 Elena Durán-González*
Elena Durán-González* Jorge Antolín Ramírez-Tejero
Jorge Antolín Ramírez-Tejero Marta Pérez-SánchezCarmen Morales-Torres
Marta Pérez-SánchezCarmen Morales-Torres Rosa Gómez-MoranoClaudia Díaz-LópezAntonio Martínez-LaraDavid Cotán
Rosa Gómez-MoranoClaudia Díaz-LópezAntonio Martínez-LaraDavid Cotán- Pronacera, Sevilla, Spain
Background: Fibromyalgia is a complex disorder whose main symptoms are chronic widespread pain and fatigue and affects between 0.2 and 6.6% of the world population. Nowadays, there are no molecular biomarkers that could facilitate diagnosis. The latest efforts by researchers have focused on studying problems at the level of central nervous system sensitivity, inflammation, and oxidative disorders.
Methods: A total of 892 women were initially enrolled in the study. For individuals who met the inclusion criteria, a plasma proteome analysis was conducted using blood samples. Briefly, blood was collected, centrifuged, and analyzed by liquid nano-chromatography coupled to tandem mass spectrometry. After the raw data analysis, proteins with statistically significant differential abundance and a fold change over 1.2 (20% increase in fibromyalgia compared with control samples) or under 0.8 (20% decrease in fibromyalgia compared with control samples) in fibromyalgia were selected. For fecal metagenome analysis, fecal samples were collected and processed for DNA extraction. Amplicon sequencing of V3–V4 regions from the 16S ribosomal RNA gene was performed using the Illumina MiSeq platform. The statistical analysis was conducted using R v4.3.2 base packages.
Results: After applying exclusion criteria, 242 women (199 patients and 43 age- and environmentally paired controls) provided plasma and feces samples, as well as properly filled health questionnaires. A total of 30 proteins and 19 taxa were differentially expressed in fibromyalgia patients, and their integration into an algorithm allows for discrimination between cases and controls. The multi-omic approach for biomarker discovery in this study proposes a multifactorial connection between gut microbiota and mitochondria-derived oxidative stress and inflammation.
Conclusions: Plasma and fecal multi-omics analysis suggest an intricate and multifactorial connection between gut microbiota and mitochondria-derived oxidative stress and inflammation in FM patients, with glyceraldehyde-3-phosphate dehydrogenase and Streptococcus salivarius as leading actors.
Trial registration: NCT05921409.
Background
Fibromyalgia (FM) is a complex disorder whose main symptoms are chronic widespread pain and fatigue. Frequently, these symptoms are accompanied by a significant variety of related physical and psychological disturbances (Longley, 2006; Clauw, 2014). Despite the lack of knowledge about the etiopathogenesis and the complex diagnosis of the disorder, the prevalence ranges between 0.2 and 6.6% of the world population (Marques et al., 2017), with women accounting for approximately 89% of patients. To date, the non-specificity of the symptoms, the huge increase in worldwide prevalence, and the economic impact that it entails have turned FM into a pressing health and social issue. This evidence, together with the absence of molecular biomarkers that could facilitate diagnosis, underlines the extreme necessity of basic research on this chronic disorder.
Several physical and psychological disturbances such as migraine, muscle cramps, intestinal discomfort, sleep alterations, lack of short- and long-term memory, inability to concentrate, depression, stress, and anxiety, are recurrently reported by FM patients (Valera-Calero et al., 2022). To unveil the source of these symptoms, the latest efforts by the researchers have focused on studying problems at the level of central nervous system (CNS) sensitivity (Giorgi et al., 2023), inflammatory and oxidative disorders (Coskun Benlidayi, 2019), and even imbalances related to the intestinal microbiota (Liu et al., 2023).
Increasing evidence indicates that inflammatory processes occurring in peripheral tissues and the CNS, primarily in the spinal cord and the brain, are responsible for the pathophysiology of FM (Bains et al., 2023; O'Mahony et al., 2021). The release of chemokines and cytokines leads to the activation of both the innate and adaptive immune systems, as well as neuroinflammation (Littlejohn and Guymer, 2018). These mechanisms are reflected in many of the peripheral clinical features reported by FM patients, such as swelling, dysesthesia, cognitive changes, and fatigue. This lack of energy that accompanies FM patients has been related to mitochondrial dysfunction and oxidative stress. In this sense, it has been shown that there is a correlation between oxidative processes and pain sensitization in FM patients. In this way, a deficiency in the function of enzymes such as superoxide dismutase 1 (SOD1), catalase (CAT), and nicotinamide adenine dinucleotide phosphate (NADPH) oxidase has been identified in FM patients (Assavarittirong et al., 2022), which correlates with the severity of pain and fatigue (Rus et al., 2021; Altindag and Celik, 2006). Likewise, certain studies have identified a lower total antioxidant capacity, accompanied by a greater presence of reactive oxygen species (ROS) in women with FM compared to the control group (Rus et al., 2021). As a whole, all these processes cause immunity deregulation and mitochondrial imbalances that, ultimately, can trigger peripheral neuroinflammatory processes (Coskun Benlidayi, 2019).
The gut-brain axis stands out as one of the recent alterations linked to FM. The main hypothesis indicates a connection between intestinal microbiota imbalance and the increase of intestinal permeability in patients, with the consequent alteration in digestion, immune response, and CNS signaling (Erdrich et al., 2020; Goebel et al., 2008). When the intestinal epithelial barrier is compromised, it can lead to increased intestinal permeability, allowing gram-negative bacteria to move through the mucosal lining and reach the blood flow. Their ability to produce neurotransmitters and their interaction with the primary function of the vagus nerve allows these bacteria to modulate the activity of the hypothalamic-pituitary-adrenal (HPA) axis (Tomasello et al., 2018). This situation allows direct interactions between intestinal microbiota components and humoral and cellular mediators from the immune system and the CNS, triggering an immune response characterized by increased production of inflammatory mediators (Moloney et al., 2016). Thus, several studies have observed changes in the composition of the gut microbiota in various disorders, including gastrointestinal, rheumatic, and metabolic conditions (Wang et al., 2022; Afzaal et al., 2022). In the FM context, a recent study has found a reduced bacterial diversity, as well as altered levels of glutamate and serine, suggesting changes in the metabolism of crucial neurotransmitters in the CNS signaling (Clos-Garcia et al., 2019). This innovative approach based on intestinal microbiota opens a new field for searching for biomarkers. Those biomarkers could also be modulated through diet therapy, which has been gaining relevance in FM management.
Advances in identifying inflammatory markers involved in FM have been largely made thanks to the development of omics techniques. Proteomics studies have been conducted in patients, primarily in saliva, serum, and cerebrospinal fluid. Therefore, (Ramírez-Tejero et al. 2018) identified 33 plasma proteins with different expression in FM patients compared with volunteers, with most proteins linked to inflammatory processes. Another study in patients revealed differences in plasma proteins involved in inflammatory, metabolic, and immunological processes and demonstrated a correlation between these proteins, patients' pain perception, and psychological distress (Wåhlén et al., 2020). Additionally, 22 differentially expressed proteins were identified in FM patients compared with pain-free volunteers, mainly associated with blood coagulation processes, immune response, and interactions with extracellular matrix receptors (Han et al., 2020). Among other findings, these results confirmed the promising utility of proteomics in the study of FM and demonstrate the roles of coagulation, inflammation, and the immune response in the disorder's pathogenesis.
In spite of the above-mentioned research, nowadays, the diagnosis and treatment of FM remain controversial. Traditionally, the diagnostic criteria of the American College of Rheumatology (ACR) have been followed (Wolfe et al., 1990, 2011, 2016), and most healthcare professionals treat their patients with numerous drugs focused on symptoms such as pain, depression, and fatigue. However, clinical recommendations from the European League Against Rheumatism (EULAR) and other organizations point to a multidisciplinary point of view of FM and its treatment (Giorgi et al., 2023; Macfarlane et al., 2017). Likewise, the latest trend is toward the use of highly individualized therapies combined with non-pharmacological approaches indeed (Kundakci et al., 2022).
In this context, this study is focused on biomarker discovery for FM in plasma and feces from 199 FM patients and 43 women as a control group, aiming to combine the analysis of both samples in a diagnostic panel for clinical practice.
Methods
Cohort recruitment
Study population
To calculate the size of the cohort, the most comprehensive data on the estimated prevalence of FM in Spain published in 2008 were considered (Mas et al., 2008). These authors concluded that approximately 2.4% of the Spanish population was suffering from FM. Specifically, this prevalence reached 8.4% in the age range of 40–49 years, whereas in women aged 50–59, it was 6.7%. A sample size calculation was performed according to the protocol described by (Naing et al. 2006), with a 95% confidence interval (CI) and an α level of 0.05. The result of this sample size calculation was n = 198 (188 plus a 5% of the estimated withdrawal), which means that the cohort had to comprise at least 198 patients with FM to obtain adequate statistical power.
Participants were recruited, and all samples were collected between March 2020 and September 2020. Both FM patients and volunteers were Caucasian. Before the beginning of the project, participants completed an informed consent form and a patients' information datasheet that have been approved by the Ethical Committee for Research with Medications (CEIM) at the Quirónsalud-Catalunya Hospital Group (protocol code IDI-20210749, approval record No. 01/2022).
Inclusion criteria
Individuals included in the study were selected through an online survey sent to FM associations, allowing for effective recruitment and reaching a larger pool of potential participants. All the personal and clinical data were recorded and processed as required by the Spanish law (Organic Law 3/2018, of December 5th) (Jefatura del Estado, 2018). A total of 892 questionnaires were received with clinical information from potential patients and control subjects, who were subsequently selected according to the following inclusion criteria for both groups:
- Women within an age range of 40–59 years, since these women are the most affected population group in Spain (Mas et al., 2008).
- Individuals with a BMI >18.5 and < 35.9.
As for exclusion criteria for both groups, the following conditions were established:
- Not being in the age or BMI range.
- Suffering from any type of cancer.
- Individuals who have undergone antibiotic treatment during the 30 days before the first sample extraction.
Additionally, for FM participants, a confirmed FM diagnosis by a healthcare professional, as defined by any ACR criteria, was required.
Although they were not considered exclusion criteria, clinical data on comorbidities and lifestyle habits were collected for analysis as potential confounding factors. Additional information was gathered about cardiovascular diseases (e.g., atherosclerosis, cardiomyopathy), autoimmune diseases (e.g., lupus, celiac disease, thyroiditis), or metabolic disorders (e.g., diabetes, metabolic syndrome). Besides, questions about pharmacological treatment with non-steroidal anti-inflammatory drugs, analgesics, antidepressants, antioxidants, lifestyle habits, diet, smoking, and alcohol consumption were also included in the questionnaires.
Clinical data collection
All clinical and personal data collected in this study were handled confidentially and in compliance with applicable data protection regulations. Technical and organizational measures were implemented to ensure the anonymization of samples and the dissociation of any identifying information. Access to clinical information was restricted to authorized personnel and stored on secure servers with access control.
The participants answered two clinical questionnaires related to their health status. All of them have completed the Spanish version of the 36-item Short Form Survey health questionnaire (SF-36) following the methodology described by (Vilagut et al. 2005). This questionnaire assesses the physical and psychological wellbeing of the participants. The 36 questions cover eight specific scales: physical functioning (PF), role limitations due to physical health (PH), pain (P), general health perceptions (GH), vitality (V), social functioning (SF), role limitations due to emotional problems (EP), and mental health (MH). The results were interpreted considering a higher score to be a better state of health.
Furthermore, only FM patients completed the revised FM Impact Questionnaire (FIQR) to assess the impact of FM on their daily lives. This questionnaire evaluates physical functioning, work performance (missed workdays and work difficulty), depression, anxiety, morning fatigue, pain, stiffness, fatigue, and overall wellbeing during the preceding week. Its 21 items are divided into three main domains: function (n = 9), overall impact (n = 2), and symptoms (n = 10). Each item is scored on a scale of zero (no impairment) to 10 (maximum impairment), and the total function score is divided by three. The sum of the scores for symptoms is divided by 2, and the overall impact score remains unchanged. The total score of the FIQR has been calculated by adding these three scores, with the maximum possible total score being 100. All the participants' recruitment and clinical data collection are detailed in our clinical data registration (NCT05921409) (Pronacera. Clinicaltrials, 2024) and protocol study already published by (Lucena Del Amo et al. 2023).
Antioxidant capacity measurement
The total antioxidant capacity of participants was determined by electrochemistry, as described in (Lucena Del Amo et al. 2023).
Plasma proteome analysis
Whole blood samples were collected in heparin-lithium tubes through venipuncture performed by a healthcare professional. To isolate the plasma fraction, the sample was transferred to a 15 ml Falcon tube and then centrifuged at 2,300 RCF for 10 min at 4 °C. Then, plasma was aliquoted and stored at −80 °C until batch analysis. Before mass spectrometry analysis, high-abundance proteins were depleted using Thermo Fisher Top 14 columns, following the manufacturer's instructions. Depleted samples were digested with trypsin and desalted before injection. To analyze the proteome, we performed liquid nano-chromatography coupled to tandem mass spectrometry (nLC-MS/MS) using an EVOSEP ONE system, which was coupled to the TIMS Tof Pro hybrid mass spectrometer. This analysis was carried out through the services provided by the BioGUNE Cooperative Research Center in Biosciences, located in the Basque Country, Spain. In brief, 250 ng of the sample were injected into the system. The 60 samples-per-day (SPD) protocol was used, with a 22-min chromatographic gradient from 5% to 50% acetonitrile [B], followed by a 2-min wash phase at 85% [B]. Acquisitions were performed on a TIMS TOF Pro system using the standard short-gradient method provided by Bruker in data-dependent acquisition (DDA) mode.
The processed data were analyzed with the search engines MASCOT (MatrixScience) and PEAKS (Bioinformatics Solutions Inc). Label-free quantification of differential proteomics was also performed by PEAKS (Bioinformatics Solutions Inc.) and MaxQuant (Cox Lab) software. The parameters were optimized considering: tolerances of 20 ppm and 0.05 Da for peptides and fragments, respectively; carbamidomethylation of cysteine considered as a fixed modification, and oxidation of methionines as a variable; Uniprot/Swissprot database limited to Homo sapiens as the organism for the searches; and selection of proteins identified with at least one unique peptide with False Discovery Rate (FDR) < 1%. Finally, raw abundances were normalized by Cyclic Loess using the NormalizerDE R package and imputed using the K-nearest neighbor algorithm matrix-wise. A differential abundance analysis was performed using the R package limma. Proteins with statistically significant differential abundance and a fold change (FC) over 1.2 (20% increase in FM compared with control samples) or under 0.8 (20% decrease in FM compared with control samples) in FM were analyzed using the Ingenuity Pathways Analysis software (IPA; Ingenuity® Systems, http://www.ingenuity.com). This bioinformatics application enables the functional analysis of differential expression data from different omics, integrating these data into molecular pathways to predict their activation state and link associated regulatory proteins.
Fecal metagenome analysis
Fecal samples were collected by participants using the DANASTOOL sample collection kit (Danagen; Barcelona, Spain) and the Fe-Col® Faecal sample collection paper (Alpha Laboratories; Hampshire, United Kingdom) as a stool-catch tool. The provided stool tube contains a DNA/RNA stabilization solution that allows samples to be stored at room temperature until batch analysis. For DNA extraction from fecal samples, the DANAGENE MICROBIOME FECAL DNA KIT (Danagen; Barcelona, Spain) was used. Briefly, 1 ml of the homogenized sample was transferred to a bead tube to ease sample disruption. Then, samples were incubated at 70 °C for 10 min. After the incubation step, thorough vortexing and centrifugation steps were applied. The upper phase was then transferred to a filtering column to extract DNA by centrifugation. DNA quality was assessed with a ThermoFisher Multiskan SkyHigh UV/Vis microplate spectrophotometer (Thermo Fisher Scientific, USA). Optimal concentrations were considered to be greater than 5 μg/μl, and 260/280 and 260/230 ratios were considered to be between 1.8 and 2.1. The resultant DNA extracts were stored at −20 °C until further analysis.
Amplicon sequencing was performed using the Illumina MiSeq platform at HelixBioS facilities (Madrid, Spain), where amplicons of V3–V4 regions from the 16S ribosomal RNA gene (16S rRNA) were sequenced. According to the manufacturer's instructions (Illumina, n.d.), tagging and barcoding processes suitable for Illumina technology were conducted. Pretreatment of the samples was performed using quantitative real-time PCR, followed by further individual quantification, pooling, washing, and titration steps. The sequencing was performed on fragments with sizes ranging from 2 × 250 to 2 × 300, in paired-end format, generating between 150,000 and 200,000 reads per sample. Quality control procedures were implemented using MultiQC software (Ewels et al., 2016), with filtering thresholds set at 50,000 reads per sample, Q30 Phred Score, and an average trimmed read length of 280 bp. After data pre-processing, sequencing results were analyzed with USEARCH V11.1. Briefly, the reads were cleaned up using the UCHIME algorithm to eliminate “singletons” (reads with only one sequence), chimeric sequences, and possible artifacts. Zero-radius Operational Taxonomic Unit (zOTU) clustering was performed using the UNOISE algorithm at a 99% identity (similarity). The sequences were then aligned against the taxonomic database specifically developed for the study of the intestinal microbiota by HelixBioS, HUMAN GUT 16S v3.2024 (HelixBioS), with an alignment cut-off point at 99% (identity) using the algorithm USE-LOCAL. The resulting taxonomic count tables were processed using the R packages DESeq2 (Love et al., 2014) (for differential abundance) and Vegan (Oksanen, 2025) (for diversity analysis). The microbiome functional profile was inferred using PICRUSt (v. 2.4.1), a bioinformatics software package designed to predict the functional content of a metagenome from identified bacteria. The zOTU table, filtered for less than 15% missingness and converted to biom format using Qiime2, underwent stratified analysis to determine zOTU contributions. Differential abundance was assessed using ggpicrust2 with DESeq2, and plotting functions were customized. Contribution data were processed with Tidyverse (R base package) to calculate the mean and standard error.
Statistical and bioinformatic analysis
The statistical analysis was conducted using R v4.3.2 base packages. Data were expressed as the sample mean ± standard deviation (SD). The null hypothesis (H0) was defined as no difference between the compared groups, with statistical significance set at 0.05 (α = 0.05). The Kolmogorov-Smirnov test was performed to determine the data distribution before each comparative analysis, thus checking whether the data followed a normal distribution. Depending on the result, either Student's t-test for unpaired samples or the non-parametric Mann–Whitney U-test was used to compare the difference between the means of groups. Pearson's chi-squared and Fisher's exact test were used for qualitative variables.
Spearman's correlation coefficients (rho, ρ) were calculated on FM and control samples separately using normalized protein and taxonomic relative abundances and clinical data. To assess differences in correlation between FM and control samples, Fisher z-tests were conducted, and p-values of less than 0.05 were considered statistically significant. Correction for multiple comparisons was implemented through Benjamini-Hochberg FDR.
To evaluate the diagnostic performance of selected omics features, we implemented a deep neural network using the keras package in R. Three datasets were analyzed: Statistically significant proteomic markers (p < 0.05), statistically significant metagenomic markers (p < 0.05) and a combined dataset integrating both. Features were scaled to the [0, 1] range, and microbial abundances were further binarized using a 0.01 threshold. The model applied was a fully connected multilayer perceptron (MLP) with four hidden layers, trained using the Adam optimizer (learning rate = 0.01) and binary cross-entropy loss. To control class imbalance, sample weights were calculated dynamically from each fold. Training was conducted over a maximum of 800 epochs, with early stopping (patience = 250, min_delta = 0.01) based on validation AUC. Model validation was performed using 10 repetitions of five-fold stratified cross-validation, ensuring balanced class distribution across folds.
AUC was used as the primary performance metric, calculated with the pROC package. Individual ROC curves were generated for each fold and repetition using the roc function. To compute the mean ROC curve, the sensitivity and specificity values across all 50 runs (five folds × 10 repeats) were interpolated. The final ROC plots include the mean curve ± 1 standard deviation. This methodology was applied to the three datasets independently (proteomics only, metagenomics only, and combined).
Results
Participants characteristics
A total of 892 potential participants were screened for this study. Of these, 642 participants were excluded during the screening phase following the inclusion/exclusion criteria. Seven participants who did not complete all the clinical questionnaires and one who did not deliver fecal samples were also excluded. The final cohort consisted of 242 women, including 199 who were FM patients and 43 who were age- and environmentally paired controls who did not present any symptom related to FM. The characteristics of the study cohort are shown in Table 1. There are no significant differences between groups for any clinical variable (p < 0.05), including BMI, physical activity, or tobacco consumption. Nevertheless, regarding comorbidities, a higher but not statistically significant incidence of cardiovascular and metabolic diseases, as well as higher consumption of antioxidants, was found in patients compared to the control group.
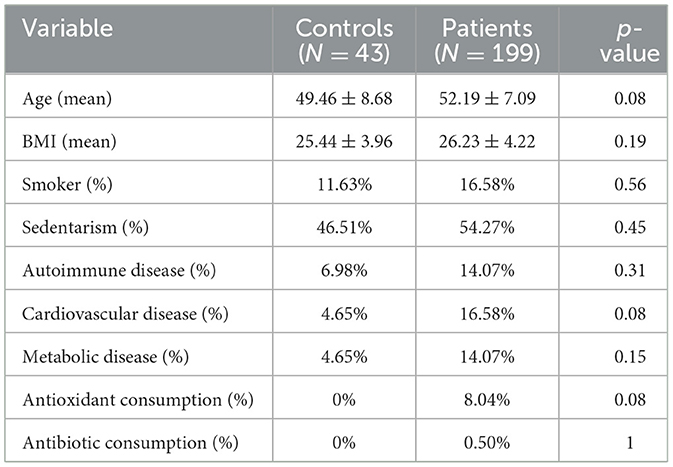
Table 1. Cohort demographic characteristics.
Depending on the variable of study Mann–Whitney U, Pearson's chi-squared, and Fisher's exact test were used, considering p-value < 0.05 as the threshold to establish a significant difference.
Data from questionnaire scores for each study group are shown in Table 2. FM impact was evaluated by FIQR, where patients obtained a 65.94 ± 17.78 score. Both groups (FM vs. C) completed the SF36 questionnaire, in which all the resulting scores on the eight items evaluated showed statistically significant differences (p-value < 0.0001) between groups. Particularly, the biggest differences were found in body pain and physical health of patients, whose scores are 77.21 ± 23.02 vs. 20.94 ± 17.77 and 82.56 ± 31.19 vs. 10.88 ± 24.69, respectively.
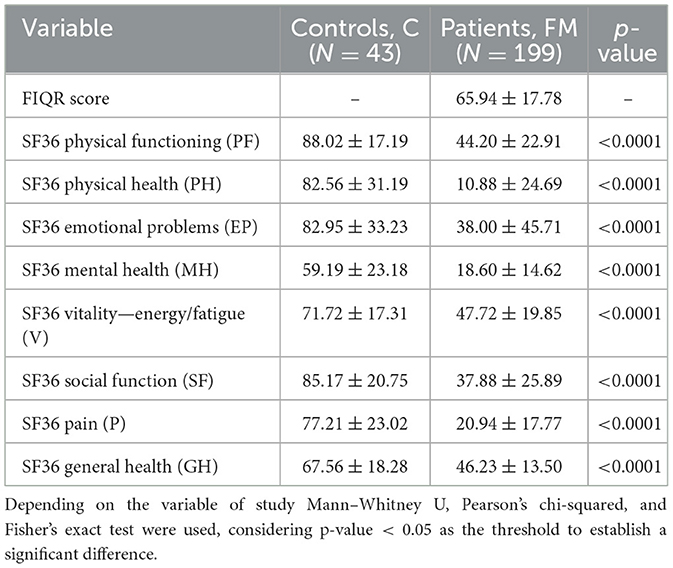
Table 2. Questionnaire scores.
Antioxidant capacity
The total antioxidant capacity of participants showed no statistically significant difference between groups (Supplementary Table S1).
Proteomic profile of plasma
Differentially expressed proteins
The proteomic profile of plasma samples from controls and patients with FM show more than 600 proteins identified. Of these, a total of 242 proteins were clearly detected in both groups. Using a cutoff p-value < 0.05 for statistical significance, a subgroup of 30 proteins were found to be differentially expressed. Finally, applying a cutoff value of 20% of FC, 14 were overexpressed (FM/C ratio ≥ 1.20) and 7 were underexpressed (FM/C ratio ≤ 0.83; Table 3). Technical validation of some of these proteins was performed by ELISA determination (Supplementary Table S1) as described in manufacturers' instructions.
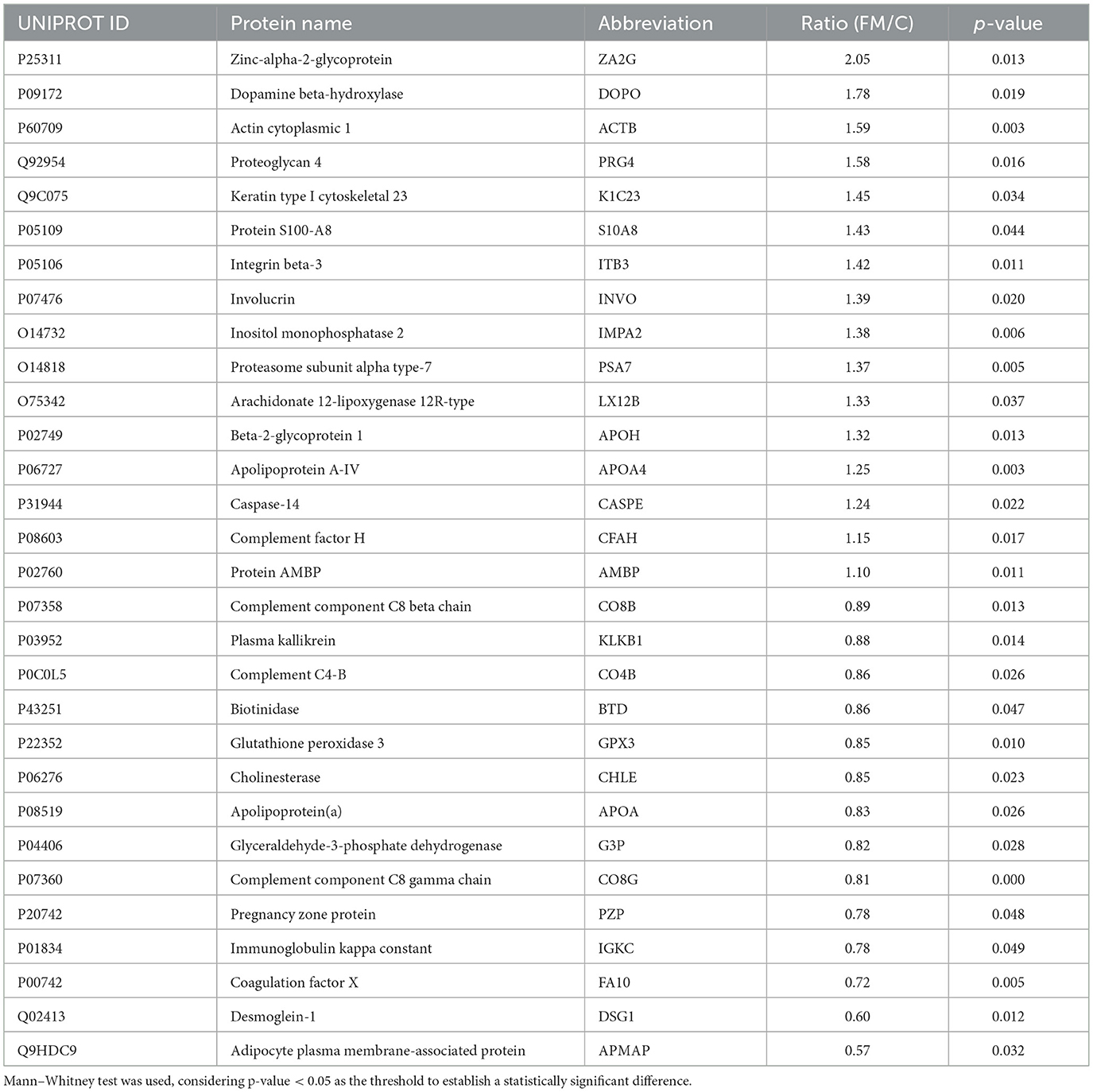
Table 3. Proteins are differentially expressed in the plasma of patients with FM.
Functional analysis of plasma proteome
IPA software was used to determine whether the proteins with different abundances were related to specific molecular pathways and networks. We analyzed the 30 deregulated plasma proteins found in FM patients, considering as reference set first, the Ingenuity Knowledge Database, and second, all the datasets obtained from plasma samples of this study, to get rid of sample-type bias. Figure 1a shows the most statistically significant enriched canonical pathways. Among these, the top 5 according to the p-value with Ingenuity Knowledge Data Base as reference set were Liver-X Receptor/Retinoid-X Receptor (LXR/RXR) activation (six proteins: AMBP, APOA4, APOH, C4A/C4B, LPA and S100A8), Complement System (four proteins: C4A/C4B, C8B, C8G, and CFH), Complement Cascade (five proteins: C4A/C4B, C8B, C8G, CFH, and IGKC), Clathrin-mediated Endocytosis Signaling (five proteins: ACTB, APOA4, ITGB3, LPA, and S100A8), and Atherosclerosis Signaling (four proteins: ALOX12B, APOA4, LPA, and S100A8). Using the study-identified proteins as a reference set, the top canonical pathways were Osteoarthritis Pathway (four proteins: CASP14, ITGB3, PRG4, and S100A8) and signaling by VEGF (2 proteins: ACTB and ITGB3).
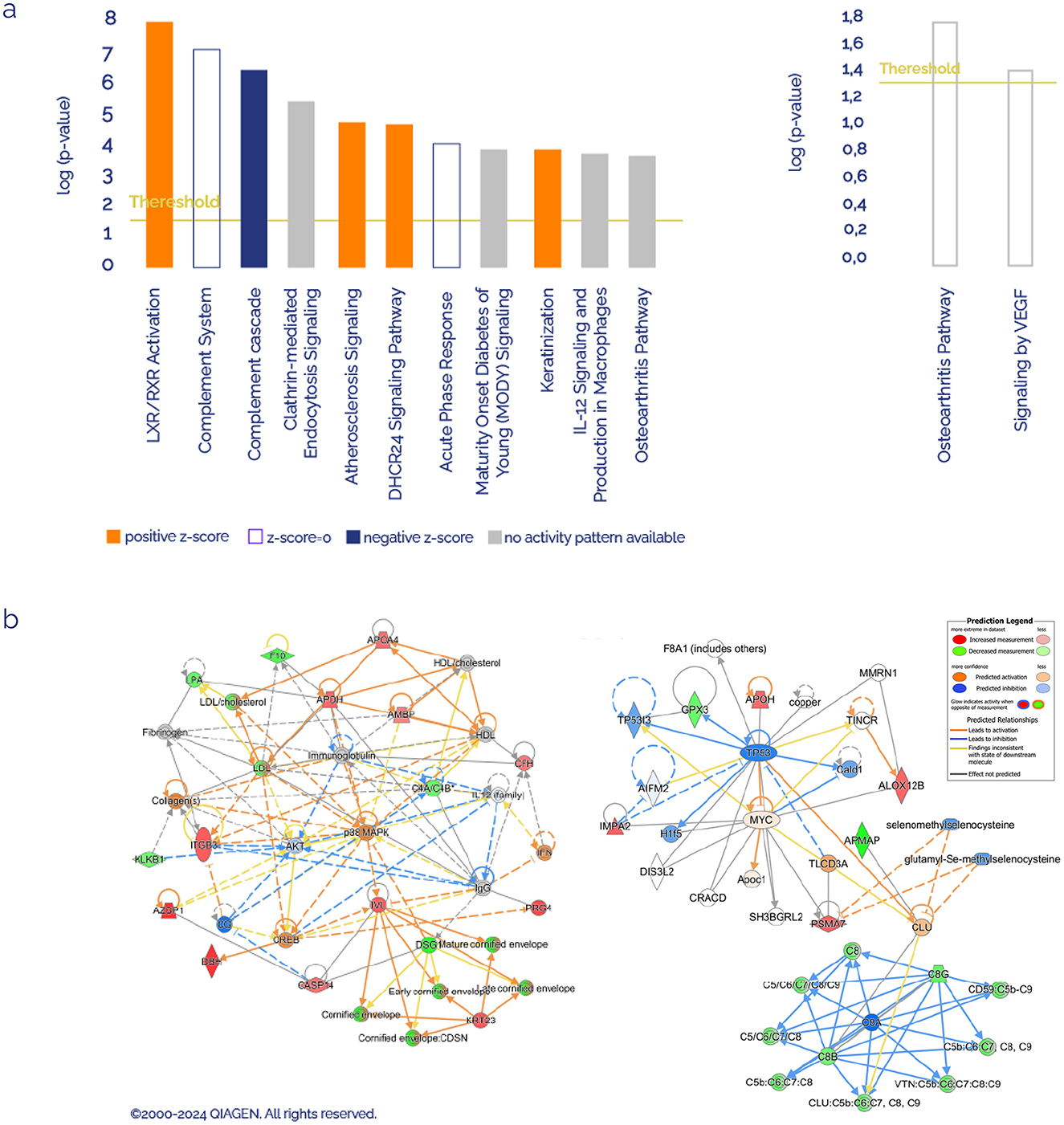
Figure 1. Ingenuity Pathways Analysis of differentially expressed proteins. (a) Canonical pathways enriched in FM sorted by significance value, using the Ingenuity Knowledge Database as reference on the left and using the identified list of proteins as reference set on the right. Orange or blue colors mean positive or negative Z-score, respectively. (b) Most significant molecular networks identified (red FC > 1.2, green FC < 0,8).
In Figure 1b, the top networks with the highest number of proteins involved were shown. On the left network, proteins associated with the inflammatory response, hematological system, and cell-to-cell signaling were gathered using the Ingenuity Knowledge Database as a reference set, where most of the proteins belong to the complement system. On the right network, using all the study-identified proteins as a reference set, most proteins and processes are associated with cancer, organismal development, and reproductive system development and function.
Metagenomic profile of feces
Differentially expressed taxa
A total of 6,030 zOTUs were identified through metagenomic profile analysis of feces samples, capturing 21,639,488 reads. All identified zOTUs were assigned to 696 different species, which were further grouped into 224 genera, 82 families, and 11 phyla.
After ubiquity and representativity filtering, the top five most predominant zOTUs families across all samples from both groups were Oscillospiraceae (28% of total raw counts), Lachnospiraceae (17%), Bacteroidaceae (14%), Clostridiaceae (6%), and Bifidobacteriaceae (5%), as indicated in Figure 2a.
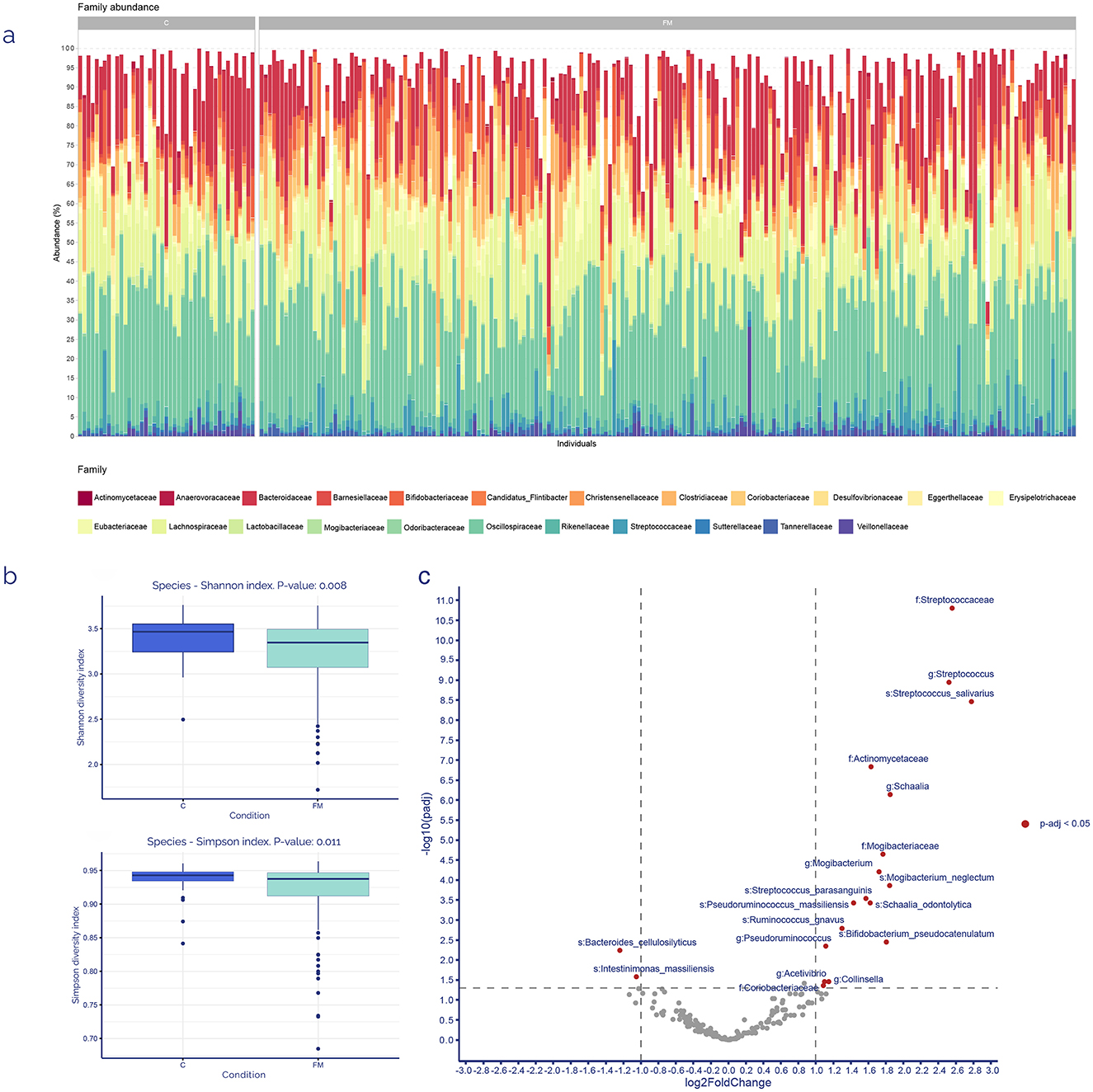
Figure 2. Core microbiome and discriminant analyses. (a) Family-level relative taxonomic composition of the gut microbiota among FM patients (n = 199) and controls (n = 43). (b) α-diversity indexes for each sample group at the species-level taxonomic classification, showing the p-value computed using the Mann–Whitney U-test. (c) Volcano plot of 194 taxonomic relatives' abundances detected in feces samples after ubiquity and representativity filtering. Positive log2FC indicates increased abundance, and negative log2FC indicates a reduction in FM patients. All p-values were adjusted using the Benjamini-Hochberg method.
Diversity was analyzed throughout 2 α-diversity scores (Shannon and Simpson), with a statistically significant decrease for both scores in FM patients (Figure 2b). Additionally, a differential zOTUs analysis (employing DESeq2) between the controls and FM core microbiomes was performed. The results showed 19 zOTUs whose abundance differed between both groups with an adjusted p-value of 0.05 (Figure 2c). Those zOTUs with higher FC in FM patients belonged mainly to Streptococcaceae, Actinomycetaceae and Mogibacteriaceae families. On the contrary, those with negative FC were just two species: Bacteroides cellulosilyticus and Intestinimonas massiliensis. Technical validation of some of these bacteria was performed by qPCR determination (Supplementary Table S1) as described in manufacturers' instructions.
Functional analysis of intestinal microbiota
PICRUSt analysis was applied to zOTUS abundances using the KEGG database to determine whether the taxonomic differences between intestinal microbiota from FM and controls might cause functional changes. The results showed that 17 KEGG pathways were significantly different between patients and controls (Figure 3a). The FM microbiota exhibited increased carbohydrate, ascorbate, pyruvate metabolism, bacterial infections, endocytosis, and aminobenzoate degradation, as well as folate, carotenoid, and flavonoid biosynthesis, type II diabetes mellitus, and glycolysis/gluconeogenesis. Conversely, the FM microbiota suggested reduced pathways were phenylpropanoid biosynthesis, PPAR signaling pathway, and cyanamide acid metabolism (p < 0.05). Enzyme activity prediction (Figure 3b) highlighted the top 5 activities according to their p-values: hydroxymethylglutaryl-CoA reductase, glyceraldehyde-3-phosphate dehydrogenase (NADP+), GMP reductase, glutathione reductase (NADPH), and 16S rRNA (guanine1207-N2)-methyltransferase, all of which showed statistically significant upregulation in the FM group. An exhaustive analysis of zOTUS contributing to the enrichment of these enzymatic activities identified Streptococcus salivarius as the primary cause of this alteration, participating in all the enzymatic pathways identified.
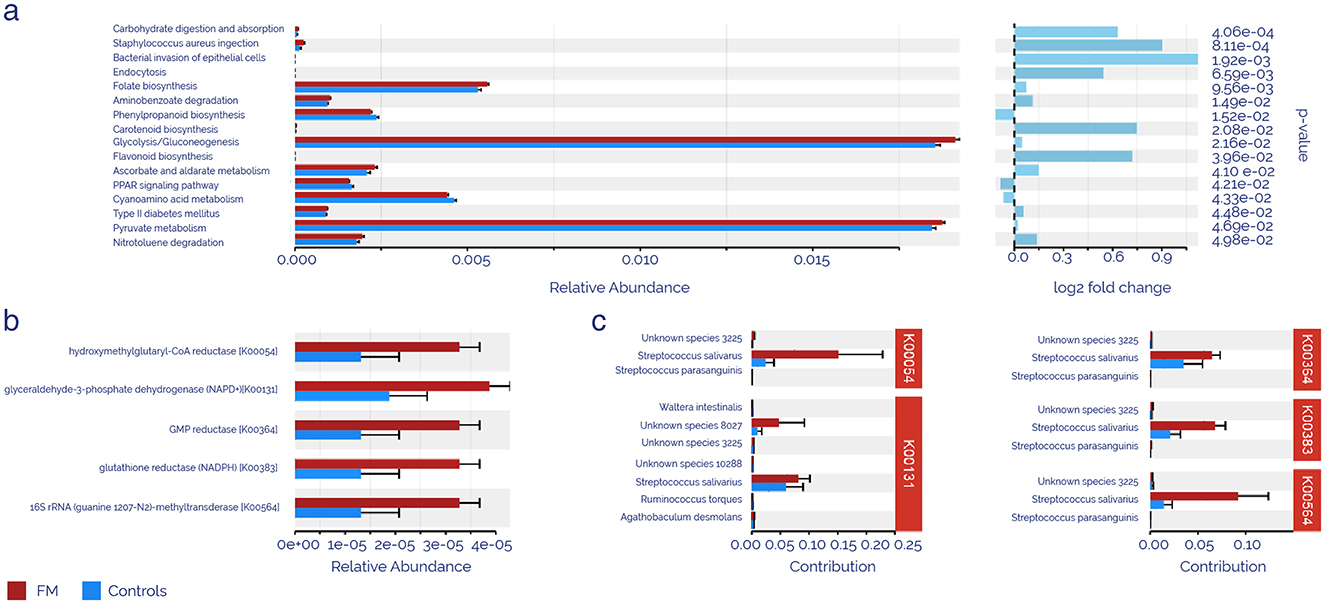
Figure 3. Predicted KEGG function analysis using PICRUSt. (a) Significantly enriched pathways according to the KEGGdatabase. (b) Top 5 significantly enriched enzyme activities. (c) zOTUS contributes to the enrichment of enzymatic activities. The red color represents the FM group, and the blue color represents the control group.
Multi-omics comparison
Correlation heatmaps
To explore the potential relationship between intestinal microbiota composition and the host plasma proteome profile, differences in ρ were studied between the FM and C group for each comparison. To depict the potential use of markers identified in both analyses as a diagnostic tool, zOTUs and proteins with statistically significant differences, independent of FC values, were correlated (Figure 4a). Several microbial taxa clustered due to their association with specific proteins. Interestingly, Bifidobacterium pseudocatenulatum and Bifidobacterium alomucense correlated with PSA7 (FM/C ratio: 1.37), LX12B (FM/C ratio: 1.33), and DOPO (FM/C ratio: 1.78), a solid result in phylogenetic terms, since the genus (Bifidobacterium) and the family (Bifidobacteriaceae) exhibited the same trend. This also occurred in other species like Mogibacterium neglectum, Collinsella aerofaciens or Schaalia odontolytica with their genera and families, respectively, consistently showing the relationship between them and the above-mentioned proteins. Moreover, certain proteins showed numerous correlations with taxonomic clusters. That was the case of PSA7 (FM/C ratio: 1.37), which had an inverse proportional correlation with zOTUs from the Bifidobacteriaceae family, and a positive one with zOTUs from the Coriobacteriaceae and Mogibacteriaceae families. So far, these two families have formed one of the most robust clusters due to their correlation with DSG1 (FM/C: ratio: 0.6), IGKC (FM/C ratio: 0.78), and PSA7 (FM/C ratio: 1.37) simultaneously. Another protein that correlated negatively with several zOTUs was BTD (FM/C ratio: 0.86), which showed similar negative correlations with the genera Agathobacter, Eghertella, and Christensenella, in addition to the species Bacteroides cellulosilyticus and Phocaeiocola massiliensis.
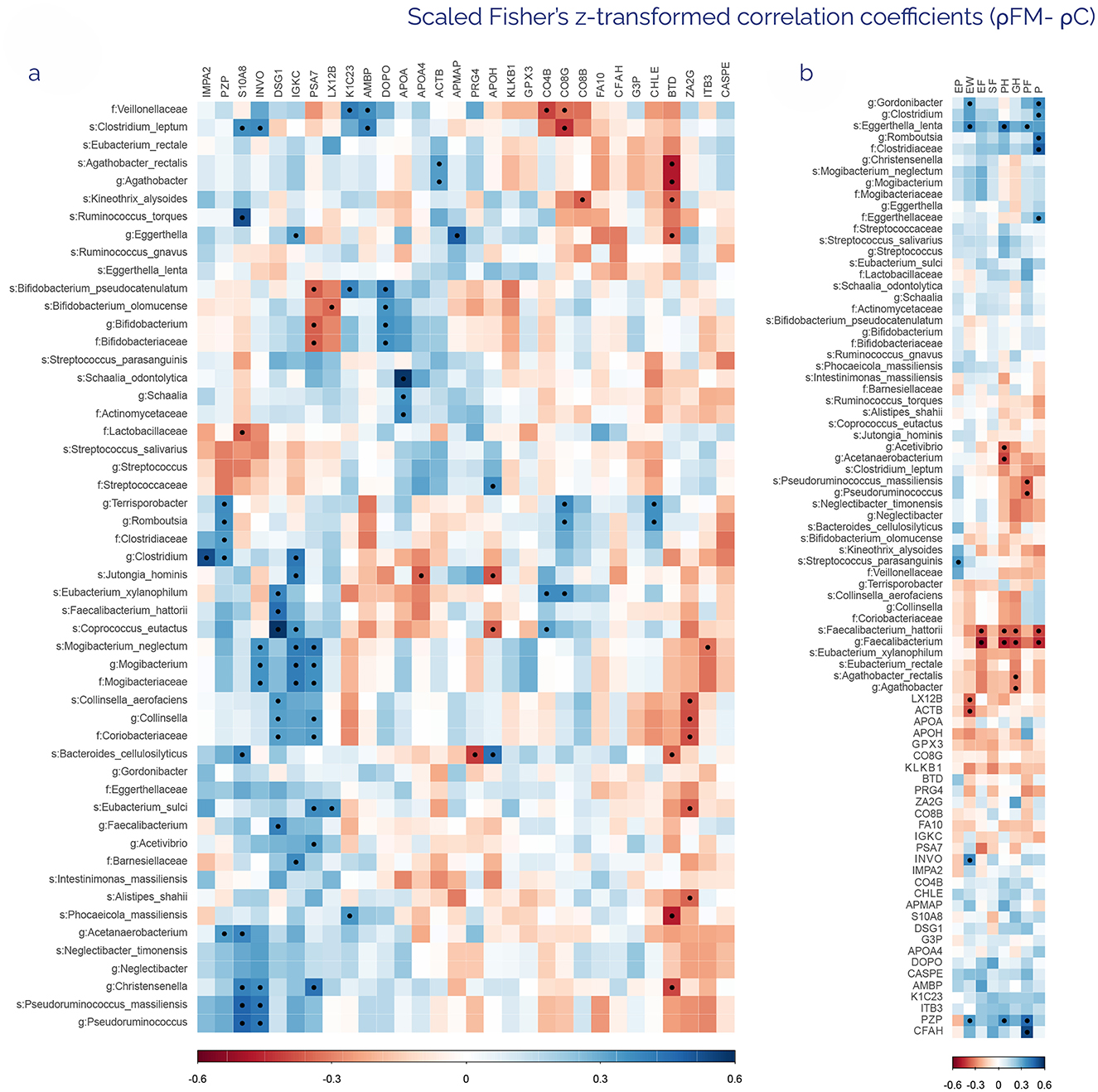
Figure 4. Heatmaps of scaled Fisher's z-transformed correlation coefficients (ρ) from the comparison of FM-related ρ vs. C-related ρ in different variables. The resulting and plotted ρ come from deducting FM-related ρ for a specific variable comparison to the C-related ρ for this same variable (ρFM- ρC). (a) Heatmap of Fisher's z-transformed ρ from ρFM-ρC between metagenomic zOTUs and proteomic markers with differentiated abundances. (b) Heatmap of Fisher's z-transformed ρ from ρFM-ρC between differentiated multi-omics results and SF36 clinical indices: emotional problems (EP), emotional wellbeing (EW), vitality-energy/fatigue (EF), social functioning (SF), physical health (PH), general health (GH), physical functioning (PF), and Pain (P). The heat map is sorted based on hierarchical clustering. Blue indicates positive correlations, whereas red indicates negative correlations (−0.6 < ρ > 0.6), with color saturation directly linked to ρ's absolute value. Those correlations with a statistically significant difference are marked by a black dot (Benjamini–Hochberg FDR < 0.05).
To determine which differences could be specifically associated with the quality of life of the participants, the correlations between the SF36 health questionnaire items and the omics data were represented (Figure 4b). Metagenomics showed the highest number of variable correlations with health scores, obtaining 15 zOTUs. A cluster composed mainly of zOTUs of the genus Faecalibacterium correlated negatively with most questionnaire items, including vitality, physical health, general health, and pain. In this sense, pain was the variable that showed a higher number of positive and negative correlations with proposed fecal markers. Although proteins had only 5 correlations, most of them were linked to the mental health score, a crucial factor in FM disorder.
Regarding the FIQR questionnaire, ρ for the omics results of FM patients and FIQR scores was calculated. The results were not significant for any of the variables, although a weak correlation (|ρ| > 0.2) suggests that depression was the item that correlated with a greater number of zOTUs, with a total of 9. Furthermore, pain showed a direct proportional correlation with all zOTUs of the Mogibacteriaceae family. As for proteins, only CASPE (FM/C ratio: 1.24) correlated negatively with stiffness, which correlates positively with the Streptococcaceae family at the same time (data not shown).
ROC curves
The diagnostic performance of the metagenomic and proteomic datasets was evaluated using supervised machine learning on 242 samples (199 FM patients and 43 controls). Models trained on each omics layer separately achieved good classification performance, indicating that both microbial and proteomic profiles contain relevant, non-redundant information for distinguishing FM patients from controls. However, the best results were obtained when both datasets were integrated into a single model, which achieved a mean AUC of 0.85 ± 0.07 across repeated stratified cross-validation (Figure 5). This integrative model, which leveraged all statistically significant microbial taxa and proteins, consistently outperformed the single-omic approaches. These findings highlight the complementary nature of the two omics layers and support the use of a combined multi-omics panel as the most effective strategy for discriminating FM patients from controls.
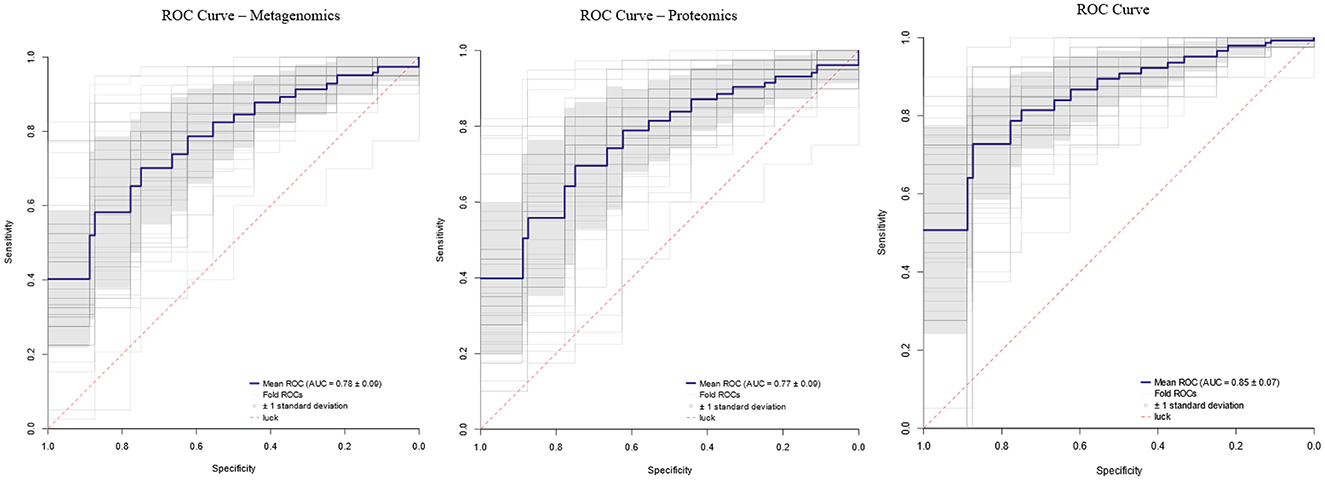
Figure 5. ROC curves. Area under the curve (AUC) values for each omics dataset and their combination.
Discussion
FM is a chronic pain-related disorder that is still underdiagnosed due to the lack of molecular markers. The present study is focused on the search for these markers in the plasma proteome and intestinal microbiota. The results showed a significant differential pattern of 30 proteins and 19 bacterial taxa between FM patients and controls, and a significantly higher α diversity in controls compared to patients, suggesting a worse intestinal environment and, consequently, a weaker intestinal and general health in FM.
Main symptoms of FM (pain and cognitive impairment) suggest an evident and deleterious nervous system disruption. In fact, several authors have agreed on classifying this disorder as one of the most common conditions among the so-called central sensitization syndromes (Boomershine, 2015), which are characterized by a CNS imbalance that affects the homeostasis of the whole nervous system. This altered neurological performance has been demonstrated in a substantial number of studies utilizing various imaging, electrodermal, and molecular techniques. For instance, different groups have found that nerve fibers participating in nociception in FM patients are less abundant, thinner, and even abnormally hyperexcited when compared with controls (Serra et al., 2014; Vecchio et al., 2020). Similarly, autonomic nervous system-dependent processes such as sweating have been reported as compromised by pathological traits in nerve fibers identified through electrodermal measurements (Reyes del Paso and de la Coba, 2020; Falco et al., 2024), adding further evidence to support the hypothesis of a complete nervous system failure in these patients.
At the molecular level, the long list of neurological imbalances is usually accompanied by chaotic neuroimmunological signaling. An extensive and varied record of altered molecular signaling in the immune system has been reported (Ovejero et al., 2020; Verma et al., 2022; Maes et al., 2024; Goebel et al., 2021), resulting in marked neuroinflammation, a common finding in FM patients (Littlejohn and Guymer, 2018; Mueller et al., 2023). In line with this hypothesis, nervous system impairment was also inferred by proteomic analysis in our cohort, with CHLE (FM/C ratio: 0.85), DOPO (FM/C ratio: 0.85) and G3P (FM/C ratio: 0.82) as affected proteins among others (Table 3); the first and the second, participate actively in the regulation of cholinergic and dopaminergic synapses (Darvesh et al., 2003; Silman, 2021; Rush and Geffen, 1980), and the third one, is annotated in the GO term neuron apoptosis process (GO:0051402). In fact, the role of G3P protein (GAPDH) seems to be essential in the gut-brain-mitochondrial axis, as discussed in the upcoming paragraphs. Inflammation traits were as well identified in our study, since some proteins among the 30 differentially expressed in the FM patients were narrowly related with pro-inflammatory pathways (Figure 1b), such as acute phase signaling, with AMBP (FM/C ratio: 1.10), APOH (FM/C ratio: 1.32), CO4B (FM/C ratio: 0.86), and KLKB1 (FM/C ratio: 0.88) as altered proteins; and IL-12 signaling and production in macrophages, represented by APOA4 (FM/C ratio: 0.83), IGKC, (FM/C ratio: 0.78), APOA (FM/C ratio: 0.83), and S100A8 (FM/C ratio: 1.43) as Table 3 showed. The last one, a component of calprotectin, has recently been proposed as a biomarker for rheumatoid arthritis and FM (Inciarte-Mundo et al., 2022; Vega-Ramírez et al., 2024), given its implication in the activation of innate immunity and inflammation.
In FM patients, the presence of neuroinflammation has been proposed as a multifactorial process driven by the co-occurrence of low vitamin D levels, mitochondrial dysfunction, and intestinal microbiota dysbiosis (Vasquez, 2016). In this sense, our proteomic results identified four proteins participating in the 24-dehydrocholesterol reductase (DHCR24) signaling pathway, according to IPA: AMBP (FM/C ratio: 1.10), APOA4 (FM/C ratio: 1.25), APOH (FM/C ratio: 1.32), and CO4B (FM/C ratio: 0.86). DHCR24 is the final enzyme in the biosynthesis pathway of cholesterol, a ubiquitous molecule that serves as a precursor to several by-products participating in a wide variety of physiological and pathological pathways, such as the metabolism of lipids and vitamin D, the synthesis of steroid hormones, and liver X-receptor (LXR)/retinoid X-receptor (RXR) activation, among others (Warren et al., 2021). LXR/RXR activation appears to be supported by stronger evidence in previous studies from our group and other authors. In the present study, the LXR/RXR activation pathway is the most differential canonical pathway between FM and C groups according to the p-value (Figure 1a), and is represented by the differential expression of AMBP (FM/C ratio: 1.10), APOA4 (FM/C ratio: 1.25), APOH (FM/C ratio: 1.32), CO4B (FM/C ratio: 0.86), APOA (FM/C ratio: 0.83), and S100A8 (FM/C ratio: 1.43), with a clear tendency toward upregulation. LXR/RXR receptors are steroid- and lipidic metabolites-dependent transcription factors involved in biological processes as diverse as lipid metabolism, immune system maintenance, neurobiological homeostasis or reproductive health homeostasis. Not in vain, this route has been significantly enriched and activated in numerous cohorts of FM patients in previous research (Ramírez-Tejero et al., 2018; Han et al., 2020; Frigo et al., 2021; Mohapatra et al., 2024), leading to altered lipid metabolism. Since steroid synthesis takes place inside the mitochondria, starting from cholesterol, the complete disruption found in lipid and steroid metabolism of FM patients that, at the end, alters the performance of the neuroendocrine system might be mainly triggered by this organelle.
Not surprisingly, mitochondria may be the common link among the various symptoms experienced by FM patients. By far, the nervous and musculoskeletal systems have the highest energy demand in the body, and they have been found to be dramatically affected by mitochondrial dysfunction (Cantó-Santos et al., 2020). Similarly, mitochondria have been repeatedly mentioned as a dysfunctional organelle in FM, not only by the scientific community, but also in previous studies from our research group (Cordero et al., 2013; Martínez-Lara et al., 2020; Castro-Marrero et al., 2013; Sánchez-Domínguez et al., 2015; Gerdle et al., 2020; Alcocer-Gómez et al., 2015; Cordero et al., 2010, 2016), which makes perfect sense, considering that pain, fatigue, joint stiffness, muscle cramps, and neurocognitive disorders stand out as main features of FM. Specifically, our previous findings suggested a profound imbalance of the mitochondrial membrane-associated porin voltage-dependent anion channel 1 (VDAC1) in FM patients (Martínez-Lara et al., 2020; Ramírez-Tejero et al., 2023), which directly cooperates with G3P in a well-defined classical mitochondrial-related apoptosis pathway (Tarze et al., 2007). Additionally, VDAC1 was identified in the plasma proteome analysis of our cohort, although it did not show statistically significant differences between groups (data not shown). This result may be due to technical limitations of the nLC-MS/MS approach, as even though a depletion step was introduced, the most abundant proteins could mask subtle changes in the less present ones. However, G3P was differentially expressed in FM patients, as shown in Table 3. This specific enzymatic activity appeared as the top functionally decreased activity in the intestinal microbiome of FM patients (Figure 3b) and is involved in the main pathways altered in FM patients, as indicated by our data. For this reason, the G3P enzyme and its by-products emerge as a promising diagnostic and therapeutic target for FM patients, and its enzymatic increase inferred from the metagenomic functional analysis could be a counteracting response to the lower plasma levels in FM patients.
Sticking to the mitochondrial environment, several other molecular functions take place inside this organelle, apart from energy metabolism. Specifically, mitochondria have been linked to oxidative stress and subsequent inflammatory processes in a wide range of pathological conditions (Chen et al., 2023). Among the differentially expressed proteins in FM patients participating in this study, some proteins related to these processes were found. For instance, GPX3 (FM/C ratio: 0.85), LX12B (FM/C ratio: 1.33), and APOA4 (FM/C ratio: 1.25) differences shown in Table 3, suggested a deficiency in oxidative stress defense as well as an increased lipid peroxidation, results already reported in FM and even linked to pain intensity and inflammation in those patients (Assavarittirong et al., 2022; Meeus et al., 2013; Akkuş et al., 2009; Ozgocmen et al., 2006). In line with these previous findings, some of the top-rated canonical pathways according to the p-value were complement cascade and complement system, implying proteins such as CO4B (FM/C ratio: 0.86), CO8B (FM/C ratio: 0.89), CO8G (FM/C ratio: 0.81), CFAH (FM/C ratio: 1.15), and IGKC (FM/C ratio: 0.78). This molecular pathway is narrowly related to chronic inflammation (Rawish et al., 2021).
All the previously identified imbalances found in our study could have a common link in the intestinal microbiome, defined as the unique signature of microbes, their genes, and metabolites, as well as their interaction with the host. Intestinal dysbiosis has been identified as a main source of CNS-related syndromes and symptoms, often via the vagus nerve and immune system (Rutsch et al., 2020). This connection has already been evaluated in FM, finding a low vagal nerve tone that might be related to pain and inflammation, as reported also for other conditions (Martins et al., 2021). One of the latest hypotheses links FM with this exact connection, dysbiosis being the main triggering cause of CNS symptoms. In FM patients, those deviations have lately been blamed for associated complications, such as irritable bowel syndrome (Garofalo et al., 2023) or psychological distress (Nhu et al., 2024). Besides that, a fast-growing hypothesis for some FM-overlapping sex-related conditions, such as endometriosis, breast and endometrial cancer, points to the role of β-glucuronidase activity from intestinal microbiota metagenome (estrobolome), specifically from the phyla Bacteroidetes (Bacteroidota) and Firmicutes (Bacillota) (Pollet et al., 2017), as the primary source of estrogens disruptions (Kwa et al., 2016; Baker et al., 2017; Ervin et al., 2019; Komorowski and Pezo, 2020; Salliss et al., 2022; Siddiqui et al., 2022; Sobstyl et al., 2022; Pai et al., 2023; Hu et al., 2023). This hypothesis reinforces the crucial role of the intestinal microbiota in a considerable list of human diseases and syndromes. In terms of hormone signaling, and following the estrobolome hypothesis, cortisol, estradiol, and progesterone levels have been investigated in FM patients and in vitro models; however, the findings on the impact of these hormones on pain and neuropsychological complications were preliminary and contradictory (Okifuji and Turk, 2006; Rus et al., 2016; Schertzinger et al., 2018; Hernandez-Leon et al., 2018; Lin et al., 2021).
Our results revealed a differential pattern in intestinal microbiota composition in FM patients, as shown in the volcano plot resulting from the metagenomic analysis (Figure 2c), with a total of 19 bacterial taxa that were differentially expressed. Specifically, at the phylum level, the distribution of zOTUS is as follows: 12 Bacillota, six Actinobacteria, and one Bacteroidota. Streptococcus salivarius appeared as the most differential bacterium between the FM and C group, with the lowest p-value and the highest FC. Interestingly, this bacterium has demonstrated regulatory properties for immunological and inflammatory responses in vitro (Kaci et al., 2014), suggesting that this marked difference may represent an outstanding research hypothesis for further studies. Several other species, such as Mogibacterium neglectum, Streptococcus parasanguinis, Schaalia odontolytica, Pseudoruminococcus massiliensis, Ruminococcus gnavus, and Bifidobacterium pseudocatenulatum showed the same trend as S. salivarius. On the other hand, Bacteroides cellulositycus and Intestinimonas massiliensis were found to be less abundant in FM patients, according to our results. To our knowledge, this is the first paper to report differential abundance in FM patients for bacteria from these genera, except for Bifidobacterium and Streptococcus. The first one was previously identified as a reduced genus in FM patients (Clos-Garcia et al., 2019), and the second one showed differences between men and women with FM in a pilot, descriptive study of our group (Ramírez-Tejero et al., 2023). In this regard, different research groups have demonstrated differentially abundant bacterial groups in the gut of FM patients compared with controls. (Minerbi et al. 2019) reported a distinct microbiota signature in FM patients, composed of a high prevalence of bacteria from the Firmicutes phylum. Besides, species such as Prevotella copri, Faecalibacterium Praustnitzii, and Bacteroides uniformis, classically associated with a healthy gut, showed less abundance in those patients. In a parallel study, (Clos-Garcia et al. 2019) found a similar pattern, where beneficial bacteria, such as Bifidobacterium spp. and Bacteroides spp., were clearly less abundant in patients compared to controls, as mentioned before. Moreover, this group demonstrated some correlation between these differences and the concentration of several metabolites with a crucial role in CNS signaling. Our results showed a striking coincidence, since Bacteroides cellulosyticus appeared with the same exact tendency as that found in both studies for their genus relatives. Several species from this genus have exerted a remarkable ability to participate in γ-aminobutyric acid (GABA) synthesis in the human gut (Otaru et al., 2021). GABA is the primary calming CNS neurotransmitter, and previous authors have demonstrated that it is present in the brain at a lower concentration in FM patients compared to controls, while its precursor and main excitatory neurotransmitter, L-glutamate, exhibits the opposite behavior (Minerbi et al., 2019; Otaru et al., 2021). In this sense, the generalized Bacteroides spp. deficiency that seems to be present in FM patients might be a promising therapeutic target for CNS imbalances in them, although further research is needed.
Functional prediction with PICRUSt of bacterial differences in our FM cohort provided promising results. Several pathways with a marked link to mitochondria were enriched in those patients. As shown in Figure 3a, glycolysis/gluconeogenesis and pyruvate metabolism were, by far, the most affected routes. Additionally, some vital routes for mitochondrial health were also compromised. For instance, the PPAR signaling pathway and folate biosynthesis (Xiu and Field, 2020; Mello et al., 2016). In line with this hypothesis, typically mitochondrial enzymatic activities also showed a differential behavior in ontology prediction based on intestinal dysbiosis found in these patients. That was the case of hydroxymethylglutaryl-CoA reductase (K00054) and glyceraldehyde-3-phosphate dehydrogenase (NADP+) (K00131). Mitochondrial dysfunction could be a triggering factor, according to further PICRUSt predictions, since antioxidant activities, such as glutathione reductase (NADPH) (K00383), also appear to be compromised. Taking into account that plasma results were suggesting a mitochondrial blockade, microbiota might be reacting as a counterbalance to this deficiency.
The multi-omics approach provided valuable insights into the relationships between proteins, bacteria, and health questionnaire results. In this sense, as shown in Figure 4, Fisher's z-transformed ρ highlighted evident differences in protein-to-bacteria and protein/bacteria-to-health questionnaire scores between controls and patients. Thus, strong positive differences in correlations were found between DSG1 and IGKC proteins and several bacterial genera, including Clostridium, Eubacterium, Faecalibacterium, Coprococcus, Collinsella, and Mogibacterium, all of which belong to the phylum Firmicutes. Desmogleins (DSG1 and DSG2) are essential proteins in cell-to-cell adhesion, serving as the primary components of desmosomes in tissues subjected to strong mechanical stress, such as the intestinal barrier layer (Gross et al., 2018; Sherrill et al., 2014). IGKC, as the constant region of the light chains from all immunoglobulin types, participates in a wide range of adaptive immune responses. Combined, these findings suggest that, in FM patients, the highest abundance of these genera may trigger an intestinal barrier breakdown, followed by an immunological response, whereas controls do not experience such a disturbance. Since FM has been classically linked to several gastrointestinal challenges (Slim et al., 2015), the intestinal microbiome composition of these patients should be a clinical indicator in their initial evaluations, as well as a therapeutic target to address. Furthermore, DOPO protein exerted a marked positive correlation with the Bifidobacteriaceae family, specifically with Bifidobacterium pseoudocatenulatum and Bifidobacterium alomucense. According to our results, the increase in these bacteria is linked to a higher abundance of Dopamine beta-hydroxylase, which is responsible for converting dopamine (a pleasure neurotransmitter) to noradrenaline (a concentration neurotransmitter). The link between these bacteria and dopaminergic signaling has been widely depicted in the scientific literature (Hamamah et al., 2022). For instance, in murine models, the supplementation with Bifidobacterium infantis elevated the levels of noradrenaline after maternal separation (Desbonnet et al., 2010). Whether these differential effects of Bifidobacterium spp. on dopamine/noradrenaline signaling could be acting as a gut-brain axis regulator checkpoint in FM symptoms needs to be addressed in further studies. An additional relevant finding concerns Bacteroides cellulosyliticus and the marked and opposite relationship established with the proteins PRG4 and APOH, both of which have higher levels in FM patients (Table 3). According to the heatmap of correlation differences, the presence of B. cellulosyliticus is somewhat related to PRG4. This proteoglycan plays a significant role in joint lubrication, but is also implicated in the management of neuroinflammation by interfering with TLR2/4 signaling (Bennett et al., 2021). In parallel, this bacterium showed a positive difference in correlations with APOH, which was previously described as a biomarker for antiphospholipid syndrome (Sibilia, 2003), a condition that worsens FM symptoms when it appears as a comorbidity (Costa et al., 2011). Given that, as previously mentioned, Bacteroides spp. was found to be a reduced genus in FM patients in both previous studies and our results, its differential correlation with these two proteins could be proposed as a molecular mechanism affected in patients.
Finally, regarding health questionnaires, the highest differences between FM and C correlation coefficients are shown by CFAH and Faecalibacterium hattori. The former showed positive differences with physical function, while the latter showed negative differences with energy/fatigue, physical health, general health, and pain, as shown in Table 2. Hence, CFAH and Faecalibacterium hattori could be potential therapeutic targets to address, thereby enhancing patients' quality of life.
Conclusions
This is the first multi-omics approach for FM patients, highlighting the feasible role of the gut-brain-mitochondrial axis in FM etiopathogenesis in such a large cohort. This study identifies several promising markers for classifying patients with FM. Plasma and fecal multi-omics analysis and its integration into an algorithm allows discrimination between cases and controls with high AUC values, pointing to an intricate and multifactorial connection between gut microbiota and mitochondria-derived oxidative stress and inflammation, with glyceraldehyde-3-phosphate dehydrogenase and Streptococcus salivarius as leading actors. All biomarkers identified in this study could play an important role in the disease, suggesting their potential as prognostic, diagnostic, and therapeutic targets for personalized, preventive, or palliative/curative treatment of FM in the future. However, further blinded studies linking these potential biomarkers to FM could define a specific signature of women suffering from FM, which would be very useful for developing a robust diagnostic tool and focusing on probiotic-based treatments or other effective procedures for the patient.
Data availability statement
The datasets presented in this study are publicly available. The metagenomic data are deposited in GSA-Human under the accession number HRA009721 (BioProject accession: PRJCA032811), available at https://bigd.big.ac.cn/gsa-human/browse/HRA009721. The proteomic data are deposited in the PRIDE database under the ProteomeXchange accession number PXD059894, with the project webpage available at https://www.ebi.ac.uk/pride/archive/projects/PXD059894 and the FTP download at https://ftp.pride.ebi.ac.uk/pride/data/archive/2025/09/PXD059894.
Ethics statement
The studies involving humans were approved by Ethical Committee for Research with Medications (CEIM) at the Quirónsalud-Catalunya Hospital Group (protocol code IDI-20210749, approval record No. 01/2022). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
Author contributions
ED-G: Methodology, Formal analysis, Investigation, Writing – original draft. JR-T: Writing – review & editing, Supervision, Conceptualization, Funding acquisition, Project administration. MP-S: Writing – review & editing, Supervision. CM-T: Methodology, Writing – review & editing. RG-M: Methodology, Writing – review & editing. CD-L: Project administration, Supervision, Writing – review & editing. AM-L: Project administration, Writing – review & editing, Conceptualization, Supervision, Funding acquisition. DC: Conceptualization, Project administration, Writing – review & editing, Funding acquisition.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The study was conducted with the co-financing of Pronacera Therapeutics S.L and the Center for the Development of Industrial Technology (CDTI) of the Spanish Ministry of Science (project code: IDI-20210749).
Acknowledgments
This project has been fully completed with the kind collaboration of volunteers and FM patients, with a crucial role being played by several FM associations that will help to study coordination. These associations, in alphabetical order, are: ACEF (Asociación Cántabra de Fibromialgia; Santander, Spain), ACOFI (Asociación Cordobesa de Fibromialgia; Córdoba, Spain), AENFIPA (Asociación de Enfermos de Fibromialgia y Síndrome de Fatiga Crónica del Principado de Asturias; Oviedo, Spain), AFIAL (Asociación de Fibromialgia y Síndrome de Fatiga Crónica; Almería, Spain), AFIBRODON (Asociación de Enfermos de FM de Don Benito; Don Benito, Badajoz, Spain), AFIBROL (Asociación de Fibromialgia y Fatiga Crónica de Olivenza; Olivenza, Badajoz, Spain), AFIBROM (Asociación de Pacientes de Fibromialgia, Síndrome de Fatiga Crónica-Encefalomielitis Miálgica y Sensibilidad Química Múltiple de la Comunidad de Madrid; Madrid, Spain), AFIBROSAL (Asociación de Fibromialgia de Salamanca; Salamanca, Spain), AFIBROSE (Asociación de Fibromialgia de Sevilla; Seville, Spain), AFIBROVI (Asociación Fibromialgia y Fatiga Crónica de Villarrobledo; Villarrobledo, Albacete, Spain), AFICROVALL (Asociación de Fibromialgia y Síndrome de Fatiga Crónica de Valladolid; Valladolid, Spain), AFINORC (Asociación de Fibromialgia del Norte de Córdoba; Pozoblanco, Córdoba, Spain), AFIJE (Asociación de Fibromialgia Jerezana; Jerez de la Frontera, Cádiz, Spain), AFITA (Asociación de Fibromialgia de Tarifa; Tarifa, Cádiz, Spain), AFIXA (Asociación de Fibromialgia de Jaén; Jaén, Spain), AGRAFIM (Asociación Granadina de Fibromialgia; Granada, Spain), ALFIEL (Asociación Lucha contra la FM de El Ejido; El Ejido, Almería, Spain), APAFER (Asociación de Pacientes con Fibromialgia, Fatiga Crónica y otras Enfermedades Reumáticas; Mijas, Málaga, Spain), ASAFA (Asociación de Fibromialgia y Fatiga Crónica de Zaragoza; Zaragoza, Spain), AVAFI (Asociación Valenciana de Afectados de Fibromialgia; Valencia, Spain); CONFESQ (Coalición Nacional de Fibromialgia, Síndrome de Fatiga Crónica, Sensibilidad Química Múltiple y Electrohipersensibilidad; Spain), BAAF (Badalona Associació Afectats De Fibromiàlgia; Badalona, Barcelona, Spain), BIZI BIDE (Asociación Guipuzcoana de Fibromialgia y Síndrome de Fatiga Crónica; Gipuzkoa, Spain), FIBROFELS (Associació de Fibromiàlgia de Castelldefels; Casteldefels, Barcelona, Spain), FIBROSEGOVIA FM-SG (Asociación de FM de Segovia; Segovia, Spain), and FIBROSIERRA (Asociación de Enfermos de Fibromialgia; Aracena, Huelva, Spain).
Conflict of interest
Pronacera Therapeutics S.L. develops and provides molecular tests for the prevention, diagnosis, and personalized treatment of several conditions, involving the analysis of genes, proteins, and microbial components in human samples. Neither the patients nor the healthcare professionals participating in this study received any payment, nor did they have any influence. Hence, the study was conducted with the co-financing of Pronacera Therapeutics S.L and the Center for the Development of Industrial Technology (CDTI) of the Spanish Ministry of Science.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1641185/full#supplementary-material
Abbreviations
FM, fibromyalgia; CNS, central nervous system; SOD1, superoxide dismutase 1; CAT, catalase; NADPH, nicotinamide adenine dinucleotide phosphate; ROS, reactive oxygen species; HPA, hypothalamic pituitary adrenal; ACR, American College of Rheumatology; EULAR, European League Against Rheumatism; CI, confidence interval; CEIM, Ethical Committee for Research with Medications; PF, physical functioning; PH, physical health; P, pain; GH, general health; V, vitality; SF, social functioning; EP, emotional problems; MH, mental health; FIQR, Revised Fibromyalgia Impact Questionnaire; FDR, false discovery rate; IPA, ingenuity pathways analysis; zOTU, Zero-Radius Operational Taxonomic Unit; SD, standard deviation; H0, null hypothesis; AUC, area under the curve; ROC, receiver operating characteristic; FC, fold change; GABA, γ-aminobutiric acid.
References
Afzaal, M., Saeed, F., Shah, Y. A., Hussain, M., Rabail, R., Socol, C. T., et al. (2022). Human gut microbiota in health and disease: unveiling the relationship. Front. Microbiol. 13:999001. doi: 10.3389/fmicb.2022.999001
Akkuş, S., Naziroglu, M., Eriş, S., Yalman, K., Yilmaz, N., Yener, M., et al. (2009). Levels of lipid peroxidation, nitric oxide, and antioxidant vitamins in plasma of patients with fibromyalgia. Cell Biochem. Funct. 27, 181–185. doi: 10.1002/cbf.1548
Alcocer-Gómez, E., Garrido-Maraver, J., Bullón, P., Marín-Aguilar, F., Cotán, D., Carrión, A. M., et al. (2015). Metformin and caloric restriction induce an AMPK-dependent restoration of mitochondrial dysfunction in fibroblasts from Fibromyalgia patients. Biochim. Biophys. Acta Mol. Basis Dis. 1852, 1257–1267. doi: 10.1016/j.bbadis.2015.03.005
Altindag, O., and Celik, H. (2006). Total antioxidant capacity and the severity of the pain in patients with fibromyalgia. Redox Rep. 11, 131–135. doi: 10.1179/135100006X116628
Assavarittirong, C., Samborski, W., and Grygiel-Górniak, B. (2022). Oxidative stress in fibromyalgia: from pathology to treatment. Oxid. Med. Cell. Longev. 2022:1582432. doi: 10.1155/2022/1582432
Bains, A., Kohrman, S., Punko, D., and Fricchione, G. A. (2023). Link between inflammatory mechanisms and fibromyalgia. Adv. Exp. Med. Biol. 1411, 357–378. doi: 10.1007/978-981-19-7376-5_16
Baker, J. M., Al-Nakkash, L., and Herbst-Kralovetz, M. M. (2017). Estrogen–gut microbiome axis: physiological and clinical implications. Maturitas 103, 45–53. doi: 10.1016/j.maturitas.2017.06.025
Bennett, M., Chin, A., Lee, H. J., Morales Cestero, E., Strazielle, N., Ghersi-Egea, J. F., et al. (2021). Proteoglycan 4 reduces neuroinflammation and protects the blood-brain barrier after traumatic brain injury. J. Neurotrauma. 38, 385–398. doi: 10.1089/neu.2020.7229
Boomershine, C. (2015). Fibromyalgia: the prototypical central sensitivity syndrome. Curr. Rheumatol. Rev. 11, 131–145. doi: 10.2174/1573397111666150619095007
Cantó-Santos, J., Grau-Junyent, J. M., and Garrabou, G. (2020). The impact of mitochondrial deficiencies in neuromuscular diseases. Antioxidants 9, 1–29. doi: 10.3390/antiox9100964
Castro-Marrero, J., Cordero, M. D., Sáez-Francas, N., Jimenez-Gutierrez, C., Aguilar-Montilla, F. J., Aliste, L., et al. (2013). Could mitochondrial dysfunction be a differentiating marker between chronic fatigue syndrome and fibromyalgia? Antioxid. Redox Signal. 19, 1855–1860. doi: 10.1089/ars.2013.5346
Chen, W., Zhao, H., and Mitochondrial, L. i. Y. (2023). dynamics in health and disease: mechanisms and potential targets. Signal Transduct. Target. Ther. 8:1–25. doi: 10.1038/s41392-023-01547-9
Clauw, D. J. (2014). Fibromyalgia: a clinical review. JAMA. 311, 1547–55. doi: 10.1001/jama.2014.3266
Clos-Garcia, M., Andrés-Marin, N., Fernández-Eulate, G., Abecia, L., Lavín, J. L., van Liempd, S., et al. (2019). Gut microbiome and serum metabolome analyses identify molecular biomarkers and altered glutamate metabolism in fibromyalgia. EBioMedicine 46, 499–511. doi: 10.1016/j.ebiom.2019.07.031
Cordero, M. D., Alcocer-Gómez, E., Marín-Aguilar, F., Rybkina, T., Cotán, D., Pérez-Pulido, A., et al. (2016). Mutation in cytochrome b gene of mitochondrial DNA in a family with fibromyalgia is associated with NLRP3-inflammasome activation. J. Med. Genet. 53, 113–122. doi: 10.1136/jmedgenet-2015-103392
Cordero, M. D., De Miguel, M., Moreno Fernández, A. M., Carmona López, I. M., Garrido Maraver, J., Cotán, D., et al. (2010). Mitochondrial dysfunction and mitophagy activation in blood mononuclear cells of fibromyalgia patients: implications in the pathogenesis of the disease. Arthritis Res. Ther. 12:R17. doi: 10.1186/ar2918
Cordero, M. D., Díaz-Parrado, E., Carrión, A. M., Alfonsi, S., Sánchez-Alcazar, J. A., Bullón, P., et al. (2013). Is inflammation a mitochondrial dysfunction-dependent event in fibromyalgia? Antioxid. Redox Signal 18, 800–807. doi: 10.1089/ars.2012.4892
Coskun Benlidayi, I. (2019). Role of inflammation in the pathogenesis and treatment of fibromyalgia. Rheumatol. Int. 39, 781–91. doi: 10.1007/s00296-019-04251-6
Costa, S. P., Lage, L. V., Da Mota, L. H., and De Carvalho, J. F. (2011). Fibromyalgia in primary antiphospholipid (Hughes) syndrome. Lupus 20, 1182–1186. doi: 10.1177/0961203311411962
Darvesh, S., Hopkins, D. A., and Geula, C. (2003). Neurobiology of butyrylcholinesterase. Nat. Rev. Neurosci. 4, 131–138. doi: 10.1038/nrn1035
Desbonnet, L., Garrett, L., Clarke, G., Kiely, B., Cryan, J. F., Dinan, T. G., et al. (2010). Effects of the probiotic Bifidobacterium infantis in the maternal separation model of depression. Neuroscience 170, 1179–1188. doi: 10.1016/j.neuroscience.2010.08.005
Erdrich, S., Hawrelak, J. A., Myers, S. P., and Harnett, J. E. (2020). Determining the association between fibromyalgia, the gut microbiome and its biomarkers: a systematic review. BMC Musculoskelet. Disord. 21:181. doi: 10.1186/s12891-020-03201-9
Ervin, S. M., Li, H., Lim, L., Roberts, L. R., Liang, X., Mani, S., et al. (2019). Gut microbial β-glucuronidases reactivate estrogens as components of the estrobolome that reactivate estrogens. J. Biol. Chem. 294, 18586–18599. doi: 10.1074/jbc.RA119.010950
Ewels, P., Magnusson, M., Lundin, S., and Käller, M. (2016). MultiQC: summarize analysis results for multiple tools and samples in a single report. Bioinformatics 32, 3047–3048. doi: 10.1093/bioinformatics/btw354
Falco, P., Galosi, E., Di Stefano, G., Leone, C., Di Pietro, G., Tramontana, L., et al. (2024). Autonomic small-fiber pathology in patients with fibromyalgia. J. Pain 25, 64–72. doi: 10.1016/j.jpain.2023.07.020
Frigo, D. E., Bondesson, M., and Williams, C. (2021). Nuclear receptors: from molecular mechanisms to therapeutics. Essays Biochem. 65, 847–856. doi: 10.1042/EBC20210020
Garofalo, C., Cristiani, C. M., Ilari, S., Passacatini, L. C., Malafoglia, V., Viglietto, G., et al. (2023). Fibromyalgia and irritable bowel syndrome interaction: a possible role for gut microbiota and gut-brain axis. Biomedicines 11:1701. doi: 10.3390/biomedicines11061701
Gerdle, B., Ghafouri, B., Lund, E., Bengtsson, A., Lundberg, P., van Ettinger-Veenstra, H., et al. (2020). Evidence of mitochondrial dysfunction in fibromyalgia: deviating muscle energy metabolism detected using microdialysis and magnetic resonance. J. Clin. Med. 9, 1–26. doi: 10.3390/jcm9113527
Giorgi, V., Bazzichi, L., Batticciotto, A., Pellegrino, G., Di Franco, M., Sirotti, S., et al. (2023). Fibromyalgia: one year in review 2023. Clin. Exp. Rheumatol. 41, 1205–1213. doi: 10.55563/clinexprheumatol/257e99
Goebel, A., Buhner, S., Schedel, R., Lochs, H., and Sprotte, G. (2008). Altered intestinal permeability in patients with primary fibromyalgia and in patients with complex regional pain syndrome. Rheumatology 47, 1223–1227. doi: 10.1093/rheumatology/ken140
Goebel, A., Krock, E., Gentry, C., Israel, M. R., Jurczak, A., Urbina, C. M., et al. (2021). Passive transfer of fibromyalgia symptoms from patients to mice. J. Clin. Invest. 131:7344. doi: 10.1172/JCI144201
Gross, A., Pack, L. A. P., Schacht, G. M., Kant, S., Ungewiss, H., Meir, M., et al. (2018). Desmoglein 2, but not desmocollin 2, protects intestinal epithelia from injury. Mucosal Immunol. 11, 1630–1639. doi: 10.1038/s41385-018-0062-z
Hamamah, S., Aghazarian, A., Nazaryan, A., Hajnal, A., and Covasa, M. (2022). Role of microbiota-gut-brain axis in regulating dopaminergic signaling. Biomedicines 10:436. doi: 10.3390/biomedicines10020436
Han, C. L., Sheng, Y. C., Wang, S. Y., Chen, Y. H., and Kang, J. H. (2020). Serum proteome profiles revealed dysregulated proteins and mechanisms associated with fibromyalgia syndrome in women. Sci. Rep. 10:12347. doi: 10.1038/s41598-020-69271-w
Hernandez-Leon, A., De la Luz-Cuellar, Y. E., Granados-Soto, V., González-Trujano, M. E., and Fernández-Guasti, A. (2018). Sex differences and estradiol involvement in hyperalgesia and allodynia in an experimental model of fibromyalgia. Horm. Behav. 97, 39–46. doi: 10.1016/j.yhbeh.2017.10.011
Hu, S., Ding, Q., Zhang, W., Kang, M., Ma, J., Zhao, L., et al. (2023). Gut microbial beta-glucuronidase: a vital regulator in female estrogen metabolism. Gut Microbes 15:2236749. doi: 10.1080/19490976.2023.2236749
Illumina (n.d.). 16S Metagenomic Sequencing Library Preparation: Preparing 16S Ribosomal RNA Gene Amplicons for the Illumina MiSeq System (Part # 15044223 Rev. B) [PDF]. Illumina. Available online at: https://support.illumina.com/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf (Retrieved September 2, 2024).
Inciarte-Mundo, J., Frade-Sosa, B., and Sanmartí, R. (2022). From bench to bedside: calprotectin (S100A8/S100A9) as a biomarker in rheumatoid arthritis. Front. Immunol. 13:1001025. doi: 10.3389/fimmu.2022.1001025
Jefatura del Estado. (2018). Ley Orgánica 3/2018, de 5 de diciembre, de Protección de Datos Personales y garantía de los derechos digitales [Ley Orgánica, Disposición 16673]. Bol. Off. Estado 294:119788. Available online at: https://www.boe.es/buscar/act.php?id=BOE-A-2018-16673 (Retrieved September 2, 2024).
Kaci, G., Goudercourt, D., Dennin, V., Pot, B., Doré, J., Ehrlich, S. D., et al. (2014). Anti-inflammatory properties of Streptococcus salivarius, a commensal bacterium of the oral cavity and digestive tract. Appl. Environ. Microbiol. 80, 928–934. doi: 10.1128/AEM.03133-13
Komorowski, A. S., and Pezo, R. C. (2020). Untapped “-omics”: the microbial metagenome, estrobolome, and their influence on the development of breast cancer and response to treatment. Breast Cancer Res. Treat. 179, 287–300. doi: 10.1007/s10549-019-05472-w
Kundakci, B., Kaur, J., Goh, S. L., Hall, M., Doherty, M., Zhang, W., et al. (2022). Efficacy of nonpharmacological interventions for individual features of fibromyalgia: a systematic review and meta-analysis of randomised controlled trials. Pain 163, 1432–45. doi: 10.1097/j.pain.0000000000002500
Kwa, M., Plottel, C. S., Blaser, M. J., and Adams, S. (2016). The intestinal microbiome and estrogen receptor-positive female breast cancer. J. Natl. Cancer Inst. 108:djw029. doi: 10.1093/jnci/djw029
Lin, Y. J., Ko, Y. C., Chow, L. H., Hsiao, F. J., Liu, H. Y., Wang, P. N., et al. (2021). Salivary cortisol is associated with cognitive changes in patients with fibromyalgia. Sci. Rep. 11:1311. doi: 10.1038/s41598-020-79349-0
Littlejohn, G., and Guymer, E. (2018). Neurogenic inflammation in fibromyalgia. Semin. Immunopathol. 40, 291–300. doi: 10.1007/s00281-018-0672-2
Liu, L., Wu, Q., Chen, Y., Ren, H., Zhang, Q., Yang, H., et al. (2023). Gut microbiota in chronic pain: Novel insights into mechanisms and promising therapeutic strategies. Int. Immunopharmacol. 115:109685. doi: 10.1016/j.intimp.2023.109685
Longley, K. (2006). Fibromyalgia: aetiology, diagnosis, symptoms and management. Br. J. Nurs. 15, 729–33. doi: 10.12968/bjon.2006.15.13.21487
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Lucena Del Amo, L., Durán-González, E., Ramírez-Tejero, J. A., Martínez-Lara, A., and Cotán, D. (2023). Study protocol for FIBROKIT: a new tool for fibromyalgia diagnosis and patient follow-up. Front. Neurol. 14:1286539. doi: 10.3389/fneur.2023.1286539
Macfarlane, G. J., Kronisch, C., Dean, L. E., Atzeni, F., Häuser, W., Flub, E., et al. (2017). EULAR revised recommendations for the management of fibromyalgia. Ann. Rheum. Dis. 76, 318–328. doi: 10.1136/annrheumdis-2016-209724
Maes, M., Almulla, A. F., Zhou, B., Algon, A. A. A., and Sodsai, P. (2024). In major dysmood disorder, physiosomatic, chronic fatigue and fibromyalgia symptoms are driven by immune activation and increased immune-associated neurotoxicity. Sci. Rep. 14:7344. doi: 10.1038/s41598-024-57350-1
Marques, A. P., Santo, A. S. E., Berssaneti, A. A., Matsutani, L. A., and Yuan, S. L. K. (2017). Prevalence of fibromyalgia: literature review update. Rev. Bras. Reumatol. 57, 356–63. doi: 10.1016/j.rbr.2016.10.004
Martínez-Lara, A., Moreno-Fernández, A. M., Jiménez-Guerrero, M., Díaz-López, C., De-Miguel, M., Cotán, D., et al. (2020). Mitochondrial imbalance as a new approach to the study of fibromyalgia. Open Access Rheumatol. 12, 175–185. doi: 10.2147/OARRR.S257470
Martins, D. F., Viseux, F. J. F., Salm, D. C., Ribeiro, A. C. A., da Silva, H. K. L., Seim, L. A., et al. (2021). The role of the vagus nerve in fibromyalgia syndrome. Neurosci. Biobehav. Rev. 131, 1136–49. doi: 10.1016/j.neubiorev.2021.10.021
Mas, A. J., Carmona, L., Valverde, M., Ribas, B., and Study Group, E. (2008). Prevalence and impact of fi bromyalgia on function and quality of life in individuals from the general population: results from a nationwide study in Spain. Clin. Exp. Rheumatol. 26, 519–526.
Meeus, M., Nijs, J., Hermans, L., Goubert, D., and Calders, P. (2013). The role of mitochondrial dysfunctions due to oxidative and nitrosative stress in the chronic pain or chronic fatigue syndromes and fibromyalgia patients: peripheral and central mechanisms as therapeutic targets? Expert Opin. Ther. Targets 17, 1081–1089. doi: 10.1517/14728222.2013.818657
Mello, T., Materozzi, M., and Galli, A. (2016). PPARs and mitochondrial metabolism: from NAFLD to HCC. PPAR Res. 2016:7403230. doi: 10.1155/2016/7403230
Minerbi, A., Gonzalez, E., Brereton, N. J. B., Anjarkouchian, A., Dewar, K., Fitzcharles, M. A., et al. (2019). Altered microbiome composition in individuals with fibromyalgia. Pain 160, 2589–2602. doi: 10.1097/j.pain.0000000000001640
Mohapatra, G., Dachet, F., Coleman, L. J., Gillis, B., and Behm, F. G. (2024). Identification of unique genomic signatures in patients with fibromyalgia and chronic pain. Sci. Rep. 14:3949. doi: 10.1038/s41598-024-53874-8
Moloney, R. D., Johnson, A. C., O'Mahony, S. M., Dinan, T. G., Greenwood-Van Meerveld, B., Cryan, J. F., et al. (2016). Stress and the microbiota-gut-brain axis in visceral pain: relevance to irritable bowel syndrome. CNS Neurosci. Ther. 22, 102–117. doi: 10.1111/cns.12490
Mueller, C., Fang, Y. H. D., Jones, C., McConathy, J. E., Raman, F., Lapi, S. E., et al. (2023). Evidence of neuroinflammation in fibromyalgia syndrome: a [18F]DPA-714 positron emission tomography study. Pain 164, 2285–2295. doi: 10.1097/j.pain.0000000000002927
Naing, L., Winn, T., and Rusli, B. N. (2006). Practical issues in calculating the sample size for prevalence studies. Arch. Orofacial Sci. 1, 9–14.
Nhu, N. T., Chen, D. Y. T., Yang, Y. C. S. H., Lo, Y. C., and Kang, J. H. (2024). Associations between brain-gut axis and psychological distress in fibromyalgia: a microbiota and magnetic resonance imaging study. J. Pain. 25, 934–945. doi: 10.1016/j.jpain.2023.10.015
Okifuji, A., and Turk, D. C. (2006). Sex hormones and pain in regularly menstruating women with fibromyalgia syndrome. J. Pain 7, 851–859. doi: 10.1016/j.jpain.2006.04.005
Oksanen, J. (2025). Vegan: Ecological Diversity [R vignette]. In vegan (Version 2.7-1) [R package]. CRAN. Available online at: https://cran.r-project.org/web/packages/vegan/vignettes/diversity-vegan.pdf (Retrieved September 2024).
O'Mahony, L. F., Srivastava, A., Mehta, P., and Ciurtin, C. (2021). Is fibromyalgia associated with a unique cytokine profile? A systematic review and meta-analysis. Rheumatology 60, 2602–2614. doi: 10.1093/rheumatology/keab146
Otaru, N., Ye, K., Mujezinovic, D., Berchtold, L., Constancias, F., Cornejo, F. A., et al. (2021). GABA production by human intestinal Bacteroides spp.: prevalence, regulation, and role in acid stress tolerance. Front. Microbiol. 12:656895. doi: 10.3389/fmicb.2021.656895
Ovejero, T., Sadones, O., Sánchez-Fito, T., Almenar-Pérez, E., Espejo, J. A., Martín-Martínez, E., et al. (2020). Activation of transposable elements in immune cells of fibromyalgia patients. Int. J. Mol. Sci. 21:1366. doi: 10.3390/ijms21041366
Ozgocmen, S., Ozyurt, H., Sogut, S., Akyol, O., Ardicoglu, O., Yildizhan, H., et al. (2006). Antioxidant status, lipid peroxidation and nitric oxide in fibromyalgia: Etiologic and therapeutic concerns. Rheumatol. Int. 26, 598–603. doi: 10.1007/s00296-005-0079-y
Pai, A. H. Y., Wang, Y. W., Lu, P. C., Wu, H. M., Xu, J. L., Huang, H. Y., et al. (2023). Gut microbiome-estrobolome profile in reproductive-age women with endometriosis. Int. J. Mol. Sci. 24:16301. doi: 10.3390/ijms242216301
Pollet, R. M., D'Agostino, E. H., Walton, W. G., Xu, Y., Little, M. S., Biernat, K. A., et al. (2017). An atlas of β-glucuronidases in the human intestinal microbiome. Structure 25, 967–977.e5. doi: 10.1016/j.str.2017.05.003
Pronacera. Clinicaltrials (2024). Tool for Fibromyalgia Diagnosis and Effect of Extra Virgin Olive Oil (FIBROKIT). Available online at: https://clinicaltrials.gov/study/NCT05921409#contacts-and-locations (Accessed July 23, 2024).
Ramírez-Tejero, J. A., Durán-González, E., Martínez-Lara, A., Lucena del Amo, L., Sepúlveda, I., Huancas-Díaz, A., et al. (2023). Microbiota and mitochondrial sex-dependent imbalance in fibromyalgia: a pilot descriptive study. Neurol. Int. 15, 868–880. doi: 10.3390/neurolint15030055
Ramírez-Tejero, J. A., Martínez-Lara, E., Rus, A., Camacho, M. V., Del Moral, M. L., Siles, E., et al. (2018). Insight into the biological pathways underlying fibromyalgia by a proteomic approach. J. Proteomics 186, 47–55. doi: 10.1016/j.jprot.2018.07.009
Rawish, E., Sauter, M., Sauter, R., Nording, H., and Langer, H. F. (2021). Complement, inflammation and thrombosis. Br. J. Pharmacol. 178, 2892–904. doi: 10.1111/bph.15476
Reyes del Paso, G. A., and de la Coba, P. (2020). Reduced activity, reactivity and functionality of the sympathetic nervous system in fibromyalgia: an electrodermal study. PLoS ONE 15:e0244830. doi: 10.1371/journal.pone.0244830
Rus, A., Molina, F., Gassó, M., Camacho, M. V., Peinado, M. Á., and del Moral, M. L. (2016). Nitric oxide, inflammation, lipid profile, and cortisol in normal- and overweight women with fibromyalgia. Biol. Res. Nurs. 18, 138–146. doi: 10.1177/1099800415591035
Rus, A., Robles-Fernandez, I., Martinez-Gonzalez, L. J., Carmona, R., and Alvarez-Cubero, M. J. (2021). Influence of oxidative stress-related genes on susceptibility to fibromyalgia. Nurs. Res. 70, 44–50. doi: 10.1097/NNR.0000000000000480
Rush, R. A., and Geffen, L. B. (1980). Dopamine beta-hydroxylase in health and disease. Crit. Rev. Clin. Lab. Sci. 12, 241–277. doi: 10.3109/10408368009108731
Rutsch, A., Kantsjö, J. B., and Ronchi, F. (2020). The gut-brain axis: how microbiota and host inflammasome influence brain physiology and pathology. Front. Immunol. 11:604179. doi: 10.3389/fimmu.2020.604179
Salliss, M. E., Farland, L. V., Mahnert, N. D., and Herbst-Kralovetz, M. M. (2022). The role of gut and genital microbiota and the estrobolome in endometriosis, infertility and chronic pelvic pain. Hum. Reprod. Update 28, 92–131. doi: 10.1093/humupd/dmab035
Sánchez-Domínguez, B., Bullón, P., Román-Malo, L., Marín-Aguilar, F., Alcocer-Gómez, E., Carrión, A. M., et al. (2015). Oxidative stress, mitochondrial dysfunction and, inflammation common events in skin of patients with Fibromyalgia. Mitochondrion 21, 69–75. doi: 10.1016/j.mito.2015.01.010
Schertzinger, M., Wesson-Sides, K., Parkitny, L., and Younger, J. (2018). Daily fluctuations of progesterone and testosterone are associated with fibromyalgia pain severity. J. Pain 19, 410–417. doi: 10.1016/j.jpain.2017.11.013
Serra, J., Collado, A., Solà, R., Antonelli, F., Torres, X., Salgueiro, M., et al. (2014). Hyperexcitable C nociceptors in fibromyalgia. Ann. Neurol. 75, 196–208. doi: 10.1002/ana.24065
Sherrill, J. D., Kc, K., Wu, D., Djukic, Z., Caldwell, J. M., Stucke, E. M., et al. (2014). Desmoglein-1 regulates esophageal epithelial barrier function and immune responses in eosinophilic esophagitis. Mucosal Immunol. 7, 718–729. doi: 10.1038/mi.2013.90
Sibilia, J. (2003). Antiphospholipid syndrome: why and how should we make the diagnosis? Joint Bone Spin 70, 97–102. doi: 10.1016/s1297-319x(03)00014-9
Siddiqui, R., Makhlouf, Z., Alharbi, A. M., Alfahemi, H., and Khan, N. A. (2022). The gut microbiome and female health. Biology 11:1683. doi: 10.3390/biology11111683
Silman, I. (2021). The multiple biological roles of the cholinesterases. Prog. Biophys. Mol. Biol. 162, 41–56. doi: 10.1016/j.pbiomolbio.2020.12.001
Slim, M., Calandre, E. P., and Rico-Villademoros, F. (2015). An insight into the gastrointestinal component of fibromyalgia: clinical manifestations and potential underlying mechanisms. Rheumatol. Int. 35, 433–444. doi: 10.1007/s00296-014-3109-9
Sobstyl, M., Brecht, P., Sobstyl, A., Mertowska, P., and Grywalska, E. (2022). The role of microbiota in the immunopathogenesis of endometrial cancer. Int. J. Mol. Sci. 23:5756. doi: 10.3390/ijms23105756
Tarze, A., Deniaud, A., Le Bras, M., Maillier, E., Molle, D., Larochette, N., et al. (2007). GAPDH, a novel regulator of the pro-apoptotic mitochondrial membrane permeabilization. Oncogene 26, 2606–2620. doi: 10.1038/sj.onc.1210074
Tomasello, G., Mazzola, M., Bosco, V., Tomasello, G., Damiani, P., Sinagra, E., et al. (2018). Intestinal dysbiosis and hormonal neuroendocrine secretion in the fibromyalgic patient. Biomed. Pap. 164, 258–262. doi: 10.5507/bp.2018.051
Valera-Calero, J. A., Arendt-Nielsen, L., Cigarán-Méndez, M., Fernández-de-las-Peñas, C., and Varol, U. (2022). Network analysis for better understanding the complex psycho-biological mechanisms behind fibromyalgia syndrome. Diagnostics 12:1845. doi: 10.3390/diagnostics12081845
Vasquez, A. (2016). Neuroinflammation in fibromyalgia and CRPS is multifactorial. Nat. Rev. Rheumatol. 12:242. doi: 10.1038/nrrheum.2016.25
Vecchio, E., Lombardi, R., Paolini, M., Libro, G., Delussi, M., Ricci, K., et al. (2020). Peripheral and central nervous system correlates in fibromyalgia. Eur. J. Pain 24, 1537–1547. doi: 10.1002/ejp.1607
Vega-Ramírez, M. T., Becerril-Villanueva, E., Maldonado-García, J. L., Pavón, L., and Pérez-Sánchez, G. (2024). S100 proteins: a new frontier in fibromyalgia research. Mol. Brain. 17:29. doi: 10.1186/s13041-024-01102-9
Verma, V., Drury, G. L., Parisien, M., Özdag Acarli, A. N., Al-Aubodah, T. A., Nijnik, A., et al. (2022). Unbiased immune profiling reveals a natural killer cell-peripheral nerve axis in fibromyalgia. Pain 163, E821–E836. doi: 10.1097/j.pain.0000000000002498
Vilagut, G., Ferrer, M., Rajmil, L., Rebollo, P., Permanyer-Miralda, G., Quintana, J. M., et al. (2005). El Cuestionario de Salud SF-36 español: una década de experiencia y nuevos desarrollos por los investigadores de la Red-IRYSS*. Gac Sanit. 19, 135–150. doi: 10.1157/13074369
Wåhlén, K., Ernberg, M., Kosek, E., Mannerkorpi, K., Gerdle, B., Ghafouri, B., et al. (2020). Significant correlation between plasma proteome profile and pain intensity, sensitivity, and psychological distress in women with fibromyalgia. Sci. Rep. 10:12508. doi: 10.1038/s41598-020-69422-z
Wang, Y., Wei, J., Zhang, W., Doherty, M., Zhang, Y., Xie, H., et al. (2022). Gut dysbiosis in rheumatic diseases: a systematic review and meta-analysis of 92 observational studies. EBioMedicine 80:104055. doi: 10.1016/j.ebiom.2022.104055
Warren, T., McAllister, R., Morgan, A., Rai, T. S., McGilligan, V., Ennis, M., et al. (2021). The interdependency and co-regulation of the vitamin D and cholesterol metabolism. Cells 10:2007. doi: 10.3390/cells10082007
Wolfe, F., Clauw, D. J., Fitzcharles, M. A., Goldenberg, D. L., Häuser, W., Katz, R. L., et al. (2016). 2016 Revisions to the 2010/2011 fibromyalgia diagnostic criteria. Semin. Arthritis Rheum. 46, 319–329. doi: 10.1016/j.semarthrit.2016.08.012
Wolfe, F., Clauw, D. J., Fitzcharles, M. A., Goldenberg, D. L., Häuser, W., Katz, R. S., et al. (2011). Fibromyalgia criteria and severity scales for clinical and epidemiological studies: a modification of the ACR preliminary diagnostic criteria for fibromyalgia. J. Rheumatol. 38, 1113–1122. doi: 10.3899/jrheum.100594
Wolfe, F., Smythe, H. A., Yunus, M. B., Bennett, R. M., Bombardier, C., Goldenberg, D. L., et al. (1990). The American collegue of rheumathology 1990 criteria for the classification of fribomyalgia. Report of the Multicenter Criteria Committee. Arthritis Rheum. 33, 160–172. doi: 10.1002/art.1780330203
Keywords: fibromyalgia, diagnostic, proteome, microbiome, mitochondria
Citation: Durán-González E, Ramírez-Tejero JA, Pérez-Sánchez M, Morales-Torres C, Gómez-Morano R, Díaz-López C, Martínez-Lara A and Cotán D (2025) Fibromyalgia diagnosis from a multi-omics approach: a gut feeling. Front. Microbiol. 16:1641185. doi: 10.3389/fmicb.2025.1641185
Received: 06 June 2025; Accepted: 29 August 2025;
Published: 02 October 2025.
Edited by:
Jianan Zhao, Temple University, United StatesReviewed by:
Faheem Ahmed Khan, National Research and Innovation Agency (BRIN), IndonesiaShreya Madhav Nuguri, The Ohio State University, United States
Copyright © 2025 Durán-González, Ramírez-Tejero, Pérez-Sánchez, Morales-Torres, Gómez-Morano, Díaz-López, Martínez-Lara and Cotán. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elena Durán-González, ZS5kdXJhbkBwcm9uYWNlcmEuY29t