 Saskia Windisch1*†
Saskia Windisch1*† Loreen Sommermann2†
Loreen Sommermann2† Doreen Babin3†
Doreen Babin3† Soumitra Paul Chowdhury4
Soumitra Paul Chowdhury4 Rita Grosch5
Rita Grosch5 Narges Moradtalab1
Narges Moradtalab1 Frank Walker6
Frank Walker6 Birgit Höglinger6
Birgit Höglinger6 Abbas El-Hasan7Wolfgang Armbruster8
Abbas El-Hasan7Wolfgang Armbruster8 Joseph Nesme9
Joseph Nesme9 Søren Johannes Sørensen9Ingo Schellenberg2Jörg Geistlinger2
Søren Johannes Sørensen9Ingo Schellenberg2Jörg Geistlinger2 Kornelia Smalla3
Kornelia Smalla3 Michael Rothballer4
Michael Rothballer4 Uwe Ludewig1
Uwe Ludewig1 Günter Neumann1*
Günter Neumann1*- 1Department of Nutritional Crop Physiology, Institute of Crop Science, University of Hohenheim, Stuttgart, Germany
- 2Institute of Bioanalytical Sciences (IBAS), Anhalt University of Applied Sciences, Bernburg, Germany
- 3Institute for Epidemiology and Pathogen Diagnostics, Julius Kühn Institute – Federal Research Centre for Cultivated Plants, Braunschweig, Germany
- 4Institute of Network Biology, Helmholtz Zentrum München, Neuherberg, Germany
- 5Plant-Microbe Systems, Leibniz Institute of Vegetable and Ornamental Crops, Großbeeren, Germany
- 6Central Chemical-Analytical Laboratory, Institute of Phytomedicine, University of Hohenheim, Stuttgart, Germany
- 7Department of Phytopathology, Institute of Phytomedicine, University of Hohenheim, Stuttgart, Germany
- 8Department of Food Chemistry and Analytical Chemistry, Institute of Food Chemistry, University of Hohenheim, Stuttgart, Germany
- 9Department of Biology, Section of Microbiology, University of Copenhagen, Copenhagen, Denmark
Fertilization management can affect plant performance and soil microbiota, involving still poorly understood rhizosphere interactions. We hypothesized that fertilization practice exerts specific effects on rhizodeposition with consequences for recruitment of rhizosphere microbiota and plant performance. To address this hypothesis, we conducted a minirhizotron experiment using lettuce as model plant and field soils with contrasting properties from two long-term field experiments (HUB-LTE: loamy sand, DOK-LTE: silty loam) with organic and mineral fertilization history. Increased relative abundance of plant-beneficial arbuscular mycorrhizal fungi and fungal pathotrophs were characteristic of the rhizospheres in the organically managed soils (HU-org; BIODYN2). Accordingly, defense-related genes were systemically expressed in shoot tissues of the respective plants. As a site-specific effect, high relative occurrence of the fungal lettuce pathogen Olpidium sp. (76–90%) was recorded in the rhizosphere, both under long-term organic and mineral fertilization at the DOK-LTE site, likely supporting Olpidium infection due to a lower water drainage potential compared to the sandy HUB-LTE soils. However, plant growth depressions and Olpidium infection were exclusively recorded in the BIODYN2 soil with organic fertilization history. This was associated with a drastic (87–97%) reduction in rhizosphere abundance of potentially plant-beneficial microbiota (Pseudomonadaceae, Mortierella elongata) and reduced concentrations of the antifungal root exudate benzoate, known to be increased in presence of Pseudomonas spp. In contrast, high relative abundance of Pseudomonadaceae (Gammaproteobacteria) in the rhizosphere of plants grown in soils with long-term mineral fertilization (61–74%) coincided with high rhizosphere concentrations of chemotactic dicarboxylates (succinate, malate) and a high C (sugar)/N (amino acid) ratio, known to support the growth of Gammaproteobacteria. This was related with generally lower systemic expression of plant defense genes as compared with organic fertilization history. Our results suggest a complex network of belowground interactions among root exudates, site-specific factors and rhizosphere microbiota, modulating the impact of fertilization management with consequences for plant health and performance.
Introduction
Fertilization practices are a central component of agricultural management having a strong impact on plant performance including crop yield and quality, plant health and resistance against abiotic and biotic stresses. However, limitations in the adaptation of fertilizer inputs to actual crop demands can cause undesirable environmental effects, such as nutrient leaching, alteration of soil pH, eutrophication of surface waters, emission of greenhouse gases, and soil degradation (Loreau et al., 2001; Foley et al., 2005; Robertson and Vitousek, 2009; Geisseler and Scow, 2014). Due to the rising demand for more sustainable agricultural production systems with reduced inputs of agrochemicals and the closing of nutrient cycles to counteract detrimental ecological side effects, investigations into alternative management practices gain in importance. The availability of essential plant nutrients in farmland is controlled by interactions between fertilization management and microbial processes (Schmidt et al., 2019). Nearly all relevant soil processes are influenced by microbial activities (Mäder et al., 2002), which underlines the importance of considering plant–microbial interactions in this context. Various beneficial aspects have been linked with organic fertilization strategies, such as increased soil organic matter, stimulation of microbial activity (Lori et al., 2017), increased microbial biomass and diversity (Esperschütz et al., 2007; Hartmann et al., 2014; Francioli et al., 2016; Schmid et al., 2018), and enrichment in plant–beneficial microorganisms (Francioli et al., 2016). However, these findings cannot be generalized and are influenced by additional factors, such as soil properties, climatic conditions or specific practices of crop management. In addition, plant roots are similarly powerful drivers of the assemblage of the rhizosphere microbial community, which exhibits distinct structural and functional differences compared with the bulk soil microbiota (Turner et al., 2013). Organic rhizodeposition shapes rhizosphere microbiota (Walters et al., 2003; Narula et al., 2009) by providing nutrients, signaling compounds, and bio-active substances against pests and pathogens (Doornbos et al., 2012; Chaparro et al., 2013; Baetz and Martinoia, 2014; Windisch et al., 2017). The quantity and composition of rhizodeposits are highly variable and influenced by soil texture, plant nutritional status, abiotic and biotic stress factors, plant genotype, and rhizosphere microbiota (Neumann and Römheld, 2007). However, the impact of fertilization management on rhizodeposits, triggering the selective recruitment of rhizosphere microbiota with potential feed-back loops on plant performance and health status, still remains an open question.
A recent study demonstrated that the fertilization legacy of the soil contributed to the assemblage of rhizosphere bacterial communities in lettuce (Chowdhury et al., 2019). The crucial role of the rhizosphere microbiota for plant performance and health has been comprehensively reviewed (Berendsen et al., 2012; Berg et al., 2016). Furthermore, a relationship between plant health and agricultural management conferred via soil microorganisms has been postulated (Lapsansky et al., 2016; van der Putten et al., 2016; Bakker et al., 2018). Chowdhury et al. (2019) provided first experimental evidence for induction of physiological adaptations under long-term organic vs. mineral fertilization that helped lettuce plants to cope with environmental stresses. A better understanding of these interactions, triggering plant–beneficial interactions but also detrimental rhizosphere effects, could therefore contribute to the development of practical approaches toward improved crop productivity and agroecosystem sustainability (Schmid et al., 2018) according to the concept of “soil biological engineering” (Bender et al., 2016). However, a more detailed understanding particularly of the critical plant factors determining these interactions still represents a major knowledge gap.
In this context, our study was initiated as a complementary approach to the recently published study by Chowdhury et al. (2019). Soils with contrasting physicochemical properties and long-term organic or mineral fertilization histories were investigated. Bacterial, archaeal and fungal communities, the composition of organic compounds in the rhizosphere soil solution as well as plant performance and expression of stress-related genes were analyzed to obtain a more holistic picture of plant-microbe interactions.
We hypothesized that long-term fertilization practices will result in characteristic patterns and chemical composition of the rhizosphere soil solution, with impact on soil microbiota and the recruitment of rhizosphere microbiota. This would affect the performance and health of the model plant lettuce. The final aim was the identification of rhizosphere metabolite profiles characteristic of the investigated long-term fertilization strategies potentially related to alterations in the assemblage of rhizosphere microbiota and plant performance.
Materials and Methods
Soil Sampling and Setup of the Minirhizotron Experiment
Field soils with contrasting properties in terms of soil type and fertilization history, originating from the two long-term fertilization experiments DOK-LTE belonging to the Research Institute of Organic Agriculture (FiBL; Therwil, Switzerland since 1978) and HUB-LTE belonging to the Humboldt-Universität zu Berlin (HUB; Thyrow, Germany since 2006) were used for plant cultivation. The field trial (i) DOK-LTE on a Haplic Luvisol (silty loam) compares bio-dynamic (compost and manure fertilizers with biodynamic preparations; BIODYN2) vs. full mineral NPK (CONMIN) fertilization. The fertilization intensity of the organic system in BIODYN2 was based on the fodder produced in the crop rotation and reflects the intensity of Swiss organic farms. The mineral fertilizer level in CONMIN was adjusted to plant-specific Swiss standard recommendations (Mäder et al., 2002). The field trial (ii) HUB-LTE on a Retisol (loamy sand) compares application of organic farmyard manure (green, and cattle manure; HU-org) vs. full mineral NPK (HU-min) fertilization. The mineral fertilizer level in HU-min was adjusted to soil fertility and yield performance in the northeast of Germany. Nitrogen was applied as calcium ammonium nitrate (KAS), phosphorus as triple-superphosphate (TSP) and potassium as Patentkali® (K + S Minerals and Agriculture GmbH, Kassel, Germany) besides an incorporation of harvested straw of winter and cover crops. The fertilization intensity of the organic system in HU-org was based on cattle and green manure from cover crops with legumes mixture. In order to obtain field soil for the minirhizotron experiment, soil sampling was performed after harvest of standing crops from the respective upper 30 cm soil layer from each field trial and combined as representative sample of 15 sampling spots. For homogenization, the soil was air-dried, sieved (4 mm mesh size) and stored in the dark at 7°C. For reactivation of the microbial communities prior to the experiment, experimental soil and soil as control without lettuce cultivation (bulk soil) was incubated for two weeks in the dark with a 20°C day/15°C night temperature regime at 100-hPA water potential (T5 tensiometer, UMS, AG, München, Germany). Detailed soil characteristics, management practices, and physicochemical parameters of the experimental soils are summarized in Table 1.
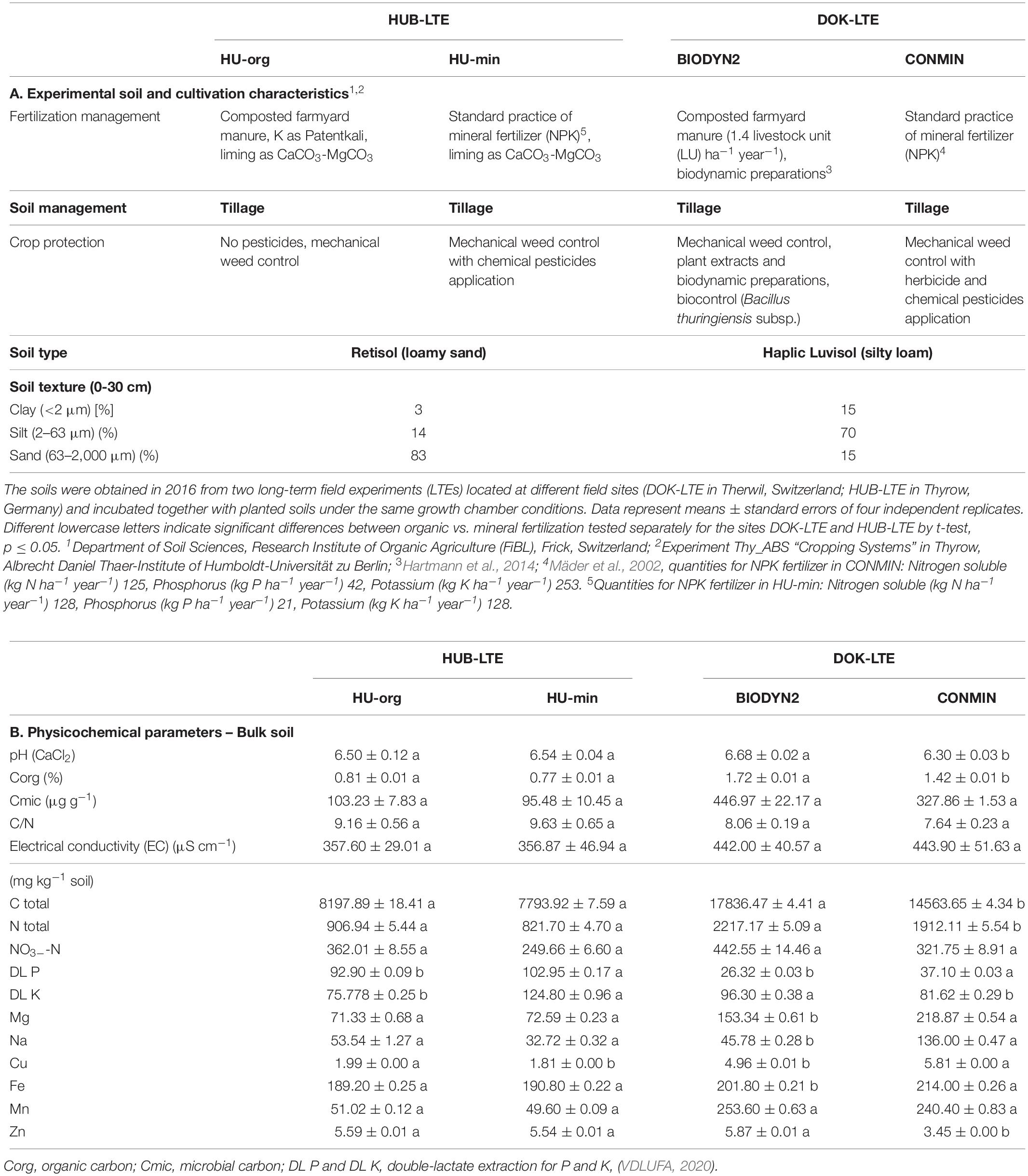
Table 1. Characteristics of the field soils (A) and physicochemical parameters of bulk soils (B).
Lettuce (Lactuca sativa L. cv. Tizian, Syngenta, Bad Salzuflen, Germany) cultivation, harvest, and root exudate sampling were performed as described by Neumann et al. (2014). To achieve homogenous plant development, lettuce seedlings were pre-cultivated until the five-leaf stage (BBCH 15) in a soil-sand mixture (70/30: w/w). Thereafter, the seedlings were transferred to minirhizotrons (0.6 kg of the soil–sand mixture 70/30) made from PVC tubes (height 22 cm; diameter 9 cm) with transparent root observation windows. The minirhizotrons were fixed at an angle of 45° to stimulate root growth along the root observation window for exudate sampling.
Full mineral N fertilization (517 mg N kg–1 substrate as YaraLiva Calcinit [Ca(NO3)2], Yara, Oslo, Norway) was supplied to cover the plant demand during the culture period. The first half of the recommended N amount for lettuce was supplied after seedling transfer to minirhizotrons, the second half provided two weeks later. The soil moisture level was adjusted to 18–20% w/w by addition of demineralized water (25 ml kg–1 soil) every second day throughout the culture period. Lettuce seedlings were cultivated in a growth chamber with a 16 h light period at 420 μmol m–2 s–1, 60% relative humidity, and a 20°C/15°C day/night temperature regime, with eight replicates per treatment in a randomized block design. Biomass of plants, root characteristics and nutritional status of lettuce shoots were analyzed. To confirm general treatment responses in plant growth and visual symptoms of pathogen infections, the experiment was performed twice with cultivation periods of six and nine weeks, respectively, with a more detailed analysis of soil microbiota, rhizosphere chemistry and gene expression after nine weeks.
Analysis of Mineral Nutrients in Soil and Shoot Tissue
The content of macro- and micronutrients in bulk soil and plant samples were analyzed according to the certified protocols of the Association of German Agricultural Analytic and Research Institutes, VDLUFA, Germany (VDLUFA, 2020). After determination of fresh shoot biomass, shoots and soil samples were oven-dried at 60°C for three days and dry biomass was recorded. Subsequently, shoots were stored in a desiccator for another two days and 200–500 mg of dry plant material and 4 g of soil were subjected to microwave digestion (Mars 6, CEM, Charlotte, NC, United States) with 5 ml of HNO3 (65%) and 3 ml of H2O2 (30%) for 20–30 min.
Potasium, Na, P, Mg, Fe, Mn, Cu, and Zn concentrations in shoot tissue and soils were determined via inductively coupled plasma optical emission spectrometry (ICP-OES); total C and N were determined via elemental analysis (Elementary Vario El cube, Elementar, Langenselbold, Germany). Soil pH was determined in calcium chloride solution and salinity via electrical conductivity.
Root Morphology
For analysis of root morphological characteristics, roots of four replicates were washed from soil using sieves (mesh size 0.5–1.0 mm) and fresh and dry biomass were recorded. Morphological characteristics were determined from fresh root samples, stored in 60% (v/v) ethanol. For analysis, root samples submerged in a water film on transparent Perspex trays, were separated with forceps and subsequently digitized using a flat-bed scanner (Epson Expression 1000 XL, Tokyo, Japan). Root length, average root diameter and the proportion of fine roots of the digitized samples were measured by applying the WinRHIZO root analysis software (Regent Instruments, Quebec, QC, Canada). Root hair length was recorded non-destructively along the root observation plane of the minirhizotrons by a video microscope (Stemi 200-c, Zeiss, Oberkochen, Germany). The digitized video images were analyzed using the AxioVision, software, Version 3.1.2.1 (Zeiss, Oberkochen, Germany).
Plant Gene Expression
For gene expression studies, leaf samples were obtained from plants in quadruplicates. Four representative leaves per plant were pooled and snap-frozen in liquid nitrogen (Chowdhury et al., 2019). Homogenized leaf material (100 mg) was subjected to total RNA extraction using the RNeasy Plant Mini Kit (QIAGEN GmbH, Hilden, Germany). RNA was quantified by a NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, United States). Target genes of lettuce (Chowdhury et al., 2019) were selected based on comparisons with functional genes from Arabidopsis thaliana using “The Arabidopsis Information Resource” (Berardini et al., 2015)1. The reference gene glyceraldehyde-3-dehydrogenase was used for normalization of qPCR results. The primer pairs for qPCR were designed using the Primer3Plus software (Untergasser et al., 2007). cDNA was synthesized from 2 μg of total RNA with the High Capacity cDNA Reverse Transcription Kit with RNase Inhibitor (Applied Biosystems, Foster City, CA, United States). The qPCR was performed with Power SYBR Green Supermix (Applied Biosystems, Foster City, CA, United States) using a peqSTAR 96Q thermal cycler (PEQLAB Biotechnologie, Erlangen, Germany). cDNA dilutions (1 μl, 1:4) were used as PCR templates. Each PCR reaction contained 12.5 μl of 2 × Power SYBR Green Supermix, 0.4 μM primers (Eurofins MVG Operon, Ebersberg, Germany), and 1 μl of template in a 25-μl reaction. PCR reactions were heated to 95°C for 3 min and then for 40 cycles with steps of 95°C for 30 s and 60°C for 30 s. The generation of specific PCR products was confirmed by melting curve analysis and gel electrophoresis. The genes, their primer pairs and qPCR conditions used in this study are described in Supplementary Table 1. The 2–ΔΔCt method (Livak and Schmittgen, 2001) was employed for relative quantification. Normalization to the endogenous control for each condition was followed by logarithmic transformation to fold change differences. The standard error of the mean was calculated from the average of technical triplicates, obtained from each of four biological replicates (n = 4).
Sampling of Rhizosphere Soil Solutions
Samples of the rhizosphere soil solution were collected with moist sorption filters (5 mm Ø, filter paper: MN815, Macherey-Nagel, Düren, Germany) placed onto the surface of lateral roots growing along the root observation window according to Haase et al. (2007). Micro-sampling was conducted nine weeks after sowing during vegetative growth of the lettuce plants, to account for most active carbohydrate partitioning to the roots and high root exudation during this phase (Marschner, 1995). For each minirhizotron, rhizosphere sampling was conducted with two 5 mm sorption filters (equivalent to 1 cm root length), applied in triplicate in subapical root zones (1–2 cm behind the root tip) and basal root zones (older, mature parts of the root system, 8–9 cm behind the root tip). As a control, sampling was performed in soil zones without visible root development (soil without root contact). Samples for each minirhizotron were pooled after an incubation time of 4 h and kept frozen at −20°C (Neumann et al., 2014). Rhizosphere soil solution was extracted from sorption filters with 0.6 ml of acetonitrile: H2O (1:1).
Analysis of Carboxylates
Aliquots of 40 μl obtained from 0.6 ml sorption filter extract were evaporated to dryness at 30°C, using a SpeedVac Concentrator (Savant, Farmington, CT, United States) and re-dissolved in 400 μl of high performance liquid chromatography (HPLC) elution buffer (18 mM KH2PO4, pH 2.1 adjusted with H3PO4; Neumann, 2006). Carboxylates were determined according to the method described by Neumann (2006), using RP-HPLC analysis in the ion suppression mode with isocratic elution (18 mM KH2PO4, pH 2.1). The identification and quantitative determination as organic acids (acetic, malic, lactic, citric, succinic, and fumaric acid) were conducted, using a reversed phase C-18 column (GROM-SIL 120 ODS ST, 5 μm particle size, 290 mm × 4.6 mm equipped with a 20 mm × 4.6 mm guard column with the same stationary phase, Grom, Herrenberg, Germany) with direct UV detection at 210 nm and comparison with known standards. In representative samples, the identity of detected carboxylates was additionally confirmed by commercial enzymatic tests (r-Biopharm, Darmstadt, Germany).
Analysis of Sugars
Aliquots of 400 μl from 0.6 ml sorption filter extract were evaporated to dryness at 55°C, using nitrogen evaporation and re-dissolved in 40 μl acetonitrile: H2O (70:30). Analyses of sugars (fructose, glucose, maltose, and sucrose) in rhizosphere soil solutions of lettuce were performed by HPLC-Evaporative Light Scattering Detector (ELSD) with isocratic elution (acetonitrile: H2O, 75:25) on a Perkin Elmer Series 200 HPLC system with a Sedex Model 80 LT ELSD system (Sedere, Orléans, France) equipped with a Shodex, Ashipak NH2P-40 3E column, 5 μm particle size, 250 mm × 3.0 mm (Shodex, München, Germany) and external standards.
Analysis of Amino Acids
A total of 20 μl aliquots from 0.6 ml sorption filter extract were mixed with 15 μl of derivatization agent (ACCQFLUOR REAG, Waters, Milford, MA, United States) and 65 μl of borate buffer, incubated 10 min at 55°C, followed by addition of 400 μl acetonitrile: H2O (1:4) with modifications of the method of Cohen and Michaud (1993). Determination of amino acids (glutamic acid, asparagine, serine, glutamine, glycine, threonine, histidine, alanine, proline, cysteine, thyrosine, methionine, isoleucine, leucine, and phenylalanine) was performed by HPLC-MS, using a Velos LTQSystem (Thermo Fisher Scientific, Waltham, MA, United States) equipped with a ACCUTAGTM column, 4 μm particle size, 150 mm × 3.9 mm (Waters, Milford, MA, United States) and additional comparison with external standards. Gradient elution was performed with (A) ammonium formate: methanol: H2O (40:9:60) and (B) acetonitrile.
Analysis of Benzoate
For determination of the antifungal compound benzoate 50 μl aliquots of sorption filter extract were mixed with 50 μl of H2O followed by UHPLC-MS analysis on a Velos LTQSystem (Thermo Fisher Scientific, Waltham, MA, United States). Identification and quantitative analysis were conducted on an ACCLAIMTMC30 column, 150 mm × 3 mm (ThermoScientific, Waltham, MA, United States) with gradient elution (solvent A): H2O: acetonitrile (95:5) and (solvent B): acetonitrile and an external standard.
Microbial Community Analyses
Total Community – DNA Extraction
The roots of the respective plants, which were studied for gene expression, were used for analysis of rhizosphere microbiota (bacteria, archaea, and fungi). At first, roots were washed with sterile tap water (Schreiter et al., 2014b). The rhizosphere fraction was obtained from 5 g of representative root samples, which were collected from complete root systems, by 1 min Stomacher treatment (Seward Ltd., Worthing, United Kingdom) followed by centrifugation (Schreiter et al., 2014a). Rhizosphere pellets were kept at −20°C for total community (TC)-DNA extraction. Root-associated soil was sampled from the soil fraction loosely adhering to the root system collected after vigorous shaking. After collection, the soil samples were immediately frozen at −20°C.
Total community-DNA was extracted from root-associated soil (0.5 g) and rhizosphere pellets using the FastPrep-24 bead-beating system and FastDNA Spin Kit for Soil. DNAs were purified with the GeneClean Spin Kit (both MP Biomedicals, Santa Ana, CA, United States).
Bacterial and Archaeal Communities
Bacterial and archaeal communities in root-associated soil and rhizosphere were characterized based on sequencing of the V3-V4 region of 16S rRNA genes amplified using the primer pair 341F (5′-CCTAYGGGRBGCASCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) originally published by Yu et al. (2005) and modified by Sundberg et al. (2013) or Caporaso et al. (2011), respectively, targeting both Bacteria and Archaea kingdoms. Detailed description of PCR amplification, purification, normalization, and amplicon sequencing on an Illumina® MiSeq® platform (2 × 250 cycles; Illumina Inc., San Diego, CA, United States) can be found in the Supplementary Text 1.
Unassembled raw amplicon data were submitted to NCBI Sequence Read Archive (SRA)2 under accession number PRJNA622892. Raw sequence reads were first trimmed of primer sequences used in first PCR using cutadapt (Martin, 2011) and only read pairs for which both primers were found were retained for subsequent analysis. Primer trimmed sequences were then merged, clustered in operational taxonomic units (OTUs) using UPARSE-OTU algorithm (Edgar, 2013) and a 97% pairwise sequence similarity threshold. The taxonomic annotation of each cluster representative sequence was performed using mothur classify.seq function (Schloss et al., 2009) with default parameters and the Ribosomal Database Project trainset 14 formatted for mothur (Cole et al., 2014)3. Only annotations with a confidence threshold above 80% were considered. Sequences classified as chloroplasts, mitochondria, or unclassified at the domain level were removed, resulting in a total of 7,294 OTUs. The average number of quality-filtered sequences per sample was 19,969.
Fungal Communities
High-throughput sequencing based on the Internal Transcribed Spacer (ITS2) region was conducted in root-associated soil and rhizosphere. PCRs using the sample-specific barcoded NGS-primer-pair ITS86F (5′-GTGAATCATCGAATCTTTGAA-3′; Op De Beeck et al., 2014) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′; White et al., 1990) as well as the processing of the ITS2 amplicon pool on an Illumina® MiSeq® platform (paired-end mode, 2 × 300bp) were carried out as previously described with a few modifications (Sommermann et al., 2018). Detailed information of PCR amplification and sequencing can be found in the Supplementary Text 2.
Unassembled raw amplicon reads were submitted to European Nucleotide Archive (ENA)4 under BioProject accession number PRJEB39853. Barcode, primer and adapter trimming were performed based on a customized in-house perl script including the FASTX toolkit5 followed by raw sequence merging using FLASH (Magoč and Salzberg, 2011). Subsequently, the analysis of the resulting sequences was carried out with a local version of the GALAXY Bioinformatics Platform6 based on a database-dependent strategy (Antweiler et al., 2017) using UNITE database v7.2 (UNITE Community, 2017) by applying the closed reference approach (Carter et al., 2017). All sequences were aligned with the database (e-value 0.001) and only results with an alignment length >200 bp and a similarity >97% to the reference were kept. Furthermore, BLAST-PARSER (Antweiler et al., 2017) was used for taxonomic assignment based on the lowest e-value. The fungal OTU abundance table was generated by counting the sequences per assignment and using the SH-numbers from the database as identifier. Sequences not classified to the kingdom “fungi” (0.17%) were removed from the fungal OTU-table. Finally, a total of 1,159 OTUs was obtained with an average of 167,113 high quality sequence reads per sample.
Detection of Pathogen Infection in the Root Tissue
Colonization of Olpidium sp. in lettuce roots was assessed according to the method described by De Cara et al. (2008) with modifications. Soil adhering to seedling roots was removed by washing with running water for 5 min. Subsequently, the roots were assessed visually for root discoloration and small sections (2-4 mm) were randomly excised from the roots of each replicate. Root specimens were then transferred to microscopic slides, mounted with lactophenol aniline blue solution (Merck, Darmstadt, Germany) and examined for the presence of sporangia and resting spores in epidermal cells using an Axioskop microscope (Carl Zeiss Microscopy GmbH, Jena, Germany) equipped with an Axiocam camera and AxioVision SE64 Rel. 4.8 software.
Statistical Analysis
For the statistical analysis of significant differences of the nutritional status of lettuce between treatment groups (separately for each LTE), a one-way ANOVA (factor fertilization) followed by a Tukey-test (p ≤ 0.05 significance level) was performed using the SAS software 9.4 (Institute Inc., Cary, NC, United States). For the statistical evaluation of physicochemical parameters (bulk soil), the expression of stress-related genes in lettuce leaves, and the chemical composition of rhizosphere soil solution, t-test and Tukey’s HSD pairwise testing was applied. Data are presented as means ± standard errors (SE).
Multivariate analyses of microbial communities were carried out by R using the packages edgeR (Robinson et al., 2010; McCarthy et al., 2012), vegan (Oksanen et al., 2019), MASS (Venables and Ripley, 2002), ggplot2 (Wickham, 2016), phyloseq (McMurdie and Holmes, 2013), pheatmap (Kolde, 2019), gplots (Warnes et al., 2019), car (Fox and Weisberg, 2019), and agricolae (De Mendiburu, 2020). Alpha-diversity indices (species richness, Shannon, Pielou) were averaged per replicate over 100 randomly taken subsamples of a size corresponding to the sample with the lowest number of reads in the complete dataset (=6,744 for 16S rRNA gene, 96,750 for ITS). Indices were tested for the effect of microhabitat (root-associated soil and rhizosphere) or fertilization management, respectively, by pairwise t-test (p ≤ 0.05). Non-metric multidimensional scaling (NMDS) was used to ordinate similarity between microbial communities based on relative abundances (Bray–Curtis distance). The effect of site, fertilization and habitat on the microbial community composition was tested by PERMANOVA analysis based on relative abundances (10,000 permutations, Bray–Curtis distance). The non-rarefied community data were checked for differentially abundant microbial taxa between different fertilization managements. Data were analyzed by likelihood ratio tests under negative binomial distribution and generalized linear models (edgeR). In doing so, only taxa with presence in at least three samples over the total dataset using a FDR-corrected p ≤ 0.05 were considered. In order to study the potential relationship between rhizosphere soil solution and the bacterial and archaeal as well as fungal community composition, canonical correspondence analyses (CCA, 999 permutations) were carried out on log10 transformed relative abundances. After checking for linear dependency, environmental variables (organic compounds averaged over basal and subapical root) were fitted onto the CCA ordination by the envfit function (999 permutations). In addition, log transformed relative abundances of Pseudomonadaceae OTUs and fungal OTUs at least classified at genus level (both relative abundance >0.5%) that were significantly enriched (FDR < 0.05) in minerally fertilized soils (tested separately per site; edgeR) were included in envfit. The relative abundance of prevalent genera was graphically displayed in a heatmap (clustering of rows using Euclidean distance). The FUNGuild database (Nguyen et al., 2016) was used to categorize fungal communities at genus level into trophic modes (saprotroph, symbiotroph and pathotroph) and into different guilds for further classification.
Biplots of principle component analysis (PCA) were used to ordinate composition of chemical compounds of the rhizosphere soil solutions, related to their habitat and tested for the effect of fertilization management. The distribution of the data was graphically displayed in a p-dimensional cartesian coordinate system with Euclidean distance by using R Studio software 3.4.1 and prcomp and autoplot functions.
Results
Plant Biomass, Root Growth Characteristics, and Nutritional Status
There were no significant differences for shoot and root dry biomasses of lettuce grown in HU-org vs. HU-min soil (Figure 1A). However, a significant decline in shoot and root dry biomass (by 41% and 81%) of lettuce grown in the BIODYN2 soil with organic fertilization was recorded compared to the CONMIN soil supplied with mineral fertilizers (Figure 1B). Growth depression was confirmed in two independent experiments in the BIODYN2 soil (Supplementary Table 2) and corresponded with a significant decline in total root length and fine root length (<0.4 mm diameter) (Figure 1B and Supplementary Table 2).
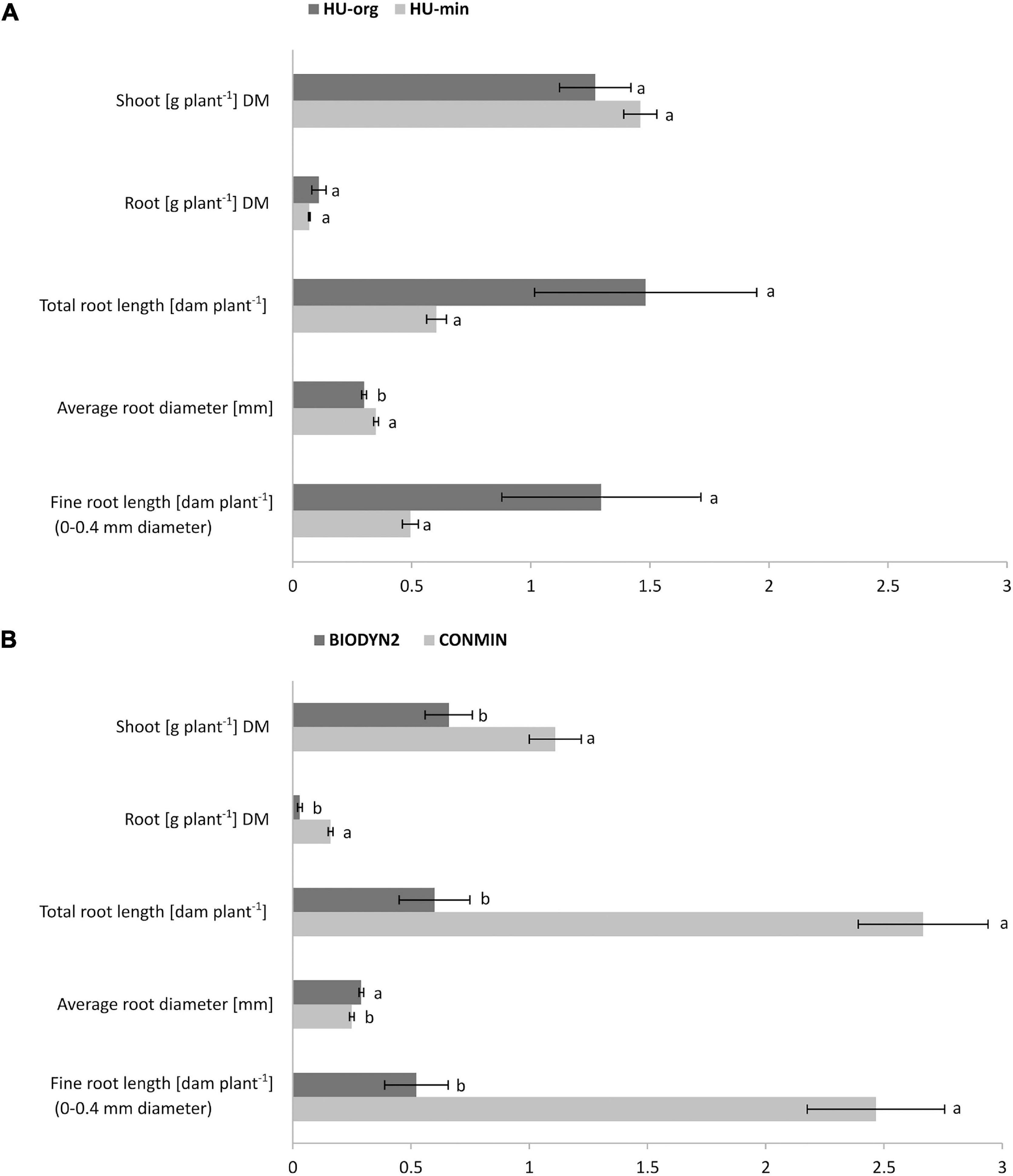
Figure 1. Shoot and root dry biomass (DM) and root morphology of lettuce (cv. Tizian). The plants were grown in minirhizotron culture for nine weeks in soils with long-term organic (HU-org, BIODYN2) or mineral (HU-min, CONMIN) fertilization history. Means ± standard errors of four independent replicates. Different lowercase letters indicate significant differences between organic vs. mineral fertilization tested separately for the sites HUB-LTE (A) and DOK-LTE (B) by one-way ANOVA, Tukey’s HSD pairwise test, p ≤ 0.05.
At the end of the culture period, shoot nutrient concentrations were recorded and deficiencies in nutrient elements such as P and K were identified in all treatments. For K, significant differences related to the fertilization management were observed, showing higher values in the HU-min vs. HU-org soil and higher levels in BIODYN2 compared to CONMIN soil. Other macro- and micronutrients in the shoot tissues, such as Ca, Mg, Mn, and Cu reached the sufficiency range in all treatments. A very moderate limitation was detected for N in all treatments. Iron concentrations were generally high, with particularly high values in DOK-LTE soils (401–842 mg kg–1 dry biomass, Table 2). Significant differences in the nutritional status were more pronounced for lettuce grown in DOK-LTE compared to HUB-LTE soils.
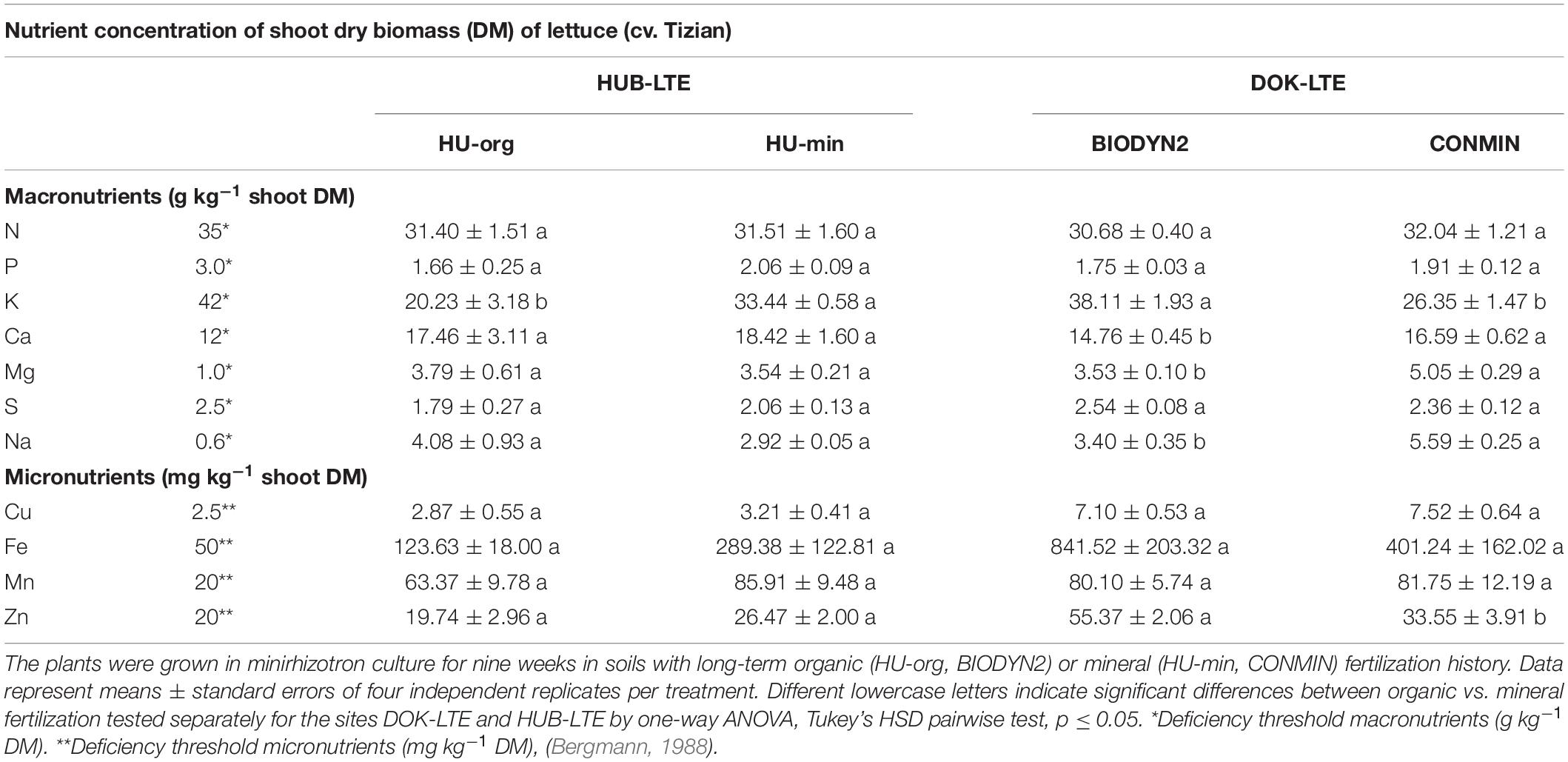
Table 2. Plant nutritional status of lettuce (cv. Tizian).
Expression of Stress-Related Genes in the Shoot Tissue
A qPCR-based method was used to investigate the relative expression of 14 genes, known to be involved in biotic or abiotic stress signaling pathways as previously investigated for lettuce (Chowdhury et al., 2019). Although various genes showed similar expression in plants grown in both soils with long-term organic fertilization (HU-org, BIODYN2) and long-term mineral fertilization (HU-min, CONMIN), certain stress-related genes were significantly upregulated in the treatments with long-term organic fertilization at both field sites. The results showed a significantly enhanced expression of the genes RbohD, PDF1.2, the Fe-transporter OPT3 gene and the nitrate reductase gene NIA1 in shoots of lettuce grown in soils with long-term organic fertilization (HU-org, BIODYN2) in comparison to plants grown in soils with long-term mineral fertilization (HU-min, CONMIN) (Figure 2). In addition, in HUB-LTE soils a significant upregulation of the genes LOX1, WRKY25, and MYB15 in shoots of lettuce grown in organically fertilized soil (HU-org) compared to mineral fertilization (HU-min) was observed.
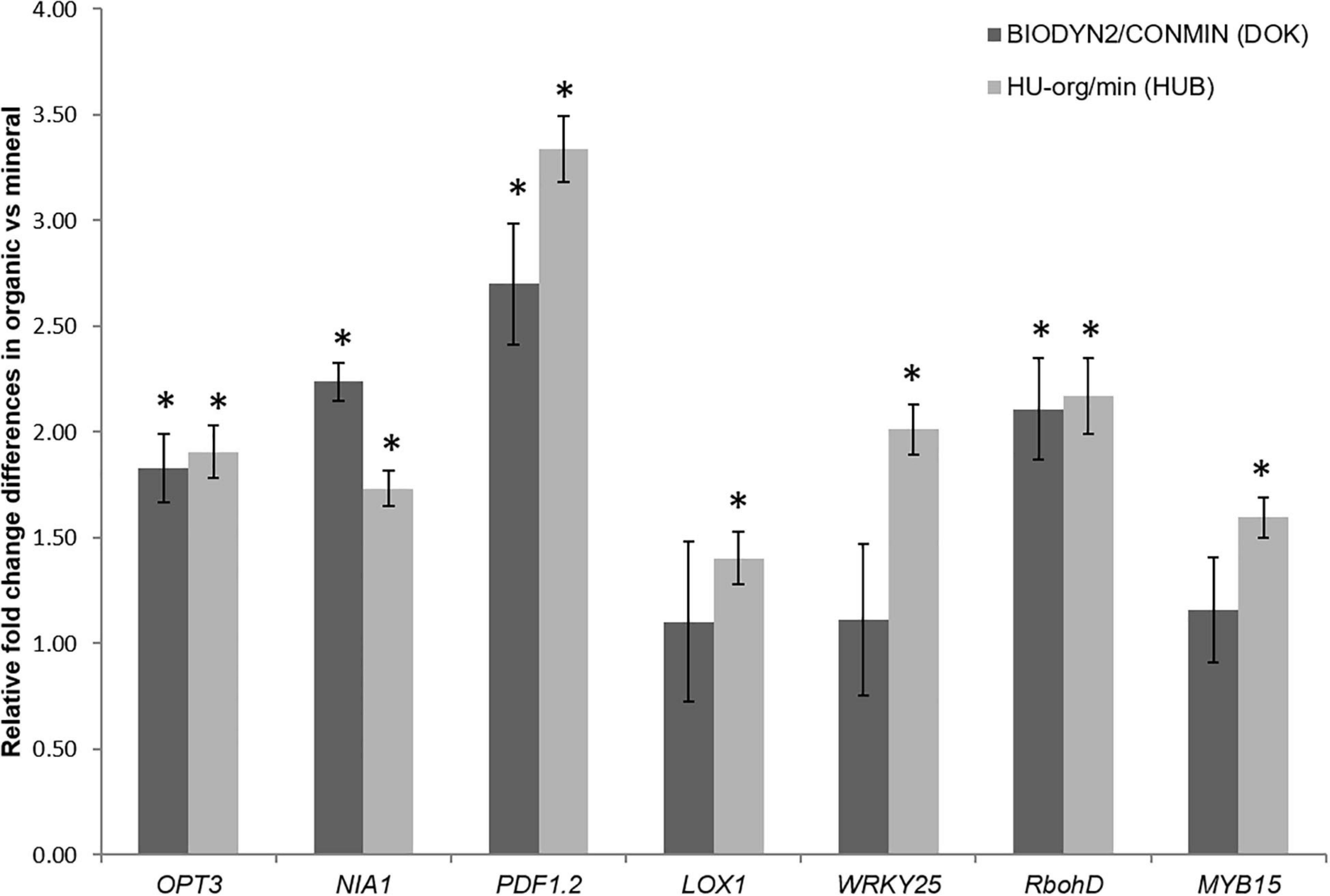
Figure 2. Upregulated genes in shoots of lettuce grown over a period of nine weeks in soils with long-term organic (HU-org, BIODYN) in comparison to mineral (HU-min, CONMIN) fertilization history. The 2–ΔΔCt method (Livak and Schmittgen, 2001) was employed for relative quantification (n = 4) by qPCR (see section “Materials and Methods”). Means ± standard errors of four independent replicates showing relative changes in gene expression of plants grown under long-term organic vs. mineral fertilization. Only genes showing significant (p ≤ 0.05) differences in ΔCt values between organic vs. mineral fertilization within each site as revealed by Tukey’s HSD pairwise test are shown (denoted by asterisks). Gene names, putative functions and primer sequences are described in Supplementary Table 1.
Chemical Composition of the Rhizosphere Soil Solution
Low molecular weight organic compounds in the rhizosphere soil solutions of lettuce grown in the various soil treatments revealed clearly different patterns depending on the fertilization history (Figure 3). The separation of organic compound patterns of organically and minerally fertilized soils were more distinct in DOK-LTE soils compared to HUB-LTE soils. In the different treatments, largely the same organic compounds were detectable with quantitative differences. In the rhizosphere of lettuce grown in BIODYN2 soil but also in BIODYN2 soil without root contact, exceptionally high levels of amino acids were detected (Table 3C and Supplementary Table 3C), leading to separate clustering compared to CONMIN and HUB-LTE soils as shown by PCA analysis (Figure 3). Moreover, higher levels particularly of low molecular weight sugars and malate in the rhizosphere soil solutions collected from lettuce plants grown in HU-min soil explained the separation from HU-org soil (Tables 3A,B and Figure 3).
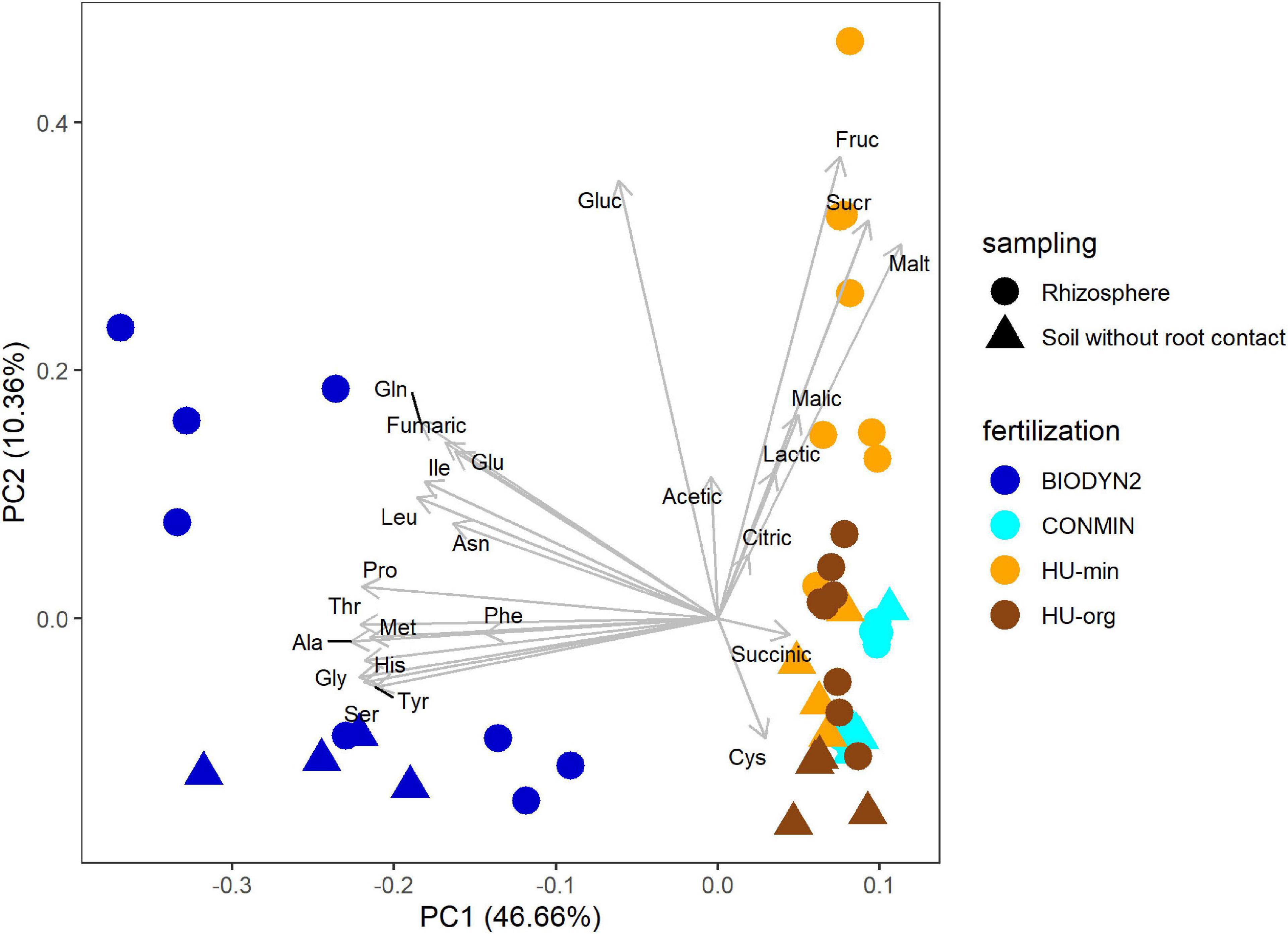
Figure 3. Biplot of PCA comparing patterns of organic compounds in the rhizosphere soil solution of lettuce (cv. Tizian) and in soil without root contact under long-term organic (HU-org, BIODYN2) or mineral (HU-min, CONMIN) fertilization. Ala, alanine; Asn, asparagine; Cys, cysteine; Gln, glutamine; Gly, glycine; Glu, glutamic acid; His, histidine; Ile, iso-leucine; Leu, leucine; Phe, phenylalanine; Pro, proline; Met, methionine; Ser, serine; Thr, threonine; Tyr, tyrosine; Fruc, fructose; Gluc, glucose; Malt, maltose, Sucr, sucrose; Acetic, acetate; Citric, citrate; Fumaric, fumarate; Malic, malate; Succinic, succinate; Lactic, lactate.
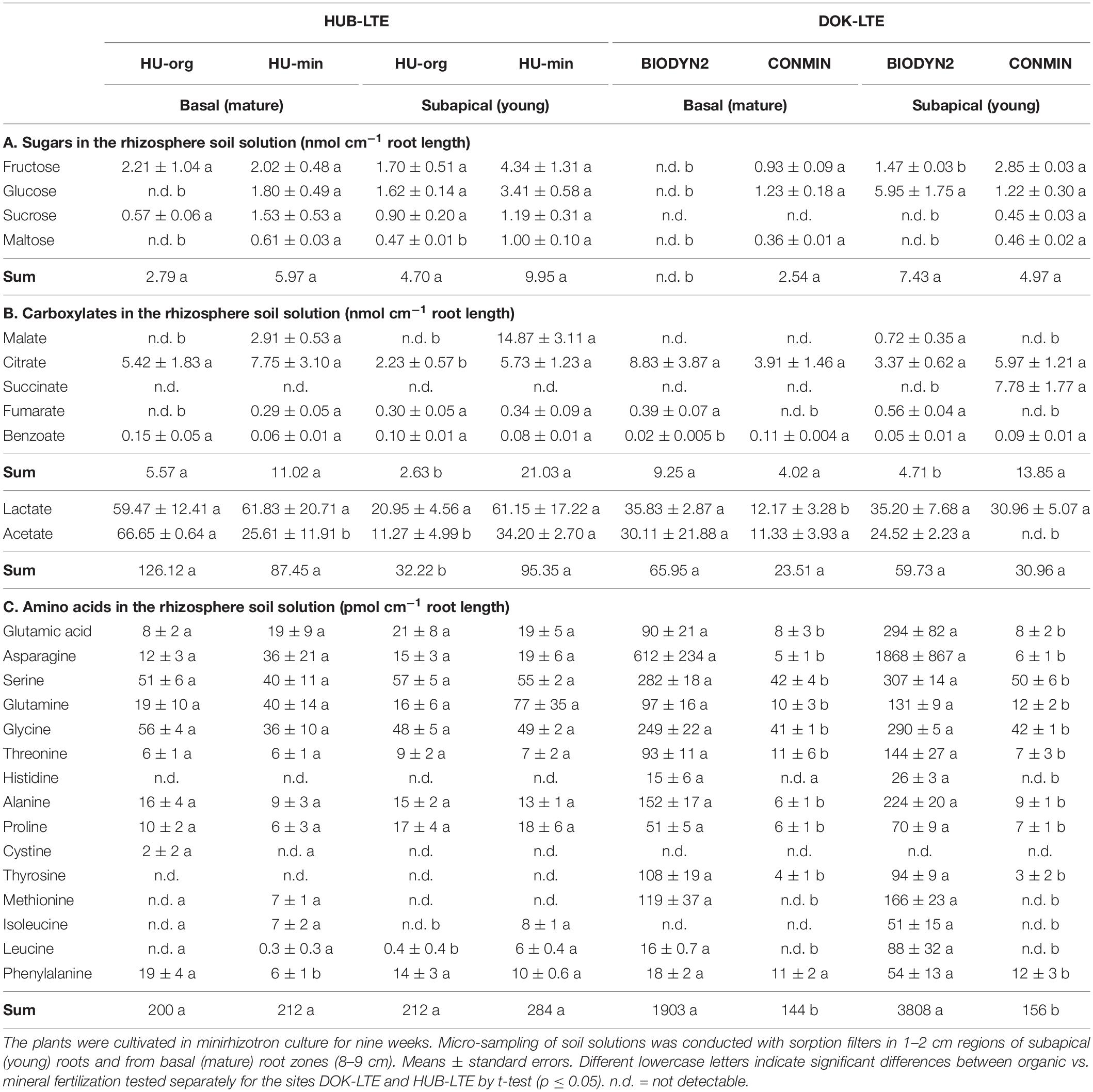
Table 3. Sugars (A), carboxylates (B), and amino acids (C) in the rhizosphere soil solution of lettuce (cv. Tizian), grown in soils with long-term organic (HU-org, BIODYN2) or mineral (HU-min, CONMIN) fertilization history.
The rhizosphere soil solution of lettuce was dominated by sugars, carboxylates, and amino acids (Table 3). Among the various mono- and di-saccharides, particularly hexoses, such as glucose and maltose, were present in significantly lower amounts in samples collected from basal (mature) root zones of lettuce plants grown in soils with long-term organic fertilization history (HU-org, BIODYN2) as compared to soils with mineral fertilization (HU-min, CONMIN). In BIODYN2 soil, all sugars ranged below the detection limit (Table 3A).
Lactate and acetate were the dominant monocarboxylates in the rhizosphere soil solutions (Table 3B) but detectable at higher levels also in samples collected from soil without root contact (Supplementary Table 3B). The concentrations of acetate were increased in the rhizosphere of lettuce grown in soils with organic fertilization history (HU-org, BIODYN2) (Table 3B). Among the di- and tri-carboxylates described also in lettuce tissues (Misaghi and Grogan, 1978), citrate was dominant in the rhizosphere soil solution of all treatments. Dicarboxylates such as malate and succinate dominated in soils with long-term mineral fertilization, with particularly high levels of succinate only in subapical (young) root zones in CONMIN soil (Table 3B). The antifungal compound benzoate was found in lower concentrations in the rhizosphere soil solution of older root zones of lettuce plants grown in the BIODYN2 soil compared with all other soils (Table 3B).
Microbial Community Analyses
Site, Habitat, and Fertilization Effects on Microbial Diversity
Alpha-diversity of bacterial and archaeal communities was lower in the rhizosphere than in root-associated soil, when assessed by Shannon, Pielou and richness indices based on 16S rRNA gene sequencing. This effect was more pronounced in HUB-LTE than in DOK-LTE soils. The long-term organic fertilization in DOK-LTE resulted in a significantly higher bacterial and archaeal diversity (Shannon, richness) in both, root-associated soil and the rhizosphere, as compared to mineral fertilization. Evenness, however, was not affected by fertilization in DOK-LTE. Organic fertilization in HUB-LTE also tended to increase the diversity (Shannon, richness) of bacterial and archaeal communities in the rhizosphere and root-associated soil but the effect was not significant (Table 4). Regarding fungal diversity, significantly lower alpha-diversity indices (Shannon, richness, Pielou) in the rhizosphere than in root-associated soil were observed in DOK-LTE based on ITS2 sequencing in both fertilization regimes. In HUB-LTE, this effect was detected only for fungal richness. HU-org significantly increased the fungal species richness compared to HU-min in the rhizosphere and root-associated soil (Shannon, Pielou). A similar trend was observed in the rhizosphere of DOK-LTE soils. No fertilization-dependent effect was observed on fungal evenness.
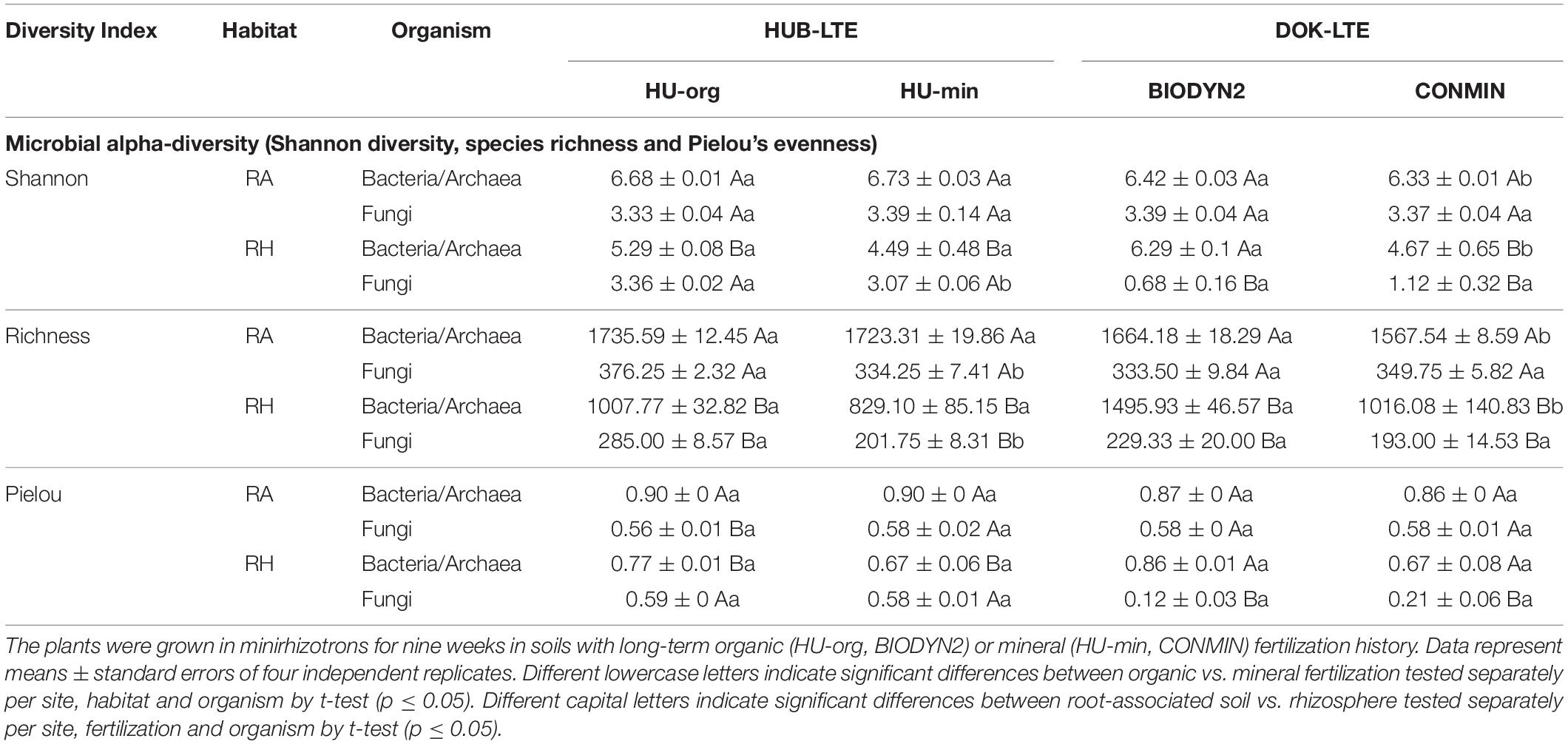
Table 4. Microbial alpha-diversity (Shannon diversity, species richness and Pielou’s evenness) in root-associated soil (RA) and rhizosphere (RH) of lettuce (cv. Tizian).
Microbial Community Composition Affected by Site, Habitat, and Fertilization History
Bacterial and archaeal community composition clearly differed depending on the habitat (R2 = 30%, p < 0.001, PERMANOVA; Figure 4A) and the site (R2 = 22%, p < 0.001). Separate clustering of rhizosphere and root-associated soils was observed for HUB-LTE and DOK-LTE depending on mineral and organic fertilization practice. This finding was confirmed by PERMANOVA analysis which revealed an interaction effect between habitat and site (R2 = 7%, p < 0.001) as well as between habitat and fertilization (R2 = 3%, p ≤ 0.05). A significant influence of the fertilization on the bacterial and archaeal communities was observed (R2 = 6%, p < 0.001). Fertilization-dependent clustering of bacterial and archaeal communities was more distinct in DOK-LTE, especially in the rhizosphere, compared to HUB-LTE (Figure 4A).
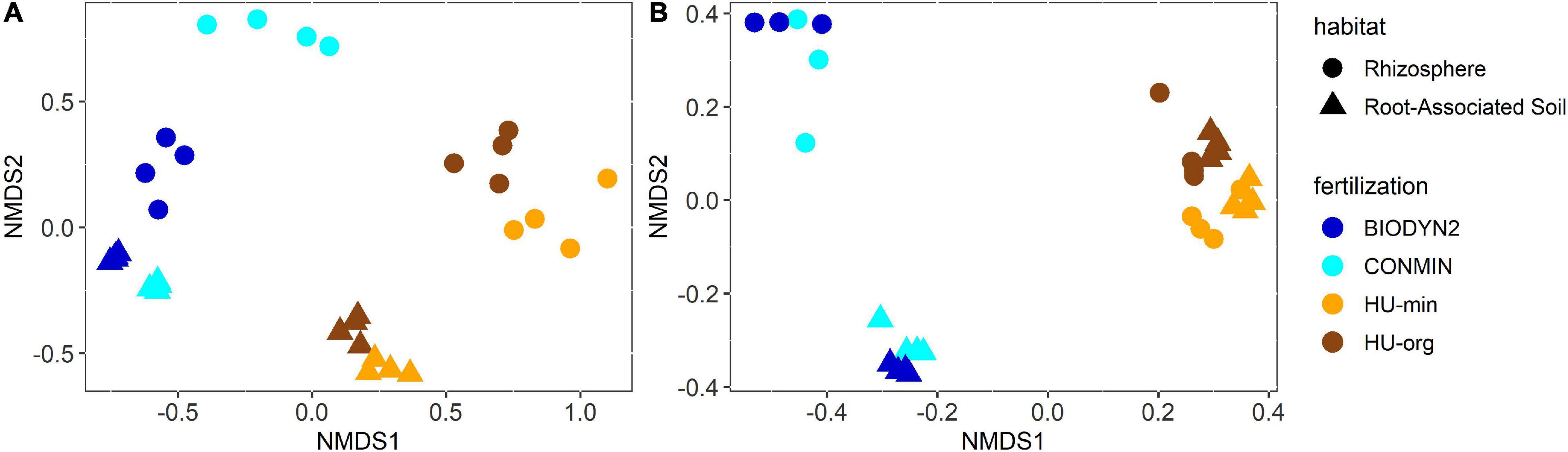
Figure 4. Two-dimensional diagram of non-metric multi-dimensional scaling (NMDS) analysis of calculated Bray-Curtis distances between (A) bacterial and archaeal community compositions or (B) fungal community compositions in root-associated soil and the rhizosphere of lettuce (cv. Tizian) grown in soils from HUB-LTE (HU-org, HU-min) or DOK-LTE (BIODYN2, CONMIN). NMDS analyses are based on relative abundances. Stress = 0.11 (bacterial and archaeal communities) and 0.07 (fungal communities).
The fungal community compositions were clearly separated by site (R2 = 42%, p < 0.001, Figure 4B). Unlike the bacterial and archaeal communities, distinct clustering of rhizosphere and root-associated soils was only observed for DOK-LTE independent of fertilization regimes due to the combined effect of site and habitat (R2 = 19%, p < 0.001). In both LTEs, the impact of fertilization was significant but marginal (R2 = 5%, p < 0.001).
Site, habitat and fertilization effects were also detected in the bacterial and archaeal taxonomic community composition. Thaumarchaeota and Verrucomicrobia had higher relative abundances in DOK-LTE compared to HUB-LTE soils (Supplementary Table 4A). The rhizosphere effect was illustrated by an enrichment in Gamma-, Alpha-, and Betaproteobacteria and by a decrease in relative abundances of Acidobacteria and Firmicutes (Supplementary Table 4A). Heatmap analysis of the most abundant genera in root-associated soils and rhizospheres from HUB-LTE and DOK-LTE showed many taxa occurring at both sites, however most of them exhibited differential relative abundances in rhizosphere and root-associated soils (Figure 5). At both sites, Bacillus and Nitrososphaera were typical genera found in root-associated soils as well as sequences belonging to the family Chitinophagaceae or classified as acidobacterial subdivisions Gp4 and Gp6. Typical rhizosphere responders to lettuce at both sites were affiliated to e.g., Pseudomonas, Rhizobium, and Massilia. When lettuce was grown in HUB-LTE soils, Variovorax, Devosia, and Asticcacaulis were highly abundant in the rhizosphere. In contrast, Duganella and sequences classified as Oxalobacteraceae were present in the rhizosphere of lettuce grown in DOK-LTE soils. Fertilization affected the relative abundance of major rhizosphere genera. For instance, Pseudomonas was highly abundant in the lettuce rhizosphere of CONMIN (up to 61%) and HU-min (up to 74%) and also HU-org (up to 35%). However, a high variability among replicates was observed. In HUB-LTE soils the relative abundance of Rhizobium reached up to 13–17% in the rhizosphere, while the remaining major abundant taxa ranged below 10% (Figure 5B).
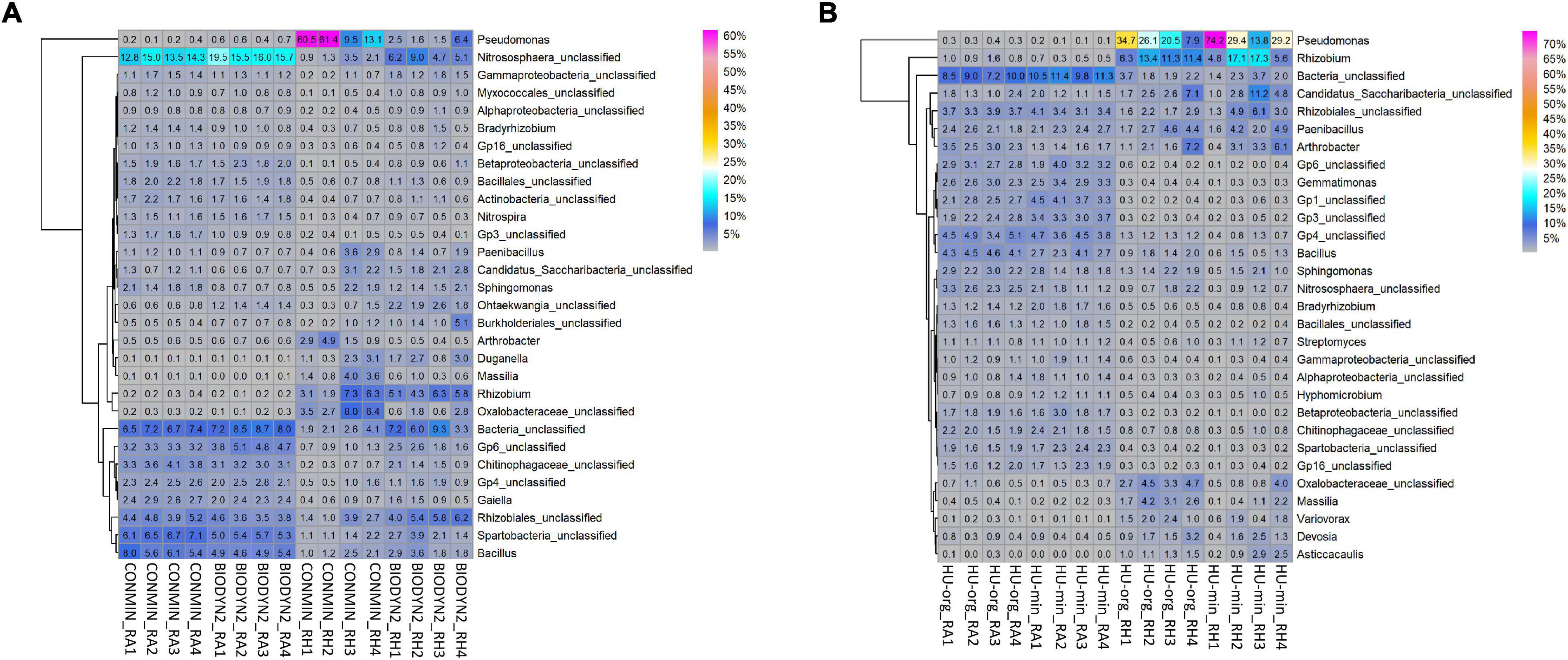
Figure 5. Heatmaps displaying relative abundance distribution of top 30 most abundant bacterial and archaeal genera in root-associated soils (RA) and in the rhizosphere (RH) of lettuce (cv. Tizian) grown in (A) DOK-LTE soils (BIODYN2, CONMIN) and (B) HUB-LTE soils (HU-org, HU-min). The numbers in cells represent relative abundances (%).
The root-associated soils of lettuce grown in HUB-LTE were enriched in the fungal phylum Ascomycota, while Mortierellomycota dominated the soils from DOK-LTE independent of fertilization regimes (Supplementary Table 4B). Mineral fertilization increased Basidiomycota at each site. A strong enrichment in Olpidiomycota characterized the rhizosphere in both treatments of DOK-LTE. The rhizosphere in organic fertilization treatments of HUB-LTE also exhibited an increase of this phylum compared to mineral fertilization. Furthermore, Glomeromycota (arbuscular mycorrhizal fungi, AMF) were enriched in the rhizosphere of organic fertilization treatments of both LTEs.
Heatmap analyses of the most abundant fungal genera in root-associated soil and rhizosphere showed that almost half of the genera could be detected in both LTEs but differing in relative abundances (Figure 6). In general, in DOK-LTE more differences were observed between habitats than between fertilization regimes (Figure 6A). The root-associated soils, especially of BIODYN2, were dominated by Mortierella and unclassified fungi (at genus level) with a strongly reduced relative abundance in the rhizosphere. Higher relative abundances of different yeasts (Solicoccozyma, Exophiala, Apiotrichum, Saitozyma, and Sloofia) as well as AMF Rhizophagus were observed in minerally fertilized soil (CONMIN). Olpidium was the dominant rhizosphere genus in both treatments of DOK-LTE. The differences between the fertilization regimes (organic vs. mineral) were distinct in HUB-LTE (Figure 6B). Typical genera positively affected by organic fertilization independent of habitat were Cercophora, Didymella, and Humicola. Mineral fertilization (HU-min) enriched not only Rhizopus and Umbelopsis but also different yeasts (Exophiala, Solicoccozyma, and Saitozyma), which was in accordance with the DOK-LTE. The rhizosphere of mineral fertilization (HU-min) showed high relative abundances of the genus Mortierella, while the rhizosphere of HU-org exhibited a high relative abundance of Retroconis and of sequences that could not be reliably classified at genus level.
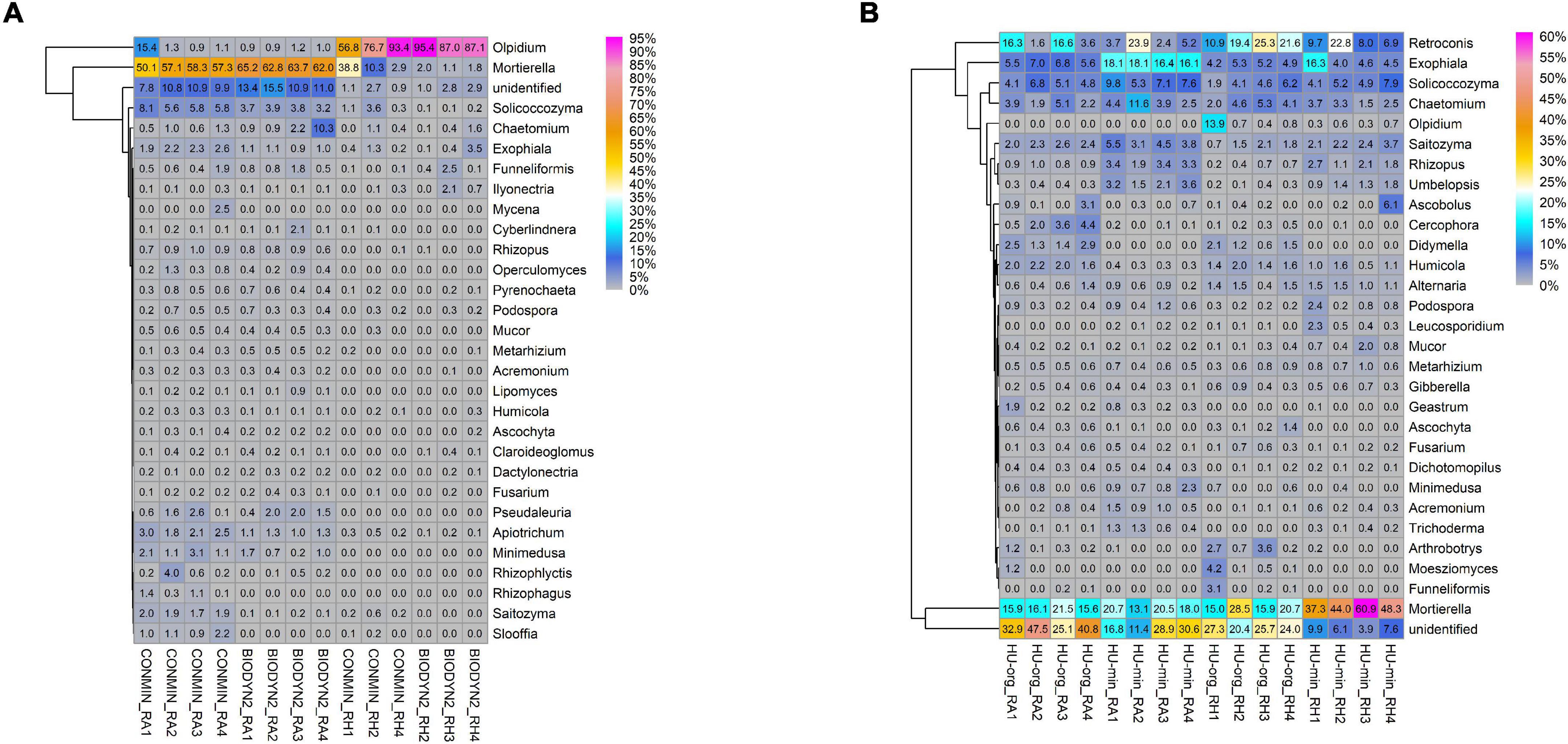
Figure 6. Heatmaps displaying relative abundance distribution of top 30 most abundant fungal genera in root-associated soils (RA) and in the rhizosphere (RH) of lettuce (cv. Tizian) grown in (A) DOK-LTE soils (BIODYN2, CONMIN) and (B) HUB-LTE soils (HU-org, HU-min). The numbers in cells represent relative abundances (%).
Fertilization Effects on Microbial Communities
As we were interested in the effect of fertilization on plant-microbe interactions, we analyzed fertilization-dependent changes in the relative abundance of microbes in the rhizosphere of lettuce. Taxa differing significantly in relative abundance between organic vs. mineral fertilization were determined separately for each site and at several taxonomic levels [OTU (only for fungi), genus, family, order, class, phylum]. More differentially abundant bacterial and archaeal taxa were found in the rhizosphere of lettuce grown in DOK-LTE soils (>1.0% relative abundance; Table 5). Pseudomonadaceae (Gammaproteobacteria) had a significantly higher relative abundance in CONMIN rhizosphere samples compared to BIODYN2 (Table 5B). A similar trend was observed in the rhizosphere of HU-min vs. HU-org, however, differences were not significant. When lettuce was grown in BIODYN2 soil, a significant enrichment in taxa belonging to the Firmicutes phylum (e.g., Clostridiales) was found compared to CONMIN (Table 5B).
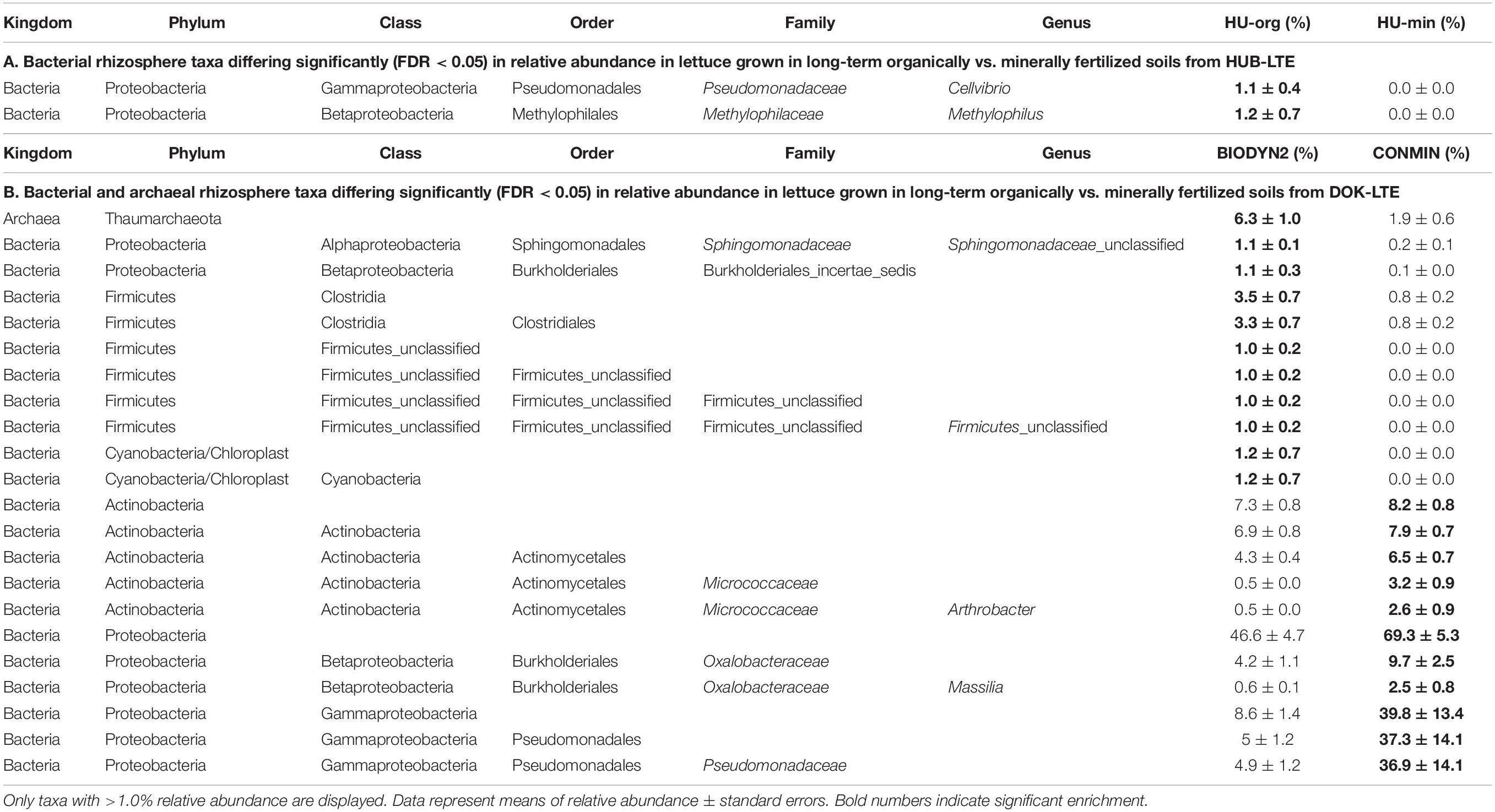
Table 5. Bacterial and archaeal taxa in the rhizosphere of lettuce (cv. Tizian) differing significantly (FDR < 0.05) in relative abundance depending on long-term organic vs. mineral fertilization practice at HUB-LTE (HU-org vs. HU-min) (A) and DOK-LTE (BIODYN2 vs. CONMIN) (B).
In contrast to bacteria and archaea, more differentially abundant fungal taxa were found in the rhizosphere of lettuce grown in HUB-LTE soils compared to DOK-LTE (Table 6). To obtain further insights into the ecological assignment of detected fungal genera, they were assessed against the FUNGuild database for classification into potential pathotrophic, saprotrophic or symbiotrophic fungi (Supplementary Table 5). In concordance with heatmaps (Figure 6), the potential pathotrophic genus Olpidium showed the highest relative abundance in the rhizosphere of lettuce when grown in both DOK-LTE soils and was enriched in the root-associated soil of CONMIN (Supplementary Tables 5, 6B). The rhizosphere of HU-org showed the highest number of significantly enriched pathotrophs including the genera Olpidium, Moesziomyces, and Ascochyta whereas the root-associated soil of HU-min was enriched with pathotrophic-saprotrophic fungi, especially Exophiala. The plant pathogen Rhizopus was increased in the root-associated soil and in the rhizosphere of HU-min, whereas Didymella was increased in the root-associated soil and in the rhizosphere of HU-org. The saprotrophic genera Cercophora and Humicola as well as Arthrobotrys and Plenodomus were highly abundant in the root-associated soil and in the rhizosphere of HU-org, respectively. Umbelopsis was enriched in both habitats of HU-min (Table 6A and Supplementary Table 6A). Significantly more sequences classified as Mortierella, a saprotrophic-symbiotrophic genus, were found in the rhizosphere of lettuce grown in minerally fertilized soils (HUB-LTE and DOK-LTE) in comparison with organic fertilization. Mycorrhizal symbiotrophs (Clariodeoglomus, Funneliformis) were enriched in both habitats of lettuce grown in HU-org whereas the relative abundance of Trichoderma increased in root-associated soil of HU-min (Supplementary Table 5). Further results for fungal taxa in the root-associated soils are shown in Supplementary Table 6.
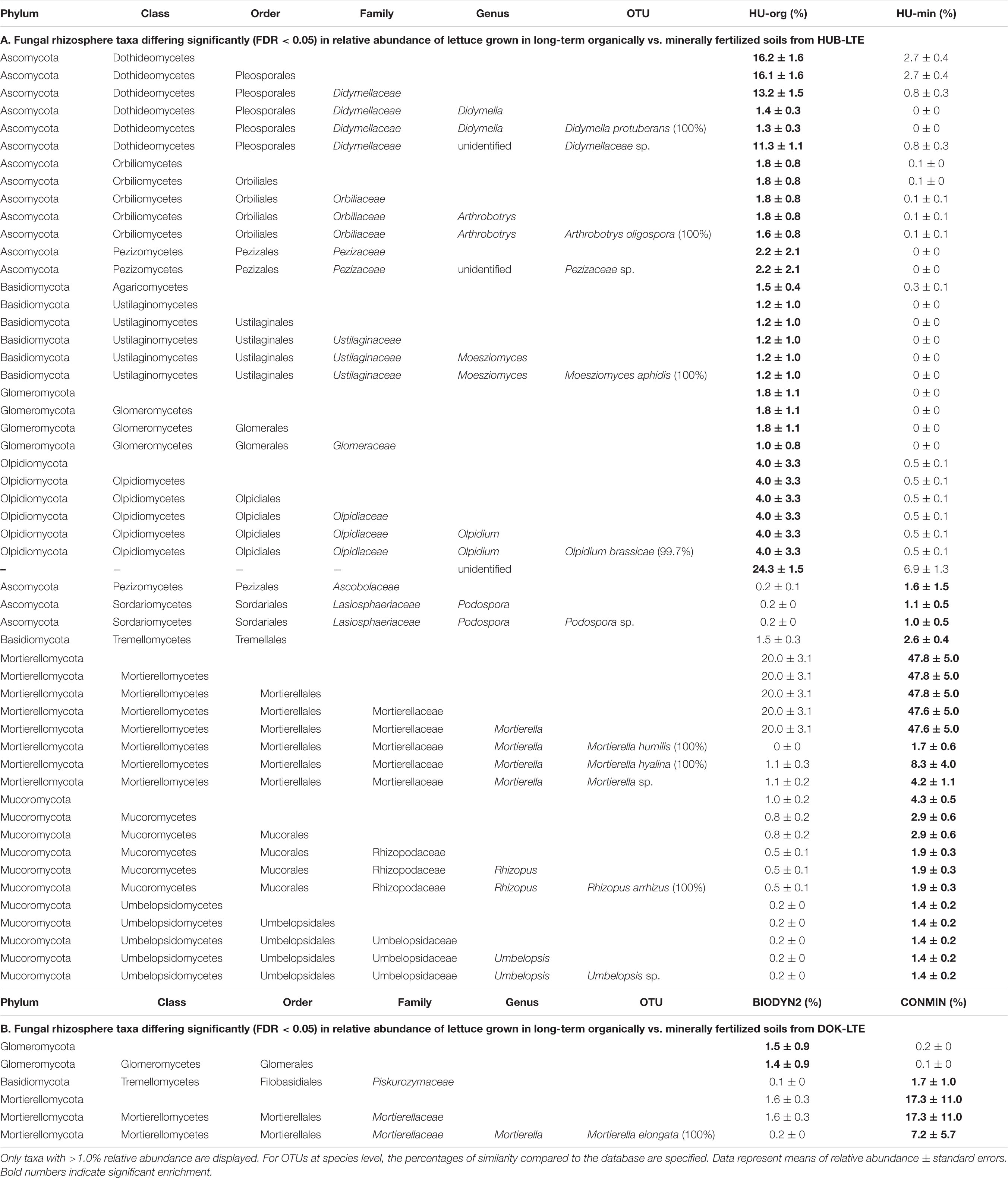
Table 6. Relative abundance of fungal taxa in the rhizosphere of lettuce (cv. Tizian) differing significantly (FDR < 0.05) in relative abundance depending on long-term organic vs. mineral fertilization practice at HUB-LTE (HU-org vs. HU-min) (A) and DOK-LTE (BIODYN2 vs. CONMIN) (B).
Canonical correspondence analyses revealed relationships in rhizosphere bacterial and archaeal and fungal community composition with organic compounds detected in the rhizosphere soil solution (Figure 7). Ordination showed that the fertilization-dependent differentiation in rhizosphere bacterial and archaeal communities of lettuce grown in DOK-LTE soils was clearly related to the concentration of succinate (CONMIN) or fumarate and amino acids (BIODYN2). HUB soils were positively associated with sugars (sucrose, maltose) which caused the separate clustering of rhizosphere bacterial and archaeal communities apart from DOK-LTE soil samples. Fertilization-dependent community differences in HUB-LTE soils were less clear as compared to DOK-LTE soils but a significant contribution of malate to the differentiation of HU-min was identified. The relative abundances of Pseudomonadaceae OTUs responding positively to mineral fertilization were negatively correlated with fumaric and amino acids (Figure 7A).
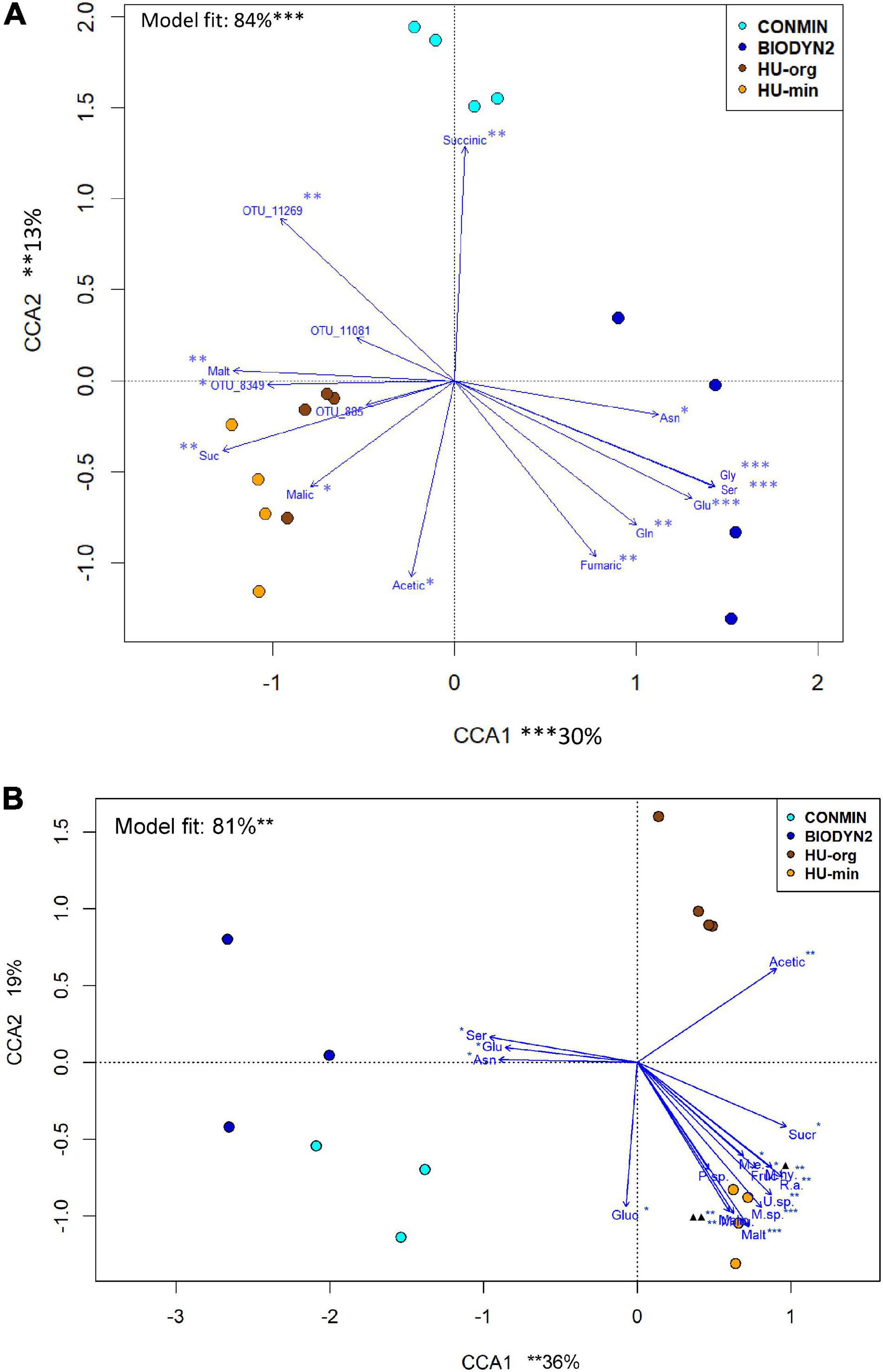
Figure 7. Canonical correspondence analysis (CCA) of bacterial and archaeal (A) and fungal (B) community composition in the rhizosphere of lettuce grown in soils with long-term organic (HU-org, BIODYN2) or mineral (HU-min, CONMIN) fertilization. CCA is based on log10 transformed relative abundances of bacterial, archaeal and fungal OTUs using organic compounds in rhizosphere soil solution averaged over basal (mature) and subapical root zones. (A) Bacterial and archaeal communities: Acetate (Acetic) + malate (Malic) + succinate (Succinic) + fumarate (Fumaric) + maltose (Malt) + sucrose (Suc) + asparagine (Asn) + serine (Ser) + glutamine (Gln) + glutamic acid (Glu) + glycine (Gly) as constraint variables. Significant vectors were fitted onto CCA ordination including log transformed relative abundances of Pseudomonadaceae OTUs that were significantly enriched (FDR < 0.05) in minerally fertilized soils and exhibited a mean relative abundance of >0.5%. Tentative taxonomic identification of OTUs based on most similar BLAST hit: OTU_11269 – Pseudomonas corrugata (99.77%); OTU_11081 – P. silesiensis (96.97%); OTU_8349 – P. corrugata (99.3%); OTU_885 – P. tolaasii (99.77%). Significant codes: ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001. (B) Fungal communities: Acetate (Acetic) + malate (Malic) + fructose (Fruc) + glucose (Gluc) + sucrose (Sucr) + maltose (Malt) + glutamic acid (Glu) + asparagine (Asn) + serine (Ser) as constraint variables. Significant vectors were fitted onto CCA ordination including log transformed relative abundances of OTUs (identified at least at genus level) that were significantly enriched (FDR < 0.05) in minerally fertilized soils and exhibited a mean relative abundance of >1.0% (M.e. – Mortierella elongata; R.a. – Rhizopus arrhizus; M.sp. – Mortierella sp.; P.sp. − Podospora sp.; U.sp. – Umbelopsis sp.; M.hy. – Mortierella hyalina; M.hu. – Mortierella humilis). Overlapping designations: ▲ Fruc∗/M.e.∗/M.hy.∗∗, ▲▲ Malic∗∗/M.hu.∗∗. Significant codes: ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001.
Fertilization-dependent differences of fungal rhizosphere communities were more pronounced in HUB-LTE compared to DOK-LTE (Figure 7B). The clear separation between organic vs. mineral fertilization (HUB-LTE) was caused by the positive correlation of several compounds in HU-min compared to only one in HU-org (acetate). Fungal communities of lettuce grown in HU-min soils showed a clear association to sugars (sucrose, maltose, and fructose) and malate, which was partly comparable with the bacterial and archaeal communities. The identified responder OTUs (Rhizopus arrhizus, Mortierella sp., Umbelopsis sp., Mortierella hyalina, Mortierella humilis, and Nigrospora oryzae, Table 6) reacted positively to mineral fertilization. Regarding all four treatments, M. elongata responded also positively to mineral fertilization in HUB-LTE, although this OTU was significantly enriched in CONMIN compared to BIODYN2 (Table 6B). Comparable with bacterial and archaeal communities, fungal rhizosphere communities of lettuce grown in DOK soils, especially BIODYN2, were clearly associated with amino acids.
Discussion
Plant roots are described as powerful drivers of microbiota assemblage (Bais et al., 2006; Bakker et al., 2014). However, to which extent site-specific factors or fertilization management interact with the recruitment of rhizosphere microbiota remains largely unclear. In a holistic approach, we tried to correlate the structure of rhizosphere microbial communities with data on organic composition of the rhizosphere soil solution, root growth characteristics and aboveground plant traits (biomass, nutrient status and expression of stress-related genes). Two strategies of fertilization management were exemplarily compared by growing lettuce plants in minirhizotrons with contrasting soils from two LTEs with organic and mineral fertilization history.
Site- and Fertilization-Dependent Plant Performance
No consistent effects of long-term fertilization practice on aboveground plant biomass and root characteristics (total and fine root length, average diameter, and length of root hairs) were detected in this study (Figure 1), as similarly reported in an earlier lettuce experiment with the same soils (Chowdhury et al., 2019). The latter authors reported similar growth in DOK-LTE soils and lower plant biomass in organically fertilized soil of the HUB-LTE (Chowdhury et al., 2019). In our experiment, the results were opposite, with similar biomass in soils of the HUB-LTE and shoot and root biomass drastically reduced by 41% and 81%, respectively, in the organically fertilized soil (BIODYN2) of the DOK-LTE. This effect could not be attributed to nutrient limitations. Although at the end of the experiment, concentrations of mineral nutrients (N, P, K) in shoot tissues were below or close to the reported deficiency thresholds (Bergmann, 1988), these effects were observed similarly for all soil treatments and not only for the BIODYN2 soil (Table 2). Similar plant growth reductions were observed in a repeated experiment with the same soils, already detectable during early plant establishment (Supplementary Table 2). This suggests the presence of additional stress factors in BIODYN2 soil independent of nutrient limitations, as a site-specific effect. The discrepancy between our findings and those of Chowdhury et al. (2019) may have resulted from the collection of the soils in different years with different pre-crops. Effects of the pre-crop on soil microbial communities have been reported previously (Sommermann et al., 2018; Babin et al., 2019) and the detritusphere microbiome of pre-crop roots can even overwrite the rhizosphere effect of the current crop (Zhou et al., 2020) with potential consequences for plant performance. Furthermore, in the study of Chowdhury et al. (2019) lettuce plants were cultivated in pots, whereas our study was conducted with minirhizotrons promoting the development of high rooting densities along the root observation windows. This may lead to locally increased exudate concentrations contributing to attraction of beneficial but also of pathogenic microorganisms with potential impact on plant performance.
Soil Microbial Communities Affected by Fertilization, Site, and Habitat Effects
The LTE site and the habitats, comprising rhizosphere and root-associated soil, distinguished bacterial, archaeal, and fungal community composition as recently reported by Chowdhury et al. (2019). A significantly higher bacterial and archaeal alpha-diversity was mainly found in the rhizosphere of lettuce when grown in organically vs. minerally fertilized soils of DOK-LTE but not of HUB-LTE (Table 4). Conversely, the alpha-diversity of the fungal rhizosphere community was significantly increased in organically fertilized soil of HUB-LTE, but not of DOK-LTE, illustrating the impact of the soil type as a major driver determining not only the soil but also the rhizosphere microbial composition (Schreiter et al., 2014b; Chowdhury et al., 2019). The selective effect of the plant on recruitment of microbial communities was apparent by a lower alpha-diversity in the rhizosphere compared to root-associated soil (Table 4) as reported in various other studies (Mendes et al., 2013; Chowdhury et al., 2019).
The majority of bacterial and archaeal rhizosphere responders were classified as Gammaproteobacteria of the genus Pseudomonas with particularly high relative abundance (up to 60–74%) in soils with mineral fertilization history, while only marginal rhizosphere enrichment (rel. abundance 2–6%) was detectable in the BIODYN2 rhizosphere with long-term organic fertilization (Figure 5). Many members of this genus exert beneficial effects on plants (Roquigny et al., 2017), among them Pseudomonas putida, Pseudomonas fluorescens, and Pseudomonas brassicacearum, although pathogens are also reported. Since we observed the lowest biomass production along with impaired fine root development in lettuce plants grown in the BIODYN2 soil (Figure 1B), this may indicate a relationship between the low relative abundance of potentially beneficial Pseudomonadaceae and plant growth, because lettuce growth was not impaired in the soils with a particularly high Pseudomonas relative abundance (Figures 1, 5). A preliminary identification showed high similarities with members of the P. fluorescens complex with plant growth-promoting properties (Garrido-Sanz et al., 2016). However, strain identification based on short-read Illumina 16S rRNA gene sequences is limited. Future studies should therefore consider cultivation-based approaches for comprehensive taxonomic and functional characterization.
With regard to fungal communities, the most remarkable rhizosphere effect was the enrichment of the pathogenic genus Olpidium in the rhizosphere of soils with organic fertilization history (HU-org, BIODYN2), which was particularly pronounced for DOK-LTE, where its relative abundance reached 76-90% (Figure 6 and Supplementary Table 4B). Lettuce is a host for Olpidium sp. (Olpidiomycota, formerly classified as Chytridiomycota). Zoospores of the fungus with random motility rapidly invade the tips of fine roots (Maccarone, 2013). Since these zoospores are not specifically attracted by root exudates (Westerlund et al., 1978), direct relationships with the observed site- and fertilization-dependent differences in the composition of the rhizosphere soil solution (Figure 3 and Table 3) are unlikely. The pathogen can survive in soils as dormant spores for up to 20 years (Campbell, 1985). Early plant infection can result in severe growth inhibition (Navarro et al., 2004). Accordingly, in our study, growth retardation and inhibition of root hair development in the BIODYN2 soil (Figure 1B and Supplementary Table 2) was associated with typical symptoms of Olpidium infection. Root infection with intracellular formation of sporangia and resting spores in fine roots (Supplementary Figure 1) was consistent with the highest relative abundance of Olpidium (90%) in the BIODYN2 rhizosphere (Figure 6 and Supplementary Table 5). Interestingly, in the CONMIN soil, symptoms caused by Olpidium were less pronounced despite similarly high relative Olpidium abundance in the rhizosphere of both treatments CONMIN (76%) and BIODYN2 soil (90%) (Supplementary Table 5). Zoospores exhibit high motility in wet clay soils to initiate infections in nearby plants (Westerlund et al., 1978). This may explain the low relative abundance of Olpidium in the sandy HUB-LTE soils (0.5–4%) and increased infection rates in the silty loam DOK-LTE soils with a higher water retention capacity as a site-specific effect. Nevertheless, this cannot explain the high prevalence of disease symptoms in plants from BIODYN2 soil (Supplementary Figure 1) with organic fertilization compared to CONMIN soil supplied with mineral fertilizers despite similar relative Olpidium abundance in the rhizosphere (Supplementary Table 5 and Figure 6).
Along with the rhizosphere enrichment in Olpidium, a particularly low relative abundance of the fungal genus Mortierella (1.6%) was found for the BIODYN2 rhizosphere, reaching 17-48% in the other investigated treatments (Figure 6 and Supplementary Table 5). Fungal endophytes of the genus Mortierella are classified as saprotrophic symbiotrophs (Supplementary Table 5). Most recently, plant growth-promoting properties and biocontrol activity against fungal pathogens via jasmonic acid-dependent plant stress signaling were reported for various Mortierella species (Kamzolova et al., 2014; Tamayo-Velez and Osorio, 2017; Li et al., 2018) including Mortierella elongata identified in our study (Table 6B). Therefore, we suggest that higher sensitivity toward Olpidium infection of lettuce plants grown in the BIODYN2 soil may be a consequence of a low relative abundance of bacterial (Pseudomonas) and fungal (Mortierella) antagonists or plant growth promoters in the rhizosphere. In contrast, a selective pathogen-suppressive effect of long-term pesticide application in the CONMIN soil (Table 1) seems to be unlikely in this case, since the relative Olpidium abundance was even higher in the root-associated CONMIN soil compared to BIODYN2 soil (Supplementary Table 5).
Apart from rhizosphere enrichment of potential plant pathotrophs and saprotrophs, AMF (Funneliformis and Claroideoglomus) as plant beneficial symbiothrophs, increased in organically fertilized soils (BIODYN2 and HU-org). A higher extent of AMF root colonization and increased AMF species diversity in organic farming soils has been frequently reported (Sattelmacher et al., 1991; Douds et al., 1997; Gosling et al., 2006) as a consequence of lower P availability (Nagahashi et al., 1996; Olsson et al., 2003) or reduced fertilizer and pesticide inputs. Accordingly, the lowest P-nutritional status was recorded for lettuce plants grown in BIODYN2 and HU-org soils (Table 2).
Components of the Rhizosphere Soil Solution Related to Rhizosphere Microbiota
In our study, significantly lower sugar concentrations (particularly glucose) were recorded in the rhizosphere soil solution of mature roots of lettuce plants grown in soils with organic compared to mineral fertilization history (Table 3A). Host plants provide up to 20% of photo-assimilates to mature arbuscular-mycorrhizal root systems, consumed preferentially in form of glucose by AMF. Consequently, this is frequently associated with lower sugar exudation into the rhizosphere (Jones et al., 2004). Accordingly, also in our study, lower glucose concentrations in the lettuce rhizosphere in soils with long-term organic fertilization coincided with a higher rhizosphere relative abundance of AMF (Table 6 and Supplementary Table 4B). Interestingly, in lettuce plants grown in BIODYN2 soil, with the highest root infection rate by the Olpidium pathogen (Figure 6 and Supplementary Table 5), sugar concentrations ranged close to or even below the detection limit in the rhizosphere of mature root zones (Table 3A). This is most likely caused by the high carbon demand and sugar consumption of the obligate biotrophic pathogen. On the other hand, some reports show antagonistic effects of AMF colonization against Olpidium (Hao et al., 2019) but it remains questionable, whether this applies for competitive root colonization of Olpidium and AMF in the establishment phase during early growth of the host plants analyzed in our study. The extremely limited availability of glucose in the rhizosphere of the BIODYN2 plants (Table 3A) may also explain the particularly low relative rhizosphere abundance of M. elongata (Table 6B) with potential antagonistic properties (Li et al., 2018), since glucose supply was identified as a major carbon source for stimulation of Mortierella mycelium growth (Chesters and Peberdy, 1965). Noteworthy, also antagonistic effects between root colonization with AMF and Mortierella sp. were recently reported in avocado nurseries (Tamayo-Velez and Osorio, 2017). This might at least partially explain the higher Mortierella rhizosphere relative abundance in the soils with long-term mineral fertilization history (Table 6 and Supplementary Table 4B).
Windisch et al. (2017) reported the ability of bacterial biocontrol inoculants (Pseudomonas sp. RU47; Serratia plymuthica 3Re-4-18) to induce root exudation of benzoate in lettuce, as a secondary metabolite with antifungal activity (Yoon et al., 2012). This was related to increased tolerance against bottom rot disease caused by Rhizoctonia solani. Surprisingly, in our study, the significantly reduced rhizosphere relative abundance of Pseudomonaceae in lettuce plants grown in the BIODYN2 soil in comparison with the CONMIN soil (Table 5 and Figure 5) was associated with Olpidium-related growth depressions and significantly reduced benzoate concentrations in the rhizosphere soil solution (Table 3B). Therefore, a low rhizosphere presence of bacterial communities with antagonistic potential in the BIODYN2 soil (e.g., Pseudomonas) may be responsible for the limited stimulation of benzoate exudation, thereby increasing the plant sensitivity to pathogen attack. Moreover, benzoate also has chemo-attractive properties for various Pseudomonas species (reviewed by Sampedro et al., 2015).
In terms of di- and tri-carboxylates, characterized as intracellular compounds in lettuce (Misaghi and Grogan, 1978), high concentrations of succinate were detected exclusively in the rhizosphere soil solution of lettuce grown in the CONMIN soil (Table 3B) and related to a high relative abundance of Pseudomonadaceae (Figure 7A). Among other dicarboxylates, such as malate and fumarate, also detected in the rhizosphere soil solutions of the investigated lettuce plants particularly in combination with mineral fertilization (Table 3B), succinate has been characterized as a potent chemoattractant with importance for root colonization by various Pseudomonas strains with plant growth-promoting properties (Oku et al., 2014; reviewed by Sampedro et al., 2015). Moreover, succinate is the major carbon source required for production of pyoverdine siderophores by fluorescent pseudomonads (Marek-Kozaczuk and Skorupska, 1997) with biocontrol functions via iron competition with pathogens (Höfte et al., 1991) and induction of systemic plant defense responses (Audenaert et al., 2002; Adesina et al., 2007). Therefore, we speculate that a potential relationship between the enrichment of beneficial Pseudomonadaceae and high succinate concentrations in the CONMIN rhizosphere counteracted Olpidium pathogenesis.
Additionally, reduced sugar concentrations in connection with extremely high amino acid concentrations resulted in a lower C/N ratio in the rhizosphere soil solution of plants grown in the BIODYN2 soil, as compared with all other soil treatments (Table 3). Gammaproteobacteria are generally characterized as fast-growing, copiotrophic microorganisms, which prefer carbon-rich environments and hence a preference for high C/N substrate ratios has been described for Gammaproteobacteria (Kuramae et al., 2012; Michaud et al., 2014; Shen et al., 2016). Consequently, the relatively high N and low C availability in the rhizosphere of lettuce grown in BIODYN2 soil may have counteracted an enrichment of Gammaproteobacteria (i.e., Pseudomonadaceae) with potentially antagonistic properties in the rhizosphere. Accordingly, amino acid accumulation in the rhizosphere was negatively correlated with rhizosphere relative abundance of Pseudomonadaceae (Figure 7A). Similarly high amino acid concentrations were recorded also in samples collected from BIODYN2 soil without root contact (Supplementary Table 3C). A possible explanation for this phenomenon is the high organic N content potentially affected by long-term input of composted farmyard manure in BIODYN2 soil with the highest Corg and total N contents among all other investigated soils (Table 1B). Thus, a high mineralization potential and elevated levels of free amino acids as intermediate products may have led to the observed increase of amino acid concentrations in the soil solution largely overwriting the rhizosphere effect. Nevertheless, a wide range of amino acids has been reported as chemoattractants for Pseudomonas strains (reviewed by Sampedro et al., 2015) but due to the absence of a rhizosphere effect for amino acids in the BIODYN2 soil, no chemoattraction can be expected in this case. Similarly, high concentrations of acetate and lactate were detectable in the rhizosphere and in soil without root contact (Table 3B and Supplementary Table 3B), suggesting that these compounds are rather products of organic carbon turnover than directly released from plant roots.
Rhizosphere Microbiota and Stress-Related Gene Expression of the Host Plant
The rhizosphere of lettuce grown in HU-org and BIODYN2 was enriched in fungal pathotrophs and patho-saprotrophs (Supplementary Table 5). This could be a possible reason for a significantly higher expression of jasmonic acid signaling-dependent genes (PDF1.2) and lipoxigenase (LOX) genes in the shoot tissue of plants grown in soils with organic vs. mineral fertilization, as previously reported also by Chowdhury et al. (2019). Recent findings suggest that the rhizosphere microbiota act as an additional immune barrier for plants and can activate long distance signaling pathways, involving ethylene, salicylic and jasmonic acid, which led to systemic resistance in distal tissues (Spoel and Dong, 2012; Hacquard et al., 2017). The RbohD gene, encodes NADPH oxidase (respiratory burst oxidase), involved in free radical (ROS) production for pathogen defense after contact with pathogen-associated molecular patterns (PAMPs) and has also been reported as systemic response to elevated pathogen abundance in the root/rhizosphere (Miura and Tada, 2014; Kadota et al., 2015). Our results showed that this gene was upregulated in both soils with organic fertilization (BIODYN2, HU-org) including high relative abundances of potential pathotrophs (Supplementary Table 5).
Redistribution of iron upon pathogen attack is another defense response in various plant species, leading to accumulation of Fe3+ in the apoplast of epidermal cells, where it induces the production of H2O2, resulting in a defensive oxidative burst (Verbon et al., 2017). This can induce local iron deficiencies in the cytosol (Liu et al., 2007) and may explain the preferential upregulation of the OPT3 gene in lettuce plants grown in organic HU-org and BIODYN2 soils (Figure 2). The gene encodes an oligopeptide transporter involved in phloem-mediated Fe transport, induced under conditions of Fe deficiency (Stacey et al., 2008). Moreover, pathogen infection can induce systemic activation of genes involved in root-induced Fe acquisition (FRO2, IRT1; Verbon et al., 2017) and may offer an explanation for the particularly high shoot Fe concentration in plants, grown in DOK-LTE soils (Table 2), associated with higher pathogen (Olpidium) enrichment in the rhizosphere compared to HUB-LTE soils (Supplementary Table 4B). Interestingly, the expression of the nitrate reductase gene (NIA1) was significantly increased in lettuce plants in soils with long-term organic fertilization history. This may be attributed to a higher N mineralization potential because of long-term manure-based fertilization in BIODYN2 and HU-org soils. However, in the experimental setup, all treatments received full N supply via nitrate fertilization, sufficient for the duration of our experiment, without any treatment differences of the N nutritional status (Table 2). An alternative explanation may be provided by the results of Vujinović et al. (2020), demonstrating superior induction of nitrate reductase genes in maize by humic substances isolated from soils with long-term organic fertilization as compared with conventional mineral fertilization.
Plant defense mechanisms are complex and in soil culture where the roots are in continuous contact with diverse microbes, it is difficult to assign a direct systemic effect of particular microbial taxa to changes in the gene expression in the shoots. The generally higher expression of stress-related genes recorded in the shoots of organically grown plants are interesting indicators for a systemic defense response. However, we were not able to detect any specific response in stress gene expression to the particularly intense infection with the root pathogen Olpidium, affecting plants grown in BIODYN2 soil. Severe growth depressions and infection symptoms suggest that the upregulation of the respective defense genes alone was not sufficient to counteract the pathogen attack. Certain pathogens are able to overcome these primary defense lines via specific elicitors with potential to inhibit signaling pathways or the synthesis and accumulation of defense compounds by the host plant (Henry et al., 2012; Fitzpatrick et al., 2020). This scenario may be reflected by the reduced accumulation of the antifungal root exudate benzoate in the rhizosphere of the Olpidium-infected plants grown in BIODYN2 soil (Table 3B). Comparison with plants in CONMIN soil, lacking disease symptoms despite a similar relative rhizosphere abundance of the Olpidium pathogen and even lower systemic expression of defense genes, suggest that additional factors are required for pathogen suppression. These factors may comprise the recruitment of beneficial microorganisms (e.g., Pseudomonas sp., Mortierella elongata) via specific root exudate patterns (i.e., increased levels of succinate, malate, glucose), indicated by a reduction in stress gene expression as part of a “cry for help” strategy employed by the host plant (Fitzpatrick et al., 2020; Liu et al., 2020). Additionally, the increased production of root exudates with antifungal potential (i.e., benzoate) was stimulated by the presence of beneficial microbes (Windisch et al., 2017). A chronological investigation of the plant responses to pathogen infection and a more comprehensive characterization of metabolic stress indicators could reveal more evidence in this concern.
Conclusion
The present study confirmed the well-documented selective effects of the soil site and the rhizosphere as important determinants shaping the diversity and composition of soil microbial communities. However, with respect to the impact of fertilization management, frequently reported benefits of long-term organic fertilization, such as increased soil organic matter, increased microbial biomass and metabolic activity, species richness and higher abundance of plant beneficial microorganisms were only partially confirmed. A significant increase of soil organic matter was only detectable for BIODYN2 soil. The rhizosphere microbial diversity under long-term organic fertilization was not consistently increased, for both bacterial/archaeal and fungal communities. Beneficial effects on the relative abundance of plant growth-promoting microorganisms were only detectable for AMF but not generally for potential plant growth-promoting microorganisms (Pseudomonas, Mortierella), and were sometimes (BIODYN2) even associated with severe growth depressions likely caused by the lettuce pathogen Olpidium. A consistently increased systemic expression of stress-related genes in shoot tissue of plants growing in soils with long-term organic fertilization, previously interpreted as stress-priming effect (Chowdhury et al., 2019), was not associated with suppression of Olpidium. In contrast, increased rhizosphere relative abundance of Pseudomonas sp. and M. elongata and increased accumulation of the antifungal root exudate benzoic acid coincided with reduction in disease severity caused by Olpidium in the CONMIN soil with long-term mineral fertilization, suggesting a function in pathogen defense. Obviously, the characteristics reported for long-term organic fertilization cannot be generalized as similarly documented in previous studies (Hartman et al., 2018; Chowdhury et al., 2019).
Furthermore, the results suggest that the effects of fertilization management on the chemical composition of the rhizosphere soil solution, as a major driver for shaping rhizosphere microbial communities, are not simply controlled by root activity and rhizodeposition but also by site-specific factors e.g., turn-over of soil organic matter and substrate competitions between different groups of rhizosphere microorganisms. Additionally, soil properties such as moisture level and water-holding capacity or the availability of specific mineral nutrients may act as modulators for the assemblage of rhizosphere microbiota.
The results point to a complex network of belowground interactions between plant roots, physicochemical soil properties and different groups of rhizosphere microbiota (schematically summarized in Figure 8), with an important role in shaping also the aboveground characteristics of the plants.
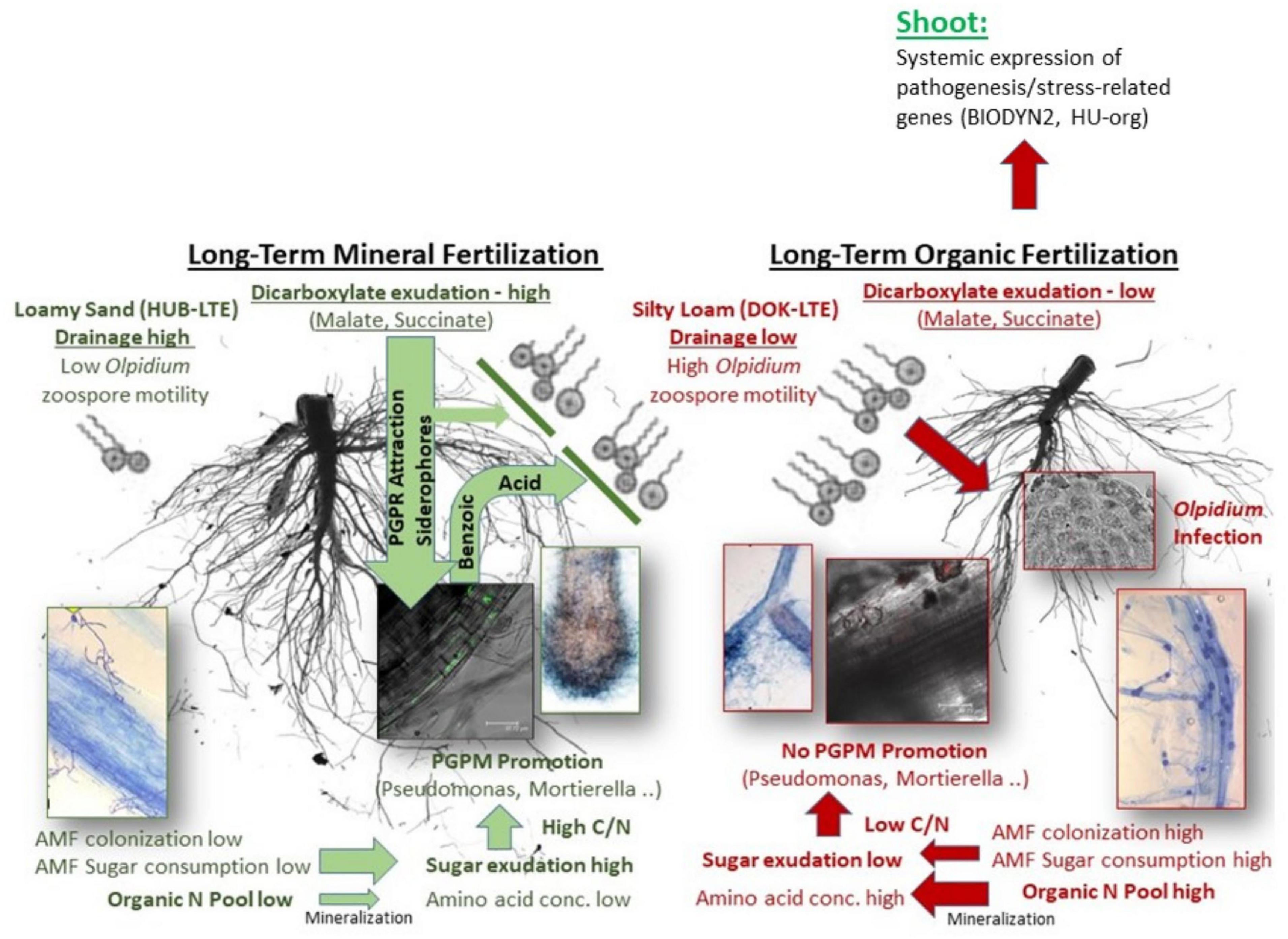
Figure 8. Hypothetic model, summarizing the potential interplay between fertilization management, soil factors, root exudation and rhizosphere microbial communities in lettuce-Olpidium interactions. Pathogen-conducive scenarios are indicated by red labels, while green labels represent pathogen-suppressive scenarios. A high drainage potential of the loamy sand soils (HUB-LTE) counteracts the motility of Olpidium zoospores, which is stimulated by low moisture draining in the silty loam soils (DOK-LTE), thereby promoting Olpidium pathogen pressure. In the pathogen suppressive scenario, long-term mineral fertilization reduces the soil AMF inoculum (arbuscular mycorrhizal fungi). Consequently, lower AMF colonization is associated with lower sugar consumption by the fungal partner and with higher sugar exudation. The higher availability of easily accessible carbohydrates attracts rhizosphere colonization by Gammaproteobacteria (i.e., Pseudomonadaceae) and Mortierella with documented plant growth-promoting and pathogen-suppressive properties. This is further promoted by higher root exudation of dicarboxylates (i.e., succinate) with known functions as chemoattractants and substrates for siderophore production in many Pseudomonas species with plant growth-promoting properties. Moreover, Pseudomonas inoculation can stimulate root exudation of benzoate with antifungal properties in lettuce. By contrast, in the pathogen-conducive scenario, high relative abundance of AMF in the rhizosphere associated with high fungal sugar consumption drastically reduces the rhizosphere sugar concentrations of lettuce plants grown in DOK-LTE soils with long-term organic fertilization (BIODYN2), counteracting root colonization by potentially beneficial Pseudomonadaceae and Mortierella species, thereby favoring Olpidium infection. A low C/N ratio in the rhizosphere soil solution, promoted by high background concentrations of amino acids probably related with more intense organic N mineralization under long-term organic fertilization further reduces the relative abundance of Pseudomonadaceae in the rhizosphere (for details, see section “Discussion”). Photos by courtesy of Markus Weinmann, Namis Eltlbany, and Khalid Hameed.
Although the presented results, so far based on model experiments under controlled conditions are mainly based on descriptions of coinciding relationships without consideration of temporal or spatial variations and lacking detailed mechanistic explanations, the study can provide a starting point for more focused investigations into the related processes. The identification of key players in these networks and their effects on plant health and crop performance may provide important information to optimize the interactive effects of fertilization management, related to rhizosphere processes and development of novel plant growth-promoting microorganisms inoculants. Finally, our results may contribute to the development of practical approaches according to the concept of “soil biological engineering” (Bender et al., 2016).
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, PRJNA622892 and http://www.ebi.ac.uk/ena/data/view/PRJEB39853.
Author Contributions
SW, LS, DB, GN, RG, JG, and KS conceived and designed the experiment. SW, LS, DB, and SC conducted the experiments and collected the data. SW, LS, and DB wrote the manuscript with GN, UL, RG, JG, and KS. FW, BH, NM, and SW developed and performed the UHPLC-MS analysis of amino acids and benzoic acid. WA and SW developed and performed the HPLC-ELSD analysis of sugars. SW and GN developed and performed the RP-HPLC analysis of organic acids. DB, KS, JN, and SS performed and evaluated the sequencing of bacterial and archaeal communities. LS, IS, and JG developed and evaluated the sequencing and analysis of fungal communities. SC and MR developed and performed the plant gene expression analysis. SW and AE performed the detection of pathogen infection in the root tissue. All authors approved the final manuscript.
Funding
This research work contains results obtained in the project DiControl (Project numbers: 031A560A-F), which has been funded by the German Federal Ministry of Education and Research (BMBF) through the program BonaRes—soil as a sustainable resource for the bioeconomy.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We express our gratitude to Konstantin Albrecht and Martin Sandmann, Helene Ochott, Charlotte Haake, Heidi Zimmermann, Elke Dachtler, and Ute Zimmerling for excellent technical assistance and help during sample preparation. Special thanks to Dr. Kathlin Schweitzer und Michael Baumecker for providing the HUB-LTE soils (Experiment Thy_ABS “Cropping Systems” in Thyrow; Albrecht Daniel Thaer – Institute of Humboldt-Universität zu Berlin) and to Andreas Fliessbach and Paul Mäder (Department of Soil Sciences, Research Institute of Organic Agriculture (FiBL), Frick, Switzerland) for the access to DOK-LTE soils.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.597745/full#supplementary-material
Supplementary Figure 1 | Habitus of lettuce plants grown for six weeks in BIODYN2 soil infected with Olpidium sp. (likely Olpidium brassicae) (A–C) infected fine roots with intracellular fungal structures (arrows) and inhibition of root hair development, (D) Olpidium resting spore ín the root tissue (E) sporangium of Olpidium in the root tissue. Photos by courtesy of Abbas El-Hasan.
Supplementary Table 1 | List of plant genes selected for expression analysis with their corresponding loci, functions in A. thaliana and primer sequences.
Supplementary Table 2 | Shoot and root biomass production and root growth parameters of lettuce (cv. Tizian). The plants were grown in minirhizotron culture in two independent experiments during a culture period of nine weeks and six weeks, respectively, on soils with long-term organic (HU-org, BIODYN2) or mineral (HU-min, CONMIN) fertilization history. Means ± standard errors of four independent replicates per treatment. Different lowercase letters indicate significant differences between organic vs. mineral fertilization tested separately for the sites DOK-LTE and HUB-LTE by one-way ANOVA, Tukey’s HSD pairwise test, p ≤ 0.05.
Supplementary Table 3 | Sugars (A), carboxylates (B) and amino acids (C) in soil solutions of samples collected without visible contact to lettuce roots grown in soils with long-term organic (HU-org, BIODYN2) or mineral (HU-min, CONMIN) fertilization history. The plants were grown in minirhizotron culture for nine weeks. Exudate collection was undertaken by micro-sampling with sorption filters on bulk soil, without any root contact. Means ± standard errors. Different lowercase letters indicate significant differences between organic vs. mineral fertilization tested separately for each long-term experimental site DOK-LTE and HUB-LTE by t-test (p ≤ 0.05). n.d. = not detectable.
Supplementary Table 4 | Taxonomic composition of (A) bacterial/archaeal phyla and proteobacterial classes and (B) fungal phyla represented as relative abundances in root-associated soil and rhizosphere of lettuce (cv. Tizian). The plants were grown in soils under long-term organic (HU-org, BIODYN2) or mineral (HU-min, CONMIN) fertilization. Different lowercase letters indicate significant differences between organic vs. mineral fertilization tested separately for each long-term experimental site and habitat by edgeR (FDR < 0.05). Means of relative abundance ± standard errors.
Supplementary Table 5 | Ecological assignment of fungal genera represented as relative abundance with FUNGuild in root-associated soils and rhizosphere of lettuce (cv. Tizian) grown in soils of DOK-LTE (BIODYN2 vs. CONMIN) and HUB-LTE (HU-org vs. HU-min). Different lowercase letters indicate significant differences (FDR < 0.05) of represented genera between organic vs. mineral fertilization tested separately for each long-term experimental site and habitat based on the results of edgeR analyses. Means of relative abundance ± standard errors. Relative abundances (> 0.5%) with significant differences are marked in bold.
Supplementary Table 6 | Fungal taxa in the root-associated soils of lettuce (cv. Tizian) differing significantly (FDR < 0.05) in relative abundance depending on long-term organic vs. mineral fertilization practice at HUB-LTE (HU-org vs. HU-min) (A) and DOK-LTE (BIODYN2 vs. CONMIN) (B). Only taxa with > 1.0% relative abundance are displayed. For OTUs on species level the similarity compared to the database are represented. Means of relative abundance ± standard errors. Bold numbers indicate significant enrichment.
Supplementary Text 1 | Materials and Methods description of microbial community analyses: Bacterial and archaeal communities.
Supplementary Text 2 | Materials and Methods description of microbial community analyses: Fungal communities.
Footnotes
- ^ www.arabidopsis.org
- ^ https://www.ncbi.nlm.nih.gov/sra
- ^ https://www.mothur.org/wiki/RDP_reference_files
- ^ https://www.ebi.ac.uk/ena/browser/home
- ^ http://hannonlab.cshl.edu/fastx_toolkit/
- ^ https://galaxyproject.org
References
Adesina, M. F., Lembke, A., Costa, R., Speksnijder, A., and Smalla, K. (2007). Screening of bacterial isolates from various European soils for in vitro antagonistic activity towards Rhizoctonia solani and Fusarium oxysporum: site-dependent composition and diversity revealed. Soil Biol. Biochem. 39, 2818–2828. doi: 10.1016/j.soilbio.2007.06.004
Antweiler, K., Schreiter, S., Keilwagen, J., Baldrian, P., Kropf, S., Smalla, S., et al. (2017). Statistical test for tolerability of effects of an antifungal biocontrol strain on fungal communities in three arable soils. Microb. Biotechnol. 10, 434–449. doi: 10.1111/1751-7915.12595
Audenaert, K., Patterey, T., Cornelis, P., and Höfte, M. (2002). Induction of systemic resistance to Botrytis cinerea in tomato by Pseudomonas aeruginosa 7NSK2: role of salicylic acid, pyochelin, and pyocyanin. Mol. Plant Microbe Interact. 15, 1147–1156.
Babin, D., Deubel, A., Jacquiod, S. J., Sørensen, S. J., Geistlinger, J., Grosch, R., et al. (2019). Impact of long-term agricultural management practices on soil prokaryotic communities. Soil Biol. Biochem. 129, 17–28. doi: 10.1016/j.soilbio.2018.11.002
Baetz, U., and Martinoia, E. (2014). Root exudates: the hidden part of plant defense. Trends Plant Sci. 19, 90–98. doi: 10.1016/j.tplants.2013.11.006
Bais, H. P., Weir, T. L., Perry, L. G., Gilroy, S., and Vivanco, J. M. (2006). The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 57, 233–266. doi: 10.1146/annurev.arplant.57.032905.105159
Bakker, M. G., Schlatter, D. C., Otto-Hanson, L., and Kinkel, L. L. (2014). Diffuse symbioses: roles of plant-plant, plant-microbe and microbe-microbe interactions in structuring the soil microbiome. Mol. Ecol. 23, 1571–1583. doi: 10.1111/mec.12571
Bakker, P. A. H. M., Pieterse, C. M. J., de Jonge, R., and Berendsen, R. L. (2018). The soil-borne legacy. Cell 172, 1178–1180. doi: 10.1016/j.cell.2018.02.024
Bender, S. F., Wagg, C., and van der Heijden, M. G. A. (2016). An underground revolution: biodiversity and soil ecological engineering for agricultural sustainability. Trends Ecol. Evol. 31, 440–452. doi: 10.1016/j.tree.2016.02.016
Berardini, T. Z., Reiser, L., Li, D., Mezheritsky, Y., Muller, R., Strait, E., et al. (2015). The Arabidopsis information resource: making and mining the “gold standard” annotated reference plant genome. Genesis 53, 474–485. doi: 10.1002/dvg.22877
Berendsen, R. L., Pieterse, C. M. J., and Bakker, P. A. H. M. (2012). The rhizosphere microbiome and plant health. Trends Plant Sci. 17, 478–486. doi: 10.1016/j.tplants.2012.04.001
Berg, G., Rybakova, D., Grube, M., and Köberl, M. (2016). The plant microbiome explored: implications for experimental botany. J. Exp. Bot. 67, 995–1002. doi: 10.1093/jxb/erv466
Campbell, R. (1985). Longevity of Olpidium brassicae in air-dry soil and the persistence of the lettuce big-vein agent. Can. J. Bot. 63, 2288–2289. doi: 10.1139/b85-326
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Lozupone, C. A., Turnbaugh, P. J., et al. (2011). Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. U.S.A. 108, 4516–4522. doi: 10.1073/pnas.1000080107
Carter, D. O., Tomberlin, J. K., Benbow, M. E., and Metcalf, J. L. (2017). Forensic Microbiology. Hoboken, NJ: John Wiley & Sons.
Chaparro, J. M., Badri, D. V., Bakker, M. G., Sugiyama, A., Manter, D. K., and Vivanco, J. M. (2013). Root exudation of phytochemicals in Arabidopsis follows specific patterns that are developmentally programmed and correlate with soil microbial functions. PLoS One 8:e55731. doi: 10.1371/journal.pone.0055731
Chesters, C., and Peberdy, J. F. (1965). Nutritional factors in relation to growth and fat synthesis in Mortierella vinacea. J. Gen. Microbiol. 41, 127–134.
Chowdhury, S. P., Babin, D., Sandmann, M., Jacquiod, S., Sommermann, L., Sørensen, S. J., et al. (2019). Effect of long-term organic and mineral fertilization strategies on rhizosphere microbiota assemblage and performance of lettuce. Environ. Microbiol. 21, 2426–2439. doi: 10.1111/1462-2920.14631
Cohen, S. A., and Michaud, D. P. (1993). Synthesis of a fluorescent derivatizing reagent, 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate, and its application for the analysis of hydrolysate amino acids via high-performance liquid chromatography. Anal. Biochem. 211, 279–287. doi: 10.1006/abio.1993.1270
Cole, J. R., Wang, Q., Fish, J. A., Chai, B., McGarrell, D. M., Sun, Y., et al. (2014). Ribosomal database project: data and tools for high throughput rRNA analysis. Nucleic Acids Res. 42, 633–642. doi: 10.1093/nar/gkt1244
De Cara, M., López, V., Córdoba, M. C., Santos, M., Jordá, C., and Tello, J. C. (2008). Association of Olpidium bornovanus and Melon necrotic spot virus with vine decline of melon in Guatemala. Plant Dis. 92, 709–713. doi: 10.1094/PDIS-92-5-0709
De Mendiburu, F. (2020). Agricolae: Statistical Procedures for Agricultural Research. Available online at: https://CRAN.R-project.org/package=agricolae (accessed November 29, 2020).
Doornbos, R. F., Van Loon, L. C., and Bakker, P. A. H. M. (2012). Impact of root exudates and plant defense signaling on bacterial communities in the rhizosphere. A review. Agron. Sustain. Dev. 32, 227–243. doi: 10.1007/s13593-011-0028-y
Douds, D. D., Galvez, L., Franke-Snyder, M., Reider, C., and Drinkwater, L. E. (1997). Effect of compost addition and crop rotation point upon VAM fungi. Agric. Ecosyst. Environ. 65, 257–266. doi: 10.1016/S0167-8809(97)00075-3
Edgar, R. (2013). UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 10, 996–998. doi: 10.1038/nmeth.2604
Esperschütz, J., Gattinger, A., Mäder, P., Schloter, M., and Fließbach, A. (2007). Response of soil microbial biomass and community structures to conventional and organic farming systems under identical crop rotations. FEMS Microbiol. Ecol. 61, 26–37. doi: 10.1111/j.1574-6941.2007.00318.x
Fitzpatrick, C. R., Salas-González, I., Conway, J. M., Finkel, O. M., Gilbert, S., Russ, D., et al. (2020). The plant microbiome: from ecology to reductionism and beyond. Annu. Rev. Microbiol. 74, 81–100. doi: 10.1146/annurev-micro-0226
Foley, J. A., DeFries, R., Asner, G. P., Barford, C., Bonan, G., Carpenter, S. R., et al. (2005). Global consequences of land use. Science 309, 570–574. doi: 10.1126/science.1111772
Fox, F., and Weisberg, S. (2019). An R Companion to Applied Regression, 3rd Edn. Thousand Oaks CA: Sage.
Francioli, D., Schulz, E., Lentendu, G., Wubet, T., Buscot, F., and Reitz, T. (2016). Mineral vs. organic amendments: microbial community structure, activity and abundance of agriculturally relevant microbes are driven by long-term fertilization strategies. Front. Microbiol. 7:1446. doi: 10.3389/fmicb.2016.01446
Garrido-Sanz, D., Meier-Kolthoff, J. P., Göker, M., Martín, M., Rivilla, R., and Redondo-Nieto, M. (2016). Genomic and genetic diversity within the Pseudomonas fluorescens complex. PLoS One 11:e0150183. doi: 10.1371/journal.pone.0150183
Geisseler, D., and Scow, K. M. (2014). Long-term effects of mineral fertilizers on soil microorganisms – a review. Soil Biol. Biochem. 75, 54–63. doi: 10.1016/j.soilbio.2014.03.023
Gosling, P., Hodge, A., Goodlass, G., and Bending, G. D. (2006). Arbuscular mycorrhizal fungi and organic farming. Agric. Ecosyst. Environ. 113, 17–35. doi: 10.1016/j.agee.2005.09.009
Haase, S., Neumann, G., Kania, A., Kuzyakov, Y., Römheld, V., and Kandeler, E. (2007). Elevation of atmospheric CO2 and N-nutritional status modify nodulation, nodule-carbon supply, and root exudation of Phaseolus vulgaris L. Soil Biol. Biochem. 39, 2208–2221. doi: 10.1016/j.soilbio.2007.03.014
Hacquard, S., Spaepen, S., Garrido-Oter, R., and Schulze-Lefert, P. (2017). Interplay between innate immunity and the plant microbiota. Annu. Rev. Phytopathol. 55, 565–589. doi: 10.1146/annurev-phyto-080516-035623
Hao, Z., Xie, W., and Chen, B. (2019). Arbuscular mycorrhizal symbiosis affects plant immunity to viral infection and accumulation. Viruses 11, 1–12. doi: 10.3390/v11060534
Hartman, K., van der Heijden, M. G. A., Wittwer, R. A., Banerjee, S., Walser, J.-C., and Schlaeppi, K. (2018). Cropping practices manipulate abundance patterns of root and soil microbiome members paving the way to smart farming. Microbiome 6, 2–14. doi: 10.1186/s40168-017-0389-9
Hartmann, M., Frey, B., Mayer, J., Mäder, P., and Widmer, F. (2014). Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 9, 1177–1194. doi: 10.1038/ismej.2014.210
Henry, G., Thonart, P., and Ongena, M. (2012). PAMPs, MAMPs, DAMPs and others: an update on the diversity of plant immunity elicitors. Biotechnol. Agron. Soc. Environ. 16, 257–268.
Höfte, M., Seong, K. Y., Jurkevitch, E., and Verstraete, W. (1991). Pyoverdin production by the plant growth beneficial Pseudomonas strain 7NSK2: ecological significance in soil. Plant Soil 130, 249–257. doi: 10.1007/BF00011880
Jones, D. L., Hodge, A., and Kuzyakov, Y. (2004). Plant and mycorrhizal regulation of rhizodeposition. New Phytol. 163, 459–480. doi: 10.1111/j.1469-8137.2004.01130.x
Kadota, Y., Shirasu, K., and Zipfel, C. (2015). Regulation of the NADPH oxidase RbohD during plant immunity. Plant Cell Physiol. 56, 1472–1480. doi: 10.1093/pcp/pcv063
Kamzolova, S. V., Vinokurova, N. G., Dedyukhina, E. G., Samoilenko, V. A., Lunina, J. N., Mironov, A. A., et al. (2014). The peculiarities of succinic acid production from rapeseed oil by Yarrowia lipolytica yeast. Appl. Microbiol. Biotechnol. 98, 4149–4157. doi: 10.1007/s00253-014-5585-y
Kolde, R. (2019). pheatmap: Pretty Heatmaps. Available online at: https://CRAN.R-project.org/package=pheatmap (accessed November 29, 2020).
Kuramae, E. E., Yergeau, E., Wong, L. C., Pijl, A. S., Van Veen, J. A., and Kowalchuk, G. A. (2012). Soil characteristics more strongly influence soil bacterial communities than land-use type. FEMS Microbiol. Ecol. 79, 12–24. doi: 10.1111/j.1574-6941.2011.01192.x
Lapsansky, E. R., Milroy, A. M., Andales, M. J., and Vivanco, J. M. (2016). Soil memory as a potential mechanism for encouraging sustainable plant health and productivity. Curr. Opin. Biotechnol. 38, 137–142. doi: 10.1016/j.copbio.2016.01.014
Li, F., Chen, L., Gordon, M. R., Zhang, J., Zhang, C., Ning, Q., et al. (2018). Mortierella elongata ‘s roles in organic agriculture and crop growth promotion in a mineral soil. Wiley 29, 1642–1651. doi: 10.1002/ldr.2965
Liu, G., Greenshields, D. L., Sammynaiken, R., Hirji, R. N., Selvaraj, G., and Wei, Y. (2007). Targeted alterations in iron homeostasis underlie plant defense responses. J. Cell Sci. 120, 596–605. doi: 10.1242/jcs.001362
Liu, H., Brettell, L. E., Qiu, Z., and Singh, B. K. (2020). Microbiome-mediated stress resistance in plants. Trends Plant Sci. 25, 733–743. doi: 10.1016/j.tplants.2020.03.014
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Loreau, M., Naeem, S., Inchausti, P., Bengtsson, J., Grime, J. P., Hector, A., et al. (2001). Ecology: biodiversity and ecosystem functioning: current knowledge and future challenges. Science 294, 804–808. doi: 10.1126/science.1064088
Lori, M., Symnaczik, S., Mäder, P., De Deyn, G., and Gattinger, A. (2017). Organic farming enhances soil microbial abundance and activity–a meta-analysis and meta-regression. PLoS One 12:e0180442. doi: 10.1371/journal.pone.0180442
Mäder, P., Fließbach, A., Dubois, D., Gunst, L., Fried, P., and Niggli, U. (2002). Soil fertility and biodiversity in organic farming. Science 296, 1694–1697. doi: 10.1126/science.1071148
Magoč, T., and Salzberg, S. L. (2011). FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics 27, 2957–2963. doi: 10.1093/bioinformatics/btr507
Marek-Kozaczuk, M., and Skorupska, A. (1997). Physiological parameters infuencing the production of siderophore by PGPR Pseudomonas sp. strain 267. Acta Microbiol. Pol. 46, 157–165.
Martin, M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 17, 10–12. doi: 10.14806/ej.17.1.200
McCarthy, D. J., Chen, Y., and Smyth, G. K. (2012). Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 40, 4288–4297. doi: 10.1093/nar/gks042
McMurdie, P. J., and Holmes, S. (2013). phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8:e61217. doi: 10.1371/journal.pone.0061217
Mendes, R., Garbeva, P., and Raaijmakers, J. M. (2013). The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 37, 634–663. doi: 10.1111/1574-6976.12028
Michaud, L., Lo Giudice, A., Interdonato, F., Triplet, S., Ying, L., and Blancheton, J. P. (2014). C/N ratio-induced structural shift of bacterial communities inside lab-scale aquaculture biofilters. Aquac. Eng. 58, 77–87. doi: 10.1016/j.aquaeng.2013.11.002
Misaghi, I. J., and Grogan, R. G. (1978). Physiological basis for tipburn development in head lettuce. Phytopathology 68:1744. doi: 10.1094/phyto-68-1744
Miura, K., and Tada, Y. (2014). Regulation of water, salinity, and cold stress responses by salicylic acid. Front. Plant Sci. 5:4. doi: 10.3389/fpls.2014.00004
Nagahashi, G., Douds, D. D., and Abney, G. D. (1996). Phosphorus amendment inhibits hyphal branching of the VAM fungus Gigaspora margarita directly and indirectly through its effect on root exudation. Mycorrhiza 6, 403–408. doi: 10.1007/s005720050139
Narula, N., Kothe, E., and Behl, R. K. (2009). Role of root exudates in plant-microbe interactions. J. Appl. Bot. Food Qual. 82, 122–130.
Navarro, J. A., Botella, F., Maruhenda, A., Sastre, P., Sánchez-Pina, M. A., and Pallas, V. (2004). Comparative infection progress analysis of Lettuce big-vein virus and Mirafiori lettuce virus in lettuce crops by developed molecular diagnosis techniques. Phytopathology 94, 470–477. doi: 10.1094/PHYTO.2004.94.5.470
Neumann, G. (2006). “Root exudates and organic composition of plant roots,” in Handbook of Methods Used in Rhizosphere Research, eds J. Luster, R. Finlay, I. Brunner, W. J. Fitz, B. Frey, A. Göttlein, et al. (Birmensdorf: Swiss Federal Institute for Forest, Snow and Landscape Research).
Neumann, G., and Römheld, V. (2007). “The release of root exudates as affected by the plant physiological status,” in The Rhizosphere: Biochemistry and Organic Substances at the Soil-Plant Interface, eds R. Pinton, Z. Varanini, and Z. Nannipieri (Boca Raton, FL: CRC Press), 22–72.
Neumann, G., Bott, S., Ohler, M. a, Mock, H.-P., Lippmann, R., Grosch, R., et al. (2014). Root exudation and root development of lettuce (Lactuca sativa L. cv. Tizian) as affected by different soils. Front. Microbiol. 5:2. doi: 10.3389/fmicb.2014.00002
Nguyen, N. H., Song, Z., Bates, S. T., Branco, S., Tedersoo, L., Menke, J., et al. (2016). FUNGuild: an open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 20, 241–248. doi: 10.1016/j.funeco.2015.06.006
Oksanen, J., Blanchet, G. F., and Friendly, M. (2019). vegan: Community Ecology Package. R package version 2.5-6. Available online at: https://cran.r-project.org/web/packages/vegan/index.html (accessed November 29, 2020).
Oku, S., Komatsu, A., Nakashimada, Y., Tajima, T., and Kato, J. (2014). Identification of Pseudomonas fluorescens chemotaxis sensory proteins for malate, succinate, and fumarate, and their involvement in root colonization. Microbes Environ. 29, 413–419. doi: 10.1264/jsme2.ME14128
Olsson, P. A., van Aarle, M. I, Allaway, W. G., Ashford, A. E., and Rouhier, H. (2003). Phosphorus effects on metabolic processes in monoxenic arbuscular mycorrhiza cultures. Plant Physiol. 130, 1162–1171.
Op De Beeck, M., Lievens, B., Busschaert, P., Declerck, S., Vangronsveld, J., and Colpaert, J. V. (2014). Comparison and validation of some ITS primer pairs useful for fungal metabarcoding studies. PLoS One 9:e97629. doi: 10.1371/journal.pone.0097629
Robertson, G. P., and Vitousek, P. M. (2009). Nitrogen in agriculture: balancing the cost of an essential resource. Annu. Rev. Environ. Resour. 34, 97–125. doi: 10.1146/annurev.environ.032108.105046
Robinson, M. D., McCarthy, D. J., and Smyth, G. K. (2010). edgeR: a bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. doi: 10.1093/bioinformatics/btp616
Roquigny, R., Novinscak, A., Biessy, A., and Filion, M. (2017). “Pseudomonadaceae: from biocontrol to plant growth promotion,” in Rhizotrophs: Plant Growth Promotion to BIOREMEDIATION, ed. S. Mehnaz (Singapore: Springer), 39–68. doi: 10.1007/978-981-10-4862-3_3
Sampedro, I., Parales, R. E., Krell, T., and Hill, J. E. (2015). Pseudomonas chemotaxis. FEMS Microbiol. Rev. 39, 17–46. doi: 10.1111/1574-6976.12081
Sattelmacher, B., Reinhard, S., and Pomikalko, A. (1991). Differences in mycorrhizal colonization of Rye (Secale cereala L.) grown in conventional or organic (Biological-dynamic) farming systems. J. Agron. Crop Sci. 167, 350–355.
Schloss, P. D., Westcott, S. L., Ryabin, T., Hall, J. R., Hartmann, M., Hollister, E. B., et al. (2009). Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541. doi: 10.1128/AEM.01541-09
Schmid, C. A. O., Schröder, P., Armbruster, M., and Schloter, M. (2018). Organic amendments in a long-term field trial—consequences for the bulk soil bacterial community as revealed by network analysis. Microb. Ecol. 76, 226–239. doi: 10.1007/s00248-017-1110-z
Schmidt, J. E., Kent, A. D., Brisson, V. L., and Gaudin, C. M. (2019). Agricultural management and plant selection interactively affect rhizosphere microbial community structure and nitrogen cycling. Microbiome 7:146. doi: 10.1186/s40168-019-0756-9
Schreiter, S., Ding, G. C., Grosch, R., Kropf, S., Antweiler, K., and Smalla, K. (2014a). Soil type-dependent effects of a potential biocontrol inoculant on indigenous bacterial communities in the rhizosphere of field-grown lettuce. FEMS Microbiol. Ecol. 90, 718–730. doi: 10.1111/1574-6941.12430
Schreiter, S., Ding, G.-C., Heuer, H., Neumann, G., Sandmann, M., Grosch, R., et al. (2014b). Effect of the soil type on the microbiome in the rhizosphere of field-grown lettuce. Front. Microbiol. 5:144. doi: 10.3389/fmicb.2014.00144
Shen, P., Murphy, D. V., George, S. J., Lapis-Gaza, H., Xu, M., and Gleeson, D. B. (2016). Increasing the size of the microbial biomass altered bacterial community structure which enhances plant phosphorus uptake. PLoS One 11:e0166062. doi: 10.1371/journal.pone.0166062
Sommermann, L., Geistlinger, J., Wibberg, D., Deubel, A., Zwanzig, J., Babin, D., et al. (2018). Fungal community profiles in agricultural soils of a long-term field trial under different tillage, fertilization and crop rotation conditions analyzed by high-throughput ITS-amplicon sequencing. PLoS One 13:e0195345. doi: 10.1371/journal.pone.0195345
Spoel, S. H., and Dong, X. (2012). How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 12, 89–100.
Stacey, M. G., Patel, A., McClain, W. E., Mathieu, M., Remley, M., Rogers, E. E., et al. (2008). The Arabidopsis AtOPT3 protein functions in metal homeostasis and movement of iron to developing seeds. Plant Physiol. 146, 589–601. doi: 10.1104/pp.107.108183
Sundberg, C., Al-Soud, W. A., Larsson, M., Alm, E., Yekta, S. S., Svensson, B. H., et al. (2013). 454 Pyrosequencing analyses of bacterial and archaeal richness in 21 full-scale biogas digesters. FEMS Microbiol. Ecol. 85, 612–626. doi: 10.1111/1574-6941.12148
Tamayo-Velez, A., and Osorio, N. W. (2017). Co-inoculation with an arbuscular mycorrhizal fungus and a phosphate-solubilizing fungus promotes the plant growth and phosphate uptake of avocado plantlets in a nursery. Botany 95, 539–545. doi: 10.1139/cjb-2016-0224
Turner, T. R., Ramakrishnan, K., Walshaw, J., Heavens, D., Alston, M., Swarbreck, D., et al. (2013). Comparative metatranscriptomics reveals kingdom level changes in the rhizosphere microbiome of plants. ISME J. 7, 2248–2258. doi: 10.1038/ismej.2013.119
UNITE Community (2017). UNITE General FASTA Release. Version 01.12.2017. UNITE Community. Available online at: https://plutof.ut.ee/#/doi/10.15156/BIO/587475 (accessed Dezember 7, 2017).
Untergasser, A., Nijveen, H., Rao, X., Bisseling, T., Geurts, R., and Leunissen, J. A. M. (2007). Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 35, 71–74. doi: 10.1093/nar/gkm306
van der Putten, W. H., Bradford, M. A., Pernilla Brinkman, E., van de Voorde, T. F. J., and Veen, G. F. (2016). Where, when and how plant–soil feedback matters in a changing world. Funct. Ecol. 30, 1109–1121. doi: 10.1111/1365-2435.12657
Venables, W. N., and Ripley, B. D. (2002). Modern Applied Statistics with S, 4th Edn. New York, NY: Springer.
Verbon, E. H., Trapet, P. L., Stringlis, I. A., Kruijs, S., Bakker, P. A. H. M., and Pieterse, M. J. (2017). Iron and Immunity. Annu. Rev. Phytopathol. 55, 255–275.
Vujinović, T., Zanin, L., Venuti, S., Contin, M., Ceccon, P., Tomasi, N., et al. (2020). Biostimulant action of dissolved humic substances from a conventionally and an organically managed soil on nitrate acquisition in maize plants. Front. Plant Sci. 10:1652. doi: 10.3389/fpls.2019.01652
Walters, D. R., Walker, R. L., and Walker, K. C. (2003). Lauric acid exhibits antifungal activity against plant pathogenic fungi. J. Phytopathol. 151, 228–230.
Warnes, G. R., Bolker, B., Bonebakker, L., Gentleman, R., Huber, W., Liaw, A., et al. (2019). gplots: Various R Programming Tools for Plotting Data. Available online at: https://CRAN.R-project.org/package=gplots (accessed November 29, 2020).
Westerlund, F. V., Campbell, R. N., Grogran, R. G., and Duniway, J. M. (1978). Soil factors affecting the reproduction and survival of Olpidium brassicae and its transmission of big vein agent to lettuce. Phytopathology 68:927. doi: 10.1094/phyto-68-927
White, T. J., Bruns, T., Lee, S., and Taylor, T. (1990). “Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics,” in PCR Protocols: A Guide to Methods and Applications, eds M. A. Innis, D. H. Gelfand, J. J. Sninsky, and T. J. White (Cambridge, MA: Academic Press), 315–322.
Windisch, S., Bott, S., Ohler, M., Mock, H., Lippmann, R., Grosch, R., et al. (2017). Rhizoctonia solani and bacterial inoculants stimulate root exudation of antifungal compounds in lettuce in a soil-type specific manner. Agronomy 7, 1–17. doi: 10.3390/agronomy7020044
Yoon, M., Seo, K. H., Lee, S. H., Choi, G. J., Jang, K. S., Choi, Y. H., et al. (2012). Antifungal activity of benzoic acid from bacillus subtilis GDYA-1 against fungal phytopathogens. Res. Plant Dis. 18, 109–116.
Yu, Y., Lee, C., Kim, J., and Hwang, S. (2005). Group-specific primer and probe sets to detect methanogenic communities using quantitative real-time polymerase chain reaction. Biotechnol. Bioeng. 89, 670–679. doi: 10.1002/bit.20347
Keywords: fertilization management, root exudates, rhizosphere microbiota, high-throughout amplicon sequencing, 16S rRNA, fungal ITS2 region, stress-related gene expression
Citation: Windisch S, Sommermann L, Babin D, Chowdhury SP, Grosch R, Moradtalab N, Walker F, Höglinger B, El-Hasan A, Armbruster W, Nesme J, Sørensen SJ, Schellenberg I, Geistlinger J, Smalla K, Rothballer M, Ludewig U and Neumann G (2021) Impact of Long-Term Organic and Mineral Fertilization on Rhizosphere Metabolites, Root–Microbial Interactions and Plant Health of Lettuce. Front. Microbiol. 11:597745. doi: 10.3389/fmicb.2020.597745
Received: 25 August 2020; Accepted: 18 November 2020;
Published: 13 January 2021.
Edited by:
Jacob George Malone, John Innes Centre, United KingdomReviewed by:
Jennifer Elise Schmidt, Mars Wrigley Davis, CA, United StatesHongmiao Wu, Fujian Agriculture and Forestry University, China
Copyright © 2021 Windisch, Sommermann, Babin, Chowdhury, Grosch, Moradtalab, Walker, Höglinger, El-Hasan, Armbruster, Nesme, Sørensen, Schellenberg, Geistlinger, Smalla, Rothballer, Ludewig and Neumann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Saskia Windisch, U2Fza2lhLldpbmRpc2NoQHVuaS1ob2hlbmhlaW0uZGU=; Günter Neumann, Z2QubmV1bWFubkB0LW9ubGluZS5kZQ==
†These authors have contributed equally to this work