 Mathias Hutzler1†
Mathias Hutzler1† Maximilian Michel1†
Maximilian Michel1† Oliver Kunz1
Oliver Kunz1 Tiina Kuusisto2,3
Tiina Kuusisto2,3 Frederico Magalhães2
Frederico Magalhães2 Kristoffer Krogerus2
Kristoffer Krogerus2 Brian Gibson4*
Brian Gibson4*- 1Research Center Weihenstephan for Brewing and Food Quality, Technical University of Munich, Freising, Germany
- 2VTT Technical Research Centre of Finland Ltd., Espoo, Finland
- 3Faculty of Science, University of Copenhagen, Copenhagen, Denmark
- 4Chair of Brewing and Beverage Technology, Technische Universität Berlin, Berlin, Germany
The successful application of Saccharomyces eubayanus and Saccharomyces paradoxus in brewery fermentations has highlighted the potential of wild Saccharomyes yeasts for brewing, and prompted investigation into the application potential of other members of the genus. Here, we evaluate, for the first time, the brewing potential of Saccharomyces jurei. The newly isolated strain from an ash tree (Fraxinus excelsior) in Upper Bavaria, Germany, close to the river Isar, was used to ferment a 12°P wort at 15°C. Performance was compared directly with that of a reference lager strain (TUM 34/70) and the S. eubayanus type strain. Both wild yeast rapidly depleted simple sugars and thereafter exhibited a lag phase before maltose utilization. This phase lasted for 4 and 10 days for S. eubayanus and S. jurei, respectively. S. eubayanus utilized fully the available maltose but, consistent with previous reports, did not use maltotriose. S. jurei, in contrast, utilized approximately 50% of the maltotriose available, making this the first report of maltotriose utilization in a wild Saccharomyces species. Maltotriose use was directly related to alcohol yield with 5.5, 4.9, and 4.5% ABV produced by Saccharomyces pastorianus, S. jurei, and S. eubayanus. Beers also differed with respect to aroma volatiles, with a high level (0.4 mg/L) of the apple/aniseed aroma ethyl hexanoate in S. jurei beers, while S. eubayanus beers had a high level of phenylethanol (100 mg/L). A trained panel rated all beers as being of high quality, but noted clear differences. A phenolic spice/clove note was prominent in S. jurei beer. This was less pronounced in the S. eubayanus beers, despite analytical levels of 4-vinylguaiacol being similar. Tropical fruit notes were pronounced in S. jurei beers, possibly resulting from the high level of ethyl hexanoate. Herein, we present results from the first intentional application of S. jurei as a yeast for beer fermentation (at the time of submission) and compare its fermentation performance to other species of the genus. Results indicate considerable potential for S. jurei application in brewing, with clear advantages compared to other wild Saccharomyces species.
Introduction
Yeasts of the genus Saccharomyces are by far the most prevalent fermentative microorganisms used in brewing. In addition to the commonly used yeasts S. pastorianus and S. cerevisiae, wild species of Saccharomyces such as S. paradoxus and S. eubayanus have recently been utilized intentionally for beer fermentations (Gibson, 2015; Osburn et al., 2016; Nikulin et al., 2020). The successful utilization of these non-domesticated species suggests that other species belonging to the genus could be profitably employed in beer production, and could facilitate differentiation of beers, or even creation of novel beer styles (Osburn et al., 2016). Alternative Saccharomyces species have the potential to introduce novel flavor profiles to beers and, in particular, to lager beers which are fermented at low temperatures (with the exception of S. cerevisiae the genus may be described as psychrophilic) (Magalhães et al., 2021). The potential benefits derived from wild yeasts are off-set by a number of characteristics that may be considered negative in the context of brewing. These include the production of phenolic flavor compounds (which typically lend a spice or smoke note to beer); poor flocculation, which hinders clarification; and inability to use all of the available sugars present in brewers wort, thereby limiting fermentation efficiency. Only S. paradoxus and S. eubayanus have been fully characterized with respect to brewing potential (Gibson et al., 2013; Mardones et al., 2020; Nikulin et al., 2020) and other species in the genus may prove to be more suitable for efficient production of flavorful beers.
Eight Saccharomyces species are currently recognized (Naseeb et al., 2017). These have been isolated from a range of habitats, but appear to be most prevalent in woodland environments. Oaks and other species within the Fagaceae family, for example, serve as a habitat for both S. paradoxus and S. cerevisiae. Sniegowski et al. (2002) have isolated several strains of S. paradoxus and S. cerevisiae from exudate, soil and bark associated with different trees belonging to the Quercus genus, while samples taken from poplar, maple and American beech trees did not yield Saccharomyces yeasts. A clear preference of Saccharomyces for oak trees compared to trees outside the Quercus genus was also shown by Sampaio and Gonçalves (2008). Other Saccharomyces species, isolated from Drosophila sp. in Brazil and from soil and decayed leaves in Japan, have been classified as Saccharomyces cariocanus, Saccharomyces kudriavzevii, and Saccharomyces mikatae, respectively (Naumov et al., 2000) (though S. cariocanus is now often designated S. paradoxus; Liti et al., 2006). S. mikatae has not been isolated from other locations to date suggesting a regional restriction of this species. S. kudriavzevii has also been isolated alongside other Saccharomyces species from oak bark and soil surrounding oak trees in Europe (Sampaio and Gonçalves, 2008). In 2008, a novel Saccharomyces species was isolated from trees in Western China. Three strains of the newly named Saccharomyces arboricola were obtained from the bark of a Quercus sp. and a Castanopsis sp. tree, both belonging to the Fagaceae family (Wang and Bai, 2008). S. eubayanus, the cryotolerant co-parent of today’s widely used lager-brewing yeast S. pastorianus, was first isolated and identified on southern-beech trees (Nothofagus spp.) in Patagonia. Along with S. eubayanus, Saccharomyces uvarum was observed in the same habitats indicating that these two cryotolerant species thrive in the cold climate of Patagonia (Libkind et al., 2011). These two species have later also been isolated in sympatry from trees in China, also including oaks (Bing et al., 2014) and Wisconsin, United States, on American beech and maple trees (Peris et al., 2014).
With the exception of the use of S. eubayanus and S. paradoxus in brewing applications, there are, to our knowledge, few to no examples of wild Saccharomyces yeasts being used directly in biotechnological applications. Indirect applications have involved the creation of interspecies hybrids and assessment of the performance of these hybrids in fermentation processes. Examples have included the use of S. cerevisiae × S. arboricola, S. cerevisiae × S. eubayanus, S. cerevisiae × S. kudriavzevii, and S. cerevisiae × S. mikatae hybrids in beverage fementations and for bioethanol production (Krogerus et al., 2017; Peris et al., 2017; Winans et al., 2020). Unaltered strains and interspecies hybrids of S. jurei, along with other Saccharomyces hybrids, have also been investigated for their baking potential in dough fermentation (Magalhães et al., 2021). Naturally occurring hybrids of S. kudriavzevii × S. cerevisiae have been proven to be abundant in man-made fermentations of several products and beverages, such as Belgian beer (Gallone et al., 2019). Peris et al. (2018) provides an extensive overview of the various occurrences of these hybrids. Molinet and Cubillos (2020) also give an overview of the use and generation of Saccharomyces interspecific hybrids and the generation of intraspecific hybrids of commercial S. cerevisiae and wild S. cerevisiae strains.
In 2017, Naseeb et al. (2018) first isolated and described S. jurei (NCYC 3947) from an oak tree (Quercus robur) in the French Pyrenees. They found a close relationship between S. mikatae and S. jurei through phylogenetic analysis and suggest shared evolutionary history of these two species (Naseeb et al., 2018). In a later study, Alsammar et al. (2019) detected DNA homologous to that of S. jurei in natural habitats using high throughput sequencing of the ITS1 region specific to Saccharomyces spp. By sampling a variety of trees (oak, beech, spruce, larch, and pine) at different altitudes in the Italian Alps, the soil surrounding the trees was scanned for evidence of the presence of members of the Saccharomyces genus. As expected, S. cerevisiae and S. paradoxus were abundant, but more interestingly, S. mikatae and S. jurei were identified in many samples as well as S. eubayanus and S. kudriavzevii (Alsammar et al., 2019). This did not yield any viable strains as only DNA was extracted from the soil. The apparent paucity of S. jurei in nature may be simply an artifact of the culturing methods used for isolation and enrichment, which may disadvantage some species relative to others. The use of wooden materials and tools in brewing has been widespread over centuries due to the relative ease of fabrication while metallic materials have only become the major material over the last century (Schnegg, 1921, 1922). The contact between wood harboring yeasts and wort containing fermentable sugars and nutrients may have been a source of yeast being used for brewing in the past or still today.
In this study, the potential of a S. jurei strain isolated from ash (Fraxinus excelsior) in Bavaria was investigated for its brewing potential. Traits studied included wort fermentation efficiency, sugar utilization, and beer flavor profile. Performance was compared to that of S. eubayanus, which has already been shown to be a capable brewing yeast, as well as the domesticated lager strain S. pastorianus TUM 34/70.
Materials and Methods
Yeast Strains
Strains used in this work are listed in Table 1. All strains with VTT codes were obtained from VTT culture collection1. The S. pastorianus strain TUM 34/70 and S. jurei strain TUM 629 were obtained from the TU Munich, Research Center Weihenstephan for Brewing and Food Quality2.

Table 1. Strains used in this study.
Saccharomyces jurei Isolation, Maintenance, and Microscopy
Samples of the bark of an ash tree (Fraxinus excelsior) located in the meadows of the Isar-river in Munich (Latitude 48.10931383333333 = 48° 6′ 33.53″ N, Longitude 11.56397166666667 = 11° 33′ 50.298″ E, 519.8 m above sea level) with attached moss were taken approximately 1 meter above ground with sterilized forceps and placed into sterile bags and stored for 2 days at 2°C until processing. 1 g samples of the collected bark were put into a flask, sealed with a sterile plug, containing 50 mL of autoclaved wort at 12°P (15 min at 121°C), pH-value 5.3 (prepared from diluted wort concentrate (Döhler, Darmstadt Germany, original gravity approximately 60°) through addition of deionized water and tetracycline (tetracycline-hydrochloride, Carl Roth, Karlsruhe, Germany) at an application concentration of 50 mg/L. 0.25 mL of a saturated alcoholic biphenyl solution were pipetted onto the sterile plug to inhibit growth of mold. Wort was used as a medium to screen for maltose and maltotriose positive yeast isolates that are able to ferment brewer’s wort. This semi-selective enrichment of the yeasts present in the sample favors yeast suited for brewing compared to an enrichment in other yeast media such as Yeast Peptone Dextrose (YPD). The flask containing wort and the sample was then incubated aerobically at 20°C for 2 weeks.
An inoculation loop of the incubated enrichment medium was streaked on a Wallerstein Nutrient Agar plate (WLN-A, Oxoid GmbH, Wesel, Germany, pH 5,6, 50 mg/L tetracycline) and incubated anaerobically for 1 week at 20°C. Single colonies were checked for cell morphology and purity of the colony with a microscope (Nikon, Düsseldorf, Germany) using dark field microscopy with a 1,000-fold magnification. The examined colonies were then streaked onto a fresh WLN-A plate as described above. The colonies subsequently identified as S. jurei appeared small, round, and umbonate with a pale greenish-white coloration. A pure culture of S. jurei was streaked on a wort agar slant which was incubated for 3 days at 28°C before storage at 2°C. For strain maintenance recultivation was performed every month. For long term storage cryotubes in cryo-vials (Roti-Store yeast cryo vials, Carl Roth, Karlsruhe, Germany) at −80°C were prepared according to the manufacturer’s directions.
Initial Identification Using Species Specific Saccharomyces Real-Time PCR Assays and ITS, D1/D2 26S rDNA Sequencing
Yeast DNA was isolated using a modified InstaGene Matrix (Biorad, Feldkirchen, Germany) protocol (Hutzler, 2009; Meier-Dörnberg et al., 2018). Yeast DNA of the single colony were analyzed using different species-specific Saccharomyces Real-Time PCR assays (Hutzler, 2009; Hutzler et al., 2015; Meier-Dörnberg et al., 2017b; Sampaio et al., 2017). Sequencing of ITS and D1/D2 26S rDNA loci was performed acccording to White et al. (1990) and Kurtzman et al. (2003) using modified protocols according to Hutzler (Kurtzman and Robnett, 1998; Hutzler, 2009, 2010). Sequences were analyzed using NCBI Blast tool (NCBI) and DNAStar, MegAlign Software (DNASTAR, Inc., Madison, Wisconsin).
Whole-Genome Sequencing
Genomic DNA from strains S. jurei C1003 and TUM 629 was isolated using Qiagen 100/G Genomic-tips (Qiagen, The Netherlands). The DNA was sequenced at the Microbial Genome Sequencing Center (Pittsburgh, PA, United States). A 150-bp paired-end Illumina Nextera library was prepared, and sequencing was carried out with a NextSeq 550 instrument. The paired-end reads were trimmed and filtered with Trimmomatic (Bolger et al., 2014). Reads were aligned to the reference genome of S. jurei NCYC 3947T (accession number GCA_900290405; Naseeb et al., 2018) using BWA-MEM (Li and Durbin, 2009). Variant analysis was performed on aligned reads using FreeBayes (Garrison and Marth, 2012). Prior to variant analysis, alignments were filtered to a minimum MAPQ of 50 with SAMtools (Li et al., 2009). Annotation and effect prediction of the variants were performed with SnpEff (Cingolani et al., 2012). The median coverage over 10,000-bp windows was calculated with mosdepth (Pedersen and Quinlan, 2017).
In addition, to test if TUM 629 was an interspecies hybrid, the trimmed reads were also aligned to a concatenated reference genome consisting of the assembled genomes of eight Saccharomyces species (Scannell et al., 2011; Liti et al., 2013; Engel et al., 2014; Naseeb et al., 2018). The median coverage over 10,000 bp windows was again calculated with mosdepth, and was visualized in R using modified scripts from sppIDer (Langdon et al., 2018).
Raw sequence reads have been deposited in the NCBI Short Read Archive under BioProject PRJNA681394.
Wort Preparation
The wort for the fermentations was produced in the VTT pilot brewery. Milled Pilsner malt (Viking Malt, Lahti, Finland) was mashed in with local Espoo City water following an infusion mashing procedure (mashing-in at 48°C; rests: 48°C 30 min—63°C 30 min—72°C 30 min—78°C 10 min), mash was filtered with a Meura (Belgium) mash filter and boiled for 60 min with Magnum hop pellets (α-acid content 15%). The wort was hopped to achieve 40 IBU and the strength at knockout was 12°Plato. The wort was collected hot (over 90°C) in stainless steel kegs and stored at 0°C before use. The sugar concentrations of the wort were 52.5 g/L maltose, 13.4 g/L maltotriose, 11.6 g/L glucose, and 2.7 g/L fructose.
Fermentation Trials and Beer Preparation
10 L-scale fermentations were carried out in duplicate with S. jurei TUM 629, S. eubayanus C902, and S. pastorianus TUM 34/70. Yeasts were first propagated by transferring an inoculation loop of yeast from a YPD agar plate to 25 mL liquid YPD culture. The culture was incubated aerobically on a shaker for 24 h, before being transferred to 500 mL YPD. After aerobic incubation with agitation (120 rpm) on an orbital shaker for 3 days, the yeast suspension was centrifuged, a 20%-slurry (200 g fresh yeast/L) was prepared in sterile Milli-Q-filtered water and yeast were inoculated into 1.5 L of 12°P wort in a 2 L Schott-bottle capped with an airlock. After 5 days of static fermentation, the yeast was removed by centrifugation (4,000 rpm; 5 min; 4°C) and a 20% slurry was again prepared. All incubations were performed at 20°C. Cell number was determined using the NucleoCounter YC-100 (ChemoMetec, Denmark) and cells were inoculated into 8 L of aerated (10 ppm dissolved oxygen) 12°P wort in 10 L-volume, stainless-steel, cylindroconical vessels, to give a starting cell density of 1 × 107 cells/mL. Fermentations were conducted at 15°C, and were monitored through regular sampling for assessment of wort pH, alcohol content and cell mass. After fermentation was complete, i.e., when minimal change in wort density was observed over consecutive days, the fresh beers were transferred from fermenters to kegs, matured for 7 days at 10°C and stabilized 7 days at 0°C before depth filtration (Seitz EK, Pall Corporation, New York, NY, United States). Prior to bottling, the beers were carbonated to 5 g/L, and the bottled beers were stored at 0°C.
Screening of Saccharomyces Type Strains for Wort Fermentation Potential
Wort fermentation screening trials included six wild Saccharomyces species and two reference brewing strains: one ale (S. cerevisiae) and one lager (S. pastorianus) strain (Table 1). Prior to fermentation, an inoculation loop was used to transfer yeast from a stock YPD agar plate to the 50 mL liquid YPD medium in a 100 mL Erlenmeyer flask. The cultures were propagated at 20°C on an orbital shaker (100× g, Infors AG TR-125). After 2 days the yeast suspensions were centrifuged (10 min, 9,000× g and 4°C) and a 20% (200 mg/mL) slurry was prepared for cell counting. The NucleoCounter YC-100 was used to calculate the cell count before the yeasts were transferred to the wort at a pitching rate of 1 × 107 cells/mL. Cells were pitched according to cell number rather than mass due to the expected differences in cell size amongst the strains. Fermentations were carried out in 100 mL of the 12°Plato all-malt wort and were conducted in 250 mL Erlenmeyer flasks, without agitation, at the typical lager brewing temperature of 15°C for 40 days. Airlocks containing 2 mL of 85% glycerol were used to seal the flasks. Fermentation progress was monitored by measuring mass loss due to CO2 release. Fermentations were performed in duplicate. When fermentations were completed, samples were taken to assess alcohol content, yeast mass and viability.
Analytical Methods
Alcohol content and pH-value of beer samples were determined using an Alcolyzer Plus with a DMA 5,000 density meter and Xsample 122 sample changer (Anton-Paar GmbH, Ostfildern, Germany). Medium chain fatty acids and medium chain fatty acid esters were determined by gas chromatography with a flame ionization detector (GC-FID) with a 50 m 0.32 mm phenomenex-FFAP-0.25 μm column. The temperature protocol was 1 min 60°C, 3 min 220°C (5°C/min), 8 min 240°C (20°C/min). Detector and injector temperatures were 250 and 200°C, respectively. Fermentation by-products were determined using headspace GC-FID analysis according to Mitteleuropäische Brautechnische Analysenkommision method 2.21.1. Briefly, an INNOWAX cross-linked polyethylene-glycol 60 m × 0.32 mm 0.5 μm column was used. Temperatures of oven, detector and injector were 250, 200, and 150°C, respectively. Injection time was 4 s and analyzing time was 17 min. Turbo-Matrix 40 headspace parameters were: sample temperature, 60°C; transfer temperature, 130°C, needle temperature 120°C. The time for GC-cycle was 22 min, thermosetting was 46 min, pressurization was 1 min and injection time was 0.03 min. A 1.0 mL sample of wort or beer was filtered through Millipore membrane (pore size of 0.45 μm) filters and frozen (−20°C). The samples were thawed and prepared for HPLC, which was used to determine concentrations of fermentable sugars (fructose, glucose, maltose and maltotriose) as well as glycerol in wort and beers. A Waters 2695 Separation Module and Waters System Interphase Module liquid chromatograph coupled with a Waters 2414 differential refractometer (Waters Co., Milford, MA, United States) were used. An Aminex HPX-87H Organic Acid Analysis Column (300 × 7.8 mm, Bio-Rad, United States) was equilibrated with 5 mM sulfuric acid (H2SO4) (Titrisol, Merck, Germany) in water at 55°C. The samples were eluted with 5 mM H2SO4 in water at a 0.3 mL/min flow rate.
Maltotriose Transport Assays
For maltotriose uptake measurement, the yeast strains were grown at 20°C in liquid YP medium containing maltose (4% w/v) or maltotriose (4% w/v) to an OD600 nm between 4 and 8. The cells were harvested by centrifugation (10 min, 5,000 rpm, 0°C), washed twice with ice-cold water and once with 0.1 M tartrate-Tris (pH 4.2) and re-suspended in the same buffer to a concentration of 200 mg/mL fresh yeast. Zero-trans rates of [U-14C]-maltotriose uptake were measured at 20°C essentially as described by Lucero et al. (1997). Briefly, aliquots of 40 μL of yeast suspension were added to 20 μL of 15 mM labeled maltotriose (for a final concentration of 5 mM [U-14C]-maltotriose) and incubated for 60 s at 20°C. The reaction was stopped with the addition of 5 mL ice-cold water. The suspension was quickly filtered and washed with an additional 5 mL of ice-cold water. The filter was submerged in 3.5 mL of Optiphase HiSafe 3 scintillation cocktail (Perkin Elmer, MA, United States) and the radioactivity measured in a Perkin Elmer Tri-carb 2810 TR scintillation counter. [U-14C]-maltotriose (ARC 627) was obtained from American Radiolabeled Chemicals (St. Louis, MO, United States) and re-purified before use as described by Dietvorst et al. (2005). Maltose (minimum purity, 99%) and maltotriose (minimum purity, 95%) were from Sigma-Aldrich (St. Louis, MO). The cell washing was expected to remove any potential extracellular carbohydrate-hydrolase that could interfere with the results. In addition, the maintenance of cells on ice until the uptake assay, the short incubation time in maltotriose, and subpar temperature and pH conditions for known extracellular glucoamylases of the Saccharomyces genus (pHOpt = 4.5–6, TOpt = 40–60°C; Hostinová and Gašperík, 2010) were expected to limit the activity of any residual carbohydrate-hydrolase that might be present.
Sensory Analysis
All beer samples were tasted and judged by a trained sensory panel of 10 panelists certified by the Deutsche Landwirtschafts-Gesellschaft (DLG). Single tasting was performed in a dedicated tasting room (single tasting chambers, white-colored room, no distracting influences, and brown glasses with three-digit number labels) to exclude all external misleading factors. The main flavor impressions were determined at a range from 1 (almost no perception) to 10 (very intense perception). Flavor impressions were chosen according to Meier-Dörnberg et al. (2017a). In addition, a tasting was performed under the same circumstances with the DLG scheme, in which the beer is judged by its aroma, taste, carbonation, body and bitterness in a range of 1–5, 1 being the lowest value (negative) and 5 being the highest value (positive).
Results
Genotype, Cell- and Colony Morphology
S. jurei TUM 629 was isolated from a piece of bark of an ash tree (Fraxinus excelsior) close to the river Isar in Munich, Bavaria, Germany. Single colonies of S. jurei TUM 629 exhibited a homogenous morphology (Figure 1). On WLN-Agar, colonies appear white with a pale green center, and exhibit a well-defined edge and a distinct umbonate morphology. On wort-agar, the colonies are round and white becoming more transparent toward the edge. The colonies of S. jurei TUM 629 differed in colony morphology from the colonies of S. eubayanus C902 and S. pastorianus TUM 34/70 (both of which were later used for the brewing trials as reference strains). The pictured colonies in Figure 1 can be referred to as strain reference pictures for the standard media used in brewing microbiology.

Figure 1. Colony morphologies of S. jurei TUM 629, S. eubayanus C902, S. pastorianus TUM 34/70 on WLN-Agar (Wallerstein-Nutrient-Agar) and Wort-Agar.
The cell morphology of S. jurei in brewer’s wort (12°P pale barley malt wort) is round with single cell-budding of round daughter-cells (dark-field microscopy with scale in Supplementary Figure S1). Cell diameter is between 4 and 8 μm. Vacuoles and cell organelle structures could be observed in the dark field microscopic picture. Cell morphology of S. jurei TUM 629 in wort is different from the cell morphologies of S. pastorianus lager strains and from some S. cerevisiae brewing strains (e.g., wheat beer strains with larger cell diameter and multilateral budding and star cluster formation).
The ITS1-5.8-ITS2 rDNA and D1/D2 units of the 26S rDNA of S. jurei TUM 629 were sequenced and NCBI Blast comparison was carried out (sequences in Supplementary Data). D1/D2 26S rDNA showed 100% sequence identity and ITS1-5.8-ITS2 rDNA 99.38% sequence identity to S. jurei type strain NCYC 3947T (data not shown). Figure 2 gives a clear indication of species identity of TUM 629 by showing the median coverage of 10 kbp windows of seqeuencing reads to a reference genome of 8 Saccharomyces species. A phylogenetic tree based on the Clustal W Alignment of the two S. jurei sequences (TUM 629 and NCYC 3947T) along S. pastorianus and S. eubayanus further supporting the species identity can be found in Supplementary Figures S2, S3.
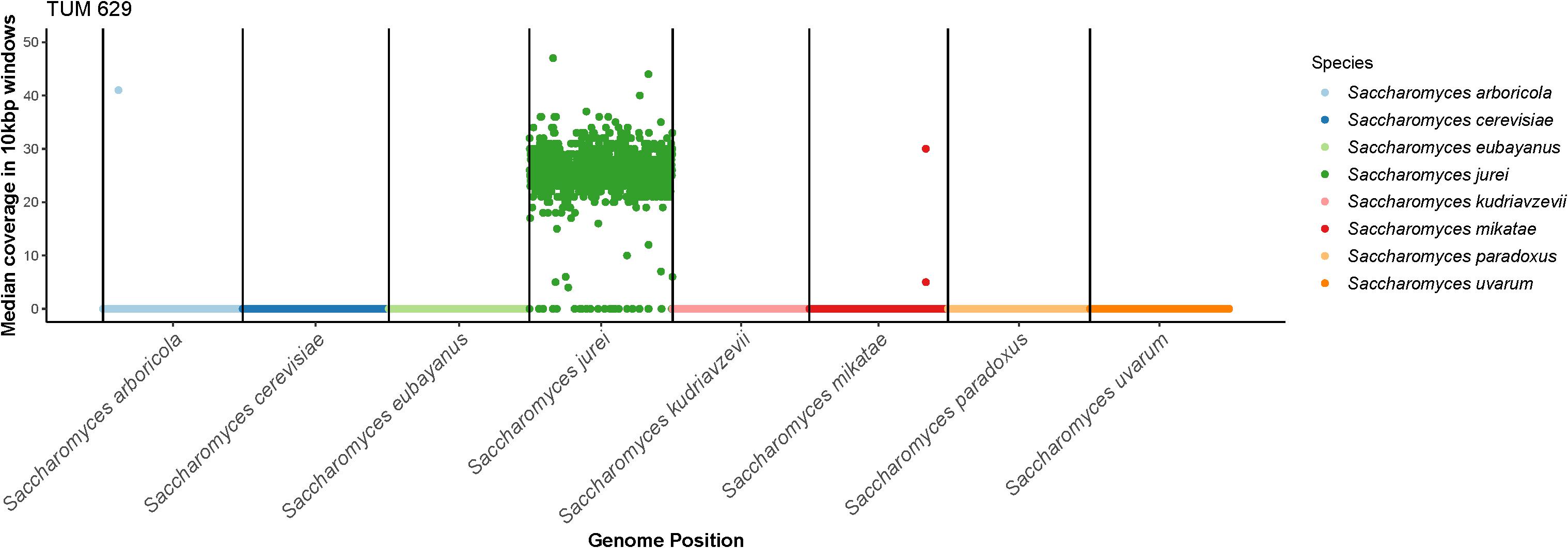
Figure 2. The median coverage in 10 kbp windows of sequencing reads from S. jurei TUM 629 aligned to a concatenated reference genome consisting of 8 species in the Saccharomyces genus. Reads align exclusively to S. jurei, except for two small regions in the sub-telomeric regions of S. arboricola chromosome 4 and S. mikatae chromosome 14. Results were visualized in R using modified scripts from sppIDer (Langdon et al., 2018).
Genome Sequencing and Analysis
To confirm the species-level identification of S. jurei TUM 629, 150 bp paired-end Illumina whole genome sequencing was carried out. S. jurei TUM 629 and VTT C-171003 (=C1003 = NCYC 3947T) were sequenced to an average coverage of 29× and 27×, respectively. A total of 9,690 single nucleotide polymorphisms (SNPs) were detected in TUM 629 compared to the reference genome of S. jurei C1003 (accession number GCA_900290405; Naseeb et al., 2018). Of these SNPs, only 303 were heterozygous, suggesting that TUM 629 is a homozygous diploid. The SNPs in TUM 629 indicate a sequence divergence of about 0.1% relative to the type strain, which was originally isolated in south-eastern France (Naseeb et al., 2017). Sequencing coverage was even across the whole reference genome, suggesting that TUM 629 is euploid (Supplementary Figures S4, S5 showing the coverage for S. jurei C1003). When sequencing reads were aligned to a concatenated reference genome of all Saccharomyces species, reads mapped exclusively to S. jurei, indicating that S. jurei TUM 629 is not an interspecies hybrid (Figure 2). As expected, only 222 SNPs (218 of which were heterozygous) were detected in the re-sequenced S. jurei type strain compared to the same reference genome. Genetic analysis allowed identification of genes potentially encoding maltose and maltotriose transporter proteins (Supplementary Table S1). Three genes ranging from 82.2 to 84.4% identity with S. cerevisiae MAL31 and one with 82.6% identity with S. cerevisiae MAL11 were identified. Additionally, 3 genes were found with 78.9–84% identity to S. cerevisiae IMA5, encoding an extracellular α-glucosidase.
Wort Fermentation
Wort fermentation was carried out to assess the brewing-relevant properties of S. jurei TUM 629. Results were compared with those of the type strain of S. eubayanus (C902), and the S. pastorianus lager strain TUM 34/70 (Figure 3). An initially rapid fermentation rate in the first 2 days was followed by a period of relative inactivity, which lasted for approximately 14 days. After this time alcohol level increased steadily for approximately 30 days, reaching a value of 4.9% (v/v) at 39 days. S. eubayanus likewise exhibited rapid fermentation in the first 2 days, and this was also followed by a lag phase. In contrast to the S. jurei fermentation, this period of inactivity only lasted until 7 days after inoculation before the fermentation rate increased. Fermentation was mostly complete after 18 days, reaching a maximum of 4.4% (v/v) alcohol. As expected, fermentation with the lager strain was rapid, with no evidence of a lag phase at any stage. 5.5% (v/v) alcohol was achieved after 9 days with this strain.
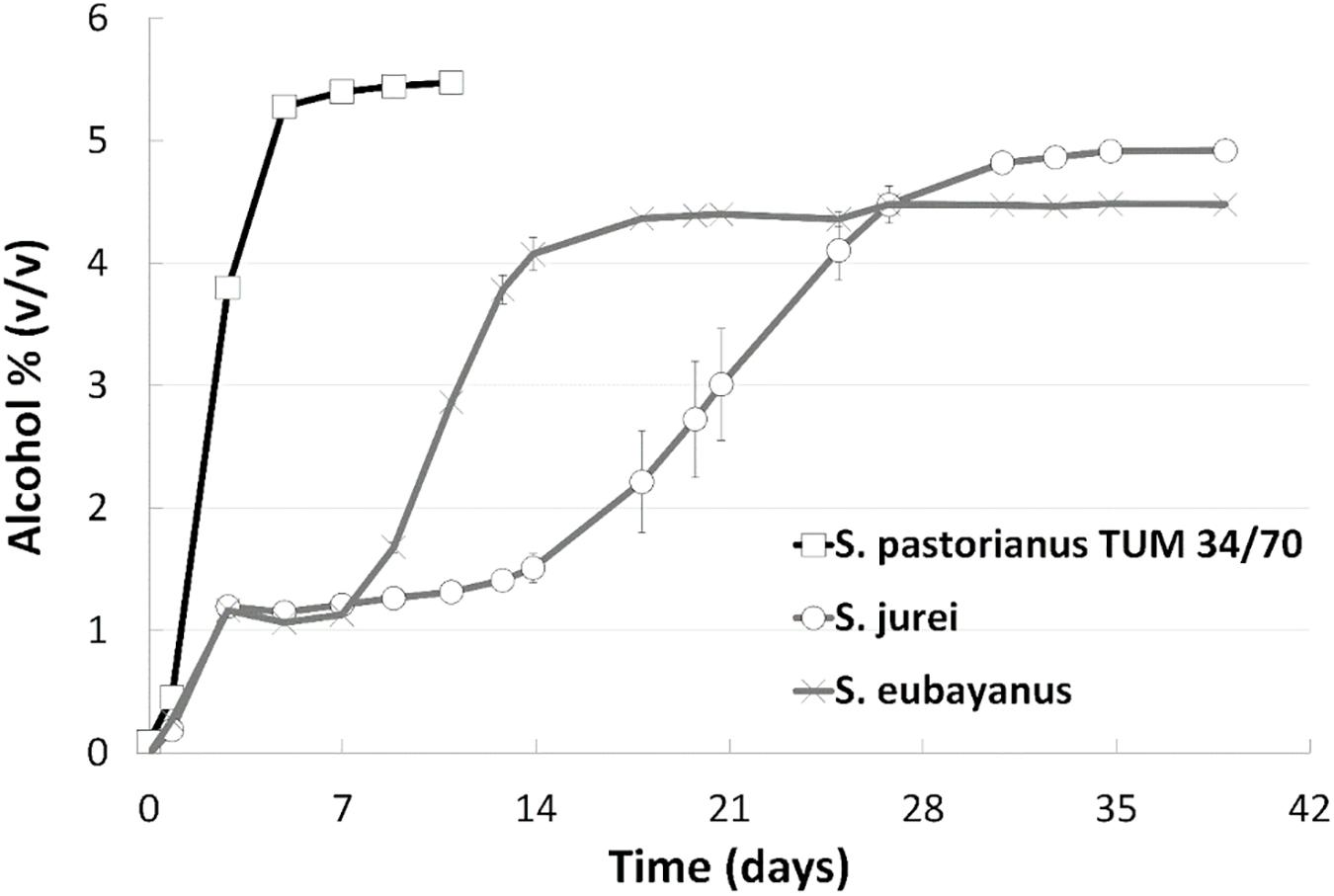
Figure 3. Fermentation progress as monitored by alcohol level (v/v) at 15°C during 39 days of fermentation of an all-malt 12°P wort. Strains include one lager reference strain (TUM 34/70), as well as the German strain of S. jurei (TUM 629), and the type strain of S. eubayanus (C902). Values are means from duplicate fermentations and error bars indicate range.
Analysis of residual sugars indicated that maltotriose consumption was a determining factor for the extent of fermentation (Figure 4). Approximately 50% of the available maltotriose was consumed by S. jurei. The corresponding depletion in the beers fermented with the lager strain was 85%. S. eubayanus, as expected, had no effect on maltotriose concentration. Maltose was completely consumed by the lager strain and by S. eubayanus, though interestingly a small portion (3.6 g/L of initially 52.5 g/L) remained in the S. jurei beer. An increase in glycerol after fermentation was observed for all strains. This was highest in the wild yeast strains both of which produced 2.9 g/L of glycerol while the corresponding concentration in the lager strain beer was 1.6 g/L. The initial concentration of glycerol in the wort was 1.05 g/L.
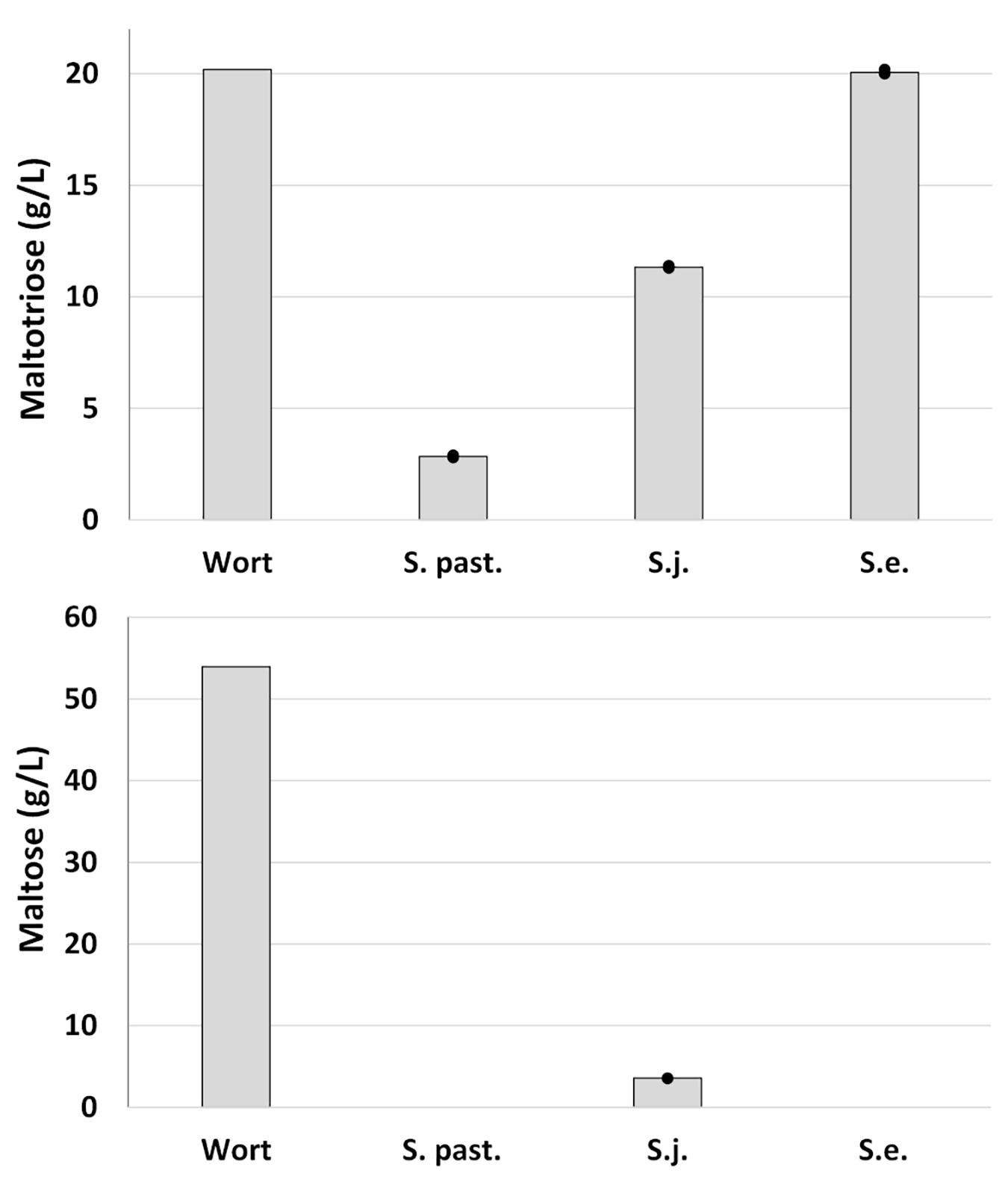
Figure 4. Maltose and Maltotriose concentrations in all-malt 12°P wort, and beer after fermentation at 15°C for up to 39 days. Strains include one lager reference strain (TUM 34/70), as well as the German strain of S. jurei (TUM 629), and the type strain of S. eubayanus (C902). Dots represent the individual fermentations and bars show the mean of the duplicates.
Characteristics of the finished beers such as their degree of attenuation, diacetyl, acetaldehyde content and pH are compiled in Table 2. The beers brewed with S. jurei and S. eubayanus do not exhibit an increased content of diacetyl compared to the established S. pastorianus strain. S. eubayanus did cause an increased concentration of acetaldehyde in the beer. The attenuation of each strain corresponds well to the aforementioned use of the wort sugars maltose and maltotriose and a high degree of attenuation with the well-adapted lager strain S. pastorianus compared to the wild yeast strains.
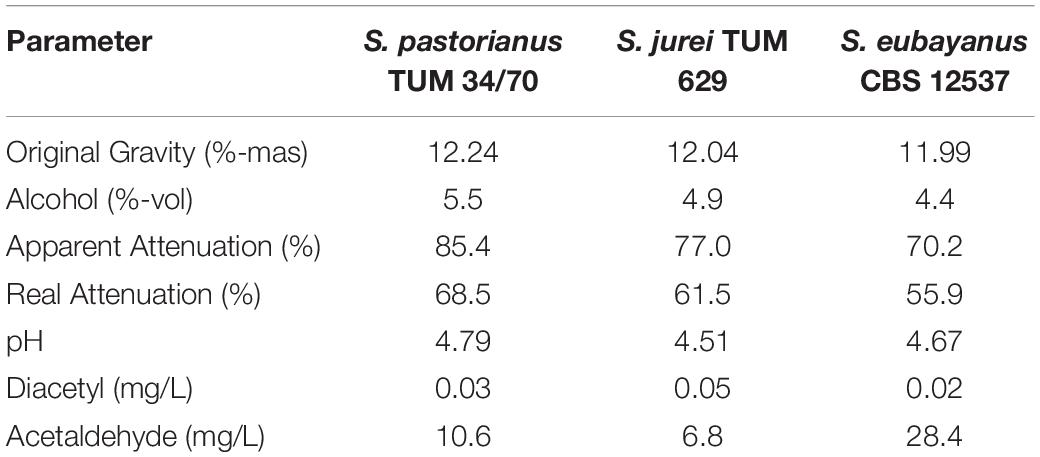
Table 2. Analysis of finished beers fermented by S. pastorianus (TUM 34/70), S. jurei (TUM 629) and S. eubayanus (CBS 12537) at a fermentation temperature of 15°C.
Maltotriose Uptake
To assess the yeast’s ability to take up maltotriose directly from the medium, uptake activities were quantified using radiolabeled maltotriose. When using maltose as propagation substrate, both S. jurei strains and S. mikatae showed maltotriose uptake activities ≤ 0.5 μmol min–1 g–1 DY–1 indicating the absence of active maltotriose transporters in the plasma membrane (Table 3). An uptake activity ≤ 0.5 μmol min–1 g–1 DY is considered negligible. S. eubayanus C902 showed an activity level slightly above 0.5 μmol min–1 g–1 DY, although it is known that this strain lacks any capacity to transport maltotriose (Gibson et al., 2013; Magalhães et al., 2016). S. pastorianus was the only species showing enough maltotriose uptake activity to ensure its consumption from the wort. The fermentation data, however, showed that S. jurei can consume maltotriose (Figure 4). If the maltotriose is taken up by the yeast cells then the transporters are either susceptible to repression by maltose or its expression requires induction by maltotriose. To validate this hypothesis the maltotriose uptake activity was also measured in cells propagated in YP medium with maltotriose as sole carbon source. Growth on maltotriose was slow, and it took 6 days to reach an OD600 value greater than 4, however, after this maltotriose exposure, the cells could grow much faster when transferred to fresh medium containing maltotriose as sole carbon source (data not shown). Uptake activity from cells grown on maltotriose confirmed that there is an active transmembrane transport occurring in S. jurei. This mechanism appears to require prolonged exposure to maltotriose, or absence of other sugars.
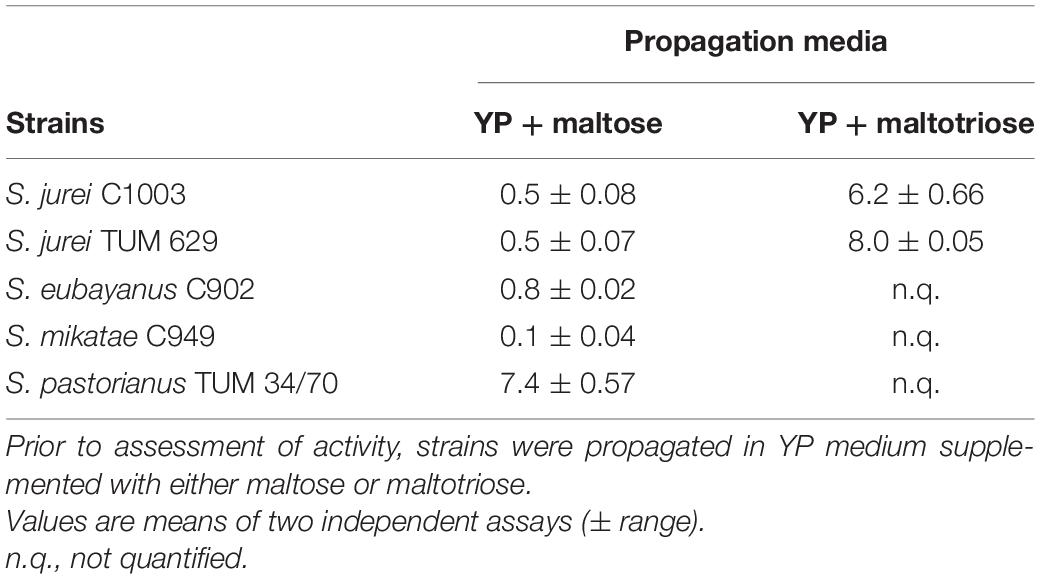
Table 3. Zero-trans rates of maltotriose uptake activity (μmol min–1 g–1 DY) of the strains in study, measured at 20°C.
Analytical and Sensory Aroma Profile
The beers produced by the three different yeast strains S. pastorianus TUM 34/70, S. eubayanus C902 and the newly isolated S. jurei TUM 629 showed significantly different sensory and analytical aroma profiles (Tables 4, 5). The sample fermented by S. pastorianus TUM 34/70 had the overall highest concentration of esters (Table 4). Of all esters detected in this sample, ethyl acetate was found at the highest concentration at 42.1 mg/L. The overall highest concentration of 3-methylbutylacetate (iso-amyl acetate) was 3.6 mg/L, and was also found in the S. pastorianus beer. Ethyl hexanoate concentration was four times higher in the sample fermented with S. jurei TUM 629 (0.40 mg/L) compared to the samples fermented with S. pastorianus (0.13 mg/L) and S. eubayanus CBS 12537 (0.11 mg/L). Ethyl hexanoate (apple, fruity flavor) has a flavor threshold of 0.23 mg/L in beer according to Meilgaard (1975a). Overall, S. jurei produced higher amounts of medium chain fatty acid esters and relatively high amounts of higher alcohols in comparison to the other two yeast strains but was outperformed in the production of acetate esters by the S. pastorianus strain, and in higher alcohol production by the S. eubayanus strain (Tables 4, 5). S. eubayanus produced a typically high amount of phenyl ethanol (121.4 mg/L) in comparison to the other two yeast strains (Gibson et al., 2013). All samples showed diacetyl levels below 0.1 mg/L and no 2,3-pentanedione (data not shown).
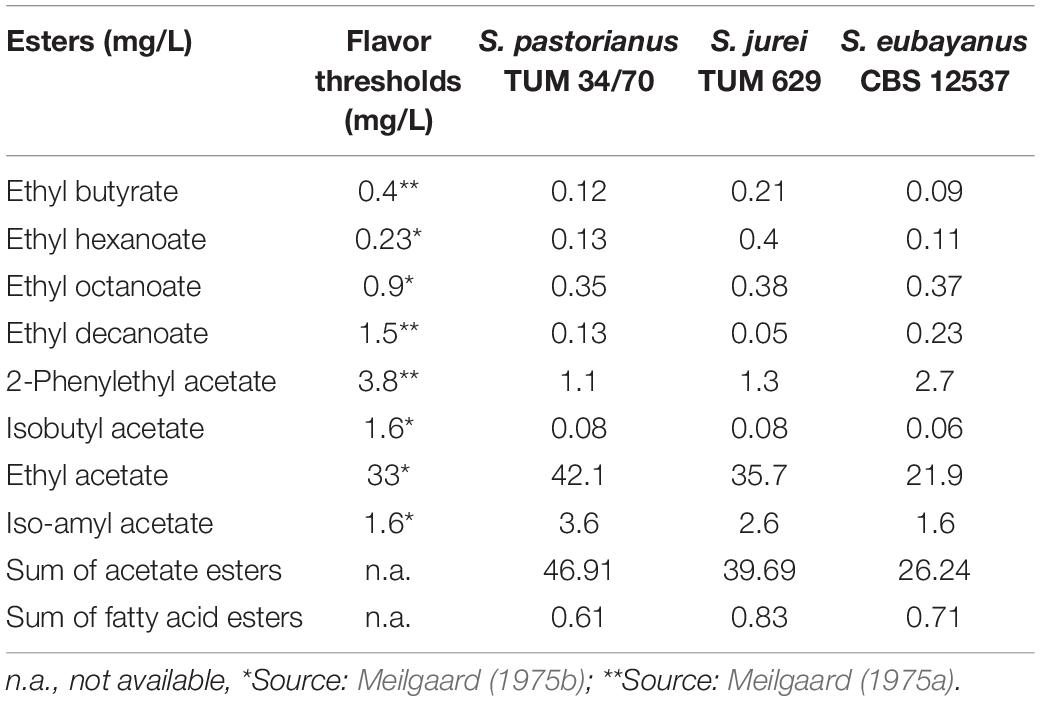
Table 4. Esters detected in the three different beer samples fermented by S. pastorianus (TUM 34/70), S. jurei (TUM 629) and S. eubayanus (CBS 12537) at a fermentation temperature of 15°C and an original gravity°12 P.
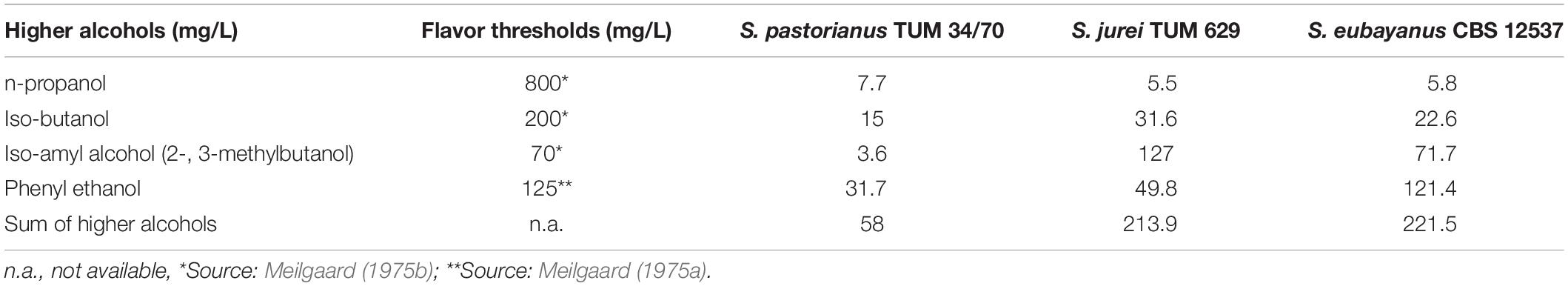
Table 5. Higher alcohols detected in the three different beer samples fermented by S. pastorianus (TUM 34/70), S. jurei (TUM 629) and S. eubayanus (CBS 12537) at a fermentation temperature of 15°C and an original gravity 12°P.
The trained panel rated all beers as high quality in the DLG scheme and found no significant difference of the purity of aroma and taste, and the quality of carbonation, body and bitterness among the three samples (data not shown). However, they noted clear differences in the descriptive analysis (Figure 5). A phenolic spice/clove note was prominent in the sample fermented by the S. jurei strain TUM 629. A significant apple flavor was detected, which can be related to the considerably higher amount of ethyl hexanoate (Table 4). The clove-like flavor was less pronounced in the sample fermented by S. eubayanus, despite the analytical levels of 4-VG being similar (S. eubayanus: 0.16 mg/L, S. jurei: 0.15 mg/L). The sample fermented by S. pastorianus TUM 34/70 was described as very fruity and berry like, which can partly be explained by generally higher acetate ester concentration (46.91 mg/L) in the sample in comparison to the other two samples (S. eubayanus 26.24 mg/L and S. jurei 39.69 mg/L) (Table 4). Results indicated considerable potential for S. jurei application in brewing, and prompted a comparative study including the type strain of S. jurei and a number of other Saccharomyces type strains.
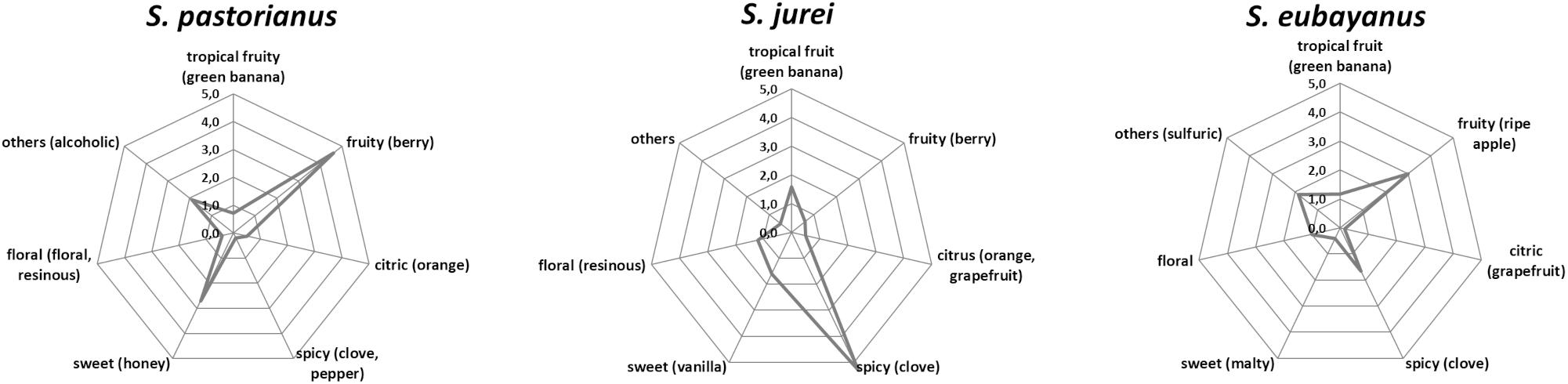
Figure 5. Tasting results of the beer samples fermented by S. pastorianus TUM 34/70, S. jurei TUM 629, and S. eubayanus at 15°C fermentation temperature and an original gravity of 12°P.
Screening of Saccharomyces Type Strains for Wort Fermentation Potential
Trial fermentations conducted with 12°P wort at 15°C revealed clear differences between the strains in terms of wort fermentation potential (Figure 6). As expected, the two reference strains fermented rapidly, with no evidence of an extended lag phase. Alcohol yield was good for both strains, 5.8% ABV for S. pastorianus and 6.1% ABV for S. cerevisiae (Figure 6). The greater efficiency of the A62 ale strain is due to a strong maltotriose fermentation capacity which has also been observed in previous studies (Krogerus et al., 2018).
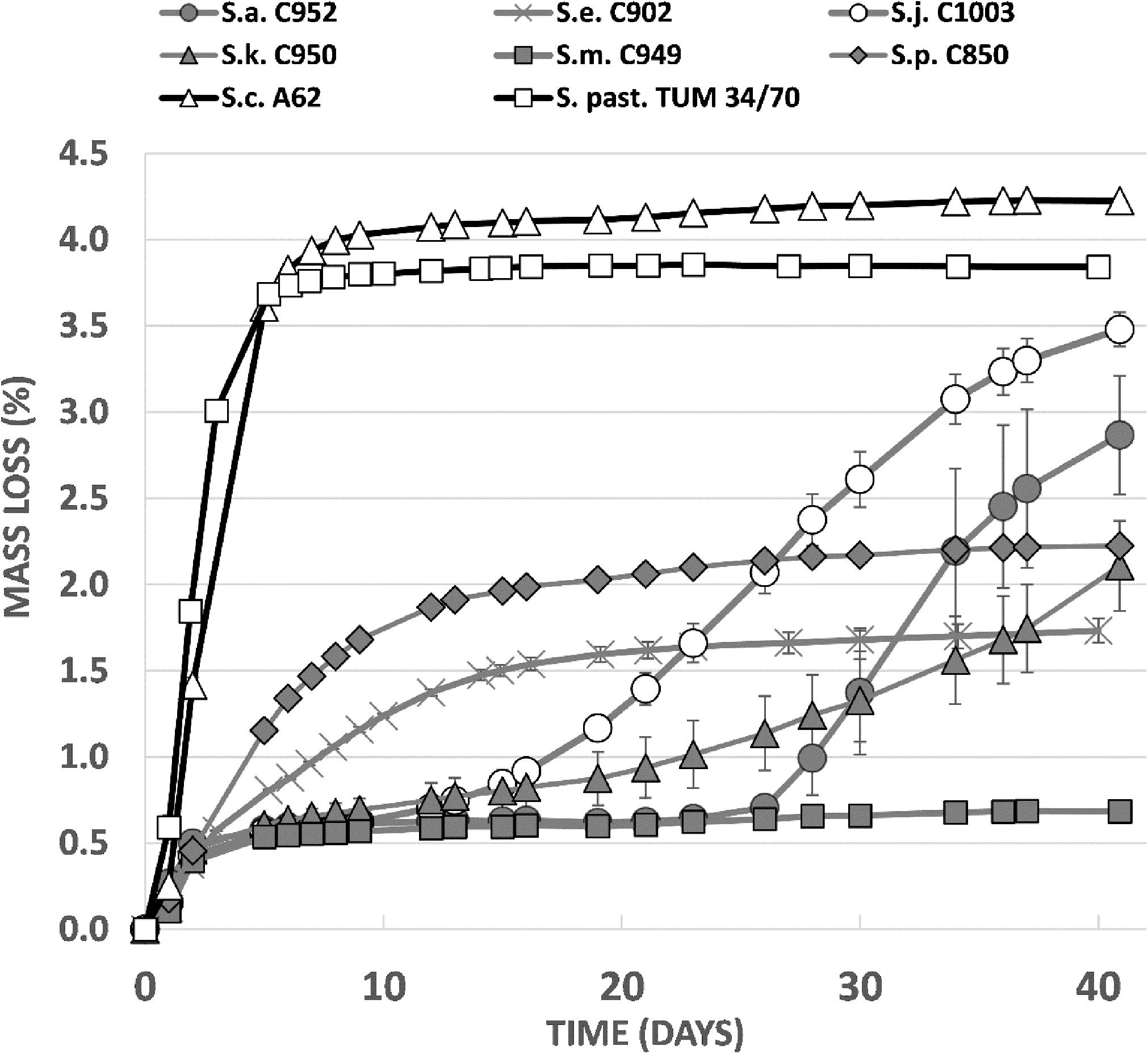
Figure 6. Fermentation performance (measured as loss of mass of the fermentor) in comparison between Saccharomyces type strains in 12°P all-malt wort at 15°C.
Fermentation characteristics of the Saccharomyces type strains were highly variable (Figure 6). All strains exhibited a relatively rapid fermentation in the first 2 days after inoculation, presumably due to utilization of the simple sugars present in the wort. S. mikatae fermentation appeared to cease after this initial period, while S. arboricola, which had an identical fermentation profile until 26 days after inoculation, appeared to start fermenting and was still actively fermenting after 41 days when it had produced an ABV level of 4.5%. S. kudriavzevii fermentations were likewise characterized by an initially slow fermentation rate, which increased over time, giving a final value of 3.6% ABV. S. paradoxus and S. eubayanus, did not exhibit any lag phase in fermentation, but overall fermentation efficiency was limited with the strains achieving 3.6 and 3.3% ABV after 41 days (Figure 7). S. jurei, despite an initially low fermentation rate in the first 2 weeks of fermentation was able to achieve an ABV of 5.3%, a value considerably higher than those found in the other wild yeast beers.
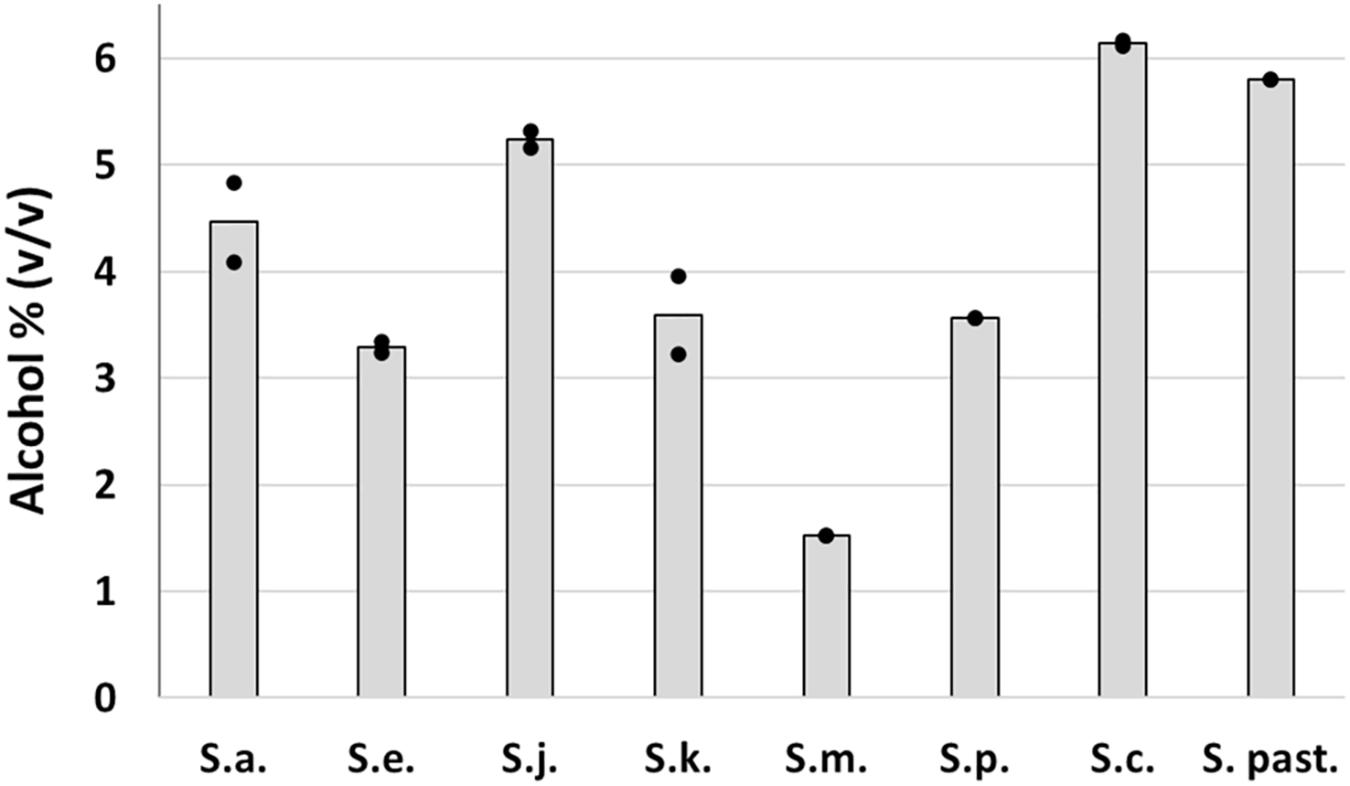
Figure 7. Final alcohol content of fermentations conducted in 12°P brewer’s wort at 15°C with Saccharomyces type strains. Dots represent the individual fermentations and bars show the mean of the duplicates.
Discussion
With the exception of recent reports on the brewing potential of two wild Saccharomyces species, S. eubayanus and S. paradoxus (Gibson et al., 2013; Cubillos et al., 2019; Nikulin et al., 2020), little is known about the brewing potential of the wild species within the Saccharomyces genus. Wild Saccharomyces species are not typically isolated from brewery fermentation environments, and when encountered in brewing, or other fermentation environments, typically occur in the form of interspecies hybrids. This has been seen for example with S. kudriavzevii, which occurs in a S. cerevisiae × S. kudriavzevii hybrid form in Belgian ales (Gallone et al., 2019), but is not otherwise isolated from brewing systems. Strains of S. cerevisiae var. diastaticus are common brewery contaminants (Meier-Dörnberg et al., 2018), but their designation as ‘wild’ species is questionable considering they group phylogenetically with one of the major ale yeast clades (Krogerus and Gibson, 2020; Pontes et al., 2020). The absence of wild yeast species in fermentation environments is indicative of deficiencies in performance. However, the successful application of S. eubayanus in brewing (Gibson et al., 2017) has demonstrated how such strains, when handled appropriately, may act as efficient fermenters, and can support product differentiation. Creative utilization of wild species may help brewers meet the consumer demand for beers with novel flavor profiles and interest in engaging background narratives.
The ability of S. jurei to utilize wort maltotriose was surprising given that this trait has not been observed previously in wild Saccharomyces yeasts and is generally considered to be a trait associated only with domesticated yeasts (Gallone et al., 2018). There are two mechanisms by which Saccharomyces yeast may utilize maltotriose (Krogerus et al., 2019). In the first, maltotriose is taken up directly across the cell membrane and hydrolyzed to glucose monomers through the action of an intracellular maltase. Active uptake is mediated by transporters such as Agt1 and Mtt1, which are also responsible for the uptake of maltose (Dietvorst et al., 2005; Vidgren and Londesborough, 2012). Despite the absence of the property in wild species, maltotriose utilization appears to be evolvable, with two separate studies demonstrating the creation of maltotriose transporters from existing maltose transporters via a series of recombination events (Baker and Hittinger, 2019; Brouwers et al., 2019). A second mechanism for maltotriose utilization involves extracellular degradation of maltotriose by “diastatic” strains of yeast. This is seen in some S. cerevisiae strains belonging, in particular, to the Beer 2 group (Gallone et al., 2016). In many of these strains, glucoamylase activity is responsible for the liberation of glucose and maltose from maltotriose, as well as dextrins and soluble starch, and obviates the requirement for transmembrane maltotriose transport (though this can still be present) (Krogerus and Gibson, 2020). Our assessment here of the uptake of radiolabeled maltotriose demonstrates that transmembrane transport occurs across cell membranes of S. jurei. Maltotriose uptake was only apparent after cells had been propagated on maltotriose, indicating that the expression of the phenotype (either through gene expression or protein configuration or localization) is either limited by the presence of other sugars or requires induction by maltotriose. Known maltotriose transporters such as Agt1 and Mtt1 can carry both maltose and maltotriose and do not require specific conditions for their expression (Magalhães et al., 2016). The mechanisms by which maltotriose is consumed by S. jurei require further investigation. Nikulin et al. (2020) in a study of brewing potential in S. paradoxus, noted that the efficient uptake of maltose was influenced by the growth medium, with growth on glucose leading to an extended lag phase prior to maltose use during fermentation. This lag phase was significantly reduced if yeasts were propagated on maltose. These results, and our observation that maltotriose use requires previous exposure to this sugar, suggest that brewing with S. jurei would benefit from carefully regulated propagation conditions. Further research is required to investigate application parameters such as pitching rate, aeration, wort composition and propagation strategies. Michel et al. (2016) describe a screening system for potential brewing yeasts, which could also be applied to further explore the characteristics of S. jurei. S. jurei’s long fermentation, as demonstrated in this study, puts it at a disadvantage compared to established brewing yeasts. In a study researching the brewing potential of S. paradoxus strains isolated from environmental samples, Nikulin et al. (2020) also demonstrated long fermentations with an extended lag phase and maltose uptake being dependent on pre-conditioning the yeast through growth on maltose. With industrial yeasts being adapted to maltose and maltotriose use, the observed lag phase only causes a short delay, if at all, in fermentation compared to environmental isolates.
The presence of an active trans-membrane-transport system does not exclude the possibility of S. jurei also hydrolyzing maltotriose extracellularly. Genes encoding both potential maltose/maltotriose transporters and extracellular α-glucosidases were found within the S. jurei genome. It is expected that a more thorough knowledge of the maltotriose utilization mechanisms may help to improve the potential brewing performance of S. jurei, both in terms of maltotriose utilization efficiency and duration of lag phase.
While brewing efficiency is a critically important trait for brewers, the contribution of yeast to beer flavor is more directly relevant for consumers. A new brewing strain should preferably offer a novel sensorial experience to beer drinkers. In this regard, the concentration of yeast-derived volatile aroma compounds is significant. The three strains included in the brewing trials had distinct flavor profiles, with each producing a high concentration of at least one important flavor-active compound. In the case of S. jurei, this was ethyl hexanoate, a generally desirable ethyl ester with an apple, cherry or aniseed aroma. The high concentration may have contributed to the tropical fruit notes detected by the sensory panel. In contrast to S. jurei, the dominant flavor volatile in the S. pastorianus beer was 3-methylbutylacetate. This is a highly desirable flavor compound in commercial brewing and contributes a banana or pear aroma to beer (Meilgaard, 1975a). In the S. eubayanus beer, the typically rose-like phenylethanol was prominent. This has previously been noted for this strain (Gibson et al., 2013). Phenylethanol has previously been found to mask the perception of other flavor compounds (Bamforth, 2020) and may explain the relatively low perception of phenolic flavor notes in the S. eubayanus beer relative to the S. jurei beer.
The discovery of relatively good fermentation efficiency in S. jurei inspired a direct comparison with fermentation performance in other members of the strain. This comparative study included an ale S. cerevisiae strain, a lager S. pastorianus strain and six type strains of wild Saccharomyces species. The only species excluded was S. uvarum, the type strain of which was shown previously to be strongly maltose-positive, but maltotriose-negative (Nikulin et al., 2018). The species included were highly variable with respect to fermentation characteristics. Relative to the reference strains, the wild strains tended to have long lag periods following the initial fermentation of monosaccharides, and when fermentation increased, it was at a relatively low rate and often limited in extent. Given these features, it is unsurprising that wild Saccharomyces species might be at a competitive disadvantage compared to domesticated yeasts, and thus rarely encountered in wort fermentations. Performance varied however, not just between brewing yeasts and wild yeasts, but also amongst the wild species. As observed previously (Nikulin et al., 2018), S. mikatae had only limited fermentation capability, with apparently no ability to metabolize maltose. Other species appeared to be able to utilize this disaccharide, albeit at different rates and after different periods of adaptation following the initial fermentation. Of note was the high level of alcohol production by the type strain of S. jurei, suggesting that a superior fermentation efficiency due to maltotriose utilization is not restricted to the Bavarian strain included in the previous fermentation trials.
As well as the association between woodland habitats and yeast, there has been traditionally a strong association between wood-based materials and brewing. Wood, in addition to being the main building material for millennia, has also been used for vessels used in food production and storage. The usage of wooden materials in brewing was not only limited to vessels, it also made up tools and additives for beer production. Oak wood was used for vats, casks and barrels; chips or shavings of hornbeam (Carpinus betulus) and hazel (Coryllus avellana) have been used as clarifying aids; and spruce and birch were commonly used for making barrel bungs (Schnegg, 1921, 1922). Schnegg, in his textbook about microscopy for brewers, describes oak wood as being hard but porous, with the pores being ideal for harboring microorganisms and residues of beer (Schnegg, 1921). Ash and oak trees have also been described as holy trees in Indo-Germanic culture (Dumont, 1992). Oaks have served as an inoculum for roman wine fermentations and ash trees were often tapped for their tree sap (Feier et al., 2019). Through its unique structure and capability of harboring microorganisms in nature, wood could potentially transfer yeasts from natural habitats to human made fermentations and thereby initiate the process of domestication. Due to the role of both ash and oak and their proven association with yeasts capable of fermenting cereal-based worts, it is not unlikely that these trees have served as an inoculum for fermentation throughout history. While many isolates of the Saccharomyces species have been associated with oak trees or other members of the order of Fagales (containing oak trees, beeches, Nothofagus spp. and aforementioned Castanopsis), F. excelsior does not belong to this order. No yeasts other than S. jurei TUM 629 were isolated from the sampled ash tree and little is known about the specific microbiome of ash trees. Other studies have revealed evidence for the presence of S. jurei in different habitats (Alsammar et al., 2019). The seemingly low abundance of S. jurei in nature may be caused by the inadequacy of culturing methods for isolating species that are present at low numbers in the sample. However, DNA-based methods of describing complex populations (metagenomics) may also run the risk of biases in this situation (Kebschull and Zador, 2015). Recently, another strain of S. jurei has been isolated from the bark of an ash in the black forest, Germany (current study, data not published). More research is needed to assess the role of S. jurei in the ecology of yeasts, its preferred habitats and geographical distribution.
The use of interspecies hybridization to fully exploit the biotechnological potential of wild yeast has been demonstrated in a number of studies, and may be instrumental in overcoming the limitations of S. jurei as a fermentative organism. It is our understanding that S. cerevisiae × S. jurei hybrids have already been created and characterized with respect to brewing potential, and that this work, led by Prof. Daniela Delneri at the University of Manchester, United Kingdom, in collaboration with Cloudwater Brew Co, United Kingdom, and Foundation Edmund Mach, Italy, will be published shortly (D. Delneri, pers comm; doi: 10.1101/2021.01.08.425916).
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author Contributions
TK: fermentation trials and analysis of fermentation data. FM: transmembrane transport assays. KK: whole-genome analyses. BG: first draft and conceptualization. MM: sensory and aroma analysis on the bottled beers, first draft, and conceptualization. MH: first draft, isolation protocol, species identification, physiological characterization, and conceptualization. OK: strain isolation and finalization of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Academy of Finland project 305453. TK’s work was supported by PBL Brewing Laboratory. This research was funded by the Wifö (Wissenschaftsförderung der Deutschen Brauwirtschaft e.V., Berlin, Germany) in the project AiF 20658 N.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Eero Mattila and Niklas Fred for assistance in the VTT Pilot Brewery, and Aila Siltala for skilled technical assistance. We thank Henrik Siegumfeldt (University of Copenhagen) for supervising the work of TK. We also thank Franziska Elisath (BLQ) for assistance with molecular biological sample preparation and analyses.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.645271/full#supplementary-material
Footnotes
- ^ http://culturecollection.vtt.fi
- ^ https://www.blq-weihenstephan.de/en/tum-yeast/yeast-and-bacteria/
References
Alsammar, H. F., Naseeb, S., Brancia, L. B., Gilman, R. T., Wang, P., and Delneri, D. (2019). Targeted metagenomics approach to capture the biodiversity of Saccharomyces genus in wild environments. Environ. Microbiol. Rep. 11, 206–214. doi: 10.1111/1758-2229.12724
Baker, E. P., and Hittinger, C. T. (2019). Evolution of a novel chimeric maltotriose transporter in Saccharomyces eubayanus from parent proteins unable to perform this function. PLoS Genetics 15:e1007786. doi: 10.1371/journal.pgen.1007786
Bamforth, C. W. (2020). The horace brown medal. Forever in focus: researches in malting and brewing sciences. J. Inst. Brew. 126, 4–13. doi: 10.1002/jib.594
Bing, J., Han, P. J., Liu, W. Q., Wang, Q. M., and Bai, F. Y. (2014). Evidence for a Far East Asian origin of lager beer yeast. Curr. Biol. 24, R380–R381. doi: 10.1016/j.cub.2014.04.031
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Brouwers, N., Gorter de Vries, A. R., van den Broek, M., Weening, S. M., Elink Schuurman, T. D., Kuijpers, N. G. A., et al. (2019). In vivo recombination of Saccharomyces eubayanus maltose-transporter genes yields a chimeric transporter that enables maltotriose fermentation. PLoS Genetics 15:e1007853. doi: 10.1371/journal.pgen.1007853
Cingolani, P., Platts, A., Wang, L. L., Coon, M., Nguyen, T., Wang, L., et al. (2012). A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff. Fly 6, 80–92. doi: 10.4161/fly.19695
Cubillos, F. A., Gibson, B., Grijalva-Vallejos, N., Krogerus, K., and Nikulin, J. (2019). Bioprospecting for brewers: exploiting natural diversity for naturally diverse beers. Yeast 36, 383–398. doi: 10.1002/yea.3380
Dietvorst, J., Londesborough, J., and Steensma, H. Y. (2005). Maltotriose utilization in lager yeast strains: MTT1 encodes a maltotriose transporter. Yeast 22, 775–788. doi: 10.1002/yea.1279
Engel, S. R., Dietrich, F. S., Fisk, D. G., Binkley, G., Balakrishnan, R., Costanzo, M. C., et al. (2014). The reference genome sequence of Saccharomyces cerevisiae: then and now. G3: Genes| Genomes| Genetics 4, 389–398. doi: 10.1534/g3.113.008995
Feier, I., Migała, A., Pietruszka, M., and Jackowski, M. (2019). Roman wine in Barbaricum. preliminary studies on ancient wine recreation. Heritage 2, 331–338. doi: 10.3390/heritage2010022
Gallone, B., Mertens, S., Gordon, J. L., Maere, S., Verstrepen, K. J., and Steensels, J. (2018). Origins, evolution, domestication and diversity of Saccharomyces beer yeasts. Curr. Opin. Biotechnol. 49, 148–155. doi: 10.1016/j.copbio.2017.08.005
Gallone, B., Steensels, J., Mertens, S., Dzialo, M. C., Gordon, J. L., Wauters, R., et al. (2019). Interspecific hybridization facilitates niche adaptation in beer yeast. Nat. Ecol. Evol. 3, 1562–1575. doi: 10.1038/s41559-019-0997-9
Gallone, B., Steensels, J., Prahl, T., Soriaga, L. B., Saels, V., Herrera-Malaver, B., et al. (2016). Domestication and divergence of Saccharomyces cerevisiae beer yeasts. Cell 166, 1397–1410.e1316.
Garrison, E., and Marth, G. (2012). Haplotype-Based Variant Detection From Short-Read Sequencing. Available online at: https://ui.adsabs.harvard.edu/abs/2012arXiv1207.3907G (accessed July 01, 2012).
Gibson, B. (2015). “Non-conventional yeast as a new tool for beer flavour modification,” in Proceedings of the 35th European Brewing Convention Congress, (Porto).
Gibson, B., Geertman, J.-M. A., Hittinger, C. T., Krogerus, K., Libkind, D., Louis, E. J., et al. (2017). New yeasts—new brews: modern approaches to brewing yeast design and development. FEMS Yeast Res. 17:fox038. doi: 10.1093/femsyr/fox038
Gibson, B. R., Storgards, E., Krogerus, K., and Vidgren, V. (2013). Comparative physiology and fermentation performance of Saaz and Frohberg lager yeast strains and the parental species Saccharomyces eubayanus. Yeast 30, 255–266. doi: 10.1002/yea.2960
Hostinová, E., and Gašperík, J. (2010). Yeast glucoamylases: molecular-genetic and structural characterization. Biologia 65, 559–568. doi: 10.2478/s11756-010-0077-8
Hutzler, M. (2009). Entwicklung und Optimierung von Methoden zur Identifizierung und Differenzierung von getränkerelevanten Hefen. Ph.D, dissertation, TU München, München.
Hutzler, M. (2010). Getränkerelevante Hefen–Identifizierung und Differenzierung. Saarbrücken: SVH-Verlag.
Hutzler, M., Koob, J., Riedl, R., Schneiderbanger, H., Mueller-Auffermann, K., and Jacob, F. (2015). “5–Yeast identification and characterization,” in Brewing Microbiology, ed. A. E. Hill (Oxford: Woodhead Publishing), 65–104.
Kebschull, J. M., and Zador, A. M. (2015). Sources of PCR-induced distortions in high-throughput sequencing data sets. Nucleic Acids Res. 43:e143. doi: 10.1093/nar/gkv717
Krogerus, K., and Gibson, B. (2020). A re-evaluation of diastatic Saccharomyces cerevisiae strains and their role in brewing. Appl. Microbiol. Biotechnol. 104, 3745–3756. doi: 10.1007/s00253-020-10531-0
Krogerus, K., Holmström, S., and Gibson, B. (2018). Enhanced Wort fermentation with De Novo lager hybrids adapted to high-ethanol environments. Appl. Environ. Microbiol. 84, e02302–e02317. doi: 10.1128/aem.02302-17
Krogerus, K., Magalhães, F., Kuivanen, J., and Gibson, B. (2019). A deletion in the STA1 promoter determines maltotriose and starch utilization in STA1+ Saccharomyces cerevisiae strains. Appl. Microbiol. Biotechnol. 103, 7597–7615. doi: 10.1007/s00253-019-10021-y
Krogerus, K., Magalhães, F., Vidgren, V., and Gibson, B. (2017). Novel brewing yeast hybrids: creation and application. Appl. Microbiol. Biotechnology 101, 65–78. doi: 10.1007/s00253-016-8007-5
Kurtzman, C. P., Boekhout, T., Robert, V., Fell, J. W., and Deak, T. (2003). “3–Methods to identify yeasts,” in Yeasts in Food, eds T. Boekhout and V. Robert (Sawston: Woodhead Publishing), 69–121.
Kurtzman, C. P., and Robnett, C. J. (1998). Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Van Leeuwenhoek 73, 331–371. doi: 10.1023/A:1001761008817
Langdon, Q. K., Peris, D., Kyle, B., and Hittinger, C. T. (2018). sppIDer: a species identification tool to investigate hybrid genomes with high-throughput sequencing. Mol. Biol. Evol. 35, 2835–2849. doi: 10.1093/molbev/msy166
Li, H., and Durbin, R. (2009). Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 25, 1754–1760. doi: 10.1093/bioinformatics/btp324
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079. doi: 10.1093/bioinformatics/btp352
Libkind, D., Hittinger, C. T., Valério, E., Gonçalves, C., Dover, J., Johnston, M., et al. (2011). Microbe domestication and the identification of the wild genetic stock of lager-brewing yeast. Proc. Natl. Acad. Sci. U.S.A. 108, 14539–14544. doi: 10.1073/pnas.1105430108
Liti, G., Barton, D. B., and Louis, E. J. (2006). Sequence diversity, reproductive isolation and species concepts in Saccharomyces. Genetics 174, 839–850. doi: 10.1534/genetics.106.062166
Liti, G., Nguyen Ba, A. N., Blythe, M., Müller, C. A., Bergström, A., Cubillos, F. A., et al. (2013). High quality de novo sequencing and assembly of the Saccharomyces arboricolus genome. BMC Genomics 14:69. doi: 10.1186/1471-2164-14-69
Lucero, P., Peñalver, E., Moreno, E., and Lagunas, R. (1997). Moderate concentrations of ethanol inhibit endocytosis of the yeast maltose transporter. Appl. Environ. Microbiol. 63, 3831–3836. doi: 10.1128/AEM.63.10.3831-3836.1997
Magalhães, F., Calton, A., Heiniö, R.-L., and Gibson, B. (2021). Frozen-dough baking potential of psychrotolerant Saccharomyces species and derived hybrids. Food Microbiol. 94:103640. doi: 10.1016/j.fm.2020.103640
Magalhães, F., Vidgren, V., Ruohonen, L., and Gibson, B. (2016). Maltose and maltotriose utilisation by group I strains of the hybrid lager yeast Saccharomyces pastorianus. FEMS Yeast Res. 16:fow053. doi: 10.1093/femsyr/fow053
Mardones, W., Villarroel, C. A., Krogerus, K., Tapia, S. M., Urbina, K., Oporto, C. I., et al. (2020). Molecular profiling of beer wort fermentation diversity across natural Saccharomyces eubayanus isolates. Microb. Biotechnol. 13, 1012–1025. doi: 10.1111/1751-7915.13545
Meier-Dörnberg, T., Hutzler, M., Michel, M., Methner, F.-J., and Jacob, F. (2017a). The importance of a comparative characterization of Saccharomyces cerevisiae and Saccharomyces pastorianus strains for brewing. Fermentation 3:41. doi: 10.3390/fermentation3030041
Meier-Dörnberg, T., Kory, O. I., Jacob, F., Michel, M., and Hutzler, M. (2018). Saccharomyces cerevisiae variety diastaticus friend or foe?-spoilage potential and brewing ability of different Saccharomyces cerevisiae variety diastaticus yeast isolates by genetic, phenotypic and physiological characterization. FEMS Yeast Res. 18:foy023. doi: 10.1093/femsyr/foy023
Meier-Dörnberg, T., Michel, M., Wagner, R., Jacob, F., and Hutzler, M. (2017b). Genetic and phenotypic characterization of different top-fermenting Saccharomyces cerevisiae Ale yeast isolates. BrewingScience 70, 9–25.
Meilgaard, M. (1975a). Flavor chemistry of beer: part II: flavor and threshold of 239 aroma volatiles. Tech. Q. MBAA 12, 151–168.
Meilgaard, M. (1975b). Flavor chemistry of beer. I. flavor interaction between principal volatiles. Tech. Q. MBAA 12, 151–168.
Michel, M., Kopecká, J., Meier-Dörnberg, T., Zarnkow, M., Jacob, F., and Hutzler, M. (2016). Screening for new brewing yeasts in the non-Saccharomyces sector with Torulaspora delbrueckii as model. Yeast 33, 129–144. doi: 10.1002/yea.3146
Molinet, J., and Cubillos, F. A. (2020). Wild yeast for the future: exploring the use of wild strains for wine and beer fermentation. Frontiers in Genetics 11:589350. doi: 10.3389/fgene.2020.589350
Naseeb, S., Alsammar, H., Burgis, T., Donaldson, I., Knyazev, N., Knight, C., et al. (2018). Whole genome sequencing, de Novo assembly and phenotypic profiling for the new budding yeast species Saccharomyces jurei. G3 (Bethesda) 8, 2967–2977. doi: 10.1534/g3.118.200476
Naseeb, S., James, S. A., Alsammar, H., Michaels, C. J., Gini, B., Nueno-Palop, C., et al. (2017). Saccharomyces jurei sp. nov., isolation and genetic identification of a novel yeast species from Quercus robur. Int. J. Syst. Evol. Microbiol. 67, 2046–2052. doi: 10.1099/ijsem.0.002013
Naumov, G. I., James, S. A., Naumova, E. S., Louis, E. J., and Roberts, I. N. (2000). Three new species in the Saccharomyces sensu stricto complex: Saccharomyces cariocanus, Saccharomyces kudriavzevii and Saccharomyces mikatae. Int. J. Syst. Evol. Microbiol. 50(Pt 5), 1931–1942. doi: 10.1099/00207713-50-5-1931
Nikulin, J., Krogerus, K., and Gibson, B. (2018). Alternative Saccharomyces interspecies hybrid combinations and their potential for low-temperature wort fermentation. Yeast 35, 113–127. doi: 10.1002/yea.3246
Nikulin, J., Vidgren, V., Krogerus, K., Magalhães, F., Valkeemäki, S., Kangas-Heiska, T., et al. (2020). Brewing potential of the wild yeast species Saccharomyces paradoxus. Eur. Food Res. Tech. 246, 2283–2297.
Osburn, K., Nisaa, N., and Bochman, M. (2016). Bio-prospecting, selection, and analysis of wild yeasts for ethanol fermentation. Zymurgy 39, 81–88.
Pedersen, B. S., and Quinlan, A. R. (2017). Mosdepth: quick coverage calculation for genomes and exomes. Bioinformatics 34, 867–868. doi: 10.1093/bioinformatics/btx699
Peris, D., Moriarty, R. V., Alexander, W. G., Baker, E., Sylvester, K., Sardi, M., et al. (2017). Hybridization and adaptive evolution of diverse Saccharomyces species for cellulosic biofuel production. Biotechnol. Biofuels 10:78. doi: 10.1186/s13068-017-0763-7
Peris, D., Pérez-Torrado, R., Hittinger, C. T., Barrio, E., and Querol, A. (2018). On the origins and industrial applications of Saccharomyces cerevisiae × Saccharomyces kudriavzevii hybrids. Yeast 35, 51–69. doi: 10.1002/yea.3283
Peris, D., Sylvester, K., Libkind, D., Gonçalves, P., Sampaio, J. P., Alexander, W. G., et al. (2014). Population structure and reticulate evolution of Saccharomyces eubayanus and its lager-brewing hybrids. Mol. Ecol. 23, 2031–2045. doi: 10.1111/mec.12702
Pontes, A., Hutzler, M., Brito, P. H., and Sampaio, J. P. (2020). Revisiting the taxonomic synonyms and populations of Saccharomyces cerevisiae—phylogeny, phenotypes, ecology and domestication. Microorganisms 8:903. doi: 10.3390/microorganisms8060903
Sampaio, J. P., and Gonçalves, P. (2008). Natural populations of Saccharomyces kudriavzevii in portugal are associated with Oak Bark and are sympatric with S. cerevisiae and S. paradoxus. Appl. Environ. Microbiol. 74, 2144–2152. doi: 10.1128/aem.02396-07
Sampaio, J. P., Pontes, A., Libkind, D., and Hutzler, M. (2017). “Taxonomy, diversity, and typing of brewing yeasts,” in Brewing Microbiology: Current Research, Omics and Microbial Ecology, eds N. A. Bokulich and C. W. Bamforth (Norfolk: Horizon Press).
Scannell, D. R., Zill, O. A., Rokas, A., Payen, C., Dunham, M. J., Eisen, M. B., et al. (2011). The awesome power of yeast evolutionary genetics: new genome sequences and strain resources for the Saccharomyces sensu stricto genus. G3: Genes| Genomes| Genetics 1, 11–25. doi: 10.1534/g3.111.000273
Schnegg, H. (1921). Das Mikroskopische Praktikum des Brauers, Anleitung zum Eingehenderen Studium der Brauereirohstoffe und Gärungsorganismen, 1. Teil: Morphologie und Anatomie der Brauereiroh-und Hilfsstoff. Stuttgart: Verlag von Ferdinand Enke.
Schnegg, H. (1922). Das Mikroskopische Praktikum des Brauers, Anleitung zum Eingehenderen Studium der Brauereirohstoffe und Gärungsorganismen, 2. Teil: Gärungsorganismen. Stuttgart: Verlag von Ferdinand Enke.
Sniegowski, P. D., Dombrowski, P. G., and Fingerman, E. (2002). Saccharomyces cerevisiae and Saccharomyces paradoxus coexist in a natural woodland site in North America and display different levels of reproductive isolation from European conspecifics. FEMS Yeast Res. 1, 299–306. doi: 10.1111/j.1567-1364.2002.tb00048.x
Vidgren, V., and Londesborough, J. (2012). Characterization of the Saccharomyces bayanus-type AGT1 transporter of lager yeast. J. Inst. Brewing 118, 148–151. doi: 10.1002/jib.22
Wang, S. A., and Bai, F. Y. (2008). Saccharomyces arboricolus sp. nov., a yeast species from tree bark. Int. J. Syst. Evol. Microbiol. 58(Pt 2), 510–514. doi: 10.1099/ijs.0.65331-0
White, T. J., Bruns, T. D., Lee, S. B., and Taylor, J. W. (1990). “Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics,” in PCR Protocols: A Guide To Methods And Amplifications, eds M. A. Innis, D. H. Gelfand, J. J. Sninsky, and T. J. White (San Diego, CA: Academic Press), 315–322.
Keywords: Saccharomyces jurei, Saccharomyces eubayanus, wild yeast, maltotriose, ethyl-hexanoate, Fraxinus excelsior (ash), beer, fermentation
Citation: Hutzler M, Michel M, Kunz O, Kuusisto T, Magalhães F, Krogerus K and Gibson B (2021) Unique Brewing-Relevant Properties of a Strain of Saccharomyces jurei Isolated From Ash (Fraxinus excelsior). Front. Microbiol. 12:645271. doi: 10.3389/fmicb.2021.645271
Received: 22 December 2020; Accepted: 08 March 2021;
Published: 31 March 2021.
Edited by:
Vittorio Capozzi, Italian National Research Council, ItalyReviewed by:
Laura Canonico, Marche Polytechnic University, ItalyMatthew Bochman, Indiana University, United States
Copyright © 2021 Hutzler, Michel, Kunz, Kuusisto, Magalhães, Krogerus and Gibson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Brian Gibson, YnJpYW4uZ2lic29uQHR1LWJlcmxpbi5kZQ==
†These authors share first authorship