains a short myosin motor-like domain (MMD). In this report, we extend our previous studies to the characterization of mutants in chsII and in the gene coding for the other MMD-Chs (chsV), and study the role of chitin synthases in the sensitivity of P. digitatum to the self-antifungal protein AfpB, and to AfpA obtained from P. expansum. The ΔchsII mutant showed no significant phenotypic and virulence differences with the wild type strain, except in the production and morphology of the conidia. In contrast, mutants in chsV showed a more dramatic phenotype than the previous ΔchsVII, with reduced growth and conidial production, increased chitin content, changes in mycelial morphology and a decrease in virulence to citrus fruit. Mutants in chsVII were specifically more tolerant than the wild type to nikkomycin Z, an antifungal inhibitor of chitin biosynthesis. Treatment of P. digitatum with its own antifungal protein AfpB resulted in an overall reduction in the expression of the chitin synthase genes. The mutants corresponding to MMD chitin synthases exhibited differential sensitivity to the antifungal proteins AfpA and AfpB, ΔchsVII being more susceptible than its parental strain and ΔchsV being slightly more tolerant despite its reduced growth in liquid broth. Taking these results together, we conclude that the MMD-containing chitin synthases affect cell wall integrity and sensitivity to antifungal proteins in P. digitatum.
Introduction
Fungi are responsible for serious diseases in plants, animals and humans, resulting in a large number of deaths and millionaire economic losses (Brown et al., 2012; Fisher et al., 2012). Effective control of phytopathogenic fungi is a major challenge due to the huge economic losses they cause, the emergence of resistance against the few classes of currently available fungicides and the cross-resistance between clinic and crop pathogens (Perfect, 2016; Fisher et al., 2018). In this context, the study of antifungal peptides and proteins has attracted much attention as promising biomolecules to control deleterious fungi (Marcos et al., 2008; Galgóczy and Marx, 2019).
The fungal cell wall (CW) is a promising target for the development of new antifungal molecules because it has a structure and composition not shared by animal or plant cells, and therefore is specific to fungi (Latgé, 2007; Lee and Sheppard, 2016). The CW is also essential as it protects the cell and interacts firstly with the host during infection and colonization, being considered important in the host-pathogen interaction (Kombrink et al., 2011; Sánchez-Vallet et al., 2014; Lee and Sheppard, 2016; Geoghegan et al., 2017; Latgé et al., 2017). Among the unique structure of the fungal CW stands out chitin, a linear homopolymer of β-(1,4)-N-acetylglucosamine neither present in plants nor higher animals, necessary for fungal survival and considered as a pathogen-associated molecular pattern (PAMP) (Kombrink et al., 2011; Thomma et al., 2011). Chitin and chitin synthesis are, therefore, targets to develop new antifungal drugs. Chitin synthesis is a complex process carried out by a repertoire of conserved enzymes known as chitin synthases (Chs) encoded by specific genes (chs). Three chs genes have been reported in the yeast Saccharomyces cerevisiae, whereas many more have been identified in filamentous fungi, as it is the case of the Blastocladiomycete Allomyces macrogynus or the Zygomycete Rhizopus chinensis with 38 and 44 different chs, respectively (Gonçalves et al., 2016; Li et al., 2016). These Chs are classified into three divisions and up to eight classes: division I contains class I, II and III chs genes; division II contains class IV, V and VII and finally division III contains class VI (Latgé, 2007; Sheng et al., 2013; Gandía et al., 2014; Li et al., 2016; Liu et al., 2016; Roncero et al., 2016). A new Chs class (VIII) in division I was defined recently (Li et al., 2016). Particularly significant are ChsV and ChsVII, which are two chitin synthases with a chimeric structure containing an N-terminal myosin motor-like domain (MMD) fused to a CHS domain, and which have been associated with hyphal growth and virulence in many filamentous fungi (Takeshita et al., 2006; Kim et al., 2009; Larson et al., 2011; Fernandes et al., 2016). Numerous studies on the biological role of the distinct chs genes in fungal pathogens have been conducted, obtaining a repertoire of deletion mutants of these genes in different filamentous fungi such as Aspergillus fumigatus (Muszkieta et al., 2014), Fusarium graminearum (Cheng et al., 2015), Magnaporthe oryzae (Kong et al., 2012) or Botrytis cinerea (Morcx et al., 2013). Overall, these studies have shown specific roles for each class/division, including the response to CW stress and involvement in virulence.
Penicillium digitatum is the main postharvest pathogen of citrus fruit and causes the green mold disease, being responsible for significant economic losses in citriculture (Marcet-Houben et al., 2012; Palou, 2014; Costa et al., 2019). We have previously identified and analyzed the expression of chitin synthase genes in P. digitatum (Gandía et al., 2012, 2014). Gene expression analyses concluded that chsIII is the most expressed and chsII is the least expressed of all the chitin synthase genes; chsI, chsIII, chsV, and chsVII were induced during fruit infection; and chsII was the most upregulated gene during axenic growth in solid PDA medium, coincident with conidia production (Gandía et al., 2012, 2014). Moreover, our group disrupted chsVII in P. digitatum and showed the importance of this gene for fungal growth, conidia production, virulence, CW integrity and mycelium development during fruit infection (Gandía et al., 2014). In addition, this ΔchsVII mutant registered an impressive compensatory induction of chsII expression and, to a lesser extent, of chsV, compared with the parental wild type, during axenic growth and in response to specific stresses such as osmotic shock caused by sorbitol.
Antifungal proteins (AFPs) produced by filamentous fungi from the class Eurotiomycetes hold great promise for the control of fungal pathogens. AFPs are small, cationic, cysteine-rich proteins that are extraordinarily stable and secreted in high amounts by producing fungal strains (Meyer, 2008; Hegedüs and Marx, 2013; Galgóczy and Marx, 2019). The two most studied AFPs are AFP from Aspergillus giganteus (Meyer, 2008) and PAF from Penicillium chrysogenum (Marx et al., 2008). Studies in our group have recently identified, produced and characterized two highly active AFPs from postharvest pathogens: AfpB from P. digitatum (Garrigues et al., 2016, 2017) and AfpA from Penicillium expansum (Garrigues et al., 2018), which are both active even against the self-fungi. Moreover, both AFPs can control experimental infections of P. digitatum and B. cinerea on citrus fruit and tomato plants, respectively (Garrigues et al., 2018; Shi et al., 2019). We have initiated the study of the mode of action of these AFPs, showing that AfpB induces the phosphorylation of mitogen-activated protein kinases (MAPKs), including Slt2 that senses CW damage and regulates the CW integrity pathway (Gandía et al., 2019), establishing a connection between CW, CW stress and the mechanism of AFPs.
In this report, we decided to expand the number of available null mutants of Chs in P. digitatum by characterizing mutants of ChsII and ChsV, which are relevant according to our previous gene expression studies, and to explore the role of chitin synthases in the mode of action of AFPs. Herein, we present data on chs gene expression in response to the treatment of P. digitatum with the self-AfpB. While the deletion of chsII did not seem to have major effects on the biology of the fungus beyond the production and morphology of conidia, the elimination of chsV generated mutant strains with severe alterations in growth and morphology, chitin content, reduction of conidia production and altered virulence. Interestingly, the two MMD-Chs mutants (ΔchsV and ΔchsVII) had differential phenotypes and showed altered sensitivity to both antifungal proteins, AfpA and AfpB.
Materials and Methods
Fungal Strains, Media and Culture Conditions
The P. digitatum parental strains used were the wild type CECT 20796 (Marcet-Houben et al., 2012) and the transformant PDMG314 (Δku70:nat1) (Gandía et al., 2016). All P. digitatum strains were routinely cultured on potato dextrose agar (PDA, Difco, Detroit, MI, United States) plates for 7–10 days at 24°C. Spores were collected, filtered, quantified with a hemocytometer, and adjusted to the appropriate concentration. Growth of strains and conidia production were evaluated by depositing 5 μL of a conidial suspension (5 × 104 conidia/mL) on PDA plates, and the growth diameter was measured daily. Conidia production was determined as previously described (Gandía et al., 2014). Statistical analyses were conducted using the statistical SPSS 22.0 package (SPSS Inc., Chicago, IL, United States) to calculate the ANOVA test (p < 0.05). The Agrobacterium tumefaciens AGL-1 was used for fungal transformation (Gandía et al., 2014).
Construction of Deletion Vectors and Fungal Transformation
To carry out the deletion of chsII and chsV, the double selection strategy (positive selection in hygromycin and negative in 5-fluoro-2’-deoxyuridine, F2dU) previously used in P. digitatum was performed (Gandía et al., 2014, 2016; Harries et al., 2015; Garrigues et al., 2016). Gene deletion constructs were obtained with the primers detailed in Supplementary Table S1. All PCR procedures were carried out with AccuPrime Taq DNA Polymerase, high fidelity (Invitrogen, Eugene, OR, United States), and the resulting DNA constructs were verified by DNA sequencing. Briefly, the hygromycin resistant cassette (hph) used as a positive selection marker, was amplified from pBHt2 (Khang et al., 2006) through PCR using the primers OJM197/OJM198. In the case of chsII, 1089 bp 5′ and 999 bp 3′ flanking regions were amplified from P. digitatum CECT 20796 genomic DNA with the primer pairs OJM408/OJM409 and OJM410/OJM411, respectively (Supplementary Figure S1). In the case of chsV, 1005 bp 5′ and 1026 bp 3′ flanking regions were amplified with the primer pairs OJM414/OJM415 and OJM416/OJM417, respectively (Supplementary Figure S2). Constructs obtained by Fusion PCR (Szewczyk et al., 2006) were cloned into pGKO2, and the obtained vectors pGKO2_ΔchsII and pGKO2_ΔchsV were transformed into A. tumefaciens AGL-1 by electroporation. CECT 20796 and PDMG314 (Δku70) strains were transformed by A. tumefaciens-mediated transformation (ATMT) as previously described (Khang et al., 2006) with minor modifications (Harries et al., 2015). Monosporic transformants were obtained through two rounds of selection on hygromycin-containing PDA plates and confirmed by PCR amplification of genomic DNA (Khang et al., 2006; Harries et al., 2015). The primers used were located at different positions around the target locus to discriminate and confirm the transformation events (Supplementary Table S1 and Supplementary Figures S1, S2). Ectopic strains, in which the transformed DNA was randomly inserted in a region different from the targeted locus, were recovered from each transformation experiment. These ectopic strains (PDMG2010 and PDMG612) were used in our experiments as additional controls.
Phenotypic Characterization and Sensitivity Assays to Chemicals and Temperature
We tested the sensitivity of fungal strains to different compounds in 24-well cell culture plates (1 mL PDA per well). PDA medium was supplemented with 1.2 M sorbitol, 0.5 M NaCl, 150 μg/mL calcofluor white (CFW) (Fluorescent Brightener 28, Sigma-Aldrich, F3543), 150 μg/mL sodium dodecyl sulphate (SDS) (Sigma-Aldrich, L4509), 2 mM H2O2 and nikkomycin Z (NZ) (16, 64 and 128 μg/mL) (Sigma-Aldrich, N8028). Five μL of serial tenfold dilution of conidia (106 to 103 conidia/mL) were applied into each well. Plates were incubated at 24°C for 3–5 days. To test sensitivity to different temperatures, the different strains were grown on standard PDA plates at 24, 26, and 28°C for 7 days.
Chitin Content
The determination of chitin content of fungal mycelia was based on methods described previously (Din et al., 1996; Gandía et al., 2014). Briefly, PDB cultures were inoculated with 2 × 105 conidia/mL of P. digitatum strains and grown at 24°C for 5 days. Digestion was carried out with 0.2 U of chitinase suspension (Sigma-Aldrich, C6137) and 2 U of Zymolyase 20T (Euromedex, UZ1000). Data are the mean of four replicates and significance was determined by the statistical SPSS 22.0 package to calculate the ANOVA test (p < 0.05). The experiment was repeated twice with similar results.
Microscopy Visualization
Each fungal strain was prepared, stained with CFW and microscopically visualized with a Nikon E90i fluorescence microscope as previously described (Garrigues et al., 2016). Bright field (BF) and fluorescence images were captured by the NIS-Elements BR v2.3 software (Nikon) and processed using FIJI software (Schindelin et al., 2012). To obtain the differential interference contrast (DIC) images of conidia, 100× objective was used and the images were captured, processed and analyzed by the NIS-Elements BR v2.3 software (Nikon). The Feret diameter (F, defined as the measure of an object size along a specified direction, or the distance between the two parallel planes restricting the object perpendicular to that direction) and the circularity (perimeter of the object) were measured to determine conidia shape. Statistical analyses were conducted using the statistical SPSS 22.0 package to calculate the ANOVA test (p < 0.05).
Antifungal Assays
Growth inhibition assays were performed in 96-well microtiter plates (Nunc, Roskilde, Denmark) in a total volume of 100 μL as described previously (Garrigues et al., 2017) with modifications. Fifty μL of 2x conidia solution (5 × 104 conidia/mL) in 1/10 diluted PDB medium containing 0.02% (w/v) chloramphenicol was mixed in each well with 50 μL of 2x concentrated protein solution (AfpA from P. expansum or AfpB from P. digitatum) from serial twofold dilutions (from 0.25 to 8 μg/mL for AfpA, or from 0.5 to 16 μg/mL for AfpB). All samples were prepared in triplicate and the mean and standard deviation (SD) were calculated. Plates were statically incubated for 4 days at 25°C, and growth was determined daily by measuring the optical density (OD) at 600 nm (OD600) using a FLUOstar Omega plate spectrophotometer (BMG labtech, Orlenberg, Germany). Dose-response curves were generated from measurements after 48 h. These experiments were repeated at least twice. The Minimum Inhibitory Concentration (MIC) is the peptide concentration that completely inhibited growth after 96 h of incubation, in all the experiments performed.
Fruit Infection Assays
Fruit infection assays with different P. digitatum strains on mature freshly harvested orange fruits (Citrus sinensis L. Osbeck cv Navelina) were conducted as previously described (González-Candelas et al., 2010). Three replicates of five fruits were inoculated with 5 μL of different conidial suspensions (104 and 106 conidia/mL) at four wounds around the equator. Each inoculated wound was scored for green mold infection symptoms and data presented are the mean value ± SD of the percentage of infected wounds per replica at different days post-inoculation (dpi).
Results
Deletion of chsII and chsV in P. digitatum
We previously described and characterized seven chitin synthase genes in the P. digitatum genome (Gandía et al., 2012, 2014). A recent re-evaluation showed that P. digitatum also has a duplication of chsIII that would code for a second class III enzyme (ChsIIIb, PDIG_76160), as occurs with other fungi (Mellado et al., 1996; Choquer et al., 2004). In a previous work, we disrupted the MMD-containing class VII chsVII gene to obtain knockout mutants and demonstrate its involvement in CW integrity and virulence (Gandía et al., 2014). In the present study, and following the same approach, we obtained deletion mutants of another two chs genes to identify their functional role. First, we selected chsV that codes for the other MMD-Chs because of its increased expression levels during fungal infection. Additionally, we chose chsII because it is one of the most affected genes by the disruption of chsVII and due to its possible role in conidiogenesis, as it had been postulated for other class II chs genes (Gandía et al., 2012; Sheng et al., 2013).
To obtain deletion mutants, a fragment of each gene was replaced by the hph cassette used as positive selection marker (Supplementary Figures S1A, S2A). Two different strains were used as parental: the wild type CECT 20796, and PDMG314 (Δku70) obtained to increase the homologous recombination (HR) frequency in P. digitatum by the inactivation of the Ku70/80 heterodimeric complex involved in the non-homologous end joining (NHEJ) DNA repair pathway (Gandía et al., 2016) (Figure 1A). The mutants ΔchsII and ΔchsV were mentioned in our previous publication to illustrate that a higher number of deletion strains were obtained when using PDMG314 as parental strain than with CECT 20796, demonstrating the increase of HR frequency (Gandía et al., 2016). Deletion mutants of chsII were easily obtained from both parental strains, and six of them (two obtained from the wild type, identified as PDMG905 and PDMG908 and four obtained from the PDMG314 strain, numbered between PDMG917 and PDMG926) were confirmed by a complete set of PCR amplifications with specific primers (Supplementary Table S1 and Supplementary Figure S1B). In the case of chsV, only two deletion transformants were obtained with PDMG314 as background and none with CECT 20796. The two deletion strains (PDMG31424 and PDMG31441) were also confirmed by PCR amplifications (Supplementary Figure S2B).
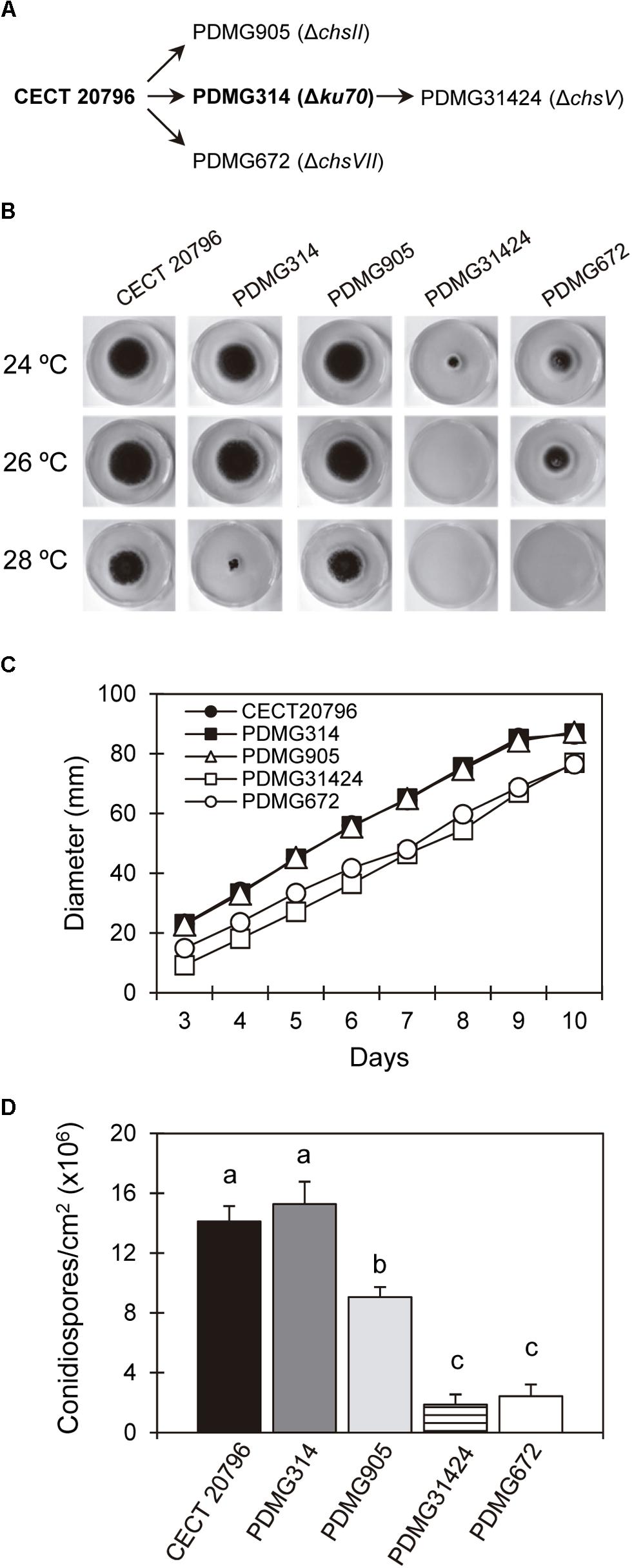
Figure 1. Growth and conidia production of different P. digitatum strains. (A) Schematic overview of the origin of different mutants, indicating the parental strains from which they come. (B) Radial growth at 7 days on PDA plates of the parental strains CECT 20796 and PDMG314 and deletion mutants ΔchsII (PDMG905); ΔchsV (PDMG31424) and ΔchsVII (PDMG672) at 24, 26, and 28°C. Note the absence of growth in PDMG31424 at 26 and 28°C and no growth of PDMG672 only at 28°C. (C) Colony diameter of different strains from 3 to 10 days of growth at 24°C in PDA. (D) Conidia production per surface area of different strains after 5 days of growth at 24°C on PDA plates. Data are mean values ± standard deviation (SD) of three replicate samples. Letters show significant differences among the strains within each day (ANOVA test, p < 0.05).
In the assays described below, we used independent mutants of each gene and found similar behavior in all of them, even among the chsII mutants obtained from two different backgrounds (data not shown). For simplicity, only one representative deletion strain of each gene is shown in most of the figures below: PDMG905 corresponding to ΔchsII, and PDMG31424 to ΔchsV. The functional complementation of these strains was not achieved: (i) in the ΔchsII mutant because it did not present major phenotypic differences with the wild type strain, (ii) in the ΔchsV, because –similar to the previous ΔchsVII (Gandía et al., 2014)– the CW and conidia production are so affected that impeded our attempts to obtain complemented strains.
Phenotypic Characterization of Mutants
Deletion of many chs genes had, as a result, alterations in CW, growth, and sensitivity to temperature or in response to different stresses in different species as Yarrowia lipolytica, P. digitatum or A. fumigatus (Sheng et al., 2013; Gandía et al., 2014; Muszkieta et al., 2014). The growth and conidia production of deletion strains PDMG905 (ΔchsII) and PDMG31424 (ΔchsV) was compared with their respective parental strains CECT 20796 and PDMG314, and with the previously analyzed PDMG672 (ΔchsVII) as well (Figure 1). PDMG905 did not present differences in growth compared to the wild type parental (Figures 1B,C). Only PDMG31424 (ΔchsV) showed a strong reduction of growth at 24°C compared to the parental PDMG314, even more reduced than ΔchsVII strain (PDMG672).
Penicillium digitatum is very sensitive to temperature and cannot grow at 30°C or above. Figure 1B shows the sensitivity of the strains at the increasing temperatures of 24 (the usual in our laboratory), 26 and 28°C. When all strains where incubated at 28°C, a slight reduction of growth was similarly detected in the parental CECT 20796 and PDMG905 (ΔchsII). PDMG314 was clearly affected at 28°C, while PDMG672 and PDMG31424 were unable to growth at this temperature. At 26°C PDMG31424 did not grow either, contrarily to all the four other strains whose growth at 26°C was similar to that at 24°C (Figure 1B). We concluded that the deletion of chsV affected the temperature sensitivity of P. digitatum more than deletion of chsVII.
We next measured the production of conidia in the different strains (Figure 1D). Results revealed a significant but moderate reduction in conidia production of PDMG905 (ΔchsII), supporting the possible involvement of this gene in conidiogenesis. The mutants in the two MMD-Chs showed a strong and similar reduction of conidia production when the number of conidia was normalized to the surface of the colony (Figure 1D) (Gandía et al., 2014).
Sensitivity of Mutants to Chemicals
To evaluate the sensitivity to chemicals of our mutants, we conducted growth assays in presence of different compounds. The osmotic stabilizer sorbitol recovered the growth of the deletion strains in chsV and chsVII (Supplementary Figure S3), as described previously in ΔchsVII mutants (Gandía et al., 2014). However, during osmotic stress caused by sodium salt (NaCl) only deletion mutant in ΔchsV was able to grow better than in PDA medium (Figure 2A).
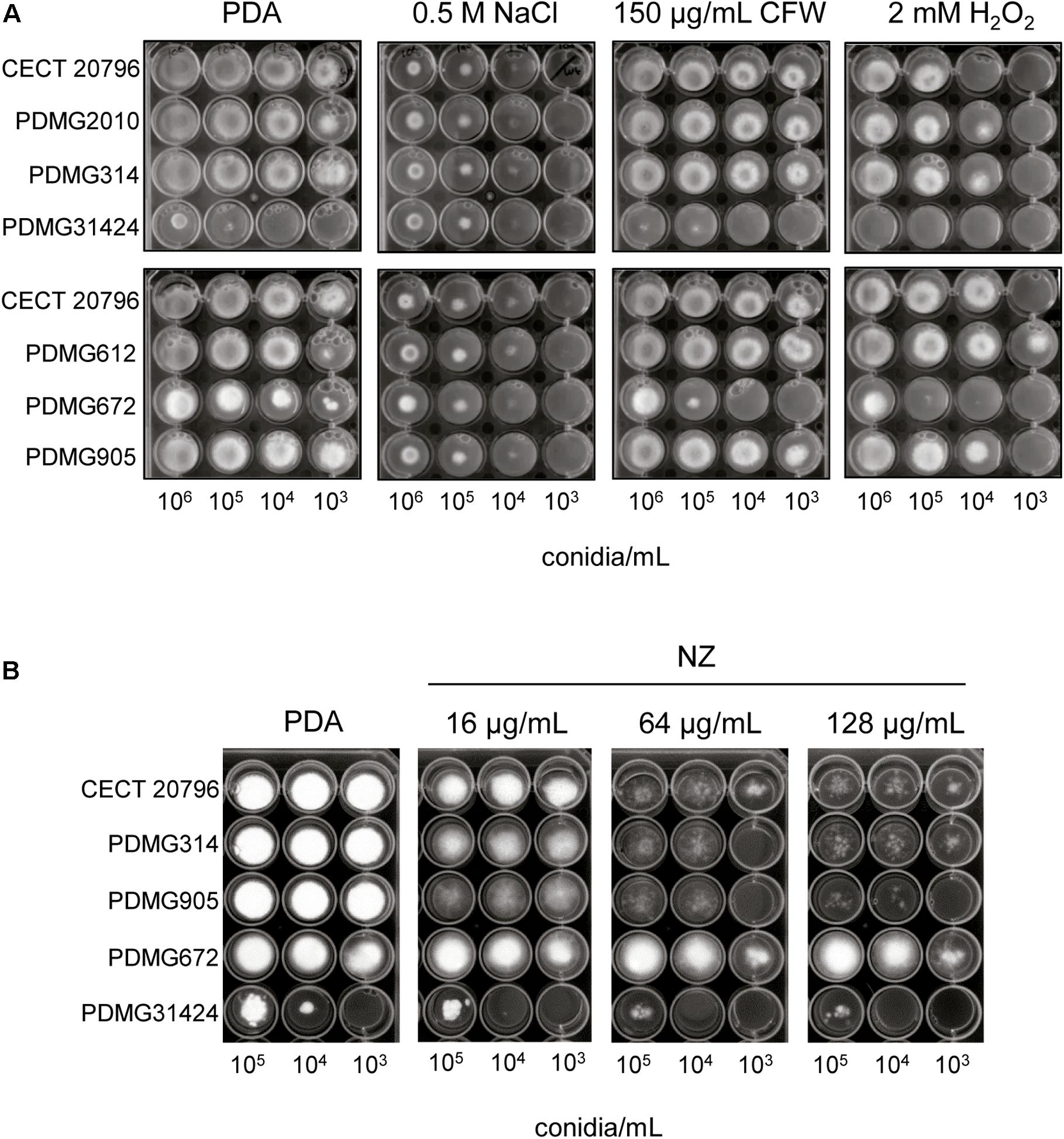
Figure 2. Sensitivity of P. digitatum strains to different compounds. (A) Comparison of different strains growth on PDA plates and PDA supplemented with the indicated compounds. Serial 1/10 dilutions of conidia of each strain were applied as indicated. (B) Growth of strains in presence of different concentrations of chitin synthase inhibitor nikkomycin Z (NZ) at 5 days of incubation at 24°C. Serial dilutions of conidia of each strain were applied as indicated. Different strains were: parental strains CECT 20796 and PDMG314; deletion strains: ΔchsII (PDMG905), ΔchsV (PDMG31424), and ΔchsVII (PDMG672) and two different ectopic strains PDMG2010 (chsV) and PDMG612 (chsVII).
The addition to PDA medium of compounds like CFW or SDS, which affect the CW and the cell membranes, respectively, produced a growth reduction in both ΔchsV and ΔchsVII mutants (Figure 2A and Supplementary Figure S3), indicative of CW damage. Similar test performed under oxidative stress (2 mM of H2O2) showed a higher sensitivity and a growth reduction of PDMG31424 (ΔchsV) (Figure 2A), similar to the results previously described for PDMG672 (ΔchsVII) (Gandía et al., 2014). In all these experiments, the strain PDMG905 (ΔchsII) did not differ in its growth profile in comparison with parentals CECT 20796 or PDMG314 (Figure 2A).
Treatments with the chitin synthase inhibitor NZ were conducted to determine changes in the sensitivity of the different strains. As shown in Figure 2B, the growth of the different strains was reduced in presence of distinct concentrations of NZ. Transformants ΔchsII and ΔchsV showed slightly higher sensitivity to NZ than that of their parental strains, whereas transformant PDMG672 (ΔchsVII) revealed increased resistance at all NZ concentrations tested.
Morphology and Chitin Content of chs Mutants
Changes in mycelial morphology of deletion mutants were analyzed by bright field and fluorescence microscopy under CFW staining that binds chitin. Mycelium of ΔchsII mutant strains (PDMG905) did not show morphological differences when it was compared with the two parental strains CECT 20796 or PDMG314 (data not shown). The mycelium of PDMG31424 (ΔchsV) exhibited strong microscopic alterations (Figures 3A,B). Germlings after 18 h of incubation presented shorter or aborted germ tubes and enlarged conidia (white arrowheads in Figure 3A); these conidia in the mutant were clearly larger than the usual enlargement that occurs in the wild type upon germination (open arrowheads in Figure 3A). At later times, balloon-like structures appeared and increased in size and number over time (arrowheads in Figure 3B), and were larger than similar structures observed previously in the ΔchsVII mutant (PDMG672) (Gandía et al., 2014). In addition, these balloon-like structures exhibited intense CFW staining and occasionally broke down and released the intracellular content (asterisks in Figure 3B). These structures and the increase in CFW staining are indicative of alterations in the CW and chitin deposition. The chitin content of the chs mutant strains was determined and PDMG31424 (ΔchsV) showed a statistically significant higher chitin content than in its parental or in the other strains (Figure 3C).
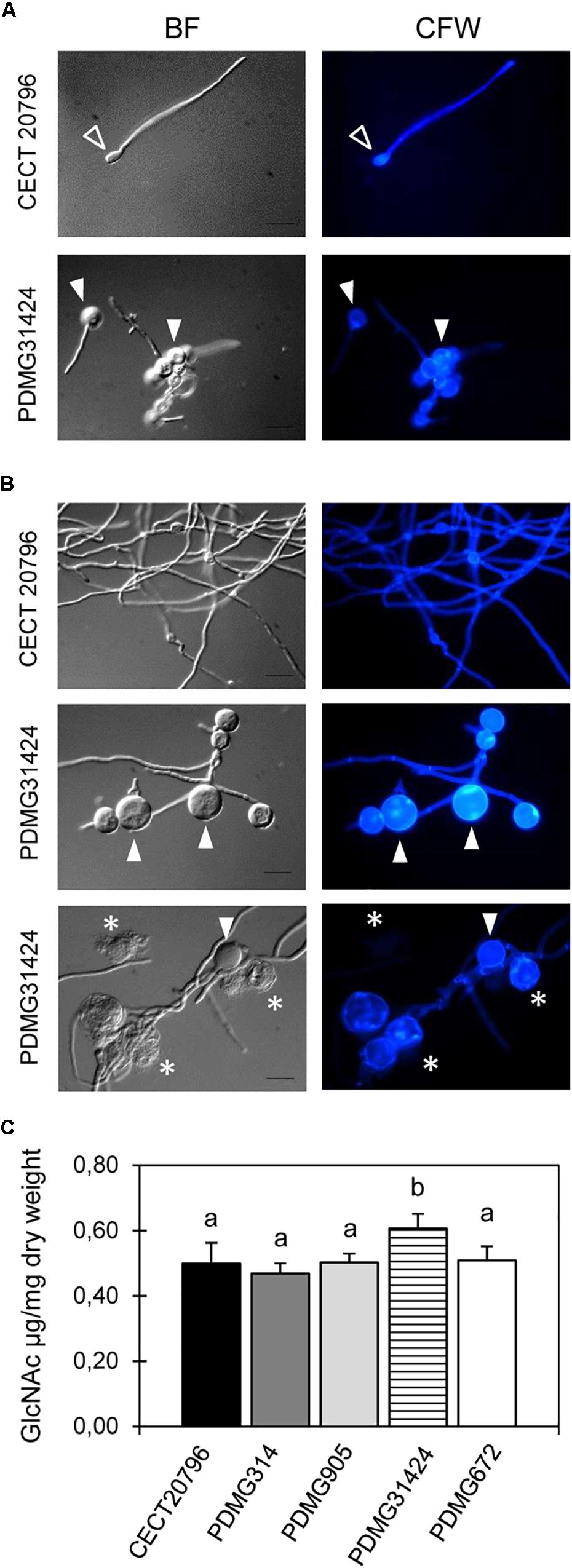
Figure 3. Morphological changes observed by microscopy and chitin content in P. digitatum strains. Representative images (BF, left and CFW stain, right) of fluorescence microscopy of the parental CECT 20796 and PDMG31424_ΔchsV strains at different times of incubation at 24°C. (A) 18 h, (B) 24 and 48 h. White arrowheads indicate balloon-like structures in mycelium of deletion strains intensively CFW stained and asterisks show shorter interseptal distances. (C) Chitin content assay of different strains determined as N-acetyl-D-glucosamine (GlcNAc) released per dry weight of tissue. Data are expressed as means ± SD of two replicates. Different letters indicate statistically significant differences among the strains as determined by ANOVA test p < 0.05. Scale black bars in BF images are 10 μm.
The conidia of the different strains were observed at 100× objective and, at first sight, ΔchsII conidia were clearly elongated compared with the parental and other mutants (Figure 4A). In an attempt to quantify any differences in the morphology of conidia, two shape parameters were analyzed in distinct mutants. The Feret diameter (F) and the circularity were measured to verify the difference among conidia shape and size in ΔchsII and other deletion mutants. Both parameters confirmed differences in two independent ΔchsII mutants (PDMG905 and PDMG908), more pronounced than in the ΔchsV mutant (PDMG31424) as compared to the parental (Figure 4B). PDMG314 and CECT 20796 parental strains did not present any differences between them (data not shown).
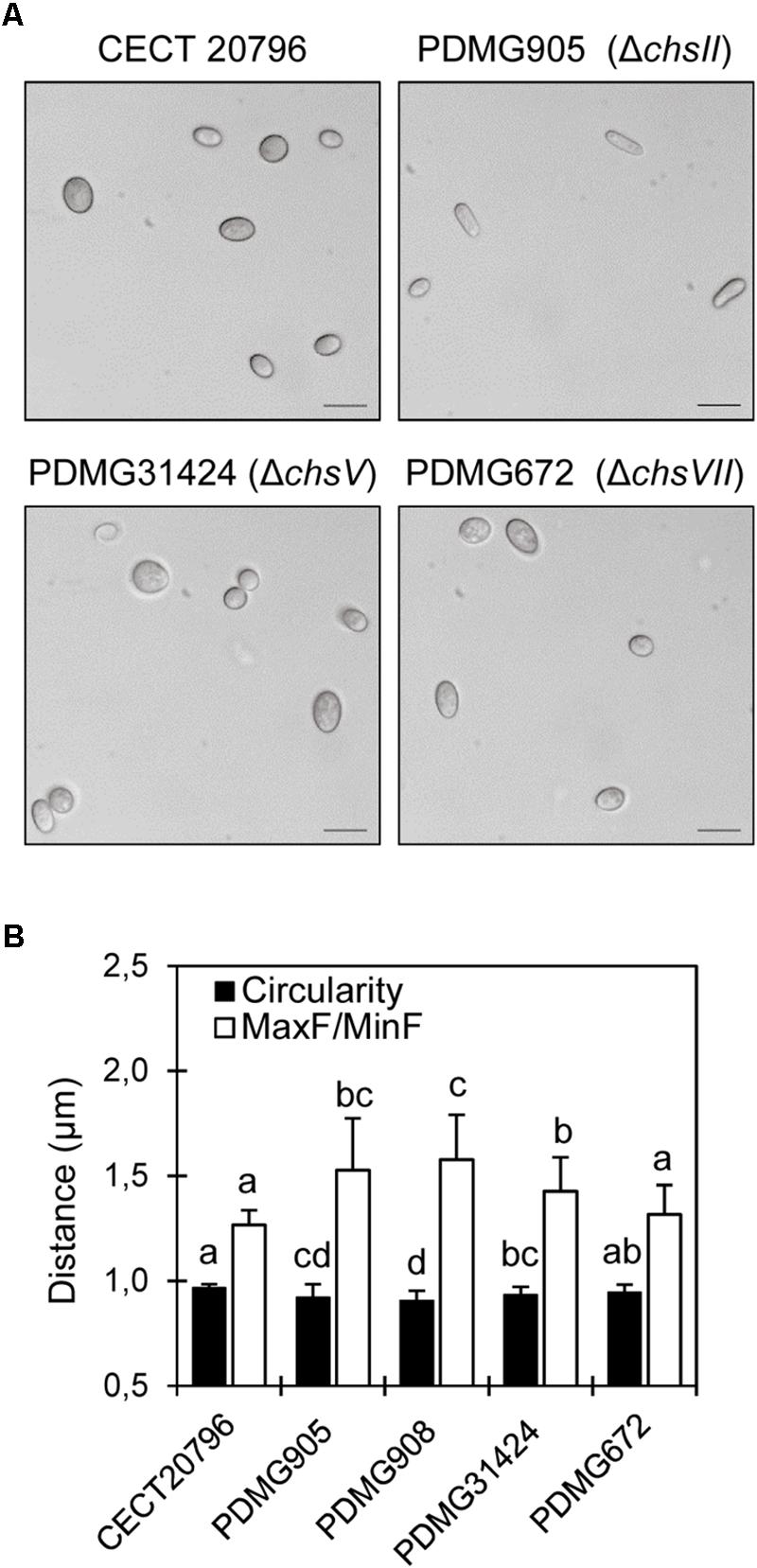
Figure 4. Phenotypic morphology and dimensions of spores of different P. digitatum strains. (A) Differential interference contrast (DIC) images obtained with 100x objective, of conidia of different strains. Scale black bars are 10 μm. (B) Analyses of two different parameters of conidia (F and circularity) in different strains. Both PDMG905 and PDMG908 are ΔchsII deletion strains. Different letters indicate statistically significant differences among the strains determined by ANOVA test p < 0.05.
Effect of Antifungal Proteins on chs Gene Expression and Deletion Mutants
Previously, chitin synthesis and chitin synthase genes have been linked to the sensitivity of filamentous fungi to AFPs (Hagen et al., 2007; Martín-Urdiroz et al., 2009). We have recently managed to produce and purify in high amounts two highly active antifungal proteins (AfpA and AfpB) (Garrigues et al., 2017, 2018), offering a great opportunity to evaluate the role of chitin synthase genes in the mode of action of these proteins. Side-by-side experiments have shown that these two AFPs are more active in vitro against P. digitatum than the previous AFP from A. giganteus and PAF from P. chrysogenum, and that both AfpA and AfpB show protective effect against the infection of fungal plant pathogens (Garrigues et al., 2017, 2018; Shi et al., 2019).
We have determined global gene expression changes by RNA-seq after treatment of P. digitatum with the self-AfpB (Bolós et al., unpublished). Table 1 shows the fold changes in gene expression corresponding to the chs genes, data extracted from the global gene expression experiment. In these data sets, treatments A and B correspond, respectively, to 1 and 3 h treatments of previously 24 h-grown P. digitatum mycelium with sub-inhibitory concentrations of AfpB; while treatment C corresponds to P. digitatum grown for 24 h in the continuous presence of sub-inhibitory concentrations of AfpB. An overall reduction of chs gene expression is observed that only reached statistical significance in treatment C and in the case of chsI, chsII, and chsIIIb.
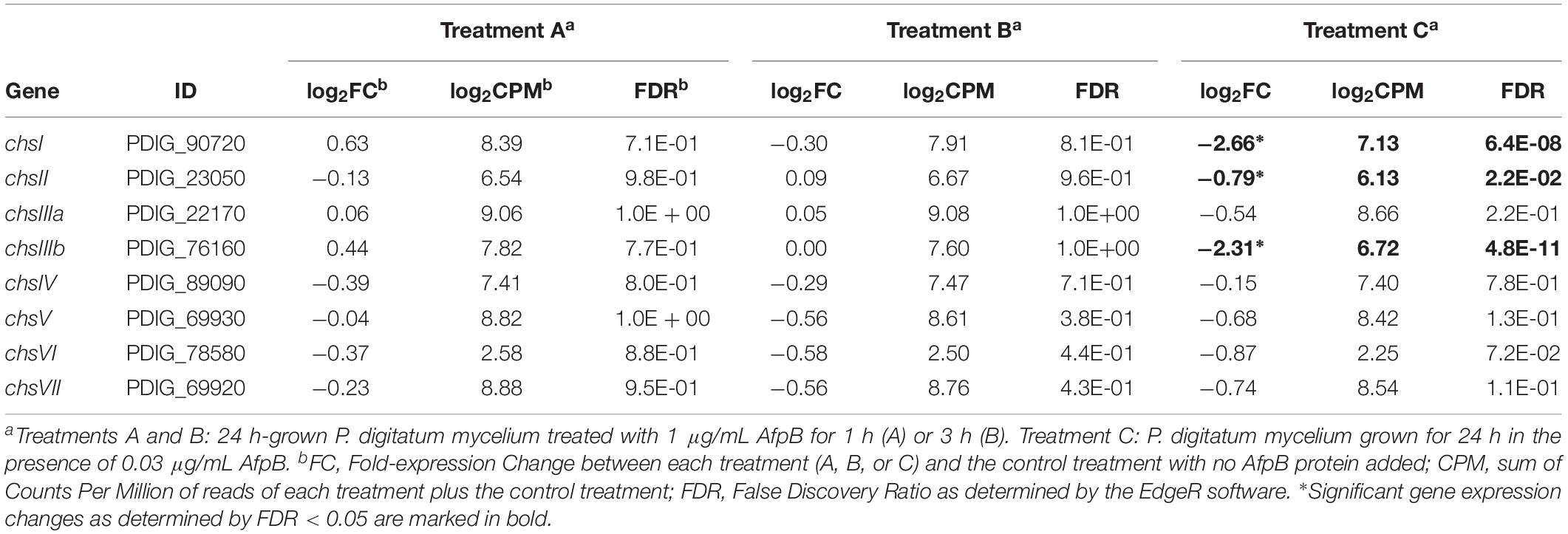
Table 1. Gene expression changes of the genes coding for chitin synthases after AfpB treatments determined by RNA-seq.
Next, we assayed the effect of the AfpB from P. digitatum and AfpA from P. expansum on the Chs mutants. Dose-response curves at 48 h of growth (Figure 5) and MIC values at the end of experiments (Table 2) showed differences among mutants. The ΔchsII mutant exhibits a behavior similar to that of the parental strain, in spite of the significant gene expression change of this gene in the presence of AfpB (Table 1). On the other hand, the MMD-chitin synthase mutant PDMG672 (ΔchsVII) showed increased sensitivity to both proteins in all the experiments carried out. At the end of the experiment (72 h), growth of PDMG672 was completely inhibited at concentrations of AfpB and AfpA twofold lower than the parental or other strains, and at lower concentrations of proteins or shorter incubation times the growth of this mutant was affected. This sensitivity was more accused in the case of the more active AfpA (establishing a MIC of 0.5 μg/mL) (Table 2). In the case of the other MMD-chitin synthase mutant PDMG31424 (ΔchsV) the data were more difficult to interpret, due to the poor growth of this mutant in liquid medium; see for instance its reduced growth at non-inhibitory 0.25 μg/mL AfpB in Figure 5B, in comparison with the other strains including its parental PDMG314. In any case, the mutant behaved as its parental strain in some experiments with the same MIC while in others showed slightly increased tolerance with a twofold increase of MIC. Nevertheless, in some experiments, this ΔchsV had increased OD values over its parental at intermediate protein concentrations and times (see for instance white and black squares in Figure 5B).
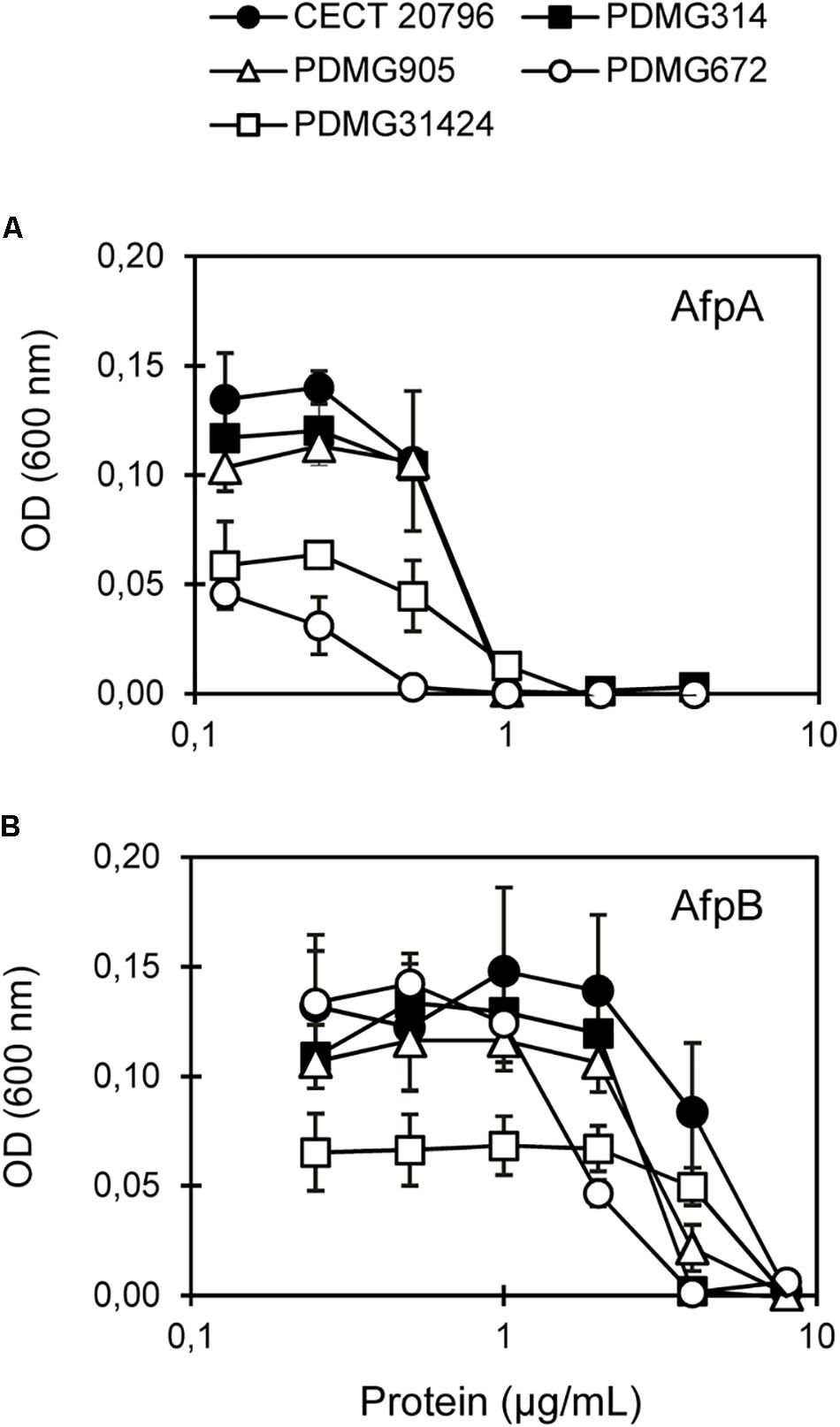
Figure 5. In vitro inhibitory activity of antifungal proteins against chs mutants. Dose-response curves of antifungal proteins against parental strains (CECT 20796 and PDMG314) and chs mutants (PDMG905_ΔchsII; PDMG672_ΔchsVII and PDMG31424_ΔchsV). (A) Response to AfpA. (B) Response to AfpB. Curves show mean ± SD after 48 h at 25°C. Strains showed: CECT 20796 black circles; PDMG314 black squares; PDMG905 white triangles; PDMG31424 white squares and PDMG672 white circles.
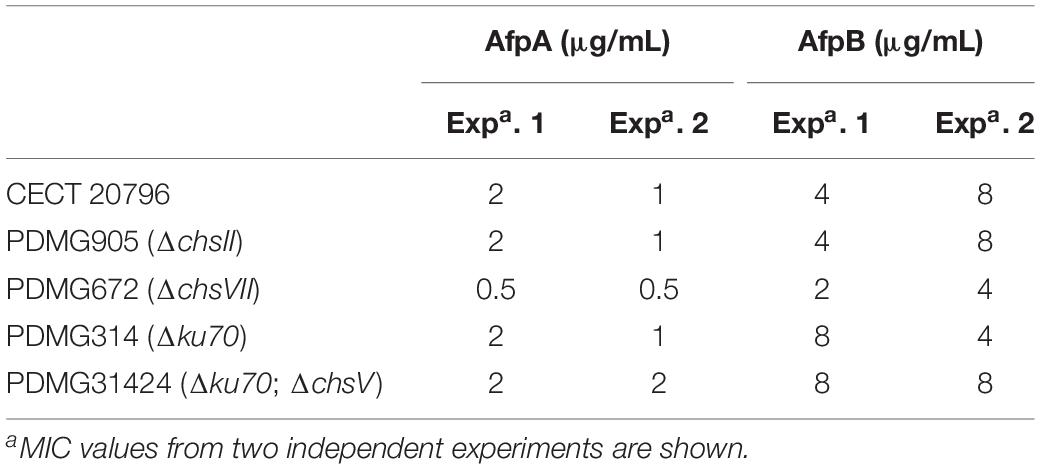
Table 2. Minimal inhibitory concentration (MIC) values (μg/mL) of two antifungal proteins AfpA and AfpB against different fungal strains tested.
Comparison of Virulence of chs Mutants
Finally, we analyzed the virulence of the different chs mutants by conducting experimental inoculation assays on orange fruits with different inoculum doses (Figure 6). Low inoculum doses (104 conidia/mL) confirmed that the deletion strain PDMG905 (ΔchsII) had the same incidence of infection as parental strains CECT 20796 and PDMG314. However, the deletion strains in MMD-chitin synthase genes chsV and chsVII had a low incidence of infection, lower in PDMG31424 than in PDMG672 (Figure 6A). At seven dpi, between 75 and 85% of wounds were infected in both parental strains and PDM905; however, PDMG672 (ΔchsVII) mutant reached 60% of infection while PDMG31424 (ΔchsV) was only able to infect 43% of wounds (Figure 6A). Both mutants showed reduced maceration area in citrus fruits (data not shown). Previous analyses had shown reduced virulence in the strain PDMG672, confirmed again in this work (Gandía et al., 2014). When higher inoculum doses were used (106 conidia/mL) all strains developed symptoms quickly and reached 100% of infection incidence at 3 dpi (Figure 6B top panel). The maceration area on citrus fruits was the same in all strains analyzed under these inoculum conditions (data not shown), and the sporulation index of infected wounds was specifically reduced in ΔchsV and ΔchsVII mutants (Figures 6B bottom panel and Figure 6C). At the end of the experiments (7 dpi), citrus fruits infected by ΔchsV and ΔchsVII at 106 conidia/mL were 100% infected and completely macerated although most of them did not develop green conidia, thus confirming that the ΔchsV mutants have a phenotype similar to that of the previously described ΔchsVII (Gandía et al., 2014) (Figures 6B,C).
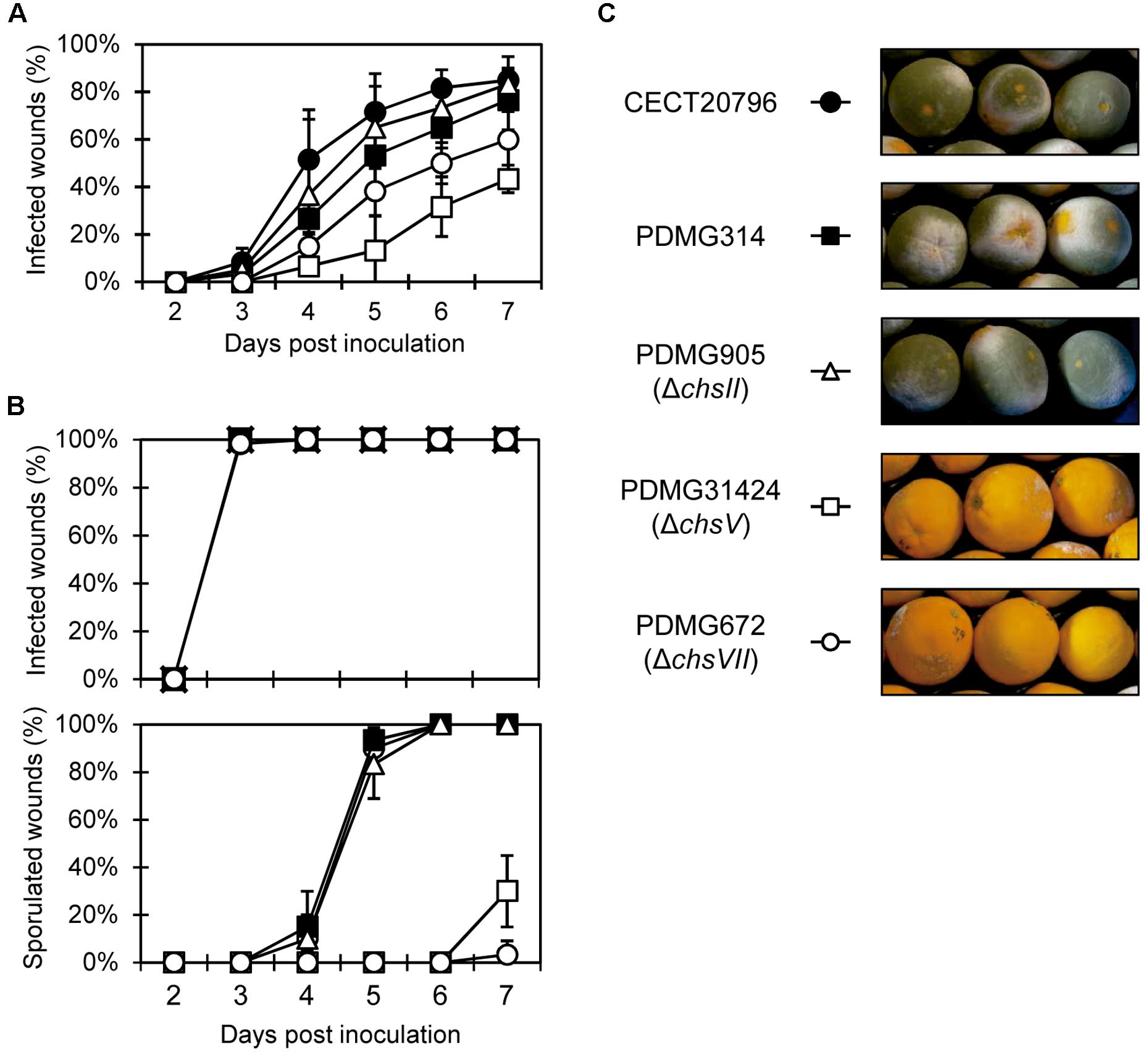
Figure 6. Virulence assays of different P. digitatum strains on orange fruits. (A) Incidence of infection caused by different strains at low inoculum dose (104 conidia/mL). Data indicate percentage of infected wounds (mean value ± SD) at each day post-inoculation. (B) Incidence of infection caused by different strains, at high inoculum dose (106 conidia/mL) (top panel) and percentage of sporulated wounds on fruits from the same experiment (bottom panel). (C) Representative images of fruits infected with different strains at 7 days post-inoculation from the experiment shown in (B). Note the absence of mycelia in fruits infected with ΔchsV and ΔchsVII deletion strains.
Discussion
The role of chitin synthase genes in fungal development and pathogenesis has been studied through gene deletions in a wide number of fungi, and the general conclusions are common in many fungal species, with just minor differences (Morcx et al., 2013; Gandía et al., 2014; Muszkieta et al., 2014; Liu et al., 2016; Zhang et al., 2016). However, the involvement of chs genes and chitin biosynthesis process in the sensitivity to AFPs from fungal origin remains controversial (Hagen et al., 2007; Martín-Urdiroz et al., 2009; Ouedraogo et al., 2011).
Our previous disruption of the chsVII gene of P. digitatum showed important effects on growth, conidia production or virulence and compensatory changes of expression in other chs genes, indicative of redundancy or cooperative role among different chs (Gandía et al., 2014), similar to other filamentous fungi (Lenardon et al., 2010; Sánchez-Vallet et al., 2014). In order to study the role of chsII, which is one of the most induced gene by chsVII disruption, and the role of the other MMD-chs gene (chsV), we obtained viable mutants in this study. Both of them belong to different classes and divisions, and results could indicate the different role of each Chs division in this fungus and could help to clarify the mechanism of action of antifungal proteins.
The Mutation of chsII in P. digitatum Reduced the Number and Changed the Morphology of Conidia
Penicillium digitatum mutants in the chsII gene were similar to the parental strain in many aspects. There were no morphological changes in growth or sensitivity to temperature (Figure 1), sensitivity to different compounds (Figure 2), chitin content (Figure 3), sensitivity to antifungal proteins AfpA or AfpB (Figure 5) or virulence (Figure 6). The phenotype of class II Chs mutants is subtle, and did not show significant growth alterations in many fungi such as Aspergillus nidulans (Motoyama et al., 1997), Aspergillus fumigatus (Munro and Gow, 2001), or Neurospora crassa (Fajardo-Somera et al., 2015). Gonçalves et al. (2016) explained that the single mutants in class I or class II did not have changes in growth or morphology due to redundant roles of these Chs classes. In the division I overlapping and cooperative functions of Chs are known, since the generation of double or multiple mutants in Chs belonging to this division produces abnormal morphologies. Double mutants in chsI/chsII in A. nidulans presented a dramatic decrease in conidia production not observed in the single mutants, despite the normal content of chitin (Fujiwara et al., 2000; Ichinomiya et al., 2005). Quadruple mutant ΔchsA/C/B/G in A. fumigatus showed reduced conidiation, growth and Chs activity (Muszkieta et al., 2014). In other studies, conducted in M. oryzae or A. nidulans, a role of this gene in conidiogenesis has been described based on the low conidia production of single mutants (Culp et al., 2000; Fujiwara et al., 2000; Kong et al., 2012). In P. digitatum a possible role of chsII in conidiogenesis was established on the basis of its strong induction during sporulation on PDA plates (Gandía et al., 2012). The mutants reported here in chsII showed a 30-40% reduction in the number of conidia (Figure 1) and abnormalities in conidia morphology (Figure 4) that demonstrate a role in conidiogenesis despite normal chitin content (Figure 3). In Wangiella dermatitidis, class II Chs was established as responsible for normal reproductive growth and mutants in this gene had sometimes enlarged cells (Zheng et al., 2006). All of these results support the possible involvement of class II Chs in P. digitatum conidiogenesis.
Disruption of MMD-Chitin Synthases Alters Morphology and Virulence of P. digitatum
A number of studies have described the important role of the MMD ChsV and ChsVII chitin synthases in the development and virulence of fungal plant pathogens (Kim et al., 2009; Treitschke et al., 2010; Larson et al., 2011; Kong et al., 2012; Fernandes et al., 2016). Previously, we obtained and characterized P. digitatum disruption strains in chsVII (Gandía et al., 2014). We did not obtain mutants in chsV in the first attempts using the wild type CECT 20796 as parental strain. We only succeeded after using a strain (PDMG314) with the non-homologous end-joining pathway (NHEJ) inactivated by deletion of ku70 gene involved in this route (Gandía et al., 2016). The increased gene targeting efficiency in P. digitatum using this strain allowed the generation of disruption mutants in chsV, which showed phenotypic alterations similar but more accused than ΔchsVII mutants, in accordance with other class V mutants that showed strongest phenotypes (Roncero et al., 2016).
Both mutants (ΔchsV and ΔchsVII) had reduced growth and conidia production, increased sensitivity to CFW, H2O2 and temperature stress, and showed alterations in the chitin content and hyphal morphology (Figures 1–3). Moreover, the ΔchsV mutant shows extremely enlarged balloon-like cells that eventually break out and release the intracellular material (Figure 3B). The number and size of these structures are higher than those described for ΔchsVII (Gandía et al., 2014), and both are intensely stained with CFW and could not be restored by osmotic stabilizers. These structures are common in other class V/class VII mutants (Takeshita et al., 2006; Jiménez-Ortigosa et al., 2012; Muszkieta et al., 2014).
All these phenotypic effects are indicative of alterations in CW (Munro, 2013; Walker et al., 2015) and, consequently, growth was recovered with osmotic stabilization with sorbitol or NaCl (Figure 2 and Supplementary Figure S3). Similar defects were present in ChsV/ChsVII mutants obtained in other filamentous fungi such as A. fumigatus (Muszkieta et al., 2014), A. nidulans (Takeshita et al., 2006), B. cinerea (Morcx et al., 2013), Fusarium oxysporum (Martín-Urdíroz et al., 2008), or M. oryzae (Kong et al., 2012). Several studies have pointed out the role of MMD-Chs in the hyphal growth, being transported by vesicles along actin filaments to the apical hyphae wherein CW-forming foci are located and contributing to polarize CW synthesis (Schuster et al., 2012; Fajardo-Somera et al., 2015; Takeshita et al., 2015). The apical growth of ΔchsV is heavily affected in germlings of the mutants (Figure 3A).
Penicillium digitatum shows temperature sensitivity. It grows and germinates at 24–25°C as optimal temperature, but at temperatures above 25°C the levels of germination are reduced up to 40–60% and at 30°C it is unable to germinate or grow (Plaza et al., 2003). The disruption of chsV using Δku70 as genetic background produced strains with extreme temperature sensitivity, part of which could be explained by the sensitivity of the parental strain from which it proceeds (Gandía et al., 2016). The class V Chs protein in W. dermatitidis is necessary for growth of this fungus at 37°C maintaining the integrity of hyphal tip (Liu et al., 2004; Abramczyk et al., 2009). Also, mutants of A. fumigatus in MMD-chs genes showed increased sensitivity to high temperature (Muszkieta et al., 2014).
It has been described that the perturbations of CW caused by mutations in chs genes trigger compensatory mechanisms such as the increase in chitin synthesis and content (Munro et al., 2007; Roncero et al., 2016). In P. digitatum, the mutant ΔchsV was again remarkable in the abnormal and high chitin content compared to wild type (Figure 3C). However, we did not find any correlation between the chitin content and sensitivity to the chitin biosynthesis inhibitor NZ. This antifungal inhibits chitin synthesis by preventing the conversion of UDP-N-acetylglucosamine into chitin. Treatment with this inhibitor revealed slight sensitivity in ΔchsII and ΔchsV (Figure 4). Both strains had different chitin levels, higher in ΔchsV (Figure 3). Only ΔchsVII registered increased resistance to NZ and its chitin content was similar to the wild type. This pattern of NZ sensitivity was not comparable to other fungi. For instance, M. oryzae mutants in class II (Chs2) and class V (Chs6) were more sensitive to NZ as in P. digitatum, but mutants in class VII (Chs5) were not different from wild type (Kong et al., 2012). In addition, none of the Chs mutants had significant changes in the chitin content (Kong et al., 2012). None of the MMD-Chs mutants in A. fumigatus showed differences in susceptibility to NZ either (Jiménez-Ortigosa et al., 2012).
Many studies have suggested the role of MMD-chitin synthases in pathogenicity or virulence (Fernandes et al., 2016). In our study, both MMD-mutants showed reduced virulence in citrus fruits (Figure 6) confirming this general behavior also in a fruit-specific pathogen. The incidence of infection was lower in ΔchsV mutant than in ΔchsVII, pointing again to the more extreme phenotype of the former. As in the case of ΔchsVII (Gandía et al., 2014), the mutant in class V showed delayed disease progression and was unable to develop mycelium in most of the infected wounds in spite of the massive colonization of the fruit tissue. These results confirmed the prominent role of these chitin synthases in virulence of P. digitatum, since are the most induced chs genes during infection of citrus fruits (Gandía et al., 2014). Altered virulence was demonstrated in other phytopathogenic fungi when MMD-Chs were eliminated (Weber et al., 2006; Treitschke et al., 2010; Larson et al., 2011; Cui et al., 2013), but, again, subtle differences were observed in fungi such as M. oryzae in which the class V was non-pathogenic but the class VII mutant behaved as the wild type (Kong et al., 2012). In F. oxysporum, the inability of mutants in MMD-chs to invade tomato plants was linked to their hypersensitivity to hydrogen peroxide, compound produced as part of the plant defense response (Madrid et al., 2003). On the contrary, Ustilago maydis mutants in class V lost the ability to infect, but were not affected in their sensitivity to H2O2, so the absence of virulence does not seem to be due exclusively to a lack of response to plant defense (Weber et al., 2006; Treitschke et al., 2010). Despite this, it seems that these MMD-chitin synthases could be involved in response to host defense mechanism (Fernandes et al., 2016). In our case, both MMD-mutants were more sensitive to H2O2 than parental strains. P. digitatum is able to neutralize the ROS response that citrus fruit generate upon infection (Macarisin et al., 2007; Costa et al., 2019). In our previous study, we speculated that the high sensitivity of ΔchsVII mutant to H2O2 was related to its decreased virulence (Gandía et al., 2014). The fact that another H2O2 sensitive mutant (ΔchsV) also has reduced virulence whereas ΔchsII with a sensitivity to H2O2 similar to the parental strain does not present changes in its virulence would further support this hypothesis.
Differential Sensitivity of MMD-Chs Mutants to Antifungal Proteins and Peptides
Antimicrobial peptides and proteins (AMPs) are a promising alternative in the fight against fungal pathogens (Hegedüs and Marx, 2013). Some of these peptides and proteins may have modes of action different from available fungicides. The involvement of CW in the interaction of AMP/AFPs with fungi is well documented. Upon interaction with the fungal cell, the A. giganteus AFP localizes first at the outer CW rich in glycoproteins (Theis et al., 2005), from where it is actively internalized (Oberparleiter et al., 2003). Other AFPs also locate intracellularly (Sonderegger et al., 2017; Huber et al., 2018), and we have similar observations for the AfpB from P. digitatum (Marcos et al., unpublished).
The observation that the antifungal protein AFP from A. giganteus can be purified by chitin affinity chromatography (Liu et al., 2002) sustained the early hypothesis that chitin or chitin synthesis are important factors in the mode of action of AFPs or in the response mechanisms of fungi to these proteins (Hagen et al., 2007). However, some contradictory results were found in the studies published so far with this AFP (Hagen et al., 2007; Martín-Urdiroz et al., 2009; Ouedraogo et al., 2011), since the results reported in these works do not completely correlate with our own observations described here. Exposure to A. giganteus AFP inhibited the biosynthesis of chitin in several sensitive filamentous fungi (Hagen et al., 2007). Chitin staining with CFW decreased and was altered in the sensitive A. nidulans treated with PAF (Binder et al., 2010) or Aspergillus flavus with PgAFP (Delgado et al., 2015). Down regulation of chs genes observed upon treatment of the sensitive P. digitatum with its self-AfpB (Table 1) is consistent with the inhibition of Chs activity observed in sensitive filamentous fungi treated with AFP (Hagen et al., 2007).
In addition, mutants in ΔchsIII and ΔchsV from F. oxysporum or A. oryzae were reported to be highly resistant to AFP, with a MIC from 1 to 400 μg/mL in the case of F. oxysporum ΔchsV. Interestingly, AFP did not inhibit chitin biosynthesis in the ΔchsV resistant strain but rather induced chitin synthase activity (Hagen et al., 2007). In contrast, a different study showed higher sensitivity to this protein in the same F. oxysporum ΔchsV strain (Martín-Urdiroz et al., 2009).
Ours is the first study that analyzes and compares the sensitivity to two AFPs in mutants from both MMD-containing Chs and provides data on ChsVII. Moreover, AfpA and AfpB are highly active AFPs, and AfpB was identified in the P. digitatum strain that is used for mutant generation. This context offers a great opportunity to refine the role of chitin biosynthesis in the action of AFPs. When our P. digitatum chs mutants were exposed to either AfpA or AfpB, only one of them (ΔchsVII) presented a subtle but clear differential response, with sensitivity twofold higher than CECT 20796 or PDMG314 parental strains (Figure 5). In this study, the ΔchsV presented an even higher chitin content than ΔchsVII. It was difficult to conclude any differential behavior of this mutant in terms of sensitivity to the proteins due to its growth penalty; if any, it is of increased tolerance that, however, did not reach the extreme resistance of F. oxysporum ΔchsV (Hagen et al., 2007).
Echinocandins are antifungals used in clinic. They are cyclic lipo-hexapeptides that are produced by ascomycetous fungi through non-ribosomal peptide synthases that target the CW by inhibiting β-glucan synthesis (Emri et al., 2013). The mutation of MMD-Chs (CsmA and CsmB) in A. fumigatus produced extreme sensitivity to several echinochandins including caspofungin (Jiménez-Ortigosa et al., 2012). We have also developed and characterized the small synthetic hexapeptide PAF26 (Muñoz et al., 2013). As occurs with AFPs, PAF26 firstly interacts with CW and is later internalized by the fungal cell to produce the cytotoxicity and antifungal effect intracellularly (Marcos and Gandía, 2009; Muñoz et al., 2013). Thus, the CW is critical for the interaction and activity of AFPs and other antifungal peptides such as PAF26. Defects in CW and protein mannosylation in the protein O-mannosyltransferase mutant Δpmt2 increase the tolerance of P. digitatum to PAF26 (Harries et al., 2015). On the contrary, the ΔchsVII mutant did not show differential sensitivity to PAF26 compared to the parental strain (Gandía et al., 2014). This is in contrast with the increased sensitivity of this mutant to the AFPs tested in this study.
The comparison between ΔchsV and ΔchsVII of growth phenotype, the morphological alterations in the hyphae, temperature sensitivity or decreased tolerance to CW interfering compounds suggest a more severe CW alteration in ΔchsV, which does not correlate with the relative sensitivity to either AfpA or AfpB, indicating that CW may not be an important (or unique) determinant for sensitivity to AFPs.
The mode of action and effects of the A. giganteus AFP were also analyzed in the model unicellular fungus S. cerevisiae, which is tolerant to the protein, with no growth inhibition up to 400 μg/mL AFP (Ouedraogo et al., 2011). The authors showed that AFP activates among others the CW integrity (CWI) pathway and induces chitin content and the expression of the three chs genes found in yeast. We have shown induction of the CWI pathway by AfpB (Gandía et al., 2019). Treatment with AfpB induces phosphorylation of Slt2 and Hog1 MAPK, revealing the involvement of CWI and high osmolarity glycerol (HOG) pathways in its mechanism of action, but not in defensive roles, since mutants in these routes were more tolerant to the action of the protein (Gandía et al., 2019). The increased gene expression in yeast exposed to AFP contrasts with the overall repression we observed with P. digitatum. The data reported here reinforces the conclusion that an increased chitin response is found in AFP-resistant fungi (S. cerevisiae or F. oxysporum ΔchsV) while AFPs reduce chitin biosynthesis in sensitive filamentous fungi (Ouedraogo et al., 2011). However, the exact role of chs gene mutation is still a matter of debate and needs to be further analyzed in more depth since different studies including this one have led to different conclusions.
Previous studies have pointed the role of MMD-Chs at the growth region of the hyphal apical tip (Steinberg, 2011; Schuster et al., 2016; Geoghegan et al., 2017). Cooperation of both MMD-Chs is needed for the final step of exocytosis that would form the new glucan and chitin chains in the growing hyphae (Geoghegan et al., 2017). Future studies with fluorescence tagged Afps would provide evidence of their location in the hyphae of parental and mutant strains and whether this location matches that described for MMD-Chs. These assays would help us to understand the mechanism of action of these antifungal proteins and the chitin biosynthesis response in sensitive or resistant strains to Afps.
Our results show that chs genes belonging to different divisions of Chs are responsible for different aspects in P. digitatum biology. Deletion of ChsII did not produce significant changes in the morphology or virulence of the fungus but affected its conidia formation process. Mutants in MMD-Chs from division II compromised growth, morphology, and virulence of the fungus, and influenced the sensitivity to antifungal proteins. Further studies about changes in P. digitatum in response to AFPs exposure will be addressed in the near future.
Data Availability Statement
All datasets generated for this study are included in the manuscript/Supplementary Files.
Author Contributions
MG, PM, and JM conceived and designed the study. MG obtained the different mutants. MG and SG phenotypically characterized the mutants. BB and JM performed the RNA-seq experiments and analyses. SG and PM obtained and purified the proteins. MG and JM prepared the first draft of the manuscript. PM and JM provided the funding. All authors contributed to manuscript revision, read and approved the submitted version.
Funding
This work was funded by BIO2015-68790-C2-1-R and RTI2018-101115-B-C21 from the “Ministerio de Ciencia, Innovación y Universidades” (Spain) (MICINN/FEDER Funds) and PROMETEO/2018/066 from ‘Conselleria d’Educació’ (Generalitat Valenciana, Comunitat Valenciana, Spain). SG was recipient of a predoctoral scholarship (FPU13/04584) within the FPU program from “Ministerio de Educación, Cultura y Deporte” (MECD, Spain). We acknowledge support of the publication fee by the CSIC Open Access Publication Support Initiative through its Unit of Information Resources for Research (URICI).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge the help with the microscopy experiments of José M. Coll-Marqués (IATA, Valencia, Spain) and Shaomei Xu for their excellent technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.02400/full#supplementary-material
References
Abramczyk, D., Park, C., and Szaniszlo, P. J. (2009). Cytolocalization of the class V chitin synthase in the yeast, hyphal and sclerotic morphotypes of Wangiella (Exophiala) dermatitidis. Fungal Genet. Biol. 46, 28–41. doi: 10.1016/j.fgb.2008.10.004
Binder, U., Oberparleiter, C., Meyer, V., and Marx, F. (2010). The antifungal protein PAF interferes with PKC/MPK and cAMP/PKA signalling of Aspergillus nidulans. Mol. Microbiol. 75, 294–307. doi: 10.1111/j.1365-2958.2009.06936.x
Brown, G. D., Denning, D. W., and Levitz, S. M. (2012). Tackling human fungal infections. Science 336:647. doi: 10.1126/science.1222236
Cheng, W., Song, X. S., Li, H. P., Cao, L. H., Sun, K., Qiu, X. L., et al. (2015). Host-induced gene silencing of an essential chitin synthase gene confers durable resistance to Fusarium head blight and seedling blight in wheat. Plant Biotechnol. J. 13, 1335–1345. doi: 10.1111/pbi.12352
Choquer, M., Boccara, M., Gonçalves, I. R., Soulie, M. C., and Vidal-Cros, A. (2004). Survey of the Botrytis cinerea chitin synthase multigenic family through the analysis of six euascomycetes genomes. Eur. J. Biochem. 271, 2153–2164. doi: 10.1111/j.1432-1033.2004.04135.x
Costa, J. H., Bazioli, J. M., de Moraes Pontes, J. G., and Fill, T. P. (2019). Penicillium digitatum infection mechanisms in citrus: what do we know so far? Fungal Biol. 123, 584–593. doi: 10.1016/j.funbio.2019.05.004
Cui, Z., Wang, Y., Lei, N., Wang, K., and Zhu, T. (2013). Botrytis cinerea chitin synthase BcChsVI is required for normal growth and pathogenicity. Curr. Genet. 59, 119–128. doi: 10.1007/s00294-013-0393-y
Culp, D. W., Dodge, C. L., Miao, Y. H., Li, L., Sag-Ozkal, D., and Borgia, P. T. (2000). The chsA gene from Aspergillus nidulans is necessary for maximal conidiation. FEMS Microbiol. Lett. 182, 349–353. doi: 10.1111/j.1574-6968.2000.tb08920.x
Delgado, J., Acosta, R., Rodriguez-Martin, A., Bermudez, E., Nunez, F., and Asensio, M. A. (2015). Growth inhibition and stability of PgAFP from Penicillium chrysogenum against fungi common on dry-ripened meat products. Int. J. Food Microbiol. 205, 23–29. doi: 10.1016/j.ijfoodmicro.2015.03.029
Din, A. B., Specht, C. A., Robbins, P. W., and Yarden, O. (1996). chs-4, a class IV chitin synthase gene from Neurospora crassa. Mol. Gen. Genet. 250, 214–222. doi: 10.1007/BF02174181
Emri, T., Majoros, L., Toth, V., and Pócsi, I. (2013). Echinocandins: production and applications. Appl. Microbiol. Biotechnol. 97, 3267–3284. doi: 10.1007/s00253-013-4761-9
Fajardo-Somera, R. A., Jöhnk, B., Bayram, Ö., Valerius, O., Braus, G. H., and Riquelme, M. (2015). Dissecting the function of the different chitin synthases in vegetative growth and sexual development in Neurospora crassa. Fungal Genet. Biol. 75, 30–45. doi: 10.1016/j.fgb.2015.01.002
Fernandes, C., Gow, N. A. R., and Gonçalves, T. (2016). The importance of subclasses of chitin synthase enzymes with myosin-like domains for the fitness of fungi. Fungal Biol. Rev. 30, 1–14. doi: 10.1016/j.fbr.2016.03.002
Fisher, M. C., Hawkins, N. J., Sanglard, D., and Gurr, S. J. (2018). Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 360, 739–742. doi: 10.1126/science.aap7999
Fisher, M. C., Henk, D. A., Briggs, C. J., Brownstein, J. S., Madoff, L. C., McCraw, S. L., et al. (2012). Emerging fungal threats to animal, plant and ecosystem health. Nature 484, 186–194. doi: 10.1038/nature10947
Fujiwara, M., Ichinomiya, M., Motoyama, T., Horiuchi, H., Ohta, A., and Takagi, M. (2000). Evidence that the Aspergillus nidulans class I and class II chitin synthase genes, chsC and chsA, share critical roles in hyphal wall integrity and conidiophore development. J. Biochem. 127, 359–366. doi: 10.1093/oxfordjournals.jbchem.a022616
Galgóczy, L., and Marx, F. (2019). Do antimicrobial proteins contribute to overcoming the hidden antifungal crisis at the dawn of a post-antibiotic era? Microorganisms 7:16. doi: 10.3390/microorganisms7010016
Gandía, M., Garrigues, S., Hernanz-Koers, M., Manzanares, P., and Marcos, J. F. (2019). Differential roles, crosstalk and response to the antifungal protein AfpB in the three mitogen-activated protein kinases (MAPK) pathways of the citrus postharvest pathogen Penicillium digitatum. Fungal Genet. Biol. 124, 17–28. doi: 10.1016/j.fgb.2018.12.006
Gandía, M., Harries, E., and Marcos, J. F. (2012). Identification and characterization of chitin synthase genes in the postharvest citrus fruit pathogen Penicillium digitatum. Fungal Biol. 116, 654–664. doi: 10.1016/j.funbio.2012.03.005
Gandía, M., Harries, E., and Marcos, J. F. (2014). The myosin motor domain-containing chitin synthase PdChsVII is required for development, cell wall Integrity and virulence in the citrus postharvest pathogen Penicillium digitatum. Fungal Genet. Biol. 67, 58–70. doi: 10.1016/j.fgb.2014.04.002
Gandía, M., Xu, S., Font, C., and Marcos, J. F. (2016). Disruption of ku70 involved in non-homologous end-joining facilitates homologous recombination but increases temperature sensitivity in the phytopatogenic fungus Penicillium digitatum. Fungal Biol. 120, 317–323. doi: 10.1016/j.funbio.2015.11.001
Garrigues, S., Gandía, M., Castillo, L., Coca, M., Marx, F., Marcos, J. F., et al. (2018). Three antifungal proteins from Penicillium expansum: different patterns of production and antifungal activity. Front. Microbiol. 9:2370. doi: 10.3389/fmicb.2018.02370
Garrigues, S., Gandía, M., and Marcos, J. F. (2016). Occurrence and function of fungal antifungal proteins: a case study of the citrus postharvest pathogen Penicillium digitatum. Appl. Microbiol. Biotechnol. 100, 2243–2256. doi: 10.1007/s00253-015-7110-3
Garrigues, S., Gandía, M., Popa, C., Borics, A., Marx, F., Coca, M., et al. (2017). Efficient production and characterization of the novel and highly active antifungal protein AfpB from Penicillium digitatum. Sci. Rep. 7:14663. doi: 10.1038/s41598-017-15277-w
Geoghegan, I., Steinberg, G., and Gurr, S. (2017). The role of the fungal cell wall in the infection of plants. Trends Microbiol. 25, 957–967. doi: 10.1016/j.tim.2017.05.015
Gonçalves, I. R., Brouillet, S., Soulié, M.-C., Gribaldo, S., Sirven, C., Charron, N., et al. (2016). Genome-wide analyses of chitin synthases identify horizontal gene transfers towards bacteria and allow a robust and unifying classification into fungi. BMC Evol. Biol. 16:252. doi: 10.1186/s12862-016-0815-9
González-Candelas, L., Alamar, S., Sánchez-Torres, P., Zacarías, L., and Marcos, J. F. (2010). A transcriptomic approach highlights induction of secondary metabolism in citrus fruit in response to Penicillium digitatum infection. BMC Plant Biol. 10:194. doi: 10.1186/1471-2229-10-194
Hagen, S., Marx, F., Ram, A. F., and Meyer, V. (2007). The antifungal protein AFP from Aspergillus giganteus inhibits chitin synthesis in sensitive fungi. Appl. Environ. Microbiol. 73, 2128–2134. doi: 10.1128/AEM.02497-06
Harries, E., Gandia, M., Carmona, L., and Marcos, J. F. (2015). The Penicillium digitatum protein O-mannosyltransferase Pmt2 is required for cell wall integrity, conidiogenesis, virulence and sensitivity to the antifungal peptide PAF26. Mol. Plant Pathol. 16, 748–761. doi: 10.1111/mpp.12232
Hegedüs, N., and Marx, F. (2013). Antifungal proteins: more than antimicrobials? Fungal Biol. Rev. 26, 132–145. doi: 10.1016/j.fbr.2012.07.002
Huber, A., Hajdu, D., Bratschun-Khan, D., Gaspari, Z., Varbanov, M., Philippot, S., et al. (2018). New antimicrobial potential and structural properties of PAFB: a cationic, cysteine-rich protein from Penicillium chrysogenum Q176. Sci. Rep. 8:1751. doi: 10.1038/s41598-018-20002-2
Ichinomiya, M., Yamada, E., Yamashita, S., Ohta, A., and Horiuchi, H. (2005). Class I and class II chitin synthases are involved in septum formation in the filamentous fungus Aspergillus nidulans. Eukaryot. Cell 4, 1125–1136. doi: 10.1128/ec.4.6.1125-1136.2005
Jiménez-Ortigosa, C., Aimanianda, V., Muszkieta, L., Mouyna, I., Alsteens, D., Pire, S., et al. (2012). Chitin synthases with a myosin motor-like domain control the resistance of Aspergillus fumigatus to echinocandins. Antimicrob. Agents Chemother. 56, 6121–6131. doi: 10.1128/AAC.00752-12
Khang, C. H., Park, S. Y., Rho, H. S., Lee, Y. H., and Kang, S. (2006). Filamentous Fungi (Magnaporthe grisea and Fusarium oxysporum). Methods Mol. Biol. 344, 403–420. doi: 10.1385/1-59745-131-2:403
Kim, J. E., Lee, H. J., Lee, J. K., Kim, K. W., Yun, S. H., Shim, W. B., et al. (2009). Gibberella zeae chitin synthase genes, GzCHS5 and GzCHS7, are required for hyphal growth, perithecia formation, and pathogenicity. Curr. Genet. 55, 449–459. doi: 10.1007/s00294-009-0258-6
Kombrink, A., Sánchez-Vallet, A., and Thomma, B. P. H. J. (2011). The role of chitin detection in plant-pathogen interactions. Microbes Infect. 13, 1168–1176. doi: 10.1016/j.micinf.2011.07.010
Kong, L. A., Yang, J., Li, G. T., Qi, L. L., Zhang, Y. J., Wang, C. F., et al. (2012). Different chitin synthase genes are required for various developmental and plant infection processes in the rice blast fungus Magnaporthe oryzae. PLoS Pathog. 8:e1002526. doi: 10.1371/journal.ppat.1002526
Larson, T. M., Kendra, D. F., Busman, M., and Brown, D. W. (2011). Fusarium verticillioides chitin synthases CHS5 and CHS7 are required for normal growth and pathogenicity. Curr. Genet. 57, 177–189. doi: 10.1007/s00294-011-0334-6
Latgé, J. P. (2007). The cell wall: a carbohydrate armour for the fungal cell. Mol. Microbiol. 66, 279–290. doi: 10.1111/j.1365-2958.2007.05872.x
Latgé, J.-P., Beauvais, A., and Chamilos, G. (2017). The cell wall of the human fungal pathogen Aspergillus fumigatus: biosynthesis, organization, immune response, and virulence. Annu. Rev. Microbiol. 71, 99–116. doi: 10.1146/annurev-micro-030117-020406
Lee, M. J., and Sheppard, D. C. (2016). Recent advances in the understanding of the Aspergillus fumigatus cell wall. J. Microbiol. 54, 232–242. doi: 10.1007/s12275-016-6045-4
Lenardon, M. D., Munro, C. A., and Gow, N. A. R. (2010). Chitin synthesis and fungal pathogenesis. Curr. Opin. Microbiol. 13, 416–423. doi: 10.1016/j.mib.2010.05.002
Li, M., Jiang, C., Wang, Q., Zhao, Z., Jin, Q., Xu, J., et al. (2016). Evolution and functional insights of different ancestral orthologous clades of chitin synthase genes in the fungal tree of life. Front. Plant Sci. 7:37. doi: 10.3389/fpls.2016.00037
Liu, H. B., Kauffman, S., Becker, J. M., and Szaniszlo, P. J. (2004). Wangiella (Exophiala) dermatitidis WdChs5p, a class V chitin synthase, is essential for sustained cell growth at temperature of infection. Eukaryot. Cell 3, 40–51. doi: 10.1128/EC.3.1.40-51.2004
Liu, R. S., Huang, H., Yang, Q., and Liu, W. Y. (2002). Purification of alpha-sarcin and an antifungal protein from mold (Aspergillus giganteus) by chitin affinity chromatography. Protein Expr. Purif. 25, 50–58. doi: 10.1006/prep.2001.1608
Liu, Z., Zhang, X., Liu, X., Fu, C., Han, X., Yin, Y., et al. (2016). The chitin synthase FgChs2 and other FgChss co-regulate vegetative development and virulence in F. graminearum. Sci. Rep. 6:34975. doi: 10.1038/srep34975
Macarisin, D., Cohen, L., Eick, A., Rafael, G., Belausov, E., Wisniewski, M., et al. (2007). Penicillium digitatum suppresses production of hydrogen peroxide in host tissue during infection of citrus fruit. Phytopathology 97, 1491–1500. doi: 10.1094/PHYTO-97-11-1491
Madrid, M. P., Di Pietro, A., and Roncero, M. I. G. (2003). Class V chitin synthase determines pathogenesis in the vascular wilt fungus Fusarium oxysporum and mediates resistance to plant defence compounds. Mol. Microbiol. 47, 257–266. doi: 10.1046/j.1365-2958.2003.03299.x
Marcet-Houben, M., Ballester, A. R., de la Fuente, B., Harries, E., Marcos, J. F., González-Candelas, L., et al. (2012). Genome sequence of the necrotrophic fungus Penicillium digitatum, the main postharvest pathogen of citrus. BMC Genomics 13:646. doi: 10.1186/1471-2164-13-646
Marcos, J. F., and Gandía, M. (2009). Antimicrobial peptides: to membranes and beyond. Expert Opin. Drug Discov. 4, 659–671. doi: 10.1517/17460440902992888
Marcos, J. F., Muñoz, A., Pérez-Payá, E., Misra, S., and López-García, B. (2008). Identification and rational design of novel antimicrobial peptides for plant protection. Annu. Rev. Phytopathol. 46, 273–301. doi: 10.1146/annurev.phyto.121307.094843
Martín-Urdiroz, M., Martínez-Rocha, A. L., Di Pietro, A., Martínez del Pozo, A., and Roncero, M. I. G. (2009). Differential toxicity of antifungal protein AFP against mutants of Fusarium oxysporum. Int. Microbiol. 12, 115–121. doi: 10.2436/20.1501.01.88
Martín-Urdíroz, M., Roncero, M. I., González-Reyes, J. A., and Ruiz-Roldán, C. (2008). ChsVb, a class VII chitin synthase involved in septation, is critical for pathogenicity in Fusarium oxysporum Eukaryot. Cell 7, 112–121. doi: 10.1128/EC.00347-07
Marx, F., Binder, U., Leiter, É, and Pócsi, I. (2008). The Penicillium chrysogenum antifungal protein PAF, a promising tool for the development of new antifungal therapies and fungal cell biology studies. Cell Mol. Life Sci. 65, 445–454. doi: 10.1007/s00018-007-7364-8
Mellado, E., AufauvreBrown, A., Gow, N. A. R., and Holden, D. W. (1996). The Aspergillus fumigatus chsC and chsG genes encode Class III chitin synthases with different functions. Mol. Microbiol. 20, 667–679. doi: 10.1046/j.1365-2958.1996.5571084.x
Meyer, V. (2008). A small protein that fights fungi: AFP as a new promising antifungal agent of biotechnological value. Appl. Microbiol. Biotechnol. 78, 17–28. doi: 10.1007/s00253-007-1291-3
Morcx, S., Kunz, C., Choquer, M., Assie, S., Blondet, E., Simond-Côte, E., et al. (2013). Disruption of Bcchs4, Bcchs6 or Bcchs7 chitin synthase genes in Botrytis cinerea and the essential role of class VI chitin synthase (Bcchs6). Fungal Genet. Biol. 52, 1–8. doi: 10.1016/j.fgb.2012.11.011
Motoyama, T., Fujiwara, M., Kojima, N., Horiuchi, H., Ohta, A., and Takagi, M. (1997). The Aspergillus nidulans genes chsA and chsD encode chitin synthases which have redundant functions in conidia formation. Mol. Gen. Genet. 253, 520–528. doi: 10.1007/s004380050353
Muñoz, A., Gandía, M., Harries, E., Carmona, L., Read, N. D., and Marcos, J. F. (2013). Understanding the mechanism of action of cell-penetrating antifungal peptides using the rationally designed hexapeptide PAF26 as a model. Fungal Biol. Rev. 26, 146–155. doi: 10.1016/j.fbr.2012.10.003
Munro, C. A. (2013). “Chitin and glucan, the yin and yang of the fungal cell wall, implications for antifungal drug discovery and therapy,” in Advances in Applied Microbiology, eds S. Sima, and M. G. Geoffrey, (Waltham, MA: Academic Press), 145–172. doi: 10.1016/b978-0-12-407678-5.00004-0
Munro, C. A., and Gow, N. A. (2001). Chitin synthesis in human pathogenic fungi. Med. Mycol. 39(Suppl. 1), 41–53. doi: 10.1080/714030998
Munro, C. A., Selvaggini, S., De Bruijn, I., Walker, L., Lenardon, M. D., Gerssen, B., et al. (2007). The PKC, HOG and Ca2+ signalling pathways co-ordinately regulate chitin synthesis in Candida albicans. Mol. Microbiol. 63, 1399–1413. doi: 10.1111/j.1365-2958.2007.05588.x
Muszkieta, L., Aimanianda, V., Mellado, E., Gribaldo, S., Alcàzar-Fuoli, L., Szewczyk, E., et al. (2014). Deciphering the role of the chitin synthase families 1 and 2 in the in vivo and in vitro growth of Aspergillus fumigatus by multiple gene targeting deletion. Cell. Microbiol. 16, 1784–1805. doi: 10.1111/cmi.12326
Oberparleiter, C., Kaiserer, L., Haas, H., Ladurner, P., Andratsch, M., and Marx, F. (2003). Active internalization of the Penicillium chrysogenum antifungal protein PAF in sensitive Aspergilli Antimicrob. Agents Chemother. 47, 3598–3601. doi: 10.1128/aac.47.11.3598-3601.2003
Ouedraogo, J. P., Hagen, S., Spielvogel, A., Engelhardt, S., and Meyer, V. (2011). Survival strategies of yeast and filamentous fungi against the antifungal protein AFP. J. Biol. Chem. 286, 13859–13868. doi: 10.1074/jbc.M110.203588
Palou, L. (2014). “Penicillium digitatum, Penicillium italicum (Green Mold, Blue Mold),” in Postharvest Decay: Control Strategies, ed. S. Bautista-Baños, (London: Elsevier), 45–102. doi: 10.1016/b978-0-12-411552-1.00002-8
Perfect, J. R. (2016). Is there an emerging need for new antifungals? Expert Opin. Emerg. Drugs 21, 129–131. doi: 10.1517/14728214.2016.1155554
Plaza, P., Usall, J., Teixido, N., and Vinas, I. (2003). Effect of water activity and temperature on germination and growth of Penicillium digitatum, P. italicum and Geotrichum candidum. J. Appl. Microbiol. 94, 549–554. doi: 10.1046/j.1365-2672.2003.01909.x
Roncero, C., Sánchez-Díaz, A., and Valdivieso, M. H. (2016). “Chitin synthesis and fungal cell morphogenesis,” in Biochemistry and Molecular Biology, ed. D. Hoffmeister, (New York, NY: Springer International Publishing), 167–190. doi: 10.1007/978-3-319-27790-5_9
Sánchez-Vallet, A., Mesters, J. R., and Thomma, B. P. H. J. (2014). The battle for chitin recognition in plant-microbe interactions. FEMS Microbiol. Rev. 39, 171–183. doi: 10.1093/femsre/fuu003
Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T., et al. (2012). Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682. doi: 10.1038/nmeth.2019
Schuster, M., Martin-Urdiroz, M., Higuchi, Y., Hacker, C., Kilaru, S., Gurr, S. J., et al. (2016). Co-delivery of cell-wall-forming enzymes in the same vesicle for coordinated fungal cell wall formation. Nat. Microbiol. 1:16149. doi: 10.1038/nmicrobiol.2016.149
Schuster, M., Treitschke, S., Kilaru, S., Molloy, J., Harmer, N. J., and Steinberg, G. (2012). Myosin-5, kinesin-1 and myosin-17 cooperate in secretion of fungal chitin synthase. EMBO J. 31, 214–227. doi: 10.1038/emboj.2011.361
Sheng, W., Yamashita, S., Ohta, A., and Horiuchi, H. (2013). Functional differentiation of chitin synthases in Yarrowia lipolytica. Biosci. Biotechnol. Biochem. 77, 1275–1281. doi: 10.1271/bbb.130111
Shi, X., Cordero, T., Garrigues, S., Marcos, J. F., Daròs, J. A., and Coca, M. (2019). Efficient production of antifungal proteins in plants using a new transient expression vector derived from tobacco mosaic virus. Plant Biotechnol. J. 17, 1069–1080. doi: 10.1111/pbi.13038
Sonderegger, C., Fizil, Á., Burtscher, L., Hajdu, D., Muñoz, A., Gáspári, Z., et al. (2017). D19S mutation of the cationic, cysteine-rich protein PAF: novel insights into its structural dynamics, thermal unfolding and antifungal function. PLoS One 12:e0169920. doi: 10.1371/journal.pone.0169920
Steinberg, G. (2011). Motors in fungal morphogenesis: cooperation versus competition. Curr. Opin. Microbiol. 14, 660–667. doi: 10.1016/j.mib.2011.09.013
Szewczyk, E., Nayak, T., Oakley, C. E., Edgerton, H., Xiong, Y., Taheri-Talesh, N., et al. (2006). Fusion PCR and gene targeting in Aspergillus nidulans. Nat. Protoc. 1, 3111–3120. doi: 10.1038/nprot.2006.405
Takeshita, N., Wernet, V., Tsuizaki, M., Grun, N., Hoshi, H. O., Ohta, A., et al. (2015). Transportation of Aspergillus nidulans Class III and V Chitin synthases to the hyphal tips depends on conventional kinesin. PLoS One 10:e0125937. doi: 10.1371/journal.pone.0125937
Takeshita, N., Yamashita, S., Ohta, A., and Horiuchi, H. (2006). Aspergillus nidulans class V and VI chitin synthases CsmA and CsmB, each with a myosin motor-like domain, perform compensatory functions that are essential for hyphal tip growth. Mol. Microbiol. 59, 1380–1394. doi: 10.1111/j.1365-2958.2006.05030.x
Theis, T., Marx, F., Salvenmoser, W., Stahl, U., and Meyer, V. (2005). New insights into the target site and mode of action of the antifungal protein of Aspergillus giganteus. Res. Microbiol. 156, 47–56. doi: 10.1016/j.resmic.2004.08.006
Thomma, B. P. H. J., Nurnberger, T., and Joosten, M. H. A. J. (2011). Of PAMPs and effectors: the blurred PTI-ETI dichotomy. Plant Cell 23, 4–15. doi: 10.1105/tpc.110.082602
Treitschke, S., Doehlemann, G., Schuster, M., and Steinberg, G. (2010). The Myosin motor domain of fungal chitin synthase V is dispensable for vesicle motility but required for virulence of the maize pathogen Ustilago maydis. Plant Cell 22, 2476–2494. doi: 10.1105/tpc.110.075028
Walker, L. A., Lee, K. K., Munro, C. A., and Gow, N. A. R. (2015). Caspofungin Treatment of Aspergillus fumigatus results in ChsG-dependent upregulation of chitin synthesis and the formation of chitin-rich microcolonies. Antimicrob. Agents Chemother. 59, 5932–5941. doi: 10.1128/aac.00862-15
Weber, I., Assmann, D., Thines, E., and Steinberg, G. (2006). Polar localizing Class V myosin chitin synthases are essential during early plant infection in the plant pathogenic fungus Ustilago maydis. Plant Cell 18, 225–242. doi: 10.1105/tpc.105.037341
Zhang, Y.-Z., Chen, Q., Liu, C.-H., Liu, Y.-B., Yi, P., Niu, K.-X., et al. (2016). Chitin synthase gene FgCHS8 affects virulence and fungal cell wall sensitivity to environmental stress in Fusarium graminearum. Fungal Biol. 120, 764–774. doi: 10.1016/j.funbio.2016.02.002
Zheng, L., Mendoza, L., Wang, Z., Liu, H., Park, C., Kauffman, S., et al. (2006). WdChs1p, a class II chitin synthase, is more responsible than WdChs2p (Class I) for normal yeast reproductive growth in the polymorphic, pathogenic fungus Wangiella (Exophiala) dermatitidis. Arch. Microbiol. 185, 316–329. doi: 10.1007/s00203-006-0101-0
Keywords: chitin, cell wall, antifungal proteins, conidia, Penicillium digitatum, virulence, postharvest
Citation: Gandía M, Garrigues S, Bolós B, Manzanares P and Marcos JF (2019) The Myosin Motor Domain-Containing Chitin Synthases Are Involved in Cell Wall Integrity and Sensitivity to Antifungal Proteins in Penicillium digitatum. Front. Microbiol. 10:2400. doi: 10.3389/fmicb.2019.02400
Received: 18 July 2019; Accepted: 04 October 2019;
Published: 18 October 2019.
Edited by:
Frank Ebel, Ludwig Maximilian University of Munich, GermanyReviewed by:
Meritxell Riquelme, Ensenada Center for Scientific Research and Higher Education (CICESE), MexicoVishukumar Aimanianda, Institut Pasteur, France
Copyright © 2019 Gandía, Garrigues, Bolós, Manzanares and Marcos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mónica Gandía, bWdhbmRpYUBpYXRhLmNzaWMuZXM=
†Present address: Sandra Garrigues, Fungal Physiology, Westerdijk Fungal Biodiversity Institute and Fungal Molecular Physiology, Utrecht University, Utrecht, Netherlands