 Malene Risum1
Malene Risum1 Rasmus Krøger Hare1
Rasmus Krøger Hare1 Jan Berg Gertsen2Lise Kristensen2Helle Krogh Johansen3,4Jannik Helweg-Larsen5Nissrine Abou-Chakra1Tacjana Pressler6Marianne Skov6Søren Jensen-Fangel7
Jan Berg Gertsen2Lise Kristensen2Helle Krogh Johansen3,4Jannik Helweg-Larsen5Nissrine Abou-Chakra1Tacjana Pressler6Marianne Skov6Søren Jensen-Fangel7 Maiken Cavling Arendrup1,3,4*
Maiken Cavling Arendrup1,3,4*- 1Unit of Mycology, Statens Serum Institut, Copenhagen, Denmark
- 2Department of Clinical Microbiology, Aarhus University Hospital, Aarhus, Denmark
- 3Department of Clinical Microbiology, Rigshospitalet, Copenhagen, Denmark
- 4Department of Clinical Medicine, University of Copenhagen, Copenhagen, Denmark
- 5Department of Infectious Diseases, Rigshospitalet, Copenhagen, Denmark
- 6Cystic Fibrosis Center Copenhagen, Department of Pediatrics and Infectious Diseases, Rigshospitalet, Copenhagen, Denmark
- 7Department of Infectious Diseases, Aarhus University Hospital, Aarhus, Denmark
Azole-resistant (azole-R) Aspergillus is an increasing challenge worldwide. Patients with cystic fibrosis (CF) are at risk of Aspergillus colonization and disease due to a favorable lung environment for microorganisms. We performed a nationwide study in 2018 of azole-non-susceptible Aspergillus in CF patients and compared with data from two prior studies. All airway samples with mold isolates from patients monitored at the two CF centers in Denmark (RH, Jan–Sept and AUH, Jan–Jun) were included. Classical species identification (morphology and thermo-tolerance) was performed and MALDI-TOF/β-tubulin sequencing was performed if needed. Susceptibility was determined using EUCAST E.Def 10.1, and E.Def 9.3.2. cyp51A sequencing and STRAf genotyping were performed for azole-non-susceptible isolates and relevant sequential isolates. In total, 340 mold isolates from 159 CF patients were obtained. The most frequent species were Aspergillus fumigatus (266/340, 78.2%) and Aspergillus terreus (26/340, 7.6%). Azole-R A. fumigatus was cultured from 7.3% (10/137) of patients, including 9.5% (9/95) of patients at RH and 2.4% at AUH (1/42), respectively. In a 10-year perspective, azole-non-susceptibility increased numerically among patients at RH (10.5% in 2018 vs 4.5% in 2007–2009). Cyp51A resistance mechanisms were found in nine azole-R A. fumigatus from eight CF patients. Five were of environmental origin (TR34/L98H), three were human medicine-driven (two M220K and one M220R), and one was novel (TR343/L98H) and found in a patient who also harbored a TR34/L98H isolate. STRAf genotyping identified 27 unique genotypes among 45 isolates and ≥2 genotypes in 8 of 12 patients. This included one patient carrying two unique TR34/L98H isolates, a rare phenomenon. Genotyping of sequential TR343/L98H and TR34/L98H isolates from the same patient showed only minor differences in 1/9 markers. Finally, azole-R A. terreus was found in three patients including two with Cyp51A alterations (M217I and G51A, respectively). Azole-R A. fumigatus is increasing among CF patients in Denmark with the environmentally associated resistance TR34/L98H mechanism being dominant. Mixed infections (wildtype/non-wildtype and several non-wildtypes) and a case of potential additional tandem repeat acquisition in vivo were found. However, similar genotypes were identified from another patient (and outside this study), potentially suggesting a predominant TR34/L98H clone in DK. These findings suggest an increasing prevalence and complexity of azole resistance in A. fumigatus.
Introduction
Azole resistance (Azole-R) in Aspergillus fumigatus is an increasing problem and complicates patient management (Lestrade et al., 2019). Azole-R A. fumigatus in patients with cystic fibrosis (CF) has been reported in several studies (Mortensen et al., 2011a; Burgel et al., 2012; Morio et al., 2012; Bader et al., 2013; Fischer et al., 2014; Stevens et al., 2016; Prigitano et al., 2017; Abdolrasouli et al., 2018; Guegan et al., 2018; Güngör et al., 2018; Seufert et al., 2018; Engel et al., 2019; Lavergne et al., 2019; Table 1). Danish CF patients are followed up monthly at the two specialized CF clinics at Copenhagen University Hospital, Rigshospitalet (RH) and at Aarhus University Hospital (AUH). From the majority of these patients, airway samples are obtained monthly. We have previously studied azole-R A. fumigatus in the Copenhagen CF cohort (Mortensen et al., 2011a, b). The first study included isolates from Jan to March (Q1) 2007 (Mortensen et al., 2011b) and the second study isolates from July to Dec (Q3-4) of 2007 and of 2009 (Mortensen et al., 2011a). These studies documented an overall azole-non-susceptibility rate of 1.6 and of 4.5%, respectively. A lower but increasing rate from 1.8 to 3.8% of azole-R A. fumigatus among clinical samples (from CF as well as non-CF patients in Denmark) was found in a subsequent reference laboratory-based study, from the years 2010 to 2014 (Jensen et al., 2016). But the epidemiology of azole-R A. fumigatus including that specifically associated with environmental origin has not been systematically studied over a longer time in our country. Due to the increasing international and political concern related to the link between environmental azole fungicide use and azole resistance in A. fumigatus, we systematically investigated the azole resistance rate in the Danish CF population in 2018 and compared it to our previous data (Mortensen et al., 2011a).
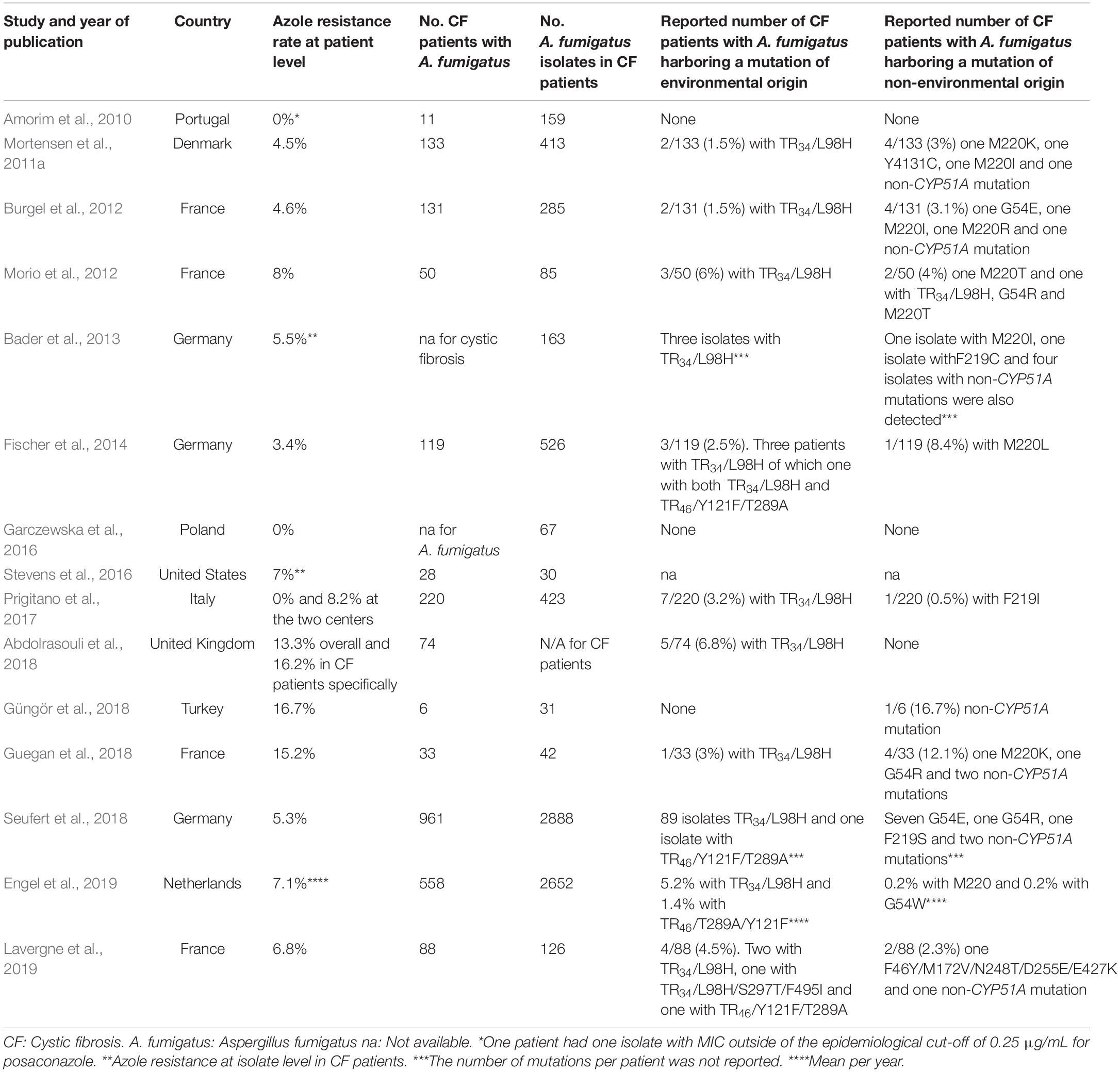
Table 1. Review of published studies on azole resistance in patients with cystic fibrosis.
CF is the most common autosomal recessive disease in Caucasians (Felton and Simmonds, 2014). Mutations in the CFTR (cystic fibrosis transmembrane regulator gene) affects the chloride transportation causing dysregulated fluid transport in the epithelial cells of multiple organs (Felton and Simmonds, 2014). Clinically, CF disease is dominated by infectious pulmonary complications (Felton and Simmonds, 2014). The respiratory tract is often colonized with molds especially A. fumigatus, which is found in 16 to 56.7% of airway samples (Pihet et al., 2009). Aspergillus may cause a diversity of manifestations ranging from asymptomatic colonization to serological sensitization, allergic bronchopulmonary aspergillosis (ABPA), Aspergillus bronchitis, and aspergilloma in CF patients (Felton and Simmonds, 2014). The most common is ABPA (Pihet et al., 2009), which occurs in approximately 10% of CF patients (Burgel et al., 2016; Carsin et al., 2017) and is the cause of hypersensitivity response to Aspergillus antigens (Williams et al., 2016). Azoles are the cornerstone in the management of CF patients with Aspergillus disease requiring antifungal therapy. Itraconazole is the first choice as an antifungal drug in the treatment of ABPA to reduce the burden of A. fumigatus and minimize use of corticosteroids (Patterson et al., 2016). Posaconazole is used as salvage therapy in ABPA or bronchitis (Skov et al., 2017; Periselneris et al., 2019), whereas voriconazole or isavuconazole are first-line agents (Maertens et al., 2016; Patterson et al., 2016) in the rare event of invasive aspergillosis (Burgel et al., 2016; Skov et al., 2017; Hamprecht et al., 2018).
Azoles target and inhibit the lanosterol 14-α-demethylase enzyme (Cyp51A) encoded by the cyp51A gene and thereby inhibit the ergosterol synthesis (Stensvold et al., 2012). Patients with recurrent or long-term need for azole therapy are at risk for azole-R Aspergillus due to selection of resistance during exposure to medical azoles (Hamprecht et al., 2018). Azole-R in A. fumigatus also occurs in patients with no prior azole therapy, caused by the inhalation of resistant mutant spores from the environment presumably selected due to azole fungicide use for plant and material protection (Astvad et al., 2014; Hagiwara et al., 2016). Well-known mechanisms behind azole resistance are target gene mutations in cyp51A. Two common resistance mechanisms, TR34/L98H and TR46/Y121F/T289A, are considered to be of environmental origin (Stensvold et al., 2012). These “environmental” mechanisms have previously been found in the Danish environment (Mortensen et al., 2010; Risum et al., 2019) and in clinical samples (Mortensen et al., 2011a; Astvad et al., 2014). Furthermore, target gene upregulation, efflux, and HapE (Camps et al., 2012) and Hmg1 (Rybak et al., 2019) alterations have been documented as underlying mechanisms of azole resistance in selected isolates.
In this study, we investigated the azole-R rate in a 10-y perspective and dissected underlying molecular resistance mechanisms and genotypes in Aspergillus in CF patients followed up at the two Danish CF centers that serve the entire country.
Materials and Methods
The two CF centers RH and AUH follow all the Danish CF patients. A total of 522 (320 and 202 adult and children) with CF were followed up at RH and AUH, respectively, in 2018. All positive cultures with mold from airway samples from the Danish CF population were included during a 6-month (Jan–June 2018) and a 9-month (Jan–Sept 2018) period, respectively, from AUH and RH. Primary culture was performed using Sabouraud glucose agar [SSI Diagnostika, Hillerød, Denmark (RH) and bioMérieux, Marcy l’Etoile, France (AUH), respectively]. Agar plates were incubated at 35–37°C and examined for 5 days (RH) and 2 days (AUH). Exclusion criteria were identical to our previous study (Mortensen et al., 2011a). In detail, repeat isolates from the same patient were excluded when found ≤30 days apart and confirmed as same species and with same susceptibility classification.
Identification was done to the Aspergillus species complex level using classical techniques, including thermo-tolerance test for A. fumigatus specifically, followed up by MALDI-TOF applying the Mass Spectrometry Identification database (Normand et al., 2017; Imbert et al., 2019) and β-tubulin sequencing (Glass and Donaldson, 1995) when necessary.
The EUCAST E.Def 10.1 method (Arendrup et al., 2017) was used for azole-R screening for A. fumigatus, and EUCAST E.Def 9.3.1 susceptibility testing (Arendrup et al., 2016) was performed for amphotericin B for the majority of the isolates and for itraconazole, posaconazole, and voriconazole for screening-positive A. fumigatus isolates and Aspergillus species other than A. fumigatus. Isolates with azole MIC(s) above the ECOFF(s) underwent cyp51A sequencing as previously described (Mortensen et al., 2011a). EUCAST clinical breakpoints v 9.0 were adopted for susceptibility classification into susceptible, non-susceptible (intermediate and resistant), and azole-R (Arendrup et al., 2013). For species and agents without breakpoints, EUCAST ECOFFs were adopted and non-wildtype isolates were regarded resistant. Sequential isolates from all patients harboring resistant A. fumigatus underwent STRAf genotyping (De Valk et al., 2005).
Results were compared to our previous Danish studies on azole-R in CF patients followed up at RH allowing a 10-year perspective (Mortensen et al., 2011a). Comparison of groups was performed with a contingency chi-square test using GraphPad Prism version 8.0.2.
(Preliminary results from RH have been presented at the European Congress on Clinical Microbiology and Infectious Diseases in 2019).
Results
In total, 340 unique mold isolates from 159 CF patients were obtained, of which 240 isolates were derived from 110 CF patients at RH (2.2 isolates per patient) and 100 isolates from 49 CF patients at AUH (2.0 isolates per patient). The median age was 30 years (6–68 years) at RH and 22 years (6–50 years) at AUH among patients with a mold isolate.
Overall, A. fumigatus was the most frequently isolated species (266/340 isolates, 78.2%), detected in 137/159 (86.2%) of the CF patients followed up by A. terreus species complex (26/340 isolates, 7.6%) in 10/159 patients (6.3%) and A. niger complex isolates (13/340 isolates, 3.8%) in 13/159 (8.2%) patients (Figure 1).
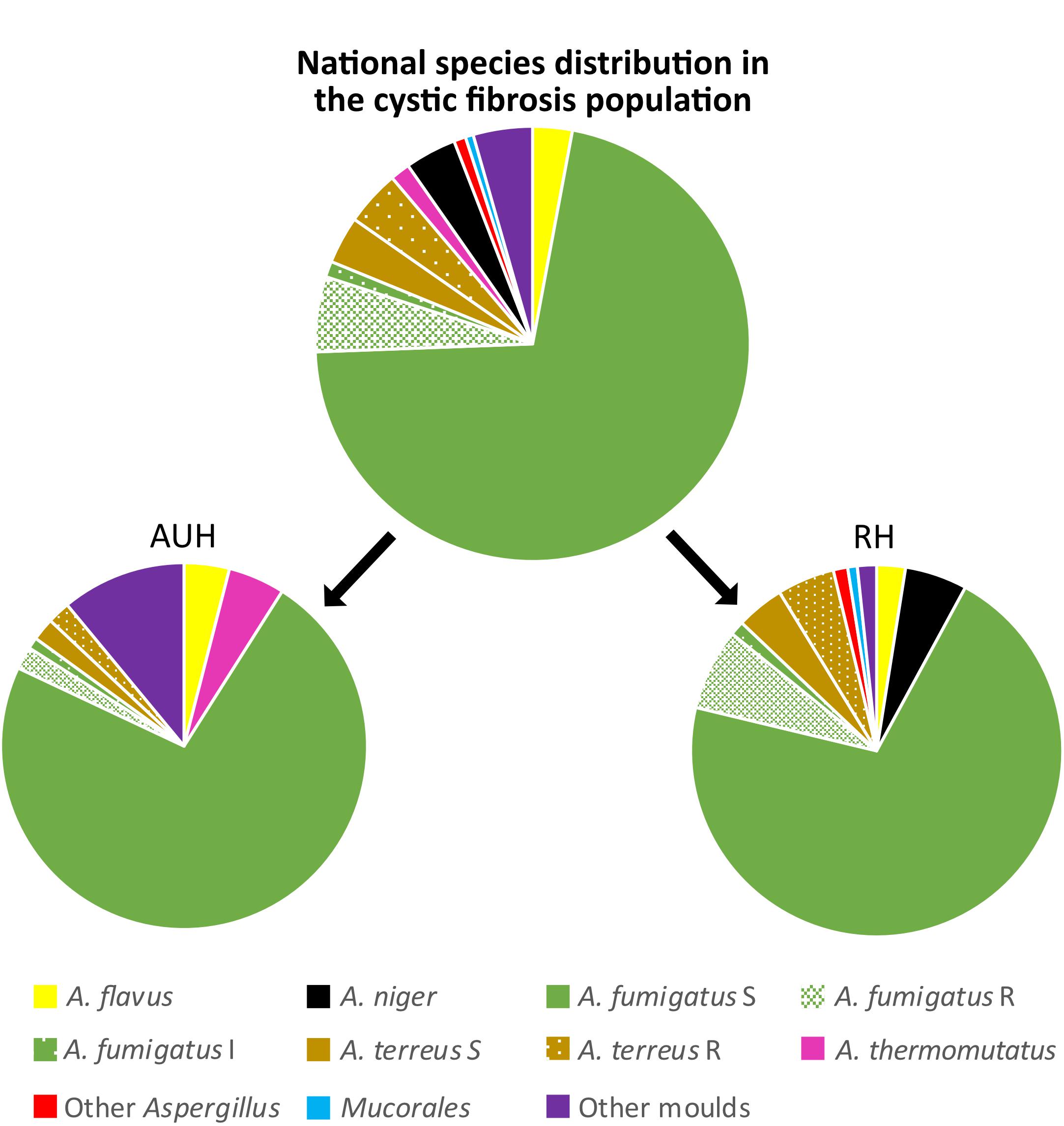
Figure 1. Species distribution at the cystic fibrosis population gathered and at the two centers separately. S, I, and R indicate the susceptibility for azoles. S: susceptible, I: intermediate, and R: resistant. Aarhus (AUH): Other molds: Scedosporium spp. (n = 8), Exophiala spp. (n = 2), Penicillium spp. (n = 1). Rigshospitalet (RH): Other Aspergillus: Aspergillus nidulans (n = 2) and Aspergillus sydowii (n = 1), Mucorales: Rhiomucor pusillus (n = 1) and Zygomycetes spp. (n = 1). Other molds: Scedosporium spp. (n = 3), and Rasamsonia spp. (n = 1).
Azole non-susceptible A. fumigatus was found in 12/137 (8.8%) and Azole-R A. fumigatus in 10/137 (7.3%) of all CF patients (Table 2). Eight patients harbored A. fumigatus with Cyp51A alterations, including one patient (RH-5) with two isolates with different Cyp51A alterations. Overall, Cyp51A alterations were thus found in 81.8% (9/11) azole-R A. fumigatus of which 45.5% (5/11) were of environmental origin (Table 2). Three A. fumigatus isolates were categorized as intermediate to one or several azoles, none of which harbored cyp51A mutations.
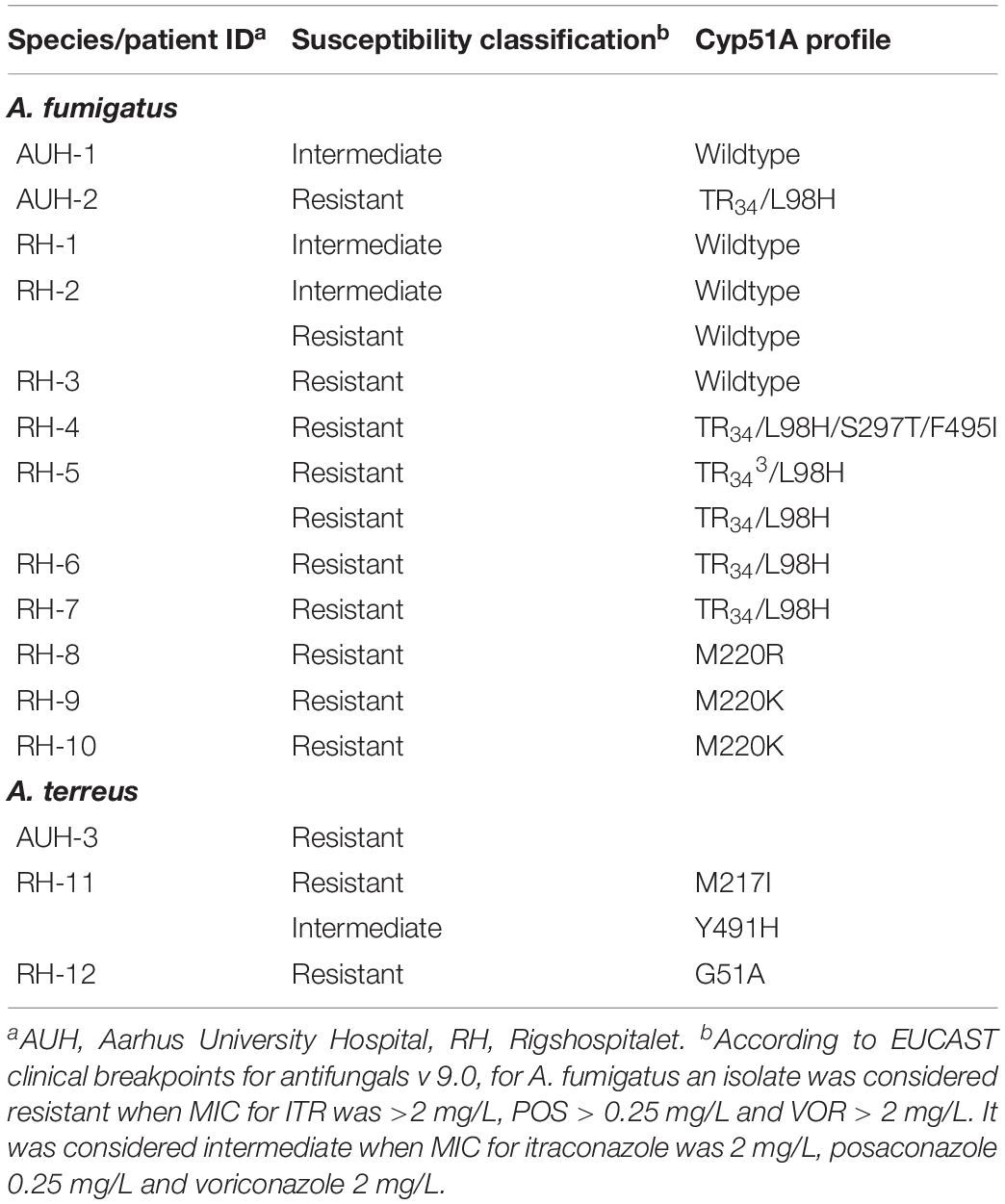
Table 2. Cyp51A profile for 18 Aspergillus fumigatus and Aspergillus terreus isolates with decreased susceptibility from 15 CF patients at the two referral centers in Denmark.
A. fumigatus was equally common among mold-colonized patients at the two centers [42/49 (85.7%) and 95/110 (86.4%)], but the proportion of patients with non-susceptible A. fumigatus isolates differed. At AUH, non-susceptible A. fumigatus isolates were observed in 2/42 (4.8%) patients (Table 2). At RH, 10/95 (10.5%) patients carried non-susceptible A. fumigatus. Nine of these patients (9.5%) carried 10 azole-R A. fumigatus. Eight patients had a cyp51A mutation (7.4% of patients), five of which with tandem repeats (TR34/L98H or TR343/L98H) and three with alterations affecting the M220 codon (Table 2). Among the 10 patients with resistant A. fumigatus, four patients had only resistant isolates obtained in the study period. These included the resistance mechanisms TR34/L98H, TR343/L98H, and wild-type. Alternating resistant and susceptible A. fumigatus isolates were found in six patients with the resistance mechanisms M220K, TR34/L98H, TR34/L98H/S297T/F495I, M220R, and wild-type (M220K two patients and one each).
STRAf genotyping identified 27 unique STRAf genotypes among the 45 A. fumigatus isolates from the 12 patients with azole non-susceptible A. fumigatus (Table 3). Eight patients harbored isolates with more than one genotype. Three of these patients, carried isolates that differed only in a single marker (RH-5, RH-7, and RH-8), and six patients carried isolates that were clearly unrelated (including two patients with both related and unrelated genotypes RH-7). Thus, patient RH-5 had six isolates with 8/9 identical STRAf markers, while marker 3A ranged from 95 repeats (TR34/L98H) to 96–101 repeats (five TR343/L98H) (Table 3). In contrast, Patient AUH-2 harbored two TR34/L98H isolates but with clearly different STRAf profiles. Patient RH-2 harbored 10 A. fumigatus with three different genotypes, while patient RH-8 had four isolates with 8/9 identical markers and a fifth isolate with 7/9 identical markers. Isolates that shared 8–9 markers were also found across patients. Thus, patient RH-7 had two TR34/L98H isolates with identical STRAf profiles as two TR343/L98H isolates from RH-5.
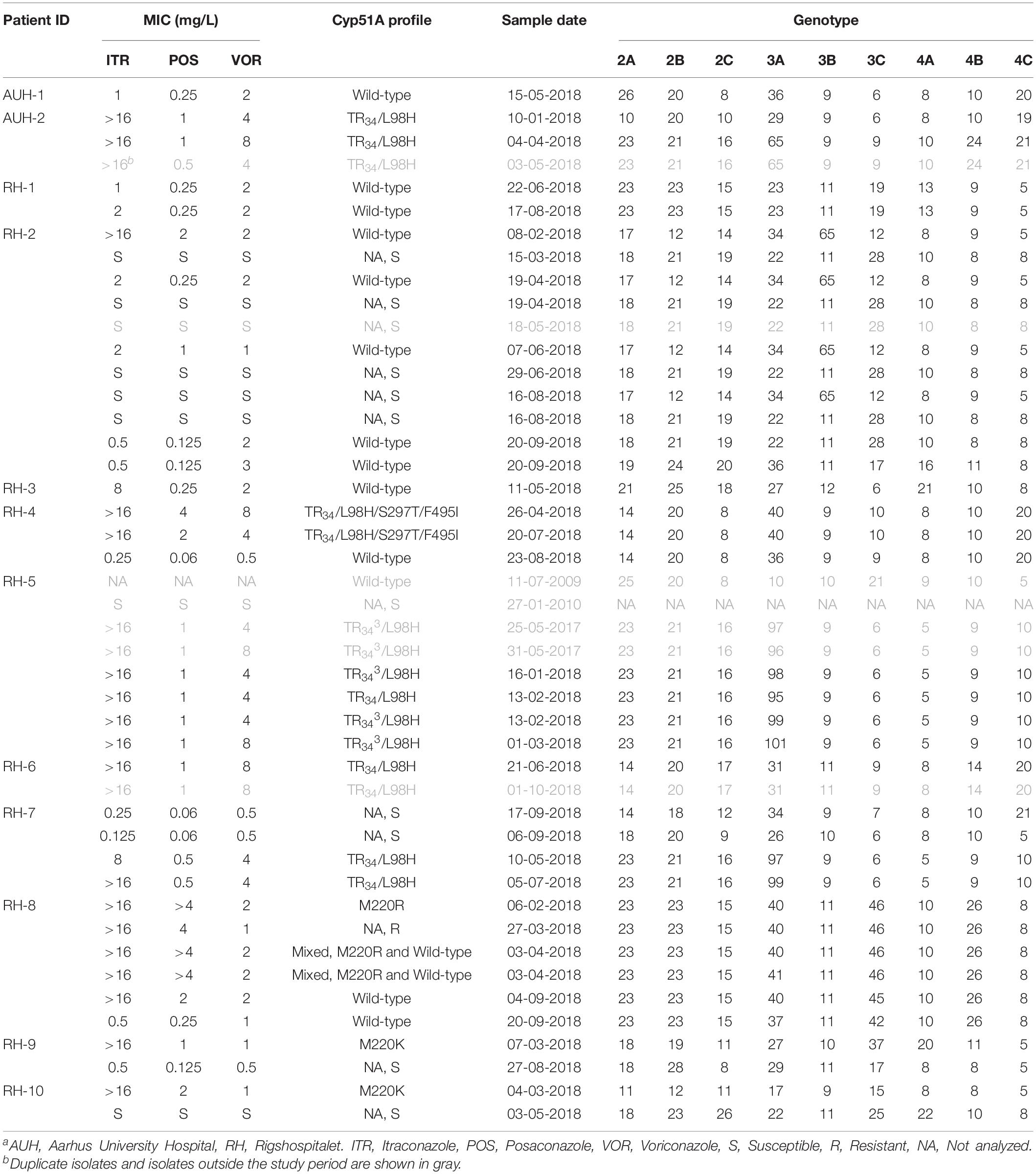
Table 3. Detailed susceptibility pattern and STRAf genotyping of all Aspergillus fumigatus isolates from patients harboring isolates with decreased azole susceptibility.
Azole resistance was also detected in other Aspergillus species. At AUH, such isolates were found in 3/49 (6.1%) patients including two patients with A. thermomutatus, and one patient with a voriconazole-resistant A. terreus isolate with a wild-type cyp51A (Table 2). At RH, two out of eight CF patients with A. terreus (1.8% of CF patients at RH) had non-susceptible isolates. One patient (RH-11) had both resistant and intermediate isolates recovered, which had Cyp51A amino acid substitutions M217I and Y491H, respectively. The other patient had a resistant A. terreus with a G51A mutation (RH-12, Table 2). MIC distributions for all Aspergillus isolates with reduced azole susceptibility at the two centers are shown in Supplementary Table 1.
Finally, we investigated potential changes in non-susceptibility and Cyp51A alteration rates in a 10-year perspective by comparison with data from the period 2007-2009 for the RH CF cohort (Mortensen et al., 2011a, b). Non-susceptible A. fumigatus was observed in 1/61 (1.6%) patients in Q1-2007, 6/133 patients in 2007–9 (4.5%) compared to 10/95 (10.5%) in 2018 (P = 0.047). Similarly, the proportion of isolates with Cyp51 alterations in Q1-2007, Q3-4-2007/9, and 2018 increased: 1.6% (95% CI: 0.1–8.7%), 3.8% (95% CI: 1.6–8.5%) and 7.4% (95% CI: 3.6–14.4%), respectively. Finally, the number of patients with resistant isolates with a tandem repeat, specifically, increased during the three study periods: 0% (95% CI: 0–5.9%), 1.5% (95% CI: 0.3–5.3%), and 4.2% (1.6–10.3%), as well as the proportion of isolates with target gene mutations associated with long-term azole treatment: 1.6% (95% CI: 0.8%-8.7%), 2.3% (95% CI: 0.6%-6.4%), and 3.2% (95% CI: 0.9%-8.9%) in Q1-2007, Q3-4-2007/9, and 2018, respectively.
Discussion
We report detailed and nationwide data on azole non-susceptibility and mold species distribution in respiratory isolates from Danish CF patients. Azole-R A. fumigatus with environmental origin was dominating and found at both centers suggesting a wide geographic distribution of TR34/L98H in Denmark. However, although the proportions of patients with A. fumigatus (85.7 and 86.3%, respectively) were similar at the two centers, an almost four-fold higher rate of azole-resistant A. fumigatus was observed at RH compared to AUH. Moreover, the resistance pattern was more diverse at RH and included both the resistance deriving from the environment and resistance mutations associated with azole treatment selection. Unfortunately, data on azole use in these patients could not be retrieved. However, it is most likely that the differences in azole-R between the two centers may be related to different prescription practices with a more extensive and longer duration of azole treatment at RH, in part due to an overall higher age and number of patients with chronic aspergillus bronchitis at RH. This is supported by the observation that human-driven target gene mutations were more common in patients at RH than at AUH.
At RH, cyp51A mutations of environmental origin accounted for half of the detected resistance, and the TR34/L98H rate has doubled over the past decade since the first detection in Q3-4 of 2007–9 (Mortensen et al., 2011a). Additionally, a subsequent laboratory study of A. fumigatus isolates received at the national reference center reported an increase during 2010–2014 (Jensen et al., 2016). Taken together, these studies suggest that TR34/L98H has gradually become more prevalent in Denmark since 2007 despite the fact that two of three environmental sampling studies in Denmark failed to detect TR34/L98H and TR46/Y121F/T289A in soil and air samples (Astvad et al., 2014; Jensen et al., 2016). This suggests either significant fluctuations in the number of resistant spores in the environment or that even low levels of resistant A. fumigatus can contribute to resistant infections in a predisposed lung environment.
Our observations of alternating or mixed resistant and susceptible isolates recovered from the same patient is a well-known phenomenon and highlight that a single sample may not be a representation for the entire lung flora (van Leer-Buter et al., 2007; Astvad et al., 2014). Not only may different phenotypes dominate in different lung sections but mixed A. fumigatus strains are also very challenging to identify and separate unless molecular analyses are performed. Of note, two TR34/L98H isolates with different and unique STRAf genotypes among our collection were recovered from patient AUH-2, a case which we have not previously seen in DK.
Of particular interest, patient RH-5 harbored five pan-azole-resistant isolates with a novel TR343/L98H resistance mechanism, which to our knowledge has not previously been found in clinical specimens. However, exposure in vitro of A. fumigatus conidia already containing a 34-bp insertion in the cyp51A-gene promoter to 8 mg/L of tebuconazole resulted in one clone with a 34-bp triplicate repeat (Snelders et al., 2012). In addition, the TR46/Y121F/T289A has also been found with additional 46-bp repeats in the promoter region in compost as well during sexual mating in in vitro studies (Zhang et al., 2017). The question remains whether this TR343/L98H resistance variant is novel in the environment and thus acquired de novo, as suggested by being isolated first, or whether the TR34/L98H was in fact first (but undiscovered in the first three specimens) and the additional TR34 repeat acquired in vivo. The STRAf profiles suggest that the five TR343/L98H isolates are isogenic with a classical example of microevolution. It is noteworthy that the TR34/L98H isolate from this patient shared 8/9 markers and had 95 repeats at marker 3A while an increasing number of repeats (96–101) were seen in the TR343/L98H isolates over time. Increasing repeat numbers have previously been found over time in vitro and in vivo (Mortensen et al., 2011a; De Groot and Meis, 2019). Furthermore, the TR34/L98H strain was discovered in a sample mixed with the TR343/L98H strain and had a white and slow-growing phenotype, which could help explain why it could potentially have been overlooked in earlier samples. Indeed, the in vivo acquisition of a tandem repeat in the promotor region has been reported from our group, where a 120-basepair tandem repeat evolved in a patient during azole therapy, supported by whole-genome sequencing (WGS) (Hare et al., 2019). It has also been suggested that the TR34 helps compensate for loss of fitness associated with the L98H change (Verweij et al., 2016) and thus the additional TR34 could potentially further improve fitness and outgrow the TR34/L98H, which in this patient appeared with a weaker phenotype (Verweij et al., 2016). A third hypothesis could be that this is a random coincidence of similar STRAf genotypes. Indeed, the finding of two TR34/L98H isolates from another patient (RH-7) displaying identical STRAf profiles as two TR343/L98H isolates was surprising and further complicates the interpretation. One concern would be lab contamination, but since the two RH-7 isolates were received months apart and with different cyp51A profiles, this seems unlikely. A final, and worrying, theory is that we may have encountered a dominating TR34/L98H clone in DK similar to the study from India (Chowdhary et al., 2012). Indeed, outside the study period we have encountered a total of 20 isolates from 12 different patients from all around DK and also in two air samples sharing the same 8/9 STRAf markers, exclusively differing in marker 3A, ranging from 35 to more than 130 repeats. The high variation in 3A (in our two patients) indicates a highly mutagenic strain type, which may help explain the rare development of the TR343/L98H variant. Further studies including WGS are desirable to further explore the origin of this novel resistance mechanism as well as the potentially novel dominating genotype.
Two CF patients had resistant A. fumigatus with wild-type cyp51A, as reported in other CF studies (Mortensen et al., 2011a; Burgel et al., 2012; Guegan et al., 2018; Seufert et al., 2018; Lavergne et al., 2019) at similar rates (Mortensen et al., 2011a; Burgel et al., 2012; Lavergne et al., 2019) as well as in patients with chronic pulmonary aspergillosis (Howard et al., 2013). Phenotypic susceptibility testing therefore remains crucial because molecular detection of resistance mechanism enables the detection of resistance, but not susceptibility. Moreover, alternating findings of susceptible and non-susceptible isolates in the same patient demonstrate the need of repeated sampling and susceptibility testing of several colonies when present in patients requiring azole therapy, as recommended in current guidelines (Ullmann et al., 2018; Guinea et al., 2019).
Azole-R A. terreus constitutes a significant challenge since A. terreus has intrinsic reduced susceptibility to amphotericin B, rendering it multidrug-resistant (Arendrup et al., 2012; Zoran et al., 2018; Rivero-Menendez et al., 2019). The present finding of A. terreus of 6.3% (10/159) nationally at patient level is quite high compared to previous studies (Mortensen et al., 2011a; Fischer et al., 2014; Engel et al., 2019), reporting 1.9% (Mortensen et al., 2011a) and 2.4% A. terreus at the isolate level (Fischer et al., 2014) and 3.9% of all Aspergillus spp. (Engel et al., 2019). Whereas M217I has been reported previously (Rivero-Menendez et al., 2019), G51A is, to our knowledge, novel. We also detected A. thermomutatus, another inherently voriconazole-resistant Aspergillus spp., at AUH. This species has been detected in one CF patient at RH previously (Mortensen et al., 2011a) illustrating that resistant Aspergillus infection is not limited to A. fumigatus in this setting.
When we compare the present study’s result to others, the current overall azole-R rate of 7.3% in the total CF population corresponded well with CF studies from other European countries [France, Germany, the Netherlands (Bader et al., 2013; Seufert et al., 2018; Engel et al., 2019; Lavergne et al., 2019), and the United States (Table 1)] (Stevens et al., 2016). Internationally, published azole resistance rates have varied greatly. No azole resistance was reported from a Portuguese center (Amorim et al., 2010), in one of two Italian centers (Prigitano et al., 2017), and in five out of 12 German centers (Seufert et al., 2018; Table 3). In contrast, Abdolrasouli et al. found a concerning high resistance rate of 16.2% in CF patients specifically, which could be reflected upon the patient group at a cardiothoracic center in United Kingdom following up CF patients (Abdolrasouli et al., 2018). Guegan et al. also found a high azole-R rate of 15.2%, but in a limited CF population of 33 patients (Guegan et al., 2018; Table 1).
The major strength of the present study is the fact that it allowed a 10-year perspective on azole-resistant Aspergillus and a nationwide surveillance perspective of the current mold epidemiology in CF patients in Denmark. Since we also included Aspergillus species other than A. fumigatus, we also reported mutations in A. terreus and furthermore detailed information on STRAf genotyping in one patient with A. fumigatus. A limitation, however, is that we do not have information regarding the clinical relevance of the retrieved A. fumigatus, nor do we have any information regarding preceding antifungal use.
In conclusion, azole-R Aspergillus is increasing in proportion and complexity among Danish CF patients. The larger and increasing proportion involved resistant A. fumigatus of environmental origin, and novel genotypes in both A. fumigatus and A. terreus were found. Although the isolation of Aspergillus may reflect contamination or transient colonization and thus include patients in whom antifungal therapy is not indicated, the continuously emerging reports of azole-resistant Aspergillus is worrisome, and resistance remains a significant challenge. This is of concern as effective alternative treatments to azoles are lacking and as it suggests that azole-resistant A. fumigatus may also be an increasing challenge in other patient populations at risk for aspergillus disease.
Data Availability Statement
The raw data can be provided from the corresponding author according to the Danish law.
Ethics Statement
Ethical review and approval was not required for the study on human participants in accordance with the local legislation and institutional requirements. Written informed consent from the patients was not required to participate in this study in accordance with the national legislation and the institutional requirements.
Author Contributions
MR, MA, and HJ designed the study. HJ, JG, and LK were responsible for primary cultures and isolation. MA, JG, and LK were responsible for the susceptibility testing. RH and NA-C performed the molecular analysis. MR performed the data management. MR and MA wrote and revised the manuscript after review from all co-authors. All authors contributed to the article and approved the submitted version.
Conflict of Interest
MR has over the past 5 years received speaker honoraria from BMS and, unrestricted research and travel grants from Novartis. JG has, over the past 5 years, received travel grants from Gilead and speaker honoraria from Gilead and MSD. RH has received unrestricted research grants from Gilead and conference meeting grants from Gilead, MSD, Pfizer, and Astellas. MA has, over the past 5 years, received research grants/contract work (paid to the SSI) from Amplyx, Basilea, Cidara, F2G, Gilead, Novabiotics, Scynexis, and T2Biosystems and speaker honoraria (personal fee) from Astellas, Gilead, Novartis, MSD, and Seges. She is the current chairman of the EUCAST-AFST.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge the staff at the laboratory at the Mycology Unit at Statens Serum Institut, the Department of Clinical Microbiology at the University of Copenhagen, Rigshospitalet, and the Department of Microbiology at Aarhus University Hospital.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.01850/full#supplementary-material
References
Abdolrasouli, A., Scourfield, A., Rhodes, J., Shah, A., Elborn, J. S., Fisher, M. C., et al. (2018). High prevalence of triazole resistance in clinical Aspergillus fumigatus isolates in a specialist cardiothoracic centre. Int. J. Antimicrob. Agents 52, 637–642. doi: 10.1016/j.ijantimicag.2018.08.004
Amorim, A., Guedes-Vaz, L., and Araujo, R. (2010). Susceptibility to five antifungals of Aspergillus fumigatus strains isolated from chronically colonised cystic fibrosis patients receiving azole therapy. Int. J. Antimicrob. Agents 35, 396–399. doi: 10.1016/j.ijantimicag.2009.12.007
Arendrup, M. C., Cuenca-Estrella, M., Lass-Flörl, C., and Hope, W. W. (2013). Breakpoints for antifungal agents: An update from EUCAST focussing on echinocandins against Candida spp. and triazoles against Aspergillus spp. Drug Resist. Updat. 16, 81–95. doi: 10.1016/j.drup.2014.01.001
Arendrup, M. C., Jensen, R. H., Grif, K., Skov, M., Pressler, T., Johansen, H. K., et al. (2012). In vivo emergence of Aspergillus terreus with reduced azole susceptibility and a Cyp51a M217I alteration. J. Infect. Dis. 206, 981–985. doi: 10.1093/infdis/jis442
Arendrup, M. C., Meletiadis, J., Mouton, J. W., Guinea, J., Cuenca-Estrella, M., Lagrou, K., et al. (2016). EUCAST technical note on isavuconazole breakpoints for Aspergillus, itraconazole breakpoints for Candida and updates for the antifungal susceptibility testing method documents. Clin. Microbiol. Infect. 22, 571.e1–571.e4. doi: 10.1016/j.cmi.2016.01.017
Arendrup, M. C., Verweij, P. E., Mouton, J. W., Lagrou, K., and Meletiadis, J. (2017). Multicentre validation of 4-well azole agar plates as a screening method for detection of clinically relevant azole-resistant Aspergillus fumigatus. J. Antimicrob. Chemother. 72, 3325–3333. doi: 10.1093/jac/dkx319
Astvad, K. M. T., Jensen, R. H., Hassan, T. M., Mathiasen, E. G., Thomsen, G. M., Pedersen, U. G., et al. (2014). First detection of TR 46/Y121F/T289A and TR34/L98H alterations in Aspergillus fumigatus isolates from azole-naive patients in Denmark despite negative findings in the environment. Antimicrob. Agents Chemother. 58, 5096–5101. doi: 10.1128/aac.02855-14
Bader, O., Weig, M., Reichard, U., Lugert, R., Kuhns, M., Christner, M., et al. (2013). cyp51A -based mechanisms of Aspergillus fumigatus azole drug resistance present in clinical samples from germany. Antimicrob. Agents Chemother. 57, 3513–3517. doi: 10.1128/aac.00167-13
Burgel, P.-R., Baixench, M.-T., Amsellem, M., Audureau, E., Chapron, J., Kanaan, R., et al. (2012). High prevalence of azole-resistant aspergillus fumigatus in adults with cystic fibrosis exposed to itraconazole. Antimicrob. Agents Chemother. 56, 869–874. doi: 10.1128/aac.05077-11
Burgel, P. R., Paugam, A., Hubert, D., and Martin, C. (2016). Aspergillus fumigatus in the cystic fibrosis lung: pros and cons of azole therapy. Infect. Drug Resist. 9, 229–238. doi: 10.2147/idr.s63621
Camps, S. M. T., Dutilh, B. E., Arendrup, M. C., Rijs, A. J. M. M., Snelders, E., Huynen, M. A., et al. (2012). Discovery of a hapE Mutation That Causes Azole Resistance in Aspergillus fumigatus through Whole Genome Sequencing and Sexual Crossing. PLoS One 7:e50034. doi: 10.1371/journal.pone.0050034
Carsin, A., Romain, T., Ranque, S., Reynaud-Gaubert, M., Dubus, J.-C., Mège, J.-L., et al. (2017). Aspergillus fumigatus in cystic fibrosis: an update on immune interactions and molecular diagnostics in allergic bronchopulmonary aspergillosis. Allergy 72, 1632–1642. doi: 10.1111/all.13204
Chowdhary, A., Kathuria, S., Xu, J., Sharma, C., Sundar, G., Singh, P. K., et al. (2012). Clonal expansion and emergence of environmental multiple-triazole-resistant Aspergillus fumigatus strains carrying the TR34/L98H mutations in the cyp51A gene in India. PLoS One 7:e52871. doi: 10.1371/journal.pone.0052871
De Groot, T., and Meis, J. F. (2019). Microsatellite stability in STR analysis Aspergillus fumigates depends on number of repeat units. Front. Cell Infect. Microbiol. 9:82. doi: 10.3389/fcimb.2019.00082
De Valk, H. A., Meis, J. F. G. M., Curfs, I. M., Muehlethaler, K., Mouton, J. W., and Klaassen, C. H. W. (2005). Use of a novel panel of nine short tandem repeats for exact and high-resolution fingerprinting of Aspergillus fumigatus isolates. J. Clin. Microbiol. 43, 4112–4120. doi: 10.1128/jcm.43.8.4112-4120.2005
Engel, T. G. P., Slabbers, L., de Jong, C., Melchers, W. J. G., Hagen, F., Verweij, P. E., et al. (2019). Prevalence and diversity of filamentous fungi in the airways of cystic fibrosis patients – A Dutch, multicentre study. J. Cyst. Fibros. 18, 221–226. doi: 10.1016/j.jcf.2018.11.012
Felton, I. C., and Simmonds, N. J. (2014). Aspergillus and cystic fibrosis: old disease – new classifications. Curr. Opin. Pulm. Med. 20, 632–638. doi: 10.1097/mcp.0000000000000106
Fischer, J., van Koningsbruggen-Rietschel, S., Rietschel, E., Vehreschild, M. J. G. T., Wisplinghoff, H., Kronke, M., et al. (2014). Prevalence and molecular characterization of azole resistance in Aspergillus spp. isolates from German cystic fibrosis patients. J. Antimicrob. Chemother. 69, 1533–1536. doi: 10.1093/jac/dku009
Garczewska, B., Jarzynka, S., Kuś, J., Skorupa, W., and Augustynowicz-Kopeć, E. (2016). Fungal infection of cystic fibrosis patients - single center experience. Pneumonol. Alergol. Pol. 84, 151–159. doi: 10.5603/piap.2016.0017
Glass, N. L., and Donaldson, G. C. (1995). Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 61, 1323–1330. doi: 10.1128/aem.61.4.1323-1330.1995
Guegan, H., Chevrier, S., Belleguic, C., Deneuville, E., Robert-Gangneux, F., and Gangneux, J. P. (2018). Performance of molecular approaches for Aspergillus detection and azole resistance surveillance in cystic fibrosis. Front. Microbiol. 9:531. doi: 10.3389/fmicb.2018.00531
Guinea, J., Verweij, P. E., Meletiadis, J., Mouton, J. W., Barchiesi, F., Arendrup, M. C., et al. (2019). How to: EUCAST recommendations on the screening procedure E.Def 10.1 for the detection of azole resistance in Aspergillus fumigatus isolates using four-well azole-containing agar plates. Clin. Microbiol. Infect. 25, 681–687. doi: 10.1016/j.cmi.2018.09.008
Güngör, Ö, Sampaio-Maia, B., Amorim, A., Araujo, R., and Erturan, Z. (2018). Determination of azole resistance and TR34/L98H mutations in isolates of Aspergillus section fumigati from Turkish cystic fibrosis patients. Mycopathologia 183, 913–920. doi: 10.1007/s11046-018-0297-y
Hagiwara, D., Watanabe, A., Kamei, K., and Goldman, G. H. (2016). Epidemiological and genomic landscape of azole resistance mechanisms in Aspergillus fungi. Front. Microbiol. 7:1382. doi: 10.3389/fmicb.2016.01382
Hamprecht, A., Morio, F., Bader, O., Le Pape, P., Steinmann, J., and Dannaoui, E. (2018). Azole resistance in aspergillus fumigatus in patients with cystic fibrosis: a matter of concern? Mycopathologia 183, 151–160. doi: 10.1007/s11046-017-0162-4
Hare, R. K., Gertsen, J. B., Astvad, K. M. T., Degn, K. B., Løkke, A., Stegger, M., et al. (2019). In vivo selection of a unique tandem repeat mediated azole resistance mechanism (TR 120) in Aspergillus fumigatus cyp51A, Denmark. Emerg. Infect. Dis. 25, 577–580. doi: 10.3201/eid2503.180297
Howard, S. J., Pasqualotto, A. C., Anderson, M. J., Leatherbarrow, H., Albarrag, A. M., Harrison, E., et al. (2013). Major variations in Aspergillus fumigatus arising within aspergillomas in chronic pulmonary aspergillosis. Mycoses 56, 434–441. doi: 10.1111/myc.12047
Imbert, S., Normand, A. C., Gabriel, F., Cassaing, S., Bonnal, C., Costa, D., et al. (2019). Multi-centric evaluation of the online MSI platform for the identification of cryptic and rare species of Aspergillus by MALDI-TOF. Med. Mycol. 57, 962–968. doi: 10.1093/mmy/myz004
Jensen, R. H., Hagen, F., Astvad, K. M. T., Tyron, A., Meis, J. F., and Arendrup, M. C. (2016). Azole-resistant Aspergillus fumigatus in Denmark: a laboratory-based study on resistance mechanisms and genotypes. Clin. Microbiol. Infect. 22, 570.e1–570.e17.
Lavergne, R.-A., Morio, F., Danner-Boucher, I., Horeau-Langlard, D., David, V., Hagen, F., et al. (2019). One year prospective survey of azole resistance in Aspergillus fumigatus at a French cystic fibrosis reference centre: prevalence and mechanisms of resistance. J. Antimicrob. Chemother. 74, 1884–1889. doi: 10.1093/jac/dkz144
Lestrade, P. P. A., Meis, J. F., Melchers, W. J. G., and Verweij, P. E. (2019). Triazole resistance in Aspergillus fumigatus: recent insights and challenges for patient management. Clin. Microbiol. Infect. 25, 799–806. doi: 10.1016/j.cmi.2018.11.027
Maertens, J. A., Raad, I. I., Marr, K. A., Patterson, T. F., Kontoyiannis, D. P., Cornely, O. A., et al. (2016). Isavuconazole versus voriconazole for primary treatment of invasive mould disease caused by Aspergillus and other filamentous fungi (SECURE): a phase 3, randomised-controlled, non-inferiority trial. Lancet 387, 760–769. doi: 10.1016/s0140-6736(15)01159-9
Morio, F., Aubin, G. G., Danner-Boucher, I., Haloun, A., Sacchetto, E., Garcia-Hermoso, D., et al. (2012). High prevalence of triazole resistance in Aspergillus fumigatus, especially mediated by TR/L98H, in a French cohort of patients with cystic fibrosis. J. Antimicrob. Chemother. 67, 1870–1873. doi: 10.1093/jac/dks160
Mortensen, K. L., Jensen, R. H., Johansen, H. K., Skov, M., Pressler, T., Howard, S. J., et al. (2011a). Aspergillus species and other molds in respiratory samples from patients with cystic fibrosis: a laboratory-based study with focus on Aspergillus fumigatus azole resistance. J. Clin. Microbiol. 49, 2243–2251. doi: 10.1128/jcm.00213-11
Mortensen, K. L., Johansen, H. K., Fuursted, K., Knudsen, J. D., Gahrn-Hansen, B., Jensen, R. H., et al. (2011b). A prospective survey of Aspergillus spp. in respiratory tract samples: prevalence, clinical impact and antifungal susceptibility. Eur. J. Clin. Microbiol. Infect. Dis. 30, 1355–1363. doi: 10.1007/s10096-011-1229-7
Mortensen, K. L., Mellado, E., Lass-Florl, C., Rodriguez-Tudela, J. L., Johansen, H. K., and Arendrup, M. C. (2010). Environmental study of azole-resistant Aspergillus fumigatus and other Aspergilli in Austria, Denmark, and Spain. Antimicrob. Agents Chemother. 54, 4545–4549. doi: 10.1128/aac.00692-10
Normand, A. C., Becker, P., Gabriel, F., Cassagne, C., Accoceberry, I., Gari-Toussaint, M., et al. (2017). Validation of a new web application for identification of fungi by use of matrix-assisted laser desorption ionization–time of flight mass spectrometry. J. Clin. Microbiol. 55, 2661–2670.
Patterson, T. F., Thompson, G. R., Denning, D. W., Fishman, J. A., Hadley, S., Herbrecht, R., et al. (2016). Practice guidelines for the diagnosis and management of Aspergillosis: 2016 update by the infectious diseases society of america. Clin. Infect. Dis. 63, e1–e60.
Periselneris, J., Nwankwo, L., Schelenz, S., Shah, A., and Armstrong-James, D. (2019). Posaconazole for the treatment of allergic bronchopulmonary aspergillosis in patients with cystic fibrosis. J. Antimicrob. Chemother. 74, 1701–1703. doi: 10.1093/jac/dkz075
Pihet, M., Carrere, J., Cimon, B., Chabasse, D., Delhaes, L., Symoens, F., et al. (2009). Occurrence and relevance of filamentous fungi in respiratory secretions of patients with cystic fibrosis – a review. Med. Mycol. 47, 387–397. doi: 10.1080/13693780802609604
Prigitano, A., Esposto, M. C., Biffi, A., De Lorenzis, G., Favuzzi, V., Koncan, R., et al. (2017). Triazole resistance in Aspergillus fumigatus isolates from patients with cystic fibrosis in Italy. J. Cyst. Fibros. 16, 64–69. doi: 10.1016/j.jcf.2016.06.006
Risum, M., Hare, R. K., Abou-Ckara, Gertsen, J. B., Kristensen, L., and Arendrup, M. C. (2019). “Azole resistance in Aspergillus spp. Preliminary six months data from the newly established surveillance in Denmark”, in DANMAP 2018. Textbox 5.4(Part 2), 77–78.
Rivero-Menendez, O., Soto-Debran, J. C., Medina, N., Lucio, J., Mellado, E., and Alastruey-Izquierdo, A. (2019). Molecular identification, antifungal susceptibility testing, and mechanisms of azole resistance in Aspergillus species received within a surveillance program on antifungal resistance in Spain. Antimicrob. Agents Chemother. 63, 1–10.
Rybak, J. M., Ge, W., Wiederhold, N. P., Parker, J. E., Kelly, S. L., Rogers, P. D., et al. (2019). Mutations in hmg1, challenging the paradigm of clinical triazole resistance in aspergillus fumigatus. mBio 10:e00437-19. doi: 10.1128/mBio.00437-19
Seufert, R., Sedlacek, L., Kahl, B., Hogardt, M., Hamprecht, A., Haase, G., et al. (2018). Prevalence and characterization of azole-resistant Aspergillus fumigatus in patients with cystic fibrosis: a prospective multicentre study in Germany. J. Antimicrob. Chemother. 73, 2047–2053. doi: 10.1093/jac/dky147
Skov, M., Mortensen, K. L., and Helweg-Larsen, J. (2017). Aspergillus lungesygdom Hos Patienter Med Cystisk Fibrose. Available online at: https://vip.regionh.dk/VIP/Redaktoer/130132.nsf/vLookupUpload/ATTACH-RHAP-AVHB3E/$FILE/29.11.17%20Aspergillus%20instruks.pdf and https://e-dok.rm.dk/edok/admin/GUI.nsf/desktop.html?Open (accessed July 31, 2020).
Snelders, E., Camps, S. M. T., Karawajczyk, A., Schaftenaar, G., Kema, G. H. J., van der Lee, H. A., et al. (2012). Triazole fungicides can induce cross-resistance to medical triazoles in Aspergillus fumigatus. PLoS One 7:e31801. doi: 10.1371/journal.pone.0031801
Stensvold, C. R., Jørgensen, L. N., and Arendrup, M. C. (2012). Azole-resistant invasive Aspergillosis: relationship to agriculture. Curr. Fungal Infect. Rep. 6, 178–191. doi: 10.1007/s12281-012-0097-7
Stevens, D. A., Moss, R. B., Hernandez, C., Clemons, K. V., and Martinez, M. (2016). Effect of media modified to mimic cystic fibrosis sputum on the susceptibility of Aspergillus fumigatus, and the frequency of resistance at one center. Antimicrob. Agents Chemother. 60, 2180–2184. doi: 10.1128/aac.02649-15
Ullmann, A. J., Aguado, J. M., Arikan-Akdagli, S., Denning, D. W., Groll, A. H., Lagrou, K., et al. (2018). Diagnosis and management of Aspergillus diseases: executive summary of the 2017 ESCMID-ECMM-ERS guideline. Clin. Microbiol. Infect. 24, e1–e38. doi: 10.1016/j.cmi.2018.01.002
van Leer-Buter, C., Takes, R. P., Hebeda, K. M., Melchers, W. J., and Verweij, P. E. (2007). Aspergillosis-and a misleading sensitivity result. Lancet 370:102. doi: 10.1016/s0140-6736(07)61055-1
Verweij, P. E., Zhang, J., Debets, A. J. M., Meis, J. F., van de Veerdonk, F. L., Schoustra, S. E., et al. (2016). In-host adaptation and acquired triazole resistance in Aspergillus fumigatus: a dilemma for clinical management. Lancet Infect. Dis. 16, e251–e260. doi: 10.1016/s1473-3099(16)30138-4
Williams, C., Ranjendran, R., and Ramage, G. (2016). Pathogenesis of fungal infections in cystic fibrosis. Curr. Fungal Infect. Rep. 10, 163–169. doi: 10.1007/s12281-016-0268-z
Zhang, J., Snelders, E., Zwaan, B. J., Schoustra, S. E., Meis, J. F., van Dijk, K., et al. (2017). A novel environmental azole resistance mutation in Aspergillus fumigatus and a possible role of sexual reproduction in its emergence. mBio. 8:e00791-17. doi: 10.1128/mBio.00791-17
Keywords: Aspergillus, cystic fibrosis, azoles, resistance, mutation
Citation: Risum M, Hare RK, Gertsen JB, Kristensen L, Johansen HK, Helweg-Larsen J, Abou-Chakra N, Pressler T, Skov M, Jensen-Fangel S and Arendrup MC (2020) Azole-Resistant Aspergillus fumigatus Among Danish Cystic Fibrosis Patients: Increasing Prevalence and Dominance of TR34/L98H. Front. Microbiol. 11:1850. doi: 10.3389/fmicb.2020.01850
Received: 12 January 2020; Accepted: 15 July 2020;
Published: 13 August 2020.
Edited by:
Uwe Groß, University Medical Center Göttingen, GermanyReviewed by:
Joerg Steinmann, Paracelsus Medical Private University, Nuremberg, GermanyEmilia Mellado, Instituto de Salud Carlos III (ISCIII), Spain
Copyright © 2020 Risum, Hare, Gertsen, Kristensen, Johansen, Helweg-Larsen, Abou-Chakra, Pressler, Skov, Jensen-Fangel and Arendrup. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maiken Cavling Arendrup, bWFjYUBzc2kuZGs=