Abstract
Seven coronaviruses (CoVs) have been isolated from humans so far. Among them, three emerging pathogenic CoVs, including severe acute respiratory syndrome coronavirus (SARS-CoV), Middle East respiratory syndrome coronavirus (MERS-CoV), and a newly identified CoV (2019-nCoV), once caused or continue to cause severe infections in humans, posing significant threats to global public health. SARS-CoV infection in humans (with about 10% case fatality rate) was first reported from China in 2002, while MERS-CoV infection in humans (with about 34.4% case fatality rate) was first reported from Saudi Arabia in June 2012. 2019-nCoV was first reported from China in December 2019, and is currently infecting more than 70000 people (with about 2.7% case fatality rate). Both SARS-CoV and MERS-CoV are zoonotic viruses, using bats as their natural reservoirs, and then transmitting through intermediate hosts, leading to human infections. Nevertheless, the intermediate host for 2019-nCoV is still under investigation and the vaccines against this new CoV have not been available. Although a variety of vaccines have been developed against infections of SARS-CoV and MERS-CoV, none of them has been approved for use in humans. In this review, we have described the structure and function of key proteins of emerging human CoVs, overviewed the current vaccine types to be developed against SARS-CoV and MERS-CoV, and summarized recent advances in subunit vaccines against these two pathogenic human CoVs. These subunit vaccines are introduced on the basis of full-length spike (S) protein, receptor-binding domain (RBD), non-RBD S protein fragments, and non-S structural proteins, and the potential factors affecting these subunit vaccines are also illustrated. Overall, this review will be helpful for rapid design and development of vaccines against the new 2019-nCoV and any future CoVs with pandemic potential. This review was written for the topic of Antivirals for Emerging Viruses: Vaccines and Therapeutics in the Virology section of Frontiers in Microbiology.
Introduction
Coronaviruses (CoVs) belong to the subfamily Othocoronavirinae, in the family Coronaviridae of the order Nidovirales. According to the 10th Report on Virus Taxonomy from the International Committee on Taxonomy of Viruses (ICTV), the Othocoronavirinae is comprised of four genera, including alphacoronavirus (alpha-CoV), betacoronavirus (beta-CoV), gammacoronavirus (gamma-CoV), and deltacoronavirus (delta-CoV) (King et al., 2018). Alpha- and beta-CoVs can infect mammals, including but not limited to bats, pigs, cats, mice, and humans (Kusanagi et al., 1992; Li et al., 2005b; Poon et al., 2005; Drexler et al., 2014; Pedersen, 2014; Kudelova et al., 2015; Cui et al., 2019). Gamma- and delta-CoVs usually infect birds, while some of them could infect mammals (Woo et al., 2009a, 2012, 2014; Ma et al., 2015). Since the late sixties, CoVs have been recognized as one of the viral sources responsible for the common cold. Among all CoVs identified so far, seven have the ability to infect humans, including human coronavirus 229E (HCoV-229E) and human coronavirus NL63 (HCoV-NL63), which belong to alpha-CoVs (Hamre and Procknow, 1966; Chiu et al., 2005), as well as human coronavirus OC43 (HCoV-OC43), human coronavirus HKU1 (HCoV-HKU1), severe acute respiratory syndrome coronavirus (SARS-CoV), Middle East respiratory syndrome coronavirus (MERS-CoV), and the newly emerged coronavirus (2019-nCoV), which are known to be beta-CoVs (Drosten et al., 2003; Ksiazek et al., 2003; Vabret et al., 2003; Woo et al., 2005; Zaki et al., 2012; Du et al., 2016b; Zhang et al., 2020; Zhu et al., 2020) (Figure 1).
FIGURE 1
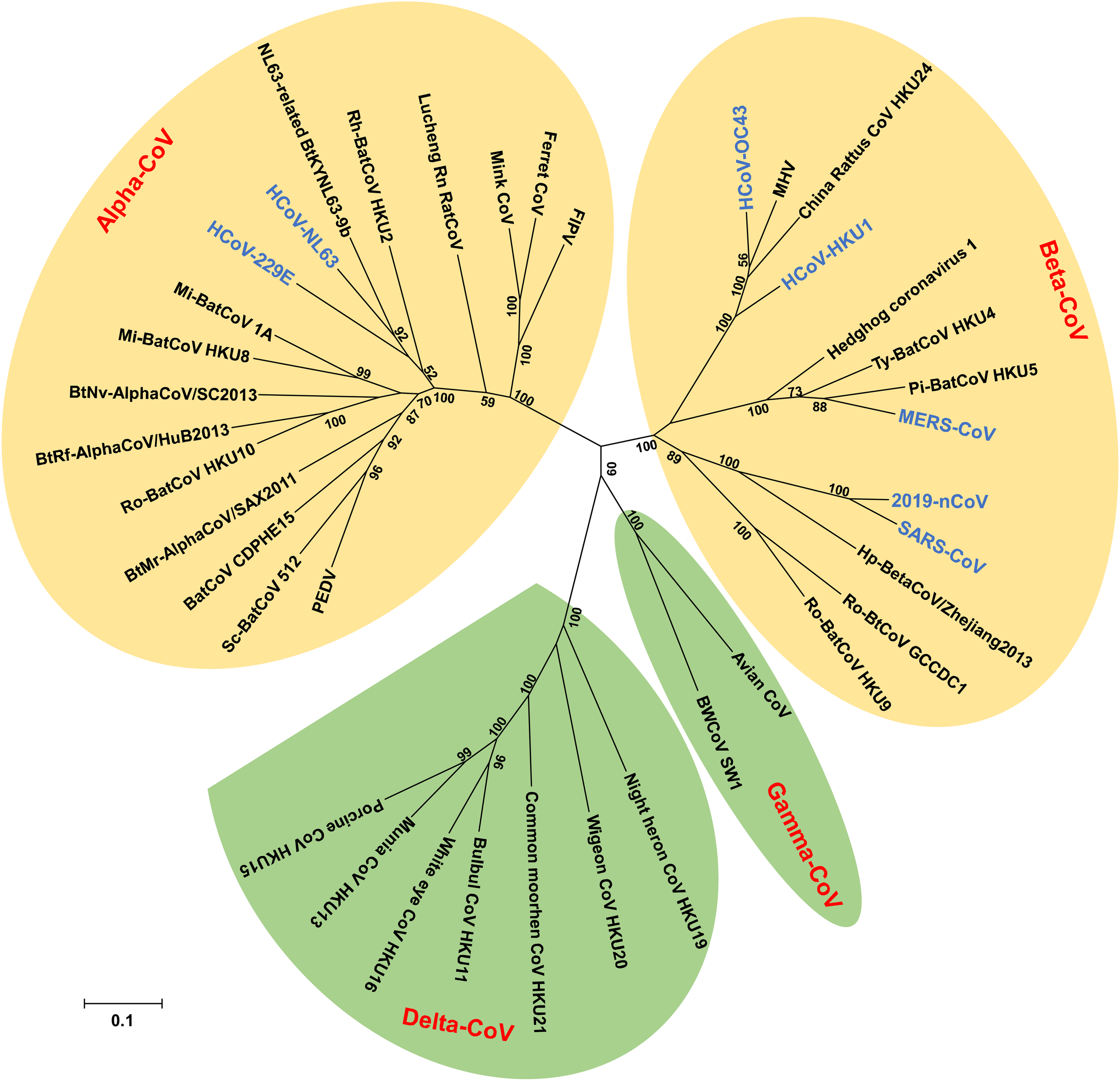
Phylogenetic tree of coronaviruses (CoVs) based on the nucleotide sequences of RNA dependent RNA polymerase (RdRp). The Tree, with 1,000 bootstrap values, was constructed by the maximum likelihood method using MEGA 6. The four main phylogenetic clusters correspond to genera alpha-CoV, beta-CoV, gamma-CoV, and delta-CoV. Each CoV genus contains different subgenera. The letters in blue indicate human CoVs.
Four human CoVs, including HCoV-229E, HCoV-NL63, HCoV-OC43, and HCoV-HKU1, have been identified in humans, but without causing severe infections. HCoV-229E was isolated from nasal secretions of medical students with minor upper respiratory disease. This virus was an original isolate, and was first reported in the 1960s (Hamre and Procknow, 1966). In addition to HCoV-229E, several studies have reported the recovery of HCoV-OC43 from patients with upper respiratory tract illness (Tyrrell and Bynoe, 1965; Hamre et al., 1967; McIntosh et al., 1967; Kapikian et al., 1969). In 2004, HCoV-NL63 was isolated from clinical species of infants suffering from pneumonia or bronchiolitis, and characterized for its ability to infect human respiratory tract (Fouchier et al., 2004; van der Hoek et al., 2004). The subsequent study in 2005 identified a new member of CoVs, named HCoV-HKU1, from a 71-year-old man with pneumonia (Woo et al., 2005). Generally, these four viruses are the most common pathogens causing mild upper respiratory infection or asymptomatic infection, and count for about 30% of all colds (Myint, 1994; Lau et al., 2006; Kim et al., 2017). In the serological surveillance on healthy adults, HCoV-229E, HCoV-NL63, and HCoV-OC43 demonstrated more than 90% seropositive with the immunological assay. It appears common for these CoVs to infect children (Mourez et al., 2007; Shao et al., 2007; Severance et al., 2008). In contrast to the above three human CoVs, HCoV-HKU1 has around 50% seropositive in healthy individuals and a relatively low exposure rate in children (Lehmann et al., 2008; Severance et al., 2008). Although the prevalence of various CoVs is different, the incidence among these viruses shows no significant difference (Woo et al., 2009b). The afore-mentioned four CoVs have been detected in 2.1–17.9% of clinical specimens (Esper et al., 2006; Lau et al., 2006; Gerna et al., 2007; Regamey et al., 2008; Matoba et al., 2015; Killerby et al., 2018). These viruses have also been associated with lower respiratory tract illness in children, elders, and immunodeficient individuals (Falsey et al., 2002; Fouchier et al., 2004; Woo et al., 2005; Gerna et al., 2006). HCoV-229E and HCoV-OC43 may lead to central nervous system infection since viral RNAs are detected in the brain of some patients (Arbour et al., 2000; Desforges et al., 2014).
Unlike the above four human CoVs, SARS-CoV, MERS-CoV, and 2019-nCoV have caused severe pneumonia and/or failure of other organs, even death, among infected populations (Nicholls et al., 2003; Zhong et al., 2003; Zaki et al., 2012; Zhu et al., 2020). The epidemic outbreak of SARS-CoV began in the Guangdong Province of China in November 2002, and spread through human-to-human transmission to other parts of the world within a few months (Ksiazek et al., 2003). From November 2002 to August 2003, SARS-CoV infected more than 8,098 people in 29 counties, resulting in over 774 deaths with ∼10% fatality rate (Du et al., 2009a). Palm civets serving as a potential intermediate host of this virus were traced immediately (Tu et al., 2004). Chinese horseshoe bats (Rhinolophus sinicus) are the natural reservoir of SARS-CoV (Li et al., 2005b). Various bat SARS-related CoVs (SARSr-CoV) have been identified in Yunnan, China, several of which can infect human cells, and have been further characterized (Ge et al., 2013; Hu et al., 2017). These discoveries indicate the threat of re-emergence of SARS-CoV or SARSr-CoV.
A decade later, another highly pathogenic human CoV, MERS-CoV, emerged, and the first patient with MERS-CoV infection was reported in Saudi Arabia in June 2012 (Zaki et al., 2012). By December 26, 2019, a total of 2,494 laboratory-confirmed cases of MERS, including 858 associated deaths in 27 countries (fatality rate 34.4%), were reported to the WHO1. Globally, the majority (about 80%) of human cases have been reported in Saudi Arabia, where people get infected through direct contact with infected dromedary camels or persons2 (Zaki et al., 2012). Isolation of MERS-CoV and detection of neutralizing antibodies from dromedary camels suggest that these camels are potentially an important intermediate host (Reusken et al., 2013; Azhar et al., 2014). Similar to SARS-CoV, MERS-CoV is also an emerging zoonotic virus (Li and Du, 2019). Bats habituate several CoVs phylogenetically related to MERS-CoV, and some of them are identical to MERS-CoVs, suggesting that MERS-CoV may originate from bats (Annan et al., 2013; Lelli et al., 2013; Lau et al., 2018; Luo et al., 2018a). Different from SARS-CoV, which has not caused infections in humans since 2004 (Du et al., 2009a), the transmission of MERS-CoV has not been interrupted, and the infected human cases continue increasing1 (Mobaraki and Ahmadzadeh, 2019). Currently, human-to-human transmission of MERS-CoV is limited.
A new CoV, 2019-nCoV, has caught worldwide attention (Liu and Saif, 2020; Zhang et al., 2020). It was first identified in Wuhan, China in December 2019, from patients with pneumonia (Zhu et al., 2020), and has infected more than 70000 people globally, including 2,009 deaths (∼2.7% fatality rate), as of February 19, 2020, particularly in China, and the other parts of the world, including Australia, Japan, Malaysia, Singapore, South Korea, Viet Nam, Cambodia, Philippines, Thailand, Nepal, Sri Lanka, India, United States, Canada, France, Finland, Germany, Italy, Russian Federation, Spain, Sweden, United Kingdom, Belgium, Egypt, and United Arab Emirates3. Different from MERS-CoV but similar to SARS-CoV, 2019-nCoV can cause human-to-human transmission, and its intermediate host that leads to the current human infection and outbreak is still under investigation.
Genome of Emerging Human Coronaviruses, as Well as Structure and Function of Their Key Proteins
The human CoVs are enveloped viruses with a positive-sense, single-stranded RNA genome. They are 80–160 nm in diameter. Like other CoVs, human CoVs contain the largest viral genome [27–32 kilobase pairs (kb)] among the RNA viruses, and they share similar genome organization (Fehr and Perlman, 2015). Two large overlapping open reading frames (ORFs), ORF 1a and ORF 1b, occupy two-thirds of the genome at the 5′-terminus, and a third of the genome at the 3′-terminus encodes four common structural proteins in the gene order of spike (S), envelope (E), membrane (M), and nucleocapsid (N) (5′–3′) (Fehr and Perlman, 2015). The large ORF 1ab is a replicase gene encoding polyproteins 1a (pp1a) and pp1b/1ab, which can be cleaved into 15–16 non-structural proteins (nsp2-nsp16 or nsp1-nsp16) by 3C-like proteinase (3CLpro, nsp5) and papain-like proteinase (PLpro, nsp3) (Bailey-Elkin et al., 2014; Fehr and Perlman, 2015; Tomar et al., 2015; Snijder et al., 2016). In addition to the genes encoding the above structural proteins, the genes encoding accessory proteins have also been detected in the 3′ region between S–E–M–N (Fehr and Perlman, 2015). Some beta-CoVs, such as HCoV-OC43 and HCoV-HKU1, contain hemagglutinin-esterase (HE) gene located between ORF 1ab and S gene encoding an additional structural protein, HE (De Groot et al., 2011; Desforges et al., 2013; Huang et al., 2015). Similar to other human CoVs, SARS-CoV possesses a ∼29-kb genome, which encodes pp1a and pp1ab, four main structural proteins (S, E, M, and N), and eight accessory proteins, such as 3a, 3b, 6, 7a, 7b, 8a, 8b, and 9b (Figure 2A) (Marra et al., 2003; Snijder et al., 2003). The MERS-CoV genome is about 30 kb in length and encodes pp1a, pp1ab, four structural proteins (S, E, M, and N), and five accessory proteins (3, 4a, 4b, 5, and 8b) (Figure 2A) (van Boheemen et al., 2012). The genomic RNA of SARS-CoV and MERS-CoV is packed inside capsid formed by the N protein, while the M, E, and S proteins form the envelope surrounding the capsid (Figure 2B). Accessory genes may incorporate into virions at low levels (Liu et al., 2014). Nevertheless, neither SARS-CoV nor MERS-CoV appears to contain the HE gene (Rota et al., 2003; Zaki et al., 2012).
FIGURE 2
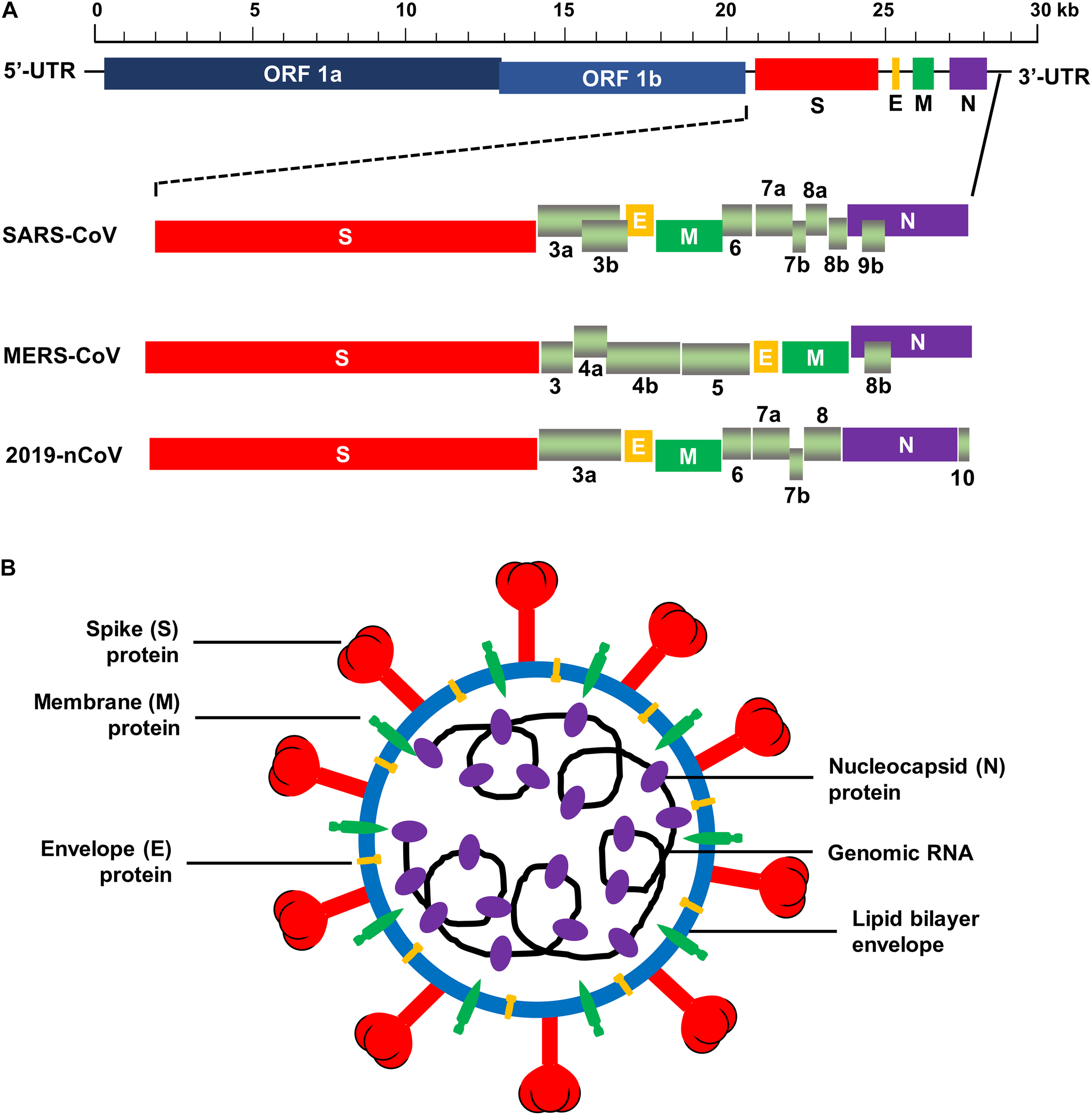
Schematic structure of SARS-CoV, MERS-CoV, and 2019-nCoV. (A) Schematic diagram of genomic organization of SARS-CoV, MERS-CoV, and 2019-nCoV. The genomic regions or open-reading frames (ORFs) are compared. Structural proteins, including spike (S), envelope (E), membrane (M) and nucleocapsid (N) proteins, as well as non-structural proteins translated from ORF 1a and ORF 1b and accessory proteins, including 3a, 3b, 6, 7a, 7b, 8a, 8b, and 9b (for SARS-CoV), 3, 4a, 4b, 5, and 8b (for MERS-CoV), and 3a, 6, 7a, 7b, 8, and 10 (for 2019-nCoV) are indicated. 5′-UTR and 3′-UTR, untranslated regions at the N- and C-terminal regions, respectively. Kb, kilobase pair. (B) Schematic structure of virion of SARS-CoV, MERS-CoV, and 2019-nCoV and its major structural proteins.
Several proteins of human CoVs are important for viral infection and/or pathogenesis. For example, most nsps participate in viral RNA replication and/or transcription (Snijder et al., 2016). The accessory proteins interact with host cells, potentially helping the viruses to evade the immune system and increase their virulence (Menachery et al., 2017). The HE protein assists in the attachment of virus–host cells, thus playing a key role in the production of infectious virions, as in the case of HCoV-OC43 (Desforges et al., 2013). The M and E proteins are responsible for virus assembly or promote virulence (Scobey et al., 2013; DeDiego et al., 2014). Different from the above proteins, the S protein of human CoVs mediates viral entry into host cells and subsequent membrane fusion, enabling viral infection (Du et al., 2009a; Lu et al., 2014). The S protein is a class I viral protein, which can be cleaved into two functional subunits, an amino-terminal S1 subunit and a carboxyl-terminal S2 subunit. The S1 subunit is responsible for virus–host cell receptor binding, whereas the S2 subunit is involved in virus–host membrane fusion (Li et al., 2005a; Lu et al., 2014). The S1 contains two major domains, an N-terminal domain (NTD) and a C-terminal domain (CTD). In general, NTDs mediate sugar binding (Li, 2016; Li et al., 2017; Ou et al., 2017; Hulswit et al., 2019; Tortorici et al., 2019), whereas CTDs facilitate protein receptor recognition (Wong et al., 2004; Hofmann et al., 2006; Lu et al., 2013). The NTDs and CTDs of the S1 subunit can bind host receptors or function as receptor-binding domains (RBDs) (Lu et al., 2013; Hulswit et al., 2019). The entry of human CoVs relies on the interaction between viral and cellular membrane proteins. Recognition of S1 subunit with a receptor and/or sugar on the cell surface initiates the infection (Li, 2015). After the initial recognition and binding, the S protein undergoes conformational changes, followed by membrane fusion through the S2 region (Li, 2015, 2016; Du et al., 2017). Consequently, the viral genetic materials are delivered into the host cell through the fusion core (Du et al., 2009a).
Similar to HCoV-NL63 (Hofmann et al., 2005; Li, 2015), SARS-CoV recognizes, through the RBD in the CTD region of its S1 subunit, angiotensin-converting enzyme 2 (ACE2) as the receptor on the target cell (Li et al., 2003). The RBD (CTD) in two states (standing or lying) has been observed in the trimeric S protein. ACE2 binds to standing RBD, specifically in the receptor-binding motif (RBM), keeping the RBD in the “standing” state (Yuan et al., 2017). In addition to human ACE2, SARS-CoV S protein could also bind to palm civet and mouse ACE2s (Li et al., 2004; Li, 2008). Mutations in the RBD of S1 subunit are required for cross-species transmission of SARS-CoV (Li et al., 2005c; Li, 2008, 2016). Several bat SARSr-CoVs have been identified, and these CoVs can utilize human ACE2 as their receptor to bind the target cells (Ge et al., 2013; Hu et al., 2017). The structure of SARS-CoV S trimer and RBD binding to the ACE2 receptor is shown in Supplementary Figures S1A,B.
MERS-CoV RBD shares a structure similar to that of the homology domain of SARS-CoV (Wang et al., 2013). However, antibodies induced by SARS-CoV RBD have no cross-reactivity and/or cross-neutralizing activity to MERS-CoV (Du et al., 2013b). Moreover, MERS-CoV utilizes dipeptidyl peptidase 4 (DPP4) as a receptor through the RBD (CTD) region (Raj et al., 2013), which is distinct from the SARS-CoV receptor ACE2. Although the core regions of SARS-CoV and MERS-CoV RBDs are similar, their RBM regions are significantly different, which explains why they recognize different receptors (Li, 2015). MERS-CoV S trimer also maintains a structure similar to that of SARS-CoV S trimer. Both standing and lying states can be detected in the MERS-CoV RBDs, whereas DPP4 only binds to the standing RBD (Pallesen et al., 2017; Yuan et al., 2017). MERS-CoV is clustered with Ty-BatCoV HKU4 and Pi-BatCoV HKU5 in the subgenus Merbecovirus. Ty-BatCoV HKU4, but not Pi-BatCoV HKU5, could use human DPP4 as a receptor (Yang et al., 2014b). Recently, some MERS-related CoVs (MERSr-CoVs) have been discovered from bats that can enter human DPP4-expressing cells (Lau et al., 2018; Luo et al., 2018a). These findings suggest that the emergence of MERSr-CoV may threaten human health owing to their potential for cross-species transmission. The MERS-CoV S protein and RBD and their complexes, along with the DPP4 receptor, are shown in Supplementary Figures S1C,D.
Recent studies have found that the new human CoV, 2019-nCoV, which belongs to the species of SARSr-CoV, shares high sequence identify (about 79.5%) to SARS-CoV (Zhou et al., 2020). The genome of 2019-nCoV encodes pp1ab (translated from ORF 1ab), four structural proteins (S, E, M, and N), and six accessory proteins (3a, 6, 7a, 7b, 8, and 10) (Figure 2A). Same as SARS-CoV and MERS-CoV, 2019-nCoV appears to have no HE gene. The virion of 2019-nCoV consists of similar structure as SARS-CoV and MERS-CoV (Figure 2B). Like SARS-CoV, 2019-nCoV also uses ACE2 as its cellular receptor to enter host cells (Zhou et al., 2020). Currently, the structure of 2019-nCoV RBD and/or its binding with the viral receptor has not yet been available.
Overview of Vaccines Against Emerging Pathogenic Human Coronaviruses
Unlike the four low pathogenic human CoVs, including HCoV-229E, HCoV-NL63, HCoV-OC43, and HCoV-HKU1, which cause mild to no pathogenesis in humans, SARS-CoV, MERS-CoV, and 2019-CoV are three highly pathogenic human CoVs (Channappanavar and Perlman, 2017; Cui et al., 2019; Zhu et al., 2020). With the increasing numbers of 2019-nCoV and MERS-CoV infections and continuous threat of re-emergence of SARS-CoV, as well as the potential of SARS- and MERS-related CoVs to cause human infections, it is critical to develop vaccines with strong efficacy and safety targeting these viruses to prevent their infections in humans. Since the vaccines against 2019-nCoV have not been available, the rest of the review will focus on the vaccines against SARS-CoV and MERS-CoV.
Although a variety of vaccines have been developed against SARS-CoV and MERS-CoV, most of them are in the preclinical studies, and only several have been tested in clinical trials4,5 (Du et al., 2016b; Cho et al., 2018). Nevertheless, no vaccines have been approved for the prevention of SARS and MERS in humans, demonstrating the need to develop effective and safe vaccines to control current MERS-CoV infection, or to be stockpiled for potential use against re-emerged SARS-CoV or SARSr-CoV. Particularly, effective and safe vaccines are urgently needed to prevent and control the current outbreak of 2019-nCoV.
Most SARS-CoV and MERS-CoV vaccines developed thus far are based on the inactivated or live attenuated viruses, DNAs, proteins, nanoparticles, viral vectors, including virus-like particles (VLPs) (Zeng et al., 2004; Jiang et al., 2005; Liu et al., 2005; Du et al., 2009a, 2016b; Pimentel et al., 2009; Al-Amri et al., 2017). Each vaccine type has different advantages and disadvantages. For instance, inactivated and live-attenuated virus-based vaccines are vaccine types developed using the most traditional approaches. Although they generally induce highly potent immune responses and/or protection, the possibility for incomplete inactivation of viruses or recovering virulence exists, resulting in significant safety concerns (Zhang et al., 2014). Also, these traditional vaccines may induce the antibody-dependent enhancement (ADE) effect, as in the case of SARS-CoV infection (Luo et al., 2018b). Similarly, some viral-vectored vaccines can elicit specific antibody and cellular immune responses with neutralizing activity and protection, but they might also induce anti-vector immunity or present pre-existing immunity, causing some harmful immune responses. Instead, DNA and nanoparticle vaccines maintain strong safety profile; however, the immunogenicity of these vaccines is usually lower than that of virus- or viral vector-based vaccines, often requiring optimization of sequences, components, or immunization routes, inclusion of appropriate adjuvants, or application of combinational immunization approaches (Zhang et al., 2014).
Subunit Vaccines Against Sars-CoV and Mers-CoV
Subunit vaccines are vaccines developed based on the synthetic peptides or recombinant proteins. Unlike inactivated or live-attenuated virus and some viral vectored vaccines, this vaccine type mainly contains specific viral antigenic fragments, but without including any components of infectious viruses, eliminating the concerns of incomplete inactivation, virulence recovery, or pre-existing immunity (Du et al., 2008; Deng et al., 2012). Similar to DNA or VLP-based vaccines, subunit vaccines are generally safe without causing potential harmful immune responses, making them promising vaccine candidates. Moreover, subunit vaccines may target specific, well-defined neutralizing epitopes with improved immunogenicity and/or efficacy (Du et al., 2008; Zhang et al., 2014).
A number of subunit vaccines against SARS-CoV and MERS-CoV have been developed, and these are described in detail in the next paragraphs. The targets used for the development of SARS-CoV and MERS-CoV subunit vaccines are also be discussed.
Potential Targets for Development of SARS-CoV and MERS-CoV Subunit Vaccines
The S protein of SARS-CoV and MERS-CoV plays a vital role in receptor binding and membrane fusion. Thus, the S protein, but not other structural proteins, is the major antigen to induce protective neutralizing antibodies to block viruses from binding their respective receptor and thus inhibit viral infection (Bisht et al., 2004; Buchholz et al., 2004; Bukreyev et al., 2004; Yang et al., 2004). As a result, the S protein is also a major target for the development of subunit vaccines against SARS-CoV and MERS-CoV. Both full-length S protein and its antigenic fragments, including S1 subunit, NTD, RBD, and S2 subunit, can serve as important targets for the development of subunit vaccines (Guo et al., 2005; Mou et al., 2013; Wang et al., 2015; Jiaming et al., 2017; Zhou et al., 2018).
Although subunit vaccines based on the full-length S protein may elicit potent immune responses and/or protection, studies have found that antibodies induced by some of these vaccines mediate enhancement of viral infection in vitro, as in the case of SARS-CoV (Kam et al., 2007; Jaume et al., 2012), raising safety concerns for the development of full-length S protein-based subunit vaccines against SARS-CoV and MERS-CoV. In contrast, RBD-based subunit vaccines comprise the major critical neutralizing domain (Du and Jiang, 2015; Zhou et al., 2019). Therefore, these vaccines may generate potent neutralizing antibodies with strong protective immunity against viral infection. S1 subunit, for example, is much shorter than the full-length S protein, but it is no less able to induce strong immune responses and/or protection against viral infection (Li et al., 2013; Adney et al., 2019). Thus, this fragment can be used as an alternative target for subunit vaccine development. Despite their ability to induce immune responses and/or neutralizing antibodies, NTD and S2 as the targets of subunit vaccines are less immunogenic, eliciting significantly lower antibody titers, cellular immune responses, and/or protection than the other regions, such as full-length, S1, and RBD (Guo et al., 2005; Jiaming et al., 2017). Therefore, in terms of safety and efficacy, the RBD and/or S1 of S protein could be applied as critical targets for the development of subunit vaccine candidates against SARS-CoV, MERS-CoV, SARSr-CoV, and MERSr-CoV. Because of its conserved amino acid sequences and high homology among different virus strains (Elshabrawy et al., 2012; Zhou et al., 2018), the S2 subunit has potential to be used as a target for the development of universal vaccines against divergent virus strains.
In addition to the S protein, the N protein of SARS-CoV and MERS-CoV may serve as an additional target for the development of subunit vaccines. Unlike S protein, the N protein has no ability to elicit neutralizing antibodies to block virus-receptor interaction and neutralize viral infection, but it may induce specific antibody and cellular immune responses (Liu et al., 2006; Zheng et al., 2009). Several immunodominant B-cell and T-cell epitopes have been identified in the N protein of SARS-CoV and MERS-CoV, some of which are conserved in mice, non-human primates, and humans (Liu et al., 2006; Chan et al., 2011; Veit et al., 2018). Other proteins, such as M protein, can be used as potential targets of SARS-CoV and MERS-CoV subunit vaccines. Notably, SARS-CoV M protein-derived peptides have immunogenicity to induce high-titer antibody responses in the immunized animals (He et al., 2005b), suggesting the potential for utilizing this protein to develop subunit vaccines.
Subunit Vaccines Against SARS-CoV
Numerous subunit vaccines against SARS-CoV have been developed since the outbreak of SARS, the majority of which use the S protein and/or its antigenic fragments, in particular, RBD, as the vaccine target (Table 1).
TABLE 1
| Name | Antigenicity and functionality | Adjuvant | Route | Animal models | Antibody response | Cellular immune response | Protection | References |
| Subunit vaccines based on SARS-CoV full-length or trimeric S protein | ||||||||
| FL-S and EC-S proteins | Bind to SARS-CoV S1, NTD, RBD, and S2-specific mAbs | MPL + TDM | S.C. | BALB/c mice | Elicit SARS-CoV S-specific Abs (IgG, > 1: 2 × 105), neutralizing (> 1:2.4 × 104) pseudotyped SARS-CoV (Tor2, GD03, and SZ3 strains) | N/A | N/A | He et al., 2006a |
| S andS-foldon proteins | N/A | TiterMax Gold; Alum Hydro+MPL | S.C. or I.M. | BALB/c mice | Elicit SARS-CoV S-specific Abs (IgG, > 1:104) in mice, neutralizing (∼2.4 × 102 for S; ∼1:7 × 102 for S-foldon) live SARS-CoV (Urbani strain) | N/A | Protect vaccinated mice from challenge of SARS-CoV (Urbani strain, 105 TCID50) with undetectable viral load in lungs | Li et al., 2013 |
| triSpike protein | N/A | Alum hydro | I.P. or S.C. | BALB/c mice; Hamsters | Elicits SARS-CoV S-specific mucosal and serum Abs (IgA and IgG) in mice and hamsters, blocking S-ACE2 receptor binding and neutralizing live SARS-CoV (HKU-39849 strain); induces ADE | N/A | Protects vaccinated hamsters from challenge of SARS-CoV (Urbani strain, 103 TCID50) with undetectable or reduced viral load in lungs | Kam et al., 2007; Jaume et al., 2012 |
| Subunit vaccines based on SARS-CoV RBD protein | ||||||||
| RBD-Fc protein | N/A | Freund’s | I.D. or I.M. | BALB/c mice; Rabbits | Elicits SARS-CoV S/RBD-specific Abs (IgG) in mice and rabbits, neutralizing pseudotyped (rabbits: ≥ 7.3 × 104) and live (mice: 1:4 × 103; rabbits: > 1:1.5 × 104) SARS-CoV (BJ01 strain) | N/A | Protects majority (4/5) of vaccinated mice from challenge of SARS-CoV (BJ01 strain, 106 TCID50), with one mouse showing mild alveolar damage in lungs | He et al., 2004; Du et al., 2007 |
| RBD193-CHO; RBD219-CHO proteins | Binds to SARS-CoV RBD-specific mAbs (neutralizing 24H8, 31H12, 35B5, 33G4, 19B2; non-neutralizing 17H9) | Freund’s | S.C. | BALB/c mice | Elicit SARS-CoV RBD-specific Abs, neutralizing pseudotyped (< 1:104 for RBD193-CHO; 1:5.8 × 104 for RBD219-CHO) and live (< 1:103 for RBD193-CHO; 1:103 for RBD219-CHO) SARS-CoV (GZ50 strain) | Induce SARS-CoV RBD-specific cellular immune responses (IFN-γ, IL-2, IL-4, IL-10) in mice | Protect all (for RBD219-CHO) or majority (3/5, for RBD219-CHO) of vaccinated mice from challenge of SARS-CoV (GZ50 strain, 100 TCID50 for RBD193-CHO; 5 × 105 TCID50 for RBD219-CHO) with undetectable viral RNA or no, to reduced, viral load in lungs | Du et al., 2009c, 2010 |
| RBD-293T protein | Binds to SARS-CoV RBD-specific mAbs (neutralizing 24H8, 31H12, 35B5, 33G4, 19B2; non-neutralizing 17H9) | SAS | S.C. | BALB/c mice | Elicits SARS-CoV RBD-specific Abs (IgG), neutralizing pseudotyped (1:6.9 × 105) and live (1:1.6 × 103) SARS-CoV (GZ50 strain) | N/A | Protects all vaccinated mice from challenge of SARS-CoV (GZ50 strain, 100 TCID50) with undetectable viral RNA and viral load in lungs | Du et al., 2009b |
| S318-510 protein | N/A | Alum; Alum + CpG | S.C. | 129S6/SvEv mice | Elicits SARS-CoV-specific Abs (IgG, IgG1, and IgG2a) in mice. Reduces neutralization after removing glycosylation | Induces SARS-CoV S-specific cellular immune responses (IFN-γ) in mice | N/A | Zakhartchouk et al., 2007 |
| Subunit vaccines based on non-RBD SARS-CoV S protein fragments | ||||||||
| S1 and S1-foldon proteins | N/A | TiterMax Gold; Alum Hydro + MPL | S.C. or I.M. | BALB/c mice | Elicit SARS-CoV S-specific Abs (IgG, > 1:104) in mice, neutralizing (1:1.7 × 102 for S1; 1:90 for S1-foldon) live SARS-CoV (Urbani strain) | N/A | Protect vaccinated mice from challenge of SARS-CoV (Urbani strain, 105 TCID50) with undetectable viral load in lungs | Li et al., 2013 |
| S2 protein | N/A | Freund’s | S.C. | BALB/c mice | Elicits SARS-CoV S2-specific Abs (IgG, 1:1.6 × 103) in mice with no neutralizing activity | Induces SARS-CoV S2-specific cellular immune responses (IFN-γ and IL-4) in mice | N/A | Guo et al., 2005 |
| Subunit vaccines based on SARS-CoV non-S structural proteins (i.e. N and M) | ||||||||
| rN protein | N/A | Freund’s | I.P. | BALB/c mice | Elicits SARS-CoV N-specific Abs (IgG (1:1.8 × 103), IgG1, and IgG2a) in mice | Induces cellular immune responses with up-regulated IFN-γ and IL-10 cytokines in mice | N/A | Zheng et al., 2009 |
| rN protein | N/A | Montanide + CpG; Freund’s | S.C. | BALB/c mice | Elicits SARS-CoV N-specific Abs (IgG) in mice | Induces SARS-CoV N-specific cellular immune responses (IFN- γ) in mice | N/A | Liu et al., 2006 |
| M1-31 and M132-161 peptides | Bind to sera from SARS patients or immunized mice and rabbits | Freund’s | I.D. | BALB/c mice; NZW rabbits | Induce SARS-CoV M-specific Abs (IgG) in rabbits | N/A | N/A | He et al., 2005b |
Subunit Vaccines against SARS-CoVa.
aAbs, antibodies; ADE, antibody-dependent enhancement; Alum hydro, aluminum hydroxide; CHO, Chinese hamster ovary; CpG, cysteine-phosphate-guanine; I.D., intradermal; I.M., intramuscular; IFN-γ, interferon gamma; IL-2, interleukin 2; IL-4, interleukin 4; IL-10, Interleukin 10; I.P., intraperitoneal; mAbs, monoclonal antibodies; Montanide, Montanide ISA-51; MPL + TDM, monophosphoryl lipid A and trehalose dicorynomycolate; N/A, not reported; NTD, N-terminal domain; NZW rabbits, New Zealand White rabbits; RBD, receptor-binding domain; SAS, Sigma adjuvant system; S.C., subcutaneous; TCID50, median tissue culture infectious dose.
SARS-CoV Subunit Vaccines Based on Full-Length S Protein
Subunit vaccines based on SARS-CoV S protein, including full-length or trimeric S protein, are immunogenic with protection against SARS-CoV infection (He et al., 2006a; Kam et al., 2007; Li et al., 2013). Either insect cell-expressed full-length (FL-S) or extracellular domain (EC-S) SARS-CoV S protein developed high-titer S-specific antibodies with neutralizing activity against pseudotyped SARS-CoV expressing S protein of representative SARS-CoV human and palm civet strains (Tor2, GD03, and SZ3) isolated during the 2002 and 2003 or 2003 and 2004 outbreaks (He et al., 2006a). In addition, full-length S-ectodomain proteins fused with or without a foldon trimeric motif (S or S-foldon) could elicit specific antibody responses and neutralizing antibodies, protecting immunized mice against SARS-CoV challenge with undetectable virus titers in the lungs (Li et al., 2013). Moreover, a subunit vaccine (triSpike) based on a full-length S protein trimer induced specific serum and mucosal antibody responses and efficient neutralizing antibodies against SARS-CoV infection (Kam et al., 2007). Nevertheless, this vaccine also resulted in Fcγ receptor II (FcγRII)-dependent and ACE2-independent ADE, particularly in human monocytic or lymphoblastic cell lines infected with pseudotyped SARS-CoV expressing viral S protein, or in Raji B cells (B-cell lymphoma line) infected with live SARS-CoV (Kam et al., 2007; Jaume et al., 2012), raising significant concerns over the use of full-length S protein as a SARS vaccine target.
SARS-CoV Subunit Vaccines Based on RBD
SARS-CoV RBD contains multiple conformation-dependent epitopes capable of eliciting high-titer neutralizing antibodies; thus, it is a major target for the development of SARS vaccines (He et al., 2004, 2005a; Jiang et al., 2012; Zhu et al., 2013). Subunit vaccines based on the SARS-CoV RBD have been extensively explored. Studies have found that a fusion protein containing RBD and the fragment crystallizable (Fc) region of human IgG1 (RBD-Fc) elicited highly potent neutralizing antibodies against SARS-CoV in the immunized rabbits and mice, which strongly blocked the binding between S1 protein and SARS-CoV receptor ACE2 (He et al., 2004). This RBD protein induced long-term, high-level SARS-CoV S-specific antibodies and neutralizing antibodies that could be maintained for 12 months after immunization, protecting most of the vaccinated mice against SARS-CoV infection (Du et al., 2007). In addition, recombinant RBDs (residues 318–510 or 318–536) stably or transiently expressed in Chinese hamster ovary (CHO) cells bound strongly to RBD-specific monoclonal antibodies (mAbs), elicited high-titer anti-SARS-CoV neutralizing antibodies, and protected most, or all, of the SARS-CoV-challenged mice, with undetectable viral RNA and undetectable or significantly reduced viral load (Du et al., 2009c, 2010). Significantly, a 293T cell-expressed RBD protein maintains excellent conformation and good antigenicity to bind SARS-CoV RBD-specific neutralizing mAbs. It elicited highly potent neutralizing antibodies that completely protected immunized mice against SARS-CoV challenge (Du et al., 2009b). Particularly, RBDs from the S proteins of Tor2, GD03, and SZ3, representative strains of SARS-CoV isolated from human 2002–2003, 2003–2004, and palm civet strains, can induce high-titer cross-neutralizing antibodies against pseudotyped SARS-CoV expressing respective S proteins (He et al., 2006c). Different from the full-length S protein-based SARS subunit vaccines, no obvious pathogenic effects have been identified in the RBD-based SARS subunit vaccines (Kam et al., 2007; Jaume et al., 2012).
SARS-CoV Subunit Vaccines Based on Non-RBD S Protein Fragments
SARS subunit vaccines based on S protein fragments (S1 and S2), other than the RBD, have shown immunogenicity and/or protective efficacy against SARS-CoV infection (Guo et al., 2005; Li et al., 2013). For example, recombinant S1 proteins fused with or without foldon elicited specific antibodies with neutralizing activity that protected immunized mice against high-dose SARS-CoV challenge (Li et al., 2013). Although some studies have demonstrated that recombinant SARS-CoV S2 (residues 681–980) protein elicits specific non-neutralizing antibody response in mice (Guo et al., 2005), others have indicated that mAbs targeting highly conserved heptad repeat 1 (HR1) and HR2 domains of SARS-CoV S protein have broad neutralizing activity against pseudotyped SARS-CoV expressing S protein of divergent strains (Elshabrawy et al., 2012), indicating the potential of utilizing the S2 region as a broad-spectrum anti-SARS-CoV vaccine target (Zheng et al., 2009).
SARS-CoV Subunit Vaccines Based on Non-S Structural Proteins
Subunit vaccines based on the N and M proteins of SARS-CoV have shown immunogenicity in vaccinated animals (Liu et al., 2006; Zheng et al., 2009). Studies have revealed that a plant-expressed SARS-CoV N protein conjugated with Freund’s adjuvant elicited specific IgG antibodies, including IgG1 and IgG2a subtypes, and cellular immune responses in mice, whereas another E. coli-expressed N protein conjugated with Montanide ISA-51 and cysteine-phosphate-guanine (CpG) adjuvants induced specific IgG antibodies toward a Th1 (IgG2a)-type response in mice (Liu et al., 2006; Zheng et al., 2009). Although N-specific antibodies have been detected in convalescent-phase SARS patient and immunized rabbit sera, they have no neutralizing activity against SARS-CoV infection (Qiu et al., 2005). In addition, immunodominant M protein peptides (M1-31 and M132-161) identified using convalescent-phase sera of SARS patients and immunized mouse and rabbit sera have immunogenicity to elicit specific IgG antibodies in rabbits (He et al., 2005b). In spite of their immunogenicity, it appears that these N- and M-based SARS subunit vaccines have not been investigated for their protective efficacy against SARS-CoV infection. Thus, it is unclear whether these non-S structural protein-based SARS subunit vaccines can prevent SARS-CoV infection.
Potential Factors Affecting SARS-CoV Subunit Vaccines
A number of factors may affect the expression of proteins to be used as SARS subunit vaccines; apart from their immunogenicity and/or protective efficacy. Understanding of these factors is important to generate subunit vaccines with good quality, high immunogenicity, and excellent protection against SARS-CoV infection.
The expression of recombinant protein-based SARS subunit vaccines may be changed by the following factors. First, addition of an intron splicing enhancer to the truncated SARS-CoV S protein fragments results in better enhancement of protein expression in mammalian cells than the exon splicing enhancers, and different cells may result in different fold increase of protein expression (Chang et al., 2006). Second, inclusion of a post-transcriptional gene silencing suppressor p19 protein from tomato bushy stunt virus to a SARS-CoV N protein may significantly increase its transient expression in tobacco (Zheng et al., 2009).
The following factors may affect the immunogenicity and protective efficacy of protein-based SARS subunit vaccines, including same proteins expressed in different expression systems, and same proteins with various lengths, amino acid mutations, or deletions (He et al., 2006b; Du et al., 2009b). For example, RBD proteins containing different lengths (193-mer: RBD193-CHO or 219-mer: RBD219-CHO) elicited different immune responses and protective efficacy against SARS-CoV challenge (Du et al., 2009c, 2010). A recombinant SARS-CoV RBD (RBD-293T) protein expressed in mammalian cell system was able to induce stronger neutralizing antibody response than those expressed in insect cells (RBD-Sf9) and E. coli (RBD-Ec) (Du et al., 2009b), suggesting that RBD purified from mammalian cells has preference for further development due to its ability to maintain native conformation. Notably, a single mutation (R441A) in the RBD of SARS-CoV disrupted its major neutralizing epitopes and affinity to bind viral receptor ACE2, thus abolishing the vaccine’s immunogenicity, and hence, its ability to induce neutralizing antibodies in immunized animals (He et al., 2006b). Additionally, deletion of a particular amino acid by changing a glycosylation site in the SARS-CoV RBD (RBD219-N1) also resulted in the alteration of subunit vaccine’s immunogenicity (Chen et al., 2014).
Other factors that potentially affect the immunogenicity of SARS subunit vaccines include immunization routes and adjuvants (Zakhartchouk et al., 2007; Li et al., 2013). Significantly high-titer antibodies were induced by monomeric or trimeric SARS-CoV S and S1 proteins through the intramuscular (I.M.) route compared to the subcutaneous (S.C.) route (Li et al., 2013). Moreover, a SARS-CoV RBD subunit vaccine conjugated with Alum plus CpG adjuvants elicited a higher level of IgG2a antibody and interferon gamma (IFN-γ) secretion than the RBD with Alum alone (Zakhartchouk et al., 2007).
Subunit Vaccines Against MERS-CoV
Subunit vaccines against MERS-CoV have been developed extensively, almost all of which are based on the S protein, including full-length S timer, NTD, S1, and S2, particularly RBD. These subunit vaccines, including their antigenicity, functionality, immunogenicity, and protective efficacy in different animal models, are summarized in Table 2.
TABLE 2
| Name | Functionality and antigenicity | Adjuvant | Route | Animal models | Antibody response | Cellular immune response | Protective efficacy | References |
| Subunit vaccines based on MERS-CoV full-length S protein | ||||||||
| MERS S-2P protein | Binds to DPP4 receptor and MERS-CoV S-NTD, RBD, and S2-specific neutralizing mAbs (G2, D12, and G4, respectively) | SAS | I.M. | BALB/c mice | Elicits neutralizing Abs in mice, neutralizing 7 pseudotyped MERS-CoV | N/A | N/A | Pallesen et al., 2017 |
| Subunit vaccines based on MERS-CoV RBD protein | ||||||||
| rRBD (S367-606) protein | N/A | Alum Hydro + CpG or poly(I:C); IFA + CpG (mouse); Alum (NHPs) | I.M. or S.C. | BALB/c mice; NHPs | Elicits MERS-CoV RBD-specific Abs in mice (IgG, IgG1, IgG2a, and IgG2b) and NHPs (IgG), neutralizing pseudotyped (mouse: < 1:5 × 102) and live (NHPs: < 1:5 × 102) MERS-CoV (EMC2012 strain) | Induces MERS-CoV RBD-specific cellular immune responses (IFN-γ, TNF-α, IL-2, IL-4, IL-6, and IL-10) in mice and/or monkeys | Partially protects vaccinated NHPs from challenge of MERS-CoV (EMC2012 strain, 6.5 × 107 TCID50) with alleviated pneumonia and decreased viral load | Lan et al., 2014, 2015 |
| RBD (S377-662)-Fc protein | Binds to DPP4 receptor | Poly(I:C); Montanide | I.N. or S.C. | BALB/c mice | Elicits MERS-CoV S1- and RBD-specific Abs (IgA, IgG (> 1:104), IgG1, IgG2a, and IgG3) in mice, neutralizing (≥ 1:2.4 × 102) live MERS-CoV (EMC2012 strain) | Induces MERS-CoV S1-specific cellular immune responses (IFN-γ and IL-2) in mice | N/A | Du et al., 2013c; Ma et al., 2014b |
| RBD (S377-588)-Fc protein | Binds to DPP4 receptor and MERS-CoV RBD specific neutralizing mAbs (Mersmab1, m336, m337, and m338) | Montanide; MF59; AddaVax | I.M. or S.C. | BALB/c mice; hDPP4-Tg mice; Rabbits | Elicits MERS-CoV S1 and RBD-specific Abs in mice (IgG (> 1:105), IgG1, and IgG2a) and rabbits (IgG), neutralizing 17 pseudotyped (≥ 1:104) and 2 live (≥ 1:103) MERS-CoV (EMC2012 and London1-2012 strains) | Induces MERS-CoV S1-specific cellular immune responses (IFN-γ and IL-2) in mice | Protects vaccinated Ad5/hDPP4-transduced BALB/c mice and majority (4/6) of vaccinated hDPP4-Tg mice from MERS-CoV (EMC2012 strain, 105 PFU for BALB/c; 103–4 TCID50 for Tg) challenge, without immunological toxicity or eosinophilic immune enhancement | Du et al., 2013a; Ma et al., 2014b; Tang et al., 2015; Zhang et al., 2016; Nyon et al., 2018 |
| RBD-Fd protein | Binds to DPP4 receptor and MERS-CoV RBD-specific neutralizing mAbs (Mersmab1, m336, m337, and m338) | MF59; Alum | I.M. or S.C. | BALB/c mice; hDPP4-Tg mice | Elicits MERS-CoV S1-specific Abs (IgG (> 1:105), IgG1, and IgG2a) in mice, neutralizing at least 9 pseudotyped (> 1:104) and live (> 1:103) MERS-CoV (EMC2012 strain) | N/A | Protects majority (5/6) of vaccinated hDPP4-Tg mice from challenge of MERS-CoV (EMC2012 strain, 104 TCID50) | Tai et al., 2016 |
| RBD (T579N) protein | Binds to receptor DPP4 and MERS-CoV RBD-specific neutralizing mAbs (hMS-1, m336, m337, and m338) | Montanide; Alum | I.M. or S.C. | BALB/c mice; hDPP4-Tg mice | Elicits neutralizing Abs (> 1:3 × 103) in mice against live MERS-CoV (EMC2012 strain) | N/A | Protects all vaccinated hDPP4-Tg mice from challenge of MERS-CoV (EMC2012 strain, 104 TCID50) | Du et al., 2016a |
| Subunit vaccines based on non-RBD MERS-CoV S protein fragments | ||||||||
| S1 protein | N/A | Ribi; Alum pho | I.M. | BALB/c mice; NHPs | Elicits MERS-CoV S1-specific Abs in mice (IgG and IgG1) and NHPs (IgG), neutralizing 8 pseudotyped and live MERS-CoV (JordanN3 strain) | N/A | Protects vaccinated NHPs from challenge of MERS-CoV (JordanN3 strain, 5 × 106 PFU) with reduced abnormalities on chest CT | Wang et al., 2015 |
| S1 protein | N/A | Advax + SAS | I.M. | Dromedary camels; Alpacas | Elicits neutralizing Abs in dromedary camels (≥ 1:80) and alpacas (≥ 1:6.4 × 102) against live MERS-CoV (EMC2012 strain) | N/A | Protects vaccinated dromedary camels and alpacas from challenge of MERS-CoV (EMC2012 strain, 107 TCID50) with reduced and delayed viral shedding in the upper airways (in camels) or complete protection (in alpacas) | Adney et al., 2019 |
| rNTD protein | N/A | Alum pho + CpG | I.M. | BALB/c mice; Ad5-hDPP4 mice | Elicits MERS-CoV S-NTD-specific Abs (IgG, ≥ 1:104) in mice, neutralizing pseudotyped and live (1:40) MERS-CoV (EMC2012 strain) | Induces MERS-CoV S-NTD-specific cellular immune responses (IFN-γ, IL-2, IL-6, IL-10, and IL-17A) in mice | Protects vaccinated Ad5-hDPP4-transduced mice from challenge of MERS-CoV (EMC2012 strain, 105 PFU) with reduced lung abnormalities and respiratory tract pathology | Jiaming et al., 2017 |
| SP3 peptide (aa736-761) | N/A | Freund’s | N/A | BALB/c mice; NZW rabbits | Elicits MERS-CoV S-specific Abs (IgG, 1:104) in rabbits, neutralizing pseudotyped MERS-CoV | N/A | N/A | Yang et al., 2014a |
Subunit Vaccines against MERS-CoVa.
aaa, amino acid; Abs, antibodies; Ad5, adenovirus serotype 5; Ad5-hDPP4 mice, Ad5-hDPP4-transuced mice; Alum hydro, aluminum hydroxide; Alum pho, Aluminum phosphate; hDPP4, human dipeptidyl peptidase 4; hDPP4-Tg mice, transgenic mice expressing MERS-CoV receptor human DPP4; IFA, incomplete Freund’s adjuvant; I.M., intramuscular; I.N., intranasal; mAbs, monoclonal antibodies; Montanide, Montanide ISA51; N/A, not reported; NHPs, non-human primates; NZW, rabbits, New Zealand White rabbits; PFU, plaque-forming unit; rRBD, recombinant RBD; SAS, Sigma Adjuvant System; S.C., subcutaneous; TCID50, median tissue culture infectious dose; TNF-α, tumor necrosis factor (TNF)-alpha.
MERS-CoV Subunit Vaccines Based on Full-Length S Protein
Subunit vaccines based on the full-length S protein cover both RBD and non-RBD neutralizing epitopes, some of which may be located in the conserved S2 subunit; thus this type of subunit vaccines are expected to induce high-titer neutralizing antibodies. Although several MERS-CoV full-length S protein-based vaccines have been reported in other vaccine types, including viral vectors and DNAs (Wang et al., 2015; Wang C. et al., 2017; Haagmans et al., 2016; Zhou et al., 2018), only a few subunit vaccines have been developed that rely on the full-length S protein. For example, a recombinant MERS-CoV S protein trimer (MERS S-2P) in prefusion conformation binds to the DPP4 receptor, as well as to the MERS-CoV NTD, RBD, and S2-specific neutralizing mAbs (Pallesen et al., 2017). Whereas this protein induces neutralizing antibodies in mice against divergent pseudotyped MERS-CoV in vitro, its in vivo protective activity against MERS-CoV infection is unknown (Pallesen et al., 2017). Therefore, more studies are needed to elucidate the potential for the development of MERS-CoV full-length S-based subunit vaccines, including understanding their protective efficacy and identifying possible harmful immune responses.
MERS-CoV Subunit Vaccines Based on RBD
Numerous MERS-CoV RBD-based subunit vaccines have been developed and extensively evaluated in available animal models since the emergence of MERS-CoV (Table 2) (Du et al., 2013c; Tai et al., 2017; Zhou et al., 2018). In general, these subunit vaccines have strong immunogenicity and are capable of inducing high neutralizing antibodies and/or protection against MERS-CoV infection (Ma et al., 2014b; Zhang et al., 2016; Tai et al., 2017; Wang Y. et al., 2017). Most subunit vaccines based on the MERS-CoV RBD have been described in detail in a previous review article (Zhou et al., 2019). In this section, we will briefly introduce these RBD-targeting MERS vaccines, and compare their functionality, antigenicity, immunogenicity, and protection against MERS-CoV infection.
Co-crystallographic analyses of MERS-CoV RBD and/or RBD/DPP4 complexes have confirmed that the RBD is attributed to residues 367–588 (Chen et al., 2013) or 367–606 (Lu et al., 2013) in the MERS-CoV S1 subunit. Indeed, a recombinant MERS-CoV RBD (rRBD) fragment (residues 367–606) elicits RBD-specific antibody and cellular immune responses and neutralizing antibodies in mice and/or non-human primates (NHPs) (Lan et al., 2014, 2015). However, it only partially protects NHPs from MERS-CoV infection by alleviating pneumonia and clinical manifestations, as well as decreasing viral load (Lan et al., 2015). In addition, an RBD protein fragment containing MERS-CoV S residues 377–622 fused with the Fc tag of human IgG can induce MERS-CoV S1- and/or RBD-specific humoral and cellular immune responses in the immunized mice with neutralizing activity against MERS-CoV infection (Du et al., 2013c; Jiang et al., 2013). However, after comparing several versions of MERS-CoV RBD fragments with different lengths, it was found that a truncated RBD (residues 377–588) had the highest DPP4-binding affinity and induced the highest-titer IgG antibodies and neutralizing antibodies against MERS-CoV, identifying its role as a critical neutralizing domain (Ma et al., 2014b).
Subsequently, several MERS-CoV subunit vaccines have been designed based on the identified critical neutralizing domain of RBD fragment, including those expressed in a stable CHO cell line (S377-588-Fc), fusing with a trimeric motif foldon (RBD-Fd), or containing single or multiple mutations in the RBD of representative human and camel strains from the 2012–2015 MERS outbreaks (Tai et al., 2016, 2017; Nyon et al., 2018). These RBD proteins maintain good conformation, functionality, antigenicity, and immunogenicity, with ability to bind the DPP4 receptor and RBD-specific neutralizing mAbs and to elicit robust neutralizing antibodies cross-neutralizing multiple strains of MERS pseudoviruses and live MERS-CoV (Tai et al., 2016, 2017; Nyon et al., 2018). It is noted that the wild-type MERS-CoV RBD proteins consisting of the identified critical neutralizing domain confer partial protection of hDPP4-transgenic (hDPP4-Tg) mice from MERS-CoV infection without causing immunological toxicity or eosinophilic immune enhancement (Tai et al., 2016; Wang Y. et al., 2017; Nyon et al., 2018); nevertheless, a structurally designed mutant version of such RBD protein with a non-neutralizing epitope masked (T579N) preserves intact conformation and significantly improves overall neutralizing activity and protective efficacy, resulting in the full protection of hDPP4-Tg mice against high-dose MERS-CoV challenge (Du et al., 2016a).
The above studies indicate that protein lengths to be chosen as MERS-CoV subunit vaccines and/or structure-based vaccine design can impact on the immunogenicity and/or protection of RBD-based subunit vaccines.
MERS-CoV Subunit Vaccines Based on Non-RBD S Protein Fragments
MERS vaccines targeting non-RBD regions of S protein have been developed and investigated in mice and NHPs. It has been shown that a MERS-CoV S1 protein formulated with Ribi (for mice) or aluminum phosphate (for NHPs) adjuvant elicited robust neutralizing antibodies in mice and NHPs against divergent strains of pseudotyped and live MERS-CoV, protecting NHPs from MERS-CoV infection (Wang et al., 2015). In addition, MERS-CoV S1 protein adjuvanted with Advax and Sigma Adjuvant System induced low-titer neutralizing antibodies in dromedary camels with reduced and delayed viral shedding after MERS-CoV challenge, but high-titer neutralizing antibodies in alpacas with complete protection of viral shedding from viral infection, indicating that protection of MERS-CoV infection is positively correlated with serum neutralizing antibody titers (Adney et al., 2019). Moreover, immunization with a recombinant MERS-CoV NTD protein (rNTD) can induce neutralizing antibodies and cell-mediated responses, protecting Ad-hDPP4-transduced mice against MERS-CoV challenge (Jiaming et al., 2017). Notably, specific antibodies with neutralizing activity have been elicited by a S2 peptide sequence (residues 736–761) of MERS-CoV in rabbits (Yang et al., 2014a), but the protective efficacy of this peptide vaccine is unknown. The above reports demonstrate the potential for the development of MERS subunit vaccines based on the non-RBD fragments of MERS-CoV S protein.
MERS-CoV Subunit Vaccines Based on Non-S Structural Proteins
Unlike SARS subunit vaccines which have been designed based on viral N and M proteins, it appears that very few subunit vaccines have been developed based on the non-S structural protein(s) of MERS-CoV. One study reports the induction of specific antibodies by MERS-CoV N peptides (Yang et al., 2014a), and another report shows that N protein is used for development of vaccines based on viral vector Vaccinia virus, modified Vaccinia Ankara (MVA) (Veit et al., 2018). This may be potentially a consequence of the weak immunogenicity and/or protective efficacy of non-S structural proteins, further confirming the role of MERS-CoV S protein as the key target for the development of MERS vaccines, including subunit vaccines.
Potential Factors Affecting MERS-CoV Subunit Vaccines
Similar to SARS-CoV subunit vaccines, the immunogenicity and/or protection of MERS-CoV subunit vaccines may also be affected by a number of factors, such as antigen sequences, fragment lengths, adjuvants, vaccination pathways, antigen doses, immunization doses and intervals used.
As described above, MERS-CoV subunit vaccines containing different antigens or fragment lengths, particularly those based on the RBD, have apparently variable immunogenicity and/or protective efficacy, and a critical neutralizing domain that contains an RBD fragment corresponding to residues 377–588 of S protein elicits the highest neutralizing antibodies among several fragments tested (Ma et al., 2014b; Zhang et al., 2015).
Adjuvants play an essential role in enhancing host immune responses to MERS-CoV subunit vaccines, including those based on the RBD, and different adjuvants can promote host immune responses to variant levels (Lan et al., 2014; Zhang et al., 2016). For example, while a MERS-CoV RBD subunit vaccine (S377-588 protein fused with Fc) alone induced detectable neutralizing antibody and T-cell responses in immunized mice, inclusion of an adjuvant enhanced its immunogenicity. Particularly, among the adjuvants (Freund’s, aluminum, Monophosphoryl lipid A, Montanide ISA51 and MF59) conjugated with this RBD protein, MF59 could best potentiate the protein to induce the highest-titer anti-S antibodies and neutralizing antibodies, protecting mice against MERS-CoV infection (Zhang et al., 2016). Moreover, a recombinant RBD (rRBD) protein plus alum and CpG adjuvants elicited the highest neutralizing antibodies against pseudotyped MERS-CoV infection, whereas the strongest T-cell responses were induced by this protein plus Freund’s and CpG adjuvants (Lan et al., 2014).
Vaccination pathways are important in inducing efficient immune responses, and different immunization routes may elicit different immune responses to the same protein antigens. For example, immunization of mice with a MERS-CoV subunit vaccine (RBD-Fc) via the intranasal route induced higher levels of cellular immune responses and stronger local mucosal neutralizing antibody responses against MERS-CoV infection than those induced by the same vaccine via the S.C. pathway (Ma et al., 2014a). In addition, while Freund’s and CpG-adjuvanted rRBD protein elicited higher-level systematic and local IFN-γ-producing T cells via the S.C. route, this protein adjuvanted with Alum and CpG induced higher-level tumor necrosis factor-alpha (TNF-α) and interleukin 4 (IL-4)-secreting T cells via the I.M. route (Lan et al., 2014).
Antigen dosage, immunization doses, and intervals may significantly affect the immunogenicity of MERS-CoV subunit vaccines. Notably, a MERS-CoV RBD (S377-588-Fc) subunit vaccine immunized at 1 μg elicited strong humoral and cellular immune responses and neutralizing antibodies in mice although the one immunized at 5 and 20 μg elicited a higher level of S1-specific antibodies (Tang et al., 2015). In addition, among the regimens at one dose and two doses at 1-, 2-, and 3-week intervals, 2 doses of this protein boosted at 4 weeks resulted in the highest antibodies and neutralizing antibodies against MERS-CoV infection (Wang Y. et al., 2017).
Potential Challenges and Future Perspectives for Sars-CoV and Mers-CoV Subunit Vaccines
Compared with other vaccine types such as inactivated virus and viral-vectored vaccines, SARS and MERS subunit vaccines are much safer and do not cause obvious side effects. However, these subunit vaccines may face some important challenges, mostly arising from their relatively low immunogenicity, which must be combined with appropriate adjuvants or optimized for suitable protein sequences, fragment lengths, and immunization schedules. In addition, structure and epitope-based vaccine design has become a promising strategy to improve the efficacy of subunit vaccines. This is evidenced by a structurally designed MERS-CoV RBD-based protein which has significantly improved neutralizing activity and protection against MERS-CoV infection (Du et al., 2016a). It is prospected that more structure-guided novel strategies will be developed to improve the overall immunogenicity and efficacy of subunit vaccines against emerging pathogenic human coronaviruses, including those targeting SARS-CoV and MERS-CoV. Although a large number of SARS and MERS subunit vaccines have been developed with potent immunogenicity and/or protection in available animal models, virtually all remain in the preclinical stage. It is thus expected that one or several of these promising subunit vaccines can be further processed into clinical trials to confirm their immunogenicity against viral infections in humans.
Rapid Development of Subunit Vaccines Against the New Pathogenic Human Coronavirus
Currently, the newly identified 2019-nCoV is spreading to infect people, resulting in significant global concerns. It is critical to rapidly design and develop effective vaccines to prevent infection of this new coronavirus. Since S protein and its fragments, such as RBD, of SARS-CoV, and MERS-CoV are prime targets for developing subunit vaccines against these two highly pathogenic human CoVs, it is expected that similar regions of 2019-nCoV can also be used as key targets for developing vaccines against this new coronavirus (Jiang et al., 2020). Similarly, other regions of 2019-nCoV, including S1 and S2 subunits of S protein and N protein, can be applied as alternative targets for vaccine development. Taken together, the approaches and strategies in the development of subunit vaccines against SARS and MERS described in this review will provide important information for the rapid design and development of safe and effective subunit vaccines against 2019-nCoV infection.
Statements
Author contributions
SJ and LD conceived the idea and revised and edited the manuscript. NW and LD collected information and drafted the manuscript. JS performed the structural analysis. All authors read and made final approval of the manuscript.
Funding
This work was supported by the NIH grants R01AI137472 and R01AI139092.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.00298/full#supplementary-material
FIGURE S1Structures of SARS-CoV and MERS-CoV S proteins, RBDs, and their complexes with respective receptor. Trimeric S proteins of SARS-CoV (PDB: 5×5b) (A) and MERS-CoV (PDB: 5×5f) (C) are colored differently for each monomer. Two conformations of the RBD in each trimeric S protein are labeled as standing and lying states. The RBDs of each S protein are shown as light blue on the right panel. ACE2 and DPP4 receptors are respectively modeled to the trimeric S proteins by match-alignment of SARS-CoV RBD-ACE2 complex (PDB: 2AJF) to SARS-CoV S trimer (B) or MERS-CoV RBD-DPP4 complex (PDB: 4kr0) to MERS-CoV S trimer (D). Each of the RBD-receptor complexes is shown on the right panel. ACE2, angiotensin-converting enzyme 2; DPP4, dipeptidyl peptidase 4; RBD, receptor-binding domain; S, spike.
Footnotes
1.^https://www.who.int/emergencies/mers-cov/en/
2.^https://www.who.int/docs/default-source/coronaviruse/situation-reports/20200219-sitrep-30-covid-19.pdf?sfvrsn=6e50645_2
3.^https://www.who.int/docs/default-source/coronaviruse/situation-reports/20200204-sitrep-15-ncov.pdf?sfvrsn=88fe8ad6_2
References
1
AdneyD. R.WangL.van DoremalenN.ShiW.ZhangY.KongW. P.et al (2019). Efficacy of an adjuvanted Middle East respiratory syndrome coronavirus spike protein vaccine in dromedary camels and alpacas.Viruses11:E212. 10.3390/v11030212
2
Al-AmriS. S.AbbasA. T.SiddiqL. A.AlghamdiA.SankiM. A.Al-MuhannaM. K.et al (2017). Immunogenicity of candidate MERS-CoV DNA vaccines based on the spike protein.Sci. Rep.7:44875. 10.1038/srep44875
3
AnnanA.BaldwinH. J.CormanV. M.KloseS. M.OwusuM.NkrumahE. E.et al (2013). Human betacoronavirus 2c EMC/2012-related viruses in Bats, Ghana and Europe.Emerg. Infect. Dis.19456–459. 10.3201/eid1903.121503
4
ArbourN.DayR.NewcombeJ.TalbotP. J. (2000). Neuroinvasion by human respiratory coronaviruses.J. Virol.748913–8921. 10.1128/jvi.74.19.8913-8921.2000
5
AzharE. I.El-KafrawyS. A.FarrajS. A.HassanA. M.Al-SaeedM. S.HashemA. M.et al (2014). Evidence for camel-to-human transmission of MERS coronavirus.N. Engl. J. Med.3702499–2505. 10.1056/NEJMoa1401505
6
Bailey-ElkinB. A.KnaapR. C.JohnsonG. G.DaleboutT. J.NinaberD. K.van KasterenP. B.et al (2014). Crystal structure of the Middle East respiratory syndrome coronavirus (MERS-CoV) papain-like protease bound to ubiquitin facilitates targeted disruption of deubiquitinating activity to demonstrate its role in innate immune suppression.J. Biol. Chem.28934667–34682. 10.1074/jbc.M114.609644
7
BishtH.RobertsA.VogelL.BukreyevA.CollinsP. L.MurphyB. R.et al (2004). Severe acute respiratory syndrome coronavirus spike protein expressed by attenuated vaccinia virus protectively immunizes mice.Proc. Natl. Acad. Sci. U.S.A.1016641–6646. 10.1073/pnas.0401939101
8
BuchholzU. J.BukreyevA.YangL. J.LamirandeE. W.MurphyB. R.SubbaraoK.et al (2004). Contributions of the structural proteins of severe acute respiratory syndrome coronavirus to protective immunity.Proc. Natl. Acad. Sci. U.S.A.1019804–9809. 10.1073/pnas.0403492101
9
BukreyevA.LamirandeE. W.BuchholzU. J.VogelL. N.ElkinsW. R.St ClaireM.et al (2004). Mucosal immunisation of African green monkeys (Cercopithecus aethiops) with an attenuated parainfluenza virus expressing the SARS coronavirus spike protein for the prevention of SARS.Lancet3632122–2127. 10.1016/s0140-6736(04)16501-x
10
ChanP. K.MaS.NgaiS. M. (2011). Identification of T-cell epitopes of SARS-coronavirus for development of peptide-based vaccines and cellular immunity assessment methods.Hong Kong Med. J.17 (Suppl. 6), 26–30.
11
ChangC. Y.HongW. W.ChongP.WuS. C. (2006). Influence of intron and exon splicing enhancers on mammalian cell expression of a truncated spike protein of SARS-CoV and its implication for subunit vaccine development.Vaccine241132–1141. 10.1016/j.vaccine.2005.09.011
12
ChannappanavarR.PerlmanS. (2017). Pathogenic human coronavirus infections: causes and consequences of cytokine storm and immunopathology.Semin. Immunopathol.39529–539. 10.1007/s00281-017-0629-x
13
ChenW. H.DuL.ChagS. M.MaC.TricocheN.TaoX.et al (2014). Yeast-expressed recombinant protein of the receptor-binding domain in SARS-CoV spike protein with deglycosylated forms as a SARS vaccine candidate.Hum. Vaccin. Immunother.10648–658. 10.4161/hv.27464
14
ChenY.RajashankarK. R.YangY.AgnihothramS. S.LiuC.LinY. L.et al (2013). Crystal structure of the receptor-binding domain from newly emerged Middle East respiratory syndrome coronavirus.J. Virol.8710777–10783. 10.1128/jvi.01756-13
15
ChiuS. S.ChanK. H.ChuK. W.KwanS. W.GuanY.PoonL. L.et al (2005). Human coronavirus NL63 infection and other coronavirus infections in children hospitalized with acute respiratory disease in Hong Kong, China.Clin. Infect. Dis.401721–1729. 10.1086/430301
16
ChoH.ExclerJ. L.KimJ. H.YoonI. K. (2018). Development of Middle East respiratory syndrome coronavirus vaccines - advances and challenges.Hum. Vaccin. Immunother.14304–313. 10.1080/21645515.2017.1389362
17
CuiJ.LiF.ShiZ. L. (2019). Origin and evolution of pathogenic coronaviruses.Nat. Rev. Microbiol.17181–192. 10.1038/s41579-018-0118-9
18
De GrootR. J.BakerS.BaricR.EnjuanesL.GorbalenyaA.HolmesK. V.et al (2011). “Family Coronaviridae, virus taxomomy: classification and nomenclature of viruses,” in Ninth Report of the International Committee on Taxonomy of Viruses, edsKingA. M. Q.AdamsM. J.CarstensE. B.LefkowitzE. J. (Amsterdam: Elsevier Academic Press), 806–826.
19
DeDiegoM. L.Nieto-TorresJ. L.Jimenez-GuardenoJ. M.Regla-NavaJ. A.Castano-RodriguezC.Fernandez-DelgadoR.et al (2014). Coronavirus virulence genes with main focus on SARS-CoV envelope gene.Virus. Res.194124–137. 10.1016/j.virusres.2014.07.024
20
DengM. P.HuZ. H.WangH. L.DengF. (2012). Developments of subunit and VLP vaccines against influenza A virus.Virol. Sin.27145–153. 10.1007/s12250-012-3241-1
21
DesforgesM.DesjardinsJ.ZhangC.TalbotP. J. (2013). The acetyl-esterase activity of the hemagglutinin-esterase protein of human coronavirus OC43 strongly enhances the production of infectious virus.J. Virol.873097–3107. 10.1128/jvi.02699-12
22
DesforgesM.Le CoupanecA.BrisonE.Meessen-PinardM.TalbotP. J. (2014). Neuroinvasive and neurotropic human respiratory coronaviruses: potential neurovirulent agents in humans.Adv. Exp. Med. Biol.80775–96. 10.1007/978-81-322-1777-0_6
23
DrexlerJ. F.CormanV. M.DrostenC. (2014). Ecology, evolution and classification of bat coronaviruses in the aftermath of SARS.Antiviral Res.10145–56. 10.1016/j.antiviral.2013.10.013
24
DrostenC.GuntherS.PreiserW.van der WerfS.BrodtH. R.BeckerS.et al (2003). Identification of a novel coronavirus in patients with severe acute respiratory syndrome.N. Engl. J. Med.3481967–1976. 10.1056/NEJMoa030747
25
DuL.HeY.JiangS.ZhengB. J. (2008). Development of subunit vaccines against severe acute respiratory syndrome.Drugs Today4463–73. 10.1358/dot.2008.44.1.1131830
26
DuL.HeY.ZhouY.LiuS.ZhengB. J.JiangS. (2009a). The spike protein of SARS-CoV - a target for vaccine and therapeutic development.Nat. Rev. Microbiol.7226–236. 10.1038/nrmicro2090
27
DuL.JiangS. (2015). Middle East respiratory syndrome: current status and future prospects for vaccine development.Expert Opin. Biol. Ther.151647–1651. 10.1517/14712598.2015.1092518
28
DuL.KouZ.MaC.TaoX.WangL.ZhaoG.et al (2013a). A truncated receptor-binding domain of MERS-CoV Spike protein potently inhibits MERS-CoV infection and induces strong neutralizing antibody responses: implication for developing therapeutics and vaccines.PLoS One8:e81587. 10.1371/journal.pone.0081587
29
DuL.MaC.JiangS. (2013b). Antibodies induced by receptor-binding domain in spike protein of SARS-CoV do not cross-neutralize the novel human coronavirus hCoV-EMC.J. Infect.67348–350. 10.1016/j.jinf.2013.05.002
30
DuL.TaiW.YangY.ZhaoG.ZhuQ.SunS.et al (2016a). Introduction of neutralizing immunogenicity index to the rational design of MERS coronavirus subunit vaccines.Nat. Commun.7:13473. 10.1038/ncomms13473
31
DuL.TaiW.ZhouY.JiangS. (2016b). Vaccines for the prevention against the threat of MERS-CoV.Expert Rev. Vaccines151123–1134. 10.1586/14760584.2016.1167603
32
DuL.YangY.ZhouY.LuL.LiF.JiangS. (2017). MERS-CoV spike protein: a key target for antivirals.Expert Opin. Ther. Targets21131–143. 10.1080/14728222.2017.1271415
33
DuL.ZhaoG.ChanC. C.LiL.HeY.ZhouY.et al (2010). A 219-mer CHO-expressing receptor-binding domain of SARS-CoV S protein induces potent immune responses and protective immunity.Viral. Immunol.23211–219. 10.1089/vim.2009.0090
34
DuL.ZhaoG.ChanC. C.SunS.ChenM.LiuZ.et al (2009b). Recombinant receptor-binding domain of SARS-CoV spike protein expressed in mammalian, insect and E. coli cells elicits potent neutralizing antibody and protective immunity.Virology393144–150. 10.1016/j.virol.2009.07.018
35
DuL.ZhaoG.HeY.GuoY.ZhengB. J.JiangS.et al (2007). Receptor-binding domain of SARS-CoV spike protein induces long-term protective immunity in an animal model.Vaccine252832–2838. 10.1016/j.vaccine.2006.10.031
36
DuL.ZhaoG.KouZ.MaC.SunS.PoonV. K.et al (2013c). Identification of a receptor-binding domain in the S protein of the novel human coronavirus Middle East Respiratory Syndrome coronavirus as an essential target for vaccine development.J. Virol.879939–9942. 10.1128/jvi.01048-13
37
DuL.ZhaoG.LiL.HeY.ZhouY.ZhengB. J.et al (2009c). Antigenicity and immunogenicity of SARS-CoV S protein receptor-binding domain stably expressed in CHO cells.Biochem. Biophys. Res. Commun.384486–490. 10.1016/j.bbre.2009.05.003
38
ElshabrawyH. A.CoughlinM. M.BakerS. C.PrabhakarB. S. (2012). Human monoclonal antibodies against highly conserved HR1 and HR2 domains of the SARS-CoV spike protein are more broadly neutralizing.PLoS One7:e50366. 10.1371/journal.pone.0050366
39
EsperF.WeibelC.FergusonD.LandryM. L.KahnJ. S. (2006). Coronavirus HKU1 infection in the United States.Emerg. Infect. Dis.12775–779. 10.3201/eid1205.051316
40
FalseyA. R.WalshE. E.HaydenF. G. (2002). Rhinovirus and coronavirus infection-associated hospitalizations among older adults.J. Infect. Dis.1851338–1341. 10.1086/339881
41
FehrA. R.PerlmanS. (2015). Coronaviruses: an overview of their replication and pathogenesis.Methods Mol. Biol.12821–23. 10.1007/978-1-4939-2438-7_1
42
FouchierR. A. M.HartwigN. G.BestebroerT. M.NiemeyerB.de JongJ. C.SimonJ. H.et al (2004). A previously undescribed coronavirus associated with respiratory disease in humans.Proc. Natl. Acad. Sci. U.S.A.1016212–6216. 10.1073/pnas.0400762101
43
GeX. Y.LiJ. L.YangX. L.ChmuraA. A.ZhuG.EpsteinJ. H.et al (2013). Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor.Nature503535–538. 10.1038/nature12711
44
GernaG.CampaniniG.RovidaF.PercivalleE.SarasiniA.MarchiA.et al (2006). Genetic variability of human coronavirus OC43-, 229E-, and NL63-like strains and their association with lower respiratory tract infections of hospitalized infants and immunocompromised patients.J. Med. Virol.78938–949. 10.1002/jmv.20645
45
GernaG.PercivalleE.SarasiniA.CampaniniG.PirallaA.RovidaF.et al (2007). Human respiratory coronavirus HKU1 versus other coronavirus infections in Italian hospitalised patients.J. Clin. Virol.38244–250. 10.1016/j.jcv.2006.12.008
46
GuoY.SunS.WangK.ZhangS.ZhuW.ChenZ. (2005). Elicitation of immunity in mice after immunization with the S2 subunit of the severe acute respiratory syndrome coronavirus.DNA Cell Biol.24510–515. 10.1089/dna.2005.24.510
47
HaagmansB. L.van den BrandJ. M.RajV. S.VolzA.WohlseinP.SmitsS. L.et al (2016). An orthopoxvirus-based vaccine reduces virus excretion after MERS-CoV infection in dromedary camels.Science35177–81. 10.1126/science.aad1283
48
HamreD.KindigD. A.MannJ. (1967). Growth and intracellular development of a new respiratory virus.J. Virol.1810–816. 10.1128/jvi.1.4.810-816.1967
49
HamreD.ProcknowJ. J. (1966). A new virus isolated from human respiratory tract.Proc. Soc. Exp. Biol. Med.121190–193. 10.3181/00379727-121-30734
50
HeY.LiJ.HeckS.LustigmanS.JiangS. (2006a). Antigenic and immunogenic characterization of recombinant baculovirus-expressed severe acute respiratory syndrome coronavirus spike protein: implication for vaccine design.J. Virol.805757–5767. 10.1128/jvi.00083-06
51
HeY.LiJ.JiangS. (2006b). A single amino acid substitution (R441A) in the receptor-binding domain of SARS coronavirus spike protein disrupts the antigenic structure and binding activity.Biochem. Biophys. Res. Commun.344106–113. 10.1016/j.bbrc.2006.03.139
52
HeY.LiJ.LiW.LustigmanS.FarzanM.JiangS. (2006c). Cross-neutralization of human and palm civet severe acute respiratory syndrome coronaviruses by antibodies targeting the receptor-binding domain of spike protein.J. Immunol.1766085–6092. 10.4049/jimmunol.176.10.6085
53
HeY.LuH.SiddiquiP.ZhouY.JiangS. (2005a). Receptor-binding domain of severe acute respiratory syndrome coronavirus spike protein contains multiple conformation-dependent epitopes that induce highly potent neutralizing antibodies.J. Immunol.1744908–4915. 10.4049/jimmunol.174.8.4908
54
HeY.ZhouY.LiuS.KouZ.LiW.FarzanM.et al (2004). Receptor-binding domain of SARS-CoV spike protein induces highly potent neutralizing antibodies: implication for developing subunit vaccine.Biochem. Biophys. Res. Commun.324773–781. 10.1016/j.bbrc.2004.09.106
55
HeY.ZhouY.SiddiquiP.NiuJ.JiangS. (2005b). Identification of immunodominant epitopes on the membrane protein of the severe acute respiratory syndrome-associated coronavirus.J. Clin. Microbiol.433718–3726. 10.1128/jcm.43.8.3718-3726.2005
56
HofmannH.PyrcK.van der HoekL.GeierM.BerkhoutB.PohlmannS. (2005). Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry.Proc. Natl. Acad. Sci. U.S.A.1027988–7993. 10.1073/pnas.0409465102
57
HofmannH.SimmonsG.RennekampA. J.ChaipanC.GrambergT.HeckE.et al (2006). Highly conserved regions within the spike proteins of human coronaviruses 229E and NL63 determine recognition of their respective cellular receptors.J. Virol.808639–8652. 10.1128/jvi.00560-06
58
HuB.ZengL. P.YangX. L.GeX. Y.ZhangW.LiB.et al (2017). Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus.PLoS Pathog.13:e1006698. 10.1371/journal.ppat.1006698
59
HuangX.DongW.MilewskaA.GoldaA.QiY.ZhuQ. K.et al (2015). Human coronavirus HKU1 spike protein uses O-Acetylated sialic acid as an attachment receptor determinant and employs hemagglutinin-esterase protein as a receptor-destroying enzyme.J. Virol.897202–7213. 10.1128/jvi.00854-15
60
HulswitR. J. G.LangY.BakkersM. J. G.LiW.LiZ.SchoutenA.et al (2019). Human coronaviruses OC43 and HKU1 bind to 9-O-acetylated sialic acids via a conserved receptor-binding site in spike protein domain A.Proc. Natl Acad. Sci. U.S.A.1162681–2690. 10.1073/pnas.1809667116
61
JaumeM.YipM. S.KamY. W.CheungC. Y.KienF.RobertsA.et al (2012). SARS CoV subunit vaccine: antibody-mediated neutralisation and enhancement.Hong Kong Med. J.18 (Suppl. 2), 31–36.
62
JiamingL.YanfengY.YaoD.YaweiH.LinlinB.BaoyingH.et al (2017). The recombinant N-terminal domain of spike proteins is a potential vaccine against Middle East respiratory syndrome coronavirus (MERS-CoV) infection.Vaccine3510–18. 10.1016/j.vaccine.2016.11.064
63
JiangS.BottazziM. E.DuL.LustigmanS.TsengC. T.CurtiE.et al (2012). Roadmap to developing a recombinant coronavirus S protein receptor-binding domain vaccine for severe acute respiratory syndrome.Expert Rev. Vaccines111405–1413. 10.1586/erv.12.126
64
JiangS.DuL.ShiZ. (2020). An emerging coronavirus causing pneumonia outbreak in Wuhan, China: calling for developing therapeutic and prophylactic strategies.Emerg Microbes Infect.9275–277. 10.1080/22221751.2020.1723441
65
JiangS.HeY.LiuS. (2005). SARS vaccine development.Emerg. Infect. Dis.111016–1020. 10.3201/1107.050219
66
JiangS.LuL.DuL.DebnathA. K. (2013). A predicted receptor-binding and critical neutralizing domain in S protein of the novel human coronavirus HCoV-EMC.J. Infect.66464–466. 10.1016/j.jinf.2012.12.003
67
KamY. W.KienF.RobertsA.CheungY. C.LamirandeE. W.VogelL.et al (2007). Antibodies against trimeric S glycoprotein protect hamsters against SARS-CoV challenge despite their capacity to mediate FcgammaRII-dependent entry into B cells in vitro.Vaccine25729–740. 10.1016/j.vaccine.2006.08.011
68
KapikianA. Z.JamesH. D.KellyS. J.DeesJ. H.TurnerH. C.McIntoshK.et al (1969). Isolation from man of avian infectious bronchitis virus-like viruses (coronavirus) similar to 229E virus with some epidemiological observations.J. Infect. Dis.119282–290. 10.1093/infdis/119.3.282
69
KillerbyM. E.BiggsH. M.HaynesA.DahlR. M.MustaquimD.GerberS. I.et al (2018). Human coronavirus circulation in the United States 2014-2017.J. Clin. Virol.10152–56. 10.1016/j.jcv.2018.01.019
70
KimK. Y.HanS. Y.KimH. S.CheongH. M.KimS. S.KimD. S. (2017). Human coronavirus in the 2014 winter season as a cause of lower respiratory tract infection.Yonsei Med. J.58174–179. 10.3349/ymj.2017.58.1.174
71
KingA. M. Q.LefkowitzE. J.MushegianA. R.AdamsM. J.DutilhB. E.GorbalenyaA. E.et al (2018). Changes to taxonomy and the international code of virus classification and nomenclature ratified by the international committee on taxonomy of viruses (2018).Arch. Virol.1632601–2631. 10.1007/s00705-018-3847-1
72
KsiazekT. G.ErdmanD.GoldsmithC. S.ZakiS. R.PeretT.EmeryS.et al (2003). A novel coronavirus associated with severe acute respiratory syndrome.N. Engl. J. Med.3481953–1966. 10.1056/NEJMoa030781
73
KudelovaM.BelvoncikovaP.VrbovaM.KovalovaA.StibraniovaI.KocakovaP.et al (2015). Detection of murine herpesvirus 68 (MHV-68) in dermacentor reticulatus ticks.Microb. Ecol.70785–794. 10.1007/s00248-015-0622-7
74
KusanagiK.KuwaharaH.KatohT.NunoyaT.IshikawaY.SamejimaT.et al (1992). Isolation and serial propagation of porcine epidemic diarrhhea virus in cell-cultures and partial characterization of the isolate.J. Vet. Med. Sci.54313–318. 10.1292/jvms.54.313
75
LanJ.DengY.ChenH.LuG.WangW.GuoX.et al (2014). Tailoring subunit vaccine immunity with adjuvant combinations and delivery routes using the Middle East respiratory coronavirus (MERS-CoV) receptor-binding domain as an antigen.PLoS One9:e112602. 10.1371/journal.pone.0112602
76
LanJ.YaoY.DengY.ChenH.LuG.WangW.et al (2015). Recombinant receptor binding domain protein induces partial protective immunity in rhesus macaques against Middle East respiratory syndrome coronavirus challenge.EBioMedicine21438–1446. 10.1016/j.ebiom.2015.08.031
77
LauS. K.WooP. C.YipC. C.TseH.TsoiH. W.ChengV. C.et al (2006). Coronavirus HKU1 and other coronavirus infections in Hong Kong.J. Clin. Microbiol.442063–2071. 10.1128/jcm.02614-05
78
LauS. K. P.ZhangL.LukH. K. H.XiongL.PengX.LiK. S. M.et al (2018). Receptor usage of a novel bat lineage C betacoronavirus reveals evolution of Middle East respiratory syndrome-related coronavirus spike proteins for human dipeptidyl peptidase 4 binding.J. Infect. Dis.218197–207. 10.1093/infdis/jiy018
79
LehmannC.WolfH.XuJ. G.ZhaoQ. B.ShaoY. M.MotzM.et al (2008). A line immunoassay utilizing recombinant nucleocapsid proteins for detection of antibodies to human coronaviruses.Diagn. Microbiol. Infect. Dis.6140–48. 10.1016/j.diagmicrobio.2007.12.002
80
LelliD.PapettiA.SabelliC.RostiE.MorenoA.BoniottiM. B. (2013). Detection of coronaviruses in bats of various species in Italy.Viruses52679–2689. 10.3390/v5112679
81
LiF. (2008). Structural analysis of major species barriers between humans and palm civets for severe acute respiratory syndrome coronavirus infections.J. Virol.826984–6991. 10.1128/jvi.00442-08
82
LiF. (2015). Receptor recognition mechanisms of coronaviruses: a decade of structural studies.J. Virol.891954–1964. 10.1128/jvi.02615-14
83
LiF. (2016). Structure, function, and evolution of coronavirus spike proteins.Annu. Rev. Virol.3237–261. 10.1146/annurev-virology-110615-042301
84
LiF.DuL. (2019). MERS coronavirus: an emerging zoonotic virus.Viruses11:E663. 10.3390/v11070663
85
LiF.LiW.FarzanM.HarrisonS. C. (2005a). Structure of SARS coronavirus spike receptor-binding domain complexed with receptor.Science3091864–1868. 10.1126/science.1116480
86
LiW.ShiZ.YuM.RenW.SmithC.EpsteinJ. H.et al (2005b). Bats are natural reservoirs of SARS-like coronaviruses.Science310676–679. 10.1126/science.1118391
87
LiW.ZhangC.SuiJ.KuhnJ. H.MooreM. J.LuoS.et al (2005c). Receptor and viral determinants of SARS-coronavirus adaptation to human ACE2.EMBO J.241634–1643. 10.1038/sj.emboj.7600640
88
LiJ.UlitzkyL.SilbersteinE.TaylorD. R.ViscidiR. (2013). Immunogenicity and protection efficacy of monomeric and trimeric recombinant SARS coronavirus spike protein subunit vaccine candidates.Viral. Immunol.26126–132. 10.1089/vim.2012.0076
89
LiW.GreenoughT. C.MooreM. J.VasilievaN.SomasundaranM.SullivanJ. L.et al (2004). Efficient replication of severe acute respiratory syndrome coronavirus in mouse cells is limited by murine angiotensin-converting enzyme 2.J. Virol.7811429–11433. 10.1128/jvi.78.20.11429-11433.2004
90
LiW.HulswitR. J. G.WidjajaI.RajV. S.McBrideR.PengW.et al (2017). Identification of sialic acid-binding function for the Middle East respiratory syndrome coronavirus spike glycoprotein.Proc. Natl Acad. Sci. U.S.A.114E8508–E8517. 10.1073/pnas.1712592114
91
LiW.MooreM. J.VasilievaN.SuiJ.WongS. K.BerneM. A.et al (2003). Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus.Nature426450–454. 10.1038/nature02145
92
LiuD. X.FungT. S.ChongK. K.ShuklaA.HilgenfeldR. (2014). Accessory proteins of SARS-CoV and other coronaviruses.Antiviral Res.10997–109. 10.1016/j.antiviral.2014.06.013
93
LiuR. Y.WuL. Z.HuangB. J.HuangJ. L.ZhangY. L.KeM. L.et al (2005). Adenoviral expression of a truncated S1 subunit of SARS-CoV spike protein results in specific humoral immune responses against SARS-CoV in rats.Virus Res.11224–31. 10.1016/j.virusres.2005.02.009
94
LiuS. J.LengC. H.LienS. P.ChiH. Y.HuangC. Y.LinC. L.et al (2006). Immunological characterizations of the nucleocapsid protein based SARS vaccine candidates.Vaccine243100–3108. 10.1016/j.vaccine.2006.01.058
95
LiuS. L.SaifL. (2020). Emerging viruses without borders: the Wuhan coronavirus.Viruses12:E130. 10.3390/v12020130
96
LuG.HuY.WangQ.QiJ.GaoF.LiY.et al (2013). Molecular basis of binding between novel human coronavirus MERS-CoV and its receptor CD26.Nature500227–231. 10.1038/nature12328
97
LuL.LiuQ.ZhuY.ChanK. H.QinL.LiY.et al (2014). Structure-based discovery of Middle East respiratory syndrome coronavirus fusion inhibitor.Nat. Commun.5:3067. 10.1038/ncomms4067
98
LuoC. M.WangN.YangX. L.LiuH. Z.ZhangW.LiB.et al (2018a). Discovery of novel bat coronaviruses in south China that use the same receptor as MERS coronavirus.J. Virol.92e116–e118. 10.1128/jvi.00116-18
99
LuoF.LiaoF. L.WangH.TangH. B.YangZ. Q.HouW. (2018b). Evaluation of antibody-dependent enhancement of SARS-CoV infection in rhesus macaques immunized with an inactivated SARS-CoV vaccine.Virol. Sin.33201–204. 10.1007/s12250-018-0009-2
100
MaC.LiY.WangL.ZhaoG.TaoX. R.TsengC. T.et al (2014a). Intranasal vaccination with recombinant receptor-binding domain of MERS-CoV spike protein induces much stronger local mucosal immune responses than subcutaneous immunization: implication for designing novel mucosal MERS vaccines.Vaccine322100–2108. 10.1016/j.vaccine.2014.02.004
101
MaC.WangL.TaoX.ZhangN.YangY.TsengC. T.et al (2014b). Searching for an ideal vaccine candidate among different MERS coronavirus receptor-binding fragments - The importance of immunofocusing in subunit vaccine design.Vaccine326170–6176. 10.1016/j.vaccine.2014.08.086
102
MaY.ZhangY.LiangX.LouF.OglesbeeM.KrakowkaS.et al (2015). Origin, evolution, and virulence of porcine deltacoronaviruses in the United States.mBio6:e00064. 10.1128/mBio.00064-15
103
MarraM. A.JonesS. J.AstellC. R.HoltR. A.Brooks-WilsonA.ButterfieldY. S.et al (2003). The genome sequence of the SARS-associated coronavirus.Science3001399–1404. 10.1126/science.1085953
104
MatobaY.AbikoC.IkedaT.AokiY.SuzukiY.YahagiK.et al (2015). Detection of the human coronavirus 229E, HKU1, NL63, and OC43 between 2010 and 2013 in Yamagata, Japan.Jpn. J. Infect. Dis.68138–141. 10.7883/yoken.JJID.2014.266
105
McIntoshK.BeckerW. B.ChanockR. M. (1967). Growth in suckling-mouse brain of IBV-like viruses from patients with upper respiratory tract disease.Proc. Natl. Acad. Sci. U.S.A.582268–2273. 10.1073/pnas.58.6.2268
106
MenacheryV. D.MitchellH. D.CockrellA. S.GralinskiL. E.YountBLJrGrahamR. L.et al (2017). MERS-CoV accessory ORFs play key role for infection and pathogenesis.mBio8:e00665-17. 10.1128/mBio.00665-17
107
MobarakiK.AhmadzadehJ. (2019). Current epidemiological status of Middle East respiratory syndrome coronavirus in the world from 1.1.2017 to 17.1.2018: a cross-sectional study.BMC Infect. Dis.19:351. 10.1186/s12879-019-3987-2
108
MouH.RajV. S.van KuppeveldF. J.RottierP. J.HaagmansB. L.BoschB. J. (2013). The receptor binding domain of the new Middle East Respiratory Syndrome coronavirus maps to a 231-residue region in the spike protein that efficiently elicits neutralizing antibodies.J. Virol.879379–9383. 10.1128/jvi.01277-13
109
MourezT.VabretA.HanY.DinaJ.LegrandL.CorbetS.et al (2007). Baculovirus expression of HCoV-OC43 nucleocapsid protein and development of a Western blot assay for detection of human antibodies against HCoV-OC43.J. Virol. Methods139175–180. 10.1016/j.jviromet.2006.09.024
110
MyintS. H. (1994). Human coronaviruses - A brief review.Rev. Med. Virol.435–46. 10.1002/rmv.1980040108
111
NichollsJ. M.PoonL. L.LeeK. C.NgW. F.LaiS. T.LeungC. Y.et al (2003). Lung pathology of fatal severe acute respiratory syndrome.Lancet3611773–1778. 10.1016/s0140-6736(03)13413-7
112
NyonM. P.DuL.TsengC. K.SeidC. A.PolletJ.NaceancenoK. S.et al (2018). Engineering a stable CHO cell line for the expression of a MERS-coronavirus vaccine antigen.Vaccine361853–1862. 10.1016/j.vaccine.2018.02.065
113
OuX.GuanH.QinB.MuZ.WojdylaJ. A.WangM.et al (2017). Crystal structure of the receptor binding domain of the spike glycoprotein of human betacoronavirus HKU1.Nat. Commun.8:2529. 10.1038/ncomms15216
114
PallesenJ.WangN.CorbettK. S.WrappD.KirchdoerferR. N.TurnerH. L.et al (2017). Immunogenicity and structures of a rationally designed prefusion MERS-CoV spike antigen.Proc. Natl. Acad. Sci. U.S.A.114E7348–E7357. 10.1073/pnas.1707304114
115
PedersenN. C. (2014). An update on feline infectious peritonitis: virology and immunopathogenesis.Vet. J.201123–132. 10.1016/j.tvjl.2014.04.017
116
PimentelT. A.YanZ.JeffersS. A.HolmesK. V.HodgesR. S.BurkhardP. (2009). Peptide nanoparticles as novel immunogens: design and analysis of a prototypic severe acute respiratory syndrome vaccine.Chem. Biol. Drug Des.7353–61. 10.1111/j.1747-0285.2008.00746.x
117
PoonL. L.ChuD. K.ChanK. H.WongO. K.EllisT. M.LeungY. H.et al (2005). Identification of a novel coronavirus in bats.J. Virol.792001–2009. 10.1128/jvi.79.4.2001-2009.2005
118
QiuM.ShiY.GuoZ.ChenZ.HeR.ChenR.et al (2005). Antibody responses to individual proteins of SARS coronavirus and their neutralization activities.Microbes Infect.7882–889. 10.1016/j.micinf.2005.02.006
119
RajV. S.MouH.SmitsS. L.DekkersD. H.MullerM. A.DijkmanR.et al (2013). Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC.Nature495251–254. 10.1038/nature12005
120
RegameyN.KaiserL.RoihaH. L.DeffernezC.KuehniC. E.LatzinP.et al (2008). Viral etiology of acute respiratory infections with cough in infancy - A community-based birth cohort study.Pediatr. Infect. Dis. J.27100–105. 10.1097/INF.0b013e31815922c8
121
ReuskenC. B.HaagmansB. L.MullerM. A.GutierrezC.GodekeG. J.MeyerB.et al (2013). Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: a comparative serological study.Lancet Infect. Dis.13859–866. 10.1016/s1473-3099(13)70164-6
122
RotaP. A.ObersteM. S.MonroeS. S.NixW. A.CampagnoliR.IcenogleJ. P.et al (2003). Characterization of a novel coronavirus associated with severe acute respiratory syndrome.Science3001394–1399. 10.1126/science.1085952
123
ScobeyT.YountB. L.SimsA. C.DonaldsonE. F.AgnihothramS. S.MenacheryV. D.et al (2013). Reverse genetics with a full-length infectious cDNA of the Middle East respiratory syndrome coronavirus.Proc. Natl. Acad. Sci. U.S.A.11016157–16162. 10.1073/pnas.1311542110
124
SeveranceE. G.BossisI.DickersonF. B.StallingsC. R.OrigoniA. E.SullensA.et al (2008). Development of a nucleocapsid-based human coronavirus immunoassay and estimates of individuals exposed to coronavirus in a US metropolitan population.Clin. Vaccine Immunol.151805–1810. 10.1128/cvi.00124-08
125
ShaoX.GuoX.EsperF.WeibelC.KahnJ. S. (2007). Seroepidemiology of group I human coronaviruses in children.J. Clin. Virol.40207–213. 10.1016/j.jcv.2007.08.007
126
SnijderE. J.BredenbeekP. J.DobbeJ. C.ThielV.ZiebuhrJ.PoonL. L.et al (2003). Unique and conserved features of genome and proteome of SARS-coronavirus, an early split-off from the coronavirus group 2 lineage.J. Mol. Biol.331991–1004. 10.1016/s0022-2836(03)00865-9
127
SnijderE. J.DecrolyE.ZiebuhrJ. (2016). The nonstructural proteins directing coronavirus RNA synthesis and processing.Adv. Virus Res.9659–126. 10.1016/bs.aivir.2016.08.008
128
TaiW.WangY.FettC. A.ZhaoG.LiF.PerlmanS.et al (2017). Recombinant receptor-binding domains of multiple middle east respiratory syndrome coronaviruses (MERS-CoVs) induce cross-neutralizing antibodies against divergent human and camel MERS-CoVs and antibody escape mutants.J. Virol91:e01651-16. 10.1128/jvi.01651-16
129
TaiW.ZhaoG.SunS.GuoY.WangY.TaoX.et al (2016). A recombinant receptor-binding domain of MERS-CoV in trimeric form protects human dipeptidyl peptidase 4 (hDPP4) transgenic mice from MERS-CoV infection.Virology499375–382. 10.1016/j.virol.2016.10.005
130
TangJ.ZhangN.TaoX.ZhaoG.GuoY.TsengC. T.et al (2015). Optimization of antigen dose for a receptor-binding domain-based subunit vaccine against MERS coronavirus.Hum. Vaccin. Immunother.111244–1250. 10.1080/21645515.2015.1021527
131
TomarS.JohnstonM. L.St JohnS. E.OsswaldH. L.NyalapatlaP. R.PaulL. N.et al (2015). Ligand-induced dimerization of Middle East respiratory syndrome (MERS) coronavirus nsp5 protease (3CLpro): implifications for nsp5 regulation and the development of antivirals.J. Biol. Chem.29019403–19422. 10.1074/jbc.M115.651463
132
TortoriciM. A.WallsA. C.LangY.WangC.LiZ.KoerhuisD.et al (2019). Structural basis for human coronavirus attachment to sialic acid receptors.Nat. Struct. Mol. Biol.26481–489. 10.1038/s41594-019-0233-y
133
TuC.CrameriG.KongX.ChenJ.SunY.YuM.et al (2004). Antibodies to SARS coronavirus in civets.Emerg. Infect. Dis.102244–2248. 10.3201/eid1012.040520
134
TyrrellD. A.BynoeM. L. (1965). Cultivation of a novel type of common-cold virus in organ cultures.Br. Med. J.11467–1470. 10.1136/bmj.1.5448.1467
135
VabretA.MourezT.GouarinS.PetitjeanJ.FreymuthF. (2003). An outbreak of coronavirus OC43 respiratory infection in Normandy, France.Clin. Infect. Dis.36985–989. 10.1086/374222
136
van BoheemenS.de GraafM.LauberC.BestebroerT. M.RajV. S.ZakiA. M.et al (2012). Genomic characterization of a newly discovered coronavirus associated with acute respiratory distress syndrome in humans.mBio3:e00473-12. 10.1128/mBio.00473-12
137
van der HoekL.PyrcK.JebbinkM. F.Vermeulen-OostW.BerkhoutR. J.WolthersK. C.et al (2004). Identification of a new human coronavirus.Nat. Med.10368–373. 10.1038/nm1024
138
VeitS.JanyS.FuxR.SutterG.VolzA. (2018). CD8+ T cells responding to the Middle East Respiratory Syndrome coronavirus nucleocapsid protein delivered by vaccinia virus MVA in mice.Viruses10:E718. 10.3390/v10120718
139
WangC.ZhengX.GaiW.ZhaoY.WangH.WangH.et al (2017). MERS-CoV virus-like particles produced in insect cells induce specific humoural and cellular imminity in rhesus macaques.Oncotarget812686–12694. 10.18632/oncotarget.8475
140
WangL.ShiW.JoyceM. G.ModjarradK.ZhangY.LeungK.et al (2015). Evaluation of candidate vaccine approaches for MERS-CoV.Nat. Commun.6:7712. 10.1038/ncomms8712
141
WangN.ShiX.JiangL.ZhangS.WangD.TongP.et al (2013). Structure of MERS-CoV spike receptor-binding domain complexed with human receptor DPP4.Cell Res.23986–993. 10.1038/cr.2013.92
142
WangY.TaiW.YangJ.ZhaoG.SunS.TsengC. K.et al (2017). Receptor-binding domain of MERS-CoV with optimal immunogen dosage and immunization interval protects human transgenic mice from MERS-CoV infection.Hum. Vaccin. Immunother.131615–1624. 10.1080/21645515.2017.1296994
143
WongS. K.LiW.MooreM. J.ChoeH.FarzanM. (2004). A 193-amino acid fragment of the SARS coronavirus S protein efficiently binds angiotensin-converting enzyme 2.J. Biol. Chem.2793197–3201. 10.1074/jbc.C300520200
144
WooP. C.LauS. K.ChuC. M.ChanK. H.TsoiH. W.HuangY.et al (2005). Characterization and complete genome sequence of a novel coronavirus, coronavirus HKU1, from patients with pneumonia.J. Virol.79884–895. 10.1128/jvi.79.2.884-895.2005
145
WooP. C.LauS. K.LamC. S.LaiK. K.HuangY.LeeP.et al (2009a). Comparative analysis of complete genome sequences of three avian coronaviruses reveals a novel group 3c coronavirus.J. Virol.83908–917. 10.1128/jvi.01977-08
146
WooP. C.LauS. K.LamC. S.LauC. C.TsangA. K.LauJ. H.et al (2012). Discovery of seven novel mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirus.J. Virol.863995–4008. 10.1128/jvi.06540-11
147
WooP. C.LauS. K.LamC. S.TsangA. K.HuiS. W.FanR. Y.et al (2014). Discovery of a novel bottlenose dolphin coronavirus reveals a distinct species of marine mammal coronavirus in gammacoronavirus.J. Virol.881318–1331. 10.1128/jvi.02351-13
148
WooP. C.LauS. K.YipC. C.HuangY.YuenK. Y. (2009b). More and more coronaviruses: human coronavirus HKU1.Viruses157–71. 10.3390/v1010057
149
YangY.DengY.WenB.WangH.MengX.LanJ.et al (2014a). The amino acids 736-761 of the MERS-CoV spike protein induce neutralizing antibodies: implications for the development of vaccines and antiviral agents.Viral Immunol.27543–550. 10.1089/vim.2014.0080
150
YangY.DuL.LiuC.WangL.MaC.TangJ.et al (2014b). Receptor usage and cell entry of bat coronavirus HKU4 provide insight into bat-to-human transmission of MERS coronavirus.Proc. Natl. Acad. Sci. U.S.A.11112516–12521. 10.1073/pnas.1405889111
151
YangZ. Y.KongW. P.HuangY.RobertsA.MurphyB. R.SubbaraoK.et al (2004). A DNA vaccine induces SARS coronavirus neutralization and protective immunity in mice.Nature428561–564. 10.1038/nature02463
152
YuanY.CaoD.ZhangY.MaJ.QiJ.WangQ.et al (2017). Cryo-EM structures of MERS-CoV and SARS-CoV spike glycoproteins reveal the dynamic receptor binding domains.Nat. Commun.8:15092. 10.1038/ncomms15092
153
ZakhartchoukA. N.SharonC.SatkunarajahM.AuperinT.ViswanathanS.MutwiriG.et al (2007). Immunogenicity of a receptor-binding domain of SARS coronavirus spike protein in mice: implications for a subunit vaccine.Vaccine25136–143. 10.1016/j.vaccine.2006.06.084
154
ZakiA. M.van BoheemenS.BestebroerT. M.OsterhausA. D.FouchierR. A. (2012). Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia.N. Engl. J. Med.3671814–1820. 10.1056/NEJMoa1211721
155
ZengF.ChowK. Y.HonC. C.LawK. M.YipC. W.ChanK. H.et al (2004). Characterization of humoral responses in mice immunized with plasmid DNAs encoding SARS-CoV spike gene fragments.Biochem. Biophys. Res. Commun.3151134–1139. 10.1016/j.bbrc.2004.01.166
156
ZhangN.ChannappanavarR.MaC.WangL.TangJ.GarronT.et al (2016). Identification of an ideal adjuvant for receptor-binding domain-based subunit vaccines against Middle East respiratory syndrome coronavirus.Cell. Mol. Immunol.13180–190. 10.1038/cmi.2015.03
157
ZhangN.JiangS.DuL. (2014). Current advancements and potential strategies in the development of MERS-CoV vaccines.Expert Rev. Vaccines13761–774. 10.1586/14760584.2014.912134
158
ZhangN.TangJ.LuL.JiangS.DuL. (2015). Receptor-binding domain-based subunit vaccines against MERS-CoV.Virus Res.202151–159. 10.1016/j.virusres.2014.11.013
159
ZhangN.WangL.DengX.LiangR.SuM.HeC.et al (2020). Recent advances in the detection of respiratory virus infection in humans.J. Med. Virol.[Epub ahead of print].
160
ZhengN.XiaR.YangC.YinB.LiY.DuanC.et al (2009). Boosted expression of the SARS-CoV nucleocapsid protein in tobacco and its immunogenicity in mice.Vaccine275001–5007. 10.1016/j.vaccine.2009.05.073
161
ZhongN. S.ZhengB. J.LiY. M.PoonL. L.XieZ. H.ChanK. H.et al (2003). Epidemiology and cause of severe acute respiratory syndrome (SARS) in Guangdong, People’s Republic of China, in February, 2003.Lancet3621353–1358. 10.1016/s0140-6736(03)14630-2
162
ZhouP.YangX. L.WangX. G.HuB.ZhangL.ZhangW.et al (2020). A pneumonia outbreak associated with a new coronavirus of probable bat origin.Nature[Epub ahead of print].
163
ZhouY.JiangS.DuL. (2018). Prospects for a MERS-CoV spike vaccine.Expert Rev. Vaccines17677–686. 10.1080/14760584.2018.1506702
164
ZhouY.YangY.HuangJ.JiangS.DuL. (2019). Advances in MERS-CoV vaccines and therapeutics based on the receptor-binding domain.Viruses11:E60. 10.3390/v11010060
165
ZhuN.ZhangD.WangW.LiX.YangB.SongJ.et al (2020). A novel coronavirus from patients with pneumonia in China, 2019.N. Engl. J. Med.[Epub ahead of print].
166
ZhuX.LiuQ.DuL.LuL.JiangS. (2013). Receptor-binding domain as a target for developing SARS vaccines.J. Thoracic Dis.5S142–S148. 10.3978/j.issn.2072-1439.2013.06.06
Summary
Keywords
human coronaviruses, pathogenesis, SARS-CoV, MERS-CoV, 2019-nCoV, subunit vaccines
Citation
Wang N, Shang J, Jiang S and Du L (2020) Subunit Vaccines Against Emerging Pathogenic Human Coronaviruses. Front. Microbiol. 11:298. doi: 10.3389/fmicb.2020.00298
Received
28 November 2019
Accepted
10 February 2020
Published
28 February 2020
Volume
11 - 2020
Edited by
Lijun Rong, The University of Illinois at Chicago, United States
Reviewed by
Ahmed Mohamed Kandeil, National Research Centre, Egypt; Liang Qiao, Loyola University Chicago, United States
Updates
Copyright
© 2020 Wang, Shang, Jiang and Du.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shibo Jiang, shibojiang@fudan.edu.cnLanying Du, ldu@nybc.org
This article was submitted to Virology, a section of the journal Frontiers in Microbiology
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.