 Claudio Alba1†
Claudio Alba1† Marina Aparicio1†
Marina Aparicio1† Felipe González-Martínez2,3María Isabel González-Sánchez2,3Jimena Pérez-Moreno2,3Blanca Toledo del Castillo2,3
Felipe González-Martínez2,3María Isabel González-Sánchez2,3Jimena Pérez-Moreno2,3Blanca Toledo del Castillo2,3 Juan Miguel Rodríguez1
Juan Miguel Rodríguez1 Rosa Rodríguez-Fernández2,3*
Rosa Rodríguez-Fernández2,3* Leonides Fernández4*
Leonides Fernández4*- 1Department of Nutrition and Food Science, Complutense University of Madrid, Madrid, Spain
- 2Department of Pediatrics, Hospital Universitario Gregorio Marañón, Madrid, Spain
- 3Instituto de Investigación Sanitaria Gregorio Marañón (IISGM), Madrid, Spain
- 4Department of Galenic Pharmacy and Food Technology, Complutense University of Madrid, Madrid, Spain
Bronchiolitis associated with the respiratory syncytial virus (RSV) is the leading cause of hospitalization among infants aged < 1 year. The main objective of this work was to assess the nasal and fecal microbiota and immune profiles in infants with RSV bronchiolitis, and to compare them with those of healthy infants. For this purpose, a total of 58 infants with RSV-positive bronchiolitis and 17 healthy infants (aged < 18 months) were recruited in this case-control study, which was approved by the Ethics Committee of the Hospital Gregorio Marañón. Nasal and fecal samples were obtained and submitted to bacterial microbiota analysis by 16S rDNA sequencing and to analysis of several immune factors related to inflammatory processes. Nasal samples in which Haemophilus and/or Moraxella accounted for > 20% of the total sequences were exclusively detected among infants of the bronchiolitis group. In this group, the relative abundances of Staphylococcus and Corynebacterium were significantly lower than in nasal samples from the control group while the opposite was observed for those of Haemophilus and Mannheimia. Fecal bacterial microbiota of infants with bronchiolitis was similar to that of healthy infants. Significant differences were obtained between bronchiolitis and control groups for both the frequency of detection and concentration of BAFF/TNFSF13B and sTNF.R1 in nasal samples. The concentration of BAFF/TNFSF13B was also significantly higher in fecal samples from the bronchiolitis group. In conclusion, signatures of RSV-associated bronchiolitis have been found in this study, including dominance of Haemophilus and a high concentration of BAFF/TNFSF13B, IL-8 and sTNF.R1 in nasal samples, and a high fecal concentration of BAFF/TNFSF13B.
Introduction
Respiratory syncytial virus bronchiolitis (RSV) is the leading cause of hospitalization in infants during their first year of life and one of the most frequent causes of visits in pediatric emergencies. RSV produces a broad spectrum of diseases, ranging from a mild infection of the upper airway to a severe infection of the lower airway (Leader and Kohlhase, 2002). The factors that explain the different severity of this disease in each patient are diverse and include both host factors, such as the existence of underlying malformations, prematurity, bronchopulmonary dysplasia or congenital heart disease and viral factors, including viral load or viral genotypes (García et al., 2010; Rodriguez-Fernandez et al., 2017). Most likely the severity of the disease is influenced by a combination of both types of factors and, also, by a dysregulated or impaired host innate immune response (de Steenhuijsen Piters et al., 2016). In addition, it is believed that the composition of the respiratory and fecal microbiotas may modulate the host immune response in the bronchiolitis setting. In this context, the principal aim of this study was to assess the nasal and fecal bacterial microbiotas and immune profiles of infants with RSV bronchiolitis and to compare them with those of healthy ones.
Materials and Methods
Study Design and Participants
A case-control study to investigate the microbiota in nasal secretions and fecal samples from infants with RSV-positive bronchiolitis and from healthy infants was conducted. Infants > 24 months of age, with a previous episode of wheezing or respiratory distress, immunodeficiency, chronic underlying diseases, or treatment with immunosuppressive agents were excluded. We recruited 58 infants with RSV-positive bronchiolitis from the Department of Pediatrics of the Hospital Universitario Gregorio Marañón (Madrid, Spain) over a 17 months period from January 2017 to May 2018 (Figure 1). Parallel, we also included healthy infants (n = 17) without allergy, infection or respiratory symptoms. Sample size was estimated according to previous results describing four different microbiota profiles in infants hospitalized with bronchiolitis and the smallest difference for the bacterial abundance of the most representative genera recorded between two profiles (Hasegawa et al., 2016a).
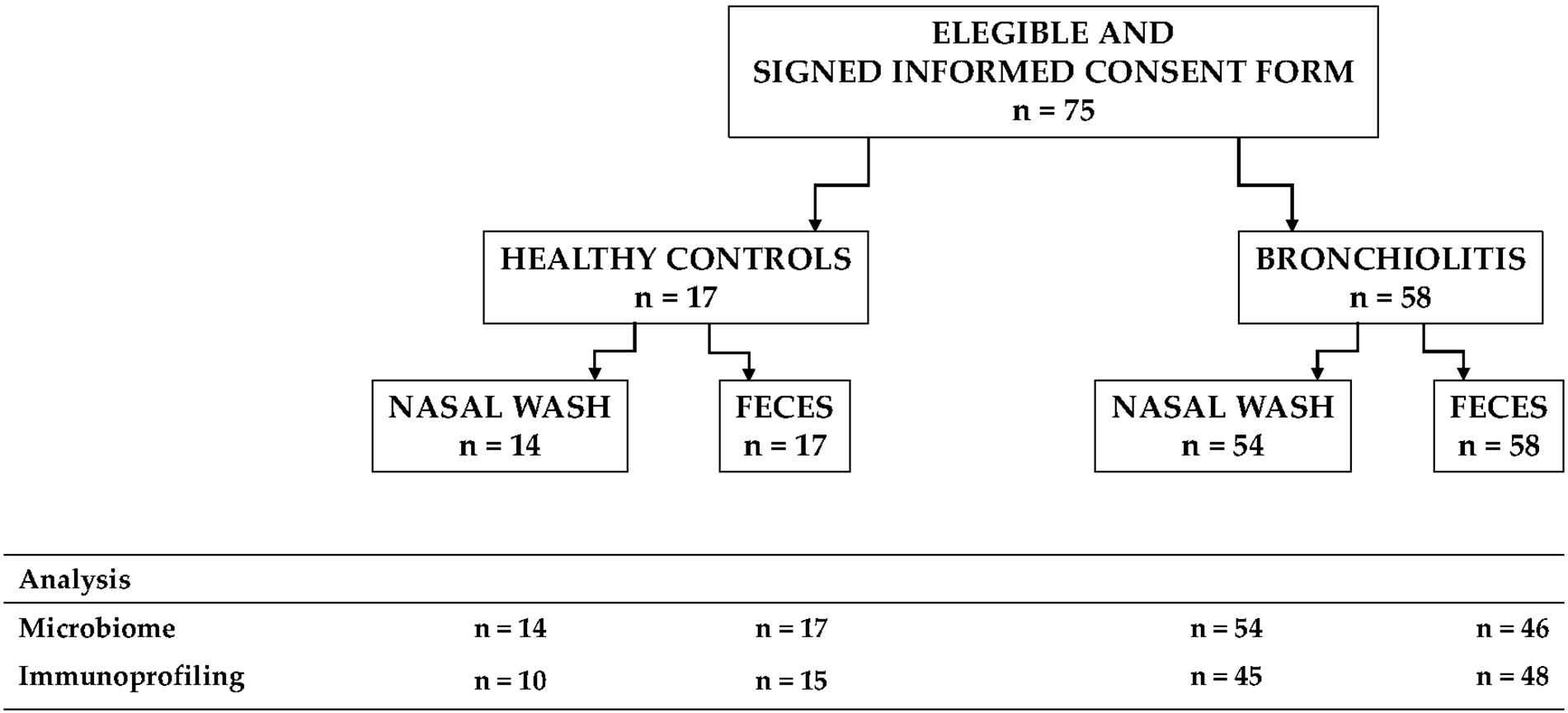
Figure 1. Flowchart of the participants included in the study, the samples collected and the analyses that were performed.
Bronchiolitis was defined as the presence of cough, rhinitis, wheezing, tachypnea, crackles, use of accessory muscles, and/or nasal flaring with or without fever according to the American Academy of Pediatrics (American Academy of Pediatrics Subcommittee on Diagnosis and Management of Bronchiolitis., 2006). Recurrent wheezing was defined by the presence of three or more episodes of physician-diagnosed wheezing within the 12 months after enrollment. RSV diagnosis was established using a direct immunofluorescence antibody (DFA) assay per standard of care and the results were confirmed by PCR in nasal wash samples obtained at enrollment (Bhandary and Boloor, 2016; Do et al., 2012). Informed consent from the parents or legal representatives of all participants was obtained. The study was approved by the Ethics Committee of the Hospital Gregorio Marañón (CEIC 276/16).
Data and Sample Collection
Clinicians recorded demographic, medical, and home environment data using a clinical questionnaire at enrollment, and by chart review. Clinical disease severity parameters were assessed using a standardized severity score at admission (González Caballero and González Pérez-Yarza, 2001), as well as need for supplemental oxygen, pediatric intensive care unit (PICU), and length of hospitalization. The most relevant characteristics of the participants are summarized in Table 1.
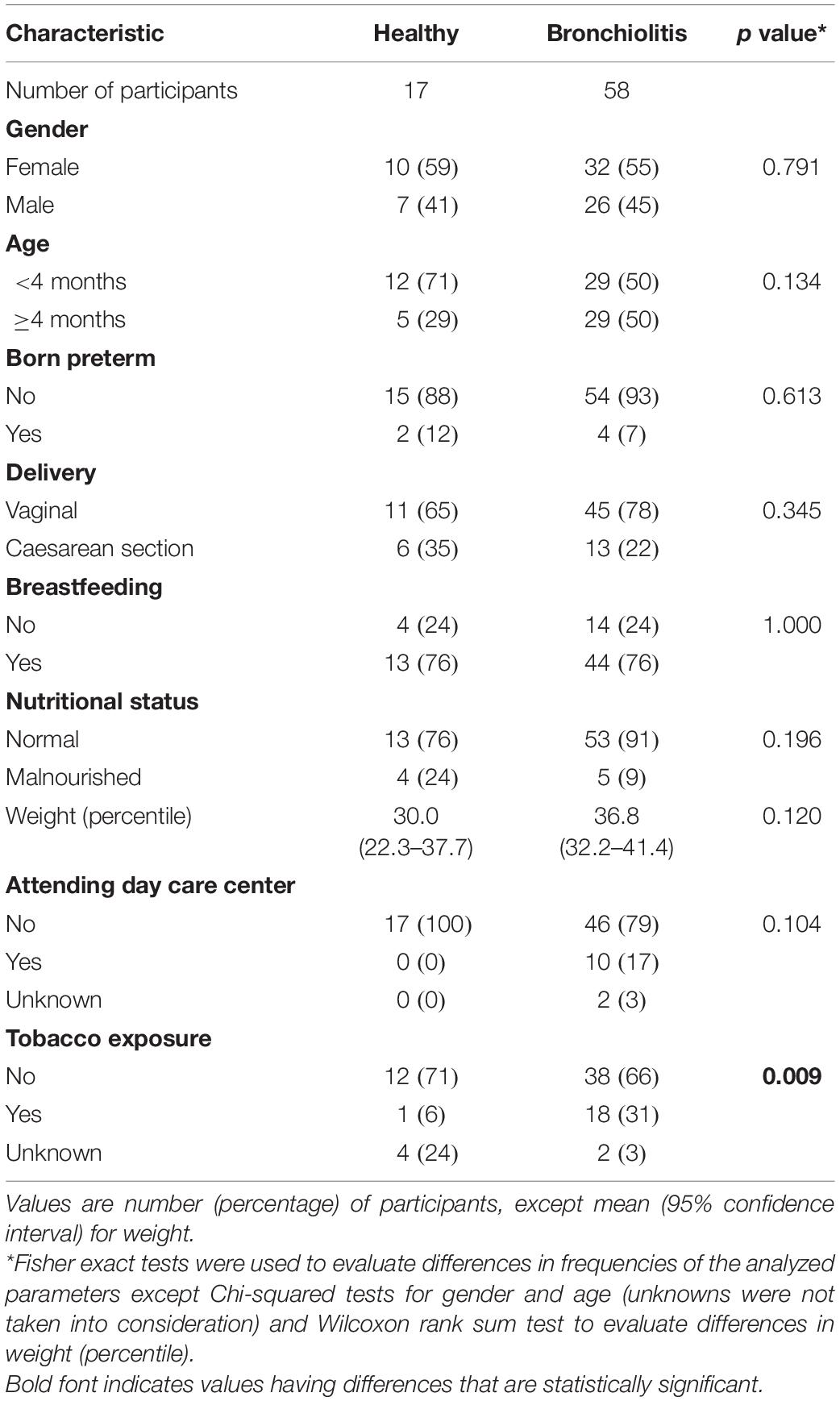
Table 1. Participant characteristics.
One nasal wash was obtained from each patient, using a standardized protocol (Stewart et al., 2017), within 24 h of hospitalization (cases) or during the routine visit (controls). Parallel, one fecal sample from each infant was collected directly from the diaper in the first 2 days of admission (cases) or during the routine visit (controls). All the collected samples were immediately stored at −20 °C.
Metataxonomic Analysis of Bacterial Microbiota
Nasal samples (1 g) from each infant were used for DNA extraction, which was performed as described (Pérez et al., 2019). Parallel, DNA was extracted from fecal samples (1 g) as already described (Lackey et al., 2019). 16S rRNA gene amplification and sequencing, targeting the V3-V4 hypervariable regions of the 16S rRNA gene, was performed in the MiSeq system of Illumina at the facilities of Parque Científico de Madrid (Tres Cantos, Spain) (Aparicio et al., 2020; Klindworth et al., 2013). The pooled, purified and barcoded DNA amplicons were sequenced using the Illumina MiSeq pair-end protocol (Illumina Inc., San Diego, CA, United States).
Taxonomical analyses of the V3-V4 region of the 16S rRNA amplicon data were conducted using MiSeq Reporter analysis software. Bioinformatic analysis was conducted combining the R version 3.5.1 (R Core Team, 2013)1, QIIME pipelines (v 1.9.1) (Caporaso et al., 2010) and Calypso (v 8.84) (Zakrzewski et al., 2017). A table of OTU counts per sample was generated, and bacterial taxa abundances were normalized to the total number of sequences in each sample. Sequences were rarified to 100,000 sequences for statistical analyses. Alpha diversity was assessed using the Shannon and Simpson diversity indices (Haegeman et al., 2013). Beta diversity studies were performed using the Principal Coordinates Analysis (PCoA) to plot patterns of bacterial community diversity through a distance matrix containing a dissimilarity value for each pairwise sample comparison. Quantitative (relative abundance) and qualitative (presence/absence) analyses were performed with the Bray-Curtis and binary Jaccard indices, respectively. PERMANOVA analysis with 999 permutations were used to reveal statistically significant differences.
Immunoprofiling
Nasal samples (1 mL) were centrifuged and the supernatants were used for the immunological assays. Fecal samples were prepared as described previously (Aparicio et al., 2020). The concentrations of a wide array of inflammation-related immune factors (APRIL/TNFSF13, BAFF/TNFSF13B, Chitinase 3-like 1, IFNα, IFNβ, IFNγ, IL2, IL8, IL10, IL11, IL12p40, IL12p70, IL19, IL20, IL22, IL26, IL27p28, IL28/IFNλ2, IL29/IFNλ1, IL32, IL34, IL35, LIGHT/TNFSF14, MMP-1, MMP-2, MMP-3, osteocalcin, osteopontin, pentraxin 3, TSLP, TWEAK/TNFSF12, gp130/sIL-6Rb, sCD30/TNFRSF8, sCD163, sIL-6Ra, sTNF-R1, and sTNF-R2) were determined using the Bio-Plex Pro Human Inflammation Assay kit (Bio-Rad, Hercules, CA, United States) in the Bio-Plex 200 instrument (Bio-Rad). Every assay was run in duplicate and standard curves were performed for each analyte.
Statistical Analysis
Quantitative data were expressed as the median and interquartile range (IQR). Differences between control and bronchiolitis groups were assessed using Fisher exact (or Chi-squared) tests for categorical variables and Wilcoxon rank sum tests with Bonferroni adjustment for multiple comparisons for quantitative variables. Principal Components Analysis (PCA) was performed to examine for similarities among nasal samples from bronchiolitis or control groups, using FactoMineR package in R. The strength and direction of association between variables was measured using the Spearman’s rank-order correlation analyses and visualized using R package corrplot. Statistical analysis and plotting were performed in the R environment (version 3.5.1 with library ggplot2 (Wickham, 2016). Differences were considered statistically significant at p < 0.05.
Results
Infants With Bronchiolitis Display an Altered Nasal Bacterial Microbiota
Baseline characteristics of the enrolled infants did not differ, but a higher proportion of infants with bronchiolitis were exposed to tobacco compared to controls (OR = 5.68; 95% CI, 0.68–47.15; p = 0.069) (Table 1).
The individual bacterial microbiota patterns of nasal samples from control group were highly similar, while those from the bronchiolitis group were more diverse (Figure 2A). The main genera contributing to the distinctive grouping of the nasal samples according to their bacterial diversity were Haemophilus, Streptococcus, Corynebacterium and Staphylococcus (Figures 2B,C). The nasal samples clustered in five groups characterized by a high abundance of Haemophilus (24% of samples), Staphylococcus (22%), Corynebacterium (13%), Streptococcus combined or not with Haemophilus or Staphylococcus (26%), and Moraxella/Streptococcus (15%) (Figure 3). All clusters included samples from both control and bronchiolitis groups, except the Haemophilus cluster that only contained samples from the bronchiolitis group. In addition, all samples containing > 20% of Haemophilus and/or Moraxella sequences corresponded to the bronchiolitis group.
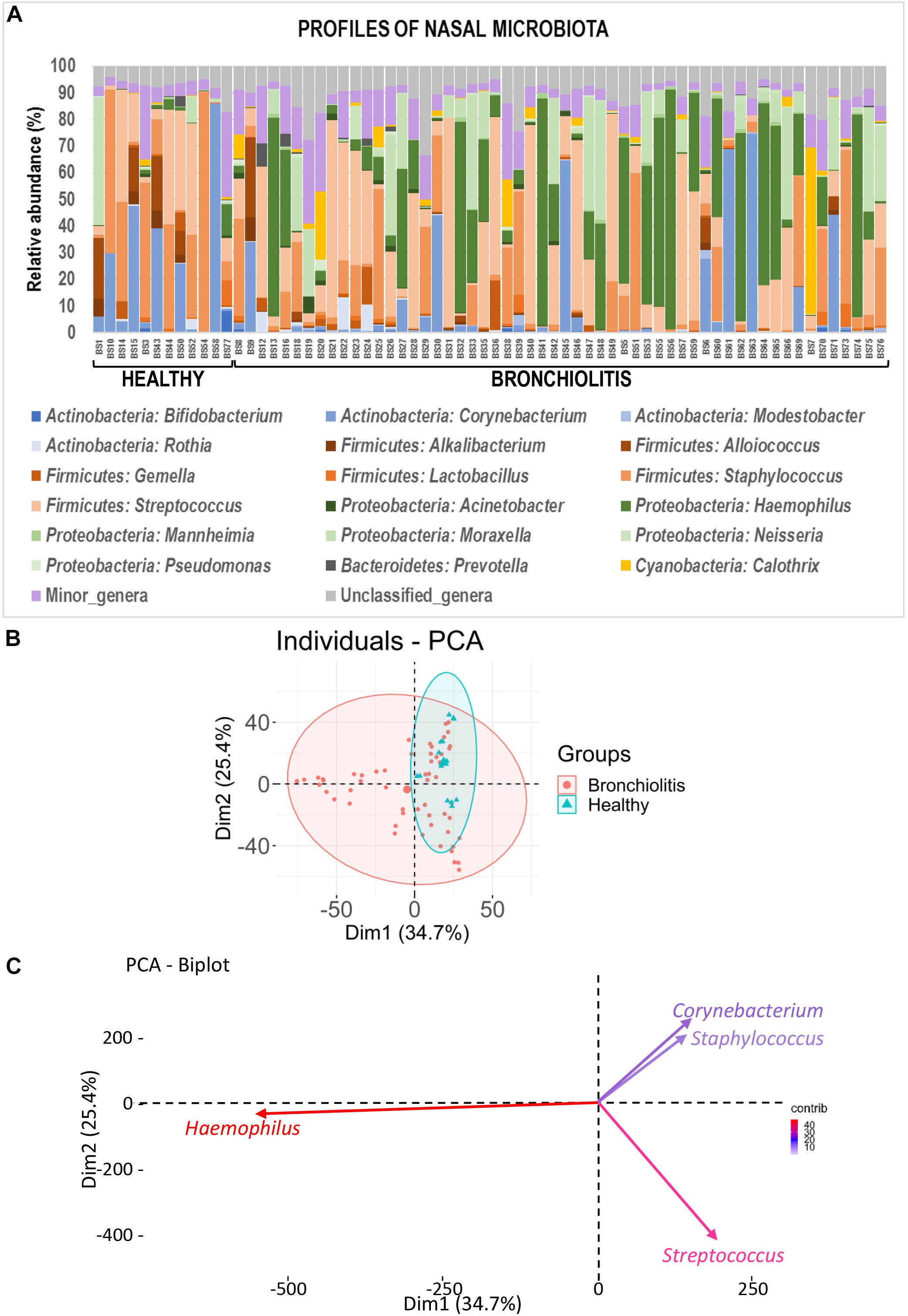
Figure 2. Nasal microbiota in healthy controls and bronchiolitis groups. (A) Individual profiles of the microbiota composition at the genus level (the 20 most abundant genera) of nasal samples from healthy control and bronchiolitis groups. Each stacked bar represents one sample and depicts the average relative abundance of the most abundant bacterial genera (color-coded). (B) Principal component analysis (PCA) for nasal samples based on the bacterial composition at the genus level showing the sample distribution around centroids (ellipses were defined by a 95% confidence interval). (C) Most influential genera (cos2 > 0.2) on sample separation presented in panel (B).
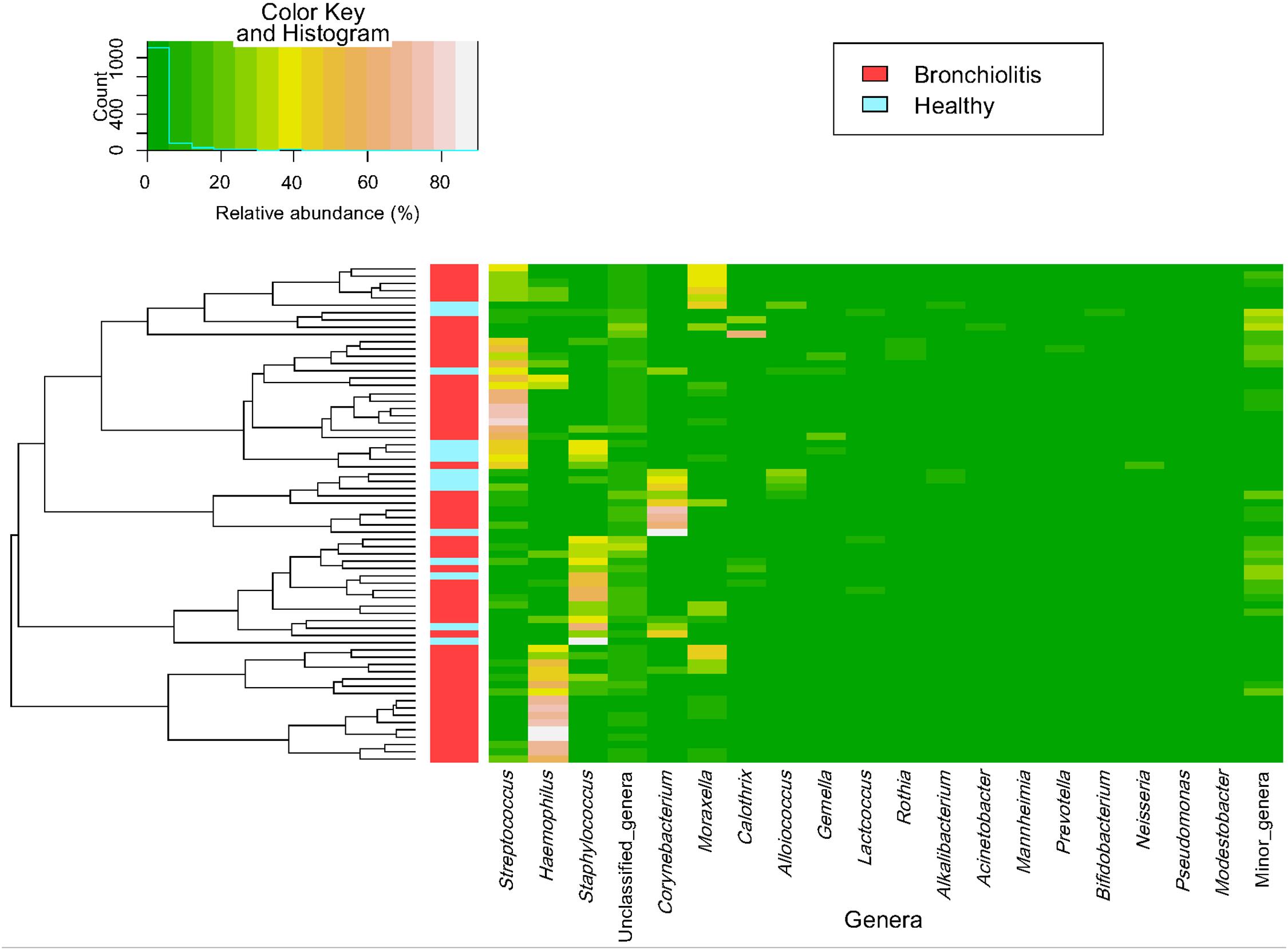
Figure 3. Heatmap of bacterial profile at the genus level in the microbiota of nasal samples. The plot depicts the relative abundance of each genera (x-axis clustering) within each sample (y-axis clustering). The relative abundances of the genera are represented by color intensity. The vertical bar (left) identifies samples from control (n = 14; blue color) and bronchiolitis (n = 54; red color) groups.
The relative abundances of Firmicutes and Actinobacteria in the nasal samples from controls (median [IQR] = 61.02% [49.84–74.03%] and 9.97% [6.98–36.62%], respectively) were nearly twice compared to those from infants with bronchiolitis (median [IQR] = 37.41% [12.09–58.69%] and 4.22% [1.16–10.96%], respectively; Wilcoxon rank sum tests, p = 0.010 and p = 0.019) (Figure 4 and Supplementary Table 1). In contrast, the relative abundance of Proteobacteria in nasal samples from infants with bronchiolitis was about 9 times higher than in the controls (median [IQR] = 32.64% [11.21–66.89%] and 3.72% [2.48–16.42%], respectively; Wilcoxon rank sum tests, p < 0.001) (Figure 4 and Supplementary Table 1). At the genus level, the relative abundances of Staphylococcus and Corynebacterium in samples from the bronchiolitis group were 10 times lower than in samples from the control group (Wilcoxon rank sum tests, p = 0.023 and p = 0.007, respectively), while those of Haemophilus and Mannheimia were 30- and 10-fold higher (Wilcoxon rank sum tests, p = 0.004 and p = 0.011, respectively) (Figure 4 and Supplementary Table 1).
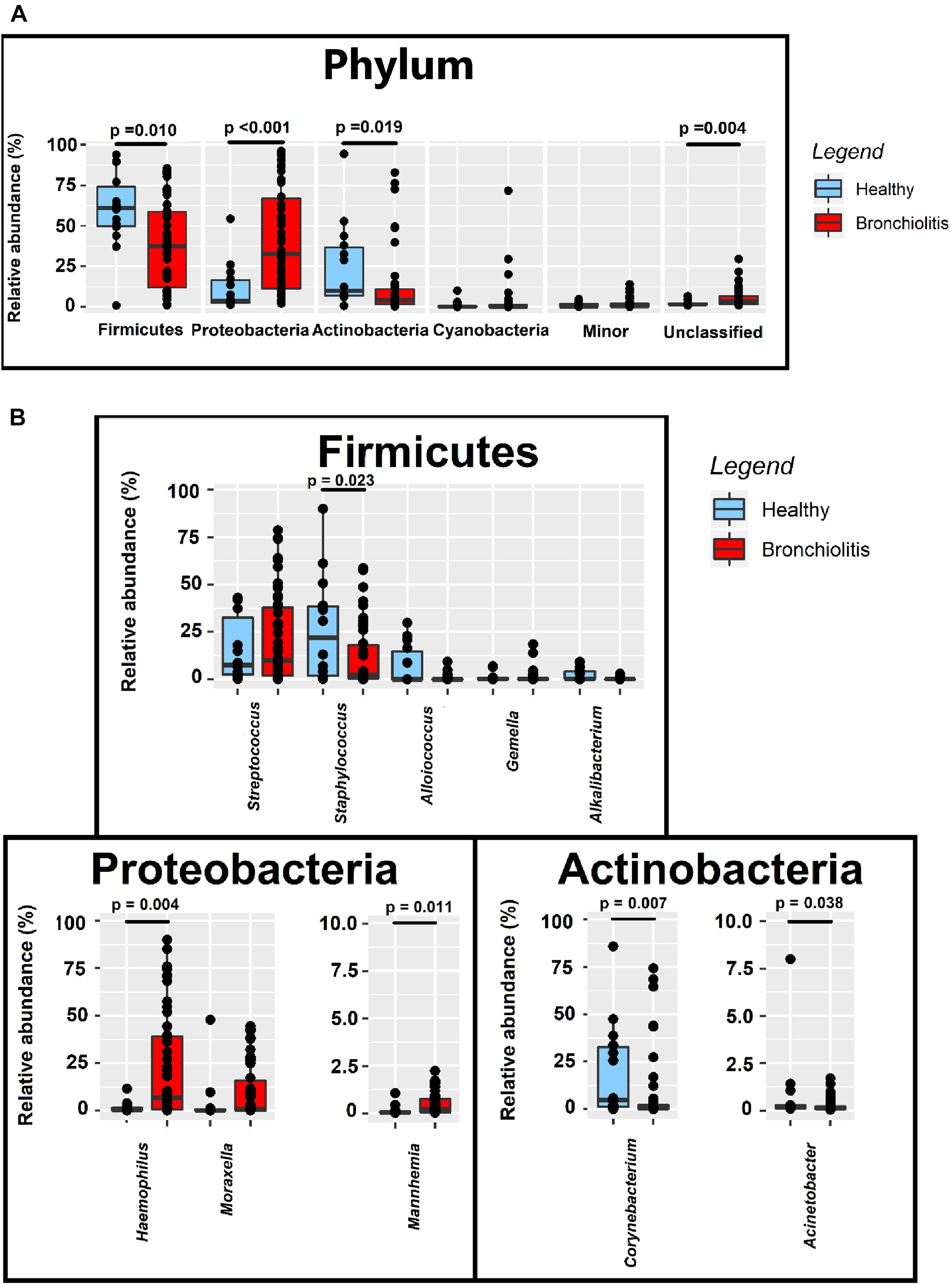
Figure 4. Relative abundance sequences at the phylum (A) and genus (B) level in nasal samples from healthy (n = 14; blue boxes) and bronchiolitis (n = 54; red boxes) infants. Significant differences between samples from both groups of infants are indicated (Wilcoxon rank sum tests).
A strong positive correlation between the relative abundance of the pairs Haemophilus/Mannheimia (ρ = 0.92) and Alkalibacterium/Alloiococcus (ρ = 0.76) and a weak negative correlation between Haemophilus/Corynebacterium (ρ = −0.41) and Mannheimia/Corynebacterium (ρ = −0.41) was recorded in nasal samples (Figure 5).
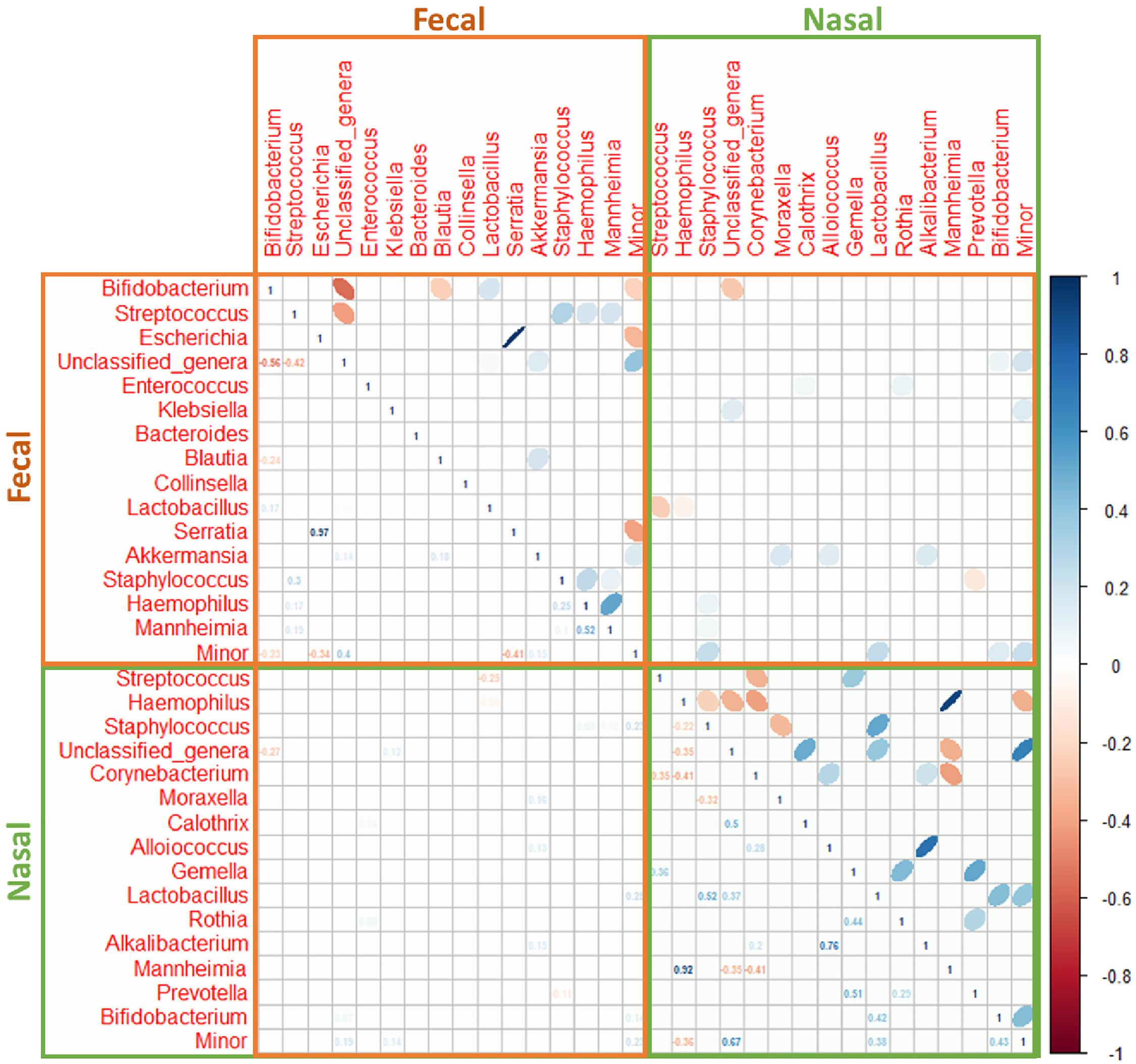
Figure 5. Spearman’s rank correlation analysis of the most abundant bacteria at the genus level in nasal (green box) and fecal (brown box) samples from both control (n = 9) and bronchiolitis (n = 34) groups. Blue ellipses represent positive correlations and red ellipses represent negative correlations. Only statistically significant correlations (p < 0.050) are shown).
Fecal Microbiota of Infants With Bronchiolitis Is Similar to That of Healthy Infants
Globally, the taxonomic profiles at the genus level of the individual fecal and nasal bacterial microbiotas were distinct (Figures 2, 6). The profiles of fecal microbiota were similar in the control and bronchiolitis groups regarding the most abundant genera (Bifidobacterium, Streptococcus, Escherichia) and changes were observed only on less abundant genera (Table 2). The relative abundance of Staphylococcus was higher in fecal samples from healthy infants than in those with bronchiolitis (median [IQR] = 0.30% [0.04–5.79%] and 0.05% [0.04–0.09%], respectively; Wilcoxon rank sum tests, p = 0.046). Haemophilus and Eggerthella were detected at a low rate in fecal samples, but at different relative abundance in infants from both groups (Table 2).
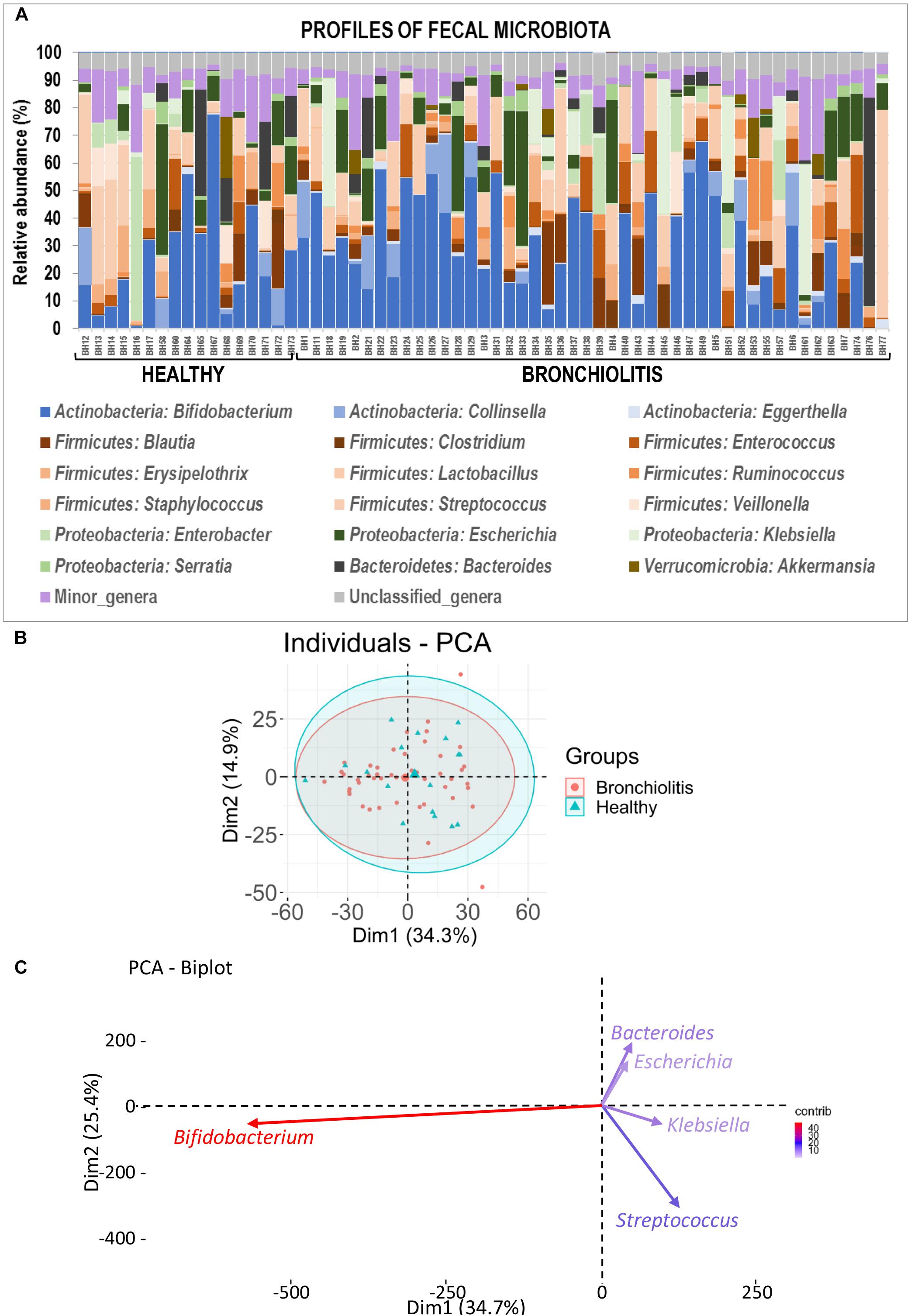
Figure 6. Fecal microbiota in healthy controls and bronchiolitis groups. (A) Individual profiles of the microbiota composition at the genus level (the 20 most abundant genera) of nasal samples from healthy control and bronchiolitis groups. Each stacked bar represents one sample and depicts the average relative abundance of the most abundant bacterial genera (color-coded). (B) Principal component analysis (PCA) for nasal samples based on the bacterial composition at the genus level showing the sample distribution around centroids (ellipses were defined by a 95% confidence interval). (C) Most influential genera (cos2 > 0.2) on sample separation presented in panel (B).
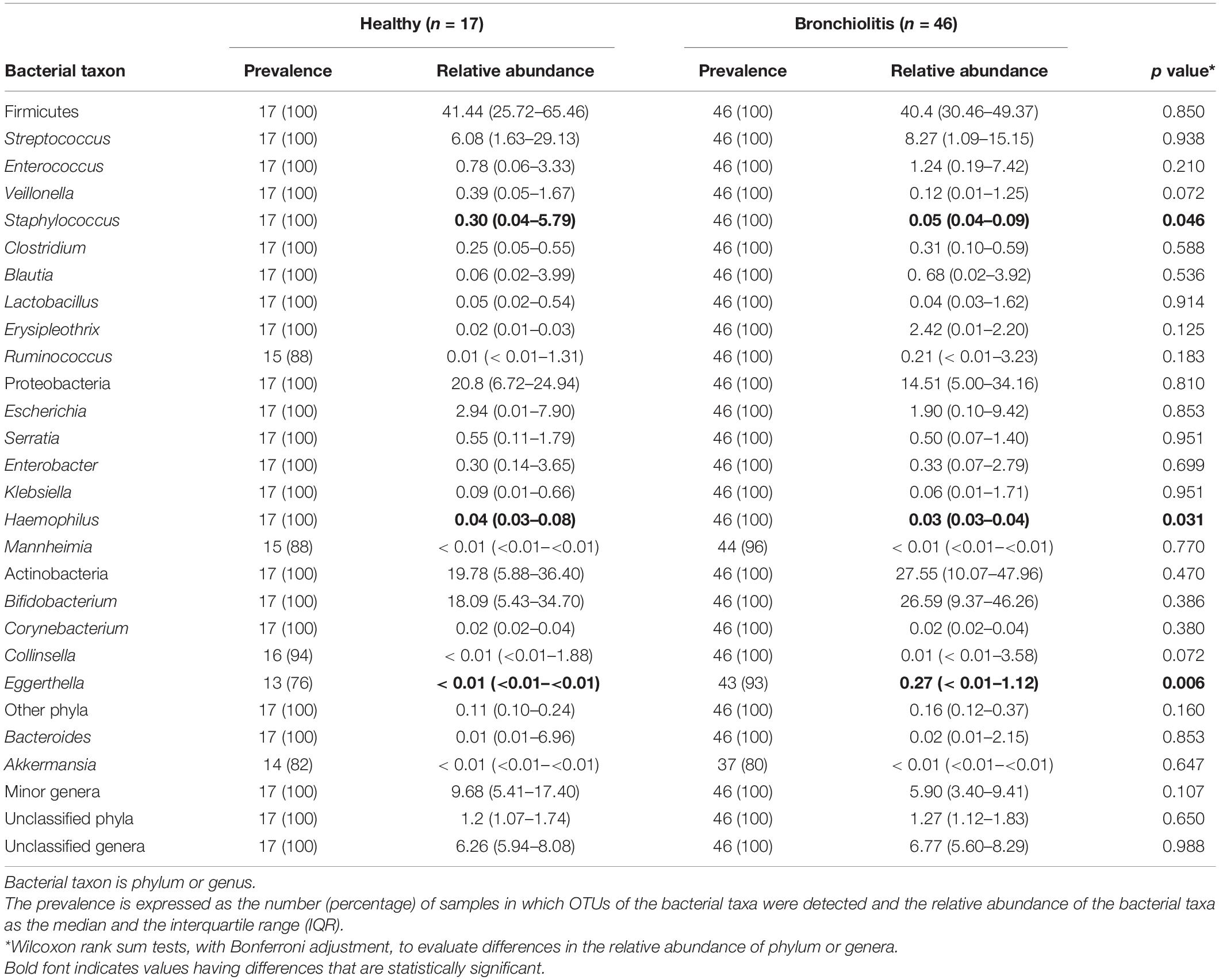
Table 2. Relative abundance of main bacterial phyla and genera in fecal samples from healthy control and bronchiolitis groups.
In fecal samples, the strongest correlation was registered in the phylum Proteobacteria, and more specifically between the pairs Escherichia/Serratia (ρ = 0.97). Haemophilus and Mannheimia were also positively correlated although the correlation coefficient (ρ = 0.52) was lower than in nasal samples (Figure 5). The relative abundances of bacterial genera in nasal and fecal samples showed very low correlation coefficients (Figure 5).
Associations Among Nasal Bacterial Microbiota and Demographic and Clinical Variables in Infants With Bronchiolitis
The nasal bacterial microbiota was compared between infants aged < 4 and ≥ 4 months because by age 4 months most infants begin eating solid food as a complementary feeding. Differences in the nasal bacterial microbiota profile between infants aged < 4 or ≥ 4 months were found for Haemophilus and Moraxella, and the minority genera Gemella, Rothia and Mannheimia (Figure 7). The relative abundance of Haemophilus in nasal secretions was higher for infants aged ≥ 4 months than for those aged < 4 months (median [IQR] = 23.10% [0.89–57.56%] and 3.47% [0.03–18.13%], respectively; Wilcoxon rank sum tests, p = 0.012) (Figure 7) and, also, when the infant had one or more siblings compared to infants who did not have any siblings (18.99% [1.96–44.47%] and 3.19% [0.17–14.18%], respectively; Wilcoxon rank sum tests, p = 0.043). The relative content of Moraxella was higher in infants attending nursery than in those who did not (median [IQR] = 13.94% [6.16–29.68%] and 0.33% [0.02–9.68%], respectively; Wilcoxon rank sum tests, p = 0.013).
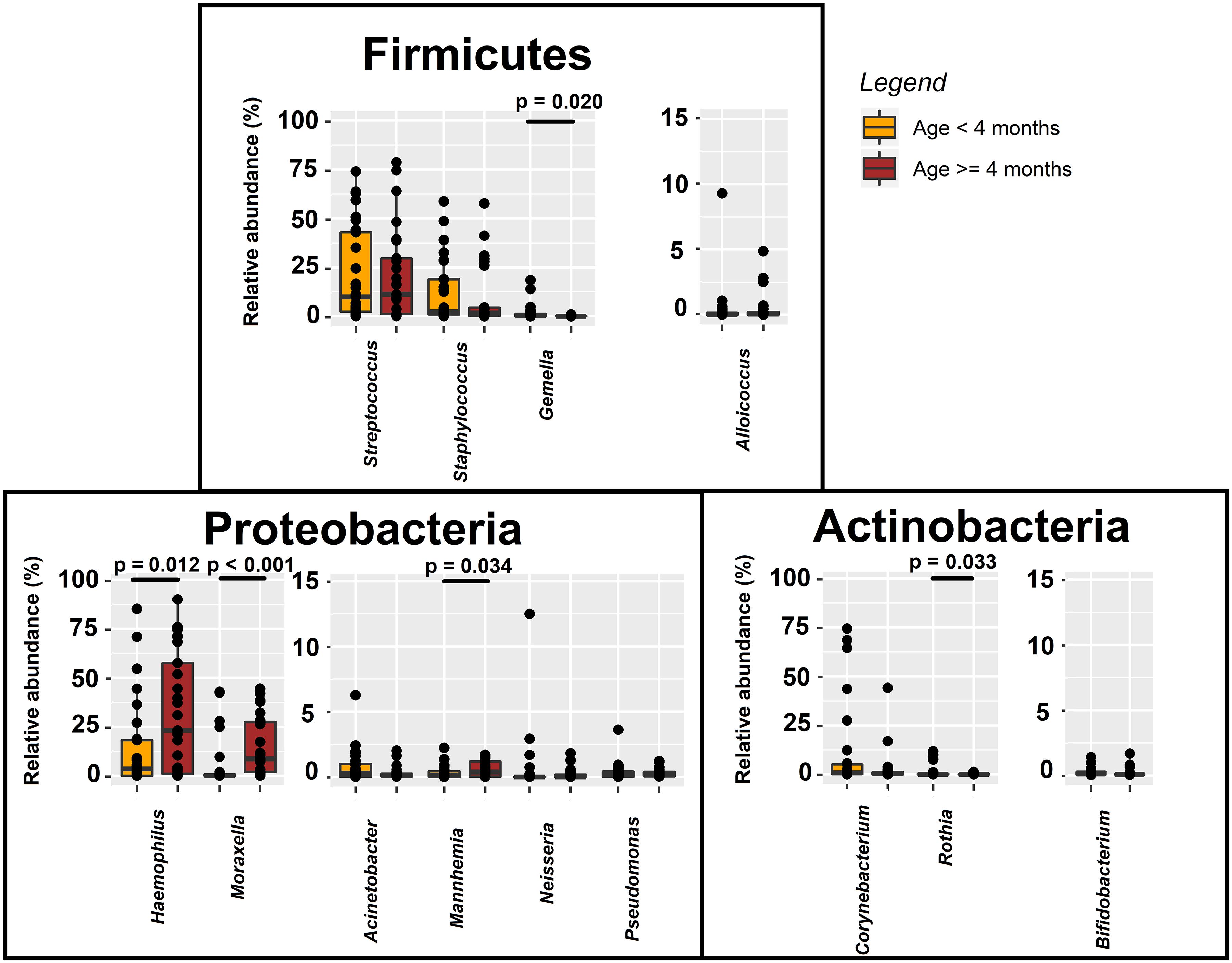
Figure 7. Relative abundance of phylum- and genus-level sequences in nasal samples from bronchiolitis infants aged < 4 years (n = 29; orange) and ≥ 4 years (n = 29; red). Significant differences between samples from both groups of infants are indicated (Wilcoxon rank sum tests).
Infants requiring hospitalization for > 1 week had about 24-, 9-, and 3-fold relative abundance of Haemophilus, Mannheimia and Corynebacterium than those requiring < 1 week (Wilcoxon rank sum tests, p = 0.013, p = 0.045, and p = 0.048, respectively) (Table 3). The relative content of Acinetobacter was about 3- or 5-fold in infants requiring PICU and/or respiratory support (Wilcoxon rank sum tests, p = 0.019 and p = 0.028, respectively) and in those diagnosed with pneumonia (Wilcoxon rank sum test, p = 0.036). Finally, wheezing was associated to higher abundance of Moraxella (median [IQR] = 8.71% (1.82–32.20%) in wheezing infants and 0.35% (0.02–10.59%) in those who did not wheeze; Wilcoxon rank sum test, p = 0.038). Wheezing infants also had higher content of Haemophilus and Mannheimia in nasal samples. On the other hand, no differences in the bacterial microbiota profile were observed in infants with bronchiolitis according to the exposure to tobacco smoke (Table 3).
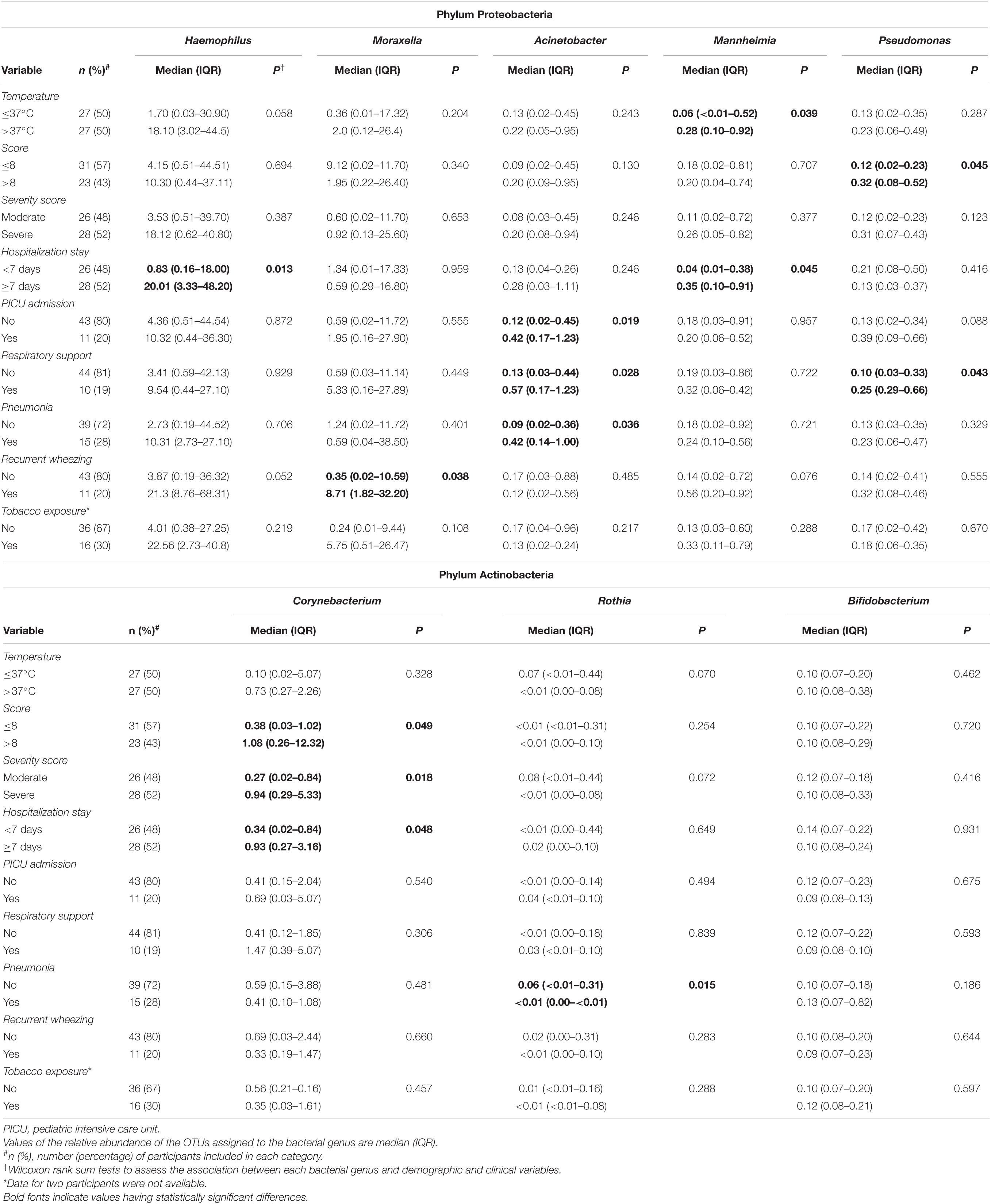
Table 3. Bivariate analysis between clinical variables and the relative abundance of the main bacterial genera detected in nasal samples of infants with bronchiolitis.
Bronchiolitis-Related Immune Signatures
Overall, the immunological profile of nasal samples from the bronchiolitis group was more complex than in healthy individuals. There were statistically significant differences in the prevalence of BAFF/TNFSF13B IL11, IL22, IL32, LIGHT/TNFSF14, MMP-2, MMP-3, TSLP, TWEAK/TNFSF12, sCD163, sIL6Rα, sTNF-R1, and sTNF-R2 (Chi-squared or Fisher exact tests, p > 0.050) (Table 4). In addition, the concentrations of BAFF/TNFSF13B and sTNF-R1 were also higher in the bronchiolitis group (Wilcoxon rank sum tests, p < 0.010) (Table 4). IL8 and osteopontin were present in a large number of samples in both groups, but IL-8 levels were about 10-fold higher in the bronchiolitis group while the opposite was noted for osteopontin (Table 4).
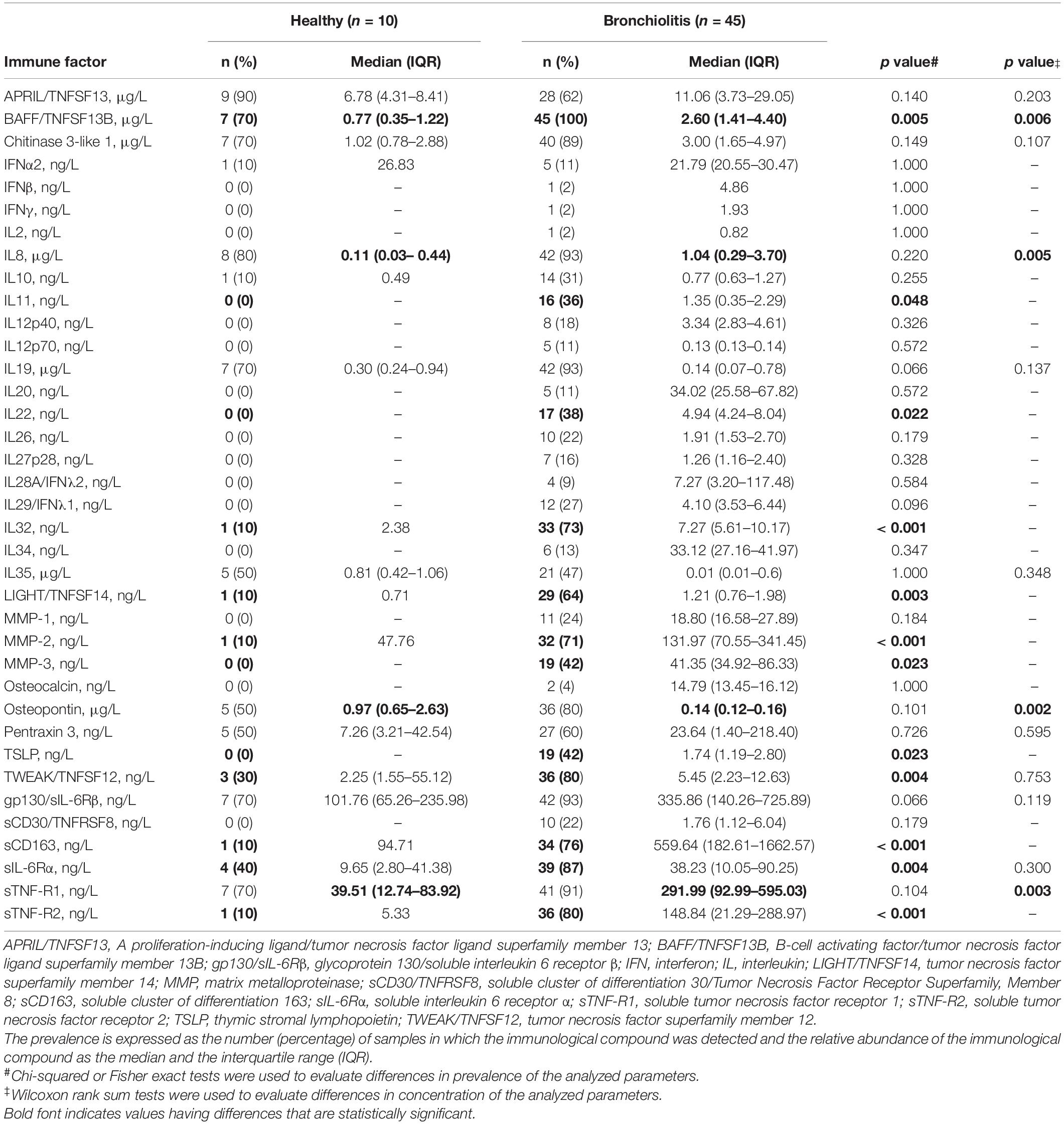
Table 4. Prevalence (%) and concentration of immune factors in nasal secretion samples from healthy control and bronchiolitis groups.
The frequency of detection of most immune factors was higher in fecal samples from healthy individuals than in the bronchiolitis group, opposite to nasal samples. BAFF/TNFSF13B was detected in almost all fecal samples but the concentration in the bronchiolitis group was approximately double that in healthy infants, similarly to nasal samples (Table 5).
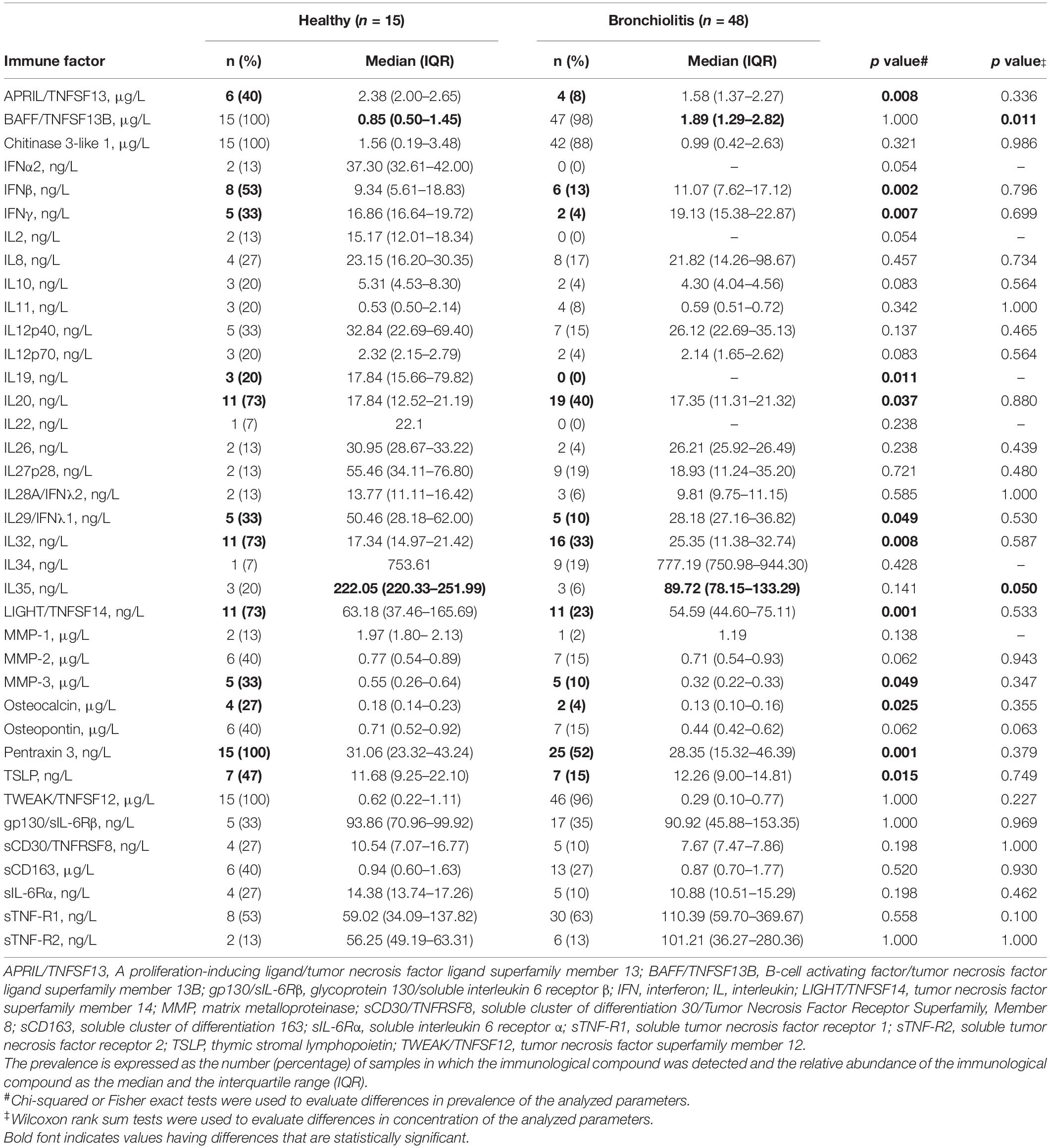
Table 5. Prevalence (%) and concentration of immune factors in fecal samples from healthy control and bronchiolitis groups.
Discussion
Although bronchiolitis is a viral infection, emerging evidence indicates that the microbiota may be associated to its pathogenesis. In this study, the composition of the bacterial microbiota and the immune profiles of nasal and fecal samples from infants with a RSV-associated bronchiolitis were assessed and compared with those from healthy ones.
All the nasal samples that were characterized by the dominance of Haemophilus or by the abundance (> 20%) of Moraxella sequences had been obtained from the bronchiolitis group. In previous studies, bronchiolitis-suffering infants with a Haemophilus- or a Moraxella-dominant profile had significantly higher rates of incidence and severity of acute respiratory infections, intensive care use or prolonged hospital stay, in comparison with the profiles of bacterial microbiota dominated by other bacteria (Hasegawa et al., 2016a; Teo et al., 2015). Our results confirm that an Haemophilus-dominant profile is a common feature of RSV-associated bronchiolitis (Hasegawa et al., 2016a; McGillivary et al., 2009; Vissing et al., 2013).
In contrast with other authors, we did not find an association between RSV-associated bronchiolitis and an abundance of Streptococcus sequences (Teo et al., 2015; de Steenhuijsen Piters et al., 2016). In addition, the abundance of Mannheimia sequences was significantly higher in the bronchiolitis group. Although this genus contains species involved in respiratory infections in a wide spectrum of mammals, such as M. (formerly Pasteurella) haemolytica, this is the first work describing an enrichment in Mannheimia sequences among infants with RSV-associated bronchiolitis. This finding must be confirmed in future studies. In the bronchiolitis group, relative abundances of Staphylococcus and Corynebacterium were significantly lower than in samples from the control group. A lower nasal abundance of Corynebacterium and Staphylococcus among infants with bronchiolitis has already been described (Hasegawa et al., 2016a; Biesbroek et al., 2014; Bosch et al., 2017). A recent study has shown that the degree of alteration of both the structure and networks of the nasal microbiota was associated to the severity of the RSV-associated bronchiolitis disease (Schippa et al., 2020).
In contrast to the nasal microbiota, the fecal microbiota of infants with bronchiolitis was similar to that of healthy infants. This finding may be limited by the sample size, especially in relation to the healthy controls. The nasal cavity drains to the pharynx, which is connected to both the trachea and the esophagus. Following the later route, nasopharyngeal bacteria may reach the gut (Proctor and Relman, 2017). However, while there are high levels of similarity between microbial communities living in the oral cavity, stomach and the gut, such similarities are not found between nasal and gut communities (Bik et al., 2006; Ding and Schloss, 2014; Bassis et al., 2015). However, Hasegawa et al. (2016b) identified 4 distinct profiles (Escherichia-dominant, Bifidobacterium-dominant, Enterobacter/Veillonella-dominant and Bacteroides-dominant) when the microbial profiles of bronchiolitis patients where compared to those of healthy controls. Interestingly, the proportion of bronchiolitis was highest among infants with the Bacteroides-dominant profile while it was lowest in those with the Enterobacter/Veillonella-dominant profile. More studies are required to elucidate or confirm potential relationships between the fecal bacteriome and bronchiolitis.
In relation to the immunological profiles, the concentration of BAFF/TNFSF13B, IL-8, and sTNF.R1 were significant higher in nasal samples from the bronchiolitis group. RSV infection upregulates expression of BAFF/TNFSF13B (Reed et al., 2009), leading to an increase in its concentration in the bronchoalveolar lavage fluid from RSV-infected infants (McNamara et al., 2013). This finding seems to be a consistent feature of the upper and lower respiratory tract responses to infections in infants, and reveals the importance of the airway epithelia in influencing immune and inflammatory responses.
In this study, BAFF was the only immune parameter which concentration was significantly higher among fecal samples from the bronchiolitis group than those from the healthy group. Airways epithelial cells and gut dendritic cells seem to play similar roles in producing BAFF in the respective mucosal surfaces, where it may support IgA production by B cells (Cerutti et al., 2011). Fecal BAFF increases when the gut is inflamed and it has been proposed as a biomarker for monitoring children with inflammatory bowel diseases (Fodor et al., 2020). The results of our study suggest that such an increase may also reflect an inflammatory state in other mucosal surfaces, such as that of the respiratory tract.
In relation to the remaining immune factors, increased IL8 and sTNF.R1 levels, and decreased osteopontin levels, have also been reported in nasal samples from infants with RSV bronchiolitis (Noah et al., 2002; Sampayo-Escobar et al., 2018; Breindahl et al., 2012). However, the number of studies addressing such immune compounds in the RSV context is still low and more studies are required to confirm their relevance as RSV biomarkers.
The main limitation of this study is the limited number of participants according to the high variability of some bacterial microbiota and immunological parameters associated to RSV bronchiolitis. In addition, all participants were recruited from a single center. However, this allowed us to determine the microbiota composition and immunoprofiling (including a very large number of immune compounds) in nasal and fecal samples of the same participants. Despite our efforts, there were missing samples and data, but this fact did not interfere with the performed analyses and the number of participants included in each comparison is adequately indicated in each table and figure. It must be taken into account that the bacteriomes of the upper respiratory and intestinal tracts are dynamic and seem to evolve rapidly during early life (Bosch et al., 2017; Durack and Christophersen, 2020); this fact may exert an influence, which is currently unknown, when results obtained from infants of different ages are compared. In conclusion, the results of this study confirm that RSV-associated bronchiolitis is correlated with an abundance of Haemophilus sequences and to an increase in the concentrations of BAFF/TNFSF13B and IL8 in nasal samples. In addition, it suggests the existence of other potential biomarkers, including an increase in Mannheimia sequences and in the concentrations of fecal BAFF/TNFSF13B and nasal sTNF.R1 and a decrease in nasal osteopontin. Such markers may be applied in the future to evaluate the response of infants with RSV-bronchiolitis to treatments.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The studies involving human participants were reviewed and approved by Ethics Committee of the Hospital Gregorio Marañón. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author Contributions
JR, RR-F, and LF conceived and designed the study, supervised the analyses of the data, drafted the initial manuscript, and reviewed and revised the manuscript. FG-M, MG-S, JP-M, and BT recruited the infants, collected the samples, the demographic and clinical data, and reviewed and revised the manuscript. CA and MA performed the metataxonomic and immunological analysis, including initial statistical and bioinformatic analysis, and reviewed and revised the manuscript. All authors discussed the results, commented on the manuscript and approved the submitted version of the manuscript.
Funding
This research was supported by grant PID2019-105606RB-100 from the Ministerio de Ciencia e Innovación (Spain).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank to all the participants for taking part in our study.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.667832/full#supplementary-material
Footnotes
References
American Academy of Pediatrics Subcommittee on Diagnosis and Management of Bronchiolitis. (2006). Diagnosis and management of bronchiolitis. Pediatrics 118, 1774–1793. doi: 10.1542/peds.2006-2223
Aparicio, M., Alba, C., CAM Public Health Area 6 PSGO, Rodríguez, J. M., and Fernández, L. (2020). Microbiological and immunological markers in milk and infant feces for common gastrointestinal disorders: a pilot study. Nutrients 12, 634. doi: 10.3390/nu12030634
Bassis, C. M., Erb-Downward, J. R., Dickson, R. P., Freeman, C. M., Schmidt, T. M., Young, V. B., et al. (2015). Analysis of the upper respiratory tract microbiotas as the source of the lung and gastric microbiotas in healthy individuals. mBio 6, e00037. doi: 10.1128/mBio.00037-15
Bhandary, R., and Boloor, R. (2016). Detection of respiratory syncytial virus using direct fluorescent antibody assay in paediatric patients with acute respiratory tract infection. J. Clin. Diagn. Res. 10, DC10–DC12. doi: 10.7860/JCDR/2016/17530.8044
Biesbroek, G., Tsivtsivadze, E., Sanders, E. A., Montijn, R., Veenhoven, R. H., Keijser, B. J., et al. (2014). Early respiratory microbiota composition determines bacterial succession patterns and respiratory health in children. Am. J. Respir. Crit. Care Med. 190, 1283–1292. doi: 10.1164/rccm.201407-1240OC
Bik, E. M., Eckburg, P. B., Gill, S. R., Nelson, K. E., Purdom, E. A., Francois, F., et al. (2006). Molecular analysis of the bacterial microbiota in the human stomach. Proc. Natl. Acad. Sci. U S A. 103, 732–737. doi: 10.1073/pnas.0506655103
Bosch, A. A. T. M., de Steenhuijsen Piters, W. A. A., van Houten, M. A., Chu, M. L. J. N., Biesbroek, G., Kool, J., et al. (2017). Maturation of the infant respiratory microbiota, environmental drivers and health consequences: a prospective cohort study. Am. J. Respir. Crit. Care Med. 196, 1582–1590. doi: 10.1164/rccm.201703-0554OC
Breindahl, M., Rieneck, K., Nielsen, C., Justesen, T., Bendtzen, K., Müller, K., et al. (2012). Cytokine responses in infants infected with respiratory syncytial virus. Open J. Immunol. 2, 40–48. doi: 10.4236/oji.2012.21005
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., Costello, E. K., et al. (2010). QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth.f.303
Cerutti, A., Chen, K., and Chorny, A. (2011). Immunoglobulin responses at the mucosal interface. Annu. Rev. Immunol. 29, 273–293. doi: 10.1146/annurev-immunol-031210-101317
de Steenhuijsen Piters, W. A., Heinonen, S., Hasrat, R., Bunsow, E., Smith, B., Suarez-Arrabal, M. C., et al. (2016). Nasopharyngeal microbiota, host transcriptome, and disease severity in children with respiratory syncytial virus infection. Am. J. Respir. Crit. Care Med. 194, 1104–1115. doi: 10.1164/rccm.201602-0220OC
Ding, T., and Schloss, P. D. (2014). Dynamics and associations of microbial community types across the human body. Nature. 509, 357–360. doi: 10.1038/nature13178
Do, L. A., van Doorn, H. R., Bryant, J. E., Nghiem, M. N., Nguyen Van, V. C., Vo, C. K., et al. (2012). A sensitive real-time PCR for detection and subgrouping of human respiratory syncytial virus. J. Virol. Methods 179, 250–255. doi: 10.1016/j.jviromet.2011.11.012
Durack, J., and Christophersen, C. T. (2020). Human respiratory and gut microbiomes-do they really contribute to respiratory health? Front. Pediatr. 8:528. doi: 10.3389/fped.2020.00528
Fodor, I., Serban, O., Serban, D. E., Farcau, D., Man, S. C., and Dumitrascu, D. L. (2020). B cell-activating factor (BAFF) in children with inflammatory bowel disease. Pediatr. Res. doi: 10.1038/s41390-020-01155-1 ∗∗ vol page,
García, C. G., Bhore, R., Soriano-Fallas, A., Trost, M., Chason, R., Ramilo, O., et al. (2010). Risk factors in children hospitalized with RSV bronchiolitis versus non-RSV bronchiolitis. Pediatrics. 126, e1453–e1460. doi: 10.1542/peds.2010-0507
González Caballero, D., and González Pérez-Yarza, E. (2001). Bronquiolitis aguda: bases para un protocolo racional [Acute bronchiolitis: fundamentals of a rational protocol]. An. Esp. Pediatr. 55, 355–364. doi: 10.1016/S1695-4033(01)77699-0
Haegeman, B., Hamelin, J., Moriarty, J., Neal, P., Dushoff, J., and Weitz, J. S. (2013). Robust estimation of microbial diversity in theory and in practice. ISME J. 7, 1092–1101. doi: 10.1038/ismej.2013.10
Hasegawa, K., Linnemann, R. W., Mansbach, J. M., Ajami, N. J., Espinola, J. A., Petrosino, J. F., et al. (2016a). The fecal microbiota profile and bronchiolitis in infants. Pediatrics. 138, e20160218. doi: 10.1542/peds.2016-0218
Hasegawa, K., Mansbach, J. M., Ajami, N. J., Espinola, J. A., Henke, D. M., Petrosino, J. F., et al. (2016b). Association of nasopharyngeal microbiota profiles with bronchiolitis severity in infants hospitalised for bronchiolitis. Eur. Respir. J. 48, 1329–1339. doi: 10.1183/13993003.00152-2016
Klindworth, A., Pruesse, E., Schweer, T., Peplies, J., Quast, C., Horn, M., et al. (2013). Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 41, e1. doi: 10.1093/nar/gks808
Lackey, K. A., Williams, J. E., Meehan, C. L., Zachek, J. A., Benda, E. D., Price, W. J., et al. (2019). What’s normal? Microbiomes in human milk and infant feces are related to each other but vary geographically: The INSPIRE study. [published correction appears in Front. Nutr. 2020 Feb 19;7:12]. Front. Nutr. 6:45. doi: 10.3389/fnut.2019.00045
Leader, S., and Kohlhase, K. (2002). Respiratory syncytial virus-coded pediatric hospitalizations, 1997 to 1999. Pediatr. Infect. Dis. J. 21, 629–632. doi: 10.1097/00006454-200207000-00005
McGillivary, G., Mason, K. M., Jurcisek, J. A., Peeples, M. E., and Bakaletz, L. O. (2009). Respiratory syncytial virus-induced dysregulation of expression of a mucosal beta-defensin augments colonization of the upper airway by non-typeable Haemophilus influenzae. Cell. Microbiol. 11, 1399–1408. doi: 10.1111/j.1462-5822.2009.01339.x
McNamara, P. S., Fonceca, A. M., Howarth, D., Correia, J. B., Slupsky, J. R., Trinick, R. E., et al. (2013). Respiratory syncytial virus infection of airway epithelial cells, in vivo and in vitro, supports pulmonary antibody responses by inducing expression of the B cell differentiation factor BAFF. Thorax. 68, 76–81. doi: 10.1136/thoraxjnl-2012-202288
Noah, T. L., Ivins, S. S., Murphy, P., Kazachkova, I., Moats-Staats, B., Henderson, F. W., et al. (2002). Chemokines and inflammation in the nasal passages of infants with respiratory syncytial virus bronchiolitis. Clin. Immunol. 104, 86–95. doi: 10.1006/clim.2002.5248
Pérez, T., Alba, C., Aparicio, M., de Andrés, J., Ruiz Santa Quiteria, J. A., Rodríguez, J. M., et al. (2019). Abundant bacteria in the proximal and distal intestine of healthy Siberian sturgeons (Acipenser baerii). Aquaculture. 506, 325–336. doi: 10.1016/j.aquaculture.2019.03.055
Proctor, D. M., and Relman, D. A. (2017). The landscape ecology and microbiota of the human nose, mouth, and throat. Cell Host Microbe. 21, 421–432. doi: 10.1016/j.chom.2017.03.011
R Core Team (2013). R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna: R Core Team.
Reed, J. L., Welliver, T. P., Sims, G. P., McKinney, L., Velozo, L., Avendano, L., et al. (2009). Innate immune signals modulate antiviral and polyreactive antibody responses during severe respiratory syncytial virus infection. J. Infect. Dis. 199, 1128–1138. doi: 10.1086/597386
Rodriguez-Fernandez, R., Tapia, L. I., Yang, C. F., Torres, J. P., Chavez-Bueno, S., Garcia, C., et al. (2017). Respiratory syncytial virus genotypes, host immune profiles, and disease severity in young children hospitalized with bronchiolitis. J. Infect. Dis. 217, 24–34. doi: 10.1093/infdis/jix543
Sampayo-Escobar, V., Green, R., Cheung, M. B., Bedi, R., Mohapatra, S., Mohapatra, S. S., et al. (2018). Osteopontin plays a pivotal role in increasing severity of respiratory syncytial virus infection. PLoS One. 13:e0192709. doi: 10.1371/journal.pone.0192709
Schippa, S., Frassanito, A., Marazzato, M., Nenna, R., Petrarca, L., Neroni, B., et al. (2020). Nasal microbiota in RSV bronchiolitis. Microorganisms. 8, 731. doi: 10.3390/microorganisms8050731
Stewart, C. J., Mansbach, J. M., Wong, M. C., Ajami, N. J., Petrosino, J. F., and Camargo, C. A. Jr., et al. (2017). Associations of nasopharyngeal metabolome and microbiome with severity among infants with bronchiolitis. A multiomic analysis. Am. J. Respir. Crit. Care Med. 196, 882–891. doi: 10.1164/rccm.201701-0071OC
Teo, S. M., Mok, D., Pham, K., Kusel, M., Serralha, M., Troy, N., et al. (2015). The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe. 17, 704–715. doi: 10.1016/j.chom.2015.03.008
Vissing, N. H., Chawes, B. L., and Bisgaard, H. (2013). Increased risk of pneumonia and bronchiolitis after bacterial colonization of the airways as neonates. Am. J. Respir. Crit. Care Med. 188, 1246–1252. doi: 10.1164/rccm.201302-0215OC
Keywords: bronchiolitis, respiratory syncytial virus, immunoprofile, nasal lavage, Haemophilus, BAFF/TNFSF13B, bacterial microbiota
Citation: Alba C, Aparicio M, González-Martínez F, González-Sánchez MI, Pérez-Moreno J, Toledo del Castillo B, Rodríguez JM, Rodríguez-Fernández R and Fernández L (2021) Nasal and Fecal Microbiota and Immunoprofiling of Infants With and Without RSV Bronchiolitis. Front. Microbiol. 12:667832. doi: 10.3389/fmicb.2021.667832
Received: 14 February 2021; Accepted: 08 April 2021;
Published: 01 June 2021.
Edited by:
Joanna Kirman, University of Otago, New ZealandReviewed by:
Alessandra Pierangeli, Sapienza University of Rome, ItalyAntonella Frassanito, Sapienza University of Rome, Italy
Jonathan Mansbach, Boston Children’s Hospital and Harvard Medical School, United States
Copyright © 2021 Alba, Aparicio, González-Martínez, González-Sánchez, Pérez-Moreno, Toledo del Castillo, Rodríguez, Rodríguez-Fernández and Fernández. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rosa Rodríguez-Fernández, cm9zYS5yb2RyaWd1ZXoyMDAwQGdtYWlsLmNvbQ==; Leonides Fernández, bGVvbmlkZXNAdWNtLmVz
†These authors have contributed equally to this work and share first authorship