 Emanoelle La Santrer1
Emanoelle La Santrer1 Cláudia Barbosa Assunção1
Cláudia Barbosa Assunção1 Thiago Miguelito Navarro de Camargo1
Thiago Miguelito Navarro de Camargo1 Izabella Rodrigues1
Izabella Rodrigues1 Sabrina Sidney Campolina1Edgar Lacerda de Aguiar2Thiago de Souza Rodrigues2
Sabrina Sidney Campolina1Edgar Lacerda de Aguiar2Thiago de Souza Rodrigues2 Rachel Basques Caligiorne1*
Rachel Basques Caligiorne1*- 1Post-graduate Program in Medicine and Biomedicine, Faculdade de Saúde Santa Casa de Belo Horizonte, Belo Horizonte, Brazil
- 2Post-graduate Program in Modelagem Matemática Computacional, Centro Federal de Educação Tecnológica de Minas Gerais – CEFET-MG, Belo Horizonte, Brazil
Protein Kinases (PKs) are a large family of enzymes that act as “molecular switches,” playing fundamental role in cellular signaling through protein phosphorylation. This process consists in transfer a phosphate group (γ-PO₄2−) from ATP (adenosine triphosphate) to specific residues in target proteins; thereby, controlling vital cellular processes, such as (i) cell proliferation and differentiation, (ii) response to environmental stimuli (stress, nutrients, hormones), (iii) metabolism, (iv) cell cycle and apoptosis, and (v) signal transduction. Among fungi, adaptability is intrinsically connected to their ability to thrive under extreme environmental stress, being morphological plasticity an example of this adaptability. While many of these adaptive responses are regulated by diverse signaling pathways involving different kinase families, as mitogen-activated protein kinase (MAPK) for example, this review places a special focus on the General Control Nonderepressible 2 kinase (GCN2), a highly conserved sensor of amino acid scarcity in many fungi, as well as the species Cryptococcus neoformans, Candida albicans, and Aspergillus fumigatus. Amino acid deprivation triggers the accumulation of uncharged tRNAs, which directly activate GCN2, and this activation leads to the phosphorylation of the eukaryotic initiation factor 2 alpha (eIF2α) at the serine in the position 51, initiating the Integrated Stress Response (ISR). Phosphorylated eIF2α suppresses global translation initiation while selectively enhancing the translation of stress-responsive genes, notably GCN4, which encodes a transcription factor that promotes amino acid biosynthesis and stress adaptation. In Cryptococcus neoformans, GCN2 emerges as the sole kinase responsible for eIF2α phosphorylation, a unique role in modulating translational responses to environmental and host-induced stressors. Previous studies have shown that the absence of GCN2 disrupts eIF2α phosphorylation, impairing stress responses and reducing pathogenicity, therefore being an important target for development of new generation antifungals. To better understand the mechanistic role of GCN2 and related kinases in amino acid sensing and stress response, we present a review based on studying the central role of kinases in fungal stress adaptation, discussing how the high conservation of their catalytic kinase domains makes them valuable as phylogenetic markers and therapeutic targets.
1 Introduction
Fungi occupy a distinct ecological and evolutionary niche characterized by a unique evolutionary lineage amongst eukaryotes (Webster and Weber, 2007; de Hoog et al., 2023; Gow et al., 2017). They inhabit soil and water, efficiently decomposing organic matter to recycle nutrients, colonizing plants or animals as pathogens, symbionts, or commensals, and their spores can be also found in airborne environments (de Hoog et al., 2023; Gow et al., 2017; Mcginnis and Tyring, 1996). As a general rule, fungi possess a rigid glucan and chitin-based cell wall, which confers structural integrity and resistance to environmental pressures, which is directly related with their evolution and adaptation (Mcginnis and Tyring, 1996; Brunke et al., 2016; Sun et al., 2020; Carbonero and Strobel, 2021; Kurokawa et al., 1998; Farrer and Fisher, 2017).
Adaptability is intrinsically connected to fungal survival and ability to thrive under extreme environmental stress as well as dimorphism is directly related to fungal adaptability (de Hoog et al., 2023; Seyedmousavi et al., 2014). Thus, dimorphic fungi are capable of transitioning between multicellular and unicellular morphs in response to changes in environmental conditions (Klein and Tebbets, 2007). The mycelial (multicellular) form is found in environmental conditions, consisting of hyphae (tubular elongated cells), while the yeast (unicellular) form is represented by oval cells (de Hoog et al., 2023; Ramírez-Sotelo et al., 2025). Fungal dimorphism is associated with changes in metabolism and cell wall remodeling and can be caused by a series of stimuli; however, temperature is often the primary regulator of dimorphism in fungi (Gauthier, 2017). In dimorphic pathogenic fungi, host temperatures (35–37°C) trigger the infectious yeast form, while cooler environmental temperatures (23–25°C) promote the mycelial form (Sacks et al., 1986). This morphological plasticity is regulated by complex signaling pathways, including kinase-mediated responses, which coordinate the cellular and metabolic changes necessary for fungal adaptation and pathogenicity (Pathan et al., 2019).
Protein Kinases (PKs) are a large family of enzymes that act as “molecular switches,” playing a fundamental role in cellular signaling through protein phosphorylation (Taylor and Kornev, 2011). This process consists of the transfer of a phosphate group (γ-PO₄2−) from ATP (adenosine triphosphate) to specific residues in target proteins and; thereby, controlling vital cellular processes, such as: (a) cell proliferation and differentiation, (b) response to environmental stimuli (stress, nutrients, hormones), (c) energy metabolism, (d) cell cycle and apoptosis, and (e) signal transduction (Taylor and Kornev, 2011; Gao et al., 2008; Wrabl and Grishin, 2001).
Considering the fungi kingdom, it is well known that kinases are commonly involved in three main biological mechanisms that directly impact the infection process: morphogenesis, virulence, and stress adaptation (Ramírez-Sotelo et al., 2025; Pathan et al., 2019). This adaptability is connected to thermotolerance, as the ability to survive at human body temperatures (37°C) is a prerequisite for pathogenicity (Mattoon et al., 2021; Money, 2024). For instance, black yeasts are a group of ascomycotan melanized fungi that perfectly exemplify these adaptive traits (de Hoog et al., 2023; Seyedmousavi et al., 2014; Assunção et al., 2020).
At the molecular level, adaptability is mediated by kinase signaling pathways, which regulate stress responses and morphological transitions (Cañete-Gibas and Wiederhold, 2018; Ene et al., 2014; Rauch et al., 2011). Our review analyzed and synthetized the current knowledge about the eIF2α-related kinases family, more specifically GCN2 kinases, which are regulators of amino acid biosynthesis and stress responses conserved across all fungal species, with potential molecular marker for studies on stress adaptability, virulence, target for new antifungal drugs and molecular evolution.
2 Kinases
Kinases are a class of highly conserved enzymes found across all the domains of life. They are activated by various regulatory signals, such as amino acid deprivation or oxidative stress (Rauch et al., 2011). These enzymes regulate cellular processes by catalyzing the transfer of phosphate groups to specific molecules in a process known as phosphorylation (Cohen, 2002). This post-translational modification occurs with a phosphate group (PO₄3−) transfer from a high-energy molecule to a specific target (Rauch et al., 2011; Hunter, 1991). Kinases selectively phosphorylate specific amino acid residues, such as serine (Ser), threonine (Thr), or tyrosine (Tyr) and are classified, considering their substrate specificity, in two types: Ser-Thr kinases, known for phosphorylating serine and threonine residues, and Thr kinases, that phosphorylate tyrosine residues. Also, some kinases are considered dual-specificity kinases for being capable of phosphorylating two substrates (Hunter, 1991; Wheeler and Yarden, 2015; Martin et al., 2010).
The family of enzymes catalyzing the phosphorylation of proteins, the protein kinases, represents a structurally diverse group of proteins (Taylor and Kornev, 2011; Martin et al., 2010; Ekhator et al., 2025; Yang, 2003; Defosse et al., 2015; Cheng et al., 2011). They are modular structures that differ widely in size, subunit structure, subcellular localization, mechanism of activation, and substrate specificity. Two general classes exist, those transferring phosphate to Ser or Thr and those transferring phosphate to Tyr (Hunter, 1991; Martin et al., 2010). According to an important study on evolution of kinases, Taylor and Kornev (2011) shows that despite this diversity, all eukaryotic protein kinases are evolutionarily related throughout a conserved catalytic core, indicating that they all share at least some common features of secondary and tertiary structures.
Ser-Thr and Tyr share the N-terminal ATP binding pocket, a conserved structural region and catalytic domain located at the N-terminal domain of the kinase, responsible for binding adenosine triphosphate (ATP) to amino acids (Hunter, 1991; Wheeler and Yarden, 2015; Martin et al., 2010). Thus, phosphorylation can be considered a molecular switch: it activates or inhibits enzyme function, affecting downstream signaling pathways, but it is not a permanent biochemical process, as it can be undone (Plattner and Bibb, 2011). There are regulatory mechanisms to ensure that phosphorylation is a reversible and controlled process (Cheng et al., 2011).
The cyclic AMP-dependent protein kinase has also been widely studied since many decades (Pir et al., 2025; Aryal and Watts, 2025; Scott, 1991; Xiong et al., 2025; Shabb, 2001; Jungmann and Russell, 1977). The cyclic AMP (cAMP) pathway represents a central signaling cascade with crucial functions in all organisms. In fungi, the cyclic AMP (cAMP) pathway plays a central role in regulating development, growth, differentiation, conidiation, germination and virulence of pathogenic species (Maeng et al., 2010; Ocampo et al., 2009; Zhao et al., 2006). Adenosine 3′,5′-phosphate (cyclic AMP) has been studied as an intracellular regulator in a wide variety of organisms. Studies of cyclic AMP function in fungi have concentrated on functions repressed by glucose as well as on diverse developmental and other specific responses reported to be influenced by cyclic AMP in animals (Sabina and Brown, 2009). Evidence has been obtained that links cyclic AMP to control of a variety of functions in fungi, including utilization of endogenous and exogenous carbon sources, conidiation (in Neurospora crassa), dimorphism in several fungi, the sexual process, and phototropism (in Phycomyces) (Davis, 1986; Judewicz et al., 1981; Wang et al., 2020). According some authors, there are some striking similarities between animal and fungal cyclic AMP-dependent protein kinases, which suggest strong evolutionary conservation of the properties of these enzymes (Müller et al., 2003; McDonough and Rodriguez, 2012).
The study of protein kinases in fungi has progressed significantly over the past decades, building a strong foundation for current models of stress response and morphogenesis. Table 1 lists some important studies on cyclic AMP-dependent protein kinases from different fungal species, which opened up perspectives for further studies.
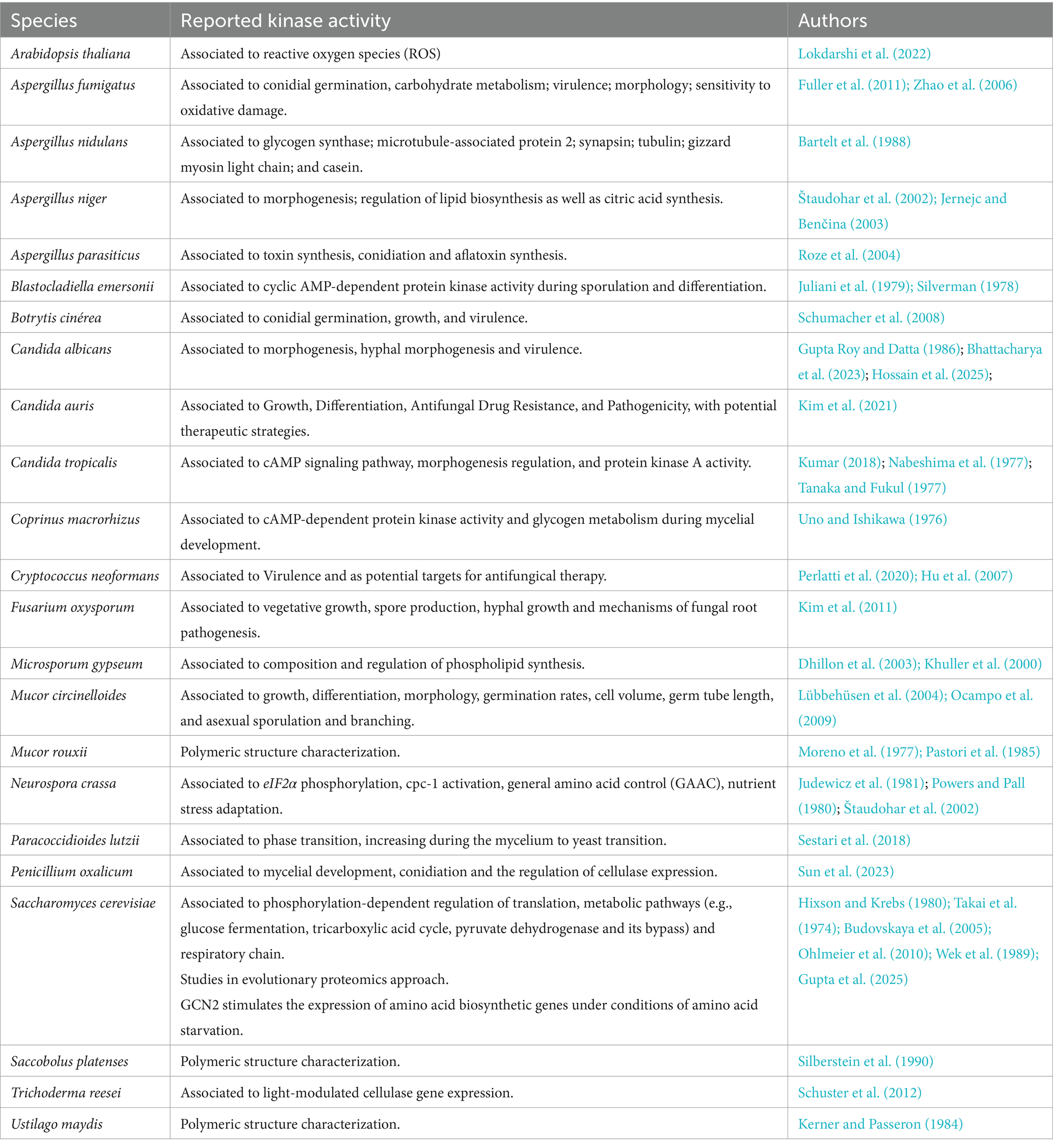
Table 1. Studies reported on cyclic AMP-dependent protein kinases in different species of fungi kingdom.
As detailed in Table 1, certain kinases play critical roles in translational control under stress conditions. For instance, the GCN2 mediated phosphorylation of eIF2α, which leads to the selective translation of transcription factors like GCN4, which is a key adaptive mechanism in fungi that will be explored in detail in a subsequent section. To better appreciate the role of kinases such as GCN2 in contemporary fungal biology, we revisit early discoveries and methodological developments that shaped this field. According to (Uno and Ishikawa (1976), the activity of cyclic AMP-dependent protein kinase in mycelial extracts of Coprinus cinerea concurrently with decrease of glycogen content in mycelial cells. Also, Silverman (1978), as in other eukaryotic cells, cyclic AMP in Blastocladiella sp. regulates cellular activity by activation of cyclic AMP-dependent protein kinase. In 1979, Juliani et al. demonstrated that cyclic AMP-dependent protein kinase activity and cyclic AMP binding components are induced during the sporulation in Blastocladiella emersonii, corroborating with the previous study by Silverman (1978), with the same fungal species (Juliani et al., 1979).
The Ca2+-dependent protein kinase seems to be associated with membranous components, whereas cyclic GMP-dependent protein kinase appears to be related to certain subcellular organelle, such as nucleus. Suggestive evidence is available implying that the cyclic AMP-, cyclic GMP- and Ca2+-activated three sets of protein kinase systems may play each specific physiological role presumably owing to their own subcellular compartments (Davis, 1986; McDonough and Rodriguez, 2012).
A Ca2+/calmodulin (CaM)-dependent multifunctional protein kinase has been isolated from Aspergillus nidulans and purified to homogeneity. Unlike any CaM-dependent multifunctional protein kinase described previously, the native enzyme from Aspergillus behaves as a monomer (Jernejc and Benčina, 2003). According to the authors, the Aspergillus kinase catalyzes the Ca2+/CaM-dependent phosphorylation of known substrates of type II Ca2+/CaM-dependent protein kinases, including glycogen synthase, microtubule-associated protein 2, synapsin, tubulin, gizzard myosin light chain, and casein (Jernejc and Benčina, 2003).
Protein phosphatases counteract kinases by removing phosphate groups, restoring proteins to their unphosphorylated state, maintaining cellular homeostasis, and preventing aberrant signaling implicated in diseases (Narayanan et al., 2023). Protein kinases, in particular, are essential regulators of cell growth, differentiation, and apoptosis, acting during normal cellular function and disease states (Hunter, 1991; Donnelly et al., 2013). Thus, diverse studies have linked kinases as targets for therapeutic intervention, and numerous kinase inhibitors has been recently developed for cancer, systemic infections, and neurodegenerative diseases (Narayanan et al., 2023; Berkes et al., 2021; Piecyk et al., 2024).
In an opposite enzymatic process, protein kinase inhibitors are molecules that hinder the activity of protein kinases by binding to the kinase’s active site or inducing conformational changes, effectively blocking phosphorylation events and modulating cellular processes. Furthermore, kinase inhibitors have been investigated for their potential as antifungal agents (Berkes et al., 2021; Hinnebusch, 2011). An important study performed by Berkes et al. (2021), which identified the cancer therapeutic dasatinib as a potent inhibitor of Histoplasma capsulatum in its pathogenic yeast form, showing the potential of repurposing kinase inhibitors for antifungal therapy.
3 Scientometric analysis on kinase studies
To measure the scientific production related to kinases in fungi, we analyzed the research output using various scientometric indicators. These included publication trends in relevant journals, keyword analysis, and the distribution of publications by country, author, and institutional relevance, as well as collaboration networks. The initial search was conducted in the PubMed database using the search terms “kinases,” “fungi,” and “proteins” (van Eck and Waltman, 2010; Aria, 2017).
The PubMed search yielded over 50,000 publications. Due to the processing capacity of the analysis software, the 10,000 most relevant articles with a specific focus on fungal kinases were selected for an initial overview. This subset was curated through manual screening to exclude reviews, duplicates, and studies with missing metadata (such as authorship, institutional affiliation, or publication year) or no abstract. The complete data selection and analysis workflow is illustrated in the scientometric analysis flowchart (Figure 1).
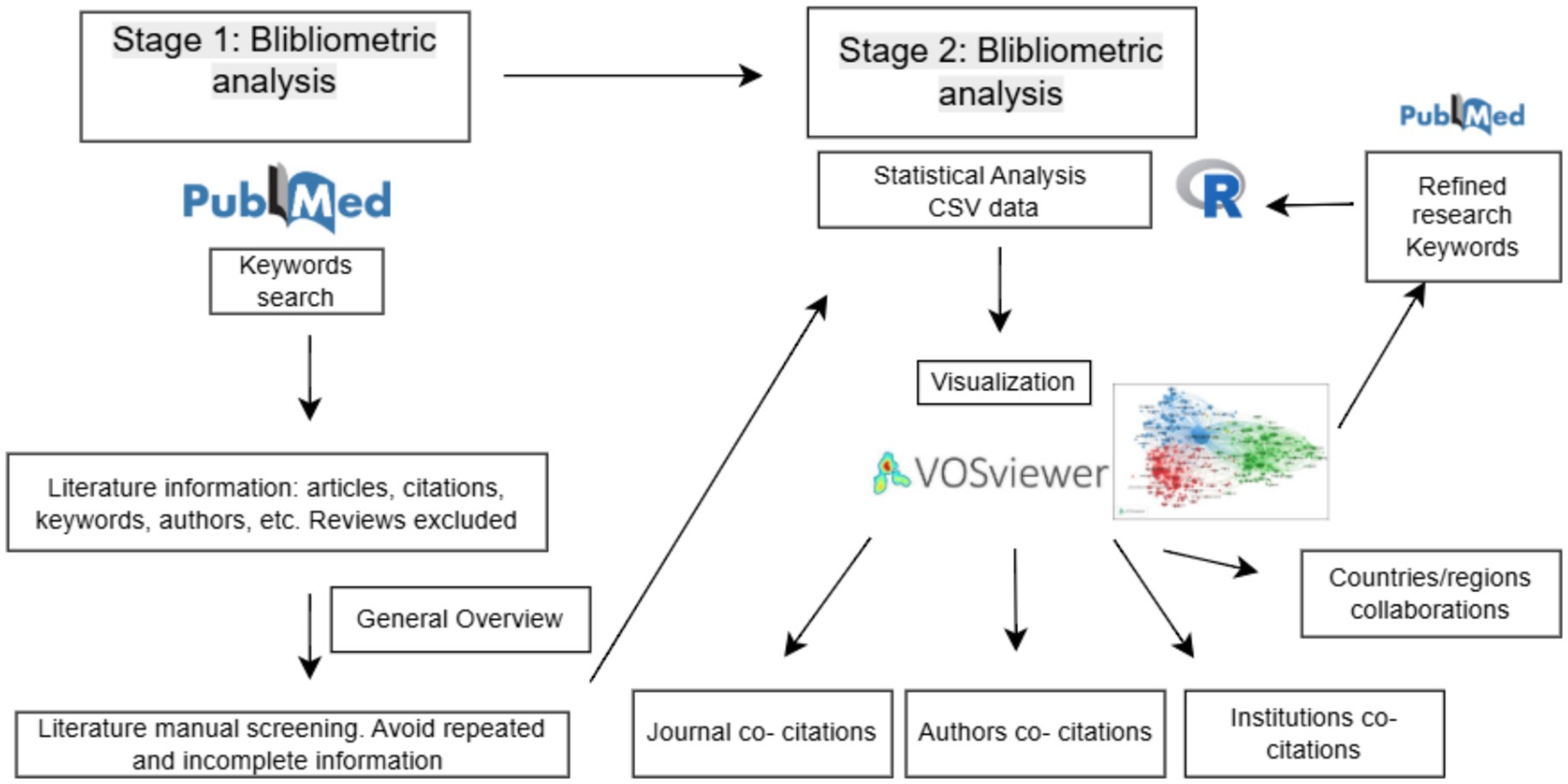
Figure 1. First, a broad overview of the kinase topic was conducted using the keywords “Fungi” and “Kinases.” Data from the 10,000 most relevant publications were manually screened to exclude reviews, duplicates, and publications with incomplete metadata. Resulting CSF generated data was analyzed using the R statistical program, and co-occurrence of major research topics was visualized with VOSviewer. Second, a refined analysis was performed using the keywords “dimorphic fungi” and “kinases.” The resulting dataset was manually screened according to the same criteria, and a second analysis was performed in R. VOSviewer was then used to visualize co-citation networks of countries, regions, journals, authors, and institutions.
Using this curated data, a CSF file was created and analyzed using R and visualized with VOSviewer (van Eck and Waltman, 2010; Aria, 2017). Keywords with at least 100 occurrences were included in this initial analysis (Lr, n.d.). Keyword analysis is critical as it indicates the focal areas of a research field and helps identify knowledge gaps. In this analysis, Saccharomyces cerevisiae was the most studied fungal species in the context of kinase proteins, followed by Schizosaccharomyces pombe (also called “fission yeast”), a common model organism in molecular and cell biology. Candida albicans was the third most studied species (Neiman et al., 1993). Regarding the biological activities of kinases, phosphorylation was the most extensively studied process. The analysis highlighted the importance of kinases in numerous cellular processes, including enzyme activation, gene expression regulation, protein binding, intracellular signaling, and transcription factor activity. Kinases are also involved in cell cycle progression, such as mitosis and DNA damage responses. Furthermore, studies on Candida albicans frequently associate kinase activity with virulence, demonstrating the functional versatility of these proteins (Figure 2).
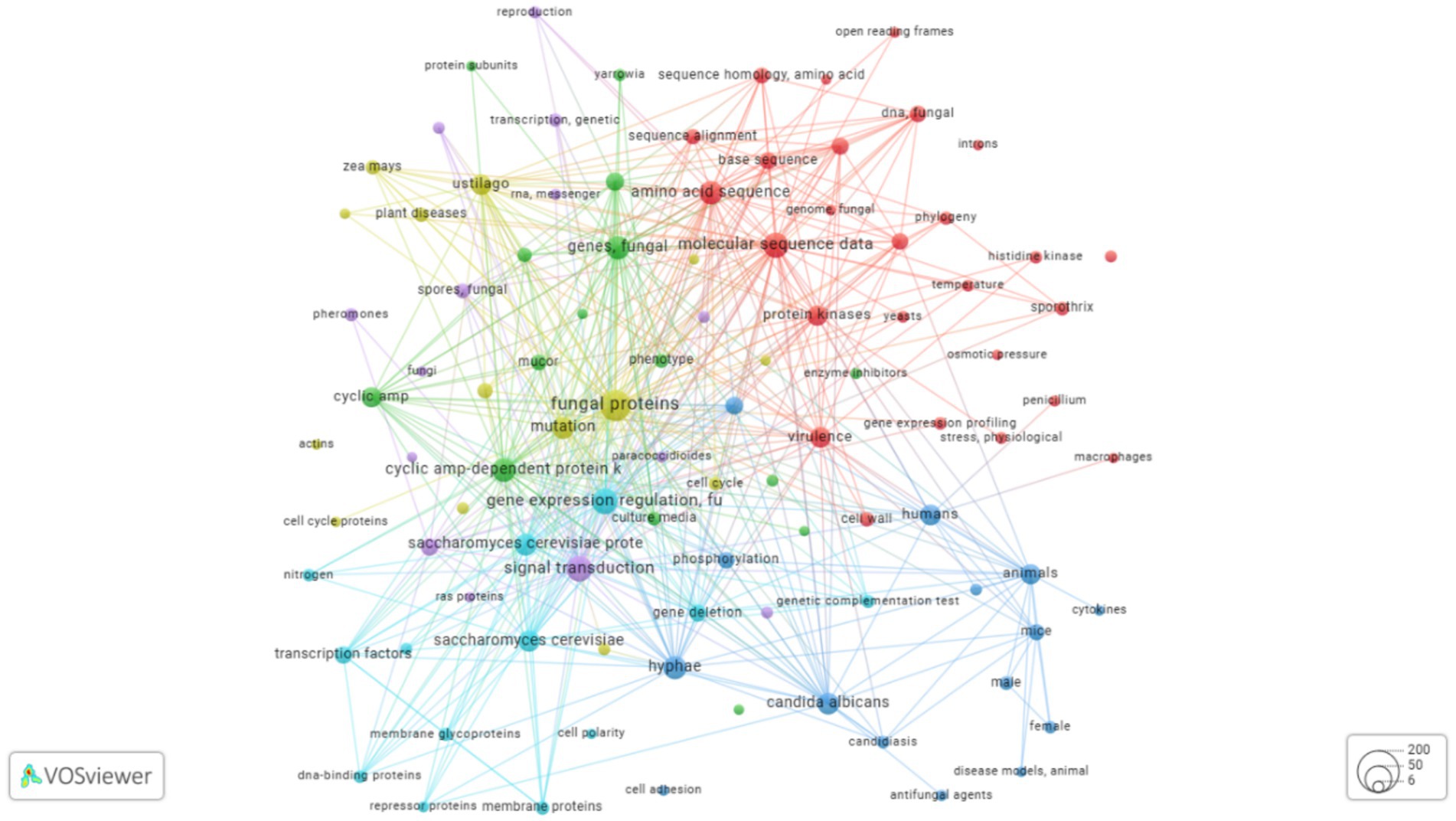
Figure 2. Keyword co-occurrence map generated using VOSviewer. The map is based on keywords extracted from approximately 270 articles on dimorphic fungal kinases retrieved from the PubMed database, published between 1982 and 2025. The analysis includes keywords that appeared at least five times. Colors denote thematic clusters of frequently co-occurring terms: the red cluster is associated with molecular data and phylogeny; the blue cluster relates to animal models and pathogenicity; and the green cluster represents cellular processes and fungal species. The size of each circle corresponds to the keyword’s frequency, with larger circles indicating terms that appeared more often.
To align with the focus of this review, a second, more targeted analysis was performed using the keywords “dimorphic fungal kinases.” Following the exclusion of reviews, duplicates, and articles with incomplete data, approximately 270 articles were selected from the PubMed database. Keywords appearing at least five times in this dataset are presented in Figure 2. In this focused analysis, Candida albicans remains the most prominent species, with Sporothrix and Paracoccidioides species also frequently appearing. Additionally, Ustilago maydis, a basidiomycete that causes corn smut, was identified (Olicón-Hernández et al., 2015). The term “Mucor” also appeared, which is associated with approximately 40 species of molds and dimorphic fungi, including both pathogenic and avirulent species (Lr, n.d.). This data demonstrates the importance of kinases in processes of morphological plasticity, such as dimorphism, and shows how molecular sequence data are crucial for uncovering kinase profiles and functions.
In addition to this keyword analysis, further scientometric analyses were conducted. The evolution of publications over the years, the most prominent authors and institutions, the most prolific journals, and existing collaboration networks were detailed. Graphical representations that complement these analyses are gathered in the Supplementary material of this study. The number of annual publications was evaluated. Publications on the subject began in the 1980s, with limited output until 1995, after which the number of publications doubled. The highest number of publications on kinases in dimorphic fungi was observed between 2000 and 2014. In 2015, there was a significant drop, followed by an increase and subsequent stabilization in 2018 and 2019 (Supplementary material).
A new drop in the number of publications was observed in 2020, likely because research institutions were focused on the COVID pandemic. The most productive institutions were also ranked, with the University of California leading (18 articles), followed by the Federal University of São Paulo (13) and the Institute of Microbiology (13). The University of Louisville (12) and the University of Buenos Aires (12) ranked fourth. The University of Murcia (11) and South China Agricultural University (11) followed, with Duke University Medical Center (10) and King’s College of London (9) completing the list (Supplementary material). Notably, two of the ranked institutions are in Latin America, likely reflecting the medical interest in dimorphic fungi in a region with a high incidence of fungal infections.
Evaluation of inter-institutional collaboration revealed no broad networks; however, collaborations between departments within Californian institutions were observed, which may contribute to the University of California’s leading rank (Supplementary material). The most prolific authors on this subject are Moreno, S. (10 articles), followed by Heitman, J. (9), and Ruiz-Herrera and Passeron, S. (seven articles each) (Supplementary material). Analysis of publication venues revealed that Eukaryotic Cell has published the highest number of papers (11), followed by Molecular Microbiology (10). Fungal Genetics and Biology and FEMS Yeast Research ranked third with six publications each (Supplementary material).
4 General Control Nonderepressible 2 (GCN2) kinase and its role in fungi
Amino acids are organic molecules fundamental to build proteins characterized by a central carbon atom bonded to an amino group (−NH3+), a carboxyl group (−COO–), a hydrogen atom and a variable side chain (R group), which determines the properties of each amino acid and consequently to protein structure and function (Donnelly et al., 2013; Douglas et al., 2024). In fungi, amino acids are fundamental for various biochemical processes, such as protein synthesis, enzyme production, and cell wall biosynthesis (Muhammad et al., 2024; Chadwick, 2023). They also act as precursors for important secondary metabolites, including pigments, toxins, and antimicrobial compounds, which are essential for fungal survival and pathogenicity (Ene et al., 2014). Amino acids represent an abundant source of nitrogen and carbon within the host and their process of acquisition by fungi interconnects sensory and uptake systems and downstream pathways (Maloy, 2013).
Amino acid starvation poses a significant challenge for fungi. Starvation is caused by an insufficient availability of amino acids due to environmental stressors (Ene et al., 2014; Chadwick, 2023; Garbe and Vylkova, 2019). Some environments are naturally poorer in nutrients, such as soil, air, or host tissue. Inside hosts, during pathogenic interaction, a sequester or degradation of essential amino acids can occur as an immunological mechanism, known as nutritional immunity, to starve the invading pathogen (Chadwick, 2023; Maloy, 2013; Garbe and Vylkova, 2019; Emery, 2012). Environmental stressors, including oxidative stress, temperature shifts, and osmotic pressure, can impair amino acid synthesis pathways or even completely deplete existing amino acid reserves (Maloy, 2013; Emery, 2012). Genetic mutations are also capable of impairing the endogenous amino acid production, leading to a starvation process even in environments rich in nutrients (Kim et al., 2011). Additionally, starvation induces autophagy, facilitating the recycling of intracellular components, including amino acids, to maintain essential metabolic processes (Maurin et al., 2022).
When faced with amino acid starvation, fungi activate stress response mechanisms to ensure survival and conserve energy, triggering a cascade of stress responses aimed at ensuring survival and adaptation (Knowles et al., 2021). The accumulation of uncharged tRNAs is a primary signal for amino acid scarcity, triggering the activation of the GCN2 kinase (Emery, 2012; McCarthy and Walsh, 2018). Recent studies reveal that fungi utilize multiple, interconnected pathways to detect and respond to amino acid scarcity (Emery, 2012; McCarthy and Walsh, 2018).
General Control Nonderepressible 2 kinase (GCN2) kinase activation by amino acid starvation also occurs in plants, mammals, and fungi through various regulation mechanisms, and it is quite conserved in those eukaryotes (Masson, 2019; Castilho et al., 2014; Maurin et al., 2005). Currently, GCN2 is identified as the sole kinase responsible for the phosphorylation of eIF2α (Leipheimer et al., 2019). In fungi, GCN2 phosphorylates eIF2α eIF2α, and, and it is composed of 1,649 amino acids, spanning from the N-terminal to the C-terminal region (Castilho et al., 2014). This phosphorylation event at serine 51 of eIF2α is a critical component of the Integrated Stress Response (ISR), allowing the fungus to modulate protein synthesis during environmental stresses (Masson, 2019; Castilho et al., 2014; Hinnebusch, 2005; Li et al., 2024).
The activation of GCN2 is triggered by the accumulation of uncharged tRNAs, leading to the phosphorylation of eIF2α, which causes a reduction in general translation initiation while selectively promoting the translation of stress-adaptive genes, more specifically GCN4 (Masson, 2019; Hinnebusch, 2005; Li et al., 2024). According to recent studies, in the absence of GCN2, C. neoformans exhibits impaired phosphorylation of eIF2α, resulting in reduced stress tolerance and diminished virulence (Leipheimer et al., 2019; Stovall et al., 2021).
In response to amino acid deprivation, yeast cells activate a signaling pathway involving GCN2-mediated phosphorylation of serine 51, which, in turn, triggers a two-pronged adaptation: global protein synthesis is down-regulated to preserve resources, while the translation of the GCN4 factor, which regulates the expression of a set of genes is preferentially promoted, stimulating the expression of genes responsible for amino acid biosynthesis (Hinnebusch, 2005; Li et al., 2024). While the fundamental architecture of GCN2 is preserved between animals and fungi, encompassing domains such as the RWD domain, pseudokinase domain (PKD), kinase domain (KD), histidyl-tRNA synthetase-like (HisRS-like) domain, and C-terminal domain (CTD), there are structural differences particularly in the CTD and dimerization interfaces since the GCN2 kinase in vertebrates has low sequence similarity when compared to the CTD of yeast GCN2, despite performing similar roles (Piecyk et al., 2024; Hamanaka et al., 2005).
As illustrated in Figure 3A, the GCN2 kinase is a multi-domain protein with five regions, a (1) RWD domain, (2) a pseudokinase domain (PKD), (3) Protein a Kinase (PK) domain, (4) a catalytic domain homologous to the histidyl-tRNA synthetase (HisRS), and a (5) C-terminal domain (Castilho et al., 2014). To better understand the structure of GCN2 protein, a predicted three-dimensional structure of the GCN2 protein from C. carrioni is presented in Figure 3 showing a detailed view of its functional architecture.
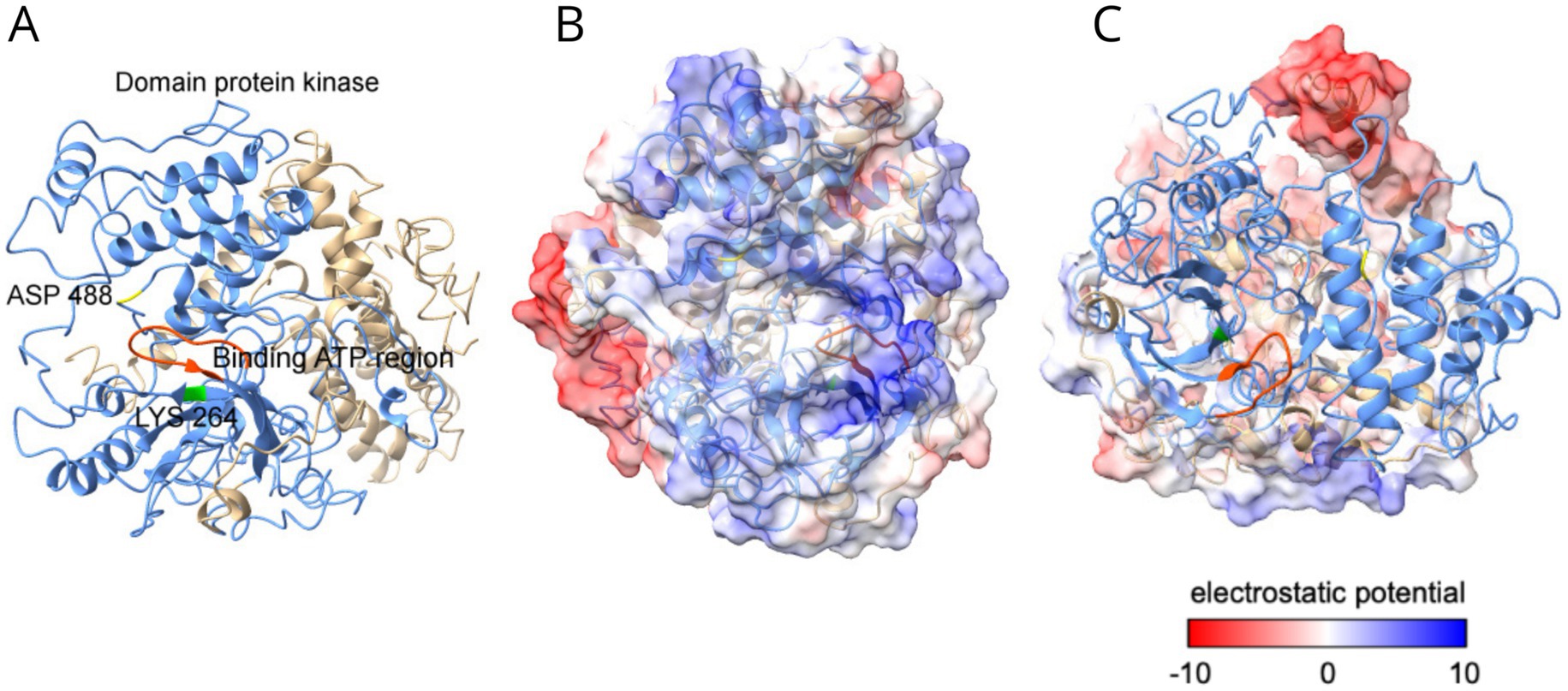
Figure 3. Structural and electrostatic analysis of the GCN2 protein from C. carrioni. (A) The overall domain architecture, illustrating the RWD, pseudokinase (PKD), catalytic kinase (PK), HisRS-like, and C-terminal domains that support its regulatory function. (B) A detailed view of the catalytic active site. Key residues lysine 264 (Lys264, shown in green) and aspartic acid 488 (Asp488, in yellow) are highlighted within the ATP-binding region (orange-red). These residues function as the proposed proton acceptor and donor. (C) Electrostatic surface potential of the kinase domain. Blue coloring represents positive charge (lower electron density), while red indicates negative charge.
The RWD domain is a structural motif located in the N-terminal region of GCN2 kinase. It is named after its occurrence in RING finger-containing proteins, WD-repeat-containing proteins, and DEAD-like helicases, but in GCN2, it serves as a protein–protein interaction module, mediating interactions with GCN1 (Marton et al., 1993; Rakesh et al., 2017). It is where the assembly of the GCN2 kinase complex occurs (Castilho et al., 2014). It is highly conserved with approximately 110 amino acids, it adopts an α + β sandwich fold, and consists of a four-stranded antiparallel β-sheet and three α-helices. Masson (2019) describes the three-dimensional structure of each domain of the GCN2 enzyme. Represented by Protein Data Bank (PDB) codes, more specifically, the RWD domain structure exhibits an α + β sandwich fold, as observed in ID: 1UKX (Masson, 2019). Residues Glu123 and Glu136 in α-helix 3 are implicated in binding to GCN1, which facilitates the interaction between GCN2 and GCN1 (Rakesh et al., 2017; Nameki et al., 2004; He et al., 2014).
Additionally, a highly positively and negatively charged region is located near the RWD domain, including a pseudokinase domain (PKD), which resembles the Protein Kinase domain but lacks the residues necessary for enzymatic activity (Castilho et al., 2014). In spite of that fact, it acts as a structural scaffold, influencing the conformation and stability of the adjacent active Kinase domain (Stovall et al., 2021; Bou-Nader et al., 2024). Therefore, in order to modulate GCN2 activity, the PKD maintains the kinase in an inactive state under non-stress conditions and undergoes conformational changes upon stress signals, thus activating the functional Kinase domain (Masson, 2019; He et al., 2014).
A Protein Kinase domain that displays a typical kinase domain structure, represented by ID: 1ZYD, while the CTD is illustrated using the dimeric CTD structure ID: 4OTN. The crystallographic structure described by Padyana et al. (2005) suggests that the kinase domain maintains a “closed” conformation due to the rigidity of the hinge at the N- and C-terminal ends, bringing these regions closer together and partially obstructing access to the catalytic site. Furthermore, the kinase can undergo autophosphorylation at two threonine residues within the activation loop, leading to stabilization of the active state when the N and C lobes open. The formation of an intermolecular salt bridge between kinase domains facilitates this activation (Masson, 2019; Nameki et al., 2004; Padyana et al., 2005; Sattlegger et al., 2011). This formation needs to occur for the catalytic activity of GCN2, specifically the phosphorylation of eIF2α. It presents structural barriers that impede active conformation, and the PK domain forms a dimer during crystallization, with its protomers arranged in an antiparallel configuration (He et al., 2014). For the kinase to become active, it must undergo isomerization, shifting to a parallel orientation (He et al., 2014; Evelyn and Hinnebusch, 2000). Moreover, it can also adopt an active dimer configuration shared by the PKR and PERK2 kinase domains (Castilho et al., 2014).
Alongside the PK domain, the catalytic domain, repeat 2, of the Serine/Threonine kinase (STK)—central catalytic domain is similar to the homodimeric class II histidyl-tRNA synthetase (HisRS). In yeast cells, the GCN2 kinase is activated under amino acid starvation conditions through the binding of uncharged tRNA to its histidyl-tRNA synthetase-like (HisRS) domain (Bou-Nader et al., 2024; Wek et al., 1995). Another essential factor for GCN2 activation is the ribosome-binding and dimerization domain located in the C-terminal region (Ramirez et al., 1991). However, interactions between the HisRS and PK domains can hinder GCN2 activation, as this interaction reduces GCN2’s affinity for tRNAs (Bou-Nader et al., 2024). A kinase-like domain near the N-terminal region, along with the PK domain and the RWD region, functions as a regulatory domain. The RWD region mediates the binding of the GCN1—GCN20 complex, which interacts with translating ribosomes, facilitating the binding of uncharged tRNA to the HisRS domain and consequently activating the kinase (Castilho et al., 2014; Stovall et al., 2021; Padyana et al., 2005).
Lastly, the C-terminal domain which maintains GCN2 in an inactive conformation under non-stress conditions, when uncharged tRNA binding to the HisRS-like domain, conformational changes in the CTD relieve this inhibition and allow kinase activation, forming a bipartite tRNA-binding structure (Nameki et al., 2004; Sattlegger et al., 2011; Dong et al., 2000). Some studies implicate that the CTD negatively regulates kinase activity and that disrupting this interaction releases inhibition potentially enhancing GCN2 activation (Nameki et al., 2004; He et al., 2014; Dong et al., 2000). The CTD also facilitates ribosome association and stress response, and GCN2 must be physically tethered to the ribosome; therefore, involving CTD and associated regions for tRNA sensing and kinase activation (Liu et al., 2019).
5 Mechanisms of GCN2 activation and regulation by GCN1
GCN2 normally exists in an autoinhibited state, where its kinase activity is kept low to prevent unnecessary signaling. This autoinhibition is relieved by an allosteric activation mechanism that senses amino acid scarcity (Donnelly et al., 2013; Komar and Merrick, 2020). When amino acids are scarce, uncharged tRNAs accumulate in the cell, binding specifically to the HisRS-like domain, which is structurally similar to histidyl-tRNA synthetase but specialized for sensing these tRNAs rather than catalyzing tRNA charging (Bou-Nader et al., 2024). This binding event induces a conformational change that relieves GCN2’s autoinhibited state, allosterically activating its kinase domain (Komar and Merrick, 2020; Bou-Nader et al., 2024).
Once active, GCN2 phosphorylates the alpha subunit of the eukaryotic initiation factor 2 (eIF2α) at the serine-51 residue. This single phosphorylation event is the critical switch for cell modulation. The functional consequence is that phosphorylated eIF2α competitively inhibits its guanine nucleotide exchange factor, eIF2B (Sattlegger and Hinnebusch, 2005; Terenin et al., 2008; Zügel et al., 1999). Since eIF2B is required to recycle eIF2 from its inactive GDP-bound form to its active GTP-bound form, its inhibition leads to a sharp decrease in the available pool of active eIF2 (Sattlegger and Hinnebusch, 2005). This results in a potent suppression of global protein synthesis, conserving cellular resources during stress, while paradoxically enabling the selective translation of stress-response mRNAs, most notably that of the transcription factor GCN4 (Terenin et al., 2008; Zügel et al., 1999).
Normally, GCN4 translation is suppressed by upstream open reading frames (uORFs) in its mRNA. However, eIF2α phosphorylation alters ribosome scanning, bypassing these uORFs and successfully initiate translation at the main GCN4 start codon. GCN4, a transcription factor, then upregulates genes involved in amino acid biosynthesis and stress adaptation, facilitating cellular recovery (Hinnebusch, 2005; Li et al., 2024; Marton et al., 1993; Sattlegger et al., 2011).
Studies indicate that GCN2 must bind to GCN1, which selectively binds to GCN2 to regulate its biological activity. The kinase GCN1 is a cytoplasmic protein that plays a crucial role in helping cells respond to different stressors, and it also contains binding sites for GCN2 (Marton et al., 1993; Sattlegger et al., 2011). GCN1 comprises three regions, A, D, and E, which ensure that GCN1 can transmit signals to GCN2, enabling the cell to adapt to nutrient stress. Among GCN1 regions, region D is essential for GCN2 binding, mutations in this region are known to disrupt this interaction, preventing activation according to Masson (2019) and Hinnebusch (2005). While Region E weakens the connection, GCN2 overexpression can partially compensate as shown by Masson (2019) and Hinnebusch (2005). Region A, on the other hand, is the least critical and mainly helps stabilize the interaction rather than being directly responsible for binding (Hinnebusch, 2005; Sattlegger et al., 2011). GCN1’s middle region is homologous to the N-terminal HEAT repeat domain in EFF3 (Fungal translation elongation factor 3), which is essential for ribosome recycling and translational elongation, facilitating tRNA release from the E-site, where deacylated tRNA exits after peptide bond formation, and enhancing ribosome efficiency (Rakesh et al., 2017; Ranjan et al., 2021).
The interaction between GCN1 and GCN2, as well as their association with the ribosome, is required for GCN2 activation in both yeast and mammals (Gottfried et al., 2022). Hinnebusch (2005) identified an amino acid in GCN1’s GCN2-binding domain through protein interaction and functional analysis assay that is essential for this interaction, Arg2259, as its mutation disrupted the association but could be rescued by GCN2 overexpression (Hinnebusch, 2005). These findings confirm that the N-terminal domain of GCN2 must bind to the C-terminal region of GCN1 for optimal GCN2 function and amino acid regulation (Marton et al., 1993; Sattlegger et al., 2011; Gottfried et al., 2022).
The data support a model where GCN1, GCN20, and GCN2 form a ribosome-associated complex, with GCN1 aiding in the transfer of uncharged tRNA from the ribosomal A-site to GCN2’s tRNA-binding domain for kinase activation. Based on inferences drawn from several studies (Wek et al., 1989; Dong et al., 2000; Wek, 2018; Masson, 2019), Figure 4 summarizes the molecular mechanism of GCN2 activation and its role in fungal stress response.
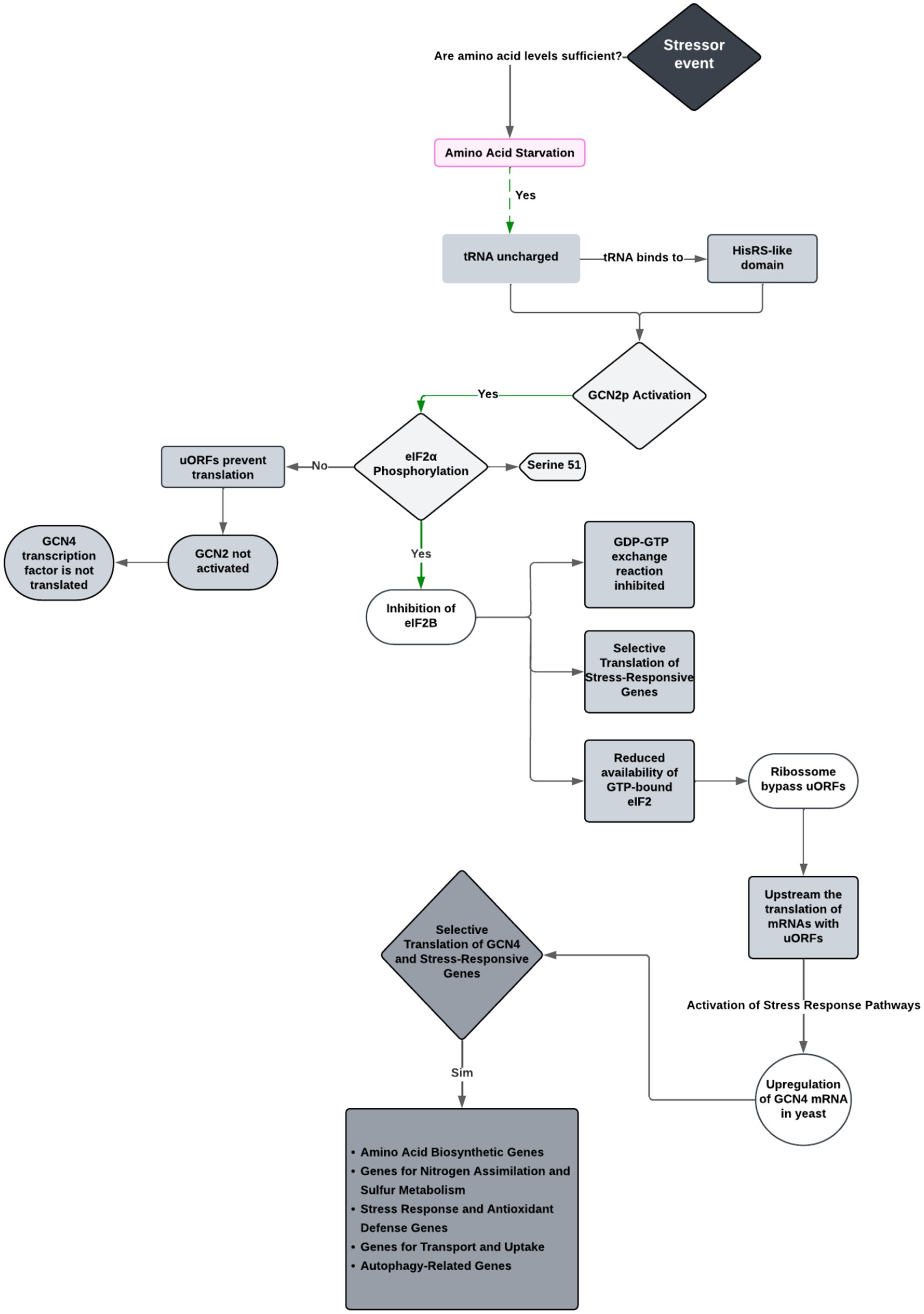
Figure 4. Integrated model of the GCN2-mediated stress response pathway in fungi. This diagram illustrates the cascade of molecular events triggered by amino acid starvation, including the activation of GCN2 by uncharged tRNA, phosphorylation of eIF2α at serine 51, inhibition of eIF2B, and the resulting selective translation of stress-responsive mRNAs such as GCN4. This pathway was constructed based on mechanistic insights from multiple authors (Hinnebusch, 2011; Masson, 2019; Castilho et al., 2014; Hinnebusch, 2005; Bou-Nader et al., 2024; Evelyn and Hinnebusch, 2000; Sattlegger and Hinnebusch, 2005; Teske et al., 2011).
6 Eukaryotic initiation factor 2 (eIF2)
Eukaryotic initiation factor 2 alpha, eIF2α, is an essential substrate within the Integrated Stress Response pathway (ISR), a conserved cellular signaling network that regulates protein synthesis in stressful conditions such as oxidative stress and amino acid deprivation (Donnelly et al., 2013; Pakos-Zebrucka et al., 2016). All this complex is considered a regulator of protein synthesis, modulating cellular responses to various stress conditions involved in phosphorylating the eIF2α subunit (Asada et al., 2025). The phosphorylation processes are necessary for the downregulation of global translation initiation, conserving resources and promoting stress adaptation allowing the selective translation of stress related protein, such as GCN4 (Wek, 2018). Eukaryotic initiation factors (eIFs) are necessary for translation initiation, and the eIF2 complex, including the regulatory subunit eIF2α is responsible for delivering initiator tRNA to the ribosome during the early stages of protein synthesis (Donnelly et al., 2013; Komar and Merrick, 2020; Burwick and Aktas, 2017).
There is an important distinction that deserves to be made: eIF2α is not a kinase since it does not add phosphate groups to other proteins. Instead, it is a substrate, phosphorylated by eIF2α related kinases at serine 51 (such as GCN2, PKR, PERK, HRI) (Donnelly et al., 2013; Komar and Merrick, 2020; Burwick and Aktas, 2017) The eIF2 Complex is a heterotrimeric GTP-binding protein composed of three subunits, eIF2α, eIF2β, and eIF2γ. The eIF2 participates in nearly all cytoplasmic mRNA translation events under normal cellular conditions (Kimball, 1999). eIF2α was initially identified as a factor capable of facilitating the binding of methionyl-initiator tRNA (Met-tRNAi) to the 40S ribosomal subunit in eukaryotic cells, in a process necessary for initiating protein synthesis through the formation of the 43S pre-initiation complex, which scans the mRNA to identify the start codon (Clemens, 1976).
Structurally, eIF2 is a heterotrimeric complex consisting of three subunits: α (36 kDa), β (38 kDa), and γ (52 kDa) (Donnelly et al., 2013; Komar and Merrick, 2020; Kimball, 1999). Each subunit shows a unique functional role in translation regulation. The α-subunit contains a key regulatory serine residue at position 51, which is a major target for phosphorylation during stress responses, leading to translation inhibition (Kimball, 1999). This subunit has an important role as translational control in fungi and will be explored in depth in this review article. The β-subunit provides binding sites for the heterotetrametric guanine nucleotide exchange factor eIF2B, which converts protein synthesis eIF2 from a GDP-bound form to the active eIF2-GTP complex, facilitating the recycling of eIF2 between its active and inactive states, and eIF5, a GTPase-activating protein that enhances translation initiation (Castilho et al., 2014; Wek, 2018; Kimball, 1999). The γ-subunit, the largest of the three, binds guanine nucleotides (GTP and GDP) and Met-tRNAi, essential for forming the ternary complex required for ribosomal initiation (Donnelly et al., 2013; Komar and Merrick, 2020; Wek, 2018).
The eIF2-GTP-Met-tRNAi ternary is a prerequisite for the assembly of the 43S pre-initiation complex, including additional factors like eIF1, eIFA, and eIF3. This complex is responsible for scanning the mRNA and locating the right AUG start codon (Komar and Merrick, 2020). The translation initiation is highly dependent on eIF2, mediating the delivery of Met-tRNAi to the 40S ribosomal subunit. Ribosomes present two subunits for protein synthesis, in eukaryotic cells the 40s ribosomal subunit, also known as the small subunit is responsible for reading mRNA, ensuring that tRNAs are paired to their respective mRNA sequences to decode genetic information (Jennings et al., 2017). The Met-tRNAi delivery occurs through a ternary complex composed of eIF2, GTP, and Met-tRNAi. eIF2 depends on its nucleotide-bound state to bind Met-tRNAi, and high affinity can be observed when eIF2 is associated with GTP (Jennings et al., 2017; Kapp and Lorsch, 2004).
The eIF5 facilitates the hydrolysis of eIF2-bound GTP by acting as a GTPase-activating protein and interacting with the 40S initiation complex, converting it to GDP. This conformational change results in the dissociation of eIF2 from the ribosome in its GDP-bound form, along with eIF5 (Unbehaun et al., 2004; Wortham and Proud, 2015). For eIF2 to start again the translation initiation, its GDP must be replaced with GTP through a process mediated by eIF2B, a guanine nucleotide exchange factor composed of five subunits (α, β, γ, δ, and ε) (Castilho et al., 2014; Komar and Merrick, 2020; Kimball, 1999; Elsby et al., 2011). This phosphorylation event inhibits eIF2B, effectively halting the GDP-GTP exchange required for forming new ternary complexes and preventing the recycling of eIF2-GDP back to its active GTP-bound form (Terenin et al., 2008; Kapp and Lorsch, 2004; Unbehaun et al., 2004). Nevertheless, certain mRNAs can bypass eIF2 dependence and continue to be translated through alternative initiation mechanisms such as internal ribosome entry sites (IRES) or re-initiation strategies (Evelyn and Hinnebusch, 2000; Ramirez et al., 1991). Selective translation of stress-response mRNA containing uORFs (upstream open reading frames), such as GCN4, continues (Hinnebusch, 2005; Li et al., 2024; Ramirez et al., 1991).
Four primary eIF2α kinases have been identified, each responding to distinct stress signals: Heme-Regulated Inhibitor Kinase (HRI), predominantly expressed in erythroid cells senses intracellular heme concentrations, exerting its functions through the dissociation of molecular chaperones such as Hsp90 and Hsc70. Heat shock proteins (Hsp) are a group of conserved chaperones that influence proteostasis and are responsible for activating the transcription of HSP genes under stress, protein folding, or proteasomal degradation (Zügel et al., 1999; Singh et al., 2024). It has been recently recognized that HRI is ubiquitously expressed and mediates eIF2α phosphorylation in various cell types exposed to multiple stressors (Lr, n.d.). Under heme-deficient conditions, heat shock, or heavy metal toxicity, HRI is activated, phosphorylating eIF2α to balance globin synthesis with available heme and; thereby, preventing the accumulation of unpaired globin chains (Donnelly et al., 2013; Burwick and Aktas, 2017; Girardin et al., 2021). This kinase responds to oxidative stress, as seen in its requirement for eIF2α phosphorylation and stress granule formation following sodium arsenite treatment, a potent inducer of oxidative damage (Burwick and Aktas, 2017; Girardin et al., 2021).
Double-Stranded RNA-activated Protein Kinase (PKR) is a vertebrate-specific kinase that serves as a key antiviral defense mechanism by recognizing viral double-stranded RNA (dsRNA), a byproduct of viral replication (Gal-Ben-Ari et al., 2019). Upon binding dsRNA, PKR undergoes dimerization and autophosphorylation via its C-terminal kinase domain, which is structurally preceded by N-terminal dsRNA-binding motifs (Gal-Ben-Ari et al., 2019). Once activated, PKR phosphorylates eIF2α, leading to the inhibition of guanine nucleotide exchange by eIF2B and subsequent suppression of cap-dependent translation initiation halting both viral and host protein synthesis (Gal-Ben-Ari et al., 2019; Yu et al., 2013). Beyond translational control, PKR can also induce apoptosis through eIF2α-independent mechanisms involving caspase pathway activation (Yu et al., 2013; Rothenburg et al., 2008). Dysregulation of PKR is associated with cancer, neurodegenerative diseases, and metabolic disorders (Gal-Ben-Ari et al., 2019; De La Cruz-Herrera et al., 2014; Dar et al., 2005). Given its central role in antiviral immunity, many viruses have evolved strategies to evade PKR activity, including direct inhibition, mislocalization, degradation, or interference with its RNA-binding capacity (Gal-Ben-Ari et al., 2019; Rothenburg et al., 2008; Dar et al., 2005).
Protein kinase RNA-like endoplasmic reticulum kinase (PERK) is a transmembrane protein residing in the ER capable or regulating cellular adaptation to ER stress. PERK is activated in response to the accumulation of misfolded proteins within the ER lumen, initiating the unfolded protein response (UPR) (Saito et al., 2011). In its inactive form, PERK is bound by the chaperone BiP (GRP78), but upon ER stress, BiP dissociates, allowing PERK dimerization and subsequent autophosphorylation at multiple serine/threonine residues, most notably at Thr980, which stabilizes its activation loop and αG helix for substrate interaction. Interestingly, PERK activation can also occur independently of misfolded proteins, possibly through fluctuations in ER luminal ATP or calcium levels sensed by BiP or associated co-chaperones (Saito et al., 2011; Cui et al., 2011). Once activated, PERK phosphorylates eIF2α and; thereby, inhibiting global cap-dependent translation and reducing protein load on the ER (Saito et al., 2011). This phosphorylation not only limits general protein synthesis but also enables selective translation of specific stress-responsive transcripts, such as ATF4, due to its unique upstream open reading frames (uORFs).
ATF4, in turn, regulates genes involved in redox balance, amino acid metabolism, and apoptosis, often converging on CHOP as a pro-apoptotic effector. PERK also directly phosphorylates the NRF2, disrupting its interaction with KEAP1 and facilitating its nuclear translocation to upregulate antioxidant genes. Notably, PERK signaling intersects with cell cycle regulation by repressing cyclin D1 translation, promoting G1 arrest to allow stress recovery (Saito et al., 2011; Cullinan and Diehl, 2004; Saptarshi et al., 2022). Recent findings further suggest PERK may act as a dual-specificity kinase, with tyrosine phosphorylation (Tyr615) contributing to its full activation. In cancer, PERK exhibits both tumor-promoting and tumor-suppressing functions depending on context (Teske et al., 2011; Saito et al., 2011; Saptarshi et al., 2022; Yan et al., 2002). Recent studies show PERK’s tumor-promoting activity in cancers such as ovarian carcinoma, where the persistent activation of the PERK-eIF2α-ATF4 signaling pathway promotes cancer cell survival, chemoresistance, and suppression of antitumor immunity via expansion and activation of myeloid-derived suppressor cells (Chen and Cubillos-Ruiz, 2021; He et al., 2024). PERK activation may also exhibit tumor-suppressive functions through induction of apoptosis by promoting expression of pro-apoptotic factors such as CHOP or by stimulating antitumor immune responses through mechanisms like paraptosis (Gibson et al., 2022).
Lastly, among the four known eIF2α-related kinases (PERK, PKR, HRI, and GCN2), GCN2 is the only kinase conserved and functionally relevant in fungi. While PERK and PKR are central to mammalian responses to ER stress and viral infection respectively, fungal cells rely exclusively on GCN2 to regulate translational control during nutrient deprivation, particularly amino acid starvation (Hamanaka et al., 2005; Li et al., 2023). This makes GCN2 the most relevant kinase that phosphorylates eIF2α in fungi, it is directly involved with fungal stress adaptation pathways and is currently the focus of this present study in order to better understand eIF2α regulation in fungi and dimorphic fungi (Masson, 2019).
Recent findings by Smirnova et al. (2025) suggest that natural compounds derived from fungi may influence stress response pathways involving eIF2α. In their study, the effects of Wuling powder, a preparation derived from the mycelium of Xylaria nigripes, was explored based on its capability of alleviating depressive-like behavior in mice (Li et al., 2016). Their findings showed that a modulation of endoplasmic reticulum (ER) stress pathways, involving the phosphorylated eIF2α and its downstream effector, activates the transcription factor 4 (ATF4) (Li et al., 2016). The direct involvement of GCN2 was not explicitly examined. Still, their findings imply a potential role for GCN2 in the observed stress response modulation, demonstrating how fungal metabolites might influence this signaling axis, potentially contributing to stress alleviation and neuroprotection (Li et al., 2016; Lin et al., 2013).
A recent phylogenetic and bioinformatic study focused on black yeasts has identified the hypothetical kinase gene in the Herpotrichiellaceae family (Assunção et al., 2024). According to this study, eIF2α is involved in stress response, suggesting that kinase proteins in black fungi could be a potential target for antifungal therapy, based on the disruption of translation regulation that negatively affects their survival under host-induced stress (Assunção et al., 2024). More recent research has shown that even in the absence of stress, GCN2 is capable of preventing excessive ribosome biogenesis and mRNA translation, maintaining proteome stability (Román-Trufero et al., 2025). Another study focused on GCN2 signaling in mammals has shown that GCN2 influences lipid homeostasis by targeting key transcriptional regulators of lipogenesis, thereby linking the integrated stress response to the control of cellular lipid metabolism, which leads to the reinforcement of GCN2’s function in regulating cellular adaptation (Guo and Cavener, 2007).
On the other hand, in specific pathological states, the GCN2-mediated stress response can be detrimental. In the context of diabetic cardiomyopathy, reducing GCN2 levels has been shown to exert a cardioprotective effect (Yuan et al., 2022; Dabravolski et al., 2021). The mechanism for this protection involves the inhibition of the canonical eIF2α-ATF4-CHOP signaling pathway, leading to a decrease in oxidative stress, apoptosis, and harmful lipid accumulation in cardiac cells (Dabravolski et al., 2021). These seemingly contradictory findings show the context-dependent and tissue-specific role of GCN2 in metabolic regulation, where its activation can be either protective or detrimental depending on the physiological state.
Another study shows that GCN2 activation during leucine deprivation suppresses hepatic lipogenesis and promotes fatty acid oxidation, helping to prevent lipid accumulation in the liver (Kurokawa et al., 1998). GCN2-deficient mice displayed increased liver triglycerides and impaired expression of genes involved in β-oxidation (Kurokawa et al., 1998). This could signify that GCN2 is a promising therapeutic target to mitigate cardiac damage in diabetic patients, with the potential to counteract the cardiotoxic side effects of certain drugs (Yuan et al., 2022; Dabravolski et al., 2021). Which would fit in GCN2’s regulatory function.
Also, further studies show that in Pestalotiopsis microspora, an endophytic fungus able to digest polyurethane-based plastic materials, GCN2 deletion mutants impaired conidiation, disrupting secondary metabolism, and causing defective cell wall integrity, which indicates GCN2’s potential role in typical morphological development and stress responses in filamentous fungi (Jin et al., 2022). Regarding GCN2 inhibition research, studies have shown promising therapeutic potential, particularly in oncology and neurodegenerative disorders. A study details the potential of a GCN2 modulator to overcome drug resistance, a major clinical challenge, based on preclinical efficacy of HC-7366, a GCN2 activator, in AML models, which induces significant anti-tumor effects both as a standalone therapy and in combination with the standard-of-care drug (Tameire et al., 2023).
Drug resistance significantly impacts treatment options, which presents a challenge, especially when treatment options are limited. In view of this cancer research has been investigating novel therapeutical options, in special, an AACR abstract introduces CRD-799, a novel oral inhibitor that targets not only GCN2 but also two other related stress-response kinases, HRI and PERK. The study shows that in multiple myeloma, a cancer of plasma cells, combining CRD-799 with proteasome inhibitors (a standard treatment) can overcome drug resistance (Inoue et al., 2024). In the same line, another study investigates the preclinical effectiveness of a new GCN2 inhibitor, KAS-1155 on rhabdomyosarcoma, a rare cancer affecting soft tissue. By inhibiting GCN2, KAS-1155 could disrupt the cancer cells’ ability to adapt to stress, leading to cell death and opening the pathway for more viable therapeutic strategies (Kang et al., 2024).
Expanding the therapeutic potential of GCN2 inhibitors beyond oncology, this study investigates their role in the neurodegenerative disease ALS. The research found that inhibiting GCN2 could reduce the toxic aggregation of mutant SOD1 protein, a crucial pathological element in some forms of ALS. In a mouse model of the disease, GCN2 inhibition delayed disease progression, suggesting that this mechanism could be a promising neuroprotective strategy (Ortiz et al., 2024). The preclinical success of these GCN2 inhibitors in complex mammalian disease models provides a strong rationale for exploring their efficacy as antifungal agents.
It is important to consider the effects of known pharmacological inhibitors when it comes to the therapeutic relevance of GCN2, studies have shown that ATP-competitive kinase inhibitors can inadvertently activate GCN2 by stabilizing its active conformation, rather than inhibiting it (Tang et al., 2022; Kato et al., 2020; Nakamura et al., 2018). This paradoxical activation suggests that drug interactions with GCN2 are more complex than previously assumed and could influence outcomes in diseases where GCN2 plays a regulatory role (Kato et al., 2020). Selective GCN2 inhibitors such as GCN2iB have demonstrated efficacy in modulating stress responses and suppressing tumor growth in preclinical models (Kato et al., 2020; Nakamura et al., 2018). It is possible to conclude that inhibiting GCN2 in fungal pathogens could offer therapeutic benefits in diverse fields. Dissecting the specific roles of this conserved pathway in pathogenic fungi remains a crucial step toward validating GCN2 as a robust and safe antifungal drug target.
7 Conclusion
• This review provides a detailed analysis of the eIF2α kinase GCN2 and its role in fungal amino acid sensing and stress response. We focus on the molecular mechanisms of GCN2 function and it signaling pathways, while also surveying the latest research trends in the field. This growing interest points toward underexplored fungi and mechanisms that could drive future discoveries, setting a path for advancing the field of microbiology.
• Adaptive capacity directly impacts fungal survival, development, and pathogenicity. As the only conserved and active eIF2α-related kinase in fungi, GCN2 has a strong potential to become a universal stress response marker. Such a marker can be particularly valuable for genetically diverse pathogenic fungi, where identifying conserved pathways is often a challenge.
• Kinase GCN2’s role and conservation in eukaryotes have shown its potential as a target for novel antifungal therapies, especially as translational control gains recognition for pathogen management. Beyond its role in sensing amino acid deprivation, recent studies indicate that GCN2 also participates in other cellular processes, such as lipid homeostasis, linking translational control to more ample metabolic regulation. This has spurred the investigation of GCN2 inhibitors, many initially developed for oncology, as potential antifungal agents.
• A critical consideration for GCN2 as a therapeutic target is its feasibility, given the existence of a human homolog, EIF2AK4. While the catalytic kinase domain is highly conserved across eukaryotes, significant structural divergences exist between fungal and human GCN2, particularly in the regulatory domains. As this review notes, regions such as the C-terminal domain (CTD) and dimerization interfaces show low sequence similarity when comparing fungal and vertebrate kinases. These differences in the non-catalytic domains, which are crucial for controlling kinase activation, offer a promising strategy for the rational design of fungus-specific inhibitors. By developing molecules that target these less-conserved regulatory regions instead of the active site, it is feasible to achieve high specificity, thereby minimizing off-target effects on the human homolog.
• Considering the rising challenge of fungal resistance, a deep understanding of how fungi adapt at the molecular level is more critical than ever. We believe this framework will guide future research and encourage collaboration across microbiology, genomics, structural biology, and drug discovery, ultimately supporting efforts to combat fungal diseases more effectively.
Author contributions
ES: Formal analysis, Writing – original draft, Visualization, Data curation, Methodology, Validation, Investigation, Software, Conceptualization, Writing – review & editing. CA: Formal analysis, Validation, Data curation, Writing – review & editing, Conceptualization, Methodology, Software, Investigation, Visualization. TC: Validation, Visualization, Formal analysis, Methodology, Investigation, Writing – review & editing. IR: Formal analysis, Visualization, Investigation, Writing – review & editing, Validation, Software, Methodology. SC: Writing – review & editing, Data curation, Visualization, Validation. EA: Validation, Methodology, Writing – review & editing, Software, Visualization. TS: Writing – review & editing, Visualization, Validation. RC: Data curation, Supervision, Investigation, Methodology, Conceptualization, Software, Validation, Formal analysis, Resources, Visualization, Funding acquisition, Project administration, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. We gratefully acknowledge the financial support provided by FAPEMIG (Fundação de Amparo à Pesquisa do Estado de Minas Gerais)—grant 03767/2023 and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico)—grant 309088/2023-2.
Acknowledgments
We would like to express our gratitude to the colleagues of the Microbiology and Bioinformatics Research Department in the Faculdade Santa Casa de Belo Horizonte, for their valuable support and contributions to this work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1630196/full#supplementary-material
References
Aria, M. (2017). Bibliometrix: an R-tool for comprehensive science mapping analysis. J. Informetr. 11, 959–975. doi: 10.1016/j.joi.2017.08.007
Aryal, S., and Watts, V. J. (2025). Molecular pharmacology studies of adenylyl cyclase 1 (AC1) regulation: calmodulin-mediated regulation and homodimerization (abstract ID: 165256). J. Pharmacol. Exp. Ther. 392:100696. doi: 10.1016/j.jpet.2024.100696
Asada, N., Ginsberg, P., Paust, H. J., Song, N., Riedel, J. H., Turner, J. E., et al. (2025). The integrated stress response pathway controls cytokine production in tissue-resident memory CD4+ T cells. Nat. Immunol. 26, 557–566. doi: 10.1038/s41590-025-02105-x
Assunção, C. B., de Aguiar, E. L., al-Hatmi, A. M. S., Silva Vieira, V. C., Machado, A. S., Junta, C., et al. (2020). New molecular marker for phylogenetic reconstruction of black yeast-like fungi (Chaetothyriales) with hypothetical EIF2AK2 kinase gene. Fungal Biol. 124, 1032–1038. doi: 10.1016/J.FUNBIO.2020.09.007
Assunção, C. B., de Aguiar, E. L., Chávez-Fumagalli, M. A., La Santrer, E. R., Dias, S. R., de Souza Rodrigues, T., et al. (2024). In silico characterization of an initiation factor 2 kinase of black fungi: a potential drug target for mycosis. Curr. Biotechnol. 13, 107–118. doi: 10.2174/0122115501285434240409040348
Bartelt, D. C., Fidelt, S., Farbert, L. H., Wolff, D. J., and Hammell, R. L. (1988). Calmodulin-dependent multifunctional protein kinase in aspergillus nidulans Proc Natl Acad Sci USA. 85: 3279–83. doi: 10.1073/pnas.85.10.3279
Berkes, C., Franco, J., Lawson, M., Brann, K., Mermelstein, J., Laverty, D., et al. (2021). Kinase inhibitor library screening identifies the cancer therapeutic sorafenib and structurally similar compounds as strong inhibitors of the fungal pathogen Histoplasma capsulatum. Antibiotics 10, 1–13. doi: 10.3390/antibiotics10101223
Bhattacharya, R., Sourirajan, A., Sharma, P., Kumar, A., Upadhyay, N. K., Shukla, R. K., et al. (2023). Bioenhancer potential of Aegle marmelos (L.) Corrêa essential oil with antifungal drugs and its mode of action against Candida albicans. Biocatal. Agric. Biotechnol. 48:102647. doi: 10.1016/j.bcab.2023.102647
Bou-Nader, C., Gaikwad, S., Bahmanjah, S., Zhang, F., Hinnebusch, A. G., and Zhang, J. (2024). Gcn2 structurally mimics and functionally repurposes the HisRS enzyme for the integrated stress response. Proc. Natl. Acad. Sci. USA 121:e2409628121. doi: 10.1073/pnas.2409628121
Brunke, S., Mogavero, S., Kasper, L., and Hube, B. (2016). Virulence factors in fungal pathogens of man. Curr. Opin. Microbiol. 32, 89–95. doi: 10.1016/j.mib.2016.05.010
Budovskaya, Y. V., Stephan, J. S., Deminoff, S. J., and Herman, P. K. (2005) An evolutionary proteomics approach identifies substrates of the cAMP-dependent protein kinase. Available online at: www.yeastgenome.org.
Burwick, N., and Aktas, B. H. (2017). The eIF2-alpha kinase HRI: a potential target beyond the red blood cell. Expert Opin. Ther. Targets 21, 1171–1177. doi: 10.1080/14728222.2017.1397133
Cañete-Gibas, C. F., and Wiederhold, N. P. (2018). The black yeasts: an update on species identification and diagnosis. Curr. Fungal Infect. Rep. 12, 59–65. doi: 10.1007/s12281-018-0314-0
Carbonero, F., and Strobel, G. (2021). Fungal ecology special issue: editorial. Microb. Ecol. 82, 1–4. doi: 10.1007/s00248-021-01784-x
Castilho, B. A., Shanmugam, R., Silva, R. C., Ramesh, R., Himme, B. M., and Sattlegger, E. (2014). Keeping the eIF2 alpha kinase Gcn2 in check. Biochim. Biophys. Acta 1843, 1948–1968. doi: 10.1016/j.bbamcr.2014.04.006
Chadwick, B. J. (2023). Investigating the biology and genetic basis of stress adaptation in Cryptococcus neoformans. Athens, Georgia: The University of Georgia.
Chen, X., and Cubillos-Ruiz, J. R. (2021). Endoplasmic reticulum stress signals in the tumour and its microenvironment. Nat. Rev. Cancer 21, 71–88. doi: 10.1038/s41568-020-00312-2
Cheng, H. C., Qi, R. Z., Paudel, H., and Zhu, H. J. (2011). Regulation and function of protein kinases and phosphatases. Enzyme Res. 2011, 1–3. doi: 10.4061/2011/794089
Clemens, M. J. (1976). Functional relationships between a reticulocyte polypeptide-chain-initiation factor (IF-MP) and the translational inhibitor involved in regulation of protein synthesis by haemin. Eur. J. Biochem. 66, 413–422. doi: 10.1111/j.1432-1033.1976.tb10531.x
Cohen, P. (2002) The origins of protein phosphorylation. Nat Cell Biol. 4, E127–E130. doi: 10.1038/ncb0502-e127
Cui, W., Li, J., Ron, D., and Sha, B. (2011). The structure of the PERK kinase domain suggests the mechanism for its activation. Acta Crystallogr. D Biol. Crystallogr. 67, 423–428. doi: 10.1107/S0907444911006445
Cullinan, S. B., and Diehl, J. A. (2004). PERK-dependent activation of Nrf2 contributes to redox homeostasis and cell survival following endoplasmic reticulum stress. J. Biol. Chem. 279, 20108–20117. doi: 10.1074/jbc.M314219200
Dabravolski, S. A., Bezsonov, E. E., Baig, M. S., Popkova, T. V., and Orekhov, A. N. (2021). Mitochondrial lipid homeostasis at the crossroads of liver and heart diseases. Int. J. Mol. Sci. 22:6949. doi: 10.3390/ijms22136949
Dar, A. C., Dever, T. E., and Sicheri, F. (2005). Higher-order substrate recognition of eIF2α by the RNA-dependent protein kinase PKR. Cell 122, 887–900. doi: 10.1016/j.cell.2005.06.044
Davis, R. H. (1986). Compartmental and regulatory mechanisms in the arginine pathways of Neurospora crassa and Saccharomyces cerevisiae. Microbiol. Rev. 50, 280–313. doi: 10.1128/mr.50.3.280-313.1986
de Hoog, G. S., Guarro, J., Gené, J., Ahmed, S. A., Al-Hatmi, A. M. S., and Figueras, M. J. (2023). Atlas of Clinical Fungi. 4th Edn. Utrecht, ND: Uppsalalaan.
De La Cruz-Herrera, C. F., Campagna, M., García, MA., Marcos-Villar, L., Lang, V., Baz-Martínez, M., et al. (2014). Activation of the double-stranded RNA-dependent protein kinase PKR by small ubiquitin-like modifier (SUMO). J. Biol. Chem. 289, 26357–26367. doi: 10.1074/jbc.M114.560961
Defosse, T. A., Sharma, A., Mondal, A. K., Dugé de Bernonville, T., Latgé, J. P., Calderone, R., et al. (2015). Hybrid histidine kinases in pathogenic fungi. Mol. Microbiol. 95, 914–924. doi: 10.1111/mmi.12911
Dhillon, N. K., Sharma, S., and Khuller, G. K. (2003). Signaling through protein kinases and transcriptional regulators in Candida albicans. Crit. Rev. Microbiol. 29, 259–275. doi: 10.1080/713610451
Dong, J., Qiu, H., Garcia-Barrio, M., Anderson, J., and Hinnebusch, A. G. (2000). Uncharged tRNA activates GCN2 by displacing the protein kinase moiety from a bipartite tRNA-binding domain. Mol. Cell 6, 269–279. doi: 10.1016/S1097-2765(00)00028-9
Donnelly, N., Gorman, A. M., Gupta, S., and Samali, A. (2013). The eIF2α kinases: their structures and functions. Cell. Mol. Life Sci. 70, 3493–3511. doi: 10.1007/s00018-012-1252-6
Douglas, J., Bouckaert, R., Carter, C. W., and Wills, P. R. (2024). Enzymic recognition of amino acids drove the evolution of primordial genetic codes. Nucleic Acids Res. 52, 558–571. doi: 10.1093/nar/gkad1160
Ekhator, E. S., Fazzari, M., and Newman, R. H. (2025). Redox regulation of cAMP-dependent protein kinase and its role in health and disease. Life 15:655. doi: 10.3390/life15040655
Elsby, R., Heiber, J. F., Reid, P., Kimball, S. R., Pavitt, G. D., and Barber, G. N. (2011). The alpha subunit of eukaryotic initiation factor 2B (eIF2B) is required for eIF2-mediated translational suppression of vesicular stomatitis virus. J. Virol. 85, 9716–9725. doi: 10.1128/jvi.05146-11
Emery, P. W. (2012). Amino Acids: Chemistry and Classification, Editor(s): Benjamin Caballero, Encyclopedia of Human Nutrition. Baltimore, MD, EUA: Academic Press. 64–71. doi: 10.1016/B978-0-12-375083-9.00009-X
Ene, I. V., Brunke, S., Brown, A. J. P., and Hube, B. (2014). Metabolism in fungal pathogenesis. Cold Spring Harb. Perspect. Med. 4:a019695. doi: 10.1101/cshperspect.a019695
Evelyn, S., and Hinnebusch, A. G. (2000). Separate domains in GCN1 for binding protein kinase GCN2 and ribosomes are required for GCN2 activation in amino acid-starved cells. EMBO J. 19, 6622–6633. doi: 10.1093/emboj/19.23.6622
Farrer, R. A., and Fisher, M. C. (2017). Describing genomic and Epigenomic traits underpinning emerging fungal pathogens. Adv. Genet. 100, 73–140. doi: 10.1016/bs.adgen.2017.09.009
Fuller, K. K., Richie, D. L., Feng, X., Krishnan, K., Stephens, T. J., Wikenheiser-Brokamp, K. A., et al. (2011). Divergent protein kinase a isoforms co-ordinately regulate conidial germination, carbohydrate metabolism and virulence in Aspergillus fumigatus. Mol. Microbiol. 79, 1045–1062. doi: 10.1111/j.1365-2958.2010.07509.x
Gal-Ben-Ari, S., Barrera, I., Ehrlich, M., and Rosenblum, K. (2019). PKR: a kinase to remember. Front. Mol. Neurosci. 11:480. doi: 10.3389/fnmol.2018.00480
Gao, X., Jin, C., Ren, J., Yao, X., and Xue, Y. (2008). Proteome-wide prediction of PKA phosphorylation sites in eukaryotic kingdom. Genomics 92, 457–463. doi: 10.1016/j.ygeno.2008.08.013
Garbe, E., and Vylkova, S. (2019). Role of amino acid metabolism in the virulence of human pathogenic fungi. Curr. Clin. Microbiol. Rep. 6:108–119. doi: 10.1007/s40588-019-00124-5
Gauthier, G. M. (2017). Fungal dimorphism and virulence: molecular mechanisms for temperature adaptation, immune evasion, and in vivo survival. Mediat. Inflamm. 2017, 1–8. doi: 10.1155/2017/8491383
Gibson, K., Chu, J. K., Zhu, S., Nguyen, D., Mrázek, J., Liu, J., et al. (2022). A tripartite efflux system affects flagellum stability in Helicobacter pylori. Int. J. Mol. Sci. 23, 1–19. doi: 10.3390/ijms231911609
Girardin, S. E., Cuziol, C., Philpott, D. J., and Arnoult, D. (2021). The eIF2α kinase HRI in innate immunity, proteostasis, and mitochondrial stress. FEBS J. 288, 3094–3107. doi: 10.1111/febs.15553
Gottfried, S., Koloamatangi, S. M. B. M. J., Daube, C., Schiemann, A. H., and Sattlegger, E. (2022). A genetic approach to identify amino acids in Gcn1 required for Gcn2 activation. PLoS One 17, 1–25. doi: 10.1371/journal.pone.0277648
Gow, N. A. R., Latge, J.-P., and Munro, C. A. (2017). The fungal cell wall: structure, biosynthesis, and function. Microbiol. Spectr. 5, 1–25. doi: 10.1128/microbiolspec.funk-0035-2016
Guo, F., and Cavener, D. R. (2007). The GCN2 eIF2α kinase regulates fatty-acid homeostasis in the liver during deprivation of an essential amino acid. Cell Metab. 5, 103–114. doi: 10.1016/j.cmet.2007.01.001
Gupta, R., Gaikwad, S., Qui, H., Bou-Nader, C., Zhang, J., and Hinnebusch, A. G. (2025). “Purification and analysis of eIF2α phosphorylation by stress-activated protein kinase Gcn2 from S. cerevisiae” in Nutrient Sensing in Eukaryotes. eds. D. G. Russell and F. M. Hardie (New York, NY: Springer US), 195–220.
Gupta Roy, B., and Datta, A. (1986). A cyclic AMP-independent protein kinase from Candida albicans. Biochem. J. 234, 543–546. doi: 10.1042/bj2340543
Hamanaka, R. B., Bennett, B. S., Cullinan, S. B., and Diehl, J. A. (2005). PERK and GCN2 contribute to eIF2α phosphorylation and cell cycle arrest after activation of the unfolded protein response pathway. Mol. Biol. Cell 16, 5493–5501. doi: 10.1091/mbc.E05-03-0268
He, H., Singh, I., Wek, S. A., Dey, S., Baird, T. D., Wek, R. C., et al. (2014). Crystal structures of GCN2 protein kinase C-terminal domains suggest regulatory differences in yeast and mammals. J. Biol. Chem. 289, 15023–15034. doi: 10.1074/jbc.M114.560789
He, J., Zhou, Y., and Sun, L. (2024). Emerging mechanisms of the unfolded protein response in therapeutic resistance: from chemotherapy to immunotherapy. BioMed Central Ltd. 22, 1–22. doi: 10.1186/s12964-023-01438-0
Hinnebusch, A. G. (2005). Translational regulation of GCN4 and the general amino acid control of yeast. Ann. Rev. Microbiol. 59, 407–450. doi: 10.1146/annurev.micro.59.031805.133833
Hinnebusch, A. G. (2011). Molecular mechanism of scanning and start codon selection in eukaryotes. Microbiol. Mol. Biol. Rev. 75, 434–467. doi: 10.1128/mmbr.00008-11
Hixson, C. S., and Krebs, E. G. (1980). Characterization of a cyclic AMP-binding protein from bakers’ yeast. Identification as a regulatory subunit of cyclic AMP-dependent protein kinase. J. Biol. Chem. 255, 2137–2145. doi: 10.1016/s0021-9258(19)86004-0
Hossain, S., Liu, Z., Robbins, N., and Cowen, L. E. (2025). Exploring the differential localization of protein kinase a isoforms in Candida albicans. mSphere 10:e0103724. doi: 10.1128/msphere.01037-24
Hu, G., Steen, B. R., Lian, T., Sham, A. P., Tam, N., Tangen, K. L., et al. (2007). Transcriptional regulation by protein kinase a in Cryptococcus neoformans. PLoS Pathog. 3:e42. doi: 10.1371/journal.ppat.0030042
Inoue, Y, Hikami, K, Ebara, S, Sugiyama, M, Oda, R., Mizutani, A., et al. (2024). Abstract 1745: combining HRI/PERK/GCN2 inhibitor CRD-799 with proteasome inhibitors provides a novel approach to overcoming resistance in multiple myeloma treatment. Cancer Res. 84:1745. doi: 10.1158/1538-7445.AM2024-1745
Jennings, M. D., Kershaw, C. J., Tomas, A., Pavitt, G. D., et al. (2017). Fail-safe control of translation initiation by dissociation of eIF2a phosphorylated ternary complexes. eLife. 6:e24542. doi: 10.7554/eLife.24542
Jernejc, K., and Benčina, M. (2003). Lipid composition of cAMP-dependent protein kinase mutants of Aspergillus niger. FEMS Microbiol. Lett. 225, 291–297. doi: 10.1016/S0378-1097(03)00532-9
Jin, J., Yang, L., Li, Q., Tao, Q., and Yang, Y. (2022). Amino acid sensor kinase Gcn2 is required for conidiation, secondary metabolism, and cell wall integrity in the Taxol-producer Pestalotiopsis microspora. J. Fungi 8:390. doi: 10.3390/jof8040390
Judewicz, N. D., Gerard, C. G., and Torres, H. N. (1981) “Protein kinase activities in Neurospora crassa”.
Juliani, M. H., Brochetto, M. R., Da, J. C., and Maia, C. (1979). Changes in cyclic AMP binding and protein kinase activities during growth and differentiation of Blastocladiella emeronh. Cell Differ. 8:421. doi: 10.1016/0045-6039(79)90038-1
Jungmann, R. A., and Russell, D. H. (1977). Cyclic AMP, cyclic AMP-dependent protein kinase, and the regulation of gene expression. Life Sci. 20, 1787–1798. doi: 10.1016/0024-3205(77)90213-2
Kang, DY, Lee, HW, Jong, WY, Jung-Nyoung, H, Choi, Y., Min Hahn, S., et al. (2024). Abstract 6447: preclinical efficacy of GCN2 inhibition by KAS-1155 in targeting the integrated stress response in rhabdomyosarcoma. Cancer Res. 84:6447. doi: 10.1158/1538-7445.AM2024-6447
Kapp, L. D., and Lorsch, J. R. (2004). GTP-dependent recognition of the methionine moiety on initiator tRNA by translation factor eIF2. J. Mol. Biol. 335, 923–936. doi: 10.1016/j.jmb.2003.11.025
Kato, Y., Kunimasa, K., Takahashi, M., Harada, A., Nagasawa, I., Osawa, M., et al. (2020). GZD824 Inhibits GCN2 and Sensitizes Cancer Cells to Amino Acid Starvation Stress. Mol Pharmacol. 98, 669–676. doi: 10.1124/molpharm.120.000070
Kerner, N., and Passeron, S. (1984). Cyclic AMP-dependent protein kinase from Ustilago maydis. Mol. Cell. Biochem. 60, 115–122. doi: 10.1007/BF00222481
Khuller, G., Sharma, S., and Deo, D. (2000). Dermatophyte lipids-composition and regulation of phospholipids. Indian J. Clin. Biochem. 15, 51–59. doi: 10.1007/BF02867544
Kim, J. S., Lee, K. T., Lee, M. H., Cheong, E., and Bahn, Y. S. (2021). Adenylyl cyclase and protein kinase a play redundant and distinct roles in growth, differentiation, antifungal drug resistance, and pathogenicity of Candida auris. MBio 12, e02729–21. doi: 10.1128/mBio.02729-21
Kim, H.-S., Park, S.-Y., Lee, S., Adams, E. L., Czymmek, K., and Kang, S. (2011). Loss of cAMP-dependent protein kinase a affects multiple traits important for root pathogenesis by fusarium oxysporum. Mol. Plant Microbe Interact. 24, 719–732. doi: 10.1094/MPMI-11-10-0267
Kimball, S. R. (1999). “Eukaryotic initiation factor eIF2.” Int J Biochem Cell Biol. 31, 25–9. doi: 10.1016/s1357-2725(98)00128-9
Klein, B. S., and Tebbets, B. Dimorphism and virulence in fungi. Curr. Opin. Microbiol., 10, 314–319 (2007). Available online at: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3412142/pdf/nihms31336.pdf
Knowles, C. M., McIntyre, K. M., and Panepinto, J. C. (2021). Tools for assessing translation in Cryptococcus neoformans. J. Fungi 7:159. doi: 10.3390/jof7030159
Komar, A. A., and Merrick, W. C. (2020). A retrospective on EIF2A—and not the alpha subunit of EIF2. Int. J. Mol. Sci. 21:2054. doi: 10.3390/ijms21062054
Kumar, A. (2018). A fungus among us: the emerging opportunistic pathogen Candida tropicalis and PKA signaling. Virulence 9, 659–661. doi: 10.1080/21505594.2018.1438026
Kurokawa, C. S., Sugizaki, M. F., and Peraçoli, M. T. (1998). Virulence factor of systemic mycoses. Rev. Inst. Med. Trop. 40, 125–135. doi: 10.1590/S0036-46651998000300001
Leipheimer, J., Bloom, A. L. M., Campomizzi, C. S., Salei, Y., and Panepinto, J. C. (2019). Translational regulation promotes oxidative stress resistance in the human fungal pathogen Cryptococcus neoformans. doi: 10.1128/mBio
Li, Y., Yang, Y., Chen, B., Zhao, M., and Zhu, J. (2024). The GCN4 transcription factor: a review of its functional progress in fungi. Horticulturae 10:1113. doi: 10.3390/horticulturae10101113
Li, Y., Zhang, J., Shen, Q., and Yu, Z. (2023). Fungi employ the GCN2 pathway to maintain the circadian clock under amino acid starvation. Innovation Life 1:100026. doi: 10.59717/j.xinn-life.2023.100026
Li, D., Zheng, J., Wang, M., Feng, L., Liu, Y., Yang, N., et al. (2016). Wuling powder prevents the depression-like behavior in learned helplessness mice model through improving the TSPO mediated-mitophagy. J. Ethnopharmacol. 186, 181–188. doi: 10.1016/j.jep.2016.03.065
Lin, Y., Wang, X. Y., Ye, R., Hu, W. H., Sun, S. C., Jiao, H. J., et al. (2013). Efficacy and safety of Wuling capsule, a single herbal formula, in Chinese subjects with insomnia: a multicenter, randomized, double-blind, placebo-controlled trial. J. Ethnopharmacol. 145, 320–327. doi: 10.1016/J.JEP.2012.11.009
Liu, X., Afrin, T., and Pajerowska-Mukhtar, K. M. (2019). Arabidopsis GCN2 kinase contributes to ABA homeostasis and stomatal immunity. Commun Biol 2:302. doi: 10.1038/s42003-019-0544-x
Lokdarshi, A., von Arnim, A. G., and Akuoko, T. K. (2022). Modulation of GCN2 activity under excess light stress by osmoprotectants and amino acids. Plant Signal. Behav. 17, 1681–1693. doi: 10.1080/15592324.2022.2115747
Lr, R. (n.d.) “Scientometric study of global mucormycosis (black fungus) research.” Available online at: https://www.researchgate.net/publication/389635958
Lübbehüsen, T., Polo, V. G., Rossi, S., Nielsen, J., Moreno, S., McIntyre, M., et al. (2004). Protein kinase a is involved in the control of morphology and branching during aerobic growth of Mucor circinelloides. Microbiology 150, 143–150. doi: 10.1099/mic.0.26708-0
Maeng, S., Ko, Y. J., Kim, G. B., Jung, K. W., Floyd, A., Heitman, J., et al. (2010). Comparative transcriptome analysis reveals novel roles of the ras and cyclic AMP signaling pathways in environmental stress response and antifungal drug sensitivity in Cryptococcus neoformans. Eukaryot. Cell 9, 360–378. doi: 10.1128/EC.00309-09
Maloy, S. (2013). “Amino Acids” in Brenner’s encyclopedia of genetics: Second edition (Elsevier Inc.), 108–110.
Martin, J., Anamika, K., and Srinivasan, N. (2010). Classification of protein kinases on the basis of both kinase and non-kinase regions. PLoS One 5, 1–12. doi: 10.1371/journal.pone.0012460
Marton, T. J., Crouch, D., and Hinnebusch, A. G. (1993). GCN1, a translational activator of GCN4 in Saccharomyces cerevisiae, is required for phosphorylation of eukaryotic translation initiation factor 2 by protein kinase GCN2. Mol Cell Biol. 13:3541–56. doi: 10.1128/mcb.13.6.3541-3556.1993
Masson, G. R. (2019). Towards a model of GCN2 activation. Biochem. Soc. Trans. 47, 1484–1488. doi: 10.1042/BST20190331
Mattoon, E. R., Casadevall, A., and Cordero, R. J. (2021). Beat the heat: correlates, compounds, and mechanisms involved in fungal thermotolerance. Fungal Biol. Rev. 36, 60–75. doi: 10.1016/J.FBR.2021.03.002
Maurin, A. C., Jousse, C., Averous, J., Parry, L., Bruhat, A., Cherasse, Y., et al. (2005). The GCN2 kinase biases feeding behavior to maintain amino acid homeostasis in omnivores. Cell Metab. 1, 273–277. doi: 10.1016/j.cmet.2005.03.004
Maurin, A.-C., Parry, L., B’chir, W., Carraro, V., Coudy-Gandilhon, C., Chaouki, G., et al. (2022). GCN2 upregulates autophagy in response to short-term deprivation of a single essential amino acid. Autophagy Rep. 1, 119–142. doi: 10.1080/27694127.2022.2049045
McCarthy, M. W., and Walsh, T. J. (2018). Amino acid metabolism and transport mechanisms as potential antifungal targets. Int. J. Mol. Sci. 19, 909–112. doi: 10.3390/ijms19030909
McDonough, K. A., and Rodriguez, A. (2012). The myriad roles of cyclic AMP in microbial pathogens: from signal to sword. Nat. Rev. Microbiol. 10, 27–38. doi: 10.1038/nrmicro2688
Mcginnis, M. R., and Tyring, S. K. (1996). “Introduction to mycology” in Medical microbiology. ed. S. Baron. 4th ed (Galveston, Texas: University of Texas Medical Branch at Galveston).
Money, N. P. (2024). Fungal thermotolerance revisited and why climate change is unlikely to be supercharging pathogenic fungi (yet). Fungal Biol. 128, 1638–1641. doi: 10.1016/j.funbio.2024.01.005
Moreno, S., Paveto, C., and Passeron, S. (1977). Multiple protein kinase activities in the dimorphic fungus Mucor rouxii: comparison with a cyclic adenosine 3′,5′-monophosphate binding protein. Arch. Biochem. Biophys. 180, 225–231. doi: 10.1016/0003-9861(77)90032-7
Muhammad, Z. U. A., Shoaib, A., Xinwei, W., Shengwei, H., and Changhong, L. (2024). Amino acids could sustain fungal life in the energy-limited anaerobic sediments below the seafloor. Appl. Environ. Microbiol. 90, e01279–e01224. doi: 10.1128/aem.01279-24
Müller, D., Exler, S., Aguilera-Vázquez, L., Guerrero-Martín, E., and Reuss, M. (2003). Cyclic AMP mediates the cell cycle dynamics of energy metabolism in Saccharomyces cerevisiae. Yeast 20, 351–367. doi: 10.1002/yea.967
Nabeshima, S., Tanaka, A., and Fukui, S. (1977). Effect of Biotin on the Level of Isocitrate Lyase in Candida tropicalis. Agric. Biol. Chem., 41, 281–285. doi: 10.1080/00021369.1977.10862488
Nakamura, A., Nambu, T., Ebara, S., Hasegawa, Y., Toyoshima, K., Tsuchiya, Y., et al. (2018). Inhibition of GCN2 sensitizes multiple myeloma to proteasome inhibitors. Proc Natl Acad Sci USA. 115:E7776–E7785. doi: 10.1073/pnas.1805523115
Nameki, N., Yoneyama, M., Koshiba, S., Tochio, N., Inoue, M., Seki, E., et al. (2004). Solution structure of the RWD domain of the mouse GCN2 protein. Protein Sci. 13, 2089–2100. doi: 10.1110/ps.04751804
Narayanan, J., Tamilanban, T., Kumar, P. S., Guru, A., Muthupandian, S., Kathiravan, M. K., et al. (2023). Role and mechanistic actions of protein kinase inhibitors as an effective drug target for cancer and COVID. Arch. Microbiol. 205–238. doi: 10.1007/s00203-023-03559-z
Neiman, A. M., Stevenson, B. J., Xu, H. P., Sprague, G. F., Herskowitz, I., Wigler, M., et al. (1993). Functional homology of protein kinases required for sexual differentiation in Schizosaccharomyces pombe and Saccharomyces cerevisiae suggests a conserved signal transduction module in eukaryotic organisms. Mol. Biol. Cell 4, 107–120. doi: 10.1091/mbc.4.1.107
Ocampo, J., Fernandez Nuñez, L., Silva, F., Pereyra, E., Moreno, S., Garre, V., et al. (2009). A subunit of protein kinase a regulates growth and differentiation in the fungus Mucor circinelloides. Eukaryot. Cell 8, 933–944. doi: 10.1128/EC.00026-09
Ohlmeier, S., Hiltunen, J. K., and Bergmann, U. (2010). Protein phosphorylation in mitochondria – a study on fermentative and respiratory growth of Saccharomyces cerevisiae. Electrophoresis 31, 2869–2881. doi: 10.1002/elps.200900759
Olicón-Hernández, D., Hernández-Lauzardo, A., Pardo, J. P., Peña, A., Valle, M., and Guerra-Sanchez, G. (2015). Influence of chitosan and its derivatives on cell development and physiology of Ustilago maydis. Int. J. Biol. Macromol. 79, 654–660. doi: 10.1016/j.ijbiomac.2015.05.057
Ortiz, D. A., Peregrín, N., Valencia, Vinueza-Gavilanes, R., Marín-Ordovas, E., Ferrero, R., Jesús Nicolás, M., et al. (2024). GCN2 inhibition reduces mutant SOD1 clustering and toxicity and delays disease progression in an amyotrophic lateral sclerosis mouse model. iScience 27:109968. doi: 10.1016/j.isci.2024.109968
Padyana, A. K., Qiu, H., Roll-Mecak, A., Hinnebusch, A. G., and Burley, S. K. (2005). Structural basis for autoinhibition and mutational activation of eukaryotic initiation factor 2α protein kinase GCN2. J. Biol. Chem. 280, 29289–29299. doi: 10.1074/jbc.M504096200
Pakos-Zebrucka, K., Koryga, I., Mnich, K., Ljujic, M., Samali, A., and Gorman, A. M. (2016). The integrated stress response. EMBO Rep. 17, 1374–1395. doi: 10.15252/embr.201642195
Pastori, R., Moreno, S., and Passeron, S. (1985) Polymeric structure of the cyclic AMP-dependent protein kinase from the dimorphic fungus Mucor rouxii and purification of its catalytic subunit. Mol Cell Biochem. 69, 55–66. doi: 10.1007/BF00225927
Pathan, E. K., Ghormade, V., Panmei, R., and Deshpande, M. V. (2019). “Biochemical and molecular aspects of dimorphism in fungi” in Advancing Frontiers in mycology and Mycotechnology: Basic and applied aspects of Fungi (Springer Singapore), 69–94.
Perlatti, B., Harris, G., Nichols, C. B., Ekanayake, D. I., Alspaugh, J. A., Gloer, J. B., et al. (2020). Campafungins: inhibitors of Candida albicans and Cryptococcus neoformans hyphal growth. J. Nat. Prod. 83, 2718–2726. doi: 10.1021/acs.jnatprod.0c00641
Piecyk, M., Ferraro-Peyret, C., Laville, D., Perros, F., and Chaveroux, C. (2024). Novel insights into the GCN2 pathway and its targeting. Therapeutic value in cancer and lessons from lung fibrosis development. FEBS J. 291, 4867–4889. doi: 10.1111/febs.17203
Pir, N. A., Mir, Z. A., Rather, A. M., Qureshi, W., and Majid, S. (2025). “Role of cAMP–PKA–CREB signaling pathway in Cancer” in Cell signaling pathways and their therapeutic implication in cancers. eds. M. U. Rehman and M. S. Khan (Singapore: Springer Nature Singapore), 265–291.
Plattner, F., and Bibb, J. A. (2011). “Serine and threonine phosphorylation” in Basic neurochemistry: Principles of molecular, cellular, and medical neurobiology: Eighth Edition (Elsevier), 467–492. doi: 10.1016/C2009-0-00066-X
Powers, P. A., and Pall, M. L. (1980). Cyclic AMP-dependent protein kinase of Neurospora crassa. Biochem. Biophys. Res. Commun. 95, 701–706. doi: 10.1016/0006-291X(80)90842-6
Rakesh, R., Krishnan, R., Sattlegger, E., and Srinivasan, N. (2017). Recognition of a structural domain (RWDBD) in Gcn1 proteins that interacts with the RWD domain containing proteins. Biol. Direct 12, 12–20. doi: 10.1186/s13062-017-0184-3
Ramirez, M., Wek, R. C., and Hinnebusch, A. G. (1991). Ribosome association of GCN2 protein kinase, a translational activator of the GCN4 gene of Saccharomyces cerevisiae. Mol. Cell. Biol. 11, 3027–3036. doi: 10.1128/MCB.11.6.3027
Ramírez-Sotelo, U., Gómez-Gaviria, M., and Mora-Montes, H. M. (2025). Signaling pathways regulating dimorphism in medically relevant fungal species. 14:350. doi: 10.20944/preprints202502.1154.v1
Ranjan, N., Pochopien, A. A., Chih-Chien Wu, C., Beckert, B., Blanchet, S., Green, R., et al. (2021). Yeast translation elongation factor eEF3 promotes late stages of tRNA translocation. EMBO J. 40:e106449. doi: 10.15252/embj.2020106449
Rauch, J., Volinsky, N., Romano, D., and Kolch, W. (2011). The secret life of kinases: functions beyond catalysis. Cell Commun. Signal 9:23. doi: 10.1186/1478-811X-9-23
Román-Trufero, M., Kleijn, I. T., Blighe, K., Zhou, J., Saavedra-García, P., Gaffar, A., et al. (2025). An ISR-independent role of GCN2 prevents excessive ribosome biogenesis and mRNA translation. Life Sci. Alliance 8:e202403014. doi: 10.26508/lsa.202403014
Rothenburg, S., Deigendesch, N., Dey, M., Dever, T. E., and Tazi, L. (2008). Double-stranded RNA-activated protein kinase PKR of fishes and amphibians: varying the number of double-stranded RNA binding domains and lineage-specific duplications. BMC Biol. 6:12. doi: 10.1186/1741-7007-6-12
Roze, L. V., Beaudry, R. M., Keller, N. P., and Linz, J. E. (2004). Regulation of aflatoxin synthesis by FadA/cAMP/protein kinase a signaling in aspergillus parasiticus. Mycopathologia 158, 219–232. doi: 10.1023/b:myco.0000041841.71648.6e
Sabina, J., and Brown, V. (2009). Glucose sensing network in Candida albicans: a sweet spot for fungal morphogenesis. Eukaryot. Cell 8, 1314–1320. doi: 10.1128/EC.00138-09
Sacks, J. J., Ajello, L., and Crockett, L. K. (1986). An outbreak and review of cave-associated histoplasmosis capsulati. Med. Mycol. 24, 313–325. doi: 10.1080/02681218680000471
Saito, A., Ochiai, K., Kondo, S., Tsumagari, K., Murakami, T., Cavener, D. R., et al. (2011). Endoplasmic reticulum stress response mediated by the PERK-eIF2α-ATF4 pathway is involved in osteoblast differentiation induced by BMP2. J. Biol. Chem. 286, 4809–4818. doi: 10.1074/jbc.M110.152900
Saptarshi, N., Porter, L. F., and Paraoan, L. (2022). PERK/EIF2AK3 integrates endoplasmic reticulum stress-induced apoptosis, oxidative stress and autophagy responses in immortalised retinal pigment epithelial cells. Sci. Rep. 12:13324. doi: 10.1038/s41598-022-16909-6
Sattlegger, E., Barbosa, J. A. R. G., Moraes, M. C. S., Martins, R. M., Hinnebusch, A. G., and Castilho, B. A. (2011). Gcn1 and actin binding to Yih1: implications for activation of the eIF2 kinase GCN2. J. Biol. Chem. 286, 10341–10355. doi: 10.1074/jbc.M110.171587
Sattlegger, E., and Hinnebusch, A. G. (2005). Polyribosome binding by GCN1 is required for full activation of eukaryotic translation initiation factor 2α kinase GCN2 during amino acid starvation. J. Biol. Chem. 280, 16514–16521. doi: 10.1074/jbc.M414566200
Schumacher, J., Kokkelink, L., Huesmann, C., Jimenez-Teja, D., Collado, IG., Barakat, R., et al. (2008). The cAMP-dependent signaling pathway and its role in conidial germination, growth, and virulence of the gray Mold Botrytis cinerea. MPMI 21, 1443–1459. doi: 10.1094/MPMI-21-11-1443
Schuster, A., Tisch, D., Seidl-Seiboth, V., Kubicek, C. P., and Schmoll, M. (2012). Roles of protein kinase a and adenylate cyclase in light-modulated cellulase regulation in Trichoderma reesei. Appl. Environ. Microbiol. 78, 2168–2178. doi: 10.1128/AEM.06959-11
Scott, J. D. (1991). Cyclic nucleotide-dependent protein kinases. Pharmacol. Ther. 50, 123–145. doi: 10.1016/0163-7258(91)90075-W
Sestari, S. J., Brito, W. A., Neves, B. J., Soares, C. M. A., and Salem-Izacc, S. M. (2018). Inhibition of protein kinase a affects Paracoccidioides lutzii dimorphism. Int. J. Biol. Macromol. 113, 1214–1220. doi: 10.1016/J.IJBIOMAC.2018.03.023
Seyedmousavi, S., Netea, M. G., Mouton, J. W., Melchers, W. J. G., Verweij, P. E., and de Hoog, G. S. (2014). Black yeasts and their filamentous relatives: principles of pathogenesis and host defense. Clin. Microbiol. Rev. 27, 527–542. doi: 10.1128/CMR.00093-13
Shabb, J. B. (2001). Physiological substrates of cAMP-dependent protein kinase. Chem. Rev. 101, 2381–2412. doi: 10.1021/cr000236l
Silberstein, S., Cantore, M. L., Galvagno, M. A., and Passeron, S. (1990). Isolation and characterization of a dimeric cAMP-dependent protein kinase from the fungus Saccobolus platensis. Arch. Biochem. Biophys. 282, 132–140. doi: 10.1016/0003-9861(90)90096-H
Silverman, P. M. (1978). Cyclic AMP binding proteins and cyclic AMP-dependent protein kinase from Blastocladiella emersonii. J. Bacteriol. 135, 976–980. doi: 10.1128/jb.135.3.976-980.1978
Singh, M. K., Shin, Y., Ju, S., Han, S., Choe, W., Yoon, K. S., et al. (2024). Heat shock response and heat shock proteins: current understanding and future opportunities in human diseases. Int. J. Mol. Sci. 25:4209. doi: 10.3390/ijms25084209
Smirnova, O. V., Sinyakov, A., and Kasparov, E. V. (2025). Application of Wuling powder in the treatment of ulcerative colitis complicated by depression as a new therapy principle. World J Gastroenterol. 31:106538. doi: 10.3748/wjg.v31.i23.106538
Štaudohar, M., Bencina, M., van de Vondervoort, PJI, Panneman, H., Legisa, M., Visser, J., et al. (2002). Cyclic AMP-dependent protein kinase is involved in morphogenesis of Aspergillus niger. Microbiology (N Y) 148, 2635–2645. doi: 10.1099/00221287-148-8-2635
Stovall, A. K., Knowles, C. M., Kalem, M. C., and Panepinto, J. C. (2021). A conserved Gcn2-Gcn4 Axis links methionine utilization and the oxidative stress response in Cryptococcus neoformans. Front. Fungal Biol. 2, 640–678. doi: 10.3389/ffunb.2021.640678
Sun, S., Hoy, M. J., and Heitman, J. (2020). Fungal pathogens. Curr. Biol. 30, R1163–R1169. doi: 10.1016/j.cub.2020.07.032
Sun, Q., Xu, G., Li, X., Li, S., Jia, Z., Yan, M., et al. (2023). Functional study of cAMP-dependent protein kinase a in Penicillium oxalicum. J. Fungi 9:1203. doi: 10.3390/jof9121203
Tameire, F., Collette, N., Fujisawaa, S., Bieging-Rolett, K., Dudgeon, C., Stokes, M. E., et al. (2023). Activation of GCN2 by HC-7366 results in significant anti-tumor efficacy as monotherapy and overcomes resistance mechanisms when combined with venetoclax in AML. Blood 142:2943. doi: 10.1182/blood-2023-189115
Takai, Y., Sakai, K., Morishita, Y., Yamamura, H., and Nishizuka, Y. (1974). Functional similarity of yeast and mammalian adenosine 3′,5′-monophosphate-dependent protein kinases. Biochem. Biophys. Res. Commun. 59, 646–652. doi: 10.1016/S0006-291X(74)80028-8
Tanaka, S. N. A., and Fukul, S. (1977). Effect of biotin on the level of isocitrate lyase in Candida tropicalis. Agricultural and Biological Chemistry. 41, 281–285.
Tang, C. P., Clark, O., Ferrarone, J. R., Campos, C., Lalani, A. S., Chodera, J. D., et al. (2022). GCN2 kinase activation by ATP-competitive kinase inhibitors. Nat. Chem. Biol. 18, 207–215. doi: 10.1038/s41589-021-00947-8
Taylor, S. S., and Kornev, A. P. (2011). Protein kinases: evolution of dynamic regulatory proteins. Trends Biochem. Sci. 36, 65–77. doi: 10.1016/j.tibs.2010.09.006
Terenin, I. M., Dmitriev, S. E., Andreev, D. E., and Shatsky, I. N. (2008). Eukaryotic translation initiation machinery can operate in a bacterial-like mode without eIF2. Nat. Struct. Mol. Biol. 15, 836–841. doi: 10.1038/nsmb.1445
Teske, B. F., Wek, S. A., Bunpo, P., Cundiff, J. K., McClintick, J. N., Anthony, T. G., et al. (2011). The eIF2 kinase PERK and the integrated stress response facilitate activation of ATF6 during endoplasmic reticulum stress. Mol. Biol. Cell 22, 4390–4405. doi: 10.1091/mbc.E11-06-0510
Unbehaun, A., Borukhov, S. I., Hellen, C. U. T., and Pestova, T. V. (2004). Release of initiation factors from 48S complexes during ribosomal subunit joining and the link between establishment of codon-anticodon base-pairing and hydrolysis of eIF2-bound GTP. Genes Dev. 18, 3078–3093. doi: 10.1101/gad.1255704
Uno, I., and Ishikawa, T. (1976) Effect of cyclic AMP on glycogen phosphorylase in Coprinus macrophizus. Biochim Biophys Acta. 452:112–20. doi: 10.1016/0005-2744(76)90062-0
van Eck, N. J., and Waltman, L. (2010). Software survey: VOSviewer, a computer program for bibliometric mapping. Scientometrics 84, 523–538. doi: 10.1007/s11192-009-0146-3
Wang, Y., Wei, X., Bian, Z., Wei, J., and Xu, J.-R. (2020). Coregulation of dimorphism and symbiosis by cyclic AMP signaling in the lichenized fungus Umbilicaria muhlenbergii. Proc. Natl. Acad. Sci. USA 117, 23847–23858. doi: 10.1073/pnas.2005109117
Webster, J., and Weber, R. (2007). Introduction to Fungi. 3rd Edn. New York: Cambridge University Press.
Wek, R. C. (2018). Role of eIF2α kinases in translational control and adaptation to cellular stress. Cold Spring Harb. Perspect. Biol. 10. doi: 10.1101/cshperspect.a032870
Wek, R. C., Jackson, B. M., and Hinnebusch, A. G. (1989). Juxtaposition of domains homologous to protein kinases and histidyl-tRNA synthetases in GCN2 protein suggests a mechanism for coupling GCN4 expression to amino acid availability (translational control/aminoacyl-tRNA synthetase). Proc Natl Acad Sci USA. 86, 4579–83. doi: 10.1073/pnas.86.12.4579
Wek, S. A., Zhu, S., and Wek, R. C. (1995). The histidyl-tRNA synthetase-related sequence in the eIF-2 protein kinase GCN2 interacts with tRNA and is required for activation in response to starvation for different amino acids. mBio. 10:e02143–19. doi: 10.1128/mBio.02143-19
Wheeler, D. L., and Yarden, Y. (Eds.) (2015). Receptor tyrosine kinases: Family and subfamilies. Wisconsin, USA: Humana Press.
Wortham, N. C., and Proud, C. G. (2015). EIF2B: recent structural and functional insights into a key regulator of translation. Biochem. Soc. Trans. 43, 1234–1240. doi: 10.1042/BST20150164
Wrabl, J. O., and Grishin, N. V. (2001). Homology between O-linked GlcNAc transferases and proteins of the glycogen phosphorylase superfamily. J. Mol. Biol. 314, 365–374. doi: 10.1006/jmbi.2001.5151
Xiong, J., Wang, L., Lin, J., Ni, L., Zhang, R., Yang, S., et al. (2025). Quantifying second-messenger information transmission in bacteria. Nat. Phys. 21, 1009–1018. doi: 10.1038/s41567-025-02848-2
Yan, W., Frank, C. L., Korth, M. J., Sopher, B. L., Novoa, I., Ron, D., et al. (2002). Control of PERK eIF2 kinase activity by the endoplasmic reticulum stress-induced molecular chaperone P58 IPK. Proc Natl Acad Sci USA. 99:15920-5. doi: 10.1073/pnas.252341799
Yu, Y., Pierciey, F. J., Maguire, T. G., and Alwine, J. C. (2013). PKR-like endoplasmic reticulum kinase is necessary for Lipogenic activation during HCMV infection. PLoS Pathog. 9:e1003266. doi: 10.1371/journal.ppat.1003266
Yuan, J., Li, F., Cui, B., Gao, J., Yu, Z., and Lu, Z. (2022). Inhibition of GCN2 Alleviates Cardiomyopathy in Type 2 Diabetic Mice via Attenuating Lipotoxicity and Oxidative Stress. Antioxidants (Basel). 11:1379. doi: 10.3390/antiox11071379
Zhao, W., Panepinto, J. C., Fortwendel, J. R., Fox, L., Oliver, B. G., Askew, D. S., et al. (2006). Deletion of the regulatory subunit of protein kinase a in Aspergillus fumigatus alters morphology, sensitivity to oxidative damage, and virulence. Infect. Immun. 74, 4865–4874. doi: 10.1128/IAI.00565-06
Keywords: kinases, GCN2, eIF2α, translational regulation, pathogenesis, virulence, stress response, amino acid deprivation
Citation: La Santrer E, Assunção CB, de Camargo TMN, Rodrigues I, Campolina SS, de Aguiar EL, de Souza Rodrigues T and Caligiorne RB (2025) The protein kinases family in fungi: adaptability, virulence and conservation between species. Front. Microbiol. 16:1630196. doi: 10.3389/fmicb.2025.1630196
Edited by:
Axel Cloeckaert, Institut National de recherche pour l’agriculture, l’alimentation et l’environnement (INRAE), FranceReviewed by:
Bernardo Franco, University of Guanajuato, MexicoCsilla Szebenyi, University of Szeged, Hungary
Na Liu, Zhejiang University, China
Copyright © 2025 La Santrer, Assunção, de Camargo, Rodrigues, Campolina, de Aguiar, de Souza Rodrigues and Caligiorne. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rachel Basques Caligiorne, cmFjaGVsYmNAZmFjdWxkYWRlc2FudGFjYXNhYmguZWR1LmJy