 Samuele Peppoloni1†
Samuele Peppoloni1† Eva Pericolini1†
Eva Pericolini1† Bruna Colombari1
Bruna Colombari1 Diego Pinetti2Claudio Cermelli1
Diego Pinetti2Claudio Cermelli1 Francesco Fini3Fabio Prati3
Francesco Fini3Fabio Prati3 Emilia Caselli3*
Emilia Caselli3* Elisabetta Blasi1*
Elisabetta Blasi1*- 1Department of Surgical, Medical, Dental and Morphological Sciences With Interest in Transplant, Oncological and Regenerative Medicine, University of Modena and Reggio Emilia, Modena, Italy
- 2Centro Interdipartimentale “Grandi Strumenti” (CIGS), University of Modena and Reggio Emilia, Modena, Italy
- 3Department of Life Sciences, University of Modena and Reggio Emilia, Modena, Italy
Pseudomonas aeruginosa is a Gram-negative nosocomial pathogen, often causative agent of severe device-related infections, given its great capacity to form biofilm. P. aeruginosa finely regulates the expression of numerous virulence factors, including biofilm production, by Quorum Sensing (QS), a cell-to-cell communication mechanism used by many bacteria. Selective inhibition of QS-controlled pathogenicity without affecting bacterial growth may represent a novel promising strategy to overcome the well-known and widespread drug resistance of P. aeruginosa. In this study, we investigated the effects of SM23, a boronic acid derivate specifically designed as β-lactamase inhibitor, on biofilm formation and virulence factors production by P. aeruginosa. Our results indicated that SM23: (1) inhibited biofilm development and production of several virulence factors, such as pyoverdine, elastase, and pyocyanin, without affecting bacterial growth; (2) decreased the levels of 3-oxo-C12-HSL and C4-HSL, two QS-related autoinducer molecules, in line with a dampened lasR/lasI system; (3) failed to bind to bacterial cells that had been preincubated with P. aeruginosa-conditioned medium; and (4) reduced both biofilm formation and pyoverdine production by P. aeruginosa onto endotracheal tubes, as assessed by a new in vitro model closely mimicking clinical settings. Taken together, our results indicate that, besides inhibiting β-lactamase, SM23 can also act as powerful inhibitor of P. aeruginosa biofilm, suggesting that it may have a potential application in the prevention and treatment of biofilm-associated P. aeruginosa infections.
Introduction
Pseudomonas aeruginosa is a Gram-negative opportunistic pathogen, causing nosocomial infections in more than 2 million patients every year (Cross et al., 1983; Rossolini and Mantengoli, 2005). Such infections are particularly frequent in immunocompromised patients, such as those with cancer, AIDS, burn wounds, and indwelling devices (Vandeputte et al., 2010; Sarabhai et al., 2015). Notably, P. aeruginosa infections are associated with unfavorable outcome in individuals with cystic fibrosis (CF), the most common life-limiting genetic disorder in the U.S. (Lyczak et al., 2002). These patients, indeed, show abnormalities in their lung tissue that promote bacterial colonization, which in turn causes long-lasting inflammation, lung injury, and eventually respiratory failure.
P. aeruginosa has a uniquely large genome, whose genes encode for several virulence factors (including LasA protease, LasB elastase, pyoverdine, pyocyanin, alginate, and exotoxin A) and regulatory mechanisms, allowing microbial adaptation to many hostile environments (Stover et al., 2000). As a consequence, P. aeruginosa is ubiquitous in nature and almost impossible to eliminate from hospitals. As many other microorganisms, besides living in a planktonic form, P. aeruginosa is able to form biofilm on medical implants or damaged tissues (Xu et al., 2013). Particularly, more than 70% of hospital-acquired infections are associated with biofilm on catheters, ventilator tubes, implants, and medical prosthetic devices (Brooun et al., 2000). Unlike planktonic cell counterpart, bacterial communities structured as biofilms exhibit an altered phenotype, with respect to growth rate, expression of virulence factors, and cell-to-cell communication system (Qu et al., 2016). Thus, when enclosed in a self-produced extracellular polymeric matrix, P. aeruginosa is protected from patient’s immune system and becomes up to 1,000 times more antibiotic resistant than the planktonic counterpart (Mah and O'Toole, 2001).
The expression of many virulence factors in P. aeruginosa is controlled by the Quorum Sensing (QS) system (Venturi, 2006), an intercellular communication mechanism that coordinates bacterial behavior and gene expression by means of signaling molecules in a cell density-dependent manner (Schuster and Greenberg, 2006). The transcriptional regulation of numerous virulence genes in Pseudomonas is under the control of two N-acyl homoserine lactone (AHL)-dependent QS systems, named lasI/lasR and rhlI/rhlR. In the lasI/lasR system, lasI encodes the synthesis of N-(3-oxo-dodecanoyl)-homoserine lactone (3-oxo-C12-HSL), which binds and activates the cognate response regulator LasR, leading to regulation of gene expression. Differently, in the rhlI/rhlR system, rhlI synthesizes the N-(butanoyl)-homoserine lactone (C4-HSL), which, in turn, by interacting with the cognate RhlR, influences the transcription of target genes. Importantly, the two QS systems are hierarchically organized, with the lasI/lasR system in turn regulating the transcription of rhlI/rhlR.
These QS systems are among the most studied in bacteria and their regulons are essential for the pathogenicity of P. aeruginosa (Schuster and Greenberg, 2006; Girard and Bloemberg, 2008; Zou and Nair, 2009). There is also a third self-inducing signal, referred to as Pseudomonas quinolone signal, that plays an integral role in the QS system (Pesci et al., 1999). Such complicated communication systems coordinate/regulate several virulence traits in P. aeruginosa, including motility, toxin production, and biofilm formation (Fuqua and Greenberg, 1998; de Kievit and Iglewski, 2000; Dong et al., 2001; Donabedian, 2003; Jakobsen et al., 2013). Recently, a fourth intercellular communication signal has been discovered, as being capable of integrating environmental stress cues with the QS network. Named as IQS, it belongs to a new class of QS signal molecules and has structurally been established to be 2-(2-hydroxyphenyl)-thiazole-4-carbaldehyde (Lee and Zhang, 2015).
The difficulty in treating biofilm-associated P. aeruginosa infections has encouraged the extensive use of antibiotics, in turn facilitating the development of multiple drug-resistant strains. Among several mechanisms of antibiotic resistance, the production of β-lactamases, enzymes able to hydrolyze β-lactams, is consistently the most concerning one (Bush and Bradford, 2016). In Pseudomonas, class C β-lactamases confer high level of resistance to penicillins, cephalosporins, and monobactams (Berrazeg et al., 2015). Besides modifying the structure of β-lactams, leading to generation of several new penicillins and cephalosporins, another relevant strategy to overcome resistance to these drugs is the co-administration of the β-lactam antibiotic together with a β-lactamase inhibitor (Bush and Bradford, 2019). Accordingly, in the last decade, new classes of β-lactamase inhibitors have entered the market giving the opportunity to restore the activity of several β-lactams (Bush and Bradford, 2019). One of the most promising class of new β-lactamase inhibitors are the boronic acids transition state analog inhibitors (BATSIs), which are known to restore the β-lactam activity both in vitro and in vivo (Drawz et al., 2011; Eidam et al., 2012; Barnes et al., 2018). Recently, a new combination of a boronic acid and the β-lactam meropenem (Vabomere®) has entered the market for treatment of infections caused by carbapenem-resistant Enterobacteriaceae and multidrug-resistant (MDR) Pseudomonas (Griffith et al., 2019). In order to obtain highly active BATSIs, we have undertaken a biomimetic approach: the boronic acid structure was decorated with chemical groups arranged in a specific stereochemistry resembling that of the natural substrate β-lactam. Among different synthesized BATSIs, one of the best inhibitors ever tested was the SM23 (Morandi et al., 2003). This compound has an R1 acylamino side chain of the β-lactam cephalothin on the boron carbon α (red colored in Supplementary Material), and a meta-carboxybenzyl side chain, that mimics the 6-carboxydihydrothiazine ring of the antibiotic (blue colored in Supplementary Material). Few commercially available phenylboronic acids have also been investigated in Vibrio harveyi, where they act as QS inhibitors with IC50 values in the low to sub-micromolar range (Ni et al., 2009).
In this study, we investigated the effects of the β-lactamase inhibitor boronic acid derivative SM23 on biofilm formation and production of QS-dependent virulence factors as well as autoinducer molecules by P. aeruginosa during biofilm formation. Overall, our results provide the first evidence on the efficacy of SM23 as a remarkable anti-biofilm agent and QS inhibitor, envisaging its use in the prevention and treatment of biofilm-associated P. aeruginosa infections.
Materials and Methods
SM23
The boronic acid SM23 was synthesized as previously described (Morandi et al., 2003). The compound was dissolved in DMSO and the different working concentrations were prepared by diluting each stock solution in PBS. In all experiments, the final DMSO concentration was less than 0.1%.
Microbial Strain
We used the bioluminescent Pseudomonas aeruginosa strain (P1242), previously engineered to express both the luciferase gene and substrate under the control of a constitutive P1 integron promoter, in order to constitutively produce a detectable bioluminescent signal (Choi and Schweizer, 2006).
Bacterial cells from −80°C glycerol stocks were initially seeded onto Tryptic Soy Agar (TSA) plates and incubated overnight at 37°C; isolated colonies were then collected, added to 10 ml of Tryptic Soil Broth (TSB), and allowed to grow overnight at 37°C with gentle shaking. Bacterial concentrations were assessed by the McFarland standard curve and diluted to the required experimental concentration.
Biofilm Formation and Quantification
To investigate the effect of SM23 on Pseudomonas biofilm formation, a bacterial cell suspension (108 cells/ml in TSB plus 2% sucrose) was seeded (100 μl/well) in a 96-well microtiter plate (Sarstedt, Nümbrecht, Germany), treated or not with 100 μl of scalar doses of SM23 (range concentrations from 0.390 to 25.0 μM) and incubated for 24 h at 37°C to allow biofilm formation. After incubation, each well was gently washed to remove planktonic cells, then crystal violet (CV) staining was performed to quantify biofilm formation as previously described (Stepanovic et al., 2000). The absorbance at 570 nm was spectrophotometrically measured by the SunRise Microplate Reader (Tecan Group Ltd., Männedorf Switzerland). The results were expressed as optical density (OD570) mean ± SEM of the biofilm biomass. In order to evaluate the metabolic activity of Pseudomonas biofilm, 100 μl of bacterial culture in TSB + 2% sucrose (5 × 104 cells/ml) were seeded (100 μl/well) in a 96-black well microtiter plate, treated or not with scalar doses of SM23 at concentrations ranging from 0.390 to 25.0 μM (100 μl/well) and then incubated for 24 h at 37°C to allow biofilm formation. After incubation, each well was gently washed twice with PBS (EuroClone, Whethereby, UK) to remove planktonic cells and then bioluminescence signal was measured by Viktor Luminescence reader (Perkin Elmer). The results were expressed as Relative Luminescence Units (RLU) mean ± SEM of metabolically active biofilm.
Confocal Microscopy Analysis of P. aeruginosa Biofilm
The fluorescence property of Pseudomonas spp. (Meyer, 2000; Cornelis and Matthijs, 2002) was exploited to perform confocal microscopy imaging. Briefly, bacterial suspensions (1 × 105 cells/ml in TSB plus 2% sucrose) were seeded on coverslips (1,000 μl/well) inserted into 24-microplate (Sarstedt, Nümbrecht Germany) and treated or not with SM23 at 0.780 and 3.125 μM. The plates were then incubated for 24 h at 37°C to allow biofilm formation. After incubation, the coverslips were washed twice with PBS, fixed with 4% paraformaldehyde (PFA) (Sigma-Aldrich, Darmstadt) for 30 min at 4°C, washed again and then analyzed by confocal microscope Leica TCS SP8 (Wetzlar, Germany) at excitation/emission wavelength 492/517 nm.
Evaluation of Live or Dead Bacterial Cells in Biofilm
In order to evaluate the live or dead cells in Pseudomonas biofilm, the bacterial cells (1 × 108 cells/ml) were seeded in 96-well black-plates and treated or not with 0.780 and 3.125 μM of SM23 for 24 h at 37°C to allow biofilm formation. After incubation, the samples were stained with the “live/dead cells stain kit” (Thermo Fisher Scientific, Waltham, Massachusetts, USA), using 5(6)-carboxyfluorescein diacetate (CFDA) to label alive cells (30 min at 37°C plus 5% CO2) and propidium iodide (PI) to stain the dead cells (15 min at 37°C plus 5% CO2). The staining procedure was conducted according to the manufacturer’s instructions. After incubation, the samples were washed twice with PBS and the fluorescence emission (CFDA excitation/emission: 485/528; PI excitation/emission: 528/645) was analyzed using a multi-well fluorescence plate reader (Synergy HTX, BIOTEK, Winooski, Vermont, USA). The results were expressed as Relative Fluorescence Units (RFU) mean ± SEM of alive/dead biofilm cells.
Elastase Activity
Elastase activity was measured in cell-free supernatants from SM23 Pseudomonas cells, treated or not with the compound (concentration range from 0.780 to 3.125 μM), during biofilm formation. After 6, 12, and 24 h of culture, the elastase activity was measured as described by (Ohman et al., 1980) using Elastin-Congo Red (ECR) (Sigma, St. Louis, USA) as a substrate. Briefly, 100 μl of untreated or SM23-treated supernatants were mixed with 900 μl of ECR buffer (100 mM Tris, 1 mM CaCl2, pH 7.5) containing 20 mg of ECR and then incubated for 3 h at 37°C. The reaction was terminated by adding 1 ml of 0.7 M sodium phosphate buffer (pH 6.0) and the tubes were placed in cold water bath. The insoluble ECR was removed by centrifugation at 10,000 rpm for 10 min and then the absorbance was measured at 495 nm by a SunRise Microplate Reader. The elastase activity was expressed as the optical density (OD495) mean ± SEM.
High-Performance Liquid Chromatography-Mass Spectrometric Analysis
Pyoverdine, pyocyanin, 3-oxo-C12-HSL and C4-HSL molecules were assessed in culture supernatants of P. aeruginosa during biofilm formation after 6, 12, and 24 h of SM23 treatment. All supernatants for the high-performance liquid chromatography-mass spectrometric (HPLC-MS) analysis were filtered on Amicon Ultra-0.5 10 K centrifugal filter devices and 1:5 diluted with 5% methanol - 0.2% formic acid in MilliQ water. The HPLC-MS instrument used was an UltiMate 3,000 system, consisting of an online degasser, a Binary Pump HPG 3400RS, a Well Plate Autosampler WPS 3000RS, and a Thermostatted Column Compartment TCC 3000RS coupled to a Q-Exactive hybrid quadrupole – orbitrap mass analyzer via a HESI-II heated electrospray ion source (Thermo Scientific). Chromatographic separation of a 5 μl sample injection was performed on a Poroshell 120 SB-C18 100 × 2.1 mm ID, 2.7 μm ps column (Agilent) at 30°C and a 0.4 ml/min flow rate. A linear gradient elution scheme was used with mobile phase components being 0.1% formic acid in water (A) and methanol (B). The gradient started at 2% B which was maintained for 0.5 min, then raised up to 30% B in 30 min, and up again to 98% B in 24.5 min. The column was then kept at 98% B for 17.9 min, then starting conditions were restored in 0.1 min and maintained for 19 min pending a successive injection. Electrospray ionization was operated in positive ion mode, using nitrogen as sheath gas (50 arbitrary units), auxiliary gas (290°C, 40 arbitrary units), and sweep gas (3 arbitrary units). The sprayer voltage was kept at 3.8 kV and the transfer capillary temperature was set at 320°C. The Q-Exactive was operated in Full MS/dd-MS2 mode. The Full MS scan range was set from m/z 170 to 1,000 at 70,000 FWHM resolution (m/z 200). The automatic gain control (AGC) target was set at 1.0 × 106 with a maximum injection time (IT) of 200 ms. Data-dependent MS2 (dd-MS2) acquisitions at 17,500 FWHM resolution (m/z 200) were triggered for the Top 3 precursor ions following each Full MS scan. The intensity threshold for precursor ion selection was set to 1.0 × 105, then dynamic exclusion was active for 20.0 s. AGC target and maximum IT for the MS2 experiments were set to 2.0 × 105 and 50 ms. Each precursor ion was fragmented using stepped normalized collision energy (NCE) values at 28, 50, and 75.
lasI/lasR Gene Transcription Analysis
A qRT-PCR was used to investigate SM23-induced changes in transcription levels of selected QS genes. Briefly, Pseudomonas cells (1 × 107 cells/ml) were seeded in 24-well flat-bottom plates (Becton Dickinson Labware Europe, Meylan Cedex, France) and untreated or treated with SM23 (1.56 μM) for 24 h at 37°C to allow biofilm formation. After incubation, each well was washed twice with PBS to remove the no-adherent bacterial cells; then, biofilm was recovered by scraping and subsequent centrifugation at +4°C; the pellet was dried and stocked at −80°C until qRT-PCR analysis. Total RNA was extracted using the HiPurA Bacterial RNA Purification (Himedia) kit and treated with DNase I on a column to remove the DNA. RNA was quantified by MaestroNano spectrophotometer readings for microvolumes. Two micrograms of total RNA and random primers were used for cDNA synthesis using EasyScript cDNA Synthesis (Abm) kit. Expression of lasI, lasR, and 16S rRNA genes was performed in triplicate by real-time PCR using 20 ng of cDNA, 0.5 μM of forward and reverse primers and BrightGreen 2x qPCR MasterMix (Abm). The primers list is shown in Supplementary Material. The amplification conditions were the following: 1 cycle for 10 min at 95°C; 40 cycles for 15 s at 95°C, 1 min at 60°C; melting curve for 30 s at 95°C, 30 s at 65°C, and 30 s at 95°C. The expression levels of the target genes in treated samples were normalized to the expression of the reference 16S rRNA gene and then compared to the control samples. The relative expression levels were calculated using the 2−ΔΔCt method.
SM23 Interaction With Bacterial Cells
Two experimental protocols, A and B, were used. Bacterial cell cultures (1 × 108 cells/ml; 200 μl/well) were seeded in a 96-black microtiter plate and incubated for 7 h at 37°C and 5% CO2, in the presence of SM23 (12.5 μM), as illustrated by the time-line of protocol A (Figure 1). In parallel experiments, microbial cells were seeded in a 96-black well microtiter plate and incubated for 5 h at 37°C and 5% CO2; then, SM23 was added to the cells and the plates were further incubated for 2 h, according to the time-line of protocol B (Figure 1).
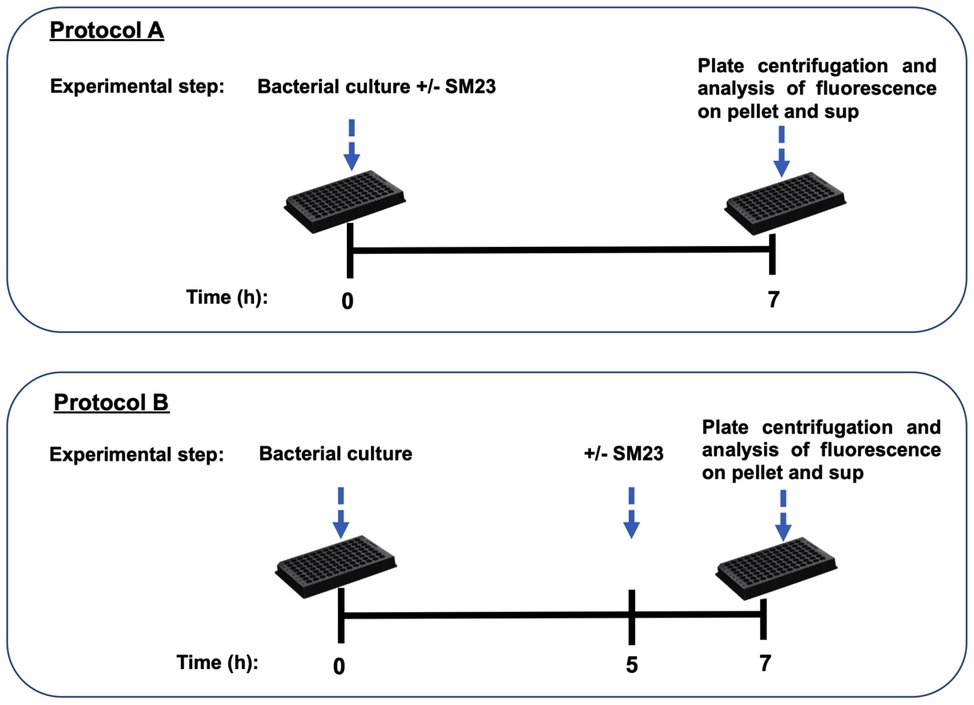
Figure 1. Time-lines of the experimental protocols used (protocol A and protocol B), as detailed in section “Materials and Methods.”
In selected experiments, fresh bacterial cells were suspended at the concentration of 1 × 108 cells/ml and pre-treated for 30 min with cell-free supernatants (P. aeruginosa-conditioned medium, recovered from an overnight culture of Pseudomonas at the starting concentration of 1 × 108 cells/ml). To exclude residual viable bacteria in such P. aeruginosa-conditioned medium, 50 μl of the supernatants was seeded onto TSA plates and incubated for 48 h at 37°C under aerobic conditions; no bacterial CFUs on TSA plates were ever observed. Bacterial cells were then incubated for 7 h with the SM23 (12.5 μM), as described in protocol A. Control wells (namely, medium alone, medium plus SM23, and untreated microbial culture) were also included in the assay to evaluate the SM23 fluorescent signal. In all the experimental protocols, at the end of the incubation period (7 h), the plates were centrifuged for 10 min at 5,000 rpm. The recovered supernatants were then collected, transferred in new wells, and the fluorescent signal in both pellets and supernatants was measured by a multi-well fluorescence plate reader (Synergy HTX, BIOTEK, Winooski, Vermont, USA). The residual fluorescence of SM23 in the cell pellets and cell-free supernatants was defined as the fluorescence values obtained by subtracting, respectively, the fluorescence of the pellet or supernatant, observed in the untreated control groups, from the fluorescence values of the corresponding treated samples. The resulting values were expressed as % of residual fluorescence of SM23 detected in the pellet versus the supernatant.
Biofilm Formation on Endotracheal Tubes and Quantification of Pyoverdine Release
Two-hundred microliter of overnight culture of Pseudomonas (5 × 104/ml) in TSB with 2% sucrose were seeded in 96-well black-plates, containing 1 endotracheal tube (ETT) piece/well; the plates were then incubated for 90 min at 37°C (adhesion period). After incubation, the ETT pieces were washed twice with PBS, transferred into new wells, and incubated for further 24 h at 37°C in fresh TSB medium in the presence of SM23 (1.560 and 3.150 μM) to allow biofilm formation. At the end of incubation, the ETT pieces were removed, washed twice with PBS, and the bioluminescence signal was measured by Viktor Luminescence reader (Perkin Elmer). The values were expressed as Relative Luminescence Units (RLU). In parallel, after removing ETT pieces, the supernatants were recovered and centrifuged twice (10,000 rpm for 15 min) to remove the remaining bacteria. Pyoverdine release was quantified in 100 μl of culture supernatants and fluorescence emission was measured with a multi-well fluorescence plate reader (excitation/emission: 360/460), according to a standard protocol (Das et al., 2016). The values were expressed as RFU.
Statistical Analysis
Quantitative variables were tested for normal distribution. Statistical differences between groups were analyzed according to one-way ANOVA or Kruskal-Wallis followed by Dunnett’s multiple comparisons tests by using GraphPad prism 8. A value of p ≤ 0.05 was considered significant.
Results
Inhibitory Effects of SM23 on Pseudomonas aeruginosa Biofilm Formation
Dose-dependent experiments were performed to assess the ability of SM23 to interfere with the biofilm formation by P. aeruginosa. Microbial cells were grown in a 96-well microtiter plate in medium only or in the presence of scalar doses of SM23 and then incubated for 24 h to allow biofilm formation. After incubation, each well was gently washed to remove planktonic cells and then CV staining was performed to quantify the biofilm biomass. The results shown in Figure 2 (left panel) indicated that the SM23, at the dose of 0.390 μM, was already able to significantly inhibit biofilm formation (about 40% of biofilm biomass), achieving its greatest effect in the dose range of 0.780–6.250 μM (about 50% of biomass reduction observed). Next, we analyzed the inhibitory effect of SM23 on biofilm, exploiting the bioluminescent properties of P. aeruginosa strain P1242, a model that allows to evaluate the effects of a given compound directly on the metabolically active biofilm (Pericolini et al., 2018). Thus, bacterial biofilm was allowed to form in 96-black well microtiter plates without or with SM23 at different concentrations. After 24 h of incubation, each well was gently washed to remove planktonic cells and, then, the RLU emitted by the metabolically active bacteria embedded in biofilm were measured. Results in Figure 2 (right panel) indicated that SM23 significantly reduced biofilm formation in a dose-dependent manner (from 0.390 to 25 μM concentration). In particular, 1.560 and 3.125 μM were the lowest SM23 doses still capable of reducing biofilm by 50–60%. Thus, from now on, we mainly used these two concentrations.
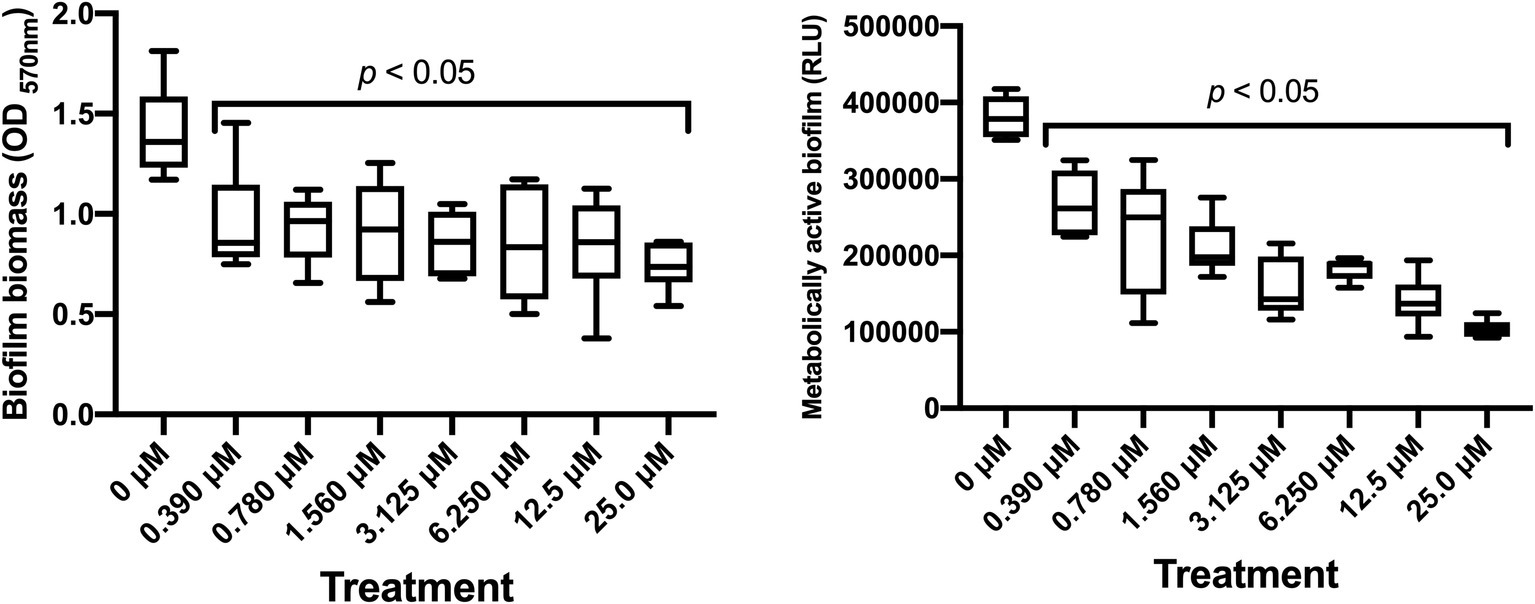
Figure 2. Dose-dependent effect of SM23 on P. aeruginosa biofilm formation. Both box-plot graphs show the mean ± SEM of microbial biofilm produced after 24 h of incubation in medium or in the presence of scalar doses of SM23. Left panel: mean ± SEM of the OD570nm (biofilm biomass) of triplicate samples from two different experiments. Right panel: mean ± SEM of the Relative Luminescence Units (RLU) of triplicate samples from two different experiments. p < 0.05; SM23-treated vs. untreated according to one-way ANOVA followed by Dunnett’s multiple comparisons test.
In parallel experiments, the influence of SM23 was evaluated on the growth of P. aeruginosa, cultured in planktonic form for 7 and 24 h. We found that SM23 failed to induce any change on bacterial growth capacity and viability as well, up to the concentration of 25.0 μM; also, the bacterial doubling times were found to be comparable, in SM23-treated and untreated controls (data not shown).
Morphological Observation of SM23-Treated Biofilm
We next performed confocal analysis in a 24 h-old biofilm, produced in medium alone or in the presence of SM23, at 0.780 and 3.125 μM. As expected, both doses strongly affected biofilm development and its morphology. Indeed, as shown in Figure 3 (left panel), unlike controls, SM23-treated biofilms showed large zones with few or no bacteria and scant extracellular matrix onto the well surface. Moreover, the residual biofilm showed drastic alterations in its architecture (Figure 3, left panel). In parallel experiments, a 24 h-old biofilm generated in the presence or absence of SM23 was stained with CFDA and PI, to discriminate live from dead cells, as detailed in section “Materials and Methods.” As shown in Figure 3 (right panel), a significant reduction of viable bacterial cells was observed in biofilms treated with SM23, irrespective of the dose used.
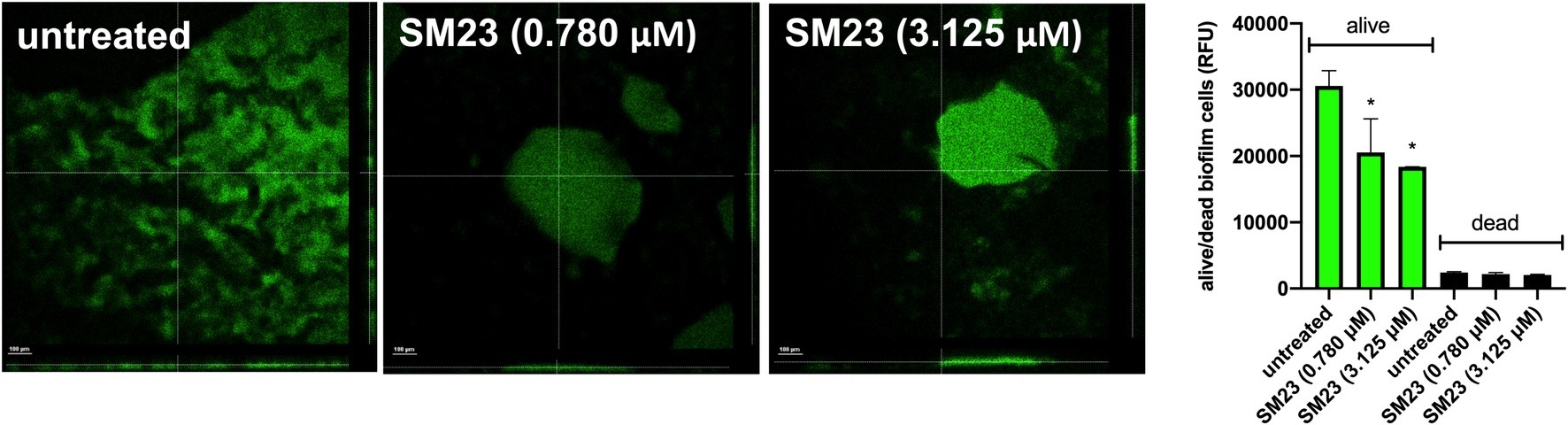
Figure 3. Confocal analysis of SM23-treated biofilm and evaluation of dead/alive cells. Left panel: confocal microscope images of a 24 h-old biofilm, untreated or treated with SM23 (0.780 and 3.125 μM). The large images illustrate three representative horizontal sections (X and Y axes) of biofilms, while the small images show biofilm sections observed along the Z axis. The micrographs are representative of two independent experiments. Scale bar: 100 μm. Magnification: 20x. Right panel: mean ± SEM of the Relative Fluorescence Units (RFU) of alive (green columns) and dead (black column) cells in a 24 h-old Pseudomonas biofilm treated with SM23 (0.780 and 3.125 μM). *p < 0.05; SM23-treated vs. untreated according to one-way ANOVA followed by Dunnett’s multiple comparisons test.
Inhibition of Pseudomonas aeruginosa Quorum Sensing-Related Virulence Factors by SM23
P. aeruginosa elastase is a zinc metalloprotease encoded by lasB gene, capable of inactivating a wide range of biological tissues and immunological agents (Preston et al., 1997; Li et al., 2019). Thus, we evaluated the elastase activity in supernatants of biofilm producing P. aeruginosa, after 6, 12 or 24 h of culture in the presence or absence of scalar doses of SM23. Our results showed a consistent elastase activity by the untreated control cells, already at 6 h persisting up to 24 h of incubation. Such activity was significantly dampened (about 45% of reduction) by SM23 treatment at all the doses tested (Table 1).
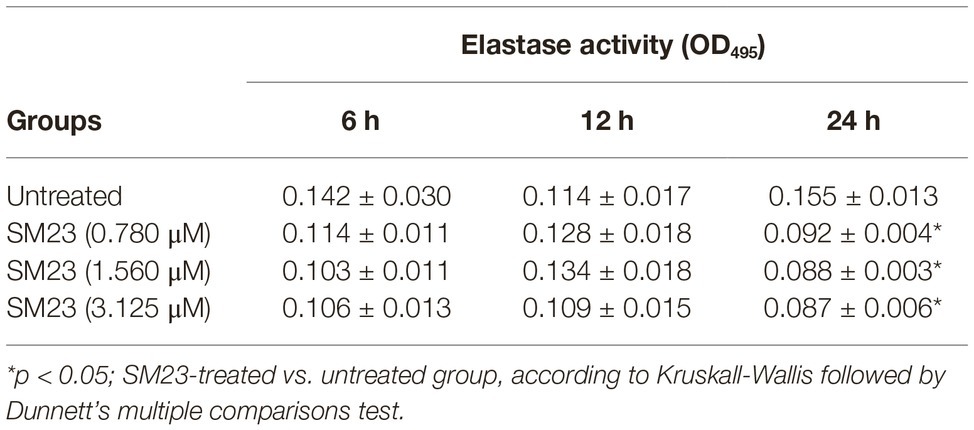
Table 1. Elastase activity of P. aeruginosa in response to SM23 treatment: time- and dose-dependence.
It is widely accepted that pyoverdine, the most potent iron-gathering siderophore in P. aeruginosa, is an important virulence factor playing an important role in biofilm formation (Meyer et al., 1996).
HPLC-MS analysis was performed to evaluate the levels of pyoverdines, pyocyanin, 3-oxo-C12-HSL and C4-HSL under the same experimental conditions used for elastase activity. Pyoverdines, in the form of doubly charged protonated molecular ions, were revealed by the presence of the specific chromophore core (P) fragment ion at m/z 204.0768 in their MS/MS spectra (Budzikiewicz et al., 2007; Wei and Aristilde, 2015). The identification of the four most intensely detected pyoverdines, achieved using their MS/MS spectra, revealed serine (Ser) as the first amino acid of the peptide sequence and either a succinyl amide (Succa, A1+ fragment ion at m/z 416.1565 Th) or a succinic acid (Succ, A1+ fragment ion at m/z 417.1405 Th) moiety as side chain of the chromophore. Two pyoverdines, with their [M + 2H]2+ ions being respectively at m/z 667.8015 and 667.3104 Th, were unambiguously identified, as previously reported for Pyoverdine D (Py D) and E (Py E) (Kilz et al., 1999); differently, other two pyoverdines, portrayed by their respective [M + 2H]2+ ions at m/z 644.7997 and 645.2914 Th, were not yet completely identified and hereinafter referred to as Succ-P-Ser-Y and Succa-P-Ser-Y, with Y being their unidentified peptide sequence side-chain.
Pyocyanin was detected as mono charged protonated ion [M + H]+ at m/z 211.0866 and its identification possibly confirmed by fragments [M + H-CH3]+ at m/z 196.0631, [M + H-CO]+ at m/z 183.0917 and [M + H-CH3-CO]+ at m/z 168.0682 appearing in its MS/MS spectrum (Watson et al., 1986). Both 3-oxo-C12-HSL and C4-HSL were revealed by the presence of their respective [M + H]+ and [M + Na]+ molecular ions (at m/z 172.0968 and 194.0788 for C4-HSL and at m/z 298.2013 and 320.1832 for 3-oxo-C12-HSL). Their identification was possibly confirmed by the fragments [M + H-C4H7NO2]+ at 71.0491 for C4-HSL and at 197.1536 for 3-oxo-C12-HSL and [C4H7NO2 + H]+ at 102.05496 appearing in their respective MS/MS spectra; these data closely recalled the profile previously established (Churchill et al., 2011).
Compounds elution provided peaks appearing in their specific chromatographic traces, showing the abundance of their respective doubly charged ions over the chromatographic run (mass range chromatograms). Peak areas were used for semi-quantitative evaluation, as previously established (Kapoore and Vaidyanathan, 2016). As depicted in Table 2, we found consistent levels of pyoverdines only after 24 h of P. aeruginosa culture. SM23 markedly reduced PyD and PyE production at all the three doses tested, whereas no effect was observed in terms of Py Succ-P-Ser-Y_isom1/2 or Py Succa-P-Ser-Y_isom1/2 levels. Under the same experimental conditions, we analyzed also the release of pyocyanin, another key virulence factor involved in P. aeruginosa biofilm formation (Li et al., 2015). Again, we found consistent levels of pyocyanin only after 24 h of P. aeruginosa culture, while the treatment with SM23, at 0.780, 1.560, and 3.125 μM, significantly impaired the pyocyanin production.
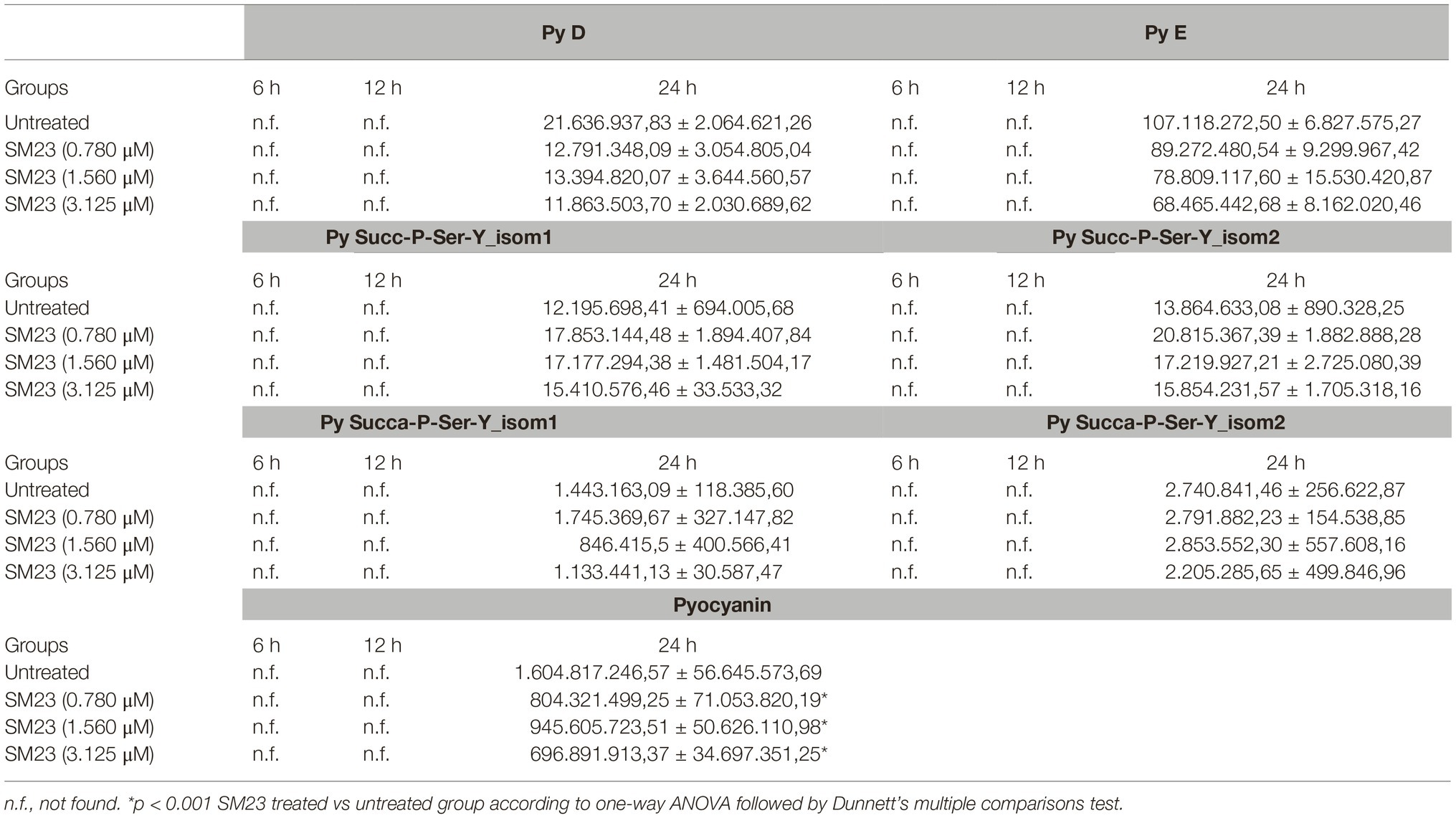
Table 2. Mass spectrometry analysis of P. aeruginosa pyoverdine and pyocyanin release in response to SM23 treatment: time- and dose-dependence.
Inhibition of Pseudomonas aeruginosa Quorum Sensing Molecules by SM23
The results in Table 3 show the levels of both 3-oxo-C12-HSL and C4-HSL autoinducers in supernatants of P. aeruginosa producing biofilm, after 6, 12, and 24 h of SM23 treatment. A consistent presence of 3-oxo-C12-HSL was measured already after 12 h that persisted up to 24 h, while C4-HSL was detectable only after 24 h of culture. All the SM23 doses significantly reduced 3-oxo-C12-HSL after 12 h of treatment; moreover, both the autoinducer molecules resulted significantly inhibited after 24 h of SM23 treatment at the highest dose (3.125 μM).
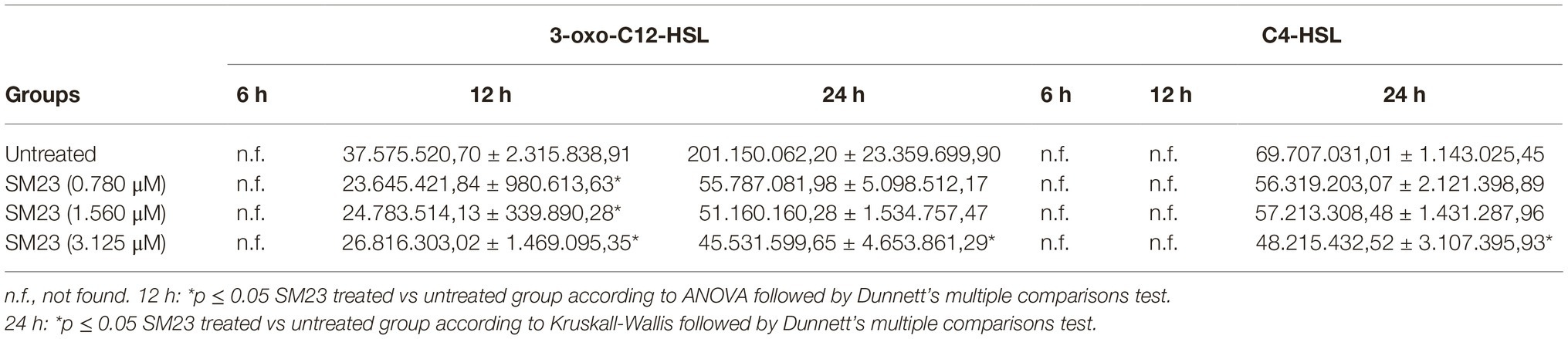
Table 3. Mass spectrometry analysis of P. aeruginosa 3-oxo-C12-HSL and C4-HSL QS molecules release in response to SM23 treatment: time- and dose-dependence.
Finally, we tested the effects of SM23 on P. aeruginosa expression of lasI/lasR genes, known to be crucially involved in QS system, biofilm formation, and virulence (Moghaddam, 2014). Our results showed that, in a 24 h-old biofilm treated with 1.560 μM SM23, both lasI and lasR gene transcript levels were downregulated by 36 and 25%, respectively (untreated vs. SM23-treated cells: lasI returned 1.0 vs. 0.644 fold change; lasR returned 1.0 vs. 0.750 fold change).
SM23 Interaction With Pseudomonas aeruginosa
Given the effects of the SM23 on P. aeruginosa biofilm formation and lasI/lasR system, we next investigated its possible mechanism of action. Thus, we developed two different experimental protocols to assess the localization of SM23 on bacterial surface, by exploiting the natural green auto-fluorescence of this compound, as detailed in section “Materials and Methods” and in Figure 1. The results, depicted in Figure 4, showed that, when the SM23 was added at Time 0 (protocol A), most of the fluorescence was associated to the cell pellet, with a minimal residual fluorescence being detectable in the supernatant. In contrast, when the SM23 was added at Time 5 h (protocol B), the fluorescence signal was detected at comparable levels both on the bacterial cell pellet and on the cell-free supernatants.
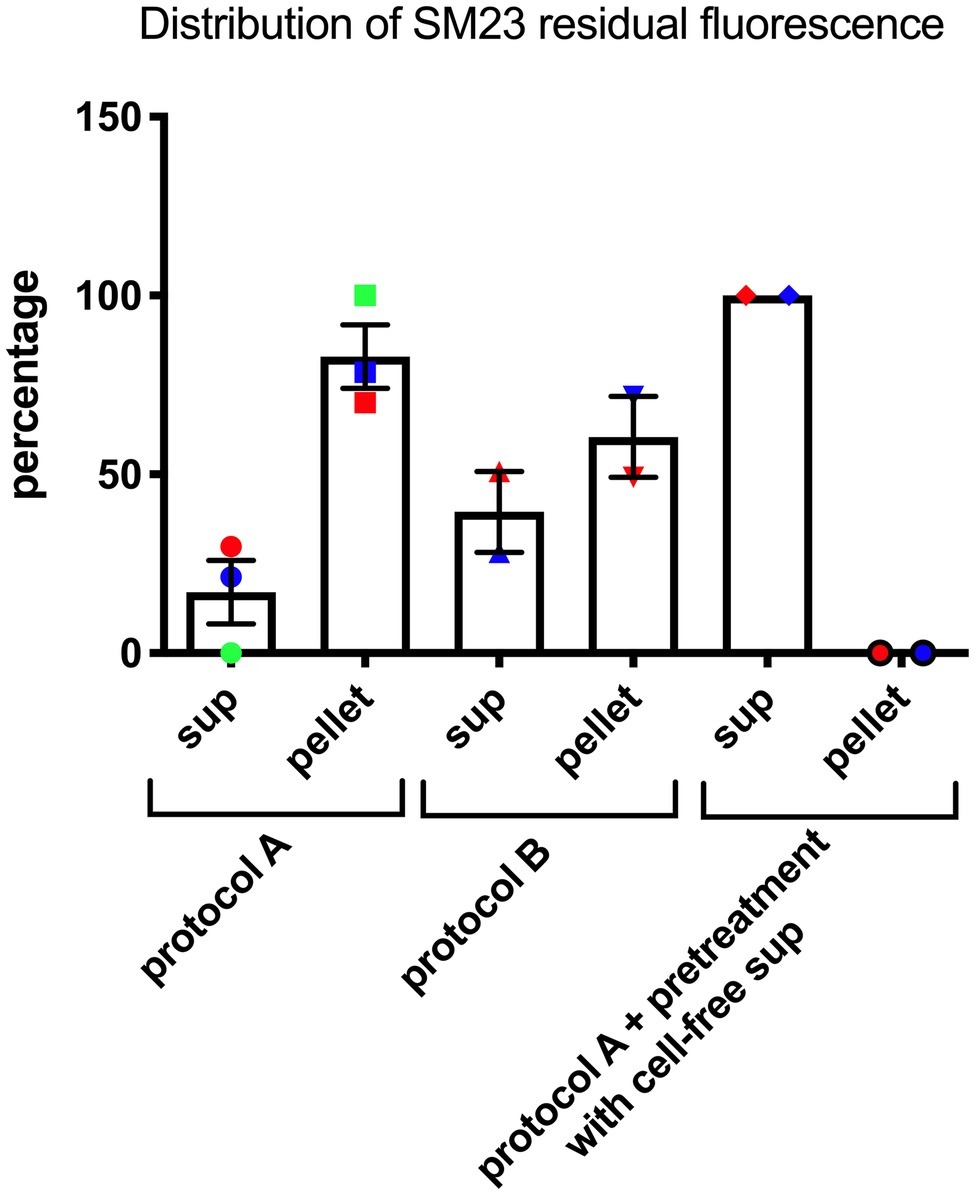
Figure 4. SM23 interaction with P. aeruginosa mean percent ± SEM of the fluorescence signal detected both in the cell-free supernatant (sup) and in the pellet. Each percent has been calculated using the data of replicate samples from two-three independent experiments. Each individual symbol identifies a single percent value.
Finally, a Pseudomonas-conditioned medium (from an overnight culture at 37°C) was used to pre-treat the bacterial cells for 30 min; then, the SM23 was added and the plates were further incubated for additional 7 h, as described in protocol A, prior to assessing the levels of free and cell-associated fluorescence. The results, depicted in Figure 4, showed that the fluorescence signal was not cell-associated, but rather only detectable into culture supernatants.
SM23 Inhibitory Effects on Endotracheal Tubes-Associated Biofilm: Impairment of Biofilm Formation and Pyoverdine Release by Pseudomonas aeruginosa
We have recently described a rapid and easy-to-perform in vitro model for the real-time monitoring of P. aeruginosa biofilm formation on endotracheal tube (ETT) pieces (Pericolini et al., 2018). Accordingly, we evaluated the effect of SM23 on a 24 h-old Pseudomonas biofilm produced on ETT pieces. As shown in Figure 5, the treatment with SM23 significantly decreased the amounts of biofilm associated with the ETT, at both the doses used, as indicated by the significant reduction in RLU (Figure 5, upper panel); in addition, the pyoverdine release by such ETT-associated biofilm was also impaired, reaching statistically significant differences at the dose of 1.56 μM (Figure 5, lower panel).
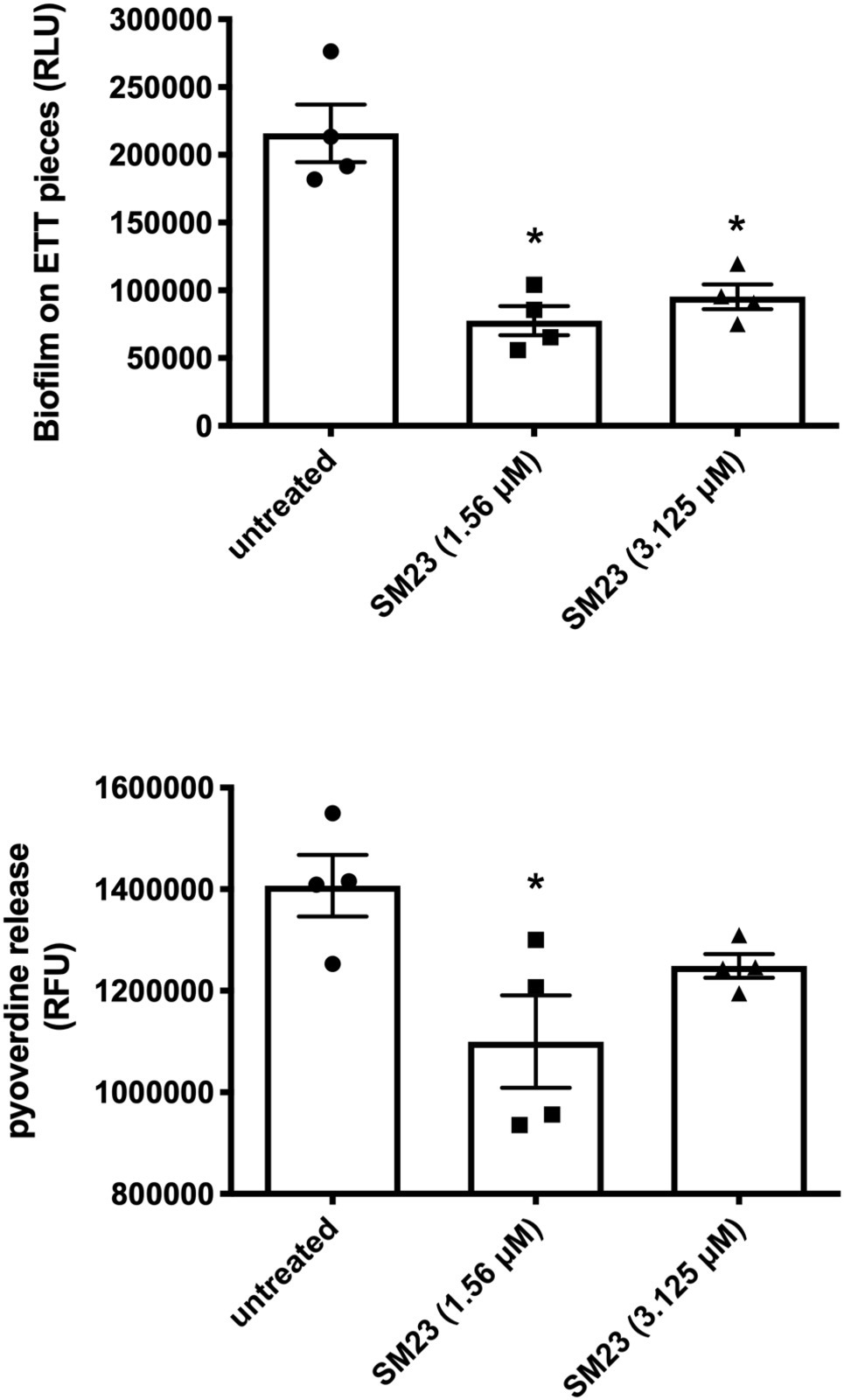
Figure 5. SM23 effects on medical devise-associated biofilm by P. aeruginosa: impairment of pyoverdine release and biofilm formation. Biofilm mass (upper panel), expressed as mean RLU ± SEM, and pyoverdine release (lower panel), expressed as mean RFU ± SEM, by a 24 h-old Pseudomonas biofilm produced on ETT pieces, in the absence and in the presence of SM23. In the figure, each individual symbol identifies the value corresponding to a single ETT piece. Data represent the mean values from two independent experiments, with duplicate samples. *p < 0.05; SM23-treated samples vs. untreated samples according to ANOVA followed by Dunnett’s multiple comparisons test.
Discussion
P. aeruginosa is a ubiquitous environmental bacterium, responsible for a wide range of severe opportunistic infections, characterized by intense neutrophil activation and significant tissue damage. Clearance is often a serious clinical challenge due to host immunodeficiency and bacterial multidrug resistance, including third-generation cephalosporin (Tenover, 2006; Moghaddam et al., 2012). Moreover, the treatment of chronic P. aeruginosa infections is further hampered by adaptive resistance, facilitated also by bacterial ability to form biofilm in vivo (Mulcahy et al., 2014). Indeed, bacteria growing as a biofilm have characteristics distinct from their planktonic cell counterpart, including an increased tolerance to antimicrobial agents and host immune response (Costerton, 1999). Furthermore, biofilm is an example of microbial community finely coordinating its behavior through QS. In this study, we provide the first evidence that, SM23, known to be a potent β-lactamase inhibitor, also impairs biofilm formation by P. aeruginosa and its production of some QS-dependent virulence factors as well as autoinducer molecules.
The boronic acid derivative SM23 has been described as a strong inhibitor of the Pseudomonas-derived cephalosporinase-3 (PDC-3), a class C β-lactamase which represents one of the major antibiotic-resistance determinants in P. aeruginosa (Ki = 4 nM) (Drawz et al., 2011). Such a boronic acid derivative is also able to lower the MIC values of cefotaxime in PDC-3 expressing E. coli, to a concentration of 4 μg/ml. Subsequent studies demonstrate that SM23 is highly active against other class C (Ki of 1 and 20 nM vs. AmpC and ADC-7, respectively) and class A (β-lactamases Ki of 420 and 64 nM vs. CTXM-16 and TEM-1, respectively), qualifying it as one of the best inhibitors of serine β-lactamase (Caselli et al., 2018). Our present results indicate that SM23 significantly inhibits P. aeruginosa biofilm formation, as established by measuring total biomass and metabolic activity of a 24 h-old biofilm. In particular, about 50% reduction in biofilm biomass occurs as assessed by CV assay (Figure 2, left panel), while a clear dose-dependent inhibition is detected by bioluminescence analysis; the latter, known to measure metabolic activity, allows to detect a gradual and consistent effect within the range of 0.390–25.0 μM (Figure 2, right panel). The impact of SM23 on biofilm is also evident by exploring its morphological structure by confocal microscopy. Indeed, unlike the control, SM23-treated biofilm is characterized by large zones with few or no bacteria and scant extracellular polymeric matrix attached to the well surface; accordingly, a noticeable reduction of viable bacteria is detected, as measured by the alive/dead cells assay.
Interestingly, the exposure of P. aeruginosa to SM23 significantly inhibits the production of some key QS-controlled virulence factors, namely pyoverdine, pyocyanin, and elastase. As a yellow-green fluorescent pigment, pyoverdine is a strong iron scavenger/iron transporter that promotes Pseudomonas pathogenicity stimulating its growth (Meyer et al., 1996). To date, about 40 structurally different pyoverdines have been identified, each one characterized by a distinct peptide chain and a specificity, peculiar to Pseudomonas species (Meyer, 2000). Pyoverdine production may also be accompanied by the appearance of related compounds, which are considered as biosynthetic precursors or later modifications; such compounds have the same peptide chain of pyoverdine, but a different chromophore group (Budzikiewicz, 2004). Another key pigment strictly regulated by Pseudomonas QS system, in particular by rhlI/rhlR, is pyocyanin. Here, we show that SM23 strongly inhibits pyocyanin production by Pseudomonas, in line with the marked reduction of pyoverdine production also observed after SM23 treatment.
For the determination of both pyoverdines and pyocyanin in culture supernatants, we have employed HPLC-MS analysis, that unambiguously identified (1) four chromophores belonging to the pyoverdine siderophore family and (2) one chromophore for pyocyanin. Interestingly, all of them happen to be markedly affected by SM23 treatment (see Table 2). Similarly, the elastase activity of P. aeruginosa is also significantly inhibited by SM23 during biofilm production. In line with these findings, the SM23 is shown to downregulate the expression of the QS-associated autoinducers 3-oxo-C12-HSL and C4-HSL during biofilm formation by dampened lasI/lasR gene expression, further suggesting that QS pathways are affected by SM23.
As described previously (Schuster and Greenberg, 2006; Girard and Bloemberg, 2008; Zou and Nair, 2009), the QS system of P. aeruginosa is mainly composed of two sets of genes. The first consists of lasI and lasR genes, encoding for HSL autoinducer synthase and R protein, respectively; the second system, named Rhl, comprises the rhlI and rhlR genes, which in turn encode for synthase and R protein. The two systems are strictly connected and hierarchically organized, with the lasI/lasR regulating the transcription of rhlI/rhlR (Schuster and Greenberg, 2006; Zou and Nair, 2009). Our present data, obtained through HPLC-MS analysis, show a significant downregulation of both the autoinducers, 3-oxo-C12-HSL and C4-HSL, in the supernatants of SM23-treated biofilm (at the dose of 3.125 μM), as compared to the untreated biofilm. These results indicate that SM23 significantly affects the synthesis of both 3-oxo-C12-HSL and C4-HSL; in line with others (Zou and Nair, 2009), we favor the idea that it may compete with the 3-oxo-C12-HSL autoinducer, likely for the same binding site on the LasR receptor. Many efforts will be necessary to deeply understand the molecular mechanisms underlying such a complex phenomenon.
By the experiments aimed at assessing SM23 interaction with bacterial cells, we demonstrate that most of its fluorescence appears to be bacteria-associated when the compound is added to a planktonic P. aeruginosa culture. Unexpectedly, such phenomenon is fully prevented once planktonic cells have been pre-incubated with a Pseudomonas-conditioned medium, prior to being exposed to SM23; in this case, the compound fluorescence is indeed detectable only in the supernatants. Also, when the compound is added to an early biofilm, approximately half of the SM23-related fluorescence signal remains unbound in the supernatant. Taken together, these data open to the hypothesis that Pseudomonas may counteract the activity of SM23 by producing something that prevents its binding to the same bacterial cell. It should be noted that, though strongly associated with the compound, planktonic cells are not affected by SM23 in their viability or rate of growth. Although remaining widely unexplained, these findings may envisage the possibility that, because of its unique mechanism of action, SM23 might be a novel compound likely insensitive to the well-known drug-resistance mechanisms.
Finally, we provide evidence that SM23 is effective also in reducing Pseudomonas biofilm biomass produced on a medical device, such as the ETT; importantly, also the release of pyoverdine by such sessile bacterial community is significantly affected by SM23. In our opinion, these data strengthen the interest on SM23, showing that it acts as anti-P. aeruginosa compound also under conditions closely mimicking biofilm infection in patients.
Overall, our results demonstrate that the boronic acid derivative SM23, besides being a strong inhibitor of β-lactamase, is able to drastically reduce biofilm formation and QS-related virulence factors in Pseudomonas. This in vitro evidence opens to future applications of SM23 in the prevention and treatment of biofilm-associated P. aeruginosa infections.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Author Contributions
SP, EP, FP, EC, and EB contributed to the conception and design of the study, revised the manuscript critically for important intellectual content, and provided approval for publication of the content. SP, EP, BC, DP, CC, and FF performed experiments and organized the results. EP performed the statistical analysis. SP wrote the first draft of the manuscript. SP, EP, EC, and DP wrote sections of the manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
Funding
EC was supported by the Department of Life Science, FAR2016DIP. EP was supported by Department of Diagnostic, Clinical and Public Health Medicine, FAR2017DIP.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.00035/full#supplementary-material
References
Barnes, M. D., Taracila, M. A., Rutter, J. D., Bethel, C. R., Galdadas, I., Hujer, A. M., et al. (2018). Deciphering the evolution of cephalosporin resistance to ceftolozane-tazobactam in Pseudomonas aeruginosa. MBio 9, 1–19. doi: 10.1128/mBio.02085-18
Berrazeg, M., Jeannot, K., Ntsogo Enguéné, V. Y., Broutin, I., Loeffert, S., Fournier, D., et al. (2015). Mutations in β-lactamase AmpC increase resistance of Pseudomonas aeruginosa isolates to antipseudomonal cephalosporins. Antimicrob. Agents Chemother. 59, 6248–6255. doi: 10.1128/AAC.00825-15
Brooun, A., Liu, S., and Lewis, K. (2000). A dose-response study of antibiotic resistance in Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 44, 640–646. doi: 10.1128/AAC.44.3.640-646.2000
Budzikiewicz, H. (2004). Siderophores of the Pseudomonadaceae sensu stricto (fluorescent and non-fluorescent Pseudomonas spp.). Fortschr. Chem. Org. Naturst. 87, 81–237. doi: 10.1007/978-3-7091-0581-8_2
Budzikiewicz, H., Schäfer, M., Fernández, D. U., Matthijs, S., and Cornelis, P. (2007). Characterization of the chromophores of pyoverdins and related siderophores by electrospray tandem mass spectrometry. Biometals 20, 135–144. doi: 10.1007/s10534-006-9021-3
Bush, K., and Bradford, P. A. (2016). β-Lactams and β-lactamase inhibitors: an overview. Cold Spring Harb. Perspect. Med. 6, 295–306. doi: 10.1101/cshperspect.a025247
Bush, K., and Bradford, P. A. (2019). Interplay between β-lactamases and new β-lactamase inhibitors. Nat. Rev. Microbiol. 17, 295–306. doi: 10.1038/s41579-019-0159-8
Caselli, E., Romagnoli, C., Powers, R. A., Taracila, M. A., Bouza, A. A., Swanson, H. C., et al. (2018). Inhibition of Acinetobacter-derived cephalosporinase: exploring the carboxylate recognition site using novel β-lactamase inhibitors. ACS Infect. Dis. 4, 337–348. doi: 10.1021/acsinfecdis.7b00153
Choi, K.-H., and Schweizer, H. P. (2006). Mini-Tn7 insertion in bacteria with single attTn7 sites: example Pseudomonas aeruginosa. Nat. Protoc. 1, 153–161. doi: 10.1038/nprot.2006.24
Churchill, M. E. A., Sibhatu, H. M., and Uhlson, C. L. (2011). Defining the structure and function of acyl-homoserine lactone autoinducers. Methods Mol. Biol. 692, 159–171. doi: 10.1007/978-1-60761-971-0_12
Cornelis, P., and Matthijs, S. (2002). Diversity of siderophore-mediated iron uptake systems in fluorescent pseudomonads: not only pyoverdines. Environ. Microbiol. 4, 787–798. doi: 10.1046/j.1462-2920.2002.00369.x
Costerton, J. W. (1999). Introduction to biofilm. Int. J. Antimicrob. Agents 11, 217–221; discussion 237–239. doi: 10.1016/S0924-8579(99)00018-7
Cross, A., Allen, J. R., Burke, J., Ducel, G., Harris, A., John, J., et al. (1983). Nosocomial infections due to Pseudomonas aeruginosa: review of recent trends. Rev. Infect. Dis. 5(Suppl. 5), S837–S845.
Das, M. C., Sandhu, P., Gupta, P., Rudrapaul, P., De, U. C., Tribedi, P., et al. (2016). Attenuation of Pseudomonas aeruginosa biofilm formation by vitexin: a combinatorial study with azithromycin and gentamicin. Sci. Rep. 6:23347. doi: 10.1038/srep23347
de Kievit, T. R., and Iglewski, B. H. (2000). Bacterial quorum sensing in pathogenic relationships. Infect. Immun. 68, 4839–4849. doi: 10.1128/IAI.68.9.4839-4849.2000
Donabedian, H. (2003). Quorum sensing and its relevance to infectious diseases. J. Infect. 46, 207–214. doi: 10.1053/jinf.2002.1120
Dong, Y. H., Wang, L. H., Xu, J. L., Zhang, H. B., Zhang, X. F., and Zhang, L. H. (2001). Quenching quorum-sensing-dependent bacterial infection by an N-acyl homoserine lactonase. Nature 411, 813–817. doi: 10.1038/35081101
Drawz, S. M., Taracila, M., Caselli, E., Prati, F., and Bonomo, R. A. (2011). Exploring sequence requirements for C₃/C₄ carboxylate recognition in the Pseudomonas aeruginosa cephalosporinase: insights into plasticity of the AmpC β-lactamase. Protein Sci. 20, 941–958. doi: 10.1002/pro.612
Eidam, O., Romagnoli, C., Dalmasso, G., Barelier, S., Caselli, E., Bonnet, R., et al. (2012). Fragment-guided design of subnanomolar β-lactamase inhibitors active in vivo. Proc. Natl. Acad. Sci. USA 109, 17448–17453. doi: 10.1073/pnas.1208337109
Fuqua, C., and Greenberg, E. P. (1998). Self perception in bacteria: quorum sensing with acylated homoserine lactones. Curr. Opin. Microbiol. 1, 183–189. doi: 10.1016/S1369-5274(98)80009-X
Girard, G., and Bloemberg, G. V. (2008). Central role of quorum sensing in regulating the production of pathogenicity factors in Pseudomonas aeruginosa. Future Microbiol. 3, 97–106. doi: 10.2217/17460913.3.1.97
Griffith, D. C., Sabet, M., Tarazi, Z., Lomovskaya, O., and Dudley, M. N. (2019). Pharmacokinetics/pharmacodynamics of vaborbactam, a novel beta-lactamase inhibitor, in combination with meropenem. Antimicrob. Agents Chemother. 63, 1–9. doi: 10.1128/AAC.01659-18
Jakobsen, T. H., Bjarnsholt, T., Jensen, P. Ø., Givskov, M., and Høiby, N. (2013). Targeting quorum sensing in Pseudomonas aeruginosa biofilms: current and emerging inhibitors. Future Microbiol. 8, 901–921. doi: 10.2217/fmb.13.57
Kapoore, R. V., and Vaidyanathan, S. (2016). Towards quantitative mass spectrometry-based metabolomics in microbial and mammalian systems. Philos. Transact. A Math. Phys. Eng. Sci. 374, 1–14. doi: 10.1098/rsta.2015.0363
Kilz, S., Lenz, C., Fuchs, R., and Budzikiewicz, H. (1999). A fast screening method for the identification of siderophores from fluorescent Pseudomonas spp. by liquid chromatography/electrospray mass spectrometry. J. Mass Spectrom. 34, 281–290. doi: 10.1002/(SICI)1096-9888(199904)34:4<281::AID-JMS750>3.0.CO;2-M
Lee, J., and Zhang, L. (2015). The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein Cell 6, 26–41. doi: 10.1007/s13238-014-0100-x
Li, H., Li, X., Wang, Z., Fu, Y., Ai, Q., Dong, Y., et al. (2015). Autoinducer-2 regulates Pseudomonas aeruginosa PAO1 biofilm formation and virulence production in a dose-dependent manner. BMC Microbiol. 15:192. doi: 10.1186/s12866-015-0529-y
Li, J., Ramezanpour, M., Fong, S. A., Cooksley, C., Murphy, J., Suzuki, M., et al. (2019). Pseudomonas aeruginosa exoprotein-induced barrier disruption correlates with elastase activity and marks chronic rhinosinusitis severity. Front. Cell. Infect. Microbiol. 9:38. doi: 10.3389/fcimb.2019.00038
Lyczak, J. B., Cannon, C. L., and Pier, G. B. (2002). Lung infections associated with cystic fibrosis. Clin. Microbiol. Rev. 15, 194–222. doi: 10.1128/CMR.15.2.194-222.2002
Mah, T. F., and O’Toole, G. A. (2001). Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 9, 34–39. doi: 10.1016/S0966-842X(00)01913-2
Meyer, J. M. (2000). Pyoverdines: pigments, siderophores and potential taxonomic markers of fluorescent Pseudomonas species. Arch. Microbiol. 174, 135–142. doi: 10.1007/s002030000188
Meyer, J. M., Neely, A., Stintzi, A., Georges, C., and Holder, I. A. (1996). Pyoverdin is essential for virulence of Pseudomonas aeruginosa. Infect. Immun. 64, 518–523. doi: 10.1128/IAI.64.2.518-523.1996
Moghaddam, M. M. (2014). Quorum sensing in bacteria and a glance on Pseudomonas aeruginosa. Clin. Microbiol. Open Access 3, 133–139. doi: 10.4172/2327-5073.1000156
Moghaddam, M. M., Abolhassani, F., Babavalian, H., Mirnejad, R., Azizi Barjini, K., and Amani, J. (2012). Comparison of in vitro antibacterial activities of two cationic peptides CM15 and CM11 against five pathogenic bacteria: Pseudomonas aeruginosa, Staphylococcus aureus, Vibrio cholerae, Acinetobacter baumannii, and Escherichia coli. Probiotics Antimicrob. Proteins 4, 133–139. doi: 10.1007/s12602-012-9098-7
Morandi, F., Caselli, E., Morandi, S., Focia, P. J., Blázquez, J., Shoichet, B. K., et al. (2003). Nanomolar inhibitors of AmpC beta-lactamase. J. Am. Chem. Soc. 125, 685–695. doi: 10.1021/ja0288338
Mulcahy, L. R., Isabella, V. M., and Lewis, K. (2014). Pseudomonas aeruginosa biofilms in disease. Microb. Ecol. 68, 1–12. doi: 10.1007/s00248-013-0297-x
Ni, N., Choudhary, G., Peng, H., Li, M., Chou, H.-T., Lu, C.-D., et al. (2009). Inhibition of quorum sensing in Vibrio harveyi by boronic acids. Chem. Biol. Drug Des. 74, 51–56. doi: 10.1111/j.1747-0285.2009.00834.x
Ohman, D. E., Cryz, S. J., and Iglewski, B. H. (1980). Isolation and characterization of Pseudomonas aeruginosa PAO mutant that produces altered elastase. J. Bacteriol. 142, 836–842. doi: 10.1128/JB.142.3.836-842.1980
Pericolini, E., Colombari, B., Ferretti, G., Iseppi, R., Ardizzoni, A., Girardis, M., et al. (2018). Real-time monitoring of Pseudomonas aeruginosa biofilm formation on endotracheal tubes in vitro. BMC Microbiol. 18:84. doi: 10.1186/s12866-018-1224-6
Pesci, E. C., Milbank, J. B., Pearson, J. P., McKnight, S., Kende, A. S., Greenberg, E. P., et al. (1999). Quinolone signaling in the cell-to-cell communication system of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 96, 11229–11234. doi: 10.1073/pnas.96.20.11229
Preston, M. J., Seed, P. C., Toder, D. S., Iglewski, B. H., Ohman, D. E., Gustin, J. K., et al. (1997). Contribution of proteases and LasR to the virulence of Pseudomonas aeruginosa during corneal infections. Infect. Immun. 65, 3086–3090. doi: 10.1128/IAI.65.8.3086-3090.1997
Qu, L., She, P., Wang, Y., Liu, F., Zhang, D., Chen, L., et al. (2016). Effects of norspermidine on Pseudomonas aeruginosa biofilm formation and eradication. MicrobiologyOpen 5, 402–412. doi: 10.1002/mbo3.338
Rossolini, G. M., and Mantengoli, E. (2005). Treatment and control of severe infections caused by multiresistant Pseudomonas aeruginosa. Clin. Microbiol. Infect. 11(Suppl. 4), 17–32. doi: 10.1111/j.1469-0691.2005.01161.x
Sarabhai, S., Harjai, K., Sharma, P., and Capalash, N. (2015). Ellagic acid derivatives from Terminalia chebula Retz. increase the susceptibility of Pseudomonas aeruginosa to stress by inhibiting polyphosphate kinase. J. Appl. Microbiol. 118, 817–825. doi: 10.1111/jam.12733
Schuster, M., and Greenberg, E. P. (2006). A network of networks: quorum-sensing gene regulation in Pseudomonas aeruginosa. Int. J. Med. Microbiol. 296, 73–81. doi: 10.1016/j.ijmm.2006.01.036
Stepanovic, S., Vukovic, D., Dakic, I., Savic, B., and Svabic-Vlahovic, M. (2000). A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 40, 175–179. doi: 10.1016/S0167-7012(00)00122-6
Stover, C. K., Pham, X. Q., Erwin, A. L., Mizoguchi, S. D., Warrener, P., Hickey, M. J., et al. (2000). Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 406, 959–964. doi: 10.1038/35023079
Tenover, F. C. (2006). Mechanisms of antimicrobial resistance in bacteria. Am. J. Infect. Control 34, S3–S10; discussion S64–73. doi: 10.1016/j.ajic.2006.05.219
Vandeputte, O. M., Kiendrebeogo, M., Rajaonson, S., Diallo, B., Mol, A., El Jaziri, M., et al. (2010). Identification of catechin as one of the flavonoids from Combretum albiflorum bark extract that reduces the production of quorum-sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol. 76, 243–253. doi: 10.1128/AEM.01059-09
Venturi, V. (2006). Regulation of quorum sensing in Pseudomonas. FEMS Microbiol. Rev. 30, 274–291. doi: 10.1111/j.1574-6976.2005.00012.x
Watson, D., MacDermot, J., Wilson, R., Cole, P. J., and Taylor, G. W. (1986). Purification and structural analysis of pyocyanin and 1-hydroxyphenazine. Eur. J. Biochem. 159, 309–313. doi: 10.1111/j.1432-1033.1986.tb09869.x
Wei, H., and Aristilde, L. (2015). Structural characterization of multiple pyoverdines secreted by two Pseudomonas strains using liquid chromatography-high resolution tandem mass spectrometry with varying dissociation energies. Anal. Bioanal. Chem. 407, 4629–4638. doi: 10.1007/s00216-015-8659-5
Xu, Z., Fang, X., Wood, T. K., and Huang, Z. J. (2013). A systems-level approach for investigating Pseudomonas aeruginosa biofilm formation. PLoS One 8:e57050. doi: 10.1371/journal.pone.0057050
Keywords: boronic acids, Pseudomonas aeruginosa biofilm, quorum sensing, virulence factors, inhibitors
Citation: Peppoloni S, Pericolini E, Colombari B, Pinetti D, Cermelli C, Fini F, Prati F, Caselli E and Blasi E (2020) The β-Lactamase Inhibitor Boronic Acid Derivative SM23 as a New Anti-Pseudomonas aeruginosa Biofilm. Front. Microbiol. 11:35. doi: 10.3389/fmicb.2020.00035
Edited by:
Silvia Buroni, University of Pavia, ItalyReviewed by:
Mona I. Shaaban, Mansoura University, EgyptRodolfo García-Contreras, National Autonomous University of Mexico, Mexico
Copyright © 2020 Peppoloni, Pericolini, Colombari, Pinetti, Cermelli, Fini, Prati, Caselli and Blasi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Emilia Caselli, ZW1pbGlhLmNhc2VsbGlAdW5pbW9yZS5pdA==; Elisabetta Blasi, ZWxpc2FiZXR0YS5ibGFzaUB1bmltb3JlLml0
†These authors have contributed equally to this work