 Lucía Saiz-Escobedo1,2
Lucía Saiz-Escobedo1,2 Mónica Ballestero-Tellez3Mireia Rajadell3Marc Garreta-Esteban3Laura Gisbert4
Mónica Ballestero-Tellez3Mireia Rajadell3Marc Garreta-Esteban3Laura Gisbert4 Irene Cadenas-Jiménez1,2,5Rosa Maria Costa1
Irene Cadenas-Jiménez1,2,5Rosa Maria Costa1 Fe Tubau1,5David Sánchez-Ramos1
Fe Tubau1,5David Sánchez-Ramos1 M. Angeles Domínguez1,2,6
M. Angeles Domínguez1,2,6 Kyriaki Xanthopoulou7,8
Kyriaki Xanthopoulou7,8 Paul G. Higgins7,8
Paul G. Higgins7,8 Carmen Ardanuy1,2,5
Carmen Ardanuy1,2,5 Aida González-Díaz1,5*
Aida González-Díaz1,5* Sara Marti1,5,9*
Sara Marti1,5,9*- 1Microbiology Department, Hospital Universitari de Bellvitge, IDIBELL-UB, Barcelona, Spain
- 2Department of Pathology and Experimental Therapeutics, University of Barcelona, Barcelona, Spain
- 3Clinical Microbiology Department, Catlab, Barcelona, Spain
- 4Infectious Diseases Department, Hospital Universitario Mutua Terrassa, Terrassa, Spain
- 5Research Network for Respiratory Diseases (CIBERES), ISCIII, Madrid, Spain
- 6Research Network for Infectious Diseases (CIBERINFEC), ISCIII, Madrid, Spain
- 7Institute for Medical Microbiology, Immunology and Hygiene, Faculty of Medicine and University Hospital Cologne, University of Cologne, Cologne, Germany
- 8German Center for Infection Research, Partner Site Bonn-Cologne, Cologne, Germany
- 9Department of Medicine, School of Medicine, University of Barcelona, Barcelona, Spain
Introduction: Haemophilus parainfluenzae is an opportunistic pathogen increasingly associated with urogenital infections and multidrug resistance. While polysaccharide capsules are known virulence factors in H. influenzae, their role in H. parainfluenzae remains poorly understood. This study aimed to characterize a new capsular operon identified in H. parainfluenzae and analyze the antimicrobial resistance profile of strains carrying this operon to provide insights into their pathogenic potential.
Methods: Two clinical isolates from genital ulcers were subjected to whole-genome sequencing. The capsular operon was identified and characterized using comparative genomics. Antimicrobial susceptibility was determined using broth microdilution, and the resistance determinants were identified through genomic analysis.
Results: A novel capsular operon, designated H. parainfluenzae HPAR_type4, was identified in both isolates. This operon spans 11,885 bp and comprises nine predicted open reading frames (ORFs) organized into the three regions characteristic of Haemophilus spp. Regions I, III, and the first ORF of region II showed high homology with the HPAR_type1 operon, while the remaining ORFs in region II shared identity with the cpsB and cpsC genes from H. sputorum HSPU_type1. Both strains exhibited multidrug resistance, with one strain carrying a CTX-M-15 extended-spectrum β-lactamase gene integrated in an integrative and conjugative element (ICE), ICEHpaHUB6. Two distinct potentially conjugative plasmids were identified, each carrying genes related to replication, mobility, and putative virulence functions.
Conclusion: The identification of a novel capsular operon in multidrug-resistant H. parainfluenzae strains highlights the species genetic plasticity and potential for increased virulence. These findings underscore the importance of ongoing surveillance in clinical settings to monitor the spread of antibiotic resistance and novel virulence factors, which may contribute to its pathogenicity and impact future treatment strategies.
1 Introduction
Haemophilus parainfluenzae is an opportunistic Gram-negative pathogen that colonizes the respiratory and urogenital tracts (Nørskov-Lauritsen, 2014). This species has been implicated as a cause for various diseases, including respiratory tract infections (Kosikowska et al., 2016), endocarditis (Revest et al., 2016) and arthritis (Baron et al., 2020), with a notably high prevalence as a cause of urethritis in recent years (Ducours et al., 2020; Hsu et al., 2015; Magdaleno-Tapial et al., 2019).
Polysaccharide capsules, a major virulence factor in H. influenzae, have also been identified in H. parainfluenzae, although their precise role in this species remains unclear. In H. influenzae, these capsules are crucial for evading host immune responses and have been used in vaccine development (González-Díaz et al., 2019; Sierra et al., 2021; Watts and Holta, 2019). The genetic blueprint for these polysaccharide capsules in the genus Haemophilus is encoded in the cap locus, which comprises three different regions. Region I has four conserved genes (bexABCD) that encode the export apparatus for the translocation of capsular polysaccharides. Region II contains serotype-specific genes (three to eight genes) involved in the polysaccharide biosynthesis. Finally, region III has two genes (hcsAB) responsible for polysaccharide transport across the outer membrane (Potts et al., 2019). To date, three different capsular operons have been described in H. parainfluenzae: HPAR-type1 (González-Díaz et al., 2019) and HPAR-type2 (Sierra et al., 2021), both detected in urogenital strains, and the serotype f-like operon recently identified in colonizing strains from nasopharyngeal samples of healthy children in Portugal (Bajanca-Lavado et al., 2022).
Along with virulence factors, antimicrobial resistance has emerged as a significant concern in H. parainfluenzae. The increasing incidence of multidrug resistance in this species is attributed to target-gene mutations and the acquisition of transferable resistance genes. Extensively drug-resistant (XDR) strains have been detected in recent years, particularly in urogenital samples, showing resistance to β-lactams, fluoroquinolones, macrolides, chloramphenicol, trimethoprim-sulfamethoxazole and tetracycline. Notably, our group previously reported the presence of a CTX-M-15 extended spectrum β-lactamase (ESBL) in four XDR H. parainfluenzae strains isolated from three urethral exudates and a genital ulcer (Saiz-Escobedo et al., 2023). This finding was subsequently corroborated by the identification of the CTX-M-15 ESBL in a urogenital XDR H. parainfluenzae strain from France (Caméléna et al., 2024).
The main objective of this study is to characterize a novel capsular operon detected in two XDR H. parainfluenzae strains isolated from genital ulcers in MSM patients. Additionally, we also aim to provide a comprehensive genotypic and phenotypic characterization of these capsulated strains, with particular emphasis on their antimicrobial resistance profiles.
2 Materials and methods
2.1 Bacterial strains and antimicrobial susceptibility
Two H. parainfluenzae strains harboring a new capsular operon, identified by whole genome sequencing (WGS), were included in this study. The first strain, HUB-HP17268, was isolated in 2022 from a genital chancre exudate of a young male patient with a genital ulcer who attended the Hospital Universitari de Bellvitge (HUB) in Barcelona (Cataluña, Spain). The second strain, HUMT-HP05, was isolated in 2024 from a genital ulcer of a young MSM patient attending the Pre-exposure Prophylaxis (PrEP) unit at Hospital Universitari Mútua de Terrassa (HUMT), also in Cataluña, Spain.
Strains were cultured on chocolate agar (BioMérieux, Marcy-l’Étoile, France) and incubated at 37 °C in a 5% CO2 atmosphere. Bacterial identification was performed by MALDI-TOF mass spectrometry (Bruker and MS-Prime BioMérieux).
Antimicrobial susceptibility was assessed by microdilution using STRHAE2 Sensititre commercial panels (Thermo Fisher Scientific, Waltham, MA, USA). Double-disk synergy test was conducted to detect ESBL production, using amoxicillin-clavulanic acid along with the β-lactams, cefotaxime, ceftazidime, aztreonam, and cefuroxime. All susceptibility testing procedures followed the European Committee on Antimicrobial Susceptibility Testing criteria for H. influenzae.1
2.2 Whole genome sequencing
Genomic DNA was extracted using the QIAmp DNA mini kit (Qiagen) and quantified with Qubit 4 (Thermo Fisher Scientific). Short-read libraries were prepared using the Illumina DNA Prep kit for paired-end sequencing (2 × 300 bp) on the MiSeq Platform (Illumina Inc.). Long-read libraries were prepared with Native Barcoding Expansion (EXP-NBD196) and Ligation Sequencing kit (SQK-LSK109), followed by sequencing on FLOMIN106D flow cells (R9.4.1) from Oxford Nanopore Technologies. Reads were assembled using Bactopia2 and hybrid assemblies combining short and long reads were generated using the Unycicler pipeline3 (Wick et al., 2017). Raw reads were uploaded to the European Nucleotide Archive (PRJEB88731). To elucidate genomic similarity between both genomes, a whole genome alignment was performed with Snippy v4.6.04 using HUMT-HP05 as reference. The final alignment was screened with Geneious R9 (version 9.1.7, Biomatters) and recombinant blocks were defined as the presence of 3 SNPs in windows of 100–1,000 bp.
2.3 Capsular operon determination
Capsular loci were identified in silico using HiCap5 (Watts and Holta, 2019). BLASTn searches were performed with Geneious R9 (version 9.1.7, Biomatters) to compare the sequences against known capsular operons from Haemophilus spp. Reference sequences included H. parainfluenzae HPAR-type1 (MH644108), HPAR-type2 (MT185932), and serotype f-like operon (JAMLEO010000002:224525-240017); H. influenzae serotype a (CP017811:328547-341597), serotype b (NC_016809:774911-790627), serotype c (HQ651151), serotype d (HM770877), serotype e (FM882247) and serotype f (CP005967:675016-687441); H. sputorum HSPU_type1 (NZ_AFNK01000031:141496-153330) and HSPU_type2 (QEPN01000003:47115-64856); and H. haemolyticus (SDPB01000019:33146-48666). Open reading frames (ORFs) were predicted, and sequence identity between putative genes/proteins from different capsular operons was calculated as the percentage of identical number of bases/residues divided by the length of the longest gene/protein.
2.4 Resistance determinants and integrative and conjugative elements (ICE)
Acquired resistance genes were screened using the AMRFinder+ v3.11.18.6 Resistance-associated mutations and mobile genetic elements were analyzed with Geneious R9 using H. parainfluenzae T3T1 (NC_015964) as a reference genome. ICE structures were examined using ICEHpaHUB6 and ICEHpaHUB7 as references (Saiz-Escobedo et al., 2023).
2.5 Plasmid description
Potential plasmids were screened with the PlasmidFinder tool7 of the Center for Genomic Epidemiology. Plasmid sequences were annotated using PROKKA8 to predict ORFs and functional elements. Homologous sequences were identified by BLASTx and BLASTn searches against public databases. Plasmid structures and virulence genes were analyzed and curated using Geneious R9 (version 9.1.7, Biomatters). For graphical representation, plasmid structures were visualized with Geneious R9, GC content was represented using Prok(see proksee.ca), and the final figure was refined with Inkscape.9
2.6 Ethics
This study complied with the Declaration of Helsinki principles from the World Medical Association and was approved by the Clinical Research Ethics Committee of Bellvitge University Hospital (PR075/21). Written informed consent was not required as this was a retrospective observational study with isolates obtained as part of routine microbiological testing. Patient confidentiality was maintained by anonymizing all personal data in accordance with Spanish legal regulations (LOPD 15/1999 and RD 1720/2007). Biological sample management adhered to Law 14/2007 on Biomedical Research.
3 Results
3.1 Clinical strains and patient background
HUB-HP17268 and HUMT-HP05 were isolated from genital ulcer swabs taken from two young men with multiple previous STIs, including chlamydia, gonorrhea, syphilis, and genital mycoplasma. Both patients lacked symptoms such as abdominal pain, diarrhea, anal bleeding, dysuria, or urogenital discharge and consulted for the mentioned genital ulcers.
Patient 1 (HUB): 34-year-old MSM with positive serology for Treponema pallidum (RPR titer of 1:8). HIV-positive undergoing antiretroviral therapy. In the current episode, the patient tested positive by PCR for Chlamydia trachomatis and Mycoplasma genitalium and was treated with a combination of intramuscular ceftriaxone and oral doxycycline.
Patient 2 (HUMT): 28-year-old MSM with positive serology for T. pallidum (RPR titer of 1:16), with adenopathy and a history of multiple STIs. This patient had been enrolled in a pre-exposure prophylaxis (PrEP) program since May 2022. In the 6 months preceding this episode, he received intramuscular ceftriaxone for gonococcal urethritis. In this episode, he was diagnosed with syphilis and treated with intramuscular penicillin G benzathine.
3.2 New capsular operon related to Haemophilus sputorum HSPU_type1
A novel capsular operon was identified by WGS as part of our hospital surveillance and was designated as HPAR_type4 (NCBI accession number PV567605). This operon was different from those previously described in H. parainfluenzae, with a total length of 11,885 bp and 9 predicted ORFs (Figure 1), following the typical genetic organization of Haemophilus spp. cap loci, comprising three regions (I, II, and III).
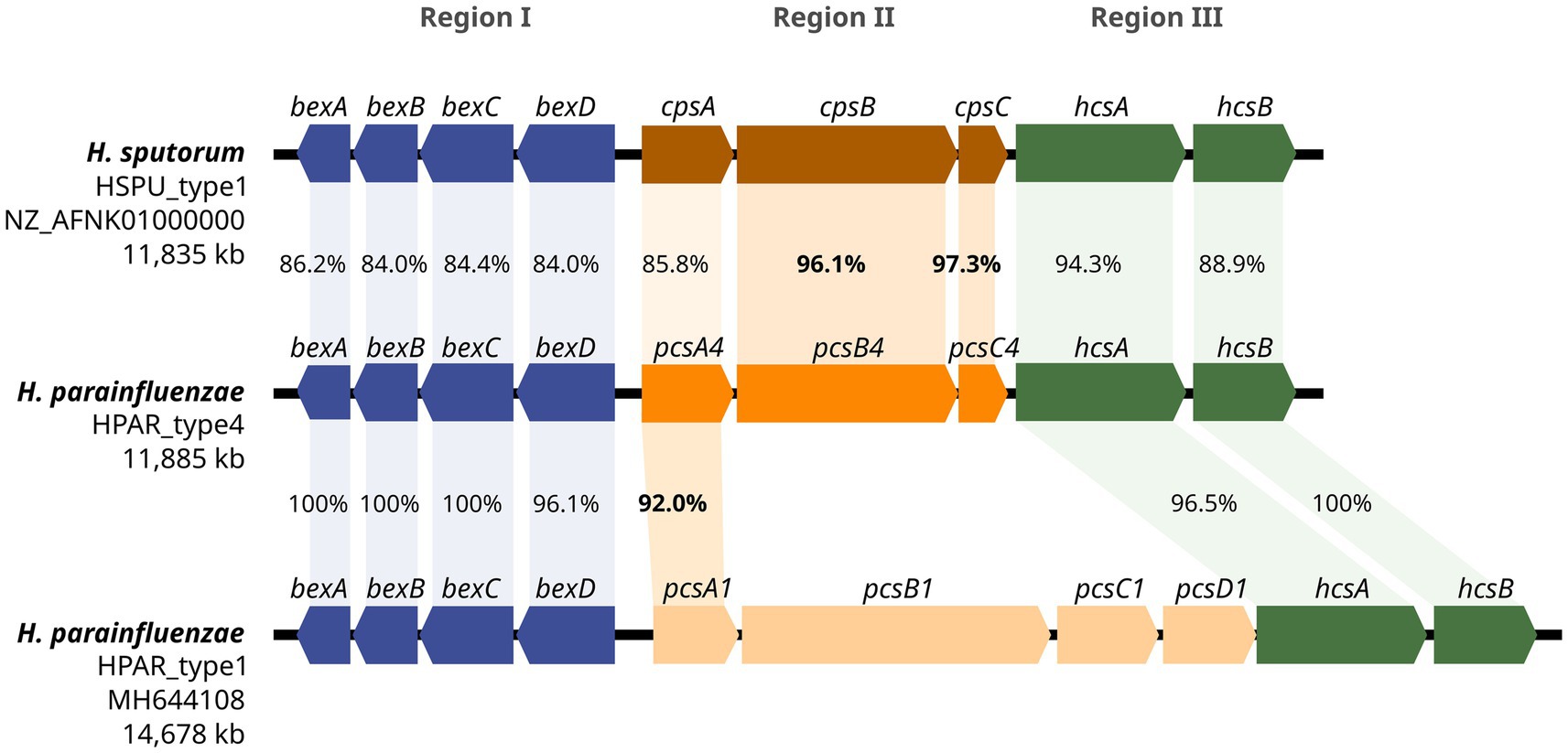
Figure 1. Genetic structure of the H. parainfluenzae HPAR_type4 operon. A comparison of the genetic organization between H. parainfluenzae HPAR_type4, H. sputorum HSPU_type1 and H. parainfluenzae HPAR_type1 operons is presented. The three main regions are represented using different colors, and the percentages of gene identity are displayed.
Regions I (bexABCD) and III (hcsAB) showed 96–100% homology to the corresponding regions in H. parainfluenzae HPAR_type1 (Supplementary Table S1). The detected ORFs were translated into protein sequences, with 98–100% identity to HPAR_type1. The serotype-specific region II contained three ORFs. The first ORF was homologous to the pscA gene found in H. parainfluenzae capsular operons, while the other two ORFs showed >96% identity to the cpsB and cpsC genes of H. sputorum HSPU_type1, encoding a glycosyltransferase and an acetyltransferase, respectively. We designated these genes as pscA4 (1,101 bp), pcsB4 (2,628 bp), and pcsC4 (621 bp).
3.3 Antimicrobial resistance
Both strains exhibited an extensively drug-resistant phenotype. HUB-HP17268 was resistant to all tested β-lactams except ceftriaxone and carbapenems (imipenem and meropenem), whereas HUMT-HP05 was resistant to all tested β-lactams except for the carbapenems. Both strains were also resistant to macrolides, fluoroquinolones, chloramphenicol, trimethoprim-sulfamethoxazole and tetracycline (Table 1).
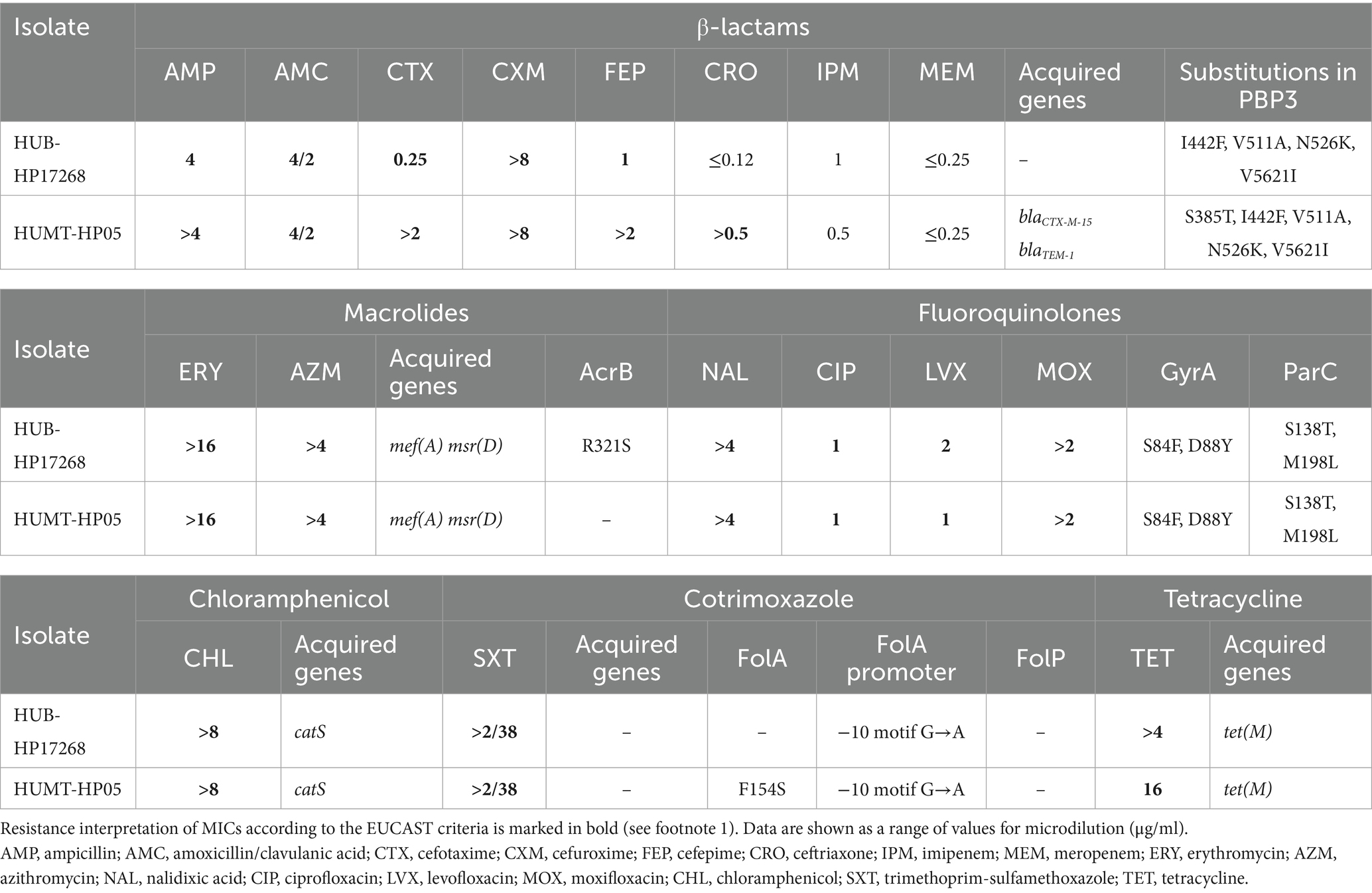
Table 1. MICs and molecular resistance mechanisms of H. parainfluenzae HPAR_type4 strains.
To further characterize the resistance mechanisms, we analyzed the genetic determinants in the genomes of both strains. Resistance to ceftriaxone in HUMT-HP05 led to the identification of the recently described CTX-M-15 extended spectrum β-lactamase, which was responsible for the high minimum inhibitory concentrations (MICs) for cephalosporins. The blaCTX-M-15 gene was inserted in a mobile element, showing >99% identity with the previously described ICEHpaHUB6. This strain also had a TEM-1 and resistance-associated modifications in PBP3 (i.e., S385T, I442F, V511A, N526K, V5621I) that contribute to β-lactam resistance. In HUB-HP17268, β-lactam resistance was only attributed to modifications in PBP3. Both strains had fluoroquinolones resistance caused by amino acid substitutions in GyrA (S84F and D88Y) and ParC (S138T and M198L). Macrolide and tetracycline resistance was conferred by the insertion of the tet(M)-MEGA element carrying the mef(E), msr(D) and tet(M) genes. Chloramphenicol resistance was due to the acquisition of catS, and trimethoprim-sulfamethoxazole resistance was attributed to mutations in the folA promoter (−10 motif [G > A]).
3.4 Genomic comparison
The genome of HUMT-HP05 consisted of 2,213,506 base pairs (bp) with a GC content of 39.5%, while the genome of HUB-HP17268 was slightly smaller, with 2,155,329 bp and a GC content of 39.6%. A total of 2,090,090 bp were shared between both genomes (94.4%). Within these shared regions, 17 recombinant blocks were identified, and only 42 SNPs were detected outside of these blocks. The presence of the ICEHpaHUB6 mobile genetic element in HUMT-HP05 accounted for 58,725 bp of the genomic differences.
3.5 Plasmid characterization
In addition to the novel capsular operon, each strain carried a plasmid, previously unreported in H. parainfluenzae (Figures 2, 3) that was undetected by PlasmidFinder but identified through long-read sequencing.
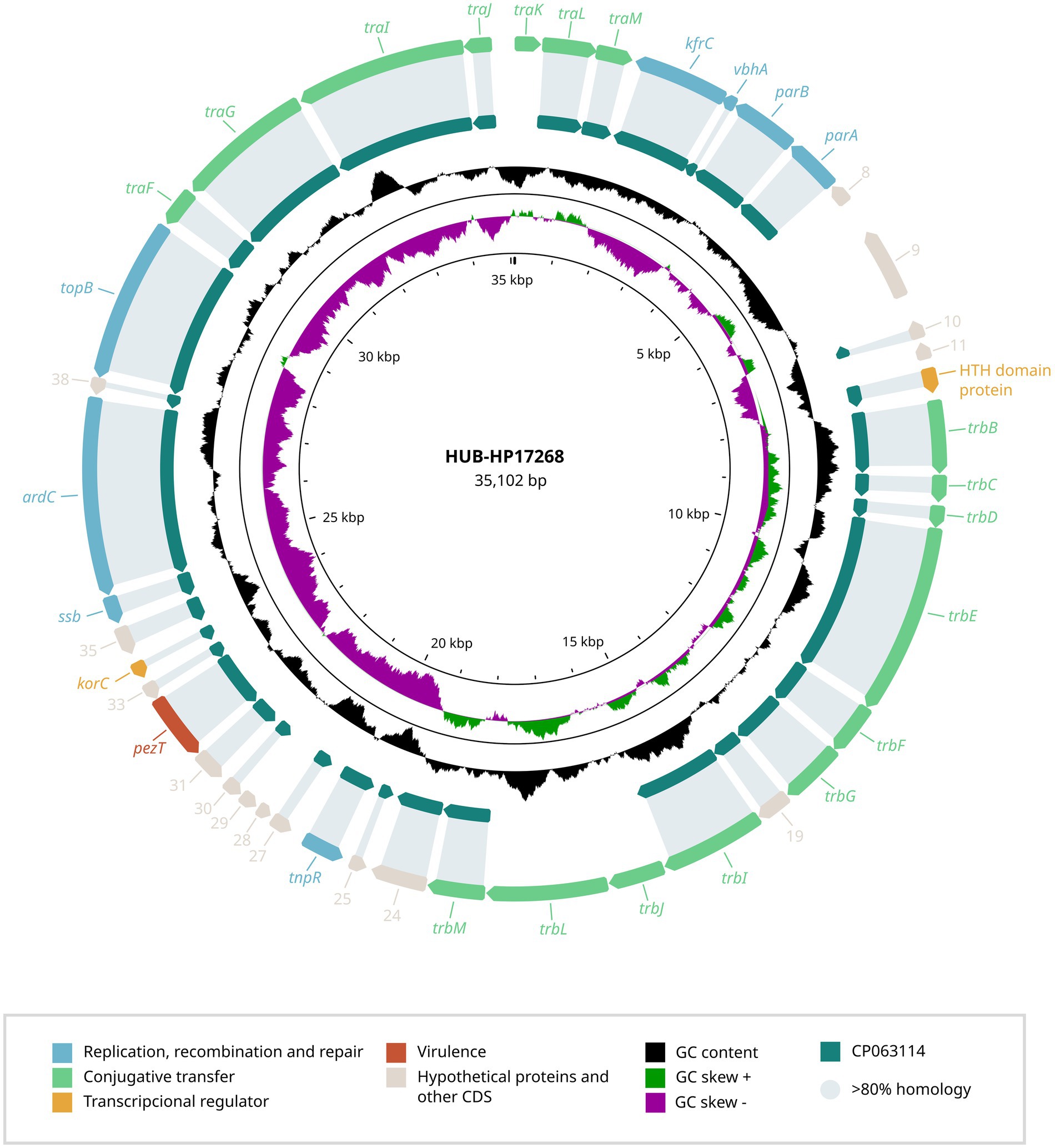
Figure 2. Genetic structure of the plasmid identified in H. parainfluenzae HUB-HP17268. The genetic organization of the HUB-HP17268 plasmid is shown, highlighting ORFs associated with replication, recombination and repair, conjugative transfer, transcriptional regulation, virulence, and hypothetical proteins. The GC content is also presented. A comparison with the most closely related plasmid found in H. parainfluenzae strain M1C147 (CP063114.1) is provided, displaying the percentage of gene identity.
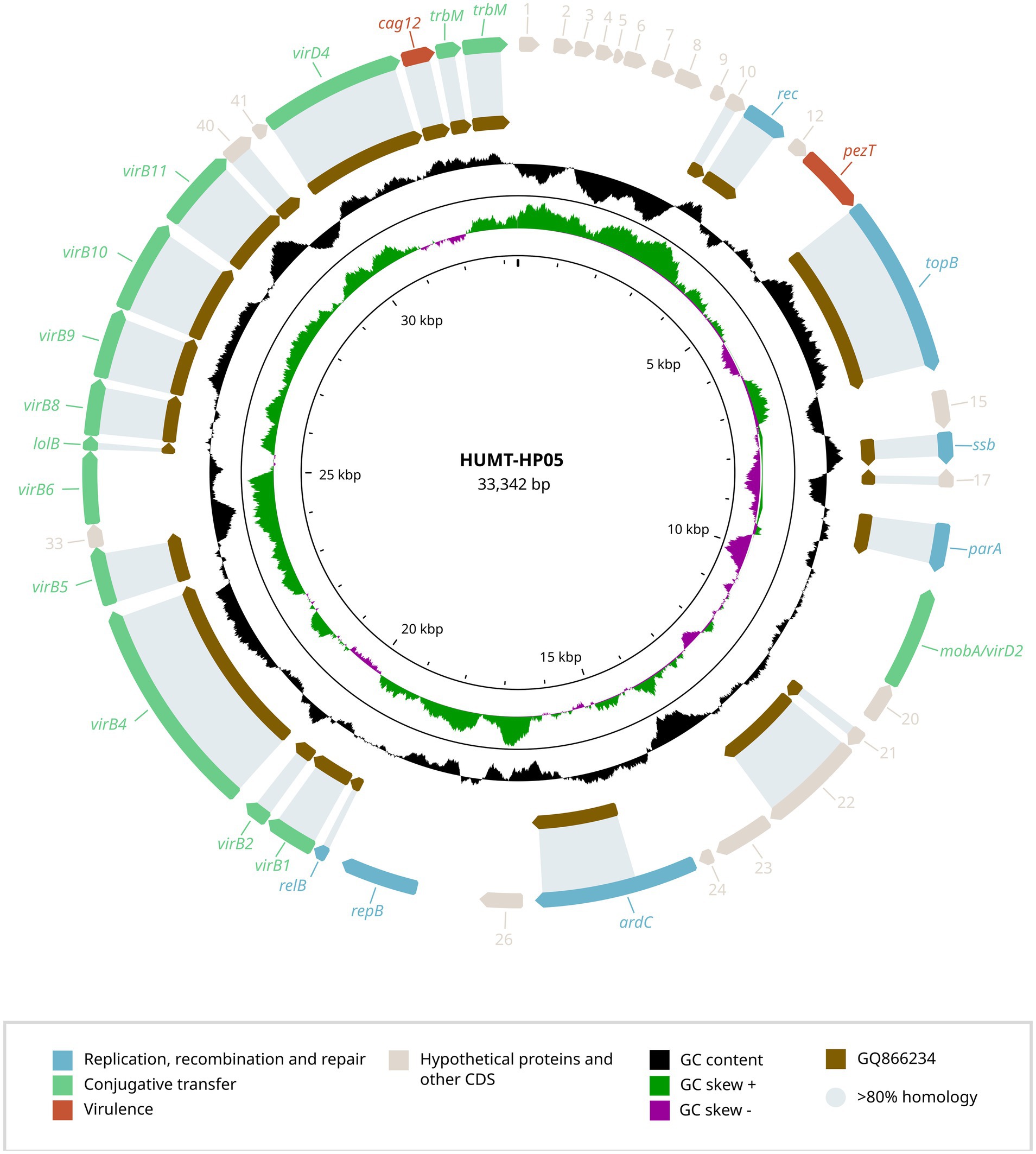
Figure 3. Genetic structure of the plasmid identified in H. parainfluenzae HUMT-HP05. The genetic organization of the HUMT-HP05 plasmid is shown, highlighting ORFs associated with replication, recombination and repair, conjugative transfer, virulence, and hypothetical proteins. The GC content is also presented. A comparison with the most closely related plasmid found in A. actinomycetemcomitans strain D11S-1 (GQ866234.1) is provided, displaying the percentage of gene identity.
To better understand these genetic elements, we conducted BLAST analyses of the plasmid sequences and their predicted ORFs against the NCBI public database, aiming to identify potential homologs and functional annotations. The plasmid carried by HUB-HP17268 (NCBI accession number PV567603) was 35,102 bp in length with a GC content of 33.0% and shared 89.0% identity with a plasmid from H. parainfluenzae strain M1C147 (CP063114.1). In contrast, HUMT-HP05 (NCBI accession number PV567604) harbored a plasmid of 33,342 bp with a GC content of 32.8%, which showed 89.1% homology with plasmid S25 from Aggregatibacter actinomycetemcomitans strain D11S-1 (GQ866234.1).
The complete annotation of predicted ORFs from both plasmids (Supplementary Table S2) identified genes strongly associated with plasmid replication, recombination and repair, including repA (replication protein), parA and parB (partitioning proteins), topB (DNA topoisomerase), and ssb (single-stranded DNA-binding protein). Neither of the plasmids contained resistance genes, but both harbored virulence-associated genes, such as pezT.
4 Discussion
The identification of a novel capsular operon, HPAR_type4, in two H. parainfluenzae strains isolated from genital ulcers in MSM patients expands our understanding of the genetic diversity of this species. This capsular operon was identified in two H. parainfluenzae strains isolated from different hospitals, with the episodes occurring 2 years apart. While there is no evidence suggesting direct contact between the patients, which initially appears to exclude a direct epidemiological link, the genetic similarity between the strains, sharing 96.7% homology, raises questions about potential common sources or transmission pathways. The absence of a standardized clonal classification system for H. parainfluenzae complicates the assessment of the epidemiological relationship between these strains. Further investigation is needed to determine if these cases represent an emerging trend or are isolated events.
Continuous genomic monitoring led to the detection of the first capsular operon in H. parainfluenzae (González-Díaz et al., 2019). Five years after its publication, four different capsular operons have already been found in this species (González-Díaz et al., 2019; Sierra et al., 2021; Bajanca-Lavado et al., 2022), including the HPAR_type4 operon described in this study. The presence of capsules in this genus may enhance their ability to evade host immune responses, as demonstrated in H. influenzae, where capsulated strains often exhibit increased pathogenicity (Guellil et al., 2022). Although the capsular operons found in H. parainfluenzae differ structurally from the virulent H. influenzae type b capsule, the notable capacity of this species to acquire new genetic material raises concerns about the potential emergence of similarly virulent capsules in the future (Sierra et al., 2021). Thus, its surveillance is important to monitor and respond to any emerging virulence factors.
The genetic organization of the capsular operon identified in HUB-HP17268 and HUMT-HP05 aligns with other Haemophilus spp. capsular loci, comprising three regions. The high homology of regions I and III with the previously characterized H. parainfluenzae HPAR_type1 operon (González-Díaz et al., 2019), indicates a conserved mechanism for capsular polysaccharide biosynthesis and export within the genus. The serotype-specific region II contains ORFs similar to genes from both H. parainfluenzae HPAR_type1 (pcsA) and H. sputorum HSPU_type1 (cpsB and cpsC), suggesting that horizontal gene transfer and recombination may play a role in the evolution of capsular diversity among Haemophilus species (Nielsen et al., 2015). Both strains were isolated from distinct Catalan towns over a 2-year period (without epidemiological connections), which excludes definitive conclusions about clonal transmission. However, the 17 recombinant blocks and the acquisition of CTX-M-15-bearing ICEHpaHUB6 in HUMT-HP05 demonstrate the capacity of H. parainfluenzae for genomic recombination.
Since the first report of a capsular H. parainfluenzae strain, we have observed an alarming increase in antibiotic resistance. Both strains in our study exhibited XDR profiles, including resistance to β-lactams and other broad-spectrum antibiotics. This trend is consistent with our previous observations of capsulated MDR and XDR strains (Sierra et al., 2021). Interestingly, the first XDR strain described in the literature also carried an HPAR_type1 operon. We were able to identify this operon through genomic analysis of the sequence deposited in NCBI, as this detail was not reported in the original article (Tinguely et al., 2013). Since then, ESBL production has also been identified in this species. Initially, the blaCTX-M-15 gene was detected in four strains at our hospital (Saiz-Escobedo et al., 2023). Subsequently, another strain (ASM3628892v1) carrying the same resistance determinant was identified in France, with the particularity that this strain also carried the H. parainfluenzae HPAR_type1 operon (Caméléna et al., 2024). The identification of the CTX-M-15 ESBL in HUMT-HP05 highlights the ability of H. parainfluenzae to acquire resistance genes through mobile genetic elements (Juhas et al., 2007). The insertion of blaCTX-M-15 into the ICEHpaHUB6 element may facilitate the spread of this resistance determinant within the species and among other bacteria co-colonizing the urogenital tract. Its detection in an MSM patient further underscores the importance of considering this pathogen in the context of STI management, especially in cases of co-infection with other pathogens, as it may serve as a reservoir for resistance genes.
To date, only transposons and ICE structures had been detected integrated into the H. parainfluenzae genome (Sierra et al., 2021). This study identified two previously unreported plasmids, both carrying genes essential for plasmid maintenance, replication and segregation, indicating their potential for propagation within host cells (Smillie et al., 2010). Among them, we identified a plasmid in H. parainfluenzae with significant homology to a plasmid from A. actinomycetemcomitans, a member of the same Pasteurellaceae family. This finding highlights the potential for horizontal transfer within this taxonomic group, which could contribute to the spread of adaptive traits, including antimicrobial resistance. Horizontal gene transfer is common in Pasteurellaceae, and aligns with recent mobilome analysis, which highlights the role of mobile genetic elements (including plasmids, transposons and integrons) in shaping the resistome of these pathogens (da Silva et al., 2022). Although none of the plasmids carried antibiotic resistance genes, unlike many clinically relevant plasmids (Watts et al., 2021), this does not preclude their potential to acquire such genes. Recently, several small plasmids have been detected in Haemophilus spp., carrying genes coding for the β-lactamases TEM-1 and TEM-40 (Watts et al., 2021). In this case, our strains were already XDR and had all the resistance determinants integrated in the genome (even the β-lactamase TEM-1 in the case of HUMT-HP05). However, the presence of these plasmids may help transfer these resistance determinants to other bacterial strains. Antimicrobial resistance genes can be incorporated through recombination events, such as the insertion of transposable elements (insertion sequences, transposons) or integrons carrying resistance determinants (Carattoli, 2013). The presence of recombination-associated genes on these plasmids suggest they have the machinery to facilitate such genetic exchanges, consistent with the role of plasmids as genetic reservoirs that can acquire and disseminate resistance genes under selective pressures (Carattoli, 2013).
In conclusion, this study highlights the genetic diversity of H. parainfluenzae and the emergence of multidrug-resistant strains in urogenital infections. The identification of new capsular operons enhancing virulence in XDR isolates, and the acquisition of resistance genes like the blaCTX-M-15, underscores the evolving threat of this pathogen. While these genomic findings are compelling, functional studies are needed to confirm the contribution of these capsular operons to pathogenicity. Nevertheless, these findings raise concerns about transmission, particularly in MSM populations, and emphasize the need for ongoing genomic surveillance to monitor the spread and pathogenic potential of these strains.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Ethics statement
The studies involving humans were approved by Clinical Research Ethics Committee of Bellvitge University Hospital. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and institutional requirements.
Author contributions
LS-E: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Visualization, Writing – original draft, Writing – review & editing. MB-T: Writing – review & editing. MR: Writing – review & editing. MG-E: Writing – review & editing. LG: Writing – review & editing. IC-J: Writing – review & editing. RC: Writing – review & editing. FT: Writing – review & editing. DS-R: Writing – review & editing. MD: Writing – review & editing. KX: Writing – review & editing. PH: Writing – review & editing. CA: Writing – review & editing. AG-D: Data curation, Formal analysis, Investigation, Methodology, Supervision, Validation, Writing – review & editing. SM: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was funded by the Fondo de Investigaciones Sanitarias (PI22/00257 to SM); Centro de Investigación Biomédica en Red de Enfermedades Respiratorias (CIBERES; CB06/06/0037) and Centro de Investigación Biomédica en Red de Enfermedades Infecciosas (CB21/13/0009), an initiative of the Instituto de Salud Carlos III (ISCIII). The European Regional Development Fund/European Social Fund (ERDF/ESF; “Investing in your future”) also provided financial support, and CERCA Program/Generalitat de Catalunya provided institutional support. SM was supported by Miguel Servet contract (CP19/00096) (ISCIII).
Acknowledgments
We would like to thank the staff of the Microbiology Laboratory of the Hospital Universitari de Bellvitge for their contribution to this project. We are also grateful to Catlab and the Hospital Universitari Mútua de Terrassa for providing and allowing us to study one of the bacterial strains essential for this work. Their support was invaluable for the completion of this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1659538/full#supplementary-material
Footnotes
1. ^www.eucast.org/clinical_breakpoints
2. ^github.com/bactopia/bactopia
3. ^github.com/rrwick/Unicycler
4. ^github.com/tseemann/snippy
7. ^cge.food.dtu.dk/services/PlasmidFinder/
References
Bajanca-Lavado, M. P., Cavaco, L., Fernandes, M., Touret, T., Candeias, C., Simões, A. S., et al. (2022). Haemophilus influenzae carriage among healthy children in Portugal, 2015-2019. Microorganisms 10:1964. doi: 10.3390/microorganisms10101964
Baron, J. E., Shamrock, A. G., Volkman, A., and Westermann, R. W. (2020). Haemophilus parainfluenzae septic arthritis following primary all-inside meniscus repair: a case report and review of the literature. Iowa Orthop. J. 40:111.
Caméléna, F., Merimèche, M., Liberge, M., Maubaret, C., Donay, J. L., Taha, M. K., et al. (2024). Detection of CTX-M-15 ESBL in XDR Haemophilus parainfluenzae from a urethral swab. J. Antimicrob. Chemother. 79, 539–545. doi: 10.1093/jac/dkad408
Carattoli, A. (2013). Plasmids and the spread of resistance. Int. J. Med. Microbiol. 303, 298–304. doi: 10.1016/j.ijmm.2013.02.001
da Silva, G. C., Gonçalves, O. S., Rosa, J. N., França, K. C., Bossé, J. T., Santana, M. F., et al. (2022). Mobile genetic elements drive antimicrobial resistance gene spread in Pasteurellaceae species. Front. Microbiol. 12:773284. doi: 10.3389/fmicb.2021.773284
Ducours, M., Puges, M., Desclaux, A., Barthod, L., Peuchant, O., and Cazanave, C. (2020). Haemophilus spp., an emerging multidrug-resistant sexually transmitted pathogen. Med. Mal. Infect. 50, 734–737. doi: 10.1016/j.medmal.2020.06.003
González-Díaz, A., Tubau, F., Pinto, M., Sierra, Y., Cubero, M., Càmara, J., et al. (2019). Identification of polysaccharide capsules among extensively drug-resistant genitourinary Haemophilus parainfluenzae isolates. Sci. Rep. 9:4481. doi: 10.1038/s41598-019-40812-2
Guellil, M., Keller, M., Dittmar, J. M., Inskip, S. A., Cessford, C., Solnik, A., et al. (2022). An invasive Haemophilus influenzae serotype b infection in an Anglo-Saxon plague victim. Genome Biol. 23:22. doi: 10.1186/s13059-021-02580-z
Hsu, M. S., Wu, M. Y., Lin, T. H., and Liao, C. H. (2015). Haemophilus parainfluenzae urethritis among homosexual men. J. Microbiol. Immunol. Infect. 48, 450–452. doi: 10.1016/j.jmii.2012.11.007
Juhas, M., Power, P. M., Harding, R. M., Ferguson, D. J. P., Dimopoulou, I. D., Elamin, A. R. E., et al. (2007). Sequence and functional analyses of Haemophilus spp. genomic islands. Genome Biol. 8:R237. doi: 10.1186/gb-2007-8-11-r237
Kosikowska, U., Biernasiuk, A., Rybojad, P., Łoś, R., and Malm, A. (2016). Haemophilus parainfluenzae as a marker of the upper respiratory tract microbiota changes under the influence of preoperative prophylaxis with or without postoperative treatment in patients with lung cancer. BMC Microbiol. 16:62. doi: 10.1186/s12866-016-0679-6
Magdaleno-Tapial, J., Valenzuela-Oñate, C., Giacaman-von der Weth, M. M., Ferrer-Guillén, B., Martínez-Domenech, A., García-Legaz Martínez, M., et al. (2019). Haemophilus species isolated in urethral exudates as a possible causative agent in acute urethritis: a study of 38 cases. Actas Dermo-Sifiliograficas 110, 38–42. doi: 10.1016/j.adengl.2018.11.011
Nielsen, S. M., de Gier, C., Dimopoulou, C., Gupta, V., Hansen, L. H., and Nørskov-Lauritsen, N. (2015). The capsule biosynthesis locus of Haemophilus influenzae shows conspicuous similarity to the corresponding locus in Haemophilus sputorum and may have been recruited from this species by horizontal gene transfer. Microbiology 161, 1182–1188. doi: 10.1099/mic.0.000081
Nørskov-Lauritsen, N. (2014). Classification, identification, and clinical significance of Haemophilus and Aggregatibacter species with host specificity for humans. Clin. Microbiol. Rev. 27, 214–240. doi: 10.1128/CMR.00103-13
Potts, C. C., Topaz, N., Rodriguez-Rivera, L. D., Hu, F., Chang, H. Y., Whaley, M. J., et al. (2019). Genomic characterization of Haemophilus influenzae: a focus on the capsule locus. BMC Genomics 20, 1–9. doi: 10.1186/s12864-019-6145-8
Revest, M., Egmann, G., Cattoir, V., and Tattevin, P. (2016). HACEK endocarditis: state-of-the-art. Expert Rev. Anti-Infect. Ther. 14, 523–530. doi: 10.1586/14787210.2016.1164032
Saiz-Escobedo, L., Cadenas-Jiménez, I., Olmos, R., Carrera-Salinas, A., Berbel, D., Càmara, J., et al. (2023). Detection of Bla CTX-M-15 in an integrative and conjugative element in four extensively drug-resistant Haemophilus parainfluenzae strains causing urethritis. Int. J. Antimicrob. Agents 62:106991. doi: 10.1016/j.ijantimicag.2023.106991
Sierra, Y., González-Díaz, A., Carrera-Salinas, A., Berbel, D., Vázquez-Sánchez, D. A., Tubau, F., et al. (2021). Genome-wide analysis of urogenital and respiratory multidrug-resistant Haemophilus parainfluenzae. J. Antimicrob. Chemother. 76, 1741–1751. doi: 10.1093/jac/dkab109
Smillie, C., Garcillán-Barcia, M. P., Francia, M. V., Rocha, E. P. C., and de la Cruz, F. (2010). Mobility of plasmids. Microbiol. Mol. Biol. Rev. 74, 434–452. doi: 10.1128/MMBR.00020-10
Tinguely, R., Seiffert, S. N., Furrer, H., Perreten, V., Droz, S., and Endimiani, A. (2013). Emergence of extensively drug-resistant Haemophilus parainfluenzae in Switzerland. Antimicrob. Agents Chemother. 57, 2867–2869. doi: 10.1128/AAC.00221-13
Watts, S. C., and Holta, K. E. (2019). HICAP: in silico serotyping of the Haemophilus influenzae capsule locus. J. Clin. Microbiol. 57:10-1128. doi: 10.1128/jcm.00190-19
Watts, S. C., Judd, L. M., Carzino, R., Ranganathan, S., and Holt, K. E. (2021). Genomic diversity and antimicrobial resistance of Haemophilus colonizing the airways of young children with cystic fibrosis. mSystems 6:10-1128. doi: 10.1128/msystems.00178-21
Keywords: Haemophilus parainfluenzae , capsular operon, antimicrobial resistance, conjugative plasmid, whole-genome sequencing, extended-spectrum β-lactamase, STIs, genomic surveillance
Citation: Saiz-Escobedo L, Ballestero-Tellez M, Rajadell M, Garreta-Esteban M, Gisbert L, Cadenas-Jiménez I, Costa RM, Tubau F, Sánchez-Ramos D, Domínguez MA, Xanthopoulou K, Higgins PG, Ardanuy C, González-Díaz A and Marti S (2025) A novel capsular operon and potentially conjugative plasmids in extensively drug-resistant urogenital Haemophilus parainfluenzae. Front. Microbiol. 16:1659538. doi: 10.3389/fmicb.2025.1659538
Edited by:
Shangshang Qin, Zhengzhou University, ChinaReviewed by:
Sandip Kumar Mukherjee, University of California, United StatesGiarlã Cunha da Silva, Universidade Federal de Viçosa, Brazil
Copyright © 2025 Saiz-Escobedo, Ballestero-Tellez, Rajadell, Garreta-Esteban, Gisbert, Cadenas-Jiménez, Costa, Tubau, Sánchez-Ramos, Domínguez, Xanthopoulou, Higgins, Ardanuy, González-Díaz and Marti. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aida González-Díaz, YWdvbnphbGV6ZEBiZWxsdml0Z2Vob3NwaXRhbC5jYXQ=; Sara Marti, c21hcnRpbm1AYmVsbHZpdGdlaG9zcGl0YWwuY2F0