 Fernanda Esposito1,2*†
Fernanda Esposito1,2*† Brenda Cardoso2,3†
Brenda Cardoso2,3† Herrison Fontana1,2
Herrison Fontana1,2 Bruna Fuga1,2,3
Bruna Fuga1,2,3 Adriana Cardenas-Arias2,3
Adriana Cardenas-Arias2,3 Quézia Moura4
Quézia Moura4 Danny Fuentes-Castillo2,5
Danny Fuentes-Castillo2,5 Nilton Lincopan1,2,3*
Nilton Lincopan1,2,3*- 1Department of Clinical Analysis, School of Pharmacy, University of São Paulo, São Paulo, Brazil
- 2One Health Brazilian Resistance Project (OneBR), São Paulo, Brazil
- 3Department of Microbiology, Institute of Biomedical Sciences, University of São Paulo, São Paulo, Brazil
- 4Federal Institute of Education, Science and Technology of Espírito Santo, Vila Velha, Brazil
- 5Department of Pathology, School of Veterinary Medicine and Animal Sciences, University of São Paulo, São Paulo, Brazil
The dissemination of antibiotic-resistant priority pathogens beyond hospital settings is both a public health and an environmental problem. In this regard, high-risk clones exhibiting a multidrug-resistant (MDR) or extensively drug-resistant (XDR) phenotype have shown rapid adaptation at the human-animal-environment interface. In this study, we report genomic data and the virulence potential of the carbapenemase, São Paulo metallo-β-lactamase (SPM-1)-producing Pseudomonas aeruginosa strains (Pa19 and Pa151) isolated from polluted urban rivers, in Brazil. Bioinformatic analysis revealed a wide resistome to clinically relevant antibiotics (carbapenems, aminoglycosides, fosfomycin, sulfonamides, phenicols, and fluoroquinolones), biocides (quaternary ammonium compounds) and heavy metals (copper), whereas the presence of exotoxin A, alginate, quorum sensing, types II, III, and IV secretion systems, colicin, and pyocin encoding virulence genes was associated with a highly virulent behavior in the Galleria mellonella infection model. These results confirm the spread of healthcare-associated critical-priority P. aeruginosa belonging to the MDR sequence type 277 (ST277) clone beyond the hospital, highlighting that the presence of these pathogens in environmental water samples can have clinical implications for humans and other animals.
Introduction
Carbapenem-resistant Pseudomonas aeruginosa are a leading cause of hospital-acquired infections and have become a health priority (Tacconelli et al., 2018). Efforts have been made to prevent colonization, infection, and decrease mortality. Based on that, the WHO proposed a global priority pathogen list of multidrug-resistant (MDR) bacteria to drive research, discovery, and development of new antibiotics. Along with MDR P. aeruginosa, the critical pathogens WHO list included Acinetobacter baumannii and bacteria from Enterobacterales group (Tacconelli et al., 2018). They were categorized as critical priority through the use of multi-criteria, including being resistant to a large number of antibiotics, such as carbapenems and third generation cephalosporins, the best available options for treating MDR pathogens (Babu et al., 2020). Worryingly, carbapenem-resistant P. aeruginosa can cause severe and often deadly infections such as bloodstream infections, pneumonia, and osteomyelitis (Fernández-Barat et al., 2017; Pliska, 2020; Jean et al., 2020; Bobrov et al., 2021; Rosales-Reyes et al., 2021). Carbapenem resistance is usually multifactorial, including overexpression of efflux pumps (i.e., mexAB-oprM), deficiency or repression of the porin gene (oprD), alterations in the penicillin-binding proteins (PBPs), and chromosomal overexpression of cephalosporinase gene ampC (Van Nguyen et al., 2018; Gajdács, 2020; Xu et al., 2020). Moreover, resistance may be acquired by the selection of mutations in chromosomal genes or horizontal uptake of resistance determinants. However, carbapenem resistance has been most associated with production of carbapenemases, which include serine β-lactamases and metallo-β-lactamases (MβLs) (Polotto et al., 2012; Lupo et al., 2018), whereas high-risk global clones have been associated with MDR or extensively drug resistant (XDR) phenotypes. Currently, global P. aeruginosa high-risk clones include sequence types (STs) ST235, ST111, ST175, ST233, ST244, ST277, ST298, ST308, ST357, and ST654 (Del Barrio-Tofiño et al., 2020; Kocsis et al., 2021). Specifically, the ST277 has been sporadically reported in Asian, North American, and European countries, whereas in Brazil is highly prevalent (Gales et al., 2003; Hopkins et al., 2016; Del Barrio-Tofiño et al., 2020; Silveira et al., 2020; Kocsis et al., 2021). The success of the Brazilian endemic clone ST277 is associated with carbapenem resistance due to production of the MβL SPM-1 (Gales et al., 2003; Cipriano et al., 2007; da Fonseca et al., 2010; Nascimento et al., 2016; Silveira et al., 2020). Worryingly, SPM-1-producing P. aeruginosa have been identified in hospital sewage and hospital wastewater treatment plants (Fuentefria et al., 2009; Miranda et al., 2015), denoting potential to spread throughout the aquatic environment, enabling human exposure and transmission. However, although whole genome sequencing (WGS) of human SPM-1-positive isolates have been performed (Nascimento et al., 2016; Galetti et al., 2019), sequence data from environmental isolates have not been provided for comparative genomic studies. Based on WHO list priority pathogens criteria, which included pathogen mortality, hospital and environment transmissibility and limited treatment options, recognition and genomic characterization of critical priority pathogens is an essential first step to understanding their dynamic of acquisition/dissemination and ultimately to development of preventive intervention strategies (Hendriksen et al., 2019). In this study, we report genomic data and the virulence potential of carbapenem-resistant SPM-1-positive P. aeruginosa strains isolated from polluted urban rivers, in Brazil.
Materials and Methods
Pseudomonas aeruginosa Strains and Antimicrobial Susceptibility Profiles
During a Brazilian surveillance study (OneBR project) conducted to investigate the burden of antimicrobial resistance in impacted aquatic environments, two P. aeruginosa strains [Pa19 (ONE609) and Pa151 (ONE610)] were isolated from two different locations along the Tietê (TIET-04900; S 23° 31' 18'', W 46° 37' 52'', S 23° 27' 16'', and W 46° 54' 36'') and Pinheiros (PINH-04900; S 23° 31' 52'' and W 46° 44' 54'') Rivers in São Paulo, Brazil (Turano et al., 2016). Tietê River stretches through São Paulo state from east to west for approximately 1,100km, while Pinheiros River is a tributary of the Tietê River that runs 25km across the city. In this study, both strains were subjected to WGS for investigation and comparative genomic studies using five public sequences from nosocomial SPM-1-positive P. aeruginosa strains, previous reported (Silveira et al., 2014, 2020; Nascimento et al., 2016; Galetti et al., 2019). Susceptibility profiles were investigated by disk-diffusion method (CLSI, 2021).
Whole Genome Sequencing and Genomic Analysis
Genomic DNA of Pa19 and Pa151 were extracted using PureLink Quick Gel Extraction & PCR Purification Combo Kit (Life Technologies, Carlsbad, CA). The Illumina paired-end libraries were constructed using a Nextera XT DNA Library Preparation Kit (Illumina Inc.), according to the manufacturer’s guidelines. Whole genome sequencing was performed using an Illumina MiSeq platform with 300-bp read lengths. Reads were de novo assembled using SPAdes 3.13,1 and the resulting contigs were automatically annotated by NCBI Prokaryotic Genome Annotation Pipeline (PGAP) version 3.2.2 Antibiotic resistance genes were predicted using ResFinder 4.13 and the Comprehensive Antibiotic Resistance Database (CARD).4 Multi-locus Sequence Typing prediction was performed using MLST v.2.0.5 Heavy metal (HM) resistance genes were manually identified using the NCBI database6 and Geneious Prime version 2020.04 (Biomatters, New Zealand). Additionally, phage prediction was performed by Genome Detective Virus Tool software.7 The rmtD gene was detected and aligned by BLASTn (Alikhan et al., 2011) against the rmtD1 allele of the P. aeruginosa (PA0905 strain), recovered from a human patient (GenBank accession number. DQ914960). Genetic context analysis of blaSPM-1 and rmtD1 resistance genes of Pa151 were performed with BLASTn algorithm and manually curated using Geneious Prime version 2020.04 (Biomatters, New Zealand).
Moreover, virulence genes, efflux systems, and regulators were determined through the Virulence Factor Database.8 Serotype was predicted using Past 1.0.9 SNP-based phylogenetic analysis was performed by using Prokka 1.13.410 for pangenome annotation, followed by Roary 3.13.011 for core genome analysis. SNP-sites tool12 was used for SNPs extraction from the core gene alignment; whereas RAxML-NG version 0.9.013 for phylogenetic construction and a maximum likelihood tree based on SNP alignment. Additionally, comparative genomic analysis of P. aeruginosa sequences was performed by BRIG v.0.95 using the BLASTn algorithm and Island viewer 4.0.
All genomic analysis were based on comparison of sequences of environmental Pa151 (Pinheiros River, GenBank accession number: PHSS00000000.1) and Pa19 (Tietê River, GenBank accession number: PHST01000000) strains, against publically available genome sequences (data obtained by using 300 bp paired-end MiSeq sequencing) of clinical SPM-1-producing P. aeruginosa CCBH4851 (catheter tip, GenBank accession number CP021380.2), PA1088 (urine, GenBank accession number CP015001.1), PA11803 (bloodstream, GenBank accession number: CP015003.1), PA12117 (bloodstream, GenBank accession number: LVXB00000000.1) and PA7790 (tracheal aspirate, GenBank accession number: CP014999.1) strains, which were retrieved from NCBI GenBank database.14 For SNP-based analysis, the genome of the P. aeruginosa strain PAO1 (ST549) was used as reference (GenBank accession number: AE004091.2).
Virulence Potential of Carbapenem-Resistant P. aeruginosa Strains in the Galleria mellonella Larvae Model
The virulence potential of P. aeruginosa Pa19 and Pa151 strains was evaluated using the Galleria mellonella infection model (Tsai et al., 2016). In brief, groups of G. mellonella containing 10 larvae of nearly 0.25–0.35g (supplied by the Institute of Biomedical Sciences of the University of São Paulo, Brazil) were infected with 104CFU/ml of each strain per larvae, by injecting a 10μl aliquot in PBS, into the body of the larvae via the last left proleg, using a sterile ultra-fine needle syringe (Fuentes-Castillo et al., 2019). Survival was monitored every hour, for 96h. Two biological replicates and two experimental replicates were performed with a group of 10 larvae per strain, in each replicate. SPM-1-producing P. aeruginosa clinical strain PA1088 was used as comparative control (Toleman et al., 2002). Moreover, a control group inoculated with sterile PBS was used in each biological and experimental replication assay, in order to verify that the larvae would not be killed by physical trauma. Survival curves were plotted using the Kaplan-Meier method, whereas statistical analyses were performed by the log rank test with p <0.05 indicating statistical significance (OriginLab Software, Northampton, Massachusetts, United States).
Results
In this study, two carbapenemase (SPM-1)-producing P. aeruginosa ST277 (Pa19 and Pa151 strains) isolated from impacted urban rivers in São Paulo, Brazil, were sequenced. As this clone has been endemic in Brazilian hospitals, being also identified in migratory birds (Figure 1), we have additionally performed a comparative analysis with publically available genomes obtained from ST277 lineages from human infections.
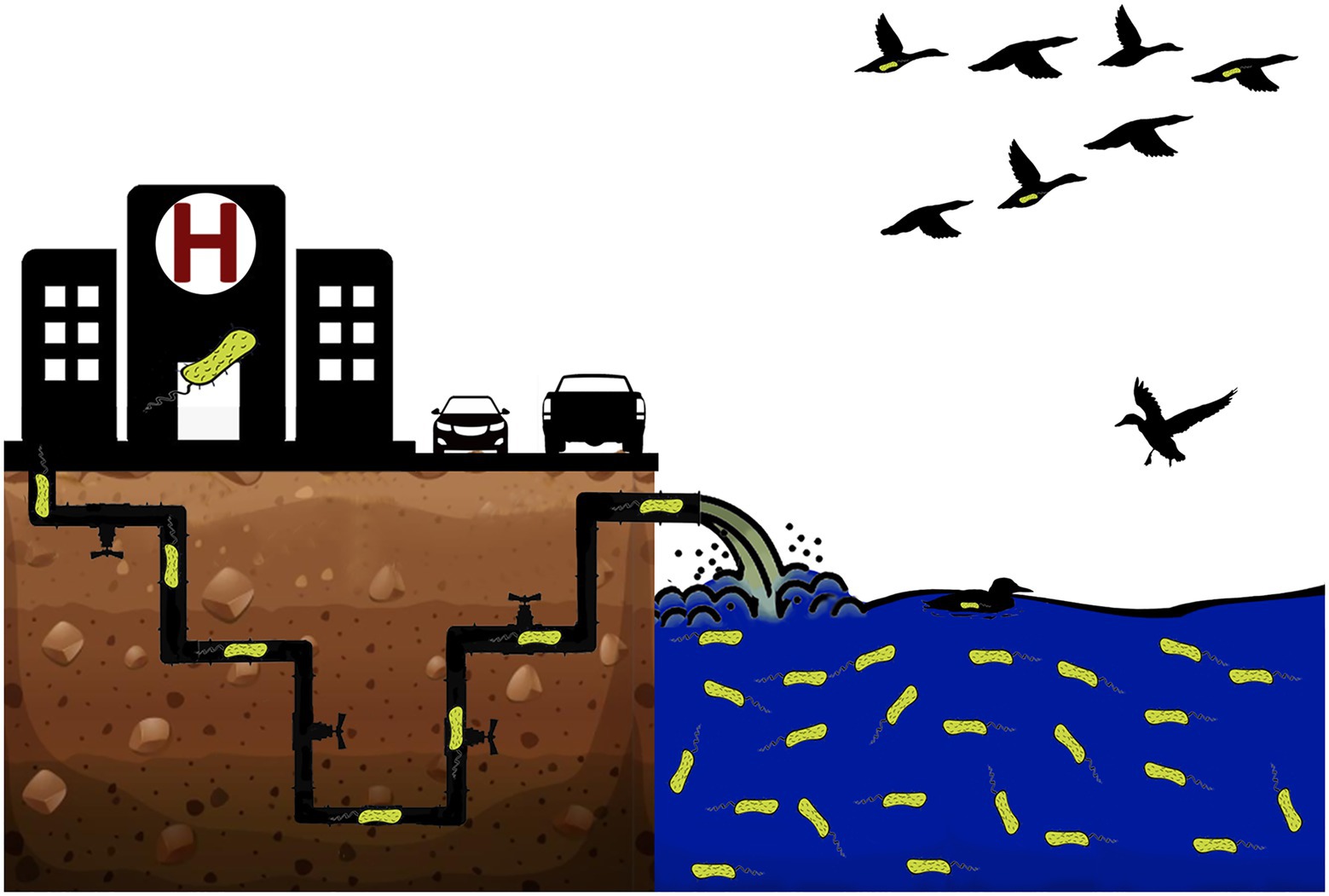
Figure 1. Schematic representation of hypothesis proposed for spread of carbapenemase (SPM-1)-producing Pseudomonas aeruginosa clone ST277 beyond the hospital, in Brazil, based on genomic data analyzed in this study.
Genome sequencing yielded a total of 968,818 and 473,825 paired-end reads assembled into 395 and 337 contigs, with 305 and 299x of coverage, to Pa19 and Pa151 strains, respectively. The genome size of Pa19 was calculated at 6,927,007bp, with a GC content of 67.8%, comprising 6,956 total genes, 60 tRNAs, three rRNAs, four ncRNAs, and 155 pseudogenes (accession number: PHST00000000.1). On the other hand, genome size of Pa151 was calculated at 6,799,801bp, with a GC content of 66.9%, comprising 6,747 total genes, 59 tRNAs, three rRNAs, four ncRNAs, and 123 pseudogenes (accession number: PHSS00000000.1). Genomic information of P. aeruginosa Pa19 and Pa151 strains are available on the OneBR platform15 under ONE609 and ONE610 ID numbers, respectively.
Environmental Pa19 and Pa151 strains displayed a MDR profile to ticarcillin-clavulanate, cefepime, ceftazidime, imipenem, meropenem, amikacin, gentamicin, nalidixic acid, ciprofloxacin, levofloxacin, and trimethoprim-sulfamethoxazole, and genomic analysis revealed a wide resistome to β-lactams (blaSPM-1, blaOXA-56, blaOXA-396, blaOXA-494, and blaPAO), aminoglycosides [aacA4, aadA7 and aph(3')-llb], fluoroquinolones [aac(6')lb-cr, and gyrA (T83I) and parC (S87L) point mutations], phenicols (cmx), sulphonamides (sul1), and fosfomycin (fosA), which was predicted in agreement with the phenotype. Additionally, Pa151 strain harbored the rmtD1 and catB7 genes related to aminoglycosides and chloramphenicol resistance, respectively (Figure 2). On the other hand, the crpP gene associated with fluoroquinolone resistance, was only identified in the Pa19 genome. Genes associated with resistance to heavy metal [copper (pcoABD)], and quaternary ammonium compounds (qacE, qacA, and sugE) were also identified in both environmental P. aeruginosa strains (Figure 2).
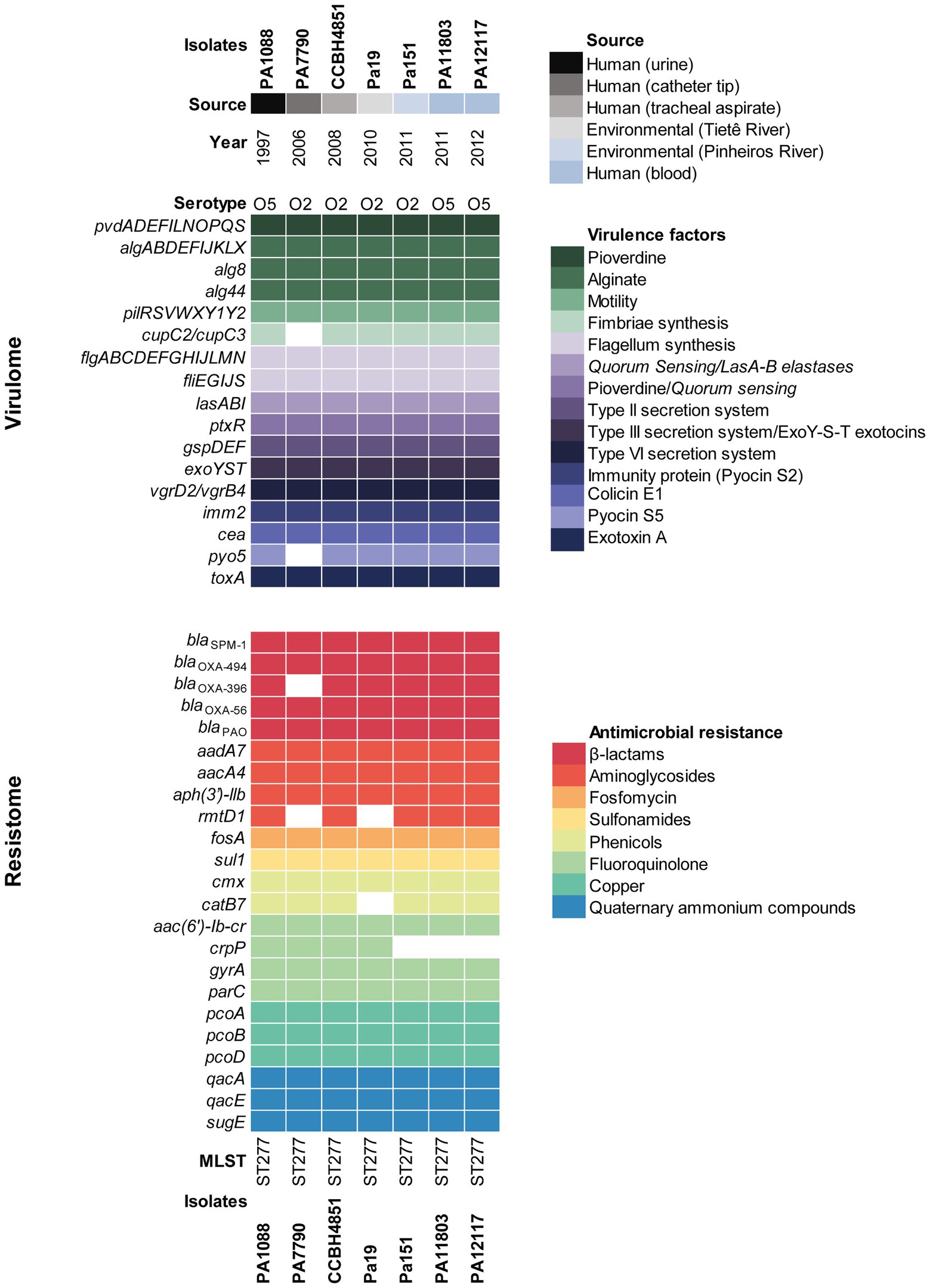
Figure 2. Heatmap showing the distribution of virulence and resistance genes in environmental and clinical SPM-1-positive P. aeruginosa strains of ST277 clone. Virulence genes are listed along with their functions and demarcated by colored squares. Resistome is demarcated by colored squares under genes names grouped by their antimicrobial resistance classes. Pseudomonas aeruginosa strains are indicated by colors displaying their source and year of isolation.
Virulome analysis of environmental Pa19 and Pa151 revealed a wide virulome. In fact, both lineages carried the quorum sensing (lasA, lasB, lasI, and ptxR), alginate (alg cluster), siderophore production (pvdA, pvdF, and pvdG), fimbriae (cup family), flagellum (flgABCDEFGHIJLMN) synthesis, immunity protein (pyo5, imm2), colicin (cea), types II (gspDEF), III (exoYST), and IV (vgrD2/vgrD4) secretion systems and exotoxin A (toxA) genes; whereas the O2 serotype was identified in both Pa19 and Pa151 environmental strains (Figure 2). In this regard, in vivo experiments using G. mellonella larvae showed that both Pa19 and Pa151 strains killed 100% of the larvae at 24h post-infection, similarly to what was observed with the clinical SPM-1-producing P. aeruginosa PA1088 strain isolated from a case of urinary tract infection (Figure 3).
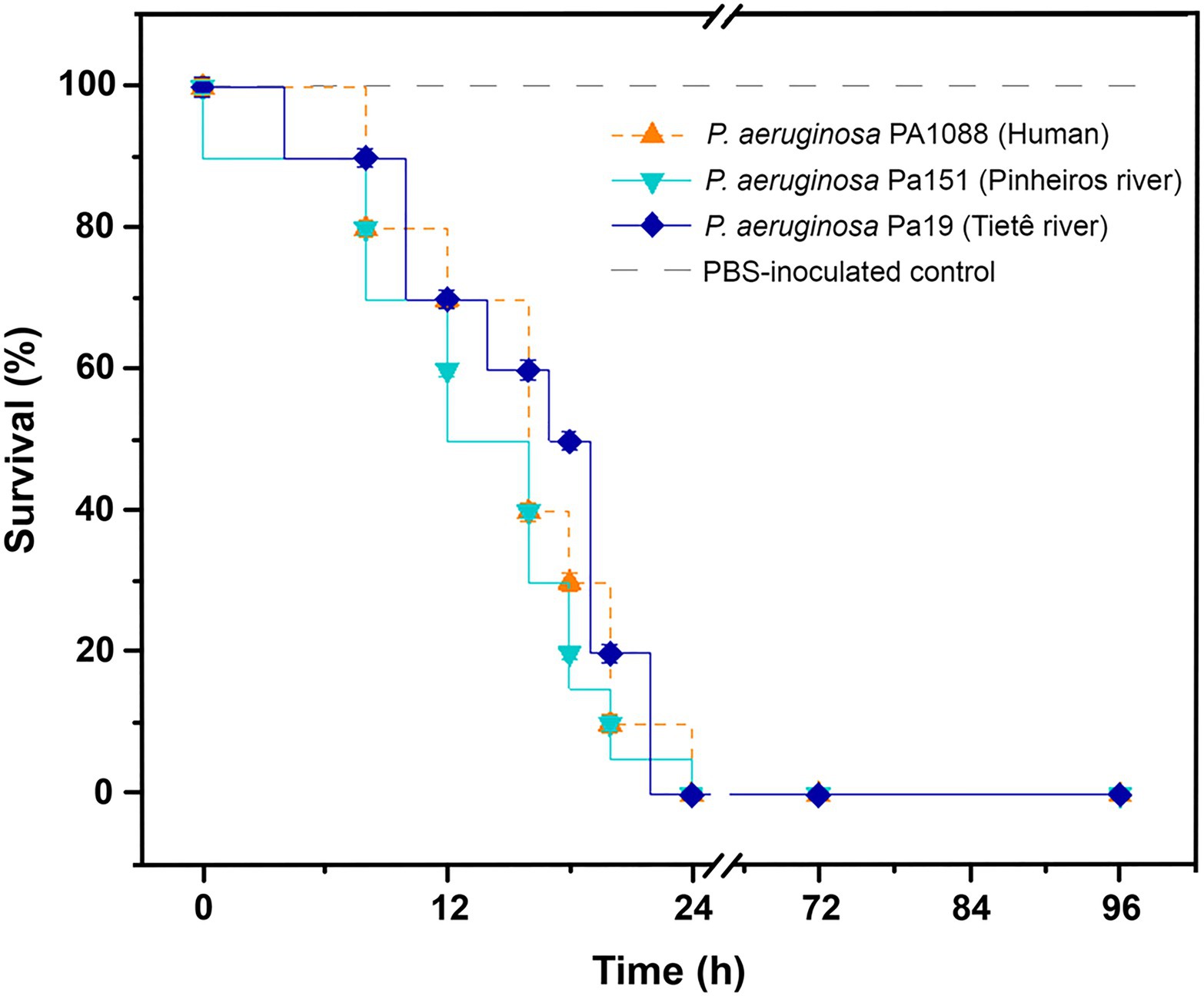
Figure 3. Virulent behavior of SPM-1-producing P. aeruginosa isolates. Kaplan-Meier survival curves of Galleria mellonella infected with 104CFU/larva of P. aeruginosa Pa19 strain (dark-blue line), P. aeruginosa Pa151 strain (blue line), and P. aeruginosa PA1088 (orange line). Environmental Pa19 and Pa151 strains, and the clinical PA1088 strain killed 100% of larvae at 24h post-infection. PBS-inoculated control group (light-gray dashed line) presented 100% of survival. For each strain, groups containing 10 G. mellonella larvae in each replicate were evaluated in two biological and experimental independent assays.
Overall comparison of five human and two environmental ST277 genomes using BLAST Ring Image Generator (BRIG) revealed high nucleotide sequence similarities among P. aeruginosa strains, even for aquatic isolates recovered at least 13years after the first clinical isolate (Toleman et al., 2002). Furthermore, SNP-based phylogenetic analysis revealed that both Pa19 and Pa151 environmental strains were closely related (>94% identity) to all human SPM-1-producing P. aeruginosa isolates (Supplementary Table S1). However, missing regions at position 5.5Mbp, named as GI-I, in environmental Pa151 and clinical PA11803 and PA12117 genomes, were identified (Figure 4). In this regard, we observed genes encoding the following proteins: integrating conjugative element protein (pill), type II secretion system protein, replicative DNA helicase (dnaB), nucleoid-associated protein YejK (yejK), NADH dehydrogenase (ndh), cell division protein ZapE (zapE), ParA family protein (parA), plasmid stabilization protein ParE (parE), integrating conjugative element protein, DNA topoisomerase I (topA), pyocin S5 (pyoS5), TetR family transcriptional regulator (tetR), conjugal transfer protein TraG (traG), regulatory protein GemA (gemA), conjugative coupling factor TraD (traD), his-Xaa-Ser repeat protein HxsA (hxsA), his-Xaa-Ser system radical SAM maturase HxsB (hxsB), his-Xaa-Ser system radical SAM maturase HxsC (hxsC), his-Xaa-Ser system protein HxsD (hxsD), chaperone protein ClpB (clpB), and genes encoding for membrane proteins, transcriptional regulator, CRISPR-associated proteins, type II secretion system protein, phage tail sheath subtilisin-like, tail fiber protein, phage tail tape measure protein, and phage head morphogenesis protein.
Schematic representations of the genetic context surrounding blaSPM-1 genes in the environmental P. aeruginosa PA151 strain is presented in Figure 5A. The blaSPM-1 was flanked by a ~4.8kbp region composed of the IS91-blaSPM-1-groEL-IS91 array. The presence of IS elements is related to horizontal gene transfer, whereas the groEL encodes for a heat-shock chaperon. Additionally, we also detected the traG (encoding a conjugal transfer protein), eexN (encoding the entry exclusion protein), traR (transcriptional regulator), bcr1 (bicyclomycin resistance), virD2 (gene encoding a relaxase), and hypothetical proteins. In Figure 5B is presented the genetic context surrounding rmtD1 gene in PA151 strain. The rmtD1 was flanked by a ~7.3kbp region composed of the IS91-rmtD1-tgt-groEL-IS91 array. In addition, aacA4, blaOXA-56, aadA7, and qacEΔ1 genes were located on a class 1 integron. Moreover, cmx and sul1 resistance genes, that encodes for chloramphenicol and sulphonamide resistance, respectively, were also identified along with genes encoding hypothetical proteins, transposase, IS110, IS481, and IS3 mobile elements.
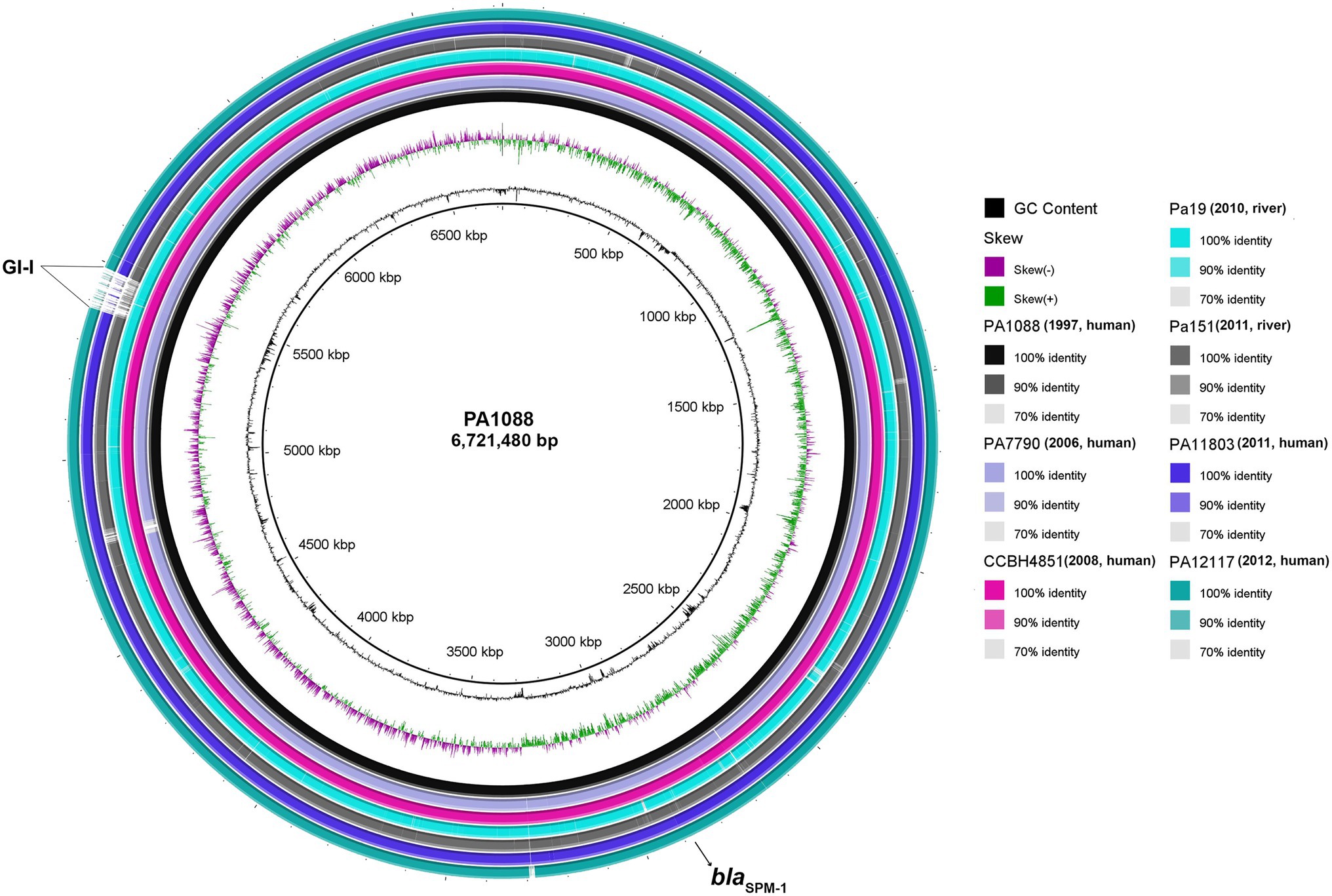
Figure 4. Circular genome maps of SPM-1-producing P. aeruginosa belonging to ST277. Circular maps were built by BLAST Ring Image Generator (BRIG) using seven P. aeruginosa genomes. All genomes were represented as individual rings and compared against the reference genome PA1088 (GenBank accession number: CP015001.1). The blaSPM-1 gene is indicated by a black arrow. Furthermore, several genes associated with DNA replication/repair/regulatory/defense and membrane proteins were identified in the major genomic island, indicated as GI-I.
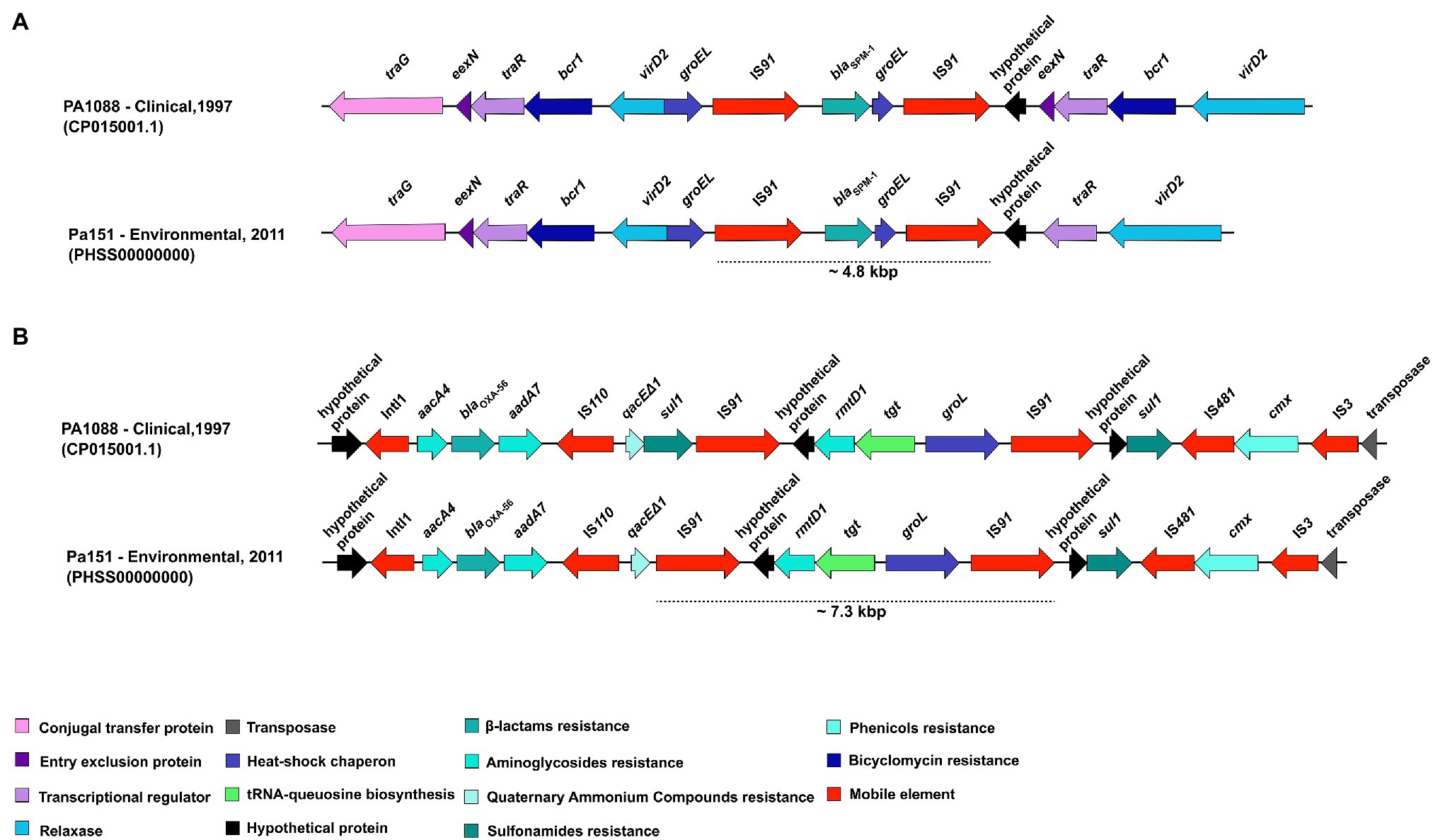
Figure 5. Overall comparison of genetic context of blaSPM-1 and rmtD1 genes carried by clinical and environmental P. aeruginosa strains belonging to ST277. (A) The blaSPM-1 was flanked by a ~4.8kbp region composed of IS91-blaSPM-1-groEL-IS91. (B) The rmtD1 was flanked by a ~7.3kbp region composed of the IS91-rmtD1-tgt-groEL-IS91. In addition, aacA4, aadA7, blaOXA-56, qacEΔ1, sul1, and cmx resistance genes were also identified along with genes encoding for hypothetical proteins, transposase; as well as IS110, IS481, and IS3 mobile genetics elements.
Discussion
Carbapenem-resistant P. aeruginosa are critical-priority pathogens associated with high mortality and morbidity (Georgescu et al., 2016; Tacconelli et al., 2018; Pang et al., 2019). In this regard, one of the major concerns has been the successful expansion and rapid spread of high-risk clones. In Brazil, the metallo-β-lactamase (SPM-1)-producing P. aeruginosa ST277 clone has gained significant attention, due to its endemicity status and further identification in migratory birds and polluted environments (Gales et al., 2003; Nascimento et al., 2016; Turano et al., 2016; Martins et al., 2018).
Worryingly, previous studies have also reported the occurrence of carbapenemase (KPC-2)-producing Klebsiella pneumoniae belonging to the clonal group CG258 and OXA-23-positive A. baumannii ST79 in the Tietê River (Oliveira et al., 2014; Turano et al., 2016), supporting an anthropogenic trend, most likely due to hospital wastewater discharge and domestic wastewaters effluents (Nascimento et al., 2017; Bartley et al., 2019; Böger et al., 2021; Popa et al., 2021). Therefore, aquatic environment could play an important role in the widespread of critical pathogens (Devarajan et al., 2017). In fact, polluted rivers could be contributing for colonization of local and migratory fauna (Martins et al., 2018; Narciso et al., 2020).
In order to elucidate the genomic aspects associate with the environmental dissemination of healthcare-associated P. aeruginosa ST277, we performed a comparative genomic analysis, extracting clinically relevant information (i.e., resistome, virulome, and phylogenomic). Interestingly, although the strains were isolated in different years (1997–2012), we observed that clinical and environmental SPM-1-producing P. aeruginosa strains share a common resistome and virulome.
Although, oral antibiotics have been successfully used in the treatment of bacterial infection, for P. aeruginosa few therapeutic options are available, being restricted to some fluoroquinolones, including ciprofloxacin, levofloxacin, and prulifloxacin, which are given alone or in combinations with a second intravenously or inhaled anti-pseudomonal antibiotic such as β-lactams (piperacillin/tazobactam, ceftolozane/tazobactam, ceftazidime, cefepime, or carbapenems) and/or aminoglycosides (tobramycin, amikacin, or gentamicin) (Tümmler, 2019; Ibrahim et al., 2020; Nisly et al., 2020). However, under a clinical perspective, even co-resistance to carbapenems and aminoglycosides in ST277 have already been reported and limited therapeutic options. This resistance profile is mediated by blaSPM-1 and rmtD genes, respectively (Doi et al., 2007). Strikingly, in some ST277, including environmental (Pa19) and human (PA7790) lineages, the rmtD gene was not found. On the other hand, the rmtD1 identified in the environmental Pa151 strain, displayed 100% identity to the rmtD1 gene from P. aeruginosa PA0905 strain, recovered from a human patient in 2005, in Brazil (Doi et al., 2007). The rmtD1 was subsequently identified in K. pneumoniae and other Enterobacterales in Latin America, Europe, and North America (Bueno et al., 2016). Since acquisition of this gene has been linked to transposition events (Doi et al., 2007; Nascimento et al., 2016), most likely genomic plasticity of P. aeruginosa has led to the dissemination of rmtD+ and rmtD-ST277 lineages (Silveira et al., 2020). In Brazil, occurrence of rmtD has also been documented in Escherichia coli and K. pneumoniae (Yamane et al., 2008; Leigue et al., 2015).
The blaSPM-1, IS91-blaSPM-1-groEL-IS91 gene array has been previously identified within a Tn4371-like integrative and conjugative element (ICETn43716061) considered stable in the chromosome loci of P. aeruginosa ST277 strains recovered from humans and animals (Fonseca et al., 2015; Nascimento et al., 2016). Since ICEs are genetic mobile platforms that play an important role during bacterial evolution, they are overlooked as vectors in the spread and resistance emergence in many bacterial species (Fonseca and Vicente, 2016). Moreover, the genetic context of rmtD1 (IS91-rmtD1-tgt-groEL-IS91) identified in the environmental strain was similar to previous descriptions, where the presence of the rmtD gene in clinical isolates was associated to the TnAs3 transposon (Fonseca et al., 2015; Nascimento et al., 2016).
In human and aquatic P. aeruginosa ST277 isolates the resistome was not restricted to antibiotics, and the presence of genes conferring tolerance to copper and QAC biocides was further detected. Currently, there is a growing concern about biocides that pollute aquatic environments, especially QACs, since these compounds are widely used in domiciliary and hospital settings, as disinfectants, soaps, toothpastes, and mouthwash formulations (Zubris et al., 2017; Fuentes-Castillo et al., 2020). Consequently, ecosystems impacted by HM and biocides could favor the selection and persistence of high-risk clones harboring a broad resistome (Baker-Austin et al., 2006; Zhao et al., 2012; Kim et al., 2018).
Although a limitation of this study was the lack of a known highly virulent P. aeruginosa to be used as a positive control in the in vivo assay; we observed that the virulent behavior of environmental strains was identical to clinical strains. Indeed, a wide virulome was also predicted in human and environmental P. aeruginosa ST277 lineages, denoting a pathogenic potential, as demonstrated in the G. mellonella infection model. Lipopolysaccharide (LPS) O-antigen, type IV pili, and flagella are components of the external cell wall structure of P. aeruginosa and play important roles in the early stage of colonization, persistence, and bacterial pathogenesis (Hauser, 2011; Behzadi et al., 2021). Furthermore, O-antigen is an important virulence factor in P. aeruginosa used for the detection of MDR/XDR high-risk clones (Del Barrio-Tofiño et al., 2019). Strikingly, among clinical strains were identified the serotypes O5 and O2. The latter was also identified among environment strains. Both serotypes have been associated with acute and chronic infections (Lu et al., 2014; Li et al., 2018).
Type secretion systems (TSSs) are mechanisms by which bacteria translocate a set of toxins into the cytosol of host cells and/or to the extracellular medium (Abby et al., 2016). Pseudomonas aeruginosa is known to have five TSSs, of which Types I (T1SS), II (T2SS), and III (T3SS) are involved in the virulence of this pathogen. Several studies have linked these TSSs with poor outcomes of patients with acute respiratory diseases (i.e., pneumonia), with T3SS being one of the most clinically relevant virulence determinants (Hauser, 2011; McMackin et al., 2019; Sarges et al., 2020). In this context, we detected ExoTSY exotoxins-encoding genes in both clinical and environmental strains. ExoTSY exotoxins are secreted by T3SS and reported to be involved in lung injury, pulmonary-vascular barrier disruption, and end-organ dysfunction in chronic infections, mainly in CF patients; as well as with mortality in animal models (Lu et al., 2014; Sarges et al., 2020; Jurado-Martín et al., 2021). Interestingly, the toxA gene (exotoxin A), which is present in the most clinically P. aeruginosa strains (Khosravi et al., 2016) was also identified in environmental strains. Exotoxin A has been associated with tissue damage related to poor outcomes of burn patients (Khosravi et al., 2016). In fact, the broad virulome harbored by P. aeruginosa ST277 seems to be associated with a remarkable ability to adapt to different human and non-human conditions (Jurado-Martín et al., 2021).
In brief, from comparative analysis, our data revealed that Pa19 and Pa151 environmental strains presented slight variations when compared against clinical strains, suggesting a high degree of genetic conservation, regardless isolation data and exposition to contaminants (antibiotics and biocides residues) present in the polluted aquatic environments.
Conclusion
In summary, we report genomic comparative data of antimicrobial-resistant P. aeruginosa isolated from aquatic environments in Brazil. The presence of SPM-1-producing P. aeruginosa ST277 in urban rivers could be associated with hospital effluents, since SNP-based phylogenomics showed high nucleotide sequence similarity between clinical and environmental genomes. Additionally, wide resistome and virulome have been conserved in environmental isolates, denoting that critical priority P. aeruginosa of the high-risk ST277 has successfully expanded beyond the hospital. Therefore, genomic surveillance is essential to rapidly identify and prevent the spread of WHO critical priority clones with One Health implications.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author Contributions
FE, BC, HF, and BF performed the data analysis. FE, BC, QM, AC-A, and DF-C conducted the experiments. NL supervised the experiments and designed and coordinated the project. FE, BC, and NL wrote, reviewed, and edited the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Bill & Melinda Gates Foundation (Grand Challenges Explorations Brazil OPP1193112). Under the grant conditions of the Foundation, a Creative Commons Attribution 4.0 Generic License has already been assigned to the Author Accepted Manuscript version that might arise from this submission. Additionally, this study was supported by the Fundação de Amparo à Pesquisa do Estado de São Paulo (2020/08224-9) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (AMR 443819/2018-1 and 433128/2018-6). FE is a research fellow of FAPESP (2019/15578-4). BC and HF are researcher fellows of CAPES (88882.333054/2019-01 and 88887.506496/2020-00). DF-C is a researcher fellow of Comisión Nacional de Investigación Científica y Tecnológica (CONICYT BCH 72170436). AC-A and BF are researcher fellows of PNPD/CAPES (88887.463414/2019-00 and 88887.358057/2019-00). NL is research fellow of CNPq (312249/2017-9).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer AB declared a shared affiliation with no collaboration with several of the authors FE, HF, BF, and NL to the handling editor at the time of the review.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors are grateful to Bill & Melinda Gates Foundation, FAPESP, CAPES, and CNPq. We also thank Cefar Diagnóstica Ltda. (São Paulo, Brazil) and Louise Cerdeira for kindly supplying the antibiotic disks for susceptibility testing and for assisting with BRIG software, respectively.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.701921/full#supplementary-material
Footnotes
1. ^https://github.com/ablab/spades
2. ^https://www.ncbi.nlm.nih.gov/genome/annotation_prok/
3. ^https://cge.cbs.dtu.dk/services/ResFinder
4. ^https://card.mcmaster.ca/analyze/rgi
5. ^https://cge.cbs.dtu.dk/services/MLST/
6. ^https://www.ncbi.nlm.nih.gov/Traces/wgs/
7. ^https://www.genomedetective.com/app/typingtool/virus/
9. ^https://cge.cbs.dtu.dk/services/PAst/
10. ^https://github.com/tseemann/prokka
11. ^https://github.com/sanger-pathogens/Roary
12. ^https://github.com/sanger-pathogens/snp-sites
13. ^https://github.com/stamatak/standard-RAxML
References
Abby, S. S., Cury, J., Guglielmini, J., Néron, B., Touchon, M., and Rocha, E. P. (2016). Identification of protein secretion systems in bacterial genomes. Sci. Rep. 6:23080. doi: 10.1038/srep23080
Alikhan, N. F., Petty, N. K., Zakour, N. L. B., and Beatson, S. A. (2011). BLAST ring image generator (BRIG): simple prokaryote genome comparisons. BMC Genomics 12:402. doi: 10.1186/1471-2164-12-402
Babu, R. N., Mutters, N. T., Marasca, G., Conti, M., Sifakis, F., Vuong, C., et al. (2020). Mandatory surveillance and outbreaks reporting of the WHO priority pathogens for research & discovery of new antibiotics in European countries. Clin. Microbiol. Infect. 26, 943.e1–943.e6. doi: 10.1016/j.cmi.2019.11.020
Baker-Austin, C., Wright, M. S., Stepanauskas, R., and McArthur, J. V. (2006). Co-selection of antibiotic and metal resistance. Trends Microbiol. 14, 176–182. doi: 10.1016/j.tim.2006.02.006
Bartley, P. S., Domitrovic, T. N., Moretto, V. T., Santos, C. S., Ponce-Terashima, R., Reis, M. G., et al. (2019). Antibiotic resistance in Enterobacteriaceae from surface waters in urban Brazil highlights the risks of poor sanitation. Am. J. Trop. Med. Hyg. 100, 1369–1377. doi: 10.4269/ajtmh.18-0726
Behzadi, P., Baráth, Z., and Gajdács, M. (2021). It’s not easy being green: a narrative review on the microbiology, virulence and therapeutic prospects of multidrug-resistant Pseudomonas aeruginosa. Antibiotics 10:42. doi: 10.3390/antibiotics10010042
Bobrov, A. G., Getnet, D., Swierczewski, B., Jacobs, A., Medina-Rojas, M., Tyner, S., et al. (2021). Evaluation of Pseudomonas aeruginosa pathogenesis and therapeutics in military-relevant animal infection models. APMIS doi: 10.1111/apm.13119 [Epub ahead of print]
Böger, B., Surek, M., Vilhena, R. O., Fachi, M. M., Junkert, A. M., Santos, J. M., et al. (2021). Occurrence of antibiotics and antibiotic resistant bacteria in subtropical urban rivers in Brazil. J. Hazard Mater. 402:123448. doi: 10.1016/j.jhazmat.2020.123448
Bueno, M. F., Francisco, G. R., de Oliveira Garcia, D., and Doi, Y. (2016). Complete sequences of multidrug resistance plasmids bearing rmtD1 and rmtD2 16S rRNA methyltransferase genes. Antimicrob. Agents Chemother. 60, 1928–1931. doi: 10.1128/AAC.02562-15
Cipriano, R., Vieira, V. V., Fonseca, É. L., Rangel, K., Freitas, F. S., and Vicente, A. C. P. (2007). Coexistence of epidemic colistin-only-sensitive clones of Pseudomonas aeruginosa, including the blaSPM clone, spread in hospitals in a Brazilian amazon city. Microb. Drug Resist. 13, 142–146. doi: 10.1089/mdr.2007.708
CLSI (2021). Performance Standards for Antimicrobial Susceptibility Testing. 31st Edn. CLSI supplement M100. Clinical and Laboratory Standards Institute.
da Fonseca, É. L., dos Santos Freitas, F., and Vicente, A. C. P. (2010). The colistin-only-sensitive Brazilian Pseudomonas aeruginosa clone SP (sequence type 277) is spread worldwide. Antimicrob. Agents Chemother. 54:2743. doi: 10.1128/AAC.00012-10
Del Barrio-Tofiño, E., López-Causapé, C., and Oliver, A. (2020). Pseudomonas aeruginosa epidemic high-risk clones and their association with horizontally acquired β-lactamases: 2020 update. Int. J. Antimicrob. Agents 56:106196. doi: 10.1016/j.ijantimicag.2020.106196
Del Barrio-Tofiño, E., Sánchez-Diener, I., Zamorano, L., Cortes-Lara, S., López-Causapé, C., Cabot, G., et al. (2019). Association between Pseudomonas aeruginosa O-antigen serotypes, resistance profiles and high-risk clones: results from a Spanish nationwide survey. J. Antimicrob. Chemother. 74, 3217–3220. doi: 10.1093/jac/dkz346
Devarajan, N., Köhler, T., Sivalingam, P., van Delden, C., Mulaji, C. K., Mpiana, P. T., et al. (2017). Antibiotic resistant Pseudomonas spp. in the aquatic environment: a prevalence study under tropical and temperate climate conditions. Water Res. 115, 256–265. doi: 10.1016/j.watres.2017.02.058
Doi, Y., de Oliveira Garcia, D., Adams, J., and Paterson, D. L. (2007). Coproduction of novel 16S rRNA methylase RmtD and metallo-β-lactamase SPM-1 in a panresistant Pseudomonas aeruginosa isolate from Brazil. Antimicrob. Agents Chemother. 51, 852–856. doi: 10.1128/AAC.01345-06
Fernández-Barat, L., Ferrer, M., De Rosa, F., Gabarrús, A., Esperatti, M., Terraneo, S., et al. (2017). Intensive care unit-acquired pneumonia due to Pseudomonas aeruginosa with and without multidrug resistance. J. Infect. 74, 142–152. doi: 10.1016/j.jinf.2016.11.008
Fonseca, E. L., Marin, M. A., Encinas, F., and Vicente, A. C. (2015). Full characterization of the integrative and conjugative element carrying the metallo-β-lactamase blaSPM-1 and bicyclomycin bcr1 resistance genes found in the pandemic Pseudomonas aeruginosa clone SP/ST277. J. Antimicrob. Chemother. 70, 2547–2550. doi: 10.1093/jac/dkv152
Fonseca, E. L., and Vicente, A. C. (2016). Commentary: clinical utilization of genomics data produced by the international Pseudomonas aeruginosa consortium. Front. Microbiol. 7:770. doi: 10.3389/fmicb.2016.00770
Fuentefria, D. B., Ferreira, A. E., Gräf, T., and Corção, G. (2009). Spread of metallo-β-lactamases: screening reveals the presence of a blaSPM-1 gene in hospital sewage in southern Brazil. Braz. J. Microbiol. 40, 82–85. doi: 10.1590/S1517-83822009000100013
Fuentes-Castillo, D., Esposito, F., Cardoso, B., Dalazen, G., Moura, Q., Fuga, B., et al. (2020). Genomic data reveal international lineages of critical priority Escherichia coli harbouring wide resistome in Andean condors (Vultur gryphus Linnaeus, 1758). Mol. Ecol. 29, 1919–1935. doi: 10.1111/mec.15455
Fuentes-Castillo, D., Farfán-López, M., Esposito, F., Moura, Q., Fernandes, M. R., Lopes, R., et al. (2019). Wild owls colonized by international clones of extended-spectrum β-lactamase (CTX-M)-producing Escherichia coli and Salmonella infantis in the southern cone of America. Sci. Total Environ. 674, 554–562. doi: 10.1016/j.scitotenv.2019.04.149
Gajdács, M. (2020). Carbapenem-resistant but cephalosporin-susceptible Pseudomonas aeruginosa in urinary tract infections: opportunity for colistin sparing. Antibiotics 9:153. doi: 10.3390/antibiotics9040153
Gales, A. C., Menezes, L. C., Silbert, S., and Sader, H. S. (2003). Dissemination in distinct Brazilian regions of an epidemic carbapenem-resistant Pseudomonas aeruginosa producing SPM metallo-β-lactamase. J. Antimicrob. Chemother. 52, 699–702. doi: 10.1093/jac/dkg416
Galetti, R., Andrade, L. N., Varani, A. M., and Darini, A. L. C. (2019). SPM-1-producing Pseudomonas aeruginosa ST277 carries a chromosomal pack of acquired resistance genes: an example of high-risk clone associated with ‘intrinsic resistome’. J. Glob. Antimicrob. Resist. 16, 183–186. doi: 10.1016/j.jgar.2018.12.009
Georgescu, M., Gheorghe, I., Curutiu, C., Lazar, V., Bleotu, C., and Chifiriuc, M. C. (2016). Virulence and resistance features of Pseudomonas aeruginosa strains isolated from chronic leg ulcers. BMC Infect. Dis. 16:92. doi: 10.1186/s12879-016-1396-3
Hauser, A. R. (2011). Pseudomonas aeruginosa: so many virulence factors, so little time. Crit. Care Med. 39, 2193–2194. doi: 10.1097/CCM.0b013e318221742d
Hendriksen, R. S., Munk, P., Njage, P., van Bunnik, B., McNally, L., Lukjancenko, O., et al. (2019). Global monitoring of antimicrobial resistance based on metagenomics analyses of urban sewage. Nat. Commun. 10:1124. doi: 10.1038/s41467-019-08853-3
Hopkins, K. L., Meunier, D., Findlay, J., Mustafa, N., Parsons, H., Pike, R., et al. (2016). SPM-1 metallo-β-lactamase-producing Pseudomonas aeruginosa ST277 in the UK. J. Med. Microbiol. 65, 696–697. doi: 10.1099/jmm.0.000269
Ibrahim, D., Jabbour, J. F., and Kanj, S. S. (2020). Current choices of antibiotic treatment for Pseudomonas aeruginosa infections. Curr. Opin. Infect. Dis. 33, 464–473. doi: 10.1097/QCO.0000000000000677
Jean, S. S., Chang, Y. C., Lin, W. C., Lee, W. S., Hsueh, P. R., and Hsu, C. W. (2020). Epidemiology, treatment, and prevention of nosocomial bacterial pneumonia. J. Clin. Med. 9:275. doi: 10.3390/jcm9010275
Jurado-Martín, I., Sainz-Mejías, M., and McClean, S. (2021). Pseudomonas aeruginosa: an audacious pathogen with an adaptable arsenal of virulence factors. Int. J. Mol. Sci. 22:3128. doi: 10.3390/ijms22063128
Khosravi, A. D., Shafie, F., Montazeri, E. A., and Rostami, S. (2016). The frequency of genes encoding exotoxin A and exoenzyme S in Pseudomonas aeruginosa strains isolated from burn patients. Burns 42, 1116–1120. doi: 10.1016/j.burns.2016.02.012
Kim, M., Weigand, M. R., Oh, S., Hatt, J. K., Krishnan, R., Tezel, U., et al. (2018). Widely used benzalkonium chloride disinfectants can promote antibiotic resistance. Appl. Environ. Microbiol. 84, e01201–e01218. doi: 10.1128/AEM.01201-18
Kocsis, B., Gulyás, D., and Szabó, D. (2021). Diversity and distribution of resistance markers in Pseudomonas aeruginosa international high-risk clones. Microorganisms 9:359. doi: 10.3390/microorganisms9020359
Leigue, L., Warth, J. F., Melo, L. C., Silva, K. C., Moura, R. A., Barbato, L., et al. (2015). MDR ST2179-CTX-M-15 Escherichia coli co-producing RmtD and AAC(6′)-Ib-cr in a horse with extraintestinal infection, Brazil. J. Antimicrob. Chemother. 70, 1263–1265. doi: 10.1093/jac/dku520
Li, H., Du, Y., Qian, C., Li, L., Jiang, L., Jiang, X., et al. (2018). Establishment of a suspension array for Pseudomonas aeruginosa O-antigen serotyping. J. Microbiol. Methods 155, 59–64. doi: 10.1016/j.mimet.2018.11.006
Lu, Q., Eggimann, P., Luyt, C. E., Wolff, M., Tamm, M., François, B., et al. (2014). Pseudomonas aeruginosa serotypes in nosocomial pneumonia: prevalence and clinical outcomes. Crit. Care 18:R17. doi: 10.1186/cc13697
Lupo, A., Haenni, M., and Madec, J. Y. (2018). Antimicrobial resistance in Acinetobacter spp. and Pseudomonas spp. Microbiol. Spectr. 6. doi: 10.1128/microbiolspec.ARBA-0007-2017
Martins, W. M. B. S., Narciso, A. C., Cayô, R., Santos, S. V., Fehlberg, L. C. C., Ramos, P. L., et al. (2018). SPM-1-producing Pseudomonas aeruginosa ST277 clone recovered from microbiota of migratory birds. Diagn. Microbiol. Infect. Dis. 90, 221–227. doi: 10.1016/j.diagmicrobio.2017.11.003
McMackin, E. A. W., Djapgne, L., Corley, J. M., and Yahr, T. L. (2019). Fitting pieces into the puzzle of Pseudomonas aeruginosa type III secretion system gene expression. J. Bacteriol. 201, e00209–e00219. doi: 10.1128/JB.00209-19
Miranda, C. C., de Filippis, I., Pinto, L. H., Coelho-Souza, T., Bianco, K., Cacci, L. C., et al. (2015). Genotypic characteristics of multidrug-resistant Pseudomonas aeruginosa from hospital wastewater treatment plant in Rio de Janeiro, Brazil. J. Appl. Microbiol. 118, 1276–1286. doi: 10.1111/jam.12792
Narciso, A. C., Martins, W., Almeida, L., Cayô, R., Santos, S. V., Ramos, P. L., et al. (2020). Healthcare-associated carbapenem-resistant OXA-72-producing Acinetobacter baumannii of the clonal complex CC79 colonizing migratory and captive aquatic birds in a Brazilian zoo. Sci. Total Environ. 726:138232. doi: 10.1016/j.scitotenv.2020.138232
Nascimento, T., Cantamessa, R., Melo, L., Fernandes, M. R., Fraga, E., Dropa, M., et al. (2017). International high-risk clones of Klebsiella pneumoniae KPC-2/CC258 and Escherichia coli CTX-M-15/CC10 in urban lake waters. Sci. Total Environ. 598, 910–915. doi: 10.1016/j.scitotenv.2017.03.207
Nascimento, A. P. B., Ortiz, M. F., Martins, W. M. B. S., Morais, G. L., Fehlberg, L. C. C., Almeida, L. G. P., et al. (2016). Intraclonal genome stability of the metallo-β-lactamase SPM-1-producing Pseudomonas aeruginosa ST277, an endemic clone disseminated in Brazilian hospitals. Front. Microbiol. 7:1946. doi: 10.3389/fmicb.2016.01946
Nisly, S. A., McClain, D. L., Fillius, A. G., and Davis, K. A. (2020). Oral antibiotics for the treatment of gram-negative bloodstream infections: a retrospective comparison of three antibiotic classes. J. Glob. Antimicrob. Resist. 20, 74–77. doi: 10.1016/j.jgar.2019.07.026
Oliveira, S., Moura, R. A., Silva, K. C., Pavez, M., McCulloch, J. A., Dropa, M., et al. (2014). Isolation of KPC-2-producing Klebsiella pneumoniae strains belonging to the high-risk multiresistant clonal complex 11 (ST437 and ST340) in urban rivers. J. Antimicrob. Chemother. 69, 849–852. doi: 10.1093/jac/dkt431
Pang, Z., Raudonis, R., Glick, B. R., Lin, T. J., and Cheng, Z. (2019). Antibiotic resistance in Pseudomonas aeruginosa: mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 37, 177–192. doi: 10.1016/j.biotechadv.2018.11.013
Pliska, N. N. (2020). Pseudomonas aeruginosa as the main causative agent of osteomyelitis and its susceptibility to antibiotics. Drug Res. 70, 280–285. doi: 10.1055/a-1150-2372
Polotto, M., Casella, T., de Lucca Oliveira, M. G., Rúbio, F. G., Nogueira, M. L., de Almeida, M. T. G., et al. (2012). Detection of P. aeruginosa harboring blaCTX-M-2, blaGES-1 and blaGES-5, blaIMP-1 and blaSPM-1 causing infections in Brazilian tertiary-care hospital. BMC Infect. Dis. 12:176. doi: 10.1186/1471-2334-12-176
Popa, L. I., Gheorghe, I., Barbu, I. C., Surleac, M., Paraschiv, S., Măruţescu, L., et al. (2021). Multidrug resistant Klebsiella pneumoniae ST101 clone survival chain from inpatients to hospital effluent after chlorine treatment. Front. Microbiol. 11:610296. doi: 10.3389/fmicb.2020.610296
Rosales-Reyes, R., Vargas-Roldán, S. Y., Lezana-Fernández, J. L., and Santos-Preciado, J. I. (2021). Pseudomonas aeruginosa: genetic adaptation, a strategy for its persistence in cystic fibrosis. Arch. Med. Res. 52, 357–361. doi: 10.1016/j.arcmed.2020.12.004
Sarges, E. D. S. N., Rodrigues, Y. C., Furlaneto, I. P., de Melo, M. V. H., Brabo, G. L. D. C., Lopes, K. C. M., et al. (2020). Type III secretion system virulotypes and their association with clinical features of cystic fibrosis patients. Infect. Drug Resist. 13, 3771–3781. doi: 10.2147/IDR.S273759
Silveira, M., Albano, R., Asensi, M., and Assef, A. P. C. (2014). The draft genome sequence of multidrug-resistant Pseudomonas aeruginosa strain CCBH4851, a nosocomial isolate belonging to clone SP (ST277) that is prevalent in Brazil. Mem. Inst. Oswaldo Cruz 109, 1086–1087. doi: 10.1590/0074-0276140336
Silveira, M. C., Rocha-de-Souza, C. M., Albano, R. M., de Oliveira Santos, I. C., and Carvalho-Assef, A. P. D. (2020). Exploring the success of Brazilian endemic clone Pseudomonas aeruginosa ST277 and its association with the CRISPR-Cas system type I-C. BMC Genomics 21:255. doi: 10.1186/s12864-020-6650-9
Tacconelli, E., Carrara, E., Savoldi, A., Harbarth, S., Mendelson, M., Monnet, D. L., et al. (2018). Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 18, 318–327. doi: 10.1016/S1473-3099(17)30753-3
Toleman, M. A., Simm, A. M., Murphy, T. A., Gales, A. C., Biedenbach, D. J., Jones, R. N., et al. (2002). Molecular characterization of SPM-1, a novel metallo-beta-lactamase isolated in Latin America: report from the SENTRY antimicrobial surveillance programme. J. Antimicrob. Chemother. 50, 673–679. doi: 10.1093/jac/dkf210
Tsai, C. J., Loh, J. M., and Proft, T. (2016). Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 7, 214–229. doi: 10.1080/21505594.2015.1135289
Tümmler, B. (2019). Emerging therapies against infections with Pseudomonas aeruginosa. F1000Res. 8:1371. doi: 10.12688/f1000research.19509.1
Turano, H., Gomes, F., Medeiros, M., Oliveira, S., Fontes, L. C., Sato, M. I. Z., et al. (2016). Presence of high-risk clones of OXA-23-producing Acinetobacter baumannii (ST79) and SPM-1-producing Pseudomonas aeruginosa (ST277) in environmental water samples in Brazil. Diagn. Microbiol. Infect. Dis. 86, 80–82. doi: 10.1016/j.diagmicrobio.2016.06.005
Van Nguyen, K., Nguyen, T. V., Nguyen, H. T. T., and Van Le, D. (2018). Mutations in the gyrA, parC, and mexR genes provide functional insights into the fluoroquinolone-resistant Pseudomonas aeruginosa isolated in Vietnam. Infect. Drug Resist. 11, 275–282. doi: 10.2147/IDR.S147581
Xu, C., Wang, D., Zhang, X., Liu, H., Zhu, G., Wang, T., et al. (2020). Mechanisms for rapid evolution of carbapenem resistance in a clinical isolate of Pseudomonas aeruginosa. Front. Microbiol. 11:1390. doi: 10.3389/fmicb.2020.01390
Yamane, K., Rossi, F., Barberino, M. G., Adams-Haduch, J. M., Doi, Y., and Paterson, D. L. (2008). 16S ribosomal RNA methylase RmtD produced by Klebsiella pneumoniae in Brazil. J. Antimicrob. Chemother. 61, 746–747. doi: 10.1093/jac/dkm526
Zhao, H., Xia, B., Fan, C., Zhao, P., and Shen, S. (2012). Human health risk from soil heavy metal contamination under different land uses near dabaoshan mine, southern China. Sci. Total Environ. 417–418, 45–54. doi: 10.1016/j.scitotenv.2011.12.047
Keywords: critical-priority pathogens, aquatic environments, carbapenemase, Galleria mellonella, resistome, virulome, genomic surveillance, One Health
Citation: Esposito F, Cardoso B, Fontana H, Fuga B, Cardenas-Arias A, Moura Q, Fuentes-Castillo D and Lincopan N (2021) Genomic Analysis of Carbapenem-Resistant Pseudomonas aeruginosa Isolated From Urban Rivers Confirms Spread of Clone Sequence Type 277 Carrying Broad Resistome and Virulome Beyond the Hospital. Front. Microbiol. 12:701921. doi: 10.3389/fmicb.2021.701921
Edited by:
Lorena Rodriguez-Rubio, University of Barcelona, SpainReviewed by:
Anelise Ballaben, University of São Paulo, BrazilPolly H. M. Leung, Hong Kong Polytechnic University, China
Márió Gajdács, University of Szeged, Hungary
Copyright © 2021 Esposito, Cardoso, Fontana, Fuga, Cardenas-Arias, Moura, Fuentes-Castillo and Lincopan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nilton Lincopan, bGluY29wYW5AdXNwLmJy; Fernanda Esposito, ZmVybmFuZGFlc3Bvc2l0b0B1c3AuYnI=
†These authors have contributed equally to this work