 Wojciech Popowski1
Wojciech Popowski1 Dominika Domanowska1Damian Koseski1Rafał Ostrowski2
Dominika Domanowska1Damian Koseski1Rafał Ostrowski2 Magdalena Zalewska2Milena Małecka-Giełdowska3
Magdalena Zalewska2Milena Małecka-Giełdowska3 Anna Łasica4
Anna Łasica4 Magdalena Popowska2*
Magdalena Popowska2*- 1Department of Oral Surgery, Medical University of Warsaw, Warsaw, Poland
- 2Department of Bacterial Physiology, Institute of Microbiology, Faculty of Biology, University of Warsaw, Warsaw, Poland
- 3Department of Laboratory Medicine, Medical University of Warsaw, Warsaw, Poland
- 4Department of Bacterial Genetics, Institute of Microbiology, Faculty of Biology, University of Warsaw, Warsaw, Poland
Platelet-rich fibrin (PRF) is a platelet concentrate widely applied in various medical fields and is considered a valuable adjunct in tissue regeneration during surgical procedures. However, infections caused by biofilm-forming bacteria at surgical sites, combined with increasing antibiotic resistance, present a major clinical concern. Current research is focused on identifying alternative therapeutic strategies to improve infection control and promote wound healing. This study aimed to characterize the oral microbiome of healthy individuals and evaluate the in vitro antimicrobial properties of two PRF formulations. The antibacterial activity, along with its temporal dynamics at different initial bacterial concentrations, was assessed against Gram-negative bacteria (Escherichia coli, Porphyromonas gingivalis) and Gram-positive bacteria exhibiting diverse morphologies (Bacillus subtilis, Micrococcus luteus, Staphylococcus lentus, Enterococcus casseliflavus, Streptococcus mutans). Our results fill gaps in knowledge concerning the spectrum of PRF’s antimicrobial activity, demonstrating efficacy against a range of opportunistic and pathogenic bacteria. Key findings include the absence of significant differences in oral microbiome composition between male and female participants, a lack of inhibitory effect of A-PRF against S. mutans, and a transient inhibitory effect against P. gingivalis observed only at low initial OD₆₀₀ and within 24 h. These findings indicate that A-PRF therapy alone may not provide a sufficiently effective antibacterial effect in patients with oral infections, and that alternative or adjunctive therapeutic approaches should be considered in such cases.
1 Introduction
Platelet concentrates are blood-derived products obtained by fractionating plasma through centrifugation using specialized equipment. By adjusting centrifugation parameters, various types of platelet concentrates can be produced, which are classified based on their cellular composition and the architecture of the fibrin matrix (Sanz et al., 2019; Pereira et al., 2023).
The development of platelet concentrate preparations dates back to the 1970s, when Matras conducted pioneering research on fibrin glues to enhance wound healing in rat skin. In the following years, several studies proposed improved methods for utilizing blood-derived preparations, achieving a higher concentration of platelets in the final product. These techniques represented the first generation of platelet-rich plasma (PRP) gels. The application of these preparations yielded promising outcomes in fields such as ophthalmology, neurosurgery, and general surgery (Dohan Ehrenfest et al., 2014). Over time, an alternative form of platelet concentrate, termed platelet-rich fibrin (PRF), was developed (Choukroun et al., 2006; Dohan Ehrenfest et al., 2014). It is free of anticoagulants, thereby eliminating the adverse effects observed with earlier platelet concentrates. Variants within this category include leukocyte- and platelet-rich fibrin (L-PRF), advanced platelet-rich fibrin (A-PRF), and injectable platelet-rich fibrin (i-PRF) (Pereira et al., 2023). Each type of PRF requires specific centrifugation parameters (Clark et al., 2018; Pereira et al., 2023; Farshidfar et al., 2025).
Platelet-rich fibrin contains a variety of biologically active molecules, including insulin-like growth factor 1 (IGF-1), platelet-derived growth factor (PDGF), vascular endothelial growth factor (VEGF), fibroblast growth factor (FGF), epidermal growth factor (EGF), platelet-derived epidermal growth factor (PDEGF), transforming growth factor beta (TGF-β), as well as proteins of the fibrin matrix, all of which are present at higher concentrations than in peripheral blood (Bagdadi et al., 2019; Pereira et al., 2023). Among these, PDGF, VEGF, and TGF-β play essential roles in angiogenesis and the formation of new tissue structures (Schär et al., 2015; Bagdadi et al., 2019). The platelets within PRF contain granules loaded with cytokines, chemokines, and other inflammatory mediators that, upon release, enhance hemostasis and promote the activation and recruitment of cells to sites of inflammation. Furthermore, leukocytes present in PRF contribute to both angiogenesis and lymphangiogenesis through intercellular interactions and the expression of various signaling molecules (Bagdadi et al., 2019). Moreover, PRF exhibits osteoinductive, anti-inflammatory, pro-angiogenic, and antibacterial properties (Liu et al., 2019; Feng et al., 2020; Jasmine et al., 2020). Platelet concentrates are widely utilized to induce and stimulate tissue repair and regeneration processes (Pereira et al., 2023).
In recent years, autologous platelet concentrates have been widely used in dentistry, maxillofacial surgery, and plastic surgery (Fabbro et al., 2016; Jasmine et al., 2020). PRF has been applied in various procedures, including maxillary sinus augmentation, bone defect regeneration around implants, guided bone regeneration (GBR), healing of post-extraction sockets, healing of bone defects following enucleation of extensive periapical cysts, temporomandibular joint disorder treatment, healing of wounds in patients after oral cancer resection surgery, and periodontal surgery (Choukroun et al., 2006; Simon et al., 2009; Jang et al., 2010; Simonpieri et al., 2012; Castro et al., 2017; Liu et al., 2019; Miron et al., 2019; Xu et al., 2023; Long et al., 2024). The use of PRF in sinus-lift procedures prior to planned implant placement can reduce healing time to 4 months; however, larger-scale studies are still needed to confirm this effect (Choukroun et al., 2006). Current evidence supporting the necessity of adding PRF to bone grafts in sinus-lift procedures is limited (Liu et al., 2019). PRF appears to be more effective in alleviating pain in patients with temporomandibular joint disorders, but additional research is required to fully determine the efficacy of such treatment (Xu et al., 2023). The application of PRF in patients undergoing oral cancer resection surgery seems promising. In these patients, PRF improves tissue regeneration, reduces postoperative discomfort, and enhances treatment outcomes. Despite encouraging findings, further high-quality, randomized, controlled clinical trials are needed (Long et al., 2024). PRF has demonstrated a beneficial effect in reducing pain, swelling, and the incidence of osteitis following the extraction of impacted lower third molars (Xiang et al., 2019).
The regenerative activity of PRF after surgical events is certainly beneficial for patients however, another condition must be addressed at the same time, which are bacterial infections. An increasing incidence of postoperative staphylococcal infections has been observed, leading to prolonged hospitalizations (Līduma et al., 2012; Jasmine et al., 2020). I-PRF, due to its content of proteins with antibacterial properties, may contribute to reducing the risk of such infections (Jasmine et al., 2020). In vitro studies available in the literature indicate that both L-PRF and high-density PRF (H-PRF) exert antibacterial activity against Staphylococcus aureus and Escherichia coli strains (Feng et al., 2020).
Another group of bacteria which may contribute to numerous diseases including post-treatment complications is the oral microbiome. The human oral cavity comprises diverse habitats—including the inner cheeks, palate, tongue, and teeth—each supporting distinct microbial communities (Morrison et al., 2023). The oral microbiome is the second most diverse microbial community after the gut and includes bacteria, viruses, fungi, protozoa, and archaea (Benn et al., 2018; Morrison et al., 2023). Among these, bacteria are the most extensively studied and are found both in saliva and on oral surfaces such as mucosa, tongue, and teeth (Morrison et al., 2023). At the phylum level, a healthy oral microbiome is dominated by Actinobacteria, Fusobacteria, Proteobacteria, Firmicutes, and Bacteroidetes, which together account for approximately 96% of all oral bacteria (Verma et al., 2018; Nearing et al., 2020). Fusobacteria and Bacteroidetes are commonly cultivable, with Fusobacteria being among the most prevalent (Nearing et al., 2020; Morrison et al., 2023). On the genus level, the microbiome is relatively stable; 11 genera are shared by over 99% of individuals and collectively make up about 77.8% of the total microbial abundance. These dominant genera include Streptococcus, Prevotella, Veillonella, Lactobacillus, Actinomyces, and Neisseria (Morrison et al., 2023). While most individuals share similar genera, species- and strain-level diversity remains highly individual-specific (Nearing et al., 2020).
Fungi also constitute a notable component of the oral microbiome. Over 100 fungal species have been identified in healthy individuals (Ghannoum et al., 2010), with Candida spp. being the most prevalent, frequently contributing to early stages of biofilm formation (Janus et al., 2017).
The diversity and stability of the oral microbiome are essential for oral and systemic health. Dysbiosis has been linked to conditions such as caries, periodontitis, and systemic diseases (Verma et al., 2018; Morrison et al., 2023). Periodontal disease arises from pathogenic biofilms that trigger gingival inflammation. Notably, certain members of the core microbiome, albeit in low abundance in healthy people,—Porphyromonas gingivalis, Tannerella forsythia, and Treponema denticola—form the so-called “red complex” and are strongly associated with periodontitis (Ximénez-Fyvie et al., 2000; Verma et al., 2018; Łasica et al., 2024). Risk factors that exacerbate disease progression include lack of proper hygiene, smoking, diabetes, obesity, and osteoporosis. Colonization of the periodontal pocket by pathogenic microorganisms promotes inflammation and tissue destruction (Łasica et al., 2024).
Although cultivation-based methods have identified many bacterial taxa, a large portion of oral microbes remains unculturable (Benn et al., 2018). The expanded Human Oral Microbiome Database (eHOMD) offers curated information on bacterial taxa inhabiting the human oral cavity and aerodigestive tract. Among the 834 listed taxa, 523 are primarily oral and 22 are primarily nasal. Of the oral taxa, 49% are formally named, 21% are cultivated but unnamed, and 29% are known solely as uncultivated phylotypes (Escapa et al., 2020). To study both culturable and unculturable members of the microbiota, nucleic acid-based techniques such as 16S rRNA gene sequencing and shotgun metagenomics are commonly used. The former targets specific hypervariable regions for taxonomic resolution, while the latter offers a broader and more functional insight into the microbial community, particularly when reference genomes are lacking. Oral rinse sampling is a practical, non-invasive approach suitable for large-scale studies, enabling effective preservation and transport of DNA (Nearing et al., 2020; Willis and Gabaldón, 2020; Morrison et al., 2023).
The aim of the study was to determine the in vitro antimicrobial properties of A-PRF (platelet-rich fibrin in the form of a membrane obtained after blood centrifugation) and LP (the liquid fraction of plasma remaining after centrifugation) against Gram-negative bacteria: E. coli, P. gingivalis and Gram-positive bacteria creating different shapes and forms: Bacillus subtilis, Micrococcus luteus, Staphylococcus lentus, Enterococcus casseliflavus, and Streptococcus mutans. PRF was obtained from people/patients with a microbiome characterized during our/this study.
2 Materials and methods
2.1 Participants and oral microbiome sample collection
The number of participants in the study was 30, including 15 men and 15 women, between 22 and 38 years old. Patients were selected based on a conducted interview (Supplementary material 1: ‘Patient survey’ and ‘Patient examination form (API)’), blood test results and after an oral cavity examination, which involved the assessment of oral hygiene using the Approximal Plaque Index (API). This index represents the percentage ratio of the number of interproximal tooth surfaces displaying biofilm to the total number of examined interproximal surfaces. Additionally, the presence of dental plaque was evaluated using a periodontal probe for interproximal surfaces from the buccal aspect in the first quarter of the dentition and from the palatal aspect in the second quarter. All participants were in good health, were nonsmokers, had no symptoms of oral infection, possessed their natural dentition and had taken no antibiotics for at least 1 month prior to the experiments.
For the analysis of the oral microbiome composition, we opted to collect site-specific samples for DNA isolation, focusing on mucosal and periodontal surfaces adjacent to areas where PRF may be applied in the treatment of periodontal diseases. All participants included in our study were systemically healthy and in good oral condition (Supplementary material 1). The sampling strategy employed in our research represents one of several validated approaches available for investigating the oral microbiome (Zaura et al., 2021). The samples for DNA isolation were collected by swabbing for approximately 30 s the oral cavity, namely teeth, insides of surface of cheeks and gums, excluding tongue, using Epicentre Buccal Swab (2 swabs in each case). After sample collection, the swabs were placed in a transport system and moved in cooling conditions (4 °C) to the laboratory. The isolation was done in sterile conditions at max. 30 min after samples were collected.
All the protocols used in this study were approved by the Ethics Committee of the Medical University of Warsaw (KB/94/2023). All methods were performed in accordance with the relevant guidelines and regulations. All study participants provided informed consent prior to study enrollment.
2.2 DNA extraction
The total DNA was isolated from swabs using the commercially available kit for DNA purification from buccal swab samples (Swab; A&A Biotechnology, Gdynia, Poland) according to the manufacturer’s recommendation. The quantity and quality of extracted DNA were determined with a Qubit 4.0 Fluorometer (dsDNA high-sensitivity assay kit; Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) and a Colibri spectrophotometer (Titertek Berthold, Pforzheim, Germany), respectively. The DNA samples were isolated in triplicate and then pooled to obtain a single DNA sample for each sample. Only samples with concentrations higher than 10 ng/μL and an A260/A280 ratio ranging from 1.8 to 2.0 were analyzed. DNA samples were stored at −20 °C for further use.
2.3 Sequencing the variable V3–V4 regions of bacterial 16S rRNA
The structure of the bacterial community was determined by sequencing the variable V3–V4 regions of bacterial 16S rRNA (according to Illumina, Part # 15044223 Rev. B) and the fungal community structure was determined using the internal transcribed spacer ITS (ITS3/ITS4; Hoggard et al., 2018) regions of fungal ribosomal DNA, both using the Illumina platform. Sequencing was performed on the Illumina NovaSeq 6,000 instrument using the NovaSeq 6,000 SP Reagent Kit v1.5 (500 cycles) in paired-end read mode (2 × 250 cycles), following the standard procedure recommended by the manufacturer with the addition of 1% PhiX control library. PCR and sequencing were done by The CeNT’s Genomics Core Facility and the analyses were performed by the DNA Sequencing and Oligonucleotide Synthesis Facility (Institute of Biochemistry and Biophysics Polish Academy of Sciences). Raw sequences were processed and analyzed by QIIME2 software (Bolyen et al., 2019) with the DADA2 option for sequence quality control and the newest release of the SILVA ribosomal RNA sequence database (SILVA SSU database 138.2) for taxonomy assignment (Yilmaz et al., 2014; Bolyen et al., 2019).
The amplicon data was visualized using the MicrobiomeAnalyst web server (Chong et al., 2020; Lu et al., 2023). The dataset was not normalized, not scaled and not rarefied. The differences in the alpha-diversity in bacterial and fungal community structure were analyzed using the QIIME2 pipeline, based on the Kruskal–Wallis H-test (Shannon, Chao1, Pielou and Simpson biodiversity indexes). The differences in beta-diversity were evaluated using the QIIME2 pipeline based on Bray-Curtis index. The correlations between different fungal phyla and different bacterial phyla were calculated and visualized using Spearman’s correlation coefficient with MicrobiomeAnalyst. The correlations between phyla were considered strong and significant when the absolute value of Spearman’s rank |r| > 0.9 and p < 0.05 (Zhu et al., 2017).
2.4 Preparation of PRF fraction
From each participant, four venous blood tubes (A-PRF matrix sterile tubes, 10 mL, Dermoaroma Italy Srl 00144), each containing 8 mL, were collected and subsequently processed and centrifuged using a centrifuge (TD4C, Yingtai Instrument) for 14 min at 18.000 rpm at room temperature according to previous reports (Cortellini et al., 2018; Miron et al., 2019). After centrifugation, the LP above the precipitated fibrin fraction was pipetted off. The collected LP was transferred to sterile 1.5 mL Eppendorf tubes. The A-PRF were compressed and converted into a standardized membrane with a thickness of 1 mm to determine their antibacterial abilities (Castro et al., 2017). The clot containing the blood’s cellular components, located below, was removed. Two A-PRF membranes from each patient’s blood were used for further analyses; in cases where high-quality A-PRF could not be obtained (approximately 20% on average), those samples were excluded from this part of the analysis. To assess the participants’ hematopoietic system, an additional blood tube was collected for further peripheral blood morphology parameters analyzed in an external certified laboratory (Supplementary material 2: Table S1).
2.5 Bacterial strains and general growth conditions
The P. gingivalis ATCC33277 strain was grown in enriched tryptic soy broth (eTSB; composition per liter: 30 g trypticase soy broth, 5 g yeast extract, at pH 7.5; further supplemented with 5 mg hemin, 0.5 g L-cysteine, and 2 mg menadione) or on eTSB blood agar (eTSB medium containing 1.5% [wt/vol] of agar; further supplemented with 5% defibrinated sheep blood). S. mutans ATCC25175 strain was grown in eTSB. Both strains were incubated at 37 °C in anaerobic conditions with an atmosphere of 90% nitrogen, 5% carbon dioxide, and 5% hydrogen provided by Anoxomat® II Anaerobic Culture System (Madej et al., 2021).
Escherichia coli, B. subtilis, M. luteus, S. lentus, E. casseliflavus (collection of Institute of Microbiology, University of Warsaw) were grown in Luria-Bertani broth (LB) or on 1.5% LB agar plates at 37 °C in aerobic conditions, depending on experiment.
2.6 Antibacterial test of the A-PRF membranes and liquid fraction LP
To assess antimicrobial properties of A-PRF clot and LP using agar plates, cell suspensions (0.5 McFarland scale) of all strains: E. coli, B. subtilis, M. luteus, S. lentus, E. casseliflavus, S. mutans were spread evenly on Mueller Hinton II agar and P. gingivalis on eTSB blood agar with cotton swab (according to the Kirby-Bauer method (Biemer, 1973)) and left aside for 15 min. Next, A-PRF clots were placed on a medium surface as a whole fragment or cut into smaller pieces (one whole A-PRF clot was cut on a maximum five smaller pieces of equal size) (Feng et al., 2020). For LP analysis, sterile cellulose discs (Oxoid, Thermo Scientific) soaked with 20 μL of the LP fraction were placed on the surface of medium covered in bacterial suspensions. Additionally, LP liquid was also directly applied (20 μL) onto plates. Disks soaked with 20 μL of sterile phosphate buffered saline (PBS, ph 7.4; Merck, Germany) served as a negative control. Plates were incubated for 24-48 h at 37 °C in aerobic conditions for E. coli, B. subtilis, M. luteus, S. lentus, E. casseliflavus (group A), and for 72 h at 37 °C in anaerobic conditions for S. mutans and P. gingivalis (group B). After incubation the inhibition zones around A-PRF membranes, discs (6 mm) with LP and liquid drops of LP were measured. Tests were done according to Figure 1.
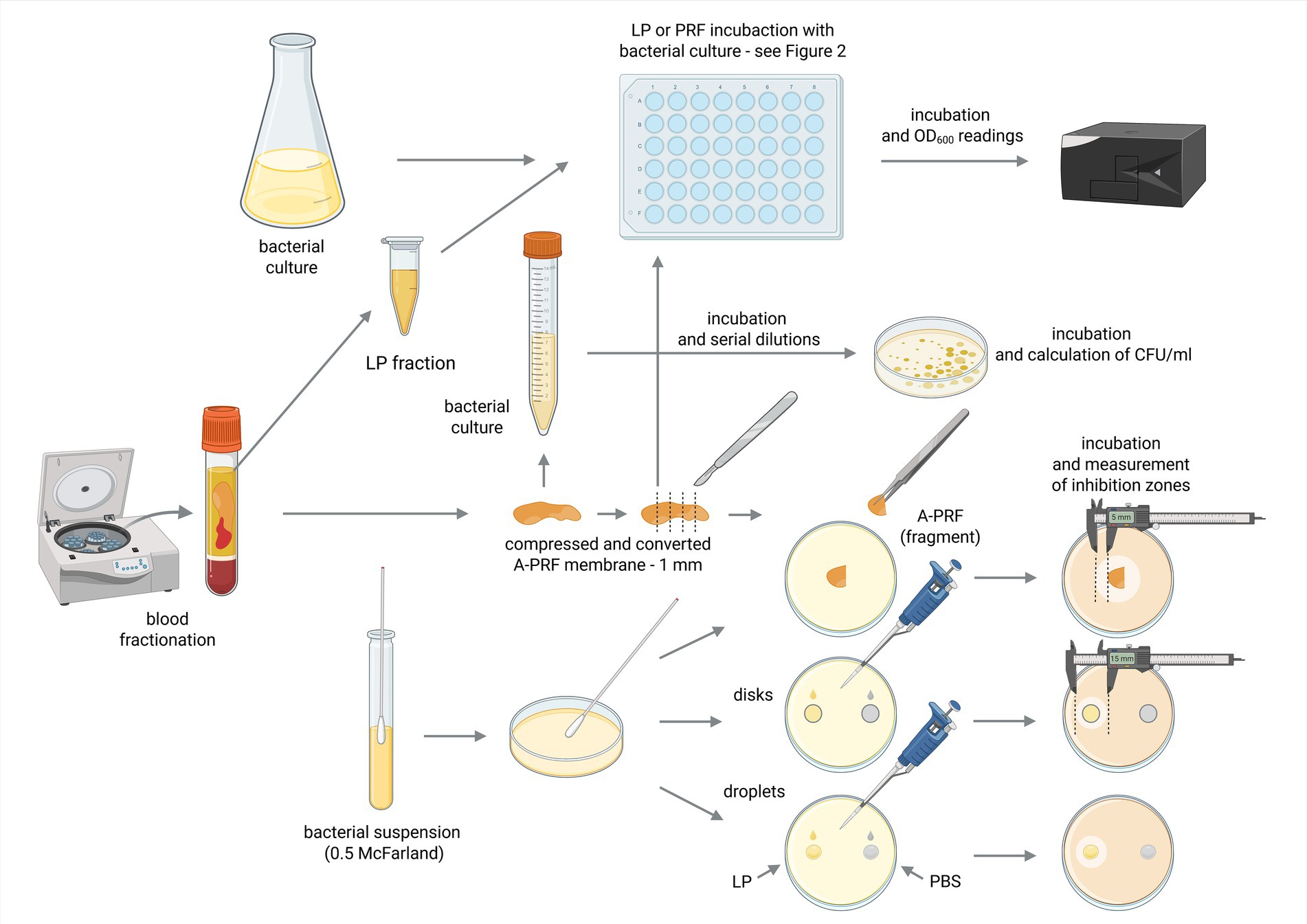
Figure 1. Schematic diagram of the conducted experiments. Created in BioRender [Zalewska M. (MZ; user: Magda Z, 2025) https://BioRender.com/mkxkcgk]. Fractions acquired from blood centrifugation (LP and A-PRF) were used for antibacterial testing performed in liquid (titration plates, OD600 spectrophotometer readings) or on solid medium (calculation of CFU/ml and inhibition zones measurements); LP, liquid fraction of plasma remaining after blood centrifugation; A-PRF, advanced platelet-rich fibrin; PBS - phosphate buffered saline.
To perform the test using liquid medium, titration plates (48-well, flat-bottom; Avantor) were prepared following Scheme 2. Briefly, 300 μL of two-fold concentrated Mueller-Hinton Broth (Oxoid) or eTSB culture medium was added to each well. Then, wells were supplemented with either 300 μL of bacterial suspension during logarithmic growth phase (OD600 = 0.8 or 0.4) or 300 μL PBS as a control. One fragment of A-PRF membrane (one whole A-PRF membrane was cut into a maximum of 5 smaller pieces of equal size) or 100 μL of LP was added to selected rows of wells with bacterial culture. One row was left as a positive control for bacterial growth. Additionally, sterility control was applied for A-PRF and LP fractions where no bacterial inoculum was added to selected wells (Figure 2).
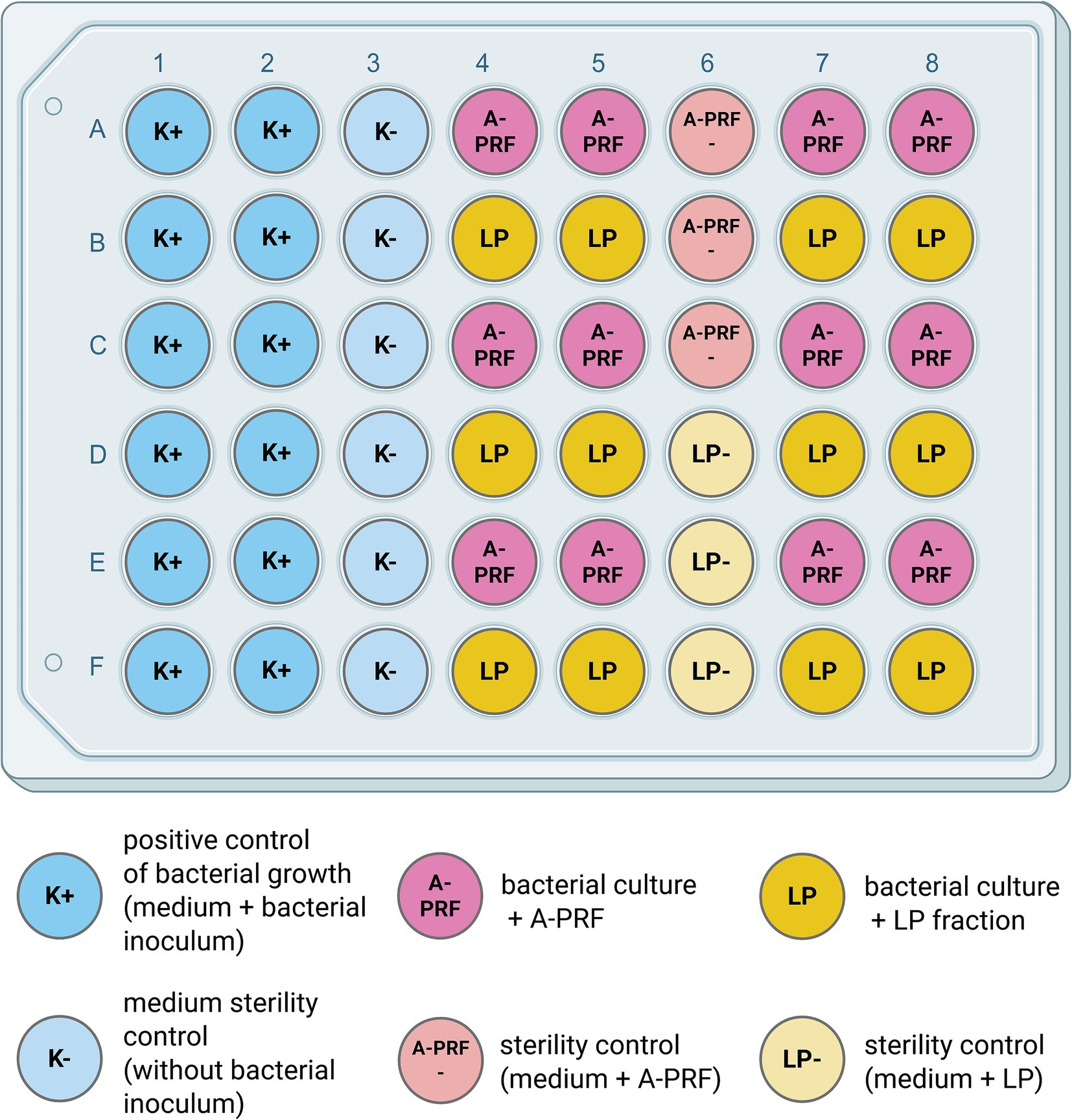
Figure 2. Schematic view of the experiment setup on the titration plate. Created in BioRender [Zalewska M. (MZ; user: Magda Z, 2025) https://BioRender.com/0j45vwx]. Antibacterial activity of LP or A-PRF was performed in a liquid medium and assessed by OD600 spectrophotometer readings. One plate was used either for 3 or 1 strain from group A or B, respectively; LP - liquid fraction of plasma remaining after blood centrifugation; A-PRF - advanced platelet-rich fibrin.
Prepared plates were incubated at 37 °C in aerobic conditions for E. coli, B. subtilis, M. luteus, S. lentus, E. casseliflavus (group A), and in anaerobic conditions for S. mutans and P. gingivalis (group B). Growth rates were determined by OD600 measurements on a spectrophotometer (SpectraMax® iD3 Multi-Mode Microplate Reader, Molecular Devices) every 60 min for 7 h and then after 24 h (group A) and every 24 h for 72 h (group B). Due to the presence of a solid A-PRF matrix in the wells, spectrophotometric measurement of the entire suspension resulted in artificially elevated OD600 values. Therefore, for samples containing A-PRF, a portion of the bacterial suspension was carefully removed and measured separately using a spectrophotometer to obtain accurate OD600 readings (Berthold Detection Systems GmbH).
For E. coli, S. mutans and P. gingivalis the experiment was repeated using the whole A-PRF membranes (from two patients in each case) in a larger volume (3 mL) of MH or eTSB culture medium, respectively. A positive control for the growth of the tested bacterial strains was also prepared (culture without A-PRF). 0.3 mL of an overnight culture of the tested strain was added to each tube. Incubation, in the specified conditions described above, was carried out in Falcon tubes (15 mL) for 24 h and then a series of dilutions of 10–1 – 10 –7 were prepared and plated (100 μL) on a dedicated solid medium (as described in p. 2.5) in duplicate. Plates were then incubated for 24 h - 72 h at 37 °C in aerobic conditions for E. coli and in anaerobic conditions for S. mutans and P. gingivalis. After incubation, colony forming units (CFU/ml) were determined.
3 Results
3.1 Study participants
All study participants were in good health, were non-smokers, had no symptoms of oral infection, possessed their natural dentition and had taken no antibiotics for at least 1 month prior to the experiments. Finally, 20 patients, including 9 men and 11 women were selected based on a conducted interview (Supplementary material 1: ‘Patient survey’ and ‘Patient examination form (API)’), blood test results (Supplementary material 2: Table S1) and after an oral cavity examination.
3.2 Bacterial community composition
Bacterial community structure at the phylum is visualized in Figure 3 and the core microbiome for the samples for phylum is shown in Figure 4.
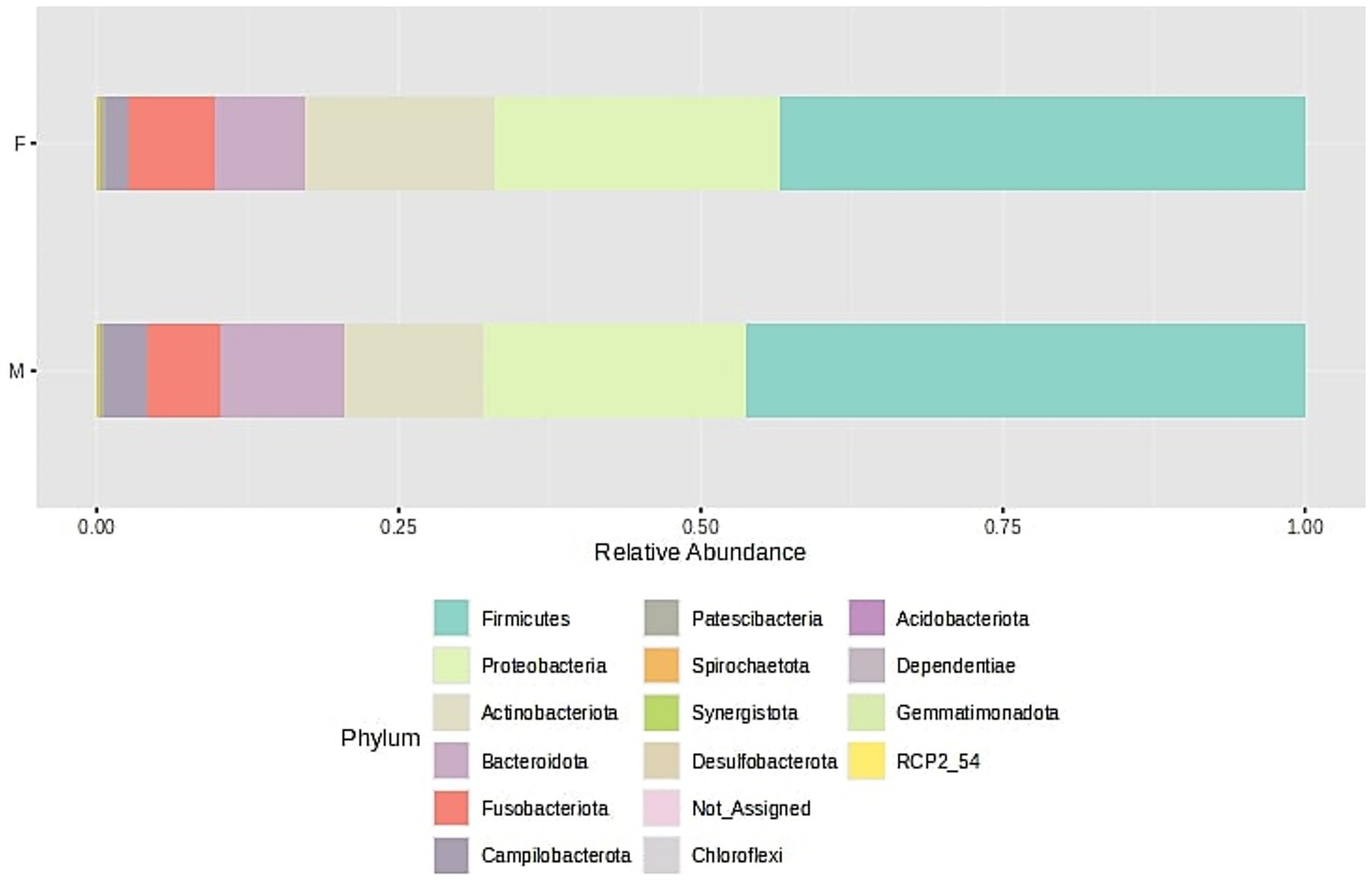
Figure 3. Bacterial community structure composition of samples at the phylum level. The samples were divided into two groups - F (female) and M (male); stacked barplots represent the average value for all samples assigned to groups.
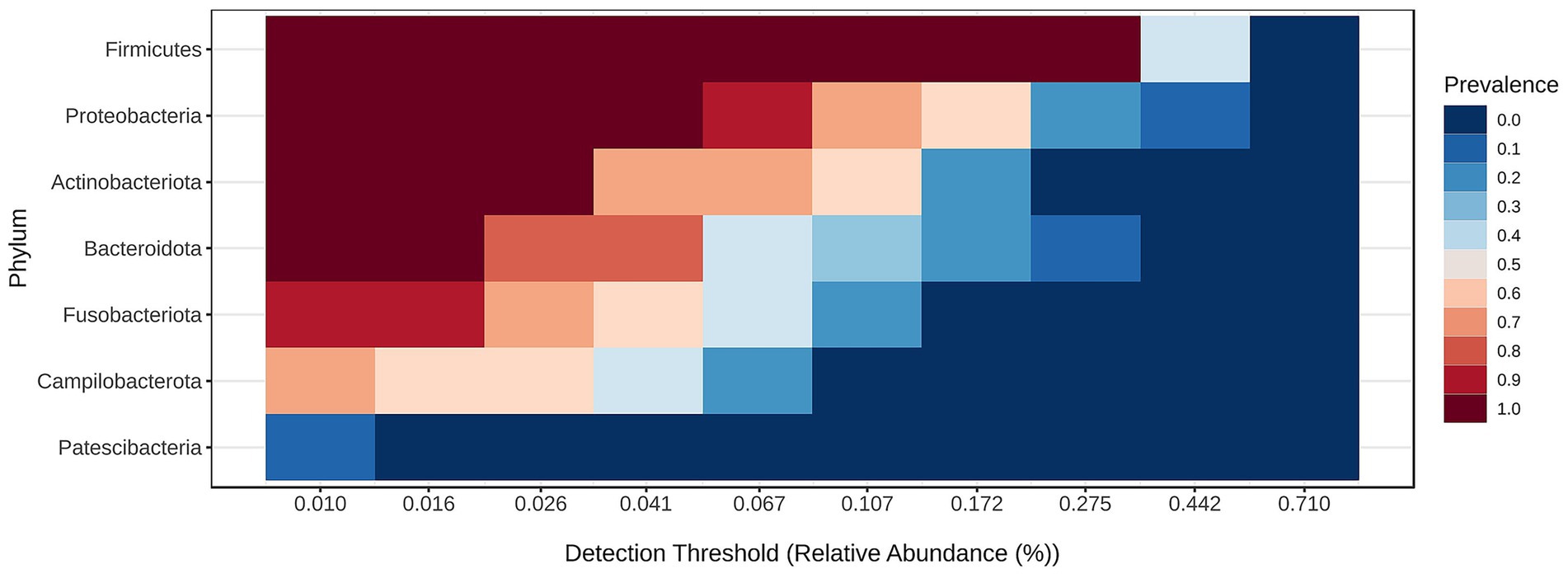
Figure 4. Core bacteria phyla across all samples.
For all samples originated from female and male, the alpha-biodiversity indexes have been calculated: Chao1 (estimates species richness, emphasizing rare species), Shannon (accounts for both richness and evenness), Simpson (measures dominance) and Pielou (quantifies the evenness of species distribution within a sample), and they do not differ between both analyzed groups, namely female and male (p > 0.05). Moreover, beta-diversity has been analyzed and the Bray-Curtis index, which quantifies community dissimilarity based on species abundance (considering both shared and unique species between samples) and the Jaccard index, which measures compositional similarity, focusing solely on species presence or absence without accounting for abundance do not differ between female and male samples (p > 0.05). Boxplots for diversity indexes data distribution are presented in Supplementary material 3: Microbiome biodiversity indexes comparison between female and male patients (Supplementary Figures S1–S12).
Microbiome analyses (Figure 3) revealed that the majority of bacterial phyla belonged to the Firmicutes, followed by Proteobacteria and Actinobacteria, with lower relative abundances of Bacteroidota, Fusobacteriota, and Campylobacteriota. The dominant bacterial classes were Bacilli, Gammaproteobacteria, Actinobacteria, Negativicutes, and Bacteroidia, followed by Fusobacteria, Campylobacteria, and Clostridia, listed in order of relative abundance. This taxonomic distribution pattern was consistent across samples obtained from both male and female participants. These findings are in line with diversity index analyses, which indicated no significant differences in microbial diversity between the groups.
The core microbiome analysis (Figure 4) demonstrated that Firmicutes represent the dominant and most consistently present phylum across samples, indicating their role as a key component of the core microbiome. Proteobacteria were also highly prevalent, though generally detected at lower relative abundances than Firmicutes. Actinobacteriota appeared to be a relatively stable, albeit less abundant, member of the microbiome. In contrast, Bacteroidota were commonly detected but rarely exceeded higher abundance thresholds. Fusobacteriota, Campylobacterota, and Patescibacteria were found infrequently and at low abundance levels, suggesting they may constitute part of the variable or transient microbiome. Notably, only Firmicutes, Proteobacteria, and Actinobacteriota maintained both high prevalence and moderate abundance, supporting their classification as core phyla.
The correlation network between bacterial classes identified in the oral cavity (Figure 5) reveals patterns of microbial co-occurrence and potential ecological interactions. Each node represents a bacterial class, with color indicating the origin of the sample: orange for female-derived samples, green for male-derived samples, and bi-colored nodes representing taxa present in both types of samples. Edges indicate statistically significant correlations in relative abundance, suggesting co-occurrence. A substantial number of bacterial classes, including Clostridia, Bacteroidia, Negativicutes, Gammaproteobacteria, Fusobacteriia, and Bacilli, appear in both male and female samples and form a dense central cluster. This suggests the presence of a core microbial community that is shared across sexes and potentially functionally integrated through ecological interactions. The dense connectivity of these bacterial classes may indicate synergistic or co-dependent metabolic relationships within the oral microbiome. In contrast, certain bacterial classes such as Desulfotobacteria, Babeliae, and the unclassified RCP2_54 appear to be specific to female samples, while Acidimicrobia, Longimicrobia, and Chloroflexia are uniquely associated with male samples. These findings suggest the existence of sex-specific components of the oral microbiota; however, the analysis of biodiversity indexes does not confirm these findings. Oral microbiome may be additionally influenced by factors such as hormonal environment, behavioral habits, or differential exposure to environmental factors.
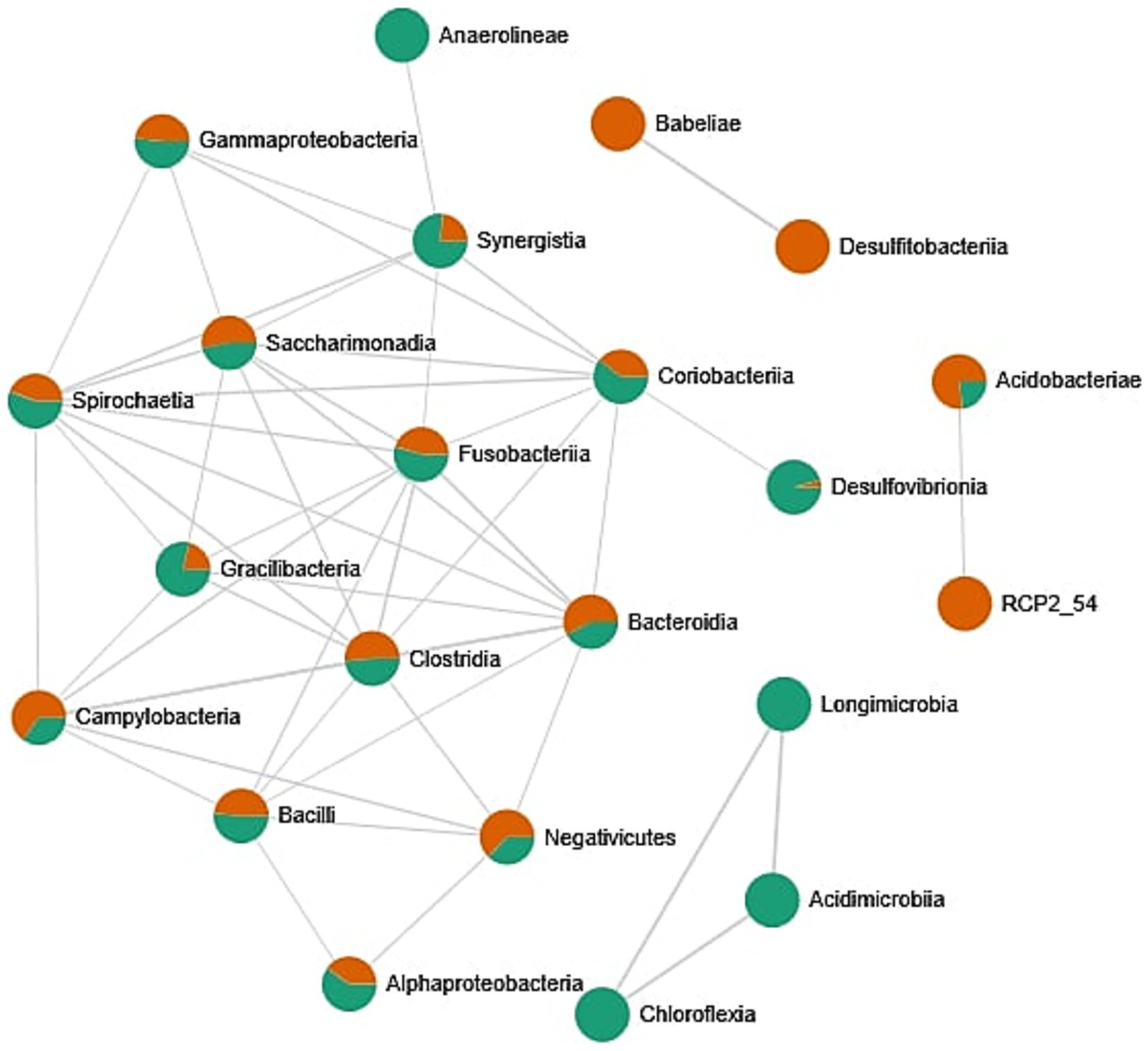
Figure 5. Correlation network between bacterial classes identified in the oral cavity. Network was created by the MicrobiomeAnalyst web tool. Only strong (|r| > 0.5) and significant (p < 0.05) correlations are presented. Nodes as a pie chart represents classes’ relative abundances. The color of the nodes indicates: female samples (green) and male samples (orange).
3.3 Fungal community structure
Fungal community structure at the phylum level is visualized in Figure 6 and the core mycobiome for the samples for phylum levels are shown in Figure 7.
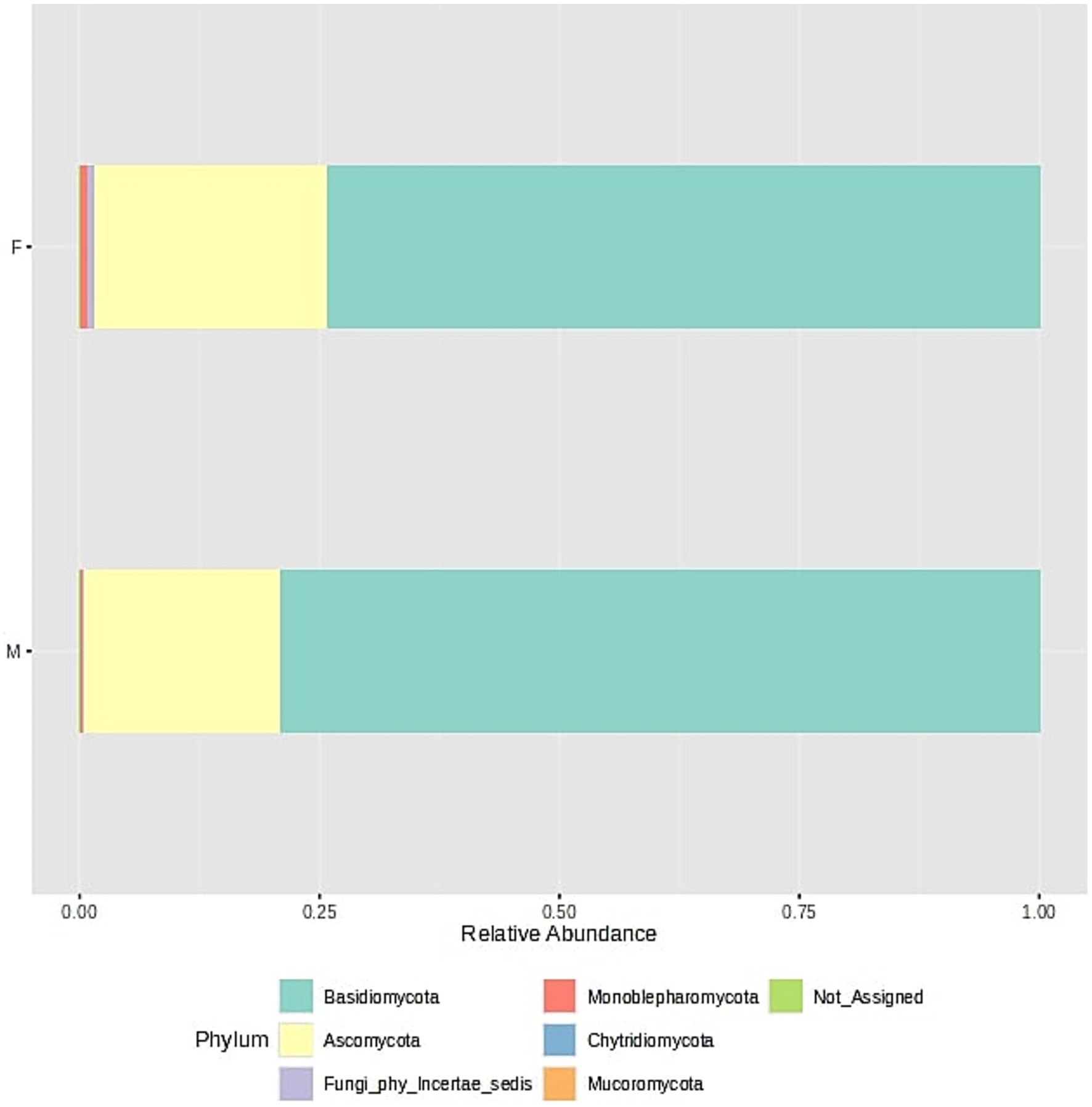
Figure 6. Fungal community structure composition of samples at the phylum level. The samples were divided into two groups - F (female) and M (male); stacked barplots represent the average value for all samples assigned to groups.
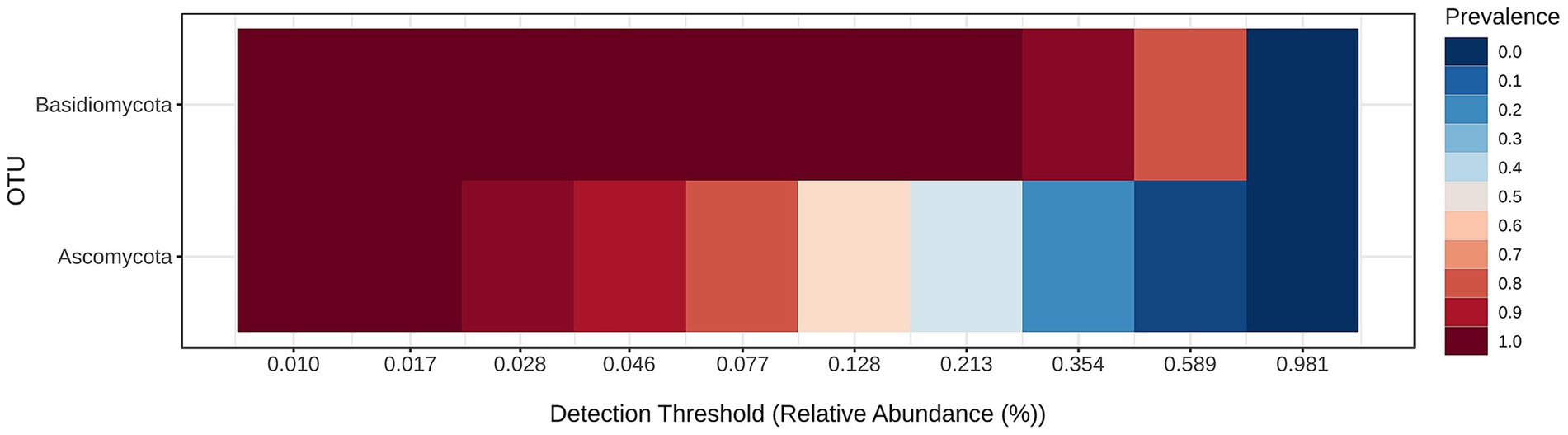
Figure 7. Core fungal phyla across all samples.
For all samples the alpha-biodiversity indexes have been calculated: Chao1, Shannon, Simpson and Pielou, and they do not differ between both analyzed groups (p > 0.05). Furthermore, beta-diversity has been analyzed and indexes do not differ between groups (p > 0.05). Boxplots for diversity indexes data distribution are presented in Supplementary material 3: Microbiome biodiversity indexes comparison between female and male patients (Supplementary Figure S1–S12).
Fungal community composition (Figure 6) in oral samples shows a dominance of two main phyla: Basidiomycota and Ascomycota. In both male and female samples, Basidiomycota represents the most abundant phylum, accounting for approximately three-quarters of the total fungal community. Ascomycota forms the second most prominent phylum, constituting about one-fourth of the total relative abundance. Other fungal phyla such as Mucoromycota, Chytridiomycota, and Monoblepharomycota are present in much lower proportions. Their detection suggests a minor and potentially transient role in the oral ecosystem. A small fraction of the sequences remains unassigned (Not_Assigned) at the phylum level, which may indicate the presence of novel or less-characterized fungal phyla. Importantly, the overall fungal composition is highly similar between sexes, with no major differences in the relative proportions of dominant phyla. This suggests a relatively conserved fungal core microbiome across male and female individuals in the studied population. This was also confirmed by analysis of different biodiversity indexes (Supplementary material 3: Microbiome biodiversity indexes comparison between female and male patients, Supplementary Figure S1–S12).
The core microbiome analysis (Figure 7) demonstrates that Basidiomycota exhibits consistently high prevalence across nearly all detection thresholds of relative abundance. This indicates that members of this phylum are both widely distributed and consistently present in the majority of samples, making Basidiomycota a clear component of the core oral mycobiome. In contrast, Ascomycota shows a broader range of prevalence, with high detection at lower abundance thresholds, but decreasing prevalence at higher thresholds. This suggests that while Ascomycota is also a frequent constituent of the oral fungal community, it may exhibit greater variability in abundance among individuals, potentially reflecting individual differences or transient colonization. Overall, this heatmap supports the conclusion that Basidiomycota forms the most stable and dominant component of the core oral mycobiome, while Ascomycota may represent a more variable, yet common, element of the community.
The fungal co-occurrence network (Figure 8) illustrates associations between different fungal classes identified in oral samples, with nodes colored according to their relative representation in male and female subjects. Mixed-colored nodes indicate classes present in both sexes, albeit at varying abundances. Saccharomycetes appears as a central and well-connected node, forming links with Taphrinomycetes and Malasseziomycetes, suggesting potential co-occurrence patterns or shared ecological niches in the oral environment. These classes are commonly found in both male and female samples and likely represent stable components of the core mycobiome. A separate cluster includes Cystobasidiomycetes and Fungi_cls_Incertae_sedis, both predominantly found in male samples. This may indicate sex-specific factors influencing the presence or abundance of these fungi, such as hormonal differences, lifestyle, or oral hygiene habits. Other classes, such as Eurotiomycetes and Dothideomycetes, show a slightly higher prevalence in female samples. Meanwhile, a distinct group composed of Pucciniomycetes, Spizellomycetes, and Dacrymycetes appears isolated and is found exclusively or primarily in male samples, possibly reflecting transient or niche-specific colonizers. The analysis of different biodiversity indexes does not confirm these assumptions (Supplementary material 3: Microbiome biodiversity indexes comparison between female and male patients, Supplementary Figures S1–S12). Overall, the network highlights the existence of a core group of fungal classes common across individuals, as well as variable or rare taxa that may reflect individual-specific or environmental factors. The patterns of co-occurrence provide insights into the complexity of the oral mycobiome.
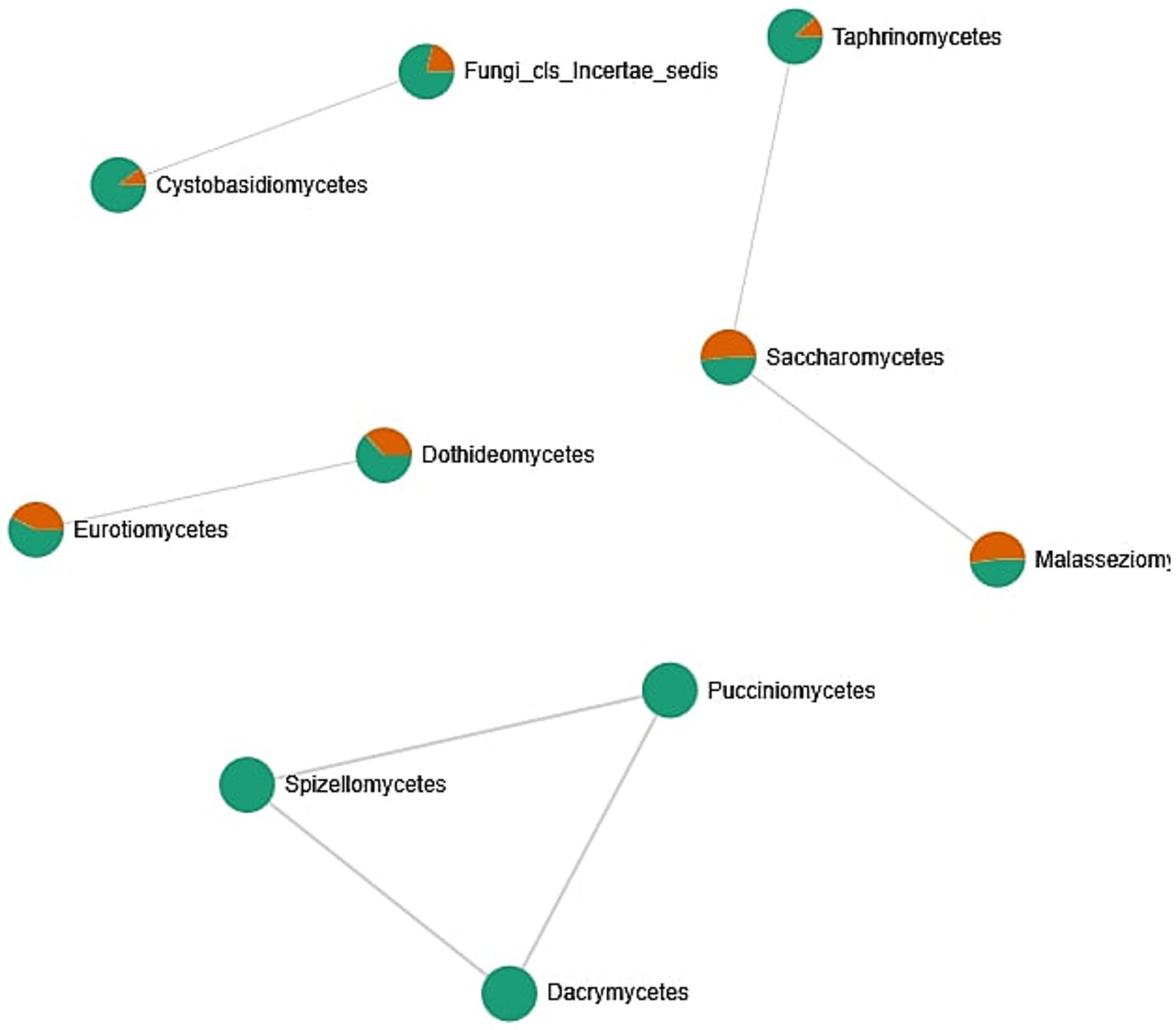
Figure 8. Correlation network between fungal classes identified in the oral cavity. The network was created by the MicrobiomeAnalyst web tool. Only strong (|r| > 0.5) and significant (p < 0.05) correlations are presented. Nodes as a pie chart represents classes’ relative abundances. The color of the nodes indicates: female samples (green) and male samples (orange).
3.4 Antibacterial effect of the A-PRF membranes and liquid fraction LP
The antibacterial activities of the A-PRF membrane and LP fraction were analyzed by directly placing them on the surface of a medium covered in bacterial suspension. Activity against bacteria with different cell wall structures and also having different shapes and creating different structures was studied. In all tested systems, after the defined incubation period and conditions, antimicrobial activity was demonstrated as a clear zone around the A-PRF membrane, the cellulose discs soaked with the LP fraction and the LP fraction dropped on the surface of medium covered in bacterial suspension. In each case, a wider inhibition zone was observed for E. coli (LP: average 10 mm ± 1 mm; A-PRF: average 4 mm ± 1 mm) compared to the tested Gram-positive bacteria (LP: 6–9 mm; A-PRF: 0–3 mm) - for E. casseliflavus – 6 mm (no inhibition zone for LP and A-PRF), M. luteus and B. subtilis - LP: 7 mm; A-PRF: 1 mm, S. lentus - LP: 9 mm; A-PRF: 3 mm. Interestingly, the clear zones were better visible after 24 h of incubation than after 48 h, where overgrowing of this zone with bacteria was observed in each case. Anaerobic bacteria: S. mutans and P. gingivalis were found to have the lowest sensitivity, the growth inhibition zone was less than 7 mm (LP: 6.8 mm; A-PRF: 0.8 mm) after 24 h and 48 h, respectively, but after further incubation, this zone became completely overgrown (Supplementary material 4: Photographic documentation – selected sample photos, A.).
For the analysis of bacterial growth in a liquid medium in the presence of A-PRF membrane or LP fraction, a growth inhibition effect was observed after 6 h for all bacteria compared to the control culture, but the strongest effect was noted for E. coli (Figures 9A,B). Interestingly, for Gram-positive bacteria, the least sensitive were cocci forming structures and capable of forming biofilms: E. casseliflavus, M. luteus, and B. subtilis, but S. lentus, although also capable of forming a biofilm, was definitely more sensitive than other Gram-positive bacteria tested (Figures 9A,B). The highest differences in OD600 readings compared to the control culture was observed after 2 h and 4 h of incubation. Antibacterial activity was also confirmed by a lower CFU/ml value at selected time points (Table 1, group A). We also evaluated the condition/size of fragmented A-PRF (titration plates) and a whole A-PRF membrane (liquid culture - 3 mL) after 24 h of incubation with aerobic bacterial cultures. All analyzed membranes remained unchanged (Supplementary material 4: Photographic documentation – selected sample photos, B).
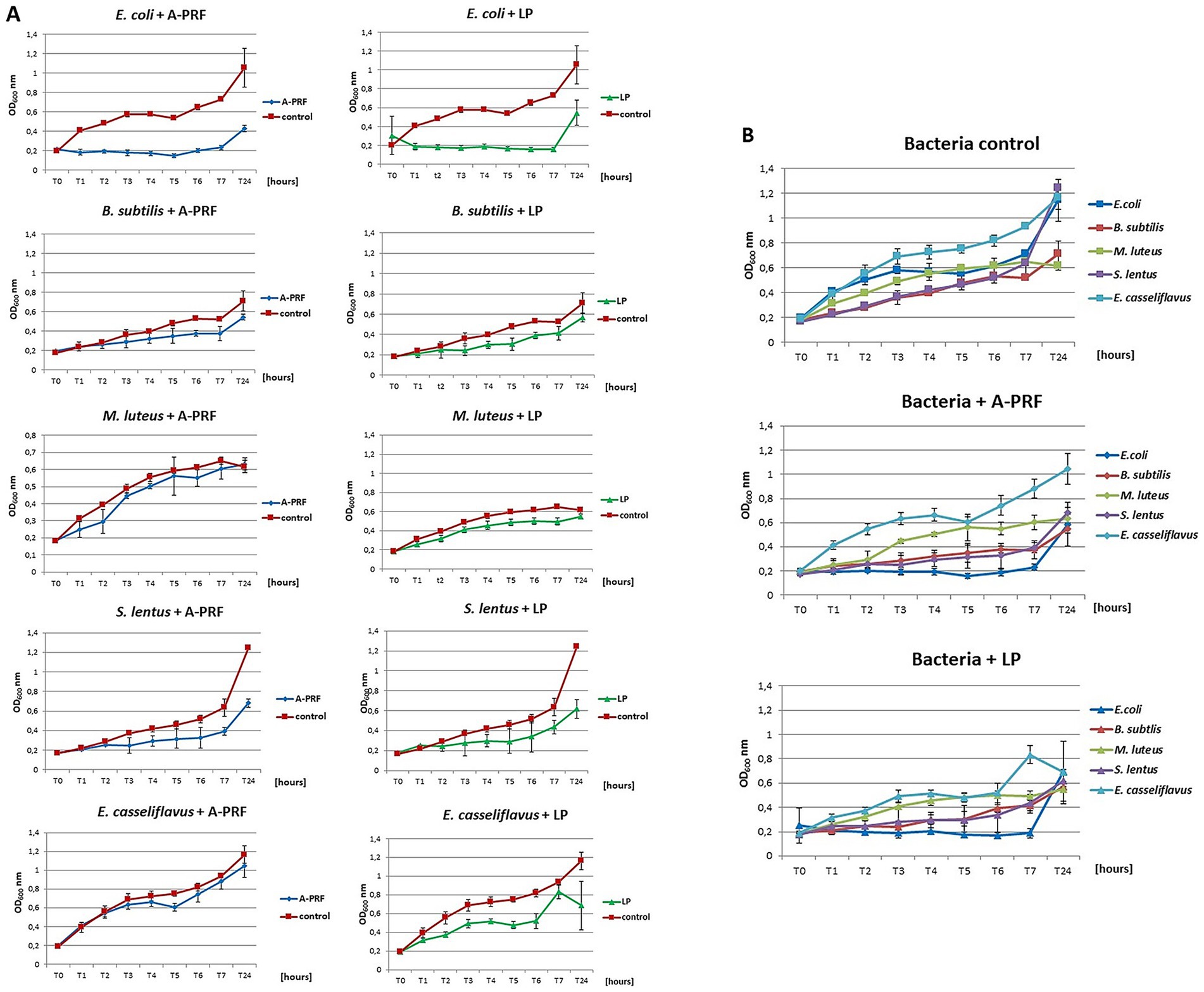
Figure 9. The growth curves of the tested bacteria from group A: E. coli, B. subtilis, M. luteus, S. lentus, E. casseliflavus in liquid medium (titration plates), in the presence of A-PRF or LP fraction, incubated at 37 °C in aerobic conditions. (A) Individual charts; (B) collective charts.
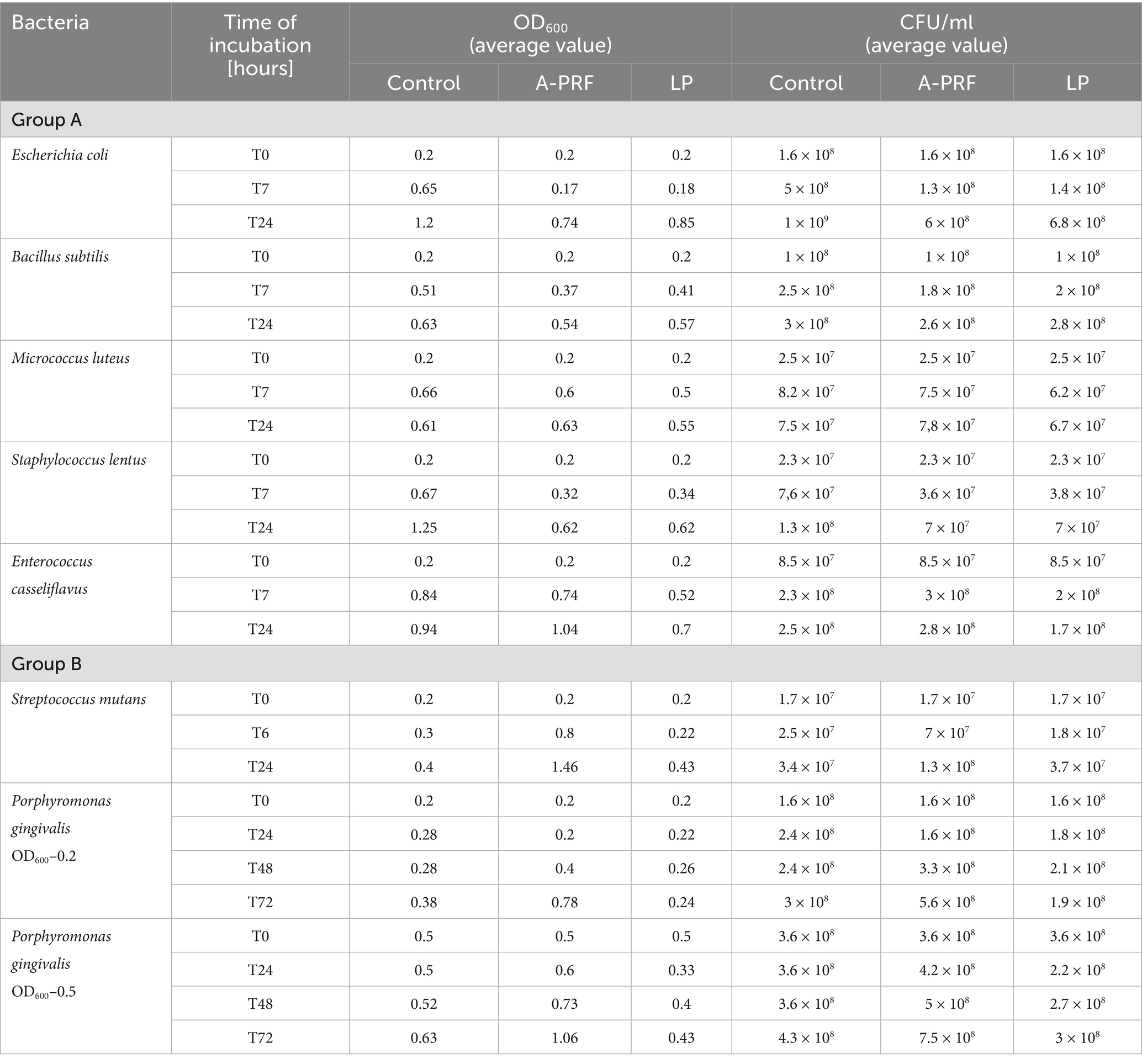
Table 1. The values of OD600 and CFU/ml obtained in the tests for group A and B bacteria, taking into account the incubation time.
In the case of P. gingivalis, where the OD600 at time point T0 was 0.2, a slight inhibition of bacterial growth was observed after 6 h with the A-PRF membrane: the OD600 increased by an average of 1.5-fold, whereas in the control sample (without A-PRF), the OD600 increased nearly threefold. In a parallel experiment with a higher initial OD600 value of 0.5 at T0, a moderate increase in OD600 was recorded in the culture supplemented with the A-PRF membrane (2.5-fold), compared to the control, where OD600 increased by an average of 1.7-fold. After 24 h, OD600 values of both culture variants converged. For S. mutans, no significant differences in OD600 values were observed between the experimental and control conditions after both 6 h and 24 h of incubation (Table 1, group B). Due to the generation time of S. mutans of 2 h and P. gingivalis of an average of 7 h, it was decided to perform an analogous study using 48-well titration plates and OD600 for S. mutans at T0, T2, T4, T6 and T24 and for P. gingivalis at T0, T24h, T48h and T72h. For P. gingivalis, with an initial OD600 around 0.2, an antibacterial effect of the A-PRF membrane was observed after 24 h of incubation. However, this effect was no longer evident after 48 h, as an increase in OD600 compared to the control culture was observed. After 72 h of incubation, the optical density had nearly doubled relative to the control. The application of the LP fraction had minimal impact on the bacterial growth level (Figure 10A). When the initial OD600 was approximately 0.5, no antibacterial effect of the A-PRF membrane was detected, and after 72 h, a more than 1.5-fold increase in OD600 compared to the control was observed. In the case of the LP fraction, a slight decrease in OD600 was noted only after 24 h of incubation (Figure 10B). The results obtained in the study using S. mutans, with an initial optical density of about 0.2, revealed a lack of sensitivity to the antibacterial activity of the A-PRF membrane and even observed growth stimulation. After 6 h of incubation, there was a more than 3-fold increase in OD600 and after 24 h as much as 4.5-fold in comparison with the control culture (Figure 10C). The use of the LP fraction had no effect on the level of growth of this bacteria (Figure 10). Antibacterial activities or their lack was also confirmed by a CFU/ml value (Table 1, group B). Interestingly, after 24 h of incubation, the A-PRF membrane was completely dissolved in cultures of both bacterial species, both in the case of whole membranes (liquid culture - 3 mL) (Supplementary material 4: Photographic documentation – selected sample photos, B.) and their fragments (titration plates).
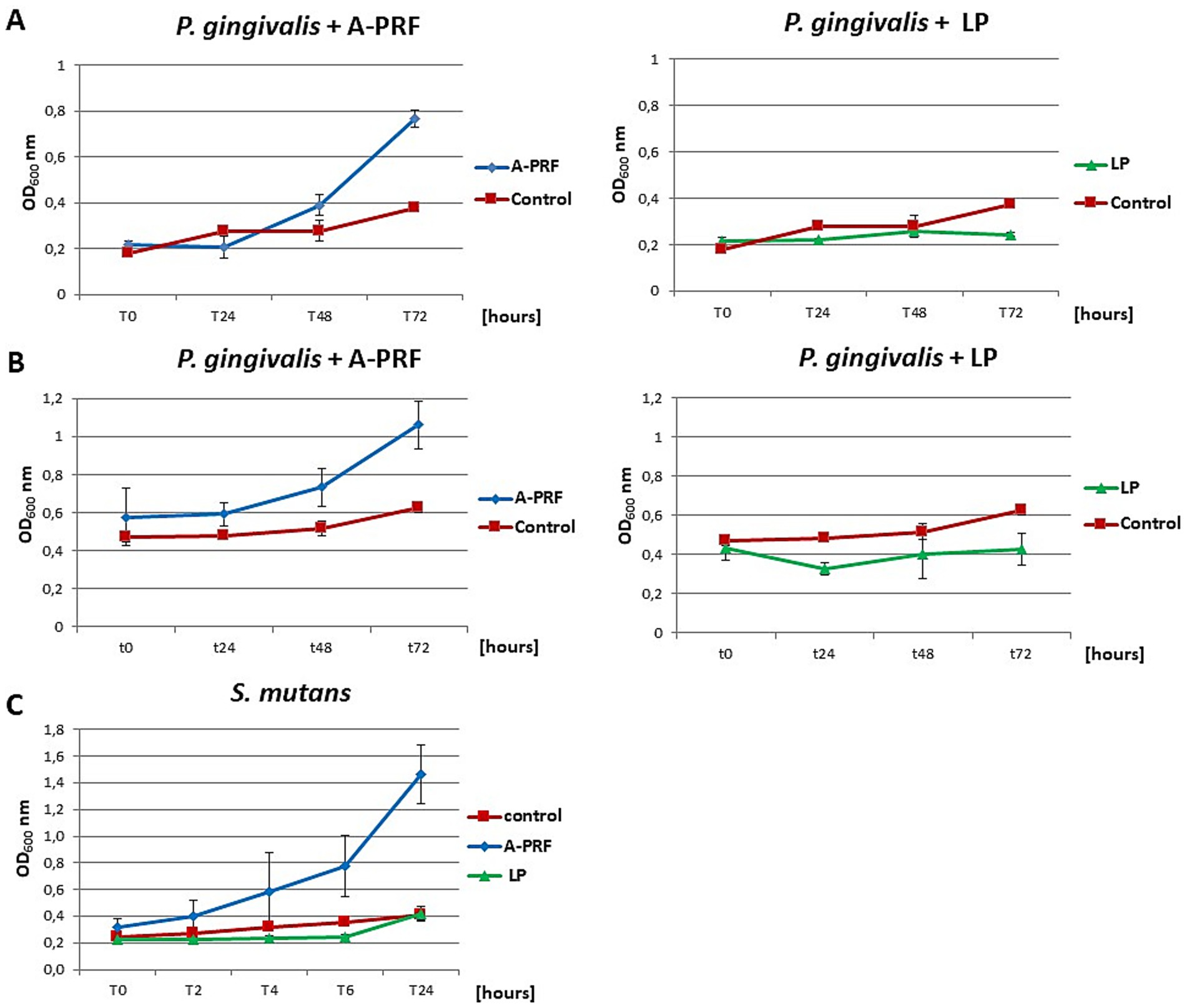
Figure 10. The growth curves of the tested bacteria from group B: P. gingivalis and S. mutans in liquid medium, in the presence of A-PRF or LP fraction, incubated at 37 °C in anaerobic conditions. (A) P. gingivalis with an initial OD600 approx. 0.2; (B) with an initial OD600 approx. 0.5; (C) S. mutans.
4 Discussion
Antibacterial resistance is currently recognized as a serious global health threat. Due to misuse or overuse of antibiotics and other medications, pathogens are no longer susceptible to available treatment that leads to disease spread and death of patients. WHO estimates that AMR contributed to 4.95 million deaths in 2019 (Antimicrobial Resistance Collaborators, 2022). The primary action to address this problem is the search for new antimicrobial substances or the modification of previously described concepts. An example of such an approach is the use of advanced PRF, described in this work. Our study design was comprehensive, consisting of clearly defined and logically sequenced stages. PRF was obtained from patients for which we analyzed the complete oral microbiome. Patient qualification involved an evaluation of survey responses, blood test results, and clinical examination of the oral cavity. The study enrolled systemically healthy, non-smoking individuals with no signs of oral infection, a complete natural dentition, and no antibiotic use for at least one month prior to sampling. Subsequently, oral microbiome analysis was performed on the qualified individuals of both sexes. After confirming that biodiversity indices did not differ significantly between males and females, the entire cohort was considered a homogeneous study group.
Our microbiome studies were largely consistent at the phylum level with those by Zaura et al. (2009) despite methodological and demographic differences (analysis of only three healthy individuals with differing geographic origins, assessment of oral health based solely on clinical periodontal status without consideration of biochemical or immunological markers). They identified Firmicutes (including Streptococcus spp., Granulicatella spp., and members of the Veillonellaceae family), Proteobacteria (Neisseria spp., Haemophilus spp.), Actinobacteria (Corynebacterium spp., Rothia spp., Actinomyces spp.), Bacteroidetes (Prevotella spp., Capnocytophaga spp., Porphyromonas spp.), and Fusobacteria (Fusobacterium spp.) as the predominant taxa in the oral microbiome as was also shown in our studies (Figures 3, 4).
Two other reports indicated the same taxa as the core oral microbiota in comparison with our results, however, their contribution was different (Verma et al., 2018; Nearing et al., 2020). They identified Actinobacteria, Fusobacteria, Proteobacteria, Firmicutes, and Bacteroidetes, respectively, as the most prevalent oral phyla. In contrast, we observed Firmicutes, Proteobacteria, Actinobacteria, Bacteroidota, and Fusobacteria as the dominant phyla. Nevertheless, it is consistent that these five phyla constitute approximately 90% of the oral microbial community.
In the present study, we analyzed 20 samples obtained from healthy male and female participants. Although the data were initially treated as a single cohort, we also investigated potential sex-based differences in microbial composition. No differences were observed in alpha- or beta-diversity indexes, likely due to high intra-sample variability or the influence of uncontrolled variables (not included in the participant survey). However, co-occurrence network analysis revealed microbial associations suggestive of sex-specific structuring. These findings imply that additional factors—such as hormonal status, behavioral differences, or environmental exposures—may contribute to shaping the oral microbiota in a sex-dependent manner. For example, hormonal fluctuations during the menstrual cycle are associated with changes in the relative abundances of Campylobacter spp., Haemophilus spp., Oribacterium spp., and Prevotella spp. (Bostanci et al., 2021), and pregnancy has been linked to increased levels of Neisseria spp., Porphyromonas spp., and Treponema spp., while Streptococcus spp. and Veillonella spp. are more abundant in non-pregnant women (Saadaoui et al., 2021).
Dietary patterns also modulate the oral microbiota: intake of fiber, medium-chain fatty acids, piscine monounsaturated fatty acids, and polyunsaturated fatty acids is associated with increased microbial diversity, while consumption of sugar and refined carbohydrates correlates with higher abundances of specific bacterial taxa. Notably, carbonated beverage consumption is positively associated with the presence of Bacteroidetes, Gammaproteobacteria, Fusobacterium, and Veillonella (Hansen et al., 2018; Chumponsuk et al., 2021).
Environmental factors such as smoking are known to significantly alter oral microbial composition, particularly by favoring anaerobic taxa (Mason et al., 2015). Oral hygiene practices also play a crucial role in shaping microbial diversity (Wade, 2021). Geographic and climatic differences can further modulate the oral microbiome, potentially contributing to inter-individual variation at the species and strain levels (Mason et al., 2015; Li et al., 2022). However, in the present study, we controlled for these two specific factors by recruiting only non-smokers residing in the same geographic area.
Despite numerous individual-specific influences on the oral microbiota, core microbiome analysis can serve as a reference point for microbial eubiosis. The oral core microbiome, defined as the set of consistently shared taxa and their genetic content across individuals, reflects a functionally important microbial backbone. Caselli et al. (2020) demonstrated a lack of statistically significant differences in core microbiome composition among study participants, supporting its potential utility as a benchmark for oral health.
It has been found that fungi account for 0.004% (approximately 100 species) (Ghannoum et al., 2010; Peters et al., 2017) of the overall oral microorganisms and have been detected in specimens from the hard palate, supragingival plaque, and oral rinses (Caselli et al., 2020), but also in saliva samples (Malassezia spp. and Candida spp.) (Li et al., 2022). Due to the lack of data aiming at a healthy human oral mycobiome, it is hard to conduct comprehensive analysis, but we found Basidiomycota and Ascomycota representing three-quarters of the total fungal community. Basidiomycota is commonly associated with environmental fungi, but some members may also be part of the normal oral microbiota (Dong et al., 2021), and Ascomycota include various yeasts and filamentous fungi, some of which are frequently found in the human oral cavity, e.g., Candida spp. The common fungi genera found in the oral cavity together with Candida spp. are Aspergillus spp., Aureobasidiums spp., Cladosporium spp., Cryptococcus spp., Fusarium spp., Gibberella spp., Penicillium spp., Rhodotorula spp., Saccharomycetales spp., and Schizophyllum spp. (Ghannoum et al., 2010; Peters et al., 2017). Moreover, Candida spp. is widely reported as a part of a healthy human oral microbiome, but also during disease (Mueller and Tainter, 2025).
Given the absence of differences in microbiome diversity indices between sexes, all samples were pooled into one dataset, and antimicrobial activity of PRF obtained from patients was assessed. To date, antibacterial activity of PRF has been demonstrated against Gram-negative bacteria: E. coli, Proteus mirabilis and Pseudomonas aeruginosa and Gram-positive bacteria: Bacillus megaterium, Enterococcus faecalis or S. aureus and S. epidermidis (Tohidnezhad et al., 2012; Feng et al., 2020; Jasmine et al., 2020). It is worth emphasizing that those were conducted on a small scale and usually using simple protocol: either determination of minimal inhibitory concentration (MIC) and minimal bactericidal concentration (MBC) for the i-PRF fraction, or analysis of growth inhibition zones around PRF using the agar diffusion technique and counting CFU/ml after the bacterial cells incubation with PRF and only after one specific incubation time. Jasmine et al. (2020) also showed that i-PRF actively inhibited the biofilm formations of tested bacterial strains at 24 h. A recently published study demonstrated antimicrobial effects of clindamycin-loaded PRF against S. aureus, Streptococcus pneumoniae, Streptococcus mitis, P. gingivalis, and Fusobacterium nucleatum using the agar-based diffusion assay (Straub et al., 2024). It is therefore difficult to refer to these findings in the context of results presented in this publication obtained from various experimental approaches.
To date, no attempts have been described elucidating the reasons for higher antibacterial activity of PRF against Gram-negative bacteria compared to Gram-positive ones. Moreover, we did not find previous studies aiming at tracking the dynamics of changes over time of this activity, nor assessing the effects at varying initial bacterial concentrations. In the case of Gram-positive bacteria, the potential effect of PRF on different structural or morphological bacterial forms has also not been assessed. In our study, we addressed these gaps by designing experiments that account for these variables, thus providing a more comprehensive understanding of PRF’s antimicrobial potential. In our research we used: Gram-negative bacteria: E. coli (rod-shaped bacteria commonly found in the gut of humans and warm-blooded animals, but some E. coli strains do cause different illnesses e.g.: diarrhea, urinary tract infections, pneumonia, and even sepsis) (Mueller and Tainter, 2025) and P. gingivalis (Gram-negative, rod-shaped, strictly anaerobic bacteria, a keystone pathogen in chronic periodontitis) (Murugaiyan et al., 2024), and Gram-positive bacteria creating different shapes: B. subtilis (rod-shaped can forming chains, spore-forming bacteria, used as a probiotic, model organism in biotechnology and medicine research, can cause various diseases in humans, including food poisoning, bacteremia, endocarditis, sepsis and pneumonia) (Koca, 2024); M. luteus (spherical bacteria, cocci occurs in tetrads, occurs all over the skin, rarely causes disease, although in people with weakened immune system can cause serious infections) (Shi et al., 2023); S. lentus (cocci, occurring singly, in pairs or tetrads, mainly an animal pathogen, however, it can colonize humans and cause a number of clinical symptoms) (Mazal and Sieger, 2010); E. casseliflavus (spherical, ovoid-shaped bacteria, forming pairs or short chains, occasionally cause opportunistic infections) (Yoshino, 2023); S. mutans (cocci, typically form pairs or chains, facultatively anaerobic bacteria related to the etiology and pathogenesis of dental caries) (Lemos et al., 2019). Gram-positive and gram-negative bacteria differ primarily in their cell wall structure. Our study demonstrated that PRF, regardless of its form, exhibits antibacterial activity, with the strongest effect observed against the Gram-negative bacteria E. coli. Among the tested Gram-positive bacteria, highest activity was noted against S. lentus.
Generally, Gram-positive bacteria possess a thick peptidoglycan layer, with many different proteins and polymers (e.g., teichoic acids), as their primary cell wall component (Pasquina-Lemonche et al., 2020), while gram-negative bacteria have a thin peptidoglycan layer surrounded by an outer membrane containing lipopolysaccharide (Silhavy et al., 2010). For small molecules, the cell wall of Gram-negative bacteria does not pose a barrier; it is also thinner, which results in a possibly faster antibacterial effect. In the case of Gram-positive bacteria—especially those forming various structures such as clusters, chains, or packets—the presence of multilayered peptidoglycan and additional polymers reduces the surface area available for interaction between the antibacterial molecule and the bacterial cell envelope. Additionally, bacteria from this group can produce capsules and modify the charge of their cell wall, making them less susceptible to antibacterial compounds (Epand et al., 2016; Malanovic and Lohner, 2016; Rajput et al., 2024).
A particularly important finding of this study is the demonstrated lack of antibacterial activity of the A-PRF membrane against S. mutans, as well as the stimulation of growth observed for P. gingivalis. Notably, for both species, complete degradation of the A-PRF membrane was observed after 24 h of incubation, in contrast to the other tested bacterial strains. Both S. mutans and P. gingivalis are known biofilm-forming species and are associated with oral health disorders. We hypothesize that bacterial proliferation was boosted by proteolytic degradation of A-PRF, which provides peptides later used as a main or additional source of nutrients. P. gingivalis is an obligate anaerobic bacteria that inhabits the gingival pocket and belongs to the red complex (Aleksijević et al., 2022). Its presence in periodontal tissues has been identified in 78% of patients with periodontitis and only in 34% of healthy people (Rafiei et al., 2018). The main source of carbon and energy for these bacteria are oligopeptides (Polgár, 2002). In the same time, among the most important virulence factors of P. gingivalis are exoproteases called gingipains (RgpA, RgpB and Kgp) (Potempa et al., 1997; Guo et al., 2010). They are present in all strains, and are responsible for 85% of the proteolytic activity of this pathogen (Potempa et al., 1997). Gingipains are very potent enzymes involved in many aspects of physiology and virulence of P. gingivalis. RgpA and Kgp have a hemagglutination-adhesion domain responsible for adhesion and penetration into erythrocytes, which consequently leads to cell disintegration, hemoglobin degradation and iron acquisition (Curtis et al., 1999). In addition, gingipains have the ability to bind and disturb extracellular matrix (ECM) components such as fibronectin or collagen, to degrade antibacterial peptides and to activate or degrade various elements of the immune system. These and many more gingipain activities result in the malfunction of the host immune response, destruction of gingival tissue, alveolar bone loss as well as weakening of blood vessels (Guo et al., 2010). The significant problem in the treatment of infections caused by this pathogen is the ability to form a biofilm. P. gingivalis in a biofilm is 500 to 1,000 times less sensitive to antimicrobial drugs than planktonic cells (Conrads et al., 2021).
S. mutans possess several key virulence factors (e.g., many different surface biological structures, surface proteins and adhesins) critically involved in the etiology and pathogenesis of dental caries. Through its ability to adhere to solid surfaces, S. mutans efficiently colonizes the oral cavity and initiates the formation of dental biofilm also known as dental plaque. This biofilm is characterized by a matrix of exopolysaccharides, which significantly influence its physical architecture and biochemical properties, thereby facilitating bacterial persistence (Lemos et al., 2019). Additional traits that enhance the colonization potential of S. mutans include its pronounced acidogenic capacity (acid production which results in dental tissue demineralization) and its ability to interact synergistically or antagonistically with other microbial species within the oral ecosystem (Fang et al., 2024). Unlike the previously mentioned P. gingivalis, the main source of carbon and energy for this bacteria are carbohydrates. However, it was shown that peptides may also serve as additional nutrients, especially during biofilm growth, when carbohydrates originated from human food become depleted. They can be acquired through mucin proteolysis (Mothey et al., 2014). S. mutans also produce exoproteases that contribute to host tissue degradation (e.g., collagen) or enable quorum sensing (Hossain and Biswas, 2012; Huang et al., 2022); therefore, deterioration of A-PRF by S. mutans could be expected.
Our findings address existing knowledge gaps regarding the antibacterial activity of PRF against both pathogenic and opportunistic bacteria representing diverse taxonomic groups and morphological forms. These microorganisms are involved in a wide range of infections affecting the oral cavity, sinuses, and skin. By systematically analyzing the effects of PRF on bacteria with different cell wall structures and creating different forms, our study provides new insights into its spectrum of activity and highlights its potential clinical relevance.
5 Conclusion
The study revealed no significant differences in the composition of the oral microbiome between male and female participants assigned to the experimental group; however other factors not included into participants’ survey may comprise differences. Regardless of its form, PRF demonstrated antibacterial activity, with the strongest effect observed against the Gram-negative bacteria E. coli, and among Gram-positive species, the highest inhibition was noted for S. lentus. In both cases, the antibacterial effect was evident up to 7 h of incubation. For P. gingivalis, such an effect was observed only at low initial OD600 up to 24 h of incubation. No inhibitory effect of A-PRF membrane was observed against S. mutans. Notably, after 24 h of incubation, complete degradation of the A-PRF membrane occurred in cultures of P. gingivalis and S. mutans, which was not observed for other tested bacterial strains, which confirms our earlier hypothesis. The LP fraction of PRF exhibited no antibacterial activity against either P. gingivalis or S. mutans. These findings indicate that A-PRF membrane therapy will not prevent oral infections which may occur after oral surgical procedures. In the presence of signs of periodontitis, or other infection on site, appropriate antimicrobial treatment should first be administered to eliminate the risk of oral pathogens, such as, P. gingivalis proliferation, which may compromise the effectiveness and safety of PRF-based therapies.
Data availability statement
Data sets created during the study covering bacterial and fungal community structure (raw data from V3-V4 variable regions of 16S rRNA gene and ITS sequencing) have been deposited in publicly available repository under the doi number: https://doi.org/10.58132/J9YP72.
Ethics statement
The studies involving humans were approved by Ethics Committee of the Medical University of Warsaw. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
WP: Project administration, Funding acquisition, Supervision, Writing – review & editing, Conceptualization. DD: Writing – original draft, Investigation. DK: Investigation, Writing – original draft. RO: Writing – original draft, Investigation, Formal analysis, Visualization. MZ: Writing – original draft, Visualization, Formal analysis. MM-G: Writing – original draft, Investigation. AŁ: Writing – original draft. MP: Project administration, Visualization, Funding acquisition, Formal analysis, Conceptualization, Investigation, Methodology, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by funds from the Statutory Grant for the University of Warsaw and the Medical University of Warsaw and partially in the frame of the “Excellence Initiative—Research University (2020–2026)” Program at the University of Warsaw.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1691046/full#supplementary-material
References
Aleksijević, L. H., Aleksijević, M., Škrlec, I., Šram, M., Šram, M., and Talapko, J. (2022). Porphyromonas gingivalis virulence factors and clinical significance in periodontal disease and coronary artery diseases. Pathogens 11:1173. doi: 10.3390/pathogens11101173
Antimicrobial Resistance Collaborators (2022). Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 399, 629–655. doi: 10.1016/S0140-6736(21)02724-0
Bagdadi, E., Kubesch, K., Yu, A., Al-Maawi, X., Orlowska, S., Dias, A., et al. (2019). Reduction of relative centrifugal forces increases growth factor release within solid platelet-rich-fibrin (PRF)-based matrices: a proof of concept of LSCC (low speed centrifugation concept). Eur. J. Trauma Emerg. Surg. 45, 467–479. doi: 10.1007/s00068-017-0785-7
Benn, A., Heng, N., Broadbent, J. M., and Thomson, W. M. (2018). Studying the human oral microbiome: challenges and the evolution of solutions. Aust. Dent. J. 63, 14–24. doi: 10.1111/adj.12565
Biemer, J. J. (1973). Antimicrobial susceptibility testing by the Kirby-Bauer disc diffusion method. Ann. Clin. Lab. Sci. 3, 135–140.
Bolyen, E., Rideout, J. R., Dillon, M. R., Bokulich, N. A., Abnet, C. C., Al-Ghalith, G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9
Bostanci, N., Krog, M. C., Hugerth, L. W., Bashir, Z., Fransson, E., Boulund, F., et al. (2021). Dysbiosis of the human Oral microbiome during the menstrual cycle and vulnerability to the external exposures of smoking and dietary sugar. Front. Cell. Infect. Microbiol. 11:625229. doi: 10.3389/fcimb.2021.625229
Caselli, E., Fabbri, C., D’Accolti, M., Soffritti, I., Bassi, C., Mazzacane, S., et al. (2020). Defining the oral microbiome by whole-genome sequencing and resistome analysis: the complexity of the healthy picture. BMC Microbiol. 20:120. doi: 10.1186/s12866-020-01801-y
Castro, A. B., Meschi, N., Temmerman, A., Pinto, N., Lambrechts, P., Teughels, W., et al. (2017). Regenerative potential of leucocyte- and platelet-rich fibrin. Part A: intra-bony defects, furcation defects and periodontal plastic surgery. A systematic review and meta-analysis. J. Clin. Periodontol. 44, 67–82. doi: 10.1111/jcpe.12643
Chong, J., Liu, P., Zhou, G., and Xia, J. (2020). Using microbiome analyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 15, 799–821. doi: 10.1038/s41596-019-0264-1
Choukroun, J., Diss, A., Simonpieri, A., Girard, M.-O., Schoeffler, C., Dohan, S. L., et al. (2006). Platelet-rich fibrin (PRF): a second-generation platelet concentrate. Part IV: clinical effects on tissue healing. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 101, e56–e60. doi: 10.1016/j.tripleo.2005.07.011
Chumponsuk, T., Gruneck, L., Gentekaki, E., Jitprasertwong, P., Kullawong, N., Nakayama, J., et al. (2021). The salivary microbiota of Thai adults with metabolic disorders and association with diet. Arch. Oral Biol. 122:105036. doi: 10.1016/j.archoralbio.2020.105036
Clark, D., Rajendran, Y., Paydar, S., Ho, S., Cox, D., Ryder, M., et al. (2018). Advanced platelet-rich fibrin and freeze-dried bone allograft for ridge preservation: A randomized controlled clinical trial. J. Periodontol. 89, 379–387. doi: 10.1002/JPER.17-0466
Conrads, G., Klomp, T., Deng, D., Wenzler, J.-S., Braun, A., and Abdelbary, M. M. H. (2021). The antimicrobial susceptibility of Porphyromonas gingivalis: genetic repertoire, global phenotype, and review of the literature. Antibiotics (Basel) 10:1438. doi: 10.3390/antibiotics10121438
Cortellini, S., Castro, A. B., Temmerman, A., Van Dessel, J., Pinto, N., Jacobs, R., et al. (2018). Leucocyte- and platelet-rich fibrin block for bone augmentation procedure: A proof-of-concept study. J. Clin. Periodontol. 45, 624–634. doi: 10.1111/jcpe.12877
Curtis, M. A., Kuramitsu, H. K., Lantz, M., Macrina, F. L., Nakayama, K., Potempa, J., et al. (1999). Molecular genetics and nomenclature of proteases of Porphyromonas gingivalis. J. Periodontal Res. 34, 464–472. doi: 10.1111/j.1600-0765.1999.tb02282.x
Dohan Ehrenfest, D. M., Andia, I., Zumstein, M. A., Zhang, C.-Q., Pinto, N. R., and Bielecki, T. (2014). Classification of platelet concentrates (platelet-rich plasma-PRP, platelet-rich fibrin-PRF) for topical and infiltrative use in orthopedic and sports medicine: current consensus, clinical implications and perspectives. Muscles Ligaments Tendons J 4, 3–9.
Dong, K., Wu, K., Zheng, T., Yue, J., Wang, W., Luo, R., et al. (2021). Comparative study of Oral Bacteria and Fungi microbiota in Tibetan and Chinese Han living at different altitude. Tohoku J. Exp. Med. 254, 129–139. doi: 10.1620/tjem.254.129
Epand, R. M., Walker, C., Epand, R. F., and Magarvey, N. A. (2016). Molecular mechanisms of membrane targeting antibiotics. Biochim. Biophys. Acta Biomembr. 1858, 980–987. doi: 10.1016/j.bbamem.2015.10.018
Escapa, F., Huang, I., Chen, Y., Lin, T., Kokaras, M., Dewhirst, A., et al. (2020). Construction of habitat-specific training sets to achieve species-level assignment in 16S rRNA gene datasets. Microbiome 8:65. doi: 10.1186/s40168-020-00841-w
Fabbro, M. D., Bortolin, M., Taschieri, S., Ceci, C., and Weinstein, R. L. (2016). Antimicrobial properties of platelet-rich preparations. A systematic review of the current pre-clinical evidence. Platelets 27, 276–285. doi: 10.3109/09537104.2015.1116686
Fang, Y., Chen, X., Chu, C. H., Yu, O. Y., He, J., and Li, M. (2024). Roles of Streptococcus mutans in human health: beyond dental caries. Front. Microbiol. 15:1503657. doi: 10.3389/fmicb.2024.1503657
Farshidfar, N., Apaza Alccayhuaman, K. A., Estrin, N. E., Ahmad, P., Sculean, A., Zhang, Y., et al. (2025). Advantages of horizontal centrifugation of platelet-rich fibrin in regenerative medicine and dentistry. Periodontol. doi: 10.1111/prd.12625
Feng, M., Wang, Y., Zhang, P., Zhao, Q., Yu, S., Shen, K., et al. (2020). Antibacterial effects of platelet-rich fibrin produced by horizontal centrifugation. Int. J. Oral Sci. 12:32. doi: 10.1038/s41368-020-00099-w
Ghannoum, M. A., Jurevic, R. J., Mukherjee, P. K., Cui, F., Sikaroodi, M., Naqvi, A., et al. (2010). Characterization of the Oral fungal microbiome (Mycobiome) in healthy individuals. PLoS Pathog. 6:e1000713. doi: 10.1371/journal.ppat.1000713
Guo, Y., Nguyen, K.-A., and Potempa, J. (2010). Dichotomy of gingipains action as virulence factors: from cleaving substrates with the precision of a surgeon’s knife to a meat chopper-like brutal degradation of proteins. Periodontol. 54, 15–44. doi: 10.1111/j.1600-0757.2010.00377.x
Hansen, T. H., Kern, T., Bak, E. G., Kashani, A., Allin, K. H., Nielsen, T., et al. (2018). Impact of a vegan diet on the human salivary microbiota. Sci. Rep. 8:5847. doi: 10.1038/s41598-018-24207-3
Hoggard, M., Vesty, A., Wong, G., Montgomery, J. M., Fourie, C., Douglas, R. G., et al. (2018). Characterizing the human Mycobiota: A comparison of small subunit rRNA, ITS1, ITS2, and large subunit rRNA genomic targets. Front. Microbiol. 9:2208. doi: 10.3389/fmicb.2018.02208
Hossain, M. S., and Biswas, I. (2012). An extracelluar protease, SepM, generates functional competence-stimulating peptide in Streptococcus mutans UA159. J. Bacteriol. 194, 5886–5896. doi: 10.1128/JB.01381-12
Huang, B., Stewart, C. A., McCulloch, C. A., Santerre, J. P., Cvitkovitch, D. G., and Finer, Y. (2022). Streptococcus mutans proteases degrade dentinal collagen. Dentistry J. 10:223. doi: 10.3390/dj10120223
Jang, E.-S., Park, J.-W., Kweon, H., Lee, K.-G., Kang, S.-W., Baek, D.-H., et al. (2010). Restoration of peri-implant defects in immediate implant installations by Choukroun platelet-rich fibrin and silk fibroin powder combination graft. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 109, 831–836. doi: 10.1016/j.tripleo.2009.10.038
Janus, M. M., Crielaard, W., Volgenant, C. M. C., van der Veen, M. H., Brandt, B. W., and Krom, B. P. (2017). Candida albicans alters the bacterial microbiome of early in vitro oral biofilms. J. Oral Microbiol. 9:1270613. doi: 10.1080/20002297.2016.1270613
Jasmine, S., A, T., Janarthanan, K., Krishnamoorthy, R., and Alshatwi, A. A. (2020). Antimicrobial and antibiofilm potential of injectable platelet rich fibrin-a second-generation platelet concentrate-against biofilm producing oral staphylococcus isolates. Saudi J. Biol. Sci. 27, 41–46. doi: 10.1016/j.sjbs.2019.04.012
Koca, Ö. (2024). Microbiological characteristics of Bacillus subtilis species and their relationship with hospital infections. IntechOpen. doi: 10.5772/intechopen.115468
Łasica, A., Golec, P., Laskus, A., Zalewska, M., Gędaj, M., and Popowska, M. (2024). Periodontitis: etiology, conventional treatments, and emerging bacteriophage and predatory bacteria therapies. Front. Microbiol. 15:1469414. doi: 10.3389/fmicb.2024.1469414
Lemos, J. A., Palmer, S. R., Zeng, L., Wen, Z. T., Kajfasz, J. K., Freires, I. A., et al. (2019). The biology of Streptococcus mutans. Microbiol. Spectr. 7. doi: 10.1128/microbiolspec.gpp3-0051-2018
Li, X., Liu, Y., Yang, X., Li, C., and Song, Z. (2022). The Oral microbiota: community composition, influencing factors, pathogenesis, and interventions. Front. Microbiol. 13:895537. doi: 10.3389/fmicb.2022.895537
Līduma, I., Tračevska, T., Bērs, U., and Žileviča, A. (2012). Phenotypic and genetic analysis of biofilm formation by Staphylococcus epidermidis. Medicina (Kaunas) 48, 305–309.
Liu, R., Yan, M., Chen, S., Huang, W., Wu, D., and Chen, J. (2019). Effectiveness of platelet-rich fibrin as an adjunctive material to bone graft in maxillary sinus augmentation: A Meta-analysis of randomized controlled trails. Biomed. Res. Int. 2019, 1–10. doi: 10.1155/2019/7267062
Long, T., Li, C., Xu, F., and Xiao, J. (2024). Therapeutic efficacy of platelet-rich fibrin on surgical site wound healing in patients undergoing oral carcinoma resection: A meta-analysis. Int. Wound J. 21:e14386. doi: 10.1111/iwj.14386
Lu, Y., Zhou, G., Ewald, J., Pang, Z., Shiri, T., and Xia, J. (2023). Microbiome analyst 2.0: comprehensive statistical, functional and integrative analysis of microbiome data. Nucleic Acids Res. 51, W310–W318. doi: 10.1093/nar/gkad407
Madej, M., Nowakowska, Z., Ksiazek, M., Lasica, A. M., Mizgalska, D., Nowak, M., et al. (2021). PorZ, an essential component of the type IX secretion system of Porphyromonas gingivalis, delivers anionic lipopolysaccharide to the PorU Sortase for Transpeptidase processing of T9SS cargo proteins. MBio 12, e02262–20. doi: 10.1128/mbio.02262-20
Malanovic, N., and Lohner, K. (2016). Antimicrobial peptides targeting gram-positive bacteria. Pharmaceuticals (Basel) 9:59. doi: 10.3390/ph9030059
Mason, M. R., Preshaw, P. M., Nagaraja, H. N., Dabdoub, S. M., Rahman, A., and Kumar, P. S. (2015). The subgingival microbiome of clinically healthy current and never smokers. ISME J. 9, 268–272. doi: 10.1038/ismej.2014.114
Mazal, C., and Sieger, B. (2010). Staphylococcus lentus: the troublemaker. Int. J. Infect. Dis. 14:e397. doi: 10.1016/j.ijid.2010.02.502
Miron, R. J., Chai, J., Zheng, S., Feng, M., Sculean, A., and Zhang, Y. (2019). A novel method for evaluating and quantifying cell types in platelet rich fibrin and an introduction to horizontal centrifugation. J. Biomed. Mater. Res. A 107, 2257–2271. doi: 10.1002/jbm.a.36734
Morrison, A. G., Sarkar, S., Umar, S., Lee, S. T. M., and Thomas, S. M. (2023). The contribution of the human Oral microbiome to Oral disease: A review. Microorganisms 11:318. doi: 10.3390/microorganisms11020318
Mothey, D., Buttaro, B. A., and Piggot, P. J. (2014). Mucin can enhance growth, biofilm formation, and survival of Streptococcus mutans. FEMS Microbiol. Lett. 350, 161–167. doi: 10.1111/1574-6968.12336
Mueller, M., and Tainter, C. R. (2025). “Escherichia coli infection” in StatPearls (Treasure Island, FL: StatPearls Publishing).
Murugaiyan, V., Utreja, S., Hovey, K. M., Sun, Y., LaMonte, M. J., Wactawski-Wende, J., et al. (2024). Defining Porphyromonas gingivalis strains associated with periodontal disease. Sci. Rep. 14:6222. doi: 10.1038/s41598-024-56849-x
Nearing, J. T., DeClercq, V., Van Limbergen, J., and Langille, M. G. I. (2020). Assessing the variation within the oral microbiome of healthy adults. mSphere 5, e00451–e00420. doi: 10.1128/mSphere.00451-20
Pasquina-Lemonche, L., Burns, J., Turner, R. D., Kumar, S., Tank, R., Mullin, N., et al. (2020). The architecture of the gram-positive bacterial cell wall. Nature 582, 294–297. doi: 10.1038/s41586-020-2236-6
Pereira, V. B. S., Lago, C. A. P., Almeida, R. d. A. C., Barbirato, D. d. S., and Vasconcelos, B. C. d. E. (2023). Biological and cellular properties of advanced platelet-rich fibrin (A-PRF) compared to other platelet concentrates: systematic review and meta-analysis. Int. J. Mol. Sci. 25:482. doi: 10.3390/ijms25010482
Peters, B. A., Wu, J., Hayes, R. B., and Ahn, J. (2017). The oral fungal mycobiome: characteristics and relation to periodontitis in a pilot study. BMC Microbiol. 17:157. doi: 10.1186/s12866-017-1064-9
Polgár, L. (2002). The prolyl oligopeptidase family. CMLS Cell. Mol. Life Sci. 59, 349–362. doi: 10.1007/s00018-002-8427-5
Potempa, J., Pike, R., and Travis, J. (1997). Titration and mapping of the active site of cysteine proteinases from Porphyromonas gingivalis (gingipains) using peptidyl chloromethanes. Biol. Chem. 378, 223–230. doi: 10.1515/bchm.1997.378.3-4.223
Rafiei, M., Kiani, F., Sayehmiri, K., Sayehmiri, F., Tavirani, M., Dousti, M., et al. (2018). Prevalence of anaerobic Bacteria (P.gingivalis) as major microbial agent in the incidence periodontal diseases by Meta-analysis. J Dent (Shiraz) 19, 232–242.
Rajput, P., Nahar, K. S., and Rahman, K. M. (2024). Evaluation of antibiotic resistance mechanisms in gram-positive bacteria. Antibiotics 13:1197. doi: 10.3390/antibiotics13121197
Saadaoui, M., Singh, P., and Al Khodor, S. (2021). Oral microbiome and pregnancy: A bidirectional relationship. J. Reprod. Immunol. 145:103293. doi: 10.1016/j.jri.2021.103293
Sanz, M., Dahlin, C., Apatzidou, D., Artzi, Z., Bozic, D., Calciolari, E., et al. (2019). Biomaterials and regenerative technologies used in bone regeneration in the craniomaxillofacial region: consensus report of group 2 of the 15th European workshop on periodontology on bone regeneration. J. Clin. Periodontol. 46, 82–91. doi: 10.1111/jcpe.13123
Schär, M. O., Diaz-Romero, J., Kohl, S., Zumstein, M. A., and Nesic, D. (2015). Platelet-rich concentrates differentially release growth factors and induce cell migration in vitro. Clin. Orthop. Relat. Res. 473, 1635–1643. doi: 10.1007/s11999-015-4192-2
Shi, X., Qiu, S., Ji, L., Lu, H., Wu, S., Chen, Q., et al. (2023). Pathogenetic characterization of a Micrococcus luteus strain isolated from an infant. Front. Pediatr. 11:1303040. doi: 10.3389/fped.2023.1303040
Silhavy, T. J., Kahne, D., and Walker, S. (2010). The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2:a000414. doi: 10.1101/cshperspect.a000414
Simon, B. I., Zatcoff, A. L., Kong, J. J. W., and O’Connell, S. M. (2009). Clinical and histological comparison of extraction socket healing following the use of autologous platelet-rich fibrin matrix (PRFM) to ridge preservation procedures employing demineralized freeze dried bone allograft material and membrane. Open Dent. J. 3, 92–99. doi: 10.2174/1874210600903010092
Simonpieri, A., Del Corso, M., Vervelle, A., Jimbo, R., Inchingolo, F., Sammartino, G., et al. (2012). Current knowledge and perspectives for the use of platelet-rich plasma (PRP) and platelet-rich fibrin (PRF) in oral and maxillofacial surgery part 2: bone graft, implant and reconstructive surgery. Curr. Pharm. Biotechnol. 13, 1231–1256. doi: 10.2174/138920112800624472
Straub, A., Stapf, M., Utz, C., Vollmer, A., Flesch, J., Kübler, A., et al. (2024). Antimicrobial effects of clindamycin-loaded platelet-rich fibrin (PRF). Clin. Oral Investig. 28:144. doi: 10.1007/s00784-024-05532-6
Tohidnezhad, M., Varoga, D., Wruck, C. J., Podschun, R., Sachweh, B. H., Bornemann, J., et al. (2012). Platelets display potent antimicrobial activity and release human beta-defensin 2. Platelets 23, 217–223. doi: 10.3109/09537104.2011.610908
Verma, D., Garg, P. K., and Dubey, A. K. (2018). Insights into the human oral microbiome. Arch. Microbiol. 200, 525–540. doi: 10.1007/s00203-018-1505-3
Wade, W. G. (2021). Resilience of the oral microbiome. Periodontol. 86, 113–122. doi: 10.1111/prd.12365
Willis, J. R., and Gabaldón, T. (2020). The human oral microbiome in health and disease: from sequences to ecosystems. Microorganisms 8:308. doi: 10.3390/microorganisms8020308
Xiang, X., Shi, P., Zhang, P., Shen, J., and Kang, J. (2019). Impact of platelet-rich fibrin on mandibular third molar surgery recovery: a systematic review and meta-analysis. BMC Oral Health 19:163. doi: 10.1186/s12903-019-0824-3
Ximénez-Fyvie, L. A., Haffajee, A. D., and Socransky, S. S. (2000). Comparison of the microbiota of supra- and subgingival plaque in health and periodontitis. J. Clin. Periodontol. 27, 648–657. doi: 10.1034/j.1600-051x.2000.027009648.x
Xu, J., Ren, H., Zhao, S., Li, Q., Li, C., Bao, G., et al. (2023). Comparative effectiveness of hyaluronic acid, platelet-rich plasma, and platelet-rich fibrin in treating temporomandibular disorders: a systematic review and network meta-analysis. Head Face Med 19:39. doi: 10.1186/s13005-023-00369-y
Yilmaz, P., Parfrey, L. W., Yarza, P., Gerken, J., Pruesse, E., Quast, C., et al. (2014). The SILVA and “all-species living tree project (LTP)” taxonomic frameworks. Nucleic Acids Res. 42, D643–D648. doi: 10.1093/nar/gkt1209
Yoshino, Y. (2023). Enterococcus casseliflavus infection: A review of clinical features and treatment. Infect. Drug Resist. 16, 363–368. doi: 10.2147/IDR.S398739
Zaura, E., Keijser, B. J. F., Huse, S. M., and Crielaard, W. (2009). Defining the healthy “core microbiome” of oral microbial communities. BMC Microbiol. 9:259. doi: 10.1186/1471-2180-9-259
Zaura, E., Pappalardo, V. Y., Buijs, M. J., Volgenant, C. M. C., and Brandt, B. W. (2021). Optimizing the quality of clinical studies on oral microbiome: a practical guide for planning, performing, and reporting. Periodontol. 85, 210–236. doi: 10.1111/prd.12359
Keywords: platelet-rich fibrin, oral microbiome, antimicrobial activity, Porphyromonas gingivalis , periodontal diseases
Citation: Popowski W, Domanowska D, Koseski D, Ostrowski R, Zalewska M, Małecka-Giełdowska M, Łasica A and Popowska M (2025) Analysis of the oral microbiome composition of healthy individuals and the in vitro antibacterial activity of platelet-rich fibrin from these individuals against oral pathogenic bacteria. Front. Microbiol. 16:1691046. doi: 10.3389/fmicb.2025.1691046
Edited by:
Tzuen-Rong Tzeng, Clemson University, United StatesReviewed by:
Cristina Coelho, Instituto Superior de Ciências de Saúde do Norte (ISCS-N), PortugalAliaa Aboulela, Alexandria University, Egypt
Copyright © 2025 Popowski, Domanowska, Koseski, Ostrowski, Zalewska, Małecka-Giełdowska, Łasica and Popowska. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Magdalena Popowska, bWEucG9wb3dza2FAdXcuZWR1LnBs