 Moritz Gröschel
Moritz Gröschel Dietmar Basta1
Dietmar Basta1 Birgit Mazurek
Birgit Mazurek Agnieszka J. Szczepek
Agnieszka J. Szczepek- 1Department of Otolaryngology, Unfallkrankenhaus Berlin, Charité Medical School, Berlin, Germany
- 2Tinnitus Center, Berlin Institute of Health, Charité-Universitätsmedizin Berlin, Corporate Member of Freie Universität Berlin, Humboldt-Universität zu Berlin, Berlin, Germany
- 3Department of Otorhinolaryngology, Head and Neck Surgery, Berlin Institute of Health, Charité-Universitätsmedizin Berlin, Corporate Member of Freie Universität Berlin, Humboldt-Universität zu Berlin, Berlin, Germany
Noise that is capable of inducing the hearing loss (NIHL) has a strong impact on the inner ear structures and causes early and most obvious pathophysiological changes in the auditory periphery. Several studies indicated that intrinsic apoptotic cell death mechanisms are the key factors inducing cellular degeneration immediately after noise exposure and are maintained for days or even weeks. In addition, studies demonstrated several changes in the central auditory system following noise exposure, consistent with early apoptosis-related pathologies. To clarify the underlying mechanisms, the present study focused on the noise-induced gene and protein expression of the pro-apoptotic protease activating factor-1 (APAF1) and the anti-apoptotic B-cell lymphoma 2 related protein a1a (BCL2A1A) in the cochlear nucleus (CN), inferior colliculus (IC) and auditory cortex (AC) of the murine central auditory pathway. The expression of Bcl2a1a mRNA was upregulated immediately after trauma in all tissues investigated, whereas the protein levels were significantly reduced at least in the auditory brainstem. Conversely, acute noise has decreased the expression of Apaf1 gene along the auditory pathway. The changes in APAF1 protein level were not statistically significant. It is tempting to speculate that the acoustic overstimulation leads to mitochondrial dysfunction and induction of apoptosis by regulation of proapoptotic and antiapoptotic proteins. The inverse expression pattern on the mRNA level of both genes might reflect a protective response to decrease cellular damage. Our results indicate the immediate presence of intrinsic apoptosis following noise trauma. This, in turn, may significantly contribute to the development of central structural deficits. Auditory pathway-specific inhibition of intrinsic apoptosis could be a therapeutic approach for the treatment of acute (noise-induced) hearing loss to prevent irreversible neuronal injury in auditory brain structures and to avoid profound deficits in complex auditory processing.
Introduction
Over 5% of the world's population (360 million people) has disabling hearing loss, i.e., they have a hearing loss of at least 40 dB (WHO, 2015). Exposure to noise is a major cause of hearing loss, particularly in the industrialized countries, and is thought to be responsible for several audiological symptoms including tinnitus or hyperacusis (Le et al., 2017; Liberman, 2017).
Noise-Induced Structural Changes in Cochlea
A lot of research regarding noise-induced changes has been performed with a focus on physiological and anatomical properties of the auditory system, particularly in the auditory periphery. Noise-induced pathologies of the central auditory system were much less studied as the peripheral ones. Noise has a strong impact on inner ear structures and induces early and most obvious pathophysiological changes by directly damaging sensory tissue due to mechanical forces and the electrochemical hyperexcitation, followed by progressive degeneration extending toward primary auditory afferents (Pujol and Puel, 1999; Hu et al., 2002, 2006; Wang et al., 2002; Hirose and Liberman, 2003; Yang et al., 2004; Henderson et al., 2006; Weisz et al., 2009; Eskiizmir et al., 2011). Although necrosis was thought to be the major cause of cochlear cell death, it became evident that apoptotic pathways play as well a fundamental role (Saunders et al., 1985; Hu et al., 2000; Pirvola et al., 2000; Bohne et al., 2007). Several studies have investigated the mechanisms underlying cochlear pathologies, providing evidence that the acoustically induced, overwhelming production of reactive oxygen species (ROS) is the key factor immediately contributing to cellular degeneration after noise exposure (Hamernik et al., 1984; Hu et al., 2002; Yamashita et al., 2004a; Yang et al., 2004; Henderson et al., 2006). Such contribution was reported to last up to several weeks after the acoustic injury.
The gene classes, in which the early changes in the expression have been reported due to a loss of auditory function include immediate early genes, inflammatory and oxidative stress responses genes and the cell death pathway genes. The relation between the ROS production and the expressional activation of various genes seems to play a crucial role in this process and was suggested to be particularly involved in cochlear apoptosis through activation of several cell-death pathways (Van De Water et al., 2004; Gross et al., 2007; Mazurek et al., 2011; Dinh et al., 2015). The expression of pro- and anti-apoptotic genes in the peripheral auditory system (i.e., the cochlea) changes within a few hours or days after pharmacologically- or noise-induced insult. During the first hours following the otological damage, the intrinsic, mitochondrial apoptotic pathway seems to be particularly involved due to oxidative stress by generation of ROS (Ohlemiller et al., 1999; Henderson et al., 2006). Contribution of the intrinsic pathway was indicated by activation of specific caspases (Nicotera et al., 2003; Han et al., 2006; García-Berrocal et al., 2007) and by expressional regulation of genes belonging to the family of B-cell lymphoma 2 (Bcl2) genes (Hu et al., 2009; Furness, 2015) or other intrinsic apoptosis-related genes (Yamashita et al., 2004a,b; Kirkegaard et al., 2006; Ding et al., 2009). Similar expression pattern has been also shown to occur in the aging cochlea, contributing to age-related hearing loss (Alam et al., 2001; Nevado et al., 2006; Tadros et al., 2008; Someya et al., 2009; Falah et al., 2016).
Central Pathophysiology Induced by Hearing Loss
Studies of the central auditory system following noise exposure have determined several alterations in neuronal properties, including spontaneous and evoked hyperactivity, increased excitatory and reduced inhibitory neurotransmission, and tonotopic reorganization (Kaltenbach et al., 1998; Rajan and Irvine, 1998; Komiya and Eggermont, 2000; Wallhäusser-Franke et al., 2003; Basta and Ernst, 2005; Holt et al., 2005; Gröschel et al., 2011, 2014). Moreover, acoustic or mechanical trauma to the auditory periphery was shown to induce cellular degeneration in the central auditory pathway including cell and tissue shrinkage, axon degeneration, and neuronal cell loss. Such damage was shown to be present after initial damage and to persist for several days or weeks (Jean-Baptiste and Morest, 1975; Benson et al., 1997; Kim et al., 1997, 2004; Aarnisalo et al., 2000; Sekiya et al., 2009, 2012). Others have pointed at the appearance of plasticity markers in auditory brain immediately following the cochlear injury (Michler and Illing, 2002; Gil-Loyzaga et al., 2010). From these studies, it became evident that the lower ascending pathway—particularly the auditory brainstem—is primarily affected by cochlear injury, and that the observed changes may fundamentally affect the auditory function.
In healthy mice, histological studies revealed that the noise exposure leads to a reduction of cell density in the key structures of the central auditory system, whereby the first arising changes are most prominent in the auditory brainstem and then occur subsequently and to a lesser extend in the thalamic and cortical areas (Basta et al., 2005; Gröschel et al., 2010). Immunohistochemistry has identified apoptosis as one of main factors responsible for the above changes. Moreover, the time course of cell death supported the histological findings (Coordes et al., 2012; Fröhlich et al., 2017). A few studies investigated changes in the gene expression of the auditory brain in response to hearing loss. An altered expression of immediate early genes and neuroplasticity markers seems to play a particular role in generation of hyperactive neuronal disorders by changing synaptic neurotransmission (Wallhäusser-Franke et al., 2003; Oh et al., 2007; Kraus et al., 2009; Manohar et al., 2016; Tan et al., 2016). Further, expression of pro- and anti-apoptotic genes belonging to the Bcl2 family has been suggested to impair hippocampal cell proliferation during pharmacological induction of hearing loss and to possibly affecting pathophysiological processes in the inferior colliculus (Manohar et al., 2014). Long-term inflammatory responses seem to be present after noise-induced hearing loss in the cochlear nucleus (Manohar et al., 2016).
It still remains unclear to which extend the central auditory system can be affected by a noise exposure, and which signaling pathways play a major role during the pathological processes. Further, it is of high relevance to characterize the mechanisms underlying pathological processes, since they lead to negative changes in the central auditory system. These changes create in turn a basis for several hearing disorders by an induction of compensatory, irreversible physiological changes (neuronal reorganization, hyperactivity, hyperexcitability), accompanied by chronic audiological disorders (tinnitus, hyperacusis, reduced ability to process complex sounds, impaired speech processing). Due to the irreversible degeneration, late therapeutic interventions aimed at prevention of noise-induced pathologies are not feasible.
Our present work was designed to clarify the role of specific genes and proteins in the early degeneration of the central auditory system. Based on our recent studies that demonstrated cell loss in the auditory brain as well as on former observations of cochlear pathologies, the experiments started with commercial PCR cell death gene arrays with 84 genes in the cochlear nucleus (CN), inferior colliculus (IC) and auditory cortex (AC) and then particularly focused on the expression of the most frequently regulated anti-apoptotic gene Bcl2a1a (B-cell lymphoma 2 related protein a1a) and the related apoptosis-inducing gene Apaf1 (apoptotic protease activating factor-1). The particular aim was to analyze the possible contribution of mitochondria-related intrinsic apoptotic pathways on acute cellular damage of auditory brain structures immediately after traumatizing noise exposure. We have chosen the time point of 15 min following the 3 h of noise exposure based on our earlier, published studies. In addition to physiological changes (neuronal hyperactivity and increased calcium-related activity), we have determined the changes in cell numbers and the types of cell death pathways in several structures of the central auditory system. Our results implied degenerative process occurring after noise exposure. Our most important finding was that the degeneration was detectable immediately after finishing acoustic trauma, which was shown for the first time in these kinds of experiments (Gröschel et al., 2010, 2011, 2014; Coordes et al., 2012; Fröhlich et al., 2017).
The results of this study should help to fill the gap of knowledge between sound-related hyperexcitation of the auditory pathway, i.e., the induction of hearing loss, and central neurodegeneration.
Materials and Methods
Animals
Eighteen 6 weeks-old female normal hearing mice (NMRI strain) were used in this study. The experimental protocol was approved by the governmental Ethics Commission for Animal Welfare (LaGeSo Berlin, Germany; approval number: G0416/10). This study was carried out in accordance with the recommendations of the EU Directive 2010/63/EU on the protection of animals used for scientific purposes.
Experimental Groups and Tissue Preparation
In total, nine animals (trauma group) were anesthetized (60 mg/kg ketamine and 6 mg/kg xylazine) and noise-exposed in a soundproof chamber (0.8 × 0.8 × 0.8 m, minimal attenuation 60 dB) for 3 h to broadband white noise (5–20 kHz) at 115 dB sound pressure level (SPL). The selected noise trauma paradigm is in accordance with what is known about the hearing range of mice (≥1 and ≤100 kHz) and people (≥20 Hz and ≤20 kHz), covering the mouse low-frequency hearing range, and is based on our earlier studies on cellular degeneration and detection of cell death mechanisms after noise exposure to ensure similar experimental conditions (Heffner and Heffner, 2007; Gröschel et al., 2010; Reynolds et al., 2010; Coordes et al., 2012; Fröhlich et al., 2017). Noise was delivered binaurally by speakers (HTC 11.19; Visaton, Haan, Germany) placed above the animal's head. The speakers were connected to an amplifier (Tangent AMP-50; Aulum, Denmark) and a DVD player. SPL was calibrated by using a sound level meter (Voltcraft 329; Conrad Electronic, Hirschau, Germany) placed near the animal's ear. The efficacy of anesthesia was controlled by using a video camera placed inside the lighted chamber. Body temperature was maintained at 37°C with a heating pad (Thermolux CM 15 W; Acculux, Murrhardt, Germany) placed under the animal. Another nine animals served as a normal hearing control group. The control animals were treated equally, that is they were anesthetized and kept in the chamber on a heating pad, but were not exposed to noise (sham exposed group).
Mice were investigated within 15 min after the noise/sham exposure. First, they were euthanized by a lethal dose of anesthetics and next they were decapitated. Brains were removed carefully from the skull, rinsed with PBS and transferred immediately into Allprotect Tissue Reagent (Qiagen, Hilden, Germany cat # 76405) at room temperature for further preparation. Tissue samples from cochlear nucleus (CN, dorsal and ventral subdivision), inferior colliculus (IC) and auditory cortex (AC) from both hemispheres were harvested. Based on our long-standing experience in anatomy and histology of the mouse central auditory system, the boundaries of the investigated structures have been recognized by their individual shape and were gently dissected under visual microscopic control and counterchecked in accordance to a mouse brain atlas (Paxinos and Franklin, 2001). For each of the different structures, tissues harvested from three animals of each group were pooled together and stored at −20°C until RNA and protein isolation. Three separate sets of experiments were performed and each included three noise-exposed animals and three controls, respectively. The mRNA isolated during the first and second experiment was used for qRT-PCR. Target protein levels were estimated using tissues from all three sets of experiments. During all experiments, noise trauma was initiated at the same time in the morning to avoid different noise impact due to diurnal variation in noise susceptibility (Meltser et al., 2014).
RNA and Protein Isolation, Reverse Transcriptase Reaction
The tissues stored at −20°C were slowly brought to the room temperature (RT) and the Allprotect reagent was removed using Eppendorf pipette with RNAse-free tip–a total volume of 50 μl was left in each tube. Next, 600 μl of RLT buffer (part of the RNeasy Mini Kit, Qiagen, cat. # 74106) was added to the samples followed by homogenization, vortexing, pulse centrifugation and transfer into QIAshredder Homogenizers (Qiagen, cat. # 79656). Following 2 min of centrifugation at 14,000 rpm, the samples were left at RT for 10 min. The mRNA was precipitated with 650 μl of 70% ethanol and the precipitate was brought to the bottom of the tube by 20 s centrifugation at 8,000 rpm. The rest of the RNA isolation was performed strictly according to the manufacturer's instructions.
The eluate, which contained protein, was collected into fresh tube and stored at −80°C until protein purification procedure. When needed, the samples were mixed with 3 ml of ice-cold acetone and incubated at −20°C for 45 min followed by centrifugation for 20 min at 10.000 rpm. The supernatant was aspirated with Eppendorf pipette and the pellet was dried at RT for 20 min. After this, the pellet was resuspended in 40 μl RIPA buffer containing 5% SDS and dissolved in thermal shaker at 95°C and 500 rpm for 8 min. Next, 80 μl of RIPA buffer was added and the samples were centrifuged in a pre-warmed Eppendorf centrifuge at 24°C, 14.000 rpm for 10 min. The supernatant was collected into fresh tube and the protein concentration was determined with Micro BCA Protein Assay Kit, (ThermoScientific, Darmstadt, Germany cat. # 23235).
The reverse transcription reaction was performed with the RT2 First Strand Kit, (Qiagen cat. # 330401) according strictly to the manufacturer's instructions. The RNA concentration was measured using NanoDrop and 200 ng of total RNA was used to synthesize cDNA. Thermal cycler 2720 (Applied Biosystems) was used for sample incubation.
RT2-PCR Array Experiments
The noise-induced expression of 84 cell death related genes have been profiled using a commercial PCR array in the first part of experiments. The cDNA templates were mixed with ready-to-use SYBR Green Mastermix (RT2 SYBR Green qPSR Mastermix, Qiagen, Cat. No. 330502), and loaded onto the 96-well RT2-PCR array (PAMM-212ZF-6- RT2 Profiler™ PCR Array Mouse Cell Death Pathway Finder, Qiagen, Cat. No. 330231; see Table 1 for detailed list of genes). In the first set of experiments, 49.50 ng of cDNA per well was used for the AC, 61.39 ng of cDNA per well was used for the IC and 29.70 ng of cDNA per well was used for the CN. In the second set of experiments the amounts of cDNA used were different and consisted of 56.44 ng of cDNA per well for the AC, 46.53 ng of cDNA per well for the IC and 36.63 ng of cDNA per well for the CN. Real-time PCR detection was initiated by heating at 95°C for 10 min, followed by 45 cycles of 95°C for 15 s and 60°C for 1 min. The cycling was performed in LightCycler® 96 Instrument (Roche, Mannheim, Germany) and the threshold value Ct was determined by LightCycler® 96 Application and Instrument Software (Roche).
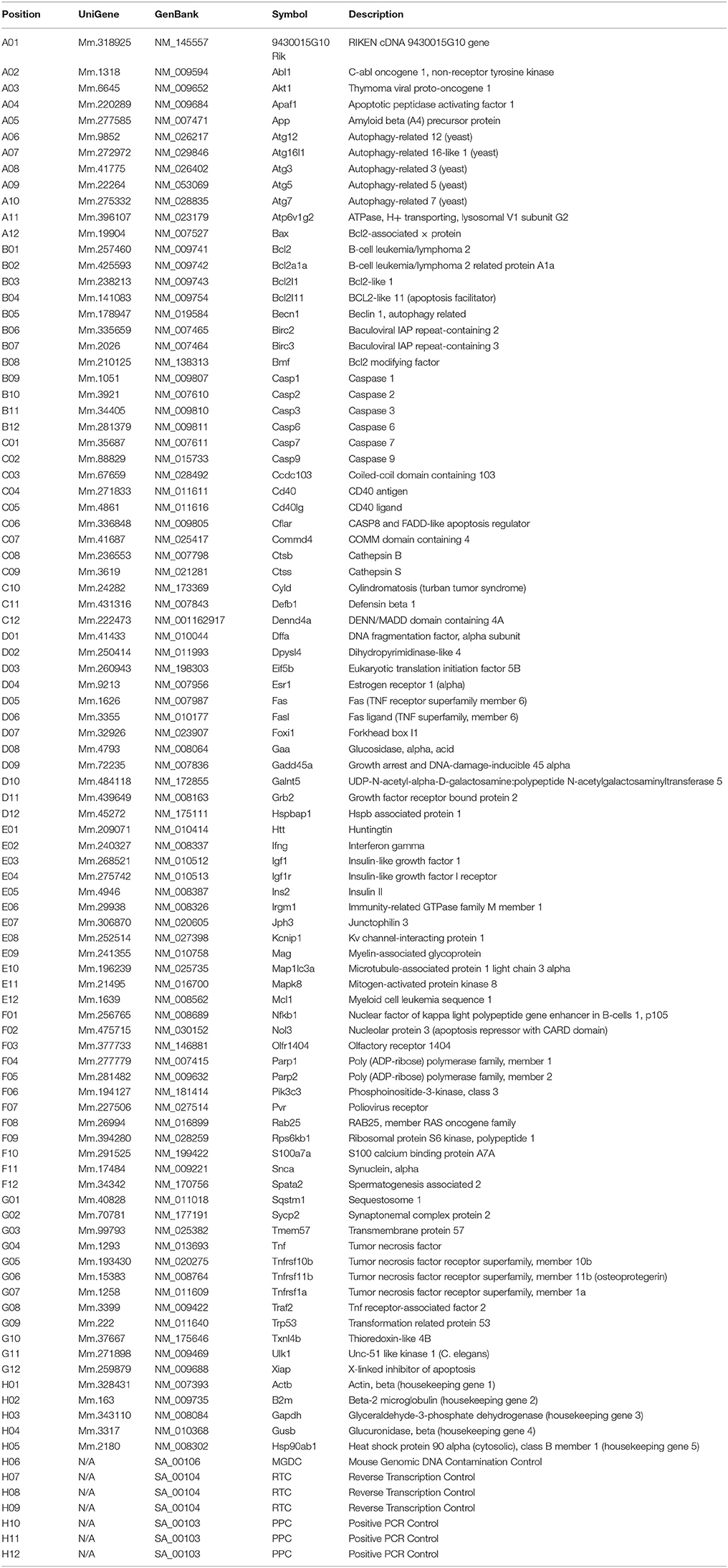
Table 1. Gene table RT2 PCR array.
Changes in mRNA levels were expressed as fold increase/decrease. The cut-off determining expression was ≥2.0 or ≤–2.0 fold changes. Genes, which met these criteria, were considered to be upregulated or downregulated.
Of the 84 cell death related genes monitored for mRNA expression in this part of the experiments, Bcl2a1a and Apaf1 were selected for further detailed quantitative analysis of noise-induced alterations in mRNA and protein levels. During all experiments, expression of three housekeeping genes present on each of the arrays was used for normalization: beta-actin, beta-2 microglobulin and heat shock protein 90 alpha (cytosolic), class B member 1.
RT2-qPCR Primer Experiments
The sets of forward and reverse primers specific for Apaf1 and Bcl2a1a were purchased from Qiagen. These primers sets were identical to the ones spotted on the expression arrays and their sequence is protected by patent. We used the primers pairs to perform confirmatory RT-PCR using this time not the pooled but individual cDNA samples. The cycling conditions were identical to the conditions used for the arrays.
Protein Analyses
The protein concentration was measured and samples containing were mixed with Roti-Load 1 (Carl Roth, Karlsruhe, Germany, cat. #K929.1). Samples containing 16 μg of total protein were pipetted into the wells of SDS-PAGE gel (Novex WedgeWell 4–20% Tris-Glycine Gel, 12 well, Thermo Fisher Scientific, cat. # XPO4202BOX). PageRuler Prestained Protein Ladder was used to estimate the molecular weight (Thermo Scientific cat. # 26618). Following the electrophoresis in Xcell SureLock Electrophoresis Cell, Invitrogen, the proteins were blotted onto Immobilon-P Transfer Membrane 0.45 μm, (Millipore cat. # IPFL 000 10) with use of XCell II Blot Module (Invitrogen cat. # EI9051). The membrane was blocked in 5% skimmed milk/PBS/0.05%Tween buffer for 1 h after which, incubation with primary and secondary antibodies followed. To detect 39 kDa BCL2A1A, a rabbit polyclonal antibody was used (Abcam # ab45414) in dilution of 1:5000. To detect 130 kDa APAF1, rabbit polyclonal antibody (Enzo # ADI-905-179) was used in dilution of 1:500. The blots were incubated with primary antibodies overnight at 4°C, followed by triple wash in PBS/0.1%Tween. Secondary antibody used was in both cases goat-anti-Rabbit IgG (H+L), conjugated with horseradish peroxidase (Promega, cat. # W4011) at the dilution of 1:5000. Protein was visualized by adding Pierce ECL Western Blotting Substrate (Thermo Scientific cat. # 32209) and scanning by C-DiGit Blot Scanner, LI-COR (Bad Homburg, Germany), followed by image analyses with Image Studio Version 5x. The unique technology of C-Digit allows direct measurement of chemiluminescence induced by a presence of enzyme-labeled secondary antibody bound to the target protein blotted on a membrane and therefore, allows a reduction of one experimental step and improves accuracy of the result.
Statistical Analyses
The data obtained from the noise-exposure animals were compared to data obtained from the control group in each investigated brain region. Delta Cq values from the expression arrays and calculated ratios to controls obtained from the western blot were used for statistical comparison between BCL2A1A and APAF1 groups. Depending on data distribution, U-test (not normally distributed data) or t-test (normally distributed data) was used. Data distribution was tested by Shapiro-Wilk test. The software SPSS (IBM SPSS Statistics Version 23, IBM Corp., Armonk, New York, USA) was used for all statistical analyses. The alpha level of significance was set for <0.05. Data are presented as mean ± standard deviation.
Results
RT2-PCR Array Experiments
In the first set of experiments, commercial RT-PCR expression arrays were used to determine changes in the expression of 84 cell death-related genes (Table 1). Upregulation (≥2.0 fold expression changes) of gene expression was observed only for one gene (Hspbap1) in the inferior colliculus and for one gene (Hspbap1) in the primary auditory cortex, whereas no changes have been detected in the cochlear nucleus. Expressional downregulation (≤–2.0 fold expression changes) affected three genes (Cd40, Esr1, Galnt5) in the CN, 11 genes (Akt1, Apaf1, Bcl2a1a, Casp6, Fas, Galnt5, Igf1, Ins2, Sycp2, Tmem57, Tnf) in the IC and eight genes (Bcl2a1a, Bmf, Cd40, Defb1, Fas, Fasl, Foxi1, Olfr1404) in the AC.
In the second set of experiments using a different amount of cDNA, we detected an upregulation of 12 genes (Atg5, Bcl2a1a, Bcl2l11, Casp1, Casp6, Ctsb, Dpysl4, Eif5b, Igf1, Ins2, Mag, Rab25) in the CN, 13 genes (Akt1, Atg7, Atp6v1g2, Bcl2l11, Casp1, Foxi1, Hspbap1, Mag, Parp1, S100a7a, Snca, Tmem57, Tnfrsf1a) in the IC and for 15 genes (Bcl2a1a, Bcl2l1, Birc2, Bmf, Cd40, Cd40lg, Defb1, Fas, Fasl, Gaa, Galnt5, Ifng, Kcnip1, Mag, Olfr1404) in the AC. Downregulation was present for 2 genes (Apaf1, Defb1) in the CN, for 17 genes (Abl1, Bcl2a1a, Bmf, Casp6, Cd40, Cd40lg, Cflar, Defb1, Dffa, Fas, Gadd45a, Galnt5, Ifng, Igf1, Ins2, Olfr1404, Rab25) in the IC and 6 genes (Casp1, Ccdc103, Foxi1, Ins2, S100a7a, Snca) in the auditory cortex (see Table 1 for list of abbreviations).
For detailed documentation, data of gene expression analysis from both sets of experiments is available in the Supplementary Material (Supplementary Tables 1–6).
Confirmatory RT-PCR Experiments
Gene expression analysis demonstrated significant changes in the expression of both investigated genes, Bcl2a1a and Apaf1, following an acute acoustic overstimulation (trauma group, 6 animals, number of samples: n = 8) compared to an unexposed control group (6 animals, number of samples: n = 8). These particular genes were selected as the anti-apoptotic gene Bcl2a1a was the most frequently regulated gene in both sets of RT2-PCR array experiments and the pro-apoptotic Apaf1 gene represents one of the central stages in the related apoptosis regulatory network.
The expression of Bcl2a1a was upregulated in all investigated structures of the central auditory pathway (Figure 1). In the cochlear nucleus, mean ΔCq value was significantly greater in the trauma group 12.089 ± 0.578 than in control animals 10.895 ± 0.307 (p < 0.001; t-test; t = −5.073; df = 14; effect size d: 2.580; power: 0.999). Similar Bcl2a1a expression pattern could be seen in the inferior colliculus as well as in the auditory cortex. In the IC, mean ΔCq values were 11.539 ± 0.687 in the controls and 13.644 ± 1.413 in the noise-exposed animals [p = 0.002; t-test (t = −3.789; df = 14; effect size d: 1.895; power: 0.975)]. In the auditory cortex, ΔCq values rose from a mean ΔCq of 13.283 ± 2.063 in the control group to 15.095± 0.650 in the noise-exposed group [p = 0.044; t-test (t = −2.370; df = 14; effect size d: 1.185; power: 0.728)].
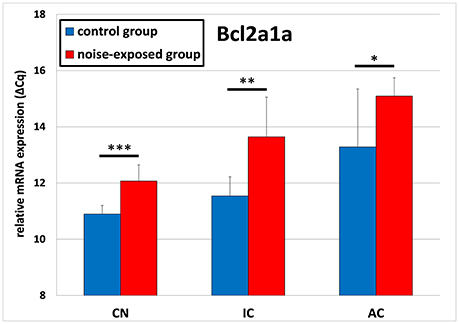
Figure 1. Relative mRNA expression of Bcl2a1a. Presented are ΔCq values (mean ± S.D.) in the cochlear nucleus (CN), inferior colliculus (IC) and auditory cortex (AC) of the mouse central auditory pathway immediately after noise trauma compared to unexposed controls. Asterisks indicate significant differences between control and trauma group (*p < 0.05; **p < 0.01; ***p < 0.001).
The gene expression analysis of Apaf1 delivered opposite results (Figure 2). The ΔCq values in the noise-exposed group were significantly lower as these in the control group. In detail, cochlear nucleus of the control group had the mean ΔCq of 9.428 ± 0.359 whereas cochlear nucleus of the noise-exposed animals had the mean ΔCq of 8.525 ± 0.282 [p < 0.001; t-test (t = 5.595; df = 14; effect size d: 2.797; power: 1)]. Similarly, the mean ΔCq in inferior colliculus of the control group was 8.710 ± 0.207 whereas in the noise-exposed group ΔCq was 8.141 ± 0.418 [p = 0.004; t-test (t = 3.450; df = 14; effect size d: 1.725; power: 0.947)]. No significant differences in the expression of Apaf1 gene have been observed between both groups in the auditory cortex [control group: 7.896 ± 0.211, trauma group: 7.860 ± 0.238; p = 0.752; t-test (t = 0.322; df = 14)].
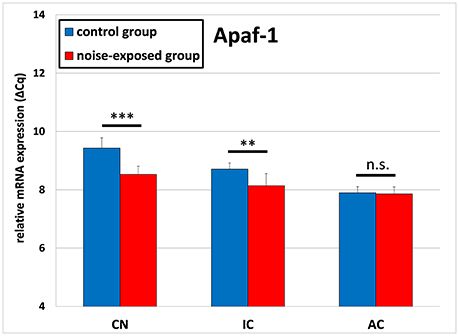
Figure 2. Relative mRNA expression of Apaf1. Presented are ΔCq values (mean ± S.D.) in the cochlear nucleus (CN), inferior colliculus (IC) and auditory cortex (AC) of the mouse central auditory pathway immediately after noise trauma compared to unexposed controls. Asterisks indicate significant differences between control and trauma group (**p < 0.01; ***p < 0.001; n.s., no significant difference between groups).
Western Blot Experiments
The relative protein levels of BCL2A1A and APAF1 were compared between the noise-exposed group (9 animals, number of samples: n = 12) and the control group (9 animals, number of samples: n = 12) for each investigated structure separately (see Supplementary Figure 1 in Supplementary Material for original data from the western blot experiments).
BCL2A1A protein levels were shown to be significantly decreased in response to acoustic trauma in the cochlear nucleus. No significant changes could be detected in the inferior colliculus or auditory cortex (Figure 3). In the CN, relative mean ratio of experimental group to control (1.0 ± 0.053) was 0.893 ± 0.116, which was significantly less than the control [p = 0.008; t-test (t = 2.901; df = 22; effect size d: 1.187; power: 0.942)]. For the inferior colliculus, there were no statistically significant differences between groups [control group: 1.0 ± 0.049, noise-exposed group: 1.034 ± 0.127; p = 0.908; U-test (U = 70; df = 22)]. The same holds true for the auditory cortex, showing a slight but not significant reduction in protein amount ratio between groups [control group: 1.0 ± 0.065, noise-exposed group: 0.934 ± 0.139; p = 0.147; t-test (t = 1.502; df = 22)].
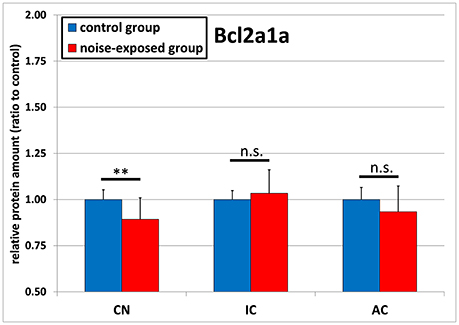
Figure 3. Relative protein amounts of BCL2A1A. Presented are ratios to respective controls from the Western blot experiments (mean ± S.D.) in the cochlear nucleus (CN), inferior colliculus (IC) and auditory cortex (AC) of the mouse central auditory pathway immediately after noise trauma compared to unexposed controls. Asterisks indicate significant differences between control and trauma group (**p < 0.01; n.s., no significant difference between groups).
The protein levels of APAF1 were slightly greater in all investigated structures of the ascending central auditory pathway, although missing statistical significance (Figure 4). In the inferior colliculus, statistical comparison of mean expression ratio between noise-exposed animals and unexposed controls showed p-value of 0.074 [control group: 1.0 ± 0.085, trauma group: 1.064 ± 0.082; t-test (t = −1.874; df = 22)]. In addition, this difference was also not statistically significant for cochlear nucleus [control group: 1.0 ± 0.041, noise-exposed group: 1.083 ± 0.325; p = 0.396; t-test (t = −0.882; df = 22)] or for the auditory cortex [control group: 1.0 ± 0.079, noise-exposed group: 1.079 ± 0.247; p = 0.773; U-test (U = 67; df = 22)].
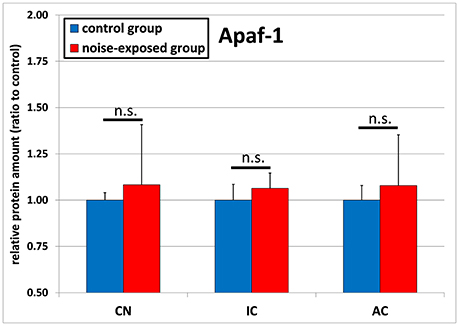
Figure 4. Relative protein amounts of APAF1. Presented are ratios to the respective controls from the Western blot experiments (mean ± S.D.) in the cochlear nucleus (CN), inferior colliculus (IC) and auditory cortex (AC) of the mouse central auditory pathway immediately after noise trauma compared to unexposed controls (n.s., no significant difference between groups).
Discussion
The present study investigates acute noise-induced changes in the expression of genes encoding members of the intrinsic apoptotic pathway, Bcl2a1a and Apaf1 in the key structures of the auditory pathway: cochlear nucleus, inferior colliculus and auditory cortex. Moreover, we investigated changes in the protein levels of BCL2A1A and APAF1 in the same structures. As it was the aim of this study to determine gene and protein expression immediately after noise exposure, therefore the auditory function in these particular animals could not be tested. However, in our earlier studies using the same noise exposure protocol (covering the mouse low-frequency hearing range) in NMRI mice, we observed greatest hearing loss immediately after noise exposure (mean threshold shift between 4 and 20 kHz: ≥60 dB). The hearing thresholds recovered partially over 1 week, and the animals were left with a stable mean permanent threshold shift of 38 dB between 4 and 20 kHz (Gröschel et al., 2011). Moreover, several other earlier studies reported that identical treatment caused the development of spontaneous neuronal hyperactivity (possibly related to tinnitus) as well as an acute and long-term upregulation in calcium activity and cellular degeneration in auditory brain structures (Gröschel et al., 2010, 2011, 2014; Coordes et al., 2012; Fröhlich et al., 2017). Our results demonstrated upregulation of Bcl2a1a gene expression immediately after acoustic trauma in all investigated structures, whereas the protein levels were reduced at least in the auditory brainstem. On the other hand, the transcriptional expression of Apaf1 mRNA was reduced after acoustic overexposure along the auditory pathway. The protein levels of APAF1 tend to be increased but without statistical significance.
Degeneration in the Auditory System
Apoptosis, the process of programmed cell death, occurs normally during development, aging or as a homeostatic mechanism and is essential for multicellular organisms. However, despite other cell death pathways, apoptosis might also occur in pathological conditions and is a key process contributing to neurodegenerative diseases, ischemia and autoimmune disorders (Elmore, 2007). Hearing disorders occurring due to acoustic trauma, pharmacological exposure or due to direct damage in the auditory periphery have the ability to induce apoptosis not only in the cochlear structures but also in the auditory nerve and the central auditory system (Coordes et al., 2012; Yu et al., 2014; Ding et al., 2015; Bohne et al., 2017). Although acute and long-term necrosis has been detected and the expression of necrosis-related genes changes, apoptosis seems to be a key mediator of cell death in NIHL particularly induced by high frequency noise at high sound intensities (Hu et al., 2000, 2002; Coordes et al., 2012; Yu et al., 2014; Bohne et al., 2017). Studies of the peripheral and central auditory system have shown the impact of noise on auditory tissues already within 1 h after exposure, being progressive for weeks and months. The degenerative effects are most prominent in cochlear outer hair cells followed by inner hair cells, supporting cells and further delayed negative effect along the central auditory pathway (Hamernik et al., 1984; Yang et al., 2004; Gröschel et al., 2010; Meltser et al., 2010; Coordes et al., 2012; Bohne et al., 2017; Fröhlich et al., 2017). Several studies investigated the expression of different cell death-related genes during cochlear degeneration to give insight into the underlying mechanisms.
Early Pathologies in the Cochlea
Apoptotic mechanisms have been characterized during exposure of cochlea to pharmacological agents in vitro and in vivo. Cisplatin-treated cochlear explants express several pro- and anti-apoptotic genes, particularly p53 and the members of Bcl2 family. In addition, caspase activation and an increase in stress response proteins within a few hours after treatment has been demonstrated (Devarajan et al., 2002; Coling et al., 2007; Ding et al., 2009; Wang et al., 2013). In vivo experiments with ototoxic drugs indicated a large contribution of genes and proteins related to intrinsic apoptosis signaling pathway (García-Berrocal et al., 2007). Moreover, an early increase in oxidative stress responses, contributing to particular cell death pathways, and accompanied by apoptotic and inflammatory reactions on the gene level in cochlear tissue, has been described in cochlea (Kirkegaard et al., 2006; Mazurek et al., 2011; Gross et al., 2014).
Also noise exposure has been shown to induce early changes in the expression of several pro- and anti-apoptotic genes in cochlea (Han et al., 2011). Noise exposure leads to neuronal hyperexcitation, which is followed by ischemia-induced generation of ROS and initiation of apoptosis in the auditory system. Oxidative stress that follows and plays an important role during intrinsic apoptosis, is a key mechanism responsible for acquired hearing loss (Henderson et al., 2006). On the cochlear level, an acute upregulation of pro-apoptotic TNF family members and a downregulation of anti-apoptotic Bcl2-related genes was demonstrated 4 h after noise exposure (Hu et al., 2009). On the other hand, an upregulation of anti-apoptotic genes was present within 1 h after sound overexposure (Han et al., 2012).
It is a common feature of a large number of early regulated genes in evoked hearing loss that they are involved in the intrinsic apoptotic pathway. A detailed examination of caspase activity in the peripheral auditory system supports the idea about the intrinsic apoptotic pathway being particularly responsible for the ototoxic and age-related cell death (upregulation of caspases 3 and 9, participation of pro-apoptotic BAK-dependent pathway), while both, the extrinsic and intrinsic pathway, contribute to the noise-induced hearing loss (activation of caspases 3, 8, and 9) (Hu et al., 2002; Nicotera et al., 2003; Van De Water et al., 2004).
Comparable changes of intrinsic pro- and anti-apoptotic signaling, including regulation of BCL2 family members and APAF1, were also shown during age-related hearing loss (ARHL) (Alam et al., 2001; Nevado et al., 2006; Tadros et al., 2008; Falah et al., 2016).
Intrinsic Apoptosis and the Central Auditory System
While the extrinsic apoptotic pathway is induced by transmembrane receptor interactions involving death ligands and corresponding death receptor domains of the tumor necrosis factor family (Ashkenazi and Dixit, 1998), the intrinsic apoptotic pathway is induced by a non-receptor-mediated stimuli, acting directly on intracellular targets and being initiated in the mitochondria (Elmore, 2007). Activation of intrinsic apoptosis occurs through changes at the inner mitochondrial membrane, which alters the transmembrane potential. The release of cytochrome c from mitochondria into the cytosol activates APAF1 to facilitate the formation of apoptosome, which in turn triggers proteolytic caspase cascades (pro-caspase 9, caspase 9 and “executioner” caspase 3) and leads to apoptosis (Sakahira et al., 1998; Hill et al., 2004; Saelens et al., 2004). APAF1 is a key molecule in the intrinsic mitochondrial pathway and conformational changes occur by binding of cytochrome c (Jiang and Wang, 2000; Shakeri et al., 2017). The direct involvement of APAF1 in ototoxicity was shown by diminishing the chemotherapy-related hearing loss trough pharmacological inhibition of APAF1 (Orzáez et al., 2014). The levels of APAF1 protein are generally low and thus, are a limiting factor in apoptosome formation and apoptotic signaling. Therefore, even a slight increase demonstrated in the present study could substantially contribute to apoptosis (Yoshida et al., 1998; Shakeri et al., 2017).
The BCL2 family consists of several pro- and anti-apoptotic genes and related proteins. BCL2 family members, including anti-apoptotic BCL2A1A, alter mitochondrial membrane permeability (Cory and Adams, 2002). Pro-apoptotic BCL2-related proteins (including BAX, BAK and BH3-only proteins) have the ability to induce the formation of pores in the outer mitochondrial membrane and to mediate cytochrome c release, which is the critical step in the intrinsic apoptotic pathway (Vogler, 2012). Imaging experiments have shown that the neuronal calcium uptake is strongly increased immediately after acoustic trauma, particularly in the auditory brainstem (Gröschel et al., 2011). Calcium plays an important role in mitochondrial homeostasis and is a key mediator of neuronal ischemia-induced excitotoxicity. Therefore, neuronal hyperexcitation can trigger intrinsic apoptosis by directly increasing the level of oxidative stress and ROS formation; thus, further influencing mitochondrial membrane potential (Yu et al., 2001; Salinska et al., 2005; Mattson, 2007).
Anti-apoptotic proteins of the BCL2 family generally counteract the activation of pro-apoptotic proteins. In the present study, this effect might be represented by the strong increase of Bcl2a1a gene expression. Such upregulated transcriptional expression may prevent further apoptosis by subsequently increasing BCL2A1A protein synthesis. According to the present results, an increase of intrinsic anti-apoptotic Bcl2a1 a signaling was demonstrated in the inferior colliculus 2 days after exposure to cisplatin (Manohar et al., 2014). Upregulation of anti-apoptotic genes, often along with increased stress protein responses, are supposed to represent a cellular mechanism of self-defense in response to pro-apoptotic gene and protein expression following acoustic injury, compensating for activation of cellular damage and balancing pro- and anti-apoptotic machineries (Wada and Penninger, 2004; Coling et al., 2007; Someya et al., 2009; Portt et al., 2011; Wang et al., 2013). This notion is supported by studies showing an acute upregulation of heat shock proteins in response to noise-induced peripheral and central degeneration, further initiating recovery from acutely developing central hyperactivity and promoting cell survival (Helfert et al., 2002; Samson et al., 2007; Sun et al., 2008).
Changes in levels of particular mRNA and proteins are related to each other in a time-dependent manner, meaning that greater amount of a particular protein could induce downregulation of the respective gene transcription (and vice versa) representing a homeostatic mechanism to rebalance the cellular proteome, referred to as proteostasis (Balch et al., 2008; Douglas and Dillin, 2010). The changes in gene and protein expression do not occur simultaneously and there may be substantial time difference between visible increase in gene expression and that of protein expression. The time window, which we studied, was rather narrow and consisted of merely 3 h. In case of possible gene expression activation, the protein synthesis may follow significantly later (Hargrove et al., 1991). For instance, we have shown earlier that the induction of HSP72 gene expression occurs already 2 h after exposure to geldanamycin, whereas statistically different protein synthesis follows 4 h after induction (Yu et al., 2009). In case of gene expression inhibition, then this also not necessarily has to be mirrored by an immediate decrease of protein concentration. In fact, half-life of most of the proteins can be measured in hours whereas that of mRNA – in minutes or even seconds, depending on if and how is mRNA protected (Friedel et al., 2009). This concept is in line with the present results, since in our study, the levels of anti-apoptotic BCL2A1A protein were significantly lower at least in the cochlear nucleus, possibly due to increased intrinsic apoptosis after acoustic trauma. Consistently, the levels of pro-apoptotic APAF1protein were slightly increased in the CN. Although missing the significance level of 5% (p = 0.074), which is possibly related to the noise trauma paradigm only covering the mouse low-frequency hearing range, an increased level of intrinsic apoptosis would be able to explain the relatively strong reduction in Apaf1 as well as the elevated Bcl2a1a mRNA expression, indicating a compensating cellular reaction to counteract cellular injury. An acute upregulation of apoptosis within the auditory brainstem is in line with earlier studies, demonstrating activation of pro-apoptotic JNK cascades, increased levels of single-stranded DNA breaks characteristic for apoptosis and an acute reduction in cell number immediately after noise exposure, with the strongest impact in the lower structures and diminishing effects toward the auditory cortex (Gröschel et al., 2010; Meltser et al., 2010; Coordes et al., 2012; Fröhlich et al., 2017).
Downregulation of pro-apoptotic and upregulation of anti-apoptotic genes was also shown in the cochlear nucleus during postnatal development in mice, presumably to prevent mature neurons from stressful insult, thereby pointing at a common underlying physiological response pattern determined during the development of central nervous system, but possibly occurring in pathological situations (Harris et al., 2005).
On the other hand, APAF1 protein is supposed to function as a pro-survival molecule, which absence impairs the cell performance and increases responsiveness to stressful conditions. Thus, a compensatory reduction of Apaf1 mRNA expression could probably be accompanied by a negative impact on the structural and functional arrangement of mitochondrial homeostasis and might further contribute to the long-term physiological deficits and pathologies after noise exposure (Ferraro et al., 2011; Sancho et al., 2014; De Zio et al., 2015).
Lastly, the dynamics of transcription/translation do not always work in the canonical way. Examples were described, where despite increase of mRNA levels, the protein levels decreased (Liu et al., 2016). In addition, the primary brain tissue of mice contains 35% of non-neuronal cells and the techniques based on tissue lysis will deliver an average signal per weight unit of tissue, but is not cell-specific (Herculano-Houzel, 2014). Furthermore, transcription and translation process in neurons occur in a different manner than in other cell types and the changes in protein level is often not combined in time and space with changes occurring in gene transcription. In fact, many mRNA species are pre-synthesized in the nucleus and instead of being immediately translated, they are transported near the synapses and there, locally translated as needed (Glock et al., 2017).
Technical Aspects
In this work, we performed two sets of experiments and obtained some overlapping and some non-overlapping results, thus, raising a question about reproducibility of our expression array results. However, we expected this kind of variability. The first reason accounting for variability of results is the technical error, multiplied by a number of manipulations (e.g., RNA isolation, the process of pooling of RNA into one sample or the efficiency of cDNA in vitro transcription). The last multiplication of possible differences occurs in logarithmic manner during the PCR. The second reason that may account for the differences in results obtained on two separate occasions is using outbred strain of mice and additional, epigenetic differences between individual animals, which could have affected gene expression pattern. The third and last reason is that the biological reaction of animals to noise may vary depending on many factors, and some of these factors may still be unknown.
It still remains unclear if and how far other cell death mechanisms contribute to the observed degeneration. However, previous studies together with the present data lead to the assumption about the intrinsic pathway playing a key role during the immediate response to traumatic insult of the auditory system. In turn, the extrinsic apoptosis and necrosis may be of greater importance at a later time following noise exposure. Comparable observations concerning the timing of apoptotic pathways have been made e.g., after induction of ischemic stroke (Broughton et al., 2009).
Future experiments should clarify the presence and development of different cell death mechanisms following noise trauma. Moreover, induction of apoptotic or necrotic cell death and neurodegeneration could lead to inflammatory responses that have indeed been observed in the cochlear and auditory brain tissues during oxidative stress or after hearing loss (Kirkegaard et al., 2006; Khan et al., 2010; Broughton et al., 2012; Magnus et al., 2012; Tan et al., 2016; Fuentes-Santamaria et al., 2017). Consequently, cellular damage and inflammation might be accompanied by (or even elicit) the neuroplasticity. Subsequently, neuroplasticity may increase excitatory neurotransmission along the auditory pathway, and possibly contribute to psychoacoustic pathologies such as tinnitus (Oh et al., 2007; Mazurek et al., 2012a,b; Manohar et al., 2016).
In conclusion, our experiments gave insight into the underlying mechanisms of early neurodegeneration of the auditory brain induced by acoustic overexposure. Moreover, for the first time we provided direct evidence about key intrinsic apoptosis genes and their products being expressed or present immediately after noise trauma, respectively. Inhibition of the intrinsic apoptotic pathway could be a promising therapeutic approach for the treatment of acute, noise-induced hearing loss. Such treatment could prevent irreversible neuronal damage and might be of particular importance to block the development of profound deficits in complex central auditory processing.
Author Contributions
MG, DB, AE, BM, and AS: designed the study; MG and AS: performed experiments and analyzed the data; DB: assisted with data analysis; MG and AS: wrote the manuscript; MG, DB, AE, BM, and AS: critically reviewed and approved the final manuscript.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge support from the German Research Foundation (DFG) and the Open Access Publication Fund of Charité–Universitätsmedizin Berlin. We are grateful to Ms. Olga Hegend, Julia Fuchs and Christiane Kölske from the ORL Research Laboratories for their excellent technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnins.2018.00312/full#supplementary-material
References
Aarnisalo, A. A., Pirvola, U., Liang, X. Q., Miller, J., and Ylikoski, J. (2000). Apoptosis in auditory brainstem neurons after a severe noise trauma of the organ of Corti: intracochlear GDNF treatment reduces the number of apoptotic cells. ORL J. Otorhinolaryngol. Relat. Spec. 62, 330–334. doi: 10.1159/000027764
Alam, S. A., Oshima, T., Suzuki, M., Kawase, T., Takasaka, T., and Ikeda, K. (2001). The expression of apoptosis-related proteins in the aged cochlea of Mongolian gerbils. Laryngoscope 111, 528–534. doi: 10.1097/00005537-200103000-00026
Ashkenazi, A., and Dixit, V. M. (1998). Death receptors: signaling and modulation. Science 281, 1305–1308.
Balch, W. E., Morimoto, R. I., Dillin, A., and Kelly, J. W. (2008). Adapting proteostasis for disease intervention. Science 319, 916–919. doi: 10.1126/science.1141448
Basta, D., and Ernst, A. (2005). Erratum to “Noise-induced changes of neuronal spontaneous activity in mice inferior colliculus brain slices.” Neurosci. Lett. 374, 74–79. doi: 10.1016/j.neulet.2004.11.002
Basta, D., Tzschentke, B., and Ernst, A. (2005). Noise-induced cell death in the mouse medial geniculate body and primary auditory cortex. Neurosci. Lett. 381, 199–204. doi: 10.1016/j.neulet.2005.02.034
Benson, C. G., Gross, J. S., Suneja, S. K., and Potashner, S. J. (1997). Synaptophysin immunoreactivity in the cochlear nucleus after unilateral cochlear or ossicular removal. Synapse 25, 243–257. doi: 10.1002/(SICI)1098-2396(199703)25:3<243::AID-SYN3>3.0.CO;2-B
Bohne, B. A., Harding, G. W., and Lee, S. C. (2007). Death pathways in noise-damaged outer hair cells. Hear. Res. 223, 61–70. doi: 10.1016/j.heares.2006.10.004
Bohne, B. A., Kimlinger, M., and Harding, G. W. (2017). Time course of organ of Corti degeneration after noise exposure. Hear. Res. 344, 158–169. doi: 10.1016/j.heares.2016.11.009
Broughton, B. R., Lim, R., Arumugam, T. V., Drummond, G. R., Wallace, E. M., and Sobey, C. G. (2012). Post-stroke inflammation and the potential efficacy of novel stem cell therapies: focus on amnion epithelial cells. Front. Cell. Neurosci. 6:66. doi: 10.3389/fncel.2012.00066
Broughton, B. R., Reutens, D. C., and Sobey, C. G. (2009). Apoptotic mechanisms after cerebral ischemia. Stroke 40, e331–339. doi: 10.1161/STROKEAHA.108.531632
Coling, D. E., Ding, D., Young, R., Lis, M., Stofko, E., Blumenthal, K. M., et al. (2007). Proteomic analysis of cisplatin-induced cochlear damage: methods and early changes in protein expression. Hear. Res. 226, 140–156. doi: 10.1016/j.heares.2006.12.017
Coordes, A., Gröschel, M., Ernst, A., and Basta, D. (2012). Apoptotic cascades in the central auditory pathway after noise exposure. J. Neurotrauma 29, 1249–1254. doi: 10.1089/neu.2011.1769
Cory, S., and Adams, J. M. (2002). The Bcl2 family: regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2, 647–656. doi: 10.1038/nrc883
Devarajan, P., Savoca, M., Castaneda, M. P., Park, M. S., Esteban-Cruciani, N., Kalinec, G., et al. (2002). Cisplatin-induced apoptosis in auditory cells: role of death receptor and mitochondrial pathways. Hear. Res. 174, 45–54. doi: 10.1016/S0378-5955(02)00634-2
De Zio, D., Molinari, F., Rizza, S., Gatta, L., Ciotti, M. T., Salvatore, A. M., et al. (2015). Apaf1-deficient cortical neurons exhibit defects in axonal outgrowth. Cell. Mol. Life Sci. 72, 4173–4191. doi: 10.1007/s00018-015-1927-x
Ding, D. L., Ping, W., Haiyan, J., Coling, D., and Salvi, R. (2009). Gene expression in cisplatin ototoxicity and protection with p53 inhibitor. J. Otol. 4, 61–70. doi: 10.1016/S1672-2930(09)50016-5
Ding, Z. J., Chen, X., Tang, X. X., Wang, X., Song, Y. L., Chen, X. D., et al. (2015). Apoptosis-inducing factor and calpain upregulation in glutamate-induced injury of rat spiral ganglion neurons. Mol. Med. Rep. 12, 1685–1692. doi: 10.3892/mmr.2015.3626
Dinh, C. T., Goncalves, S., Bas, E., Van De Water, T. R., and Zine, A. (2015). Molecular regulation of auditory hair cell death and approaches to protect sensory receptor cells and/or stimulate repair following acoustic trauma. Front. Cell. Neurosci. 9:96. doi: 10.3389/fncel.2015.00096
Douglas, P. M., and Dillin, A. (2010). Protein homeostasis and aging in neurodegeneration. J. Cell Biol. 190, 719–729. doi: 10.1083/jcb.201005144
Elmore, S. (2007). Apoptosis: a review of programmed cell death. Toxicol. Pathol. 35, 495–516. doi: 10.1080/01926230701320337
Eskiizmir, G., Yücetürk, A. V., Inan, S., and Gürgen, S. G. (2011). Acute spiral ganglion cell degeneration following acoustic overstimulation: an experimental study. ORL J. Otorhinolaryngol. Relat. Spec. 73, 24–30. doi: 10.1159/000321116
Falah, M., Najafi, M., Houshmand, M., and Farhadi, M. (2016). Expression levels of the BAK1 and BCL2 genes highlight the role of apoptosis in age-related hearing impairment. Clin. Interv. Aging 11, 1003–1008. doi: 10.2147/CIA.S109110
Ferraro, E., Pesaresi, M. G., De Zio, D., Cencioni, M. T., Gortat, A., Cozzolino, M., et al. (2011). Apaf1 plays a pro-survival role by regulating centrosome morphology and function. J. Cell Sci. 124(Pt 20), 3450–3463. doi: 10.1242/jcs.086298
Friedel, C. C., Dölken, L., Ruzsics, Z., Koszinowski, U. H., and Zimmer, R. (2009). Conserved principles of mammalian transcriptional regulation revealed by RNA half-life. Nucleic Acids Res. 37:e115. doi: 10.1093/nar/gkp542
Fröhlich, F., Basta, D., Strübing, I., Ernst, A., and Gröschel, M. (2017). Time course of cell death due to acoustic overstimulation in the mouse medial geniculate body and primary auditory cortex. Noise Health 19, 133–139. doi: 10.4103/nah.NAH_10_17
Fuentes-Santamaria, V., Alvarado, J. C., Melgar-Rojas, P., Gabaldon-Ull, M. C., Miller, J. M., and Juiz, J. M. (2017). The role of glia in the peripheral and central auditory system following noise overexposure: contribution of TNF-α and IL-1β to the pathogenesis of hearing loss. Front. Neuroanat. 11:9. doi: 10.3389/fnana.2017.00009
Furness, D. N. (2015). Molecular basis of hair cell loss. Cell Tissue Res. 361, 387–399. doi: 10.1007/s00441-015-2113-z
García-Berrocal, J. R., Nevado, J., Ramírez-Camacho, R., Sanz, R., González-García, J. A., Sánchez-Rodríguez, C., et al. (2007). The anticancer drug cisplatin induces an intrinsic apoptotic pathway inside the inner ear. Br. J. Pharmacol. 152, 1012–1020. doi: 10.1038/sj.bjp.0707405
Gil-Loyzaga, P., Carricondo, F., Bartolomé, M. V., Iglesias, M. C., Rodríguez, F., and Poch-Broto, J. (2010). Cellular and molecular bases of neuroplasticity: brainstem effects after cochlear damage. Acta Otolaryngol. 130, 318–325. doi: 10.3109/00016480903127468
Glock, C., Heumuller, M., and Schuman, E. M. (2017). mRNA transport & local translation in neurons. Curr. Opin. Neurobiol. 45, 169–177. doi: 10.1016/j.conb.2017.05.005
Gröschel, M., Götze, R., Ernst, A., and Basta, D. (2010). Differential impact of temporary and permanent noise-induced hearing loss on neuronal cell density in the mouse central auditory pathway. J. Neurotrauma 27, 1499–1507. doi: 10.1089/neu.2009.1246
Gröschel, M., Müller, S., Götze, R., Ernst, A., and Basta, D. (2011). The possible impact of noise-induced Ca2+-dependent activity in the central auditory pathway: a manganese-enhanced MRI study. Neuroimage 57, 190–197. doi: 10.1016/j.neuroimage.2011.04.022
Gröschel, M., Ryll, J., Götze, R., Ernst, A., and Basta, D. (2014). Acute and long-term effects of noise exposure on the neuronal spontaneous activity in cochlear nucleus and inferior colliculus brain slices. Biomed Res. Int. 2014:909260. doi: 10.1155/2014/909260
Gross, J., Machulik, A., Amarjargal, N., Moller, R., Ungethüm, U., Kuban, R. J., et al. (2007). Expression of apoptosis-related genes in the organ of Corti, modiolus and stria vascularis of newborn rats. Brain Res. 1162, 56–68. doi: 10.1016/j.brainres.2007.05.061
Gross, J., Olze, H., and Mazurek, B. (2014). Differential expression of transcription factors and inflammation-, ROS-, and cell death-related genes in organotypic cultures in the modiolus, the organ of Corti and the stria vascularis of newborn rats. Cell. Mol. Neurobiol. 34, 523–538. doi: 10.1007/s10571-014-0036-y
Hamernik, R. P., Turrentine, G., Roberto, M., Salvi, R., and Henderson, D. (1984). Anatomical correlates of impulse noise-induced mechanical damage in the cochlea. Hear. Res. 13, 229–247. doi: 10.1016/0378-5955(84)90077-7
Han, W., Shi, X., and Nuttall, A. L. (2006). AIF and endoG translocation in noise exposure induced hair cell death. Hear. Res. 211, 85–95. doi: 10.1016/j.heares.2005.10.004
Han, Y., Hong, L., Chen, Y., Zhong, C., Wang, Y., Zhao, D., et al. (2011). Up-regulation of Nob1 in the rat auditory system with noise-induced hearing loss. Neurosci. Lett. 491, 79–82. doi: 10.1016/j.neulet.2011.01.010
Han, Y., Hong, L., Zhong, C., Chen, Y., Wang, Y., Mao, X., et al. (2012). Identification of new altered genes in rat cochleae with noise-induced hearing loss. Gene 499, 318–322. doi: 10.1016/j.gene.2012.02.042
Hargrove, J. L., Hulsey, M. G., and Beale, E. G. (1991). The kinetics of mammalian gene expression. Bioessays 13, 667–674. doi: 10.1002/bies.950131209
Harris, J. A., Hardie, N. A., Bermingham-McDonogh, O., and Rubel, E. W. (2005). Gene expression differences over a critical period of afferent-dependent neuron survival in the mouse auditory brainstem. J. Comp. Neurol. 493, 460–474. doi: 10.1002/cne.20776
Heffner, H. E., and Heffner, R. S. (2007). Hearing ranges of laboratory animals. J. Am. Assoc. Lab. Anim. Sci. 46, 20–22.
Helfert, R. H., Glatz, F. R. III., Wilson, T. S., Ramkumar, V., and Hughes, L. F. (2002). Hsp70 in the inferior colliculus of Fischer-344 rats: effects of age and acoustic stress. Hear. Res. 170, 155–165. doi: 10.1016/S0378-5955(02)00487-2
Henderson, D., Bielefeld, E. C., Harris, K. C., and Hu, B. H. (2006). The role of oxidative stress in noise-induced hearing loss. Ear Hear. 27, 1–19. doi: 10.1097/01.aud.0000191942.36672.f3
Herculano-Houzel, S. (2014). The glia/neuron ratio: how it varies uniformly across brain structures and species and what that means for brain physiology and evolution. Glia 62, 1377–1391. doi: 10.1002/glia.22683
Hill, M. M., Adrain, C., Duriez, P. J., Creagh, E. M., and Martin, S. J. (2004). Analysis of the composition, assembly kinetics and activity of native Apaf-1 apoptosomes. EMBO J. 23, 2134–2145. doi: 10.1038/sj.emboj.7600210
Hirose, K., and Liberman, M. C. (2003). Lateral wall histopathology and endocochlear potential in the noise-damaged mouse cochlea. J. Assoc. Res. Otolaryngol. 4, 339–352. doi: 10.1007/s10162-002-3036-4
Holt, A. G., Asako, M., Lomax, C. A., MacDonald, J. W., Tong, L., Lomax, M. I., et al. (2005). Deafness-related plasticity in the inferior colliculus: gene expression profiling following removal of peripheral activity. J. Neurochem. 93, 1069–1086. doi: 10.1111/j.1471-4159.2005.03090.x
Hu, B. H., Cai, Q., Manohar, S., Jiang, H., Ding, D., Coling, D. E., et al. (2009). Differential expression of apoptosis-related genes in the cochlea of noise-exposed rats. Neuroscience 161, 915–925. doi: 10.1016/j.neuroscience.2009.03.072
Hu, B. H., Guo, W., Wang, P. Y., Henderson, D., and Jiang, S. C. (2000). Intense noise-induced apoptosis in hair cells of guinea pig cochleae. Acta Otolaryngol. 120, 19–24. doi: 10.1080/000164800760370774
Hu, B. H., Henderson, D., and Nicotera, T. M. (2002). Involvement of apoptosis in progression of cochlear lesion following exposure to intense noise. Hear. Res. 166, 62–71. doi: 10.1016/S0378-5955(02)00286-1
Hu, B. H., Henderson, D., and Nicotera, T. M. (2006). Extremely rapid induction of outer hair cell apoptosis in the chinchilla cochlea following exposure to impulse noise. Hear. Res. 211, 16–25. doi: 10.1016/j.heares.2005.08.006
Jean-Baptiste, M., and Morest, D. K. (1975). Transneuronal changes of synaptic endings and nuclear chromatin in the trapezoid body following cochlear ablations in cats. J.Comp Neurol. 162, 111–134. doi: 10.1002/cne.901620107
Jiang, X., and Wang, X. (2000). Cytochrome c promotes caspase-9 activation by inducing nucleotide binding to Apaf-1. J. Biol. Chem. 275, 31199–31203. doi: 10.1074/jbc.C000405200
Kaltenbach, J. A., Godfrey, D. A., Neumann, J. B., McCaslin, D. L., Afman, C. E., and Zhang, J. (1998). Changes in spontaneous neural activity in the dorsal cochlear nucleus following exposure to intense sound: relation to threshold shift. Hear. Res. 124, 78–84. doi: 10.1016/S0378-5955(98)00119-1
Khan, M., Szczepek, A. J., Haupt, H., Olze, H., and Mazurek, B. (2010). Expression of the proinflammatory cytokines in cochlear explant cultures: influence of normoxia and hypoxia. Neurosci. Lett. 479, 249–252. doi: 10.1016/j.neulet.2010.05.072
Kim, J. J., Gross, J., Morest, D. K., and Potashner, S. J. (2004). Quantitative study of degeneration and new growth of axons and synaptic endings in the chinchilla cochlear nucleus after acoustic overstimulation. J. Neurosci. Res. 77, 829–842. doi: 10.1002/jnr.20211
Kim, J., Morest, D. K., and Bohne, B. A. (1997). Degeneration of axons in the brainstem of the chinchilla after auditory overstimulation. Hear. Res. 103, 169–191. doi: 10.1016/S0378-5955(96)00173-6
Kirkegaard, M., Murai, N., Risling, M., Suneson, A., Järlebark, L., and Ulfendahl, M. (2006). Differential gene expression in the rat cochlea after exposure to impulse noise. Neuroscience 142, 425–435. doi: 10.1016/j.neuroscience.2006.06.037
Komiya, H., and Eggermont, J. J. (2000). Spontaneous firing activity of cortical neurons in adult cats with reorganized tonotopic map following pure-tone trauma. Acta Otolaryngol. 120, 750–756. doi: 10.1080/000164800750000298
Kraus, K. S., Ding, D., Zhou, Y., and Salvi, R. J. (2009). Central auditory plasticity after carboplatin-induced unilateral inner ear damage in the chinchilla: up-regulation of GAP-43 in the ventral cochlear nucleus. Hear. Res. 255, 33–43. doi: 10.1016/j.heares.2009.05.001
Le, T. N., Straatman, L. V., Lea, J., and Westerberg, B. (2017). Current insights in noise-induced hearing loss: a literature review of the underlying mechanism, pathophysiology, asymmetry, and management options. J. Otolaryngol. Head Neck Surg. 46:41. doi: 10.1186/s40463-017-0219-x
Liberman, M. C. (2017). Noise-induced and age-related hearing loss: new perspectives and potential therapies. F1000Res 6:927. doi: 10.12688/f1000research.11310.1
Liu, Y., Beyer, A., and Aebersold, R. (2016). On the Dependency of Cellular Protein Levels on mRNA Abundance. Cell 165, 535–550. doi: 10.1016/j.cell.2016.03.014
Magnus, T., Wiendl, H., and Kleinschnitz, C. (2012). Immune mechanisms of stroke. Curr. Opin. Neurol. 25, 334–340. doi: 10.1097/WCO.0b013e328352ede6
Manohar, S., Dahar, K., Adler, H. J., Dalian, D., and Salvi, R. (2016). Noise-induced hearing loss: neuropathic pain via Ntrk1 signaling. Mol. Cell. Neurosci. 75, 101–112. doi: 10.1016/j.mcn.2016.07.005
Manohar, S., Jamesdaniel, S., and Salvi, R. (2014). Cisplatin inhibits hippocampal cell proliferation and alters the expression of apoptotic genes. Neurotox. Res. 25, 369–380. doi: 10.1007/s12640-013-9443-y
Mattson, M. P. (2007). Calcium and neurodegeneration. Aging Cell 6, 337–350. doi: 10.1111/j.1474-9726.2007.00275.x
Mazurek, B., Amarjargal, N., Haupt, H., Fuchs, J., Olze, H., Machulik, A., et al. (2011). Expression of genes implicated in oxidative stress in the cochlea of newborn rats. Hear. Res. 277, 54–60. doi: 10.1016/j.heares.2011.03.011
Mazurek, B., Haupt, H., Klapp, B. F., Szczepek, A. J., and Olze, H. (2012a). Exposure of Wistar rats to 24-h psycho-social stress alters gene expression in the inferior colliculus. Neurosci. Lett. 527, 40–45. doi: 10.1016/j.neulet.2012.08.019
Mazurek, B., Haupt, H., Olze, H., and Szczepek, A. J. (2012b). Stress and tinnitus-from bedside to bench and back. Front. Syst. Neurosci. 6:47. doi: 10.3389/fnsys.2012.00047
Meltser, I., Cederroth, C. R., Basinou, V., Savelyev, S., Lundkvist, G. S., and Canlon, B. (2014). TrkB-mediated protection against circadian sensitivity to noise trauma in the murine cochlea. Curr. Biol. 24, 658–663. doi: 10.1016/j.cub.2014.01.047
Meltser, I., Tahera, Y., and Canlon, B. (2010). Differential activation of mitogen-activated protein kinases and brain-derived neurotrophic factor after temporary or permanent damage to a sensory system. Neuroscience 165, 1439–1446. doi: 10.1016/j.neuroscience.2009.11.025
Michler, S. A., and Illing, R. B. (2002). Acoustic trauma induces reemergence of the growth- and plasticity-associated protein GAP-43 in the rat auditory brainstem. J. Comp. Neurol. 451, 250–266. doi: 10.1002/cne.10348
Nevado, J., Sanz, R., Casqueiro, J. C., Ayala, A., García-Berrocal, J. R., and Ramírez-Camacho, R. (2006). Ageing evokes an intrinsic pro-apoptotic signalling pathway in rat cochlea. Acta Otolaryngol. 126, 1134–1139. doi: 10.1080/00016480600672592
Nicotera, T. M., Hu, B. H., and Henderson, D. (2003). The caspase pathway in noise-induced apoptosis of the chinchilla cochlea. J. Assoc. Res. Otolaryngol. 4, 466–477. doi: 10.1007/s10162-002-3038-2
Oh, S. H., Kim, C. S., and Song, J. J. (2007). Gene expression and plasticity in the rat auditory cortex after bilateral cochlear ablation. Acta Otolaryngol. 127, 341–350. doi: 10.1080/00016480701275246
Ohlemiller, K. K., Wright, J. S., and Dugan, L. L. (1999). Early elevation of cochlear reactive oxygen species following noise exposure. Audiol. Neurootol. 4, 229–236. doi: 10.1159/000013846
Orzáez, M., Sancho, M., Marchán, S., Mondragón, L., Montava, R., Valero, J. G., et al. (2014). Apaf-1 inhibitors protect from unwanted cell death in in vivo models of kidney ischemia and chemotherapy induced ototoxicity. PLoS ONE 9:e110979. doi: 10.1371/journal.pone.0110979
Paxinos, G., and Franklin, K. B. J. (2001). The Mouse Brain in Stereotaxic Coordinates. Oxford: Elsevier Science & Technology.
Pirvola, U., Xing-Qun, L., Virkkala, J., Saarma, M., Murakata, C., Camoratto, A. M., et al. (2000). Rescue of hearing, auditory hair cells, and neurons by CEP-1347/KT7515, an inhibitor of c-Jun N-terminal kinase activation. J. Neurosci. 20, 43–50. doi: 10.1523/JNEUROSCI.20-01-00043.2000
Portt, L., Norman, G., Clapp, C., Greenwood, M., and Greenwood, M. T. (2011). Anti-apoptosis and cell survival: a review. Biochim. Biophys. Acta 1813, 238–259. doi: 10.1016/j.bbamcr.2010.10.010
Pujol, R., and Puel, J. L. (1999). Excitotoxicity, synaptic repair, and functional recovery in the mammalian cochlea: a review of recent findings. Ann. N.Y Acad. Sci. 884, 249–254. doi: 10.1111/j.1749-6632.1999.tb08646.x
Rajan, R., and Irvine, D. R. (1998). Absence of plasticity of the frequency map in dorsal cochlear nucleus of adult cats after unilateral partial cochlear lesions. J. Comp. Neurol. 399, 35–46. doi: 10.1002/(SICI)1096-9861(19980914)399:1<35::AID-CNE3>3.0.CO;2-B
Reynolds, R. P., Kinard, W. L., Degraff, J. J., Leverage, N., and Norton, J. N. (2010). Noise in a laboratory animal facility from the human and mouse perspectives. J. Am. Assoc. Lab. Anim. Sci. 49, 592–597.
Saelens, X., Festjens, N., Vande Walle, L., van Gurp, M., van Loo, G., and Vandenabeele, P. (2004). Toxic proteins released from mitochondria in cell death. Oncogene 23, 2861–2874. doi: 10.1038/sj.onc.1207523
Sakahira, H., Enari, M., and Nagata, S. (1998). Cleavage of CAD inhibitor in CAD activation and DNA degradation during apoptosis. Nature 391, 96–99.
Salinska, E., Danysz, W., and Łazarewicz, J. W. (2005). The role of excitotoxicity in neurodegeneration. Folia Neuropathol. 43, 322–339.
Samson, J., Sheeladevi, R., Ravindran, R., and Senthilvelan, M. (2007). Stress response in rat brain after different durations of noise exposure. Neurosci. Res. 57, 143–147. doi: 10.1016/j.neures.2006.09.019
Sancho, M., Gortat, A., Herrera, A. E., Andreu-Fernández, V., Ferraro, E., Cecconi, F., et al. (2014). Altered mitochondria morphology and cell metabolism in Apaf1-deficient cells. PLoS ONE 9:e84666. doi: 10.1371/journal.pone.0084666
Saunders, J. C., Dear, S. P., and Schneider, M. E. (1985). The anatomical consequences of acoustic injury: a review and tutorial. J. Acoust. Soc. Am. 78, 833–860. doi: 10.1121/1.392915
Sekiya, T., Canlon, B., Viberg, A., Matsumoto, M., Kojima, K., Ono, K., et al. (2009). Selective vulnerability of adult cochlear nucleus neurons to de-afferentation by mechanical compression. Exp. Neurol. 218, 117–123. doi: 10.1016/j.expneurol.2009.04.014
Sekiya, T., Viberg, A., Kojima, K., Sakamoto, T., Nakagawa, T., Ito, J., et al. (2012). Trauma-specific insults to the cochlear nucleus in the rat. J. Neurosci. Res. 90, 1924–1931. doi: 10.1002/jnr.23093
Shakeri, R., Kheirollahi, A., and Davoodi, J. (2017). Apaf-1: regulation and function in cell death. Biochimie 135, 111–125. doi: 10.1016/j.biochi.2017.02.001
Someya, S., Xu, J., Kondo, K., Ding, D., Salvi, R. J., Yamasoba, T., et al. (2009). Age-related hearing loss in C57BL/6J mice is mediated by Bak-dependent mitochondrial apoptosis. Proc. Natl. Acad. Sci. U.S.A. 106, 19432–19437. doi: 10.1073/pnas.0908786106
Sun, W., Zhang, L., Lu, J., Yang, G., Laundrie, E., and Salvi, R. (2008). Noise exposure-induced enhancement of auditory cortex response and changes in gene expression. Neuroscience 156, 374–380. doi: 10.1016/j.neuroscience.2008.07.040
Tadros, S. F., D'Souza, M., Zhu, X., and Frisina, R. D. (2008). Apoptosis-related genes change their expression with age and hearing loss in the mouse cochlea. Apoptosis 13, 1303–1321. doi: 10.1007/s10495-008-0266-x
Tan, W. J., Thorne, P. R., and Vlajkovic, S. M. (2016). Characterisation of cochlear inflammation in mice following acute and chronic noise exposure. Histochem. Cell Biol. 146, 219–230. doi: 10.1007/s00418-016-1436-5
Van De Water, T. R., Lallemend, F., Eshraghi, A. A., Ahsan, S., He, J., Guzman, J., et al. (2004). Caspases, the enemy within, and their role in oxidative stress-induced apoptosis of inner ear sensory cells. Otol. Neurotol. 25, 627–632. doi: 10.1097/00129492-200407000-00035
Vogler, M. (2012). BCL2A1: the underdog in the BCL2 family. Cell Death Differ. 19, 67–74. doi: 10.1038/cdd.2011.158
Wada, T., and Penninger, J. M. (2004). Mitogen-activated protein kinases in apoptosis regulation. Oncogene 23, 2838–2849. doi: 10.1038/sj.onc.1207556
Wallhäusser-Franke, E., Mahlke, C., Oliva, R., Braun, S., Wenz, G., and Langner, G. (2003). Expression of c-fos in auditory and non-auditory brain regions of the gerbil after manipulations that induce tinnitus. Exp. Brain Res. 153, 649–654. doi: 10.1007/s00221-003-1614-2
Wang, P., Zhang, P., Huang, J., Li, M., and Chen, X. (2013). Trichostatin A protects against cisplatin-induced ototoxicity by regulating expression of genes related to apoptosis and synaptic function. Neurotoxicology 37, 51–62. doi: 10.1016/j.neuro.2013.03.007
Wang, Y., Hirose, K., and Liberman, M. C. (2002). Dynamics of noise-induced cellular injury and repair in the mouse cochlea. J. Assoc. Res. Otolaryngol. 3, 248–268. doi: 10.1007/s101620020028
Weisz, C., Glowatzki, E., and Fuchs, P. (2009). The postsynaptic function of type II cochlear afferents. Nature 461, 1126–1129. doi: 10.1038/nature08487
WHO (2015). World Health Organization Department for Management of Nuncommunicable Diseases - 1.1 Billion People at Risk of Hearing Loss. Available Online at: http://www.who.int/mediacentre/news/releases/2015/ear-care/en/ (Accessed Dec 1, 2017).
Yamashita, D., Jiang, H. Y., Schacht, J., and Miller, J. M. (2004a). Delayed production of free radicals following noise exposure. Brain Res. 1019, 201–209. doi: 10.1016/j.brainres.2004.05.104
Yamashita, D., Miller, J. M., Jiang, H. Y., Minami, S. B., and Schacht, J. (2004b). AIF and EndoG in noise-induced hearing loss. Neuroreport 15, 2719–2722.
Yang, W. P., Henderson, D., Hu, B. H., and Nicotera, T. M. (2004). Quantitative analysis of apoptotic and necrotic outer hair cells after exposure to different levels of continuous noise. Hear. Res. 196, 69–76. doi: 10.1016/j.heares.2004.04.015
Yoshida, H., Kong, Y. Y., Yoshida, R., Elia, A. J., Hakem, A., Hakem, R., et al. (1998). Apaf1 is required for mitochondrial pathways of apoptosis and brain development. Cell 94, 739–750. doi: 10.1016/S0092-8674(00)81733-X
Yu, J., Ding, D., Wang, F., Jiang, H., Sun, H., and Salvi, R. (2014). Pattern of hair cell loss and delayed peripheral neuron degeneration in inner ear by a high-dose intratympanic gentamicin. J. Otol. 9, 126–135. doi: 10.1016/j.joto.2014.12.001
Yu, S. P., Canzoniero, L. M., and Choi, D. W. (2001). Ion homeostasis and apoptosis. Curr.Opin.Cell Biol. 13, 405–411. doi: 10.1016/S0955-0674(00)00228-3
Keywords: noise-induced hearing loss, central auditory system, acute noise exposure, APAF1, BCL2A1A
Citation: Gröschel M, Basta D, Ernst A, Mazurek B and Szczepek AJ (2018) Acute Noise Exposure Is Associated With Intrinsic Apoptosis in Murine Central Auditory Pathway. Front. Neurosci. 12:312. doi: 10.3389/fnins.2018.00312
Received: 23 February 2018; Accepted: 23 April 2018;
Published: 09 May 2018.
Edited by:
Victoria M. Bajo Lorenzana, University of Oxford, United KingdomReviewed by:
Dora Reglodi, University of Pécs, HungaryJuan Carlos Alvarado, Universidad de Castilla-La Mancha, Spain
Copyright © 2018 Gröschel, Basta, Ernst, Mazurek and Szczepek. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Moritz Gröschel, bW9yaXR6Lmdyb2VzY2hlbEBiaW9sb2dpZS5odS1iZXJsaW4uZGU=
Agnieszka J. Szczepek, YWduZXMuc3pjemVwZWtAY2hhcml0ZS5kZQ==