 Micaël Carrier1
Micaël Carrier1 Marie-Ève Robert1Fernando González Ibáñez1
Marie-Ève Robert1Fernando González Ibáñez1 Michèle Desjardins2,3
Michèle Desjardins2,3 Marie-Ève Tremblay1,4,5*
Marie-Ève Tremblay1,4,5*- 1Axe Neurosciences, Centre de Recherche du CHU de Québec, Université Laval, Québec City, QC, Canada
- 2Axe Oncologie, Centre de Recherche du CHU de Québec, Université Laval, Québec City, QC, Canada
- 3Department of Physics, Physical Engineering and Optics, Université Laval, Québec City, QC, Canada
- 4Department of Molecular Medicine, Université Laval, Québec City, QC, Canada
- 5Division of Medical Sciences, University of Victoria, Victoria, BC, Canada
The immune system is essential for maintaining homeostasis, as well as promoting growth and healing throughout the brain and body. Considering that immune cells respond rapidly to changes in their microenvironment, they are very difficult to study without affecting their structure and function. The advancement of non-invasive imaging methods greatly contributed to elucidating the physiological roles performed by immune cells in the brain across stages of the lifespan and contexts of health and disease. For instance, techniques like two-photon in vivo microscopy were pivotal for studying microglial functional dynamics in the healthy brain. Through these observations, their interactions with neurons, astrocytes, blood vessels and synapses were uncovered. High-resolution electron microscopy with immunostaining and 3D-reconstruction, as well as super-resolution fluorescence microscopy, provided complementary insights by revealing microglial interventions at synapses (phagocytosis, trogocytosis, synaptic stripping, etc.). In addition, serial block-face scanning electron microscopy has provided the first 3D reconstruction of a microglial cell at nanoscale resolution. This review will discuss the technical toolbox that currently allows to study microglia and other immune cells in the brain, as well as introduce emerging methods that were developed and could be used to increase the spatial and temporal resolution of neuroimmune imaging. A special attention will also be placed on positron emission tomography and the development of selective functional radiotracers for microglia and peripheral macrophages, considering their strong potential for research translation between animals and humans, notably when paired with other imaging modalities such as magnetic resonance imaging.
Introduction
The immune system of the central nervous system (CNS) is essential to maintain its homeostasis [reviewed in Tay et al. (2017)]. Typically, this task is mainly performed by the resident immune cell population, microglia (Ginhoux et al., 2010; Tay et al., 2019). The role of microglia and other immune cells in the brain was uncovered using innovative methods allowing to image them in situ (Savage et al., 2019a) and in vivo (Davalos et al., 2005; Herz et al., 2012). Considering that immune cells like microglia are highly sensitive to changes in their microenvironment, non-invasive techniques that prevent their reaction during tissue preparation and imaging are necessary (Cătălin et al., 2017; Augusto-Oliveira et al., 2019). Immune cell reactivity can be assessed using morphological features (Savage et al., 2019a) or more sensitively by ultrastructure (El Hajj et al., 2019; Savage et al., 2019c). Overall, imaging techniques revealed that microglia play essential roles in the brain: as immune sentinels that clean toxic extracellular debris and repair damage to neighboring cells, but also as glial cells regulating neuronal and astrocytic/oligodendrocytic cell survival, differentiation, maturation, migration, as well as network formation and refinement with relation to neuronal activity (Ter Veer et al., 2017; Bordeleau et al., 2019; Tay et al., 2019). Once reactive, microglia play important roles in neuroinflammation –beneficial or detrimental depending on the context of disease or injury (Plemel et al., 2020). These roles of reactive microglia are exerted in cooperation with other glial and other innate immune cells (e.g., monocytes, neutrophils) as well as endothelial cells forming the blood-brain-barrier (BBB) (Sofroniew, 2015; Van Dyken and Lacoste, 2018; Bellver-Landete et al., 2019). Although monocytes were not shown to be present in the healthy adult mouse brain (Ginhoux et al., 2013; Prinz and Priller, 2014, 2017) they can be recruited rapidly in a wide panoply of contexts (D’Mello et al., 2009; Wohleb et al., 2015; Varvel et al., 2016; Niraula et al., 2018; Schneider et al., 2019). Once in the brain, monocytes tend to adopt a similar morphology as microglia and are often referred to as microglia-like cells even if they have a different origin, from the bone marrow while microglia come from the embryonic yolk sac (Leone et al., 2006; Varvel et al., 2012; Katsumoto et al., 2014). The adaptive immune system is also highly complex, with multiple cell types such as lymphocytes (T cells, B cells, and NK cells) playing a possibly important role in brain development, activity and plasticity notably through their secretion of neurotrophins (Kipnis et al., 2004; Lewitus and Schwartz, 2009; Morimoto and Nakajima, 2019). T cells in particular are often seen in the brain parenchyma in multiple sclerosis, both in human samples and its animal models of experimental autoimmune encephalomyelitis. In these autoimmune diseases, T cells are considered to change their behavior in a destructive way that causes demyelination and inflammation (Lehmann, 1998). Here, we will review the main imaging techniques that are currently available to study brain immunity, focusing on microglia as they are mainly studied, but also on monocytes and T cells, while providing examples of their complementary applications, and discuss the power of their combination into integrative studies.
Imaging the Homeostatic Brain Using Photons
Microscopy techniques (brightfield, fluorescence, confocal, multiphoton, etc.) are commonly used in neuroimmunology to analyze the density, distribution and morphology of different cell types, including microglia, and their interactions one with another (Savage et al., 2019a). The examined cells often require markers to be visualized. Immunostaining is widely used to label the cells of interest across various types of samples, from animal models to human postmortem CNS. For example, double immunostaining against IBA1 and TMEM119 can be used to distinguish microglia (double positive; derived from embryonic yolk sac) from infiltrating peripheral macrophages (expressing IBA1 but not TMEM119; from bone marrow) (González Ibanez et al., 2019). One should note that TMEM119 expression is not stable in early development (until P20 in mouse) and can change in disease context such as multiple sclerosis, highlighting the importance of using a combination of markers including Fcrls, siglec-h, sall1, P2RY12 and more recently HEXB to identify microglia (Butovsky et al., 2014; Buttgereit et al., 2016; Konishi et al., 2017; Zrzavy et al., 2017; Masuda et al., 2020). Fluorescence can also label immune cells directly, without immunostaining, when markers are biologically expressed in transgenic animals (Martell et al., 2017; Daigle et al., 2018). Fate mapping of immune cells origin and trajectory, from the embryonic yolk sac or bone marrow to the CNS, and then function in the CNS can then be performed (Gomez Perdiguero et al., 2015). Reporter mice are especially useful when they are crossed one with another. As an example, the CX3CR1+/GFP;Thy1H+/YFP model (Feng et al., 2000; Jung et al., 2000) labels microglia and neurons including synapses in two different colors. It allows to study dynamic relationships between the two cell types, which is particularly relevant for studying microglia-synapse structural interactions (Tremblay et al., 2010). Furthermore, fluorescent dyes can be injected in vivo, while cells can be stained ex vivo and introduced back into animals, which can be transgenic, to allow for their discrimination (Helmchen and Denk, 2005). Many different mouse models, discussed in detail in a comprehensive review (Theret et al., 2019), were developed to study the function of resident microglia and macrophages within the CNS. Parallel to this, photonic microscopy greatly evolved, providing always improved images, bypassing the spatial resolution limit from 350 nm to around 65 nm with super-resolution microscopy (Klar et al., 2001; Evilsizor et al., 2015), which is especially useful for studying synaptic interactions. We will next explore the main microscopy techniques that are commonly used in neuroimmunology and present emerging ones that could provide new insights. Advantages and limitations are compared in Table 1.
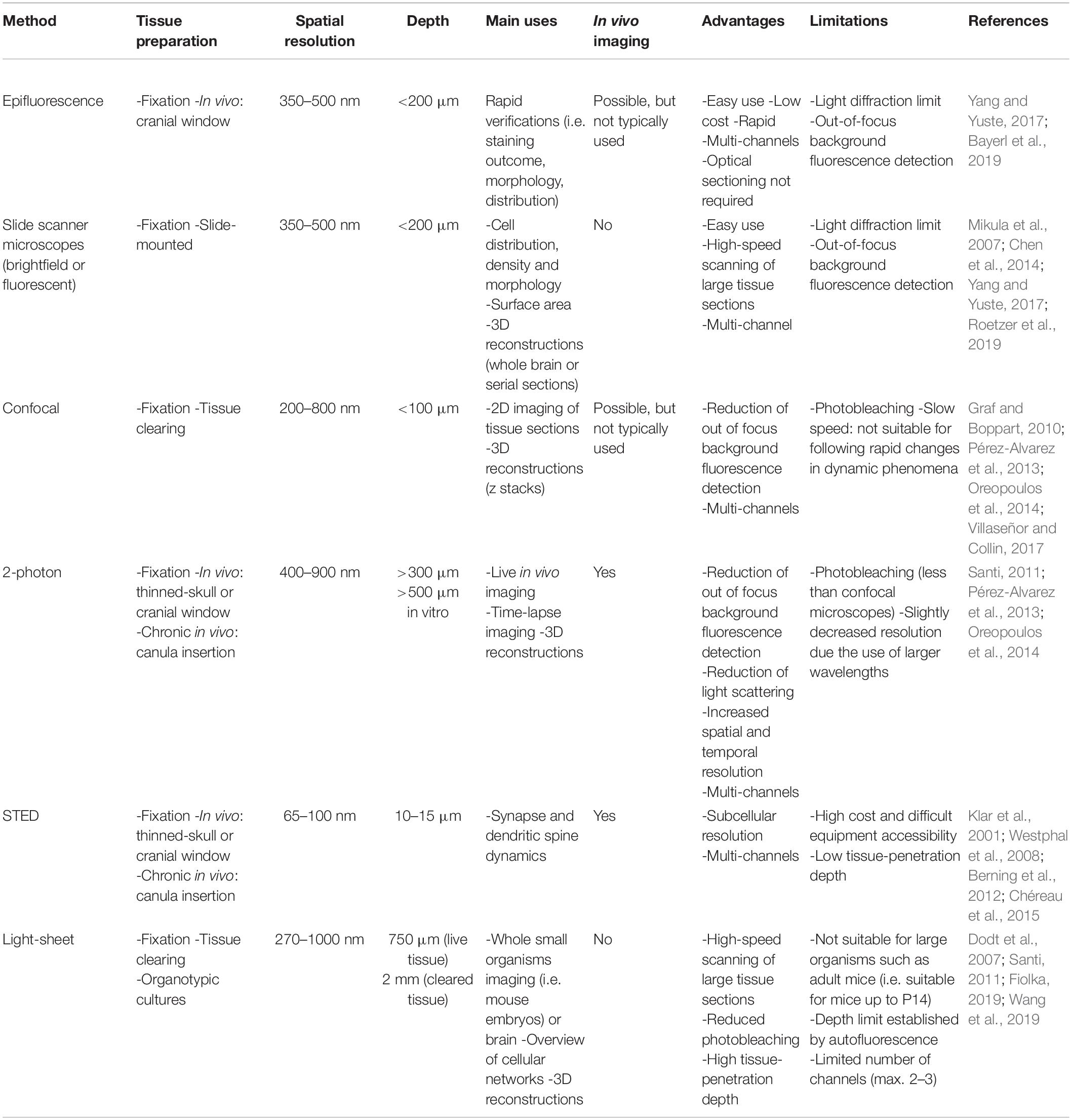
Table 1. Comparison of light microscopy techniques based on their advantages and limitations.
Slide Scanning
When answering questions about immune cell functions without knowing the specific CNS regions of interest involved, one can turn to high throughput imaging techniques such as slide scanning. This very convenient technique uses a mobile stage to automatically image regions of interest, from a series of CNS sections mounted on an entire microscopy glass slide, with a theoretical resolution of 900 nm (x-y). Whole mounts of mouse brain (Roetzer et al., 2019) and spinal cord (Bellver-Landete et al., 2019) were successfully imaged using this technique. It is ideal to perform quantitative analysis of microglial density and distribution, for instance, across large CNS tissue areas, as proposed within an experimental workflow in Figure 1A. One edge of the technique is the large amount of information provided in slide scanning images. Being formed of multiple tiles, these images allow the user to zoom in and out to visualize cellular density, distribution, morphology, intercellular relationships and other features of interest. The mosaic can be used to see the bigger picture of the cellular alterations or phenotypic changes as mentioned in Figure 1B. It can also be used to create new files containing smaller regions of interest with softwares such as QUPATH that could also be used for automatic analysis of density, distribution or morphology and intercellular relationships with machine learning (Bankhead et al., 2017). For instance, automatic analysis of slide scanning images was performed to characterize IBA1-immunopositive microglia/monocytes in traumatic brain injured mice at the site of injuries. The mosaic generated was used to count and classify these cells as “ramified” or “amoeboid” using machine learning, in less than 15 min compared to 5 h for a manual analyst. This faster process suggests strong potential for similar high-throughput analysis using human brain slices (Kyriazis et al., 2019). Slide scanning combined with machine learning is then a prime choice method for whole slices imaging in animal and human postmortem samples (Budde and Annese, 2013), considering that it removes the human bias when counting or tracing the contour of infiltrating monocytes or microglia from hundreds of slices.
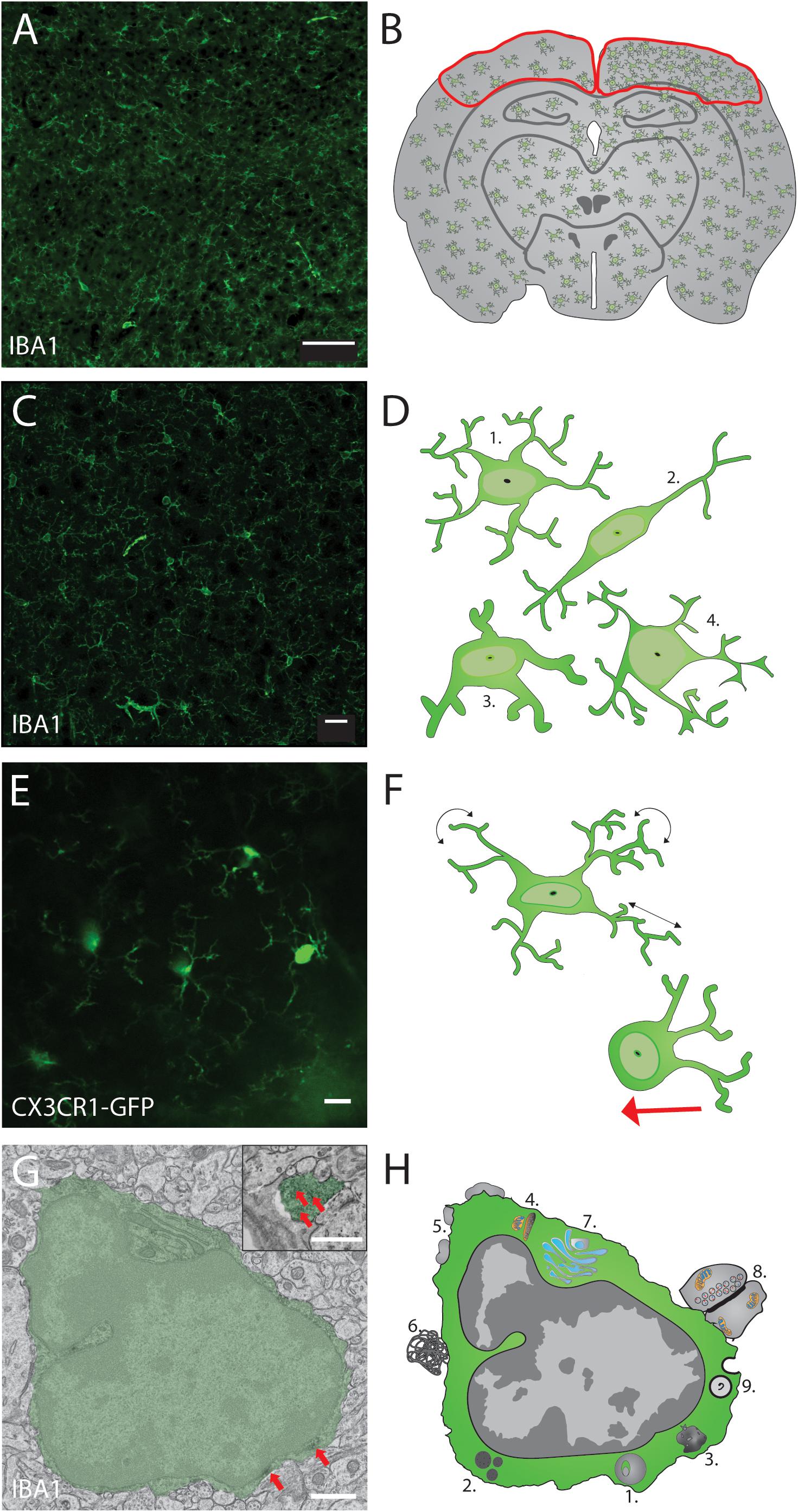
Figure 1. Proposed thorough structural analysis of the immune cells in the brain, exemplified with microglia. Optimal analysis of microglial density, distribution, morphology and ultrastructure using 4 different imaging modalities can allow to evaluate their cellular network, organellar function and health status. (A) Slide scanning can be performed to image microglial density and distribution throughout the brain (scale bar = 100 μm) while discriminating between different regions and layers of interest (B). Microglial morphology can be assessed in (C) using confocal microscopy to measure quantitative changes in their cell body and arborization area (scale bar = 20 μm) to be then classified, as illustrated starting from the top in (D) into ramified, rod shape, hypertrophic and dystrophic phenotypes, an assessment not always feasible by slide scanner imaging due to the lower resolution and lack of 3D information. Microglial dynamics can be investigated using non-invasive two-photon in vivo imaging of CX3CR1+/GFP reporter mice (scale bar = 10 μm), as shown in (E) with a thinned-skull preparation allowing to assess microglial cell body mobility (red arrow in F) and process motility (black arrows) during normal physiological conditions in the intact mouse cerebral cortex. The ultrastructure of microglial cell body and processes can be analyzed using (G) transmission electron microscopy, as illustrated by a pseudocolored microglia (scale bar = 1 μm) that displays peroxidase staining in its cell body and processes against the IBA1 protein (red arrows). Cytologic events that can be quantified in microglia using different electron microscopy techniques are illustrated in (H) and include, notably, 1. Lysosomal inclusions, 2. Lipid bodies, 3. Lipofuscin granules, 4. Elongated or altered mitochondria, 5. Extracellular space pockets, 6. Extracellular digestion, 7. Endoplasmic reticulum/Golgi dilation, 8. Synaptic contacts, and 9. Phagolysosomal inclusions.
Confocal Microscopy
The biggest foe of fluorescent microscopy is light scattering, which limits its spatial resolution to about 300 nm (x-y) and 600 nm (z) at best. To circumscribe this problem, confocal microscopes increase resolution by only detecting the fluorescence emitted by the sample at the focal point. This feat is achieved by using a pinhole to reject light originating from outside the focal plane (Elliott, 2020). In neuroimmunology, confocal microscopy is commonly used to image the morphology of immune cells in three dimensions (3D), by acquiring z-stacks as presented in Figure 1C. Z-stacks are the key functionality of confocal microscopy. Allowing the 3D analysis of immune cells can give deeper insight when characterizing the shape of microglia, monocytes and T cells (Perego et al., 2013; Mittal et al., 2019). In Alzheimer’s disease pathology, for instance, T cell morphology can be 3D rendered in relation to neurons, which allowed the identification of “emperipolesis,” a phenomenon by which the T cell is actually inside the neuron, which might suggest neuronal damage associated with the foreign cell entry (Lodygin et al., 2019). In injuries, the morphology of microglia can also be effectively assessed from 3D reconstruction where the ramification density and arborization area can be analyzed for microglia close to injury site versus located in the periphery (Otxoa-de-Amezaga et al., 2019). Such changes in morphology, associated with microglial surveillance capacity and interactions with other parenchymal elements, including synapses, can be missed in typical epifluorescence images as smaller ramifications are difficult to discern with the reduced resolution. Thus, confocal microscopy is an effective tool to reveal the morphology of microglia and other cell types in 3D, as required to identify phenotypic changes, cellular alterations and functions, as illustrated in Figure 1D. In the case of microglia, their morphology is considered a direct indicator of their activity. For instance, amoeboid microglia tend to migrate in the brain while hypertrophic microglia tend to perform more phagocytosis than surveillant ones (Savage et al., 2019a; Tay et al., 2019). Using confocal microscopy, researchers can analyze various morphological changes like the formation of phagocytic cups at the end of microglia/monocyte processes, associated with the removal of engulfed newly proliferated cells and responsible for the development of juvenile play in adolescent rat (Sierra et al., 2010; VanRyzin et al., 2019). However, confocal microscopy is not often used in in vivo applications because of its limited penetration depth, which depends on the usable excitation wavelength (Graf and Boppart, 2010).
Stimulated Emission Depletion (STED)
Microscopy increases image resolution by overcoming the light diffraction limit. Similar to other commonly used fluorescence microscopy approaches, STED involves the detection of fluorescence by point-scanning. A doughnut-like shape beam is used to deplete surrounding fluorophores in order to only read the fluorescence of the fluorophore of interest (Hell and Wichmann, 1994; Tardif et al., 2019) STED microscopy’s nanoscale resolution (65 nm in x-y and 150 nm in z) resulted in multiple applications, including the structural imaging of microglial and synaptic dynamics in fixed and live thick brain slices (Hein et al., 2008; Chéreau et al., 2015). Recent progresses further allowed to generate the first STED able to image efficiently in vivo, by adding components of a two-photon system. The use of a pulsed laser source provided better penetration, combined with the high spatial resolution of STED microscopy. With this approach, a greater number of small dendritic spines was imaged than with conventional two-photon microscopy, thus allowing to show a higher turnover of dendritic spines than previously envisioned (Pfeiffer et al., 2018). Other high-resolution microscopy techniques have been developed such as structured illumination microscopy (SIM) that uses a spatial modulation pattern to illuminate the sample, in which a software demodulation and filtering allows to overcome the diffraction limit with a factor two (Schermelleh et al., 2008). Stochastic optical reconstruction microscopy (STORM), where only a portion of fluorophores are excited on each cycle of laser emission, gives insight into their precise location at the nanoscale (Rust et al., 2006).
Light-Sheet
While super-resolution techniques revolutionized the use of fluorescence microscopy by increasing its spatial resolution, light-sheet microscopy is a technique that was developed for faster imaging. It allows to perform high speed scanning of large CNS tissue volumes. The system requires a fluorescence detection setup that differs slightly from epifluorescence microscopy, since detection is performed transverse to the illumination by a thin sheet of laser light (Keller and Ahrens, 2015). This non-destructive imaging technique prevents sectioning interpretation artifacts. These artifacts arise when cells are cut in cross-section by vibratome, freezing microtome or cryostat. In addition, it offers the possibility to perform molecular analyses after imaging the fixed tissue (Glaser et al., 2017). Light-sheet fluorescence microscopy achieves a resolution of ∼26 μm (x-y) (Colombelli and Lorenzo, 2014) with 3D optical sectioning (up to ∼300 nm in z) and high-speed of imaging (2.7 × 104 μm3s–1) (Lu C.-H. et al., 2019) that limits the effects of photobleaching compared to confocal microscopy (Santi, 2011; Power and Huisken, 2017). Light-sheet microscopy was used to visualize the entire adult mouse brain in health (Qi et al., 2019) and Alzheimer’s disease pathology (Liebmann et al., 2016). This emerging technique showed the 3D-distribution of neurons, microglia, the vasculature and tau proteins, throughout the entire brain parenchyma, using IDISCO to clear the tissue (Qi et al., 2019). Further iterations of IDISCO such as FDISCO or SHIELD could be employed as well (Renier et al., 2014; Park et al., 2018). However, light-sheet imaging techniques with in vivo whole mouse brain imaging capacity were not yet proposed (Dodt et al., 2007; Susaki et al., 2014).
Multiphoton Microscopy
Is currently the technique of choice in neuroimmunology for in vivo studies. This technique uses a femtosecond pulsed laser that produces a very high density of photons in a very short time allowing for 2 or 3 photons, respectively, to play the same excitation role as a single photon of full energy. These 2 or 3 photons have half or one third of the quantum energy of a single photon since their wavelength is about two or three times longer than a single excitation photon (typically 800–950 nm for two-photon and 1100–1300 nm for three-photon microscopy) (Svoboda and Yasuda, 2006). Because longer wavelengths are more penetrating, one can explore phenomena deeper into biological tissues (Wang et al., 2018). The addition of energy from several photons takes place at the focal plane of the objective where a large number of photons is needed to make emerge this rare phenomenon, in a single point instead of a large double cone-like structure for single photon excitation. Therefore, the multiphoton technique preserves tissue integrity outside of the focal plane. With two-photon microscopy, the single focal point achieves a spatial resolution approximating 0.42 μm (x-y) and 0.81 μm (z) according to the numerical aperture of the objective (Soeller and Cannell, 1996; Diaspro et al., 2005; Zheng et al., 2019).
Seminal findings that dramatically changed the field of neuroimmunology were obtained with two-photon in vivo microscopy. A pioneer experiment conducted with this technique, in healthy conditions versus after a focal lesion generated with a two-photon laser, debunked the previous paradigm. Microglia are not resting cells in the absence of injury. Instead, they are extremely dynamic, with their processes constantly surveying the entire brain parenchyma with a time course that reaches 5.5 h in adult mice (Davalos et al., 2005; Nimmerjahn et al., 2005). In this seminal work conducted in anesthetized CX3CR1+/GFP reporter mice, time-lapse images of fluorescent microglia were obtained through the skull, using minimally invasive thinned-skull method (Marker et al., 2010; Parkhurst et al., 2013). This achievement is reproduced in Figure 1E. A growing technology for two-photon microscopy is the resonant scanner, which allows acquisition speed to reach 30 frames per second. This high imaging speed is particularly useful for studying the real time activity of neuronal and glial cell populations via in vivo calcium imaging (Grewe et al., 2011; Olmedillas del Moral et al., 2019; Verkhratsky et al., 2019). Other types of microscopy techniques providing time-lapse imaging with high temporal resolution are extensively reviewed here (Mondal, 2014; Ueda et al., 2020).
Beating the Spatial Resolution Limit Using Electrons
Electron microscopy (EM) generates images by interrogating a dehydrated sample with a beam of electrons, which are transmitted (transmission electron microscopy; TEM) or bounced back (scanning electron microscopy; SEM) using specialized detectors. The resulting beam reveals the ultrastructure of the investigated object, e.g., structure of protein, organelle, subcellular compartment or cell. By uncovering these elements from the atomic to the millimeter (and even centimeter) level, ultrastructural techniques have provided important information in neuroscience and neuroimmunology (Genoud et al., 2006; Knott and Genoud, 2013; Savage et al., 2018, 2019a). EM has allowed to shed light on microglia-neuron communication, revealing that 94% of microglial processes directly contact synaptic elements in the healthy adolescent mouse visual cortex (Tremblay et al., 2010). These ultrastructural insights and others contributed to defining the role of microglia in developmental synaptic pruning (Tremblay et al., 2010; Paolicelli et al., 2011; Schafer et al., 2012). The quadripartite synapse model, in which microglia and astrocytes regulate together synaptic function and plasticity, also arose from ultrastructural observations (Bennett, 2007; Tremblay et al., 2011, 2014; Schafer et al., 2013; Sierra and Tremblay, 2014).
TEM
Electron microscopy can reach the resolution at <50 pm (x-y) for TEM compared to 3 Å for cryo-EM (Earl et al., 2017). This allows TEM to provide fine details regarding immune cells morphology, from cell body to finest and most distal processes. As illustrated in Figures 1G,H, this nanoscale resolution allows to identify signs of ‘intracellular’ activity (e.g., organellar changes, phagosomal inclusions) and extracellular activity (e.g., extracellular digestion of cellular debris, intercellular contacts with other glial cells, neurons, and synapses). The quantification of events for each cell type studied is essential to assess the situation of the cell, whether it is in steady state or overwhelmed by phagocytic inclusions requiring digestion, accumulated misfolded proteins in the endoplasmic reticulum or Golgi apparatus, stressed mitochondria or making increased synaptic contacts as previously evaluated for microglia across a range of contexts, including Huntington’s disease pathology (Savage et al., 2020). Additionally, an increasing body of evidence supports the idea that the microglial population is composed of diverse subpopulations endowed with unique intrinsic properties that perform different functions, and display a high degree of spatial and temporal specialization (Stratoulias et al., 2019). Evidence of this microglial heterogeneity was notably provided with TEM. The dark microglia, identified at the ultrastructural level by their electron dense cytoplasm and other markers of cellular stress, as well as extensive interactions with synapses (Bisht et al., 2016b), are rare in healthy young adult mice, but increase in number up to 10-fold with chronic unpredictable or social defeat stress, aging and other pathological contexts, including Alzheimer’s disease pathology (St-Pierre et al., 2020), showing the importance of this approach for the in situ study of microglial functional diversity.
SBF-SEM
3D-EM allows to reconstruct organelles, cytoskeletal elements, subcellular compartments, and cellular relationships, among different cell types including immune cells in situ (Bolasco et al., 2018). Blocks of CNS tissue can be imaged using serial block face SEM, in which the tissue is sequentially imaged at a resolution of ∼10 nm (x-y), cut (25–50 nm thick)(z) with an ultramicrotome mounted inside the SEM chamber, realigned and imaged, to automatically generate z-stacks of serial images (Denk and Horstmann, 2004; Briggman and Bock, 2012; Peddie and Collinson, 2014; Yamasaki et al., 2014). This allows SBF-SEM to reconstruct multiple immune cells, neurons and astrocytes at the nanoscale (Calì et al., 2019). This technique is much faster than the method used in pioneering studies: serial section TEM to image synapses (Fiala and Harris, 2001) or microglial processes interacting with synapses (Tremblay et al., 2010), by aligning and segmenting series of ultrathin sections cut manually and reconstructed semi-manually with Reconstruct (Fiala, 2005). This method allowed the accurate comparison of nucleus to cell body ratio and mitochondrial distribution among the different brain cells, including between individual microglial cells, giving insight into the functional status of microglia and other cells in their 3D microenvironment (Savage et al., 2018). Microglial mitochondria analysis is promising based on reports of mitochondria defects in multiple neurodegenerative disease such as Parkinson’s disease, Huntington’s disease, Alzheimer’s disease and amyotrophic lateral sclerosis (Chaturvedi and Flint Beal, 2013). This whole cell analysis was further used to discriminate monocyte-derived cells from microglia based on their distinctive ultrastructural features in the brain of experimental autoimmune encephalomyelitis mouse models of multiple sclerosis. This distinction between microglia and monocyte-derived cells is normally difficult to achieve in a 2D plane but was possible in 3D while showing the specific interaction of monocytes at the Ranvier node, where they could be initiating the demyelination (Yamasaki et al., 2014).
FIB-SEM
3D-EM is also possible using focused ion beam scanning electron microscopy (FIB-SEM) that removes as little as 3 nm of tissue after each block face image acquisition to create z-stack of images. FIB-SEM produces a higher 3D resolution reaching 3–5 nm (x, y, and z) (Briggman and Bock, 2012), which is required to study the fine geometry of organelles, phagosomes and autophagosomes and cytoskeletal elements at the expense of having a reduced field of view and slower acquisition speed compared with SBF-SEM (Heymann et al., 2006; Knott and Genoud, 2013; Peddie and Collinson, 2014; Savage et al., 2019b). With this level of resolution, FIB-SEM imaging of immune cells allows to discern between cellular elements that are partially surrounded by a microglial process, for instance during synaptic stripping, from the ones that are fully engulfed and internalized (phagocytosed). Different steps of phagocytosis process could be identified, from partial to complete engulfment, given the observation of microglia nibbling small pieces of axon terminals (i.e., performing trogocytosis) in the postnatal mouse hippocampus (Weinhard et al., 2018).
Correlative Light and Electron Microscopy (CLEM)
Has seen its usage escalating over the past years (Begemann and Galic, 2016). Combining the minimally invasive in vivo imaging capacity of two-photon imaging with the nanometer resolution of serial section TEM revealed microglial ability to interact actively with pre- and post-synaptic elements, while some microglial cells had synaptic elements inside their phagosomes under normal physiological conditions (Tremblay et al., 2010). In the context of studying Alzheimer’s disease pathology, brightfield microscopy combined with TEM also allowed to image microglial diversity around amyloid plaques (Bisht et al., 2016a; El Hajj et al., 2019). Figure 1 proposes a correlative approach that combines four different modalities of microscopy to paint the most insightful portrait of microglial diversity. CLEM is a most promising method for investigating in 2D and even 3D microglial relationships with synapses throughout life, from development (Schafer et al., 2012) to aging (Beckman et al., 2019). Synapses can be imaged using confocal microscopy and their ultrastructural relationships with microglia and other immune cells can be imaged using TEM or even SEM with array tomography. This workflow with array tomography allows the captured images to be stitched into larger 2D mosaic, thus providing a better view of their general organization within CNS regions and layers. These images can be correlated with the evaluation of cognitive decline by measuring the loss of synapses but also the integrity of their structure in the prefrontal cortex of human donors (Henstridge et al., 2018). Other important protocols such as the Nanobody-Assisted Tissue Immunostaining for Volumetric Electron microscopy (NATIVE) can provide enhanced staining using single-domain nanobodies with better penetration into the tissue for large scale 3D reconstruction over the entire mouse hippocampus (Fang et al., 2018), and would allow to correlate data from the nanoscale to the brain-region level.
Clinical View on Neuroimmune Imaging
Studying the human brain offers a different set of challenges compared to studying animal models. Several non-invasive techniques developed between 1960 and 1980 (Nutma et al., 2019), especially, allow to have a look at immune system activity inside the human brain. Some of the techniques currently used are positron emission tomography (PET) (Harrison et al., 2014) and magnetic resonance imaging (MRI) (Arnò et al., 2014; Harrison et al., 2015; Rollins et al., 2018) to analyze the integrity of the brain parenchyma and BBB (Kenk et al., 2015; Montagne et al., 2016).
PET
Allows to visualize microglial activity by measuring the dynamic coupling of the radiotracer [11C]PBR28 or [11C]PK11195 to the translocator protein (TSPO). This protein localized to microglial mitochondria can be used as a proxy of their neuroinflammatory and phagocytic activity (Sandiego et al., 2015; Sucksdorff et al., 2019). This makes PET imaging useful to study immune cells in vivo using these radiolabels, but it lacks in spatial resolution (Albrecht et al., 2016). While [11C]PBR28 or [11C]PK11195 tracers are widely used, a range of different microglial receptors and signaling proteins can be targeted via PET radiotracers (Tronel et al., 2017). This approach allowed to observe live changes in microglial activity (notably associated with phagocytosis and/or release of pro-inflammatory mediators) in the human brain after peripheral injection of lipopolysaccharide, a bacterial endotoxin, that was studied to determine how the adaptive immune response exerts a direct effect on behavior between health and disease (Sandiego et al., 2015). For PET imaging, the nature of the radiotracer targets and their low expression levels at steady-state generally limit their use to study reactive functions (e.g., neuroinflammation and exacerbated phagocytosis), and not microglial functions in normal homeostatic conditions (Owen et al., 2012). Using PET in schizophrenic patients with [18 F]-FEPPA, another ligand for TSPO, neuroinflammation was assessed in schizophrenia patients after antipsychotic treatment, revealing no difference, thus suggesting that neuroinflammation happens in earlier stages of the pathology (Kenk et al., 2015). Efforts to find and develop new tracers for the study of non-inflamed microglia would represent a step of major importance for the future investigation of the microglial subpopulations engaged in different functions across a variety of homeostatic and neuroinflammatory states (Beaino et al., 2017; Villa et al., 2018). Other innate immune cells such as monocytes have also been imaged using PET with the tracer [111Indium] oxyquinoline which enters leukocyte cell membrane and provides an effective visualization in the mouse and human blood (Kircher et al., 2008). More mouse PET investigations have shown T cells which can also be labeled by [111Indium] oxyquinoline in animal models (Gong et al., 2011). Further investigation of monocytes and T cells is required in human to translate the findings from animal research. Clinical investigation using high spatial resolution imaging typically uses MRI.
MRI
Is an established non-invasive imaging technique that is used in clinical and preclinical research studies, with various animal models, making it an excellent tool for translational research (Desjardins et al., 2019). MRI with gadolinium has been used in rodents to study BBB integrity and its role in depression together with blood oxygenation dynamics. This is particularly interesting as the BBB is emerging as an active interface between the periphery and the brain that modulates neuroimmune interactions differently between health and disease conditions (Menard et al., 2017). Further than BBB investigation, MRI is a crucial tool that provides a high-resolution view of brain structures across the whole parenchyma in the same session (Ladd et al., 2018). While it is not able to directly image immune cells, MRI is used to image the whole human or animal brain during neuroinflammation, giving insight into immune cells effects on the disruption of the neurovascular unit, demyelination as well as gray and white matter volume reduction (Quarantelli, 2015; Albrecht et al., 2016). One MRI imaging method, diffusion MRI, is widely used to image white matter tracts. This is achieved by measuring water molecule arbitrary movements in the parenchyma to map the cellular architecture of the white matter (Le Bihan and Iima, 2015). This technique was used similarly to PET for the analysis of neuroinflammation in schizophrenia patients. By measuring free-water abnormities with the MRI machine, authors showed limited extracellular space indicating less inflammation in the parenchyma during late schizophrenia stages (Pasternak et al., 2015), which supports the previous evidence.
With its ability to image during the lifetime of the subject, whether animal or human, MRI can be used in experimental settings to investigate disease progression longitudinally, which can then be correlated with postmortem immunostaining for microglia and monocytes (Neuwelt et al., 2009), in addition to various disease hallmarks (Justicia et al., 2008; Jacobs et al., 2012; Tang et al., 2018; Yi et al., 2019). In a rat model of glioma, tumors were imaged during growth progression while studying the microglial/macrophagic changes visualized using ultrasmall superparamagnetic iron oxides (USPIOs). Microglia/macrophages store iron, a mineral necessary for oxidative metabolism, synaptic plasticity, myelination and neurotransmitter synthesis (Nnah and Wessling-Resnick, 2018). USPIOs is also internalized by microglia and macrophages, making the USPIOs-filled cells appearing as dark spots on MRI images. The observations were then correlated with confocal imaging of microglia/macrophages taking up Texas Red-labeled USPIOs in the tumor zone using high spatial resolution confocal imaging (Fleige et al., 2001). Ultimately, correlative [18F]DPA-714-PET and MRI-USPIOs together with confocal imaging allowed to investigate microglia/macrophage properties in an experimental autoimmune encephalomyelitis mouse model of multiple sclerosis. This work showed that microglia/macrophages upregulate TSPO and IBA1 in the demyelinating regions during acute phases, suggesting their dysfunction as a contributing factor to the inflammation seen in multiple sclerosis (Coda et al., 2020).
Conclusion
A Case for Correlative Microscopy and New Development
Research focusing on the neuroimmune component of the CNS is fundamental to provide further information on the properly functioning system with the goal of identifying and understanding the complex subcellular, cellular and intercellular processes that become compromised, dysregulated, or exacerbated in disease, whether the involved cells are stressed, degenerating or senescent. There is a tremendous need to pair clinical PET and MRI together with postmortem CLEM, notably using slide scanning, confocal, TEM or SEM together within an integrative investigation. CLEM can bring researchers closer to the goal of providing further insights into the mechanisms that govern brain immunity and its important consequences on neuronal circuits and behavior across life. This collaborative endeavor aims to maintain health, as well as prevent and treat cancer, but also neurodevelopmental, neuropsychiatric, neurological and neurodegenerative diseases, in which the innate and adaptive immune cells have been critically implicated (Sierra and Tremblay, 2014; Rangachari et al., 2017; Lecours et al., 2018; Bordeleau et al., 2019). By bringing together complementary imaging approaches CLEM can allow researchers to pair the observations in order to provide unprecedented insights into the mechanistic underpinnings of CNS development, function and plasticity. From slide scanning and confocal microscopy to more specialized tools like two-photon imaging and the different emerging types of 3D-EM modalities (Briggman and Bock, 2012; Peddie and Collinson, 2014), STED and light-sheet microscopy, which are now on the rise, will likely become common practice as the technology becomes more accessible.
The perspective of the field is now at a point where several new technologies are just waiting to be used in neuroimmunology. One example of new technology that could provide insights into the role of T cells, in particular, is 4D electron microscopy. This cutting-edge type of microscopy detects photon-induced near-field signals measured at T cell surfaces. This principle allowed the evaluation by electromagnetic measurement of T cell activation and to correlate this finding with the major compatibility binding complexes measured from the near-field signals at the cell surface. These results demonstrated structural changes that serve as biomarkers of T cell sensitivity to immune challenges (Lu Y. et al., 2019). Other techniques are still not offering their advantages in neuroimmunology yet, such as holographic microscopy which brings the resolution of electron microscopy to the order of the Armstrong. This would raise the imaging of phagosomal inclusions highlighted in Figure 1H to a new level, where could be determined the content of these vesicles. Multi-isotope imaging mass spectrometry would also bring many possibilities to the field. This imaging technique is able to image and quantify molecules in the samples, which presents great potential to identify new molecular targets in the neuroimmunology field (Arrojo E Drigo et al., 2019; Madan et al., 2019).
Author Contributions
MC was responsible for the manuscript plan and management, writing of the correlative and clinical imaging sections while taking care of the overall revision and formatting of the manuscript, and creating the figure included in the manuscript. M-ÈR was responsible for writing the fluorescent imaging section and creating the table. FG was in charge of writing the electron microscopy section of the manuscript. MD and M-ÈT were in charge of revising the manuscript and providing theoretical and writing support to all authors while M-ÈT contributed significantly to the structure and ideas content of the manuscript.
Funding
This work was supported by grants from the Canadian Institutes of Health Research (CIHR; Foundation Scheme), Natural Sciences and Engineering Research Council of Canada (Discovery Grant), and NEURON-ERANET Synaptic Dysfunction in Disorders of the Nervous System (MicroSynDep consortium) to M-ÈT. MC was supported by a Master’s Training Award from Fonds de Recherche du Québec – Santé (FRQS). FG was supported by a Mexican Council of Science and Technology (CONACYT) training grant. M-ÈT holds a Canada Research Chair (Tier 2) in Neurobiology of Aging and Cognition.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to our colleagues Sammy Weiser Novak and Jean-Christophe Comte for sharing their insights and contributing to our manuscript through their expertise in electron microscopy and multiphoton microscopy.
References
Albrecht, D. S., Granziera, C., Hooker, J. M., and Loggia, M. L. (2016). In vivo imaging of human neuroinflammation. ACS Chem. Neurosci. 7, 470–483. doi: 10.1021/acschemneuro.6b00056
Arnò, B., Grassivaro, F., Rossi, C., Bergamaschi, A., Castiglioni, V., Furlan, R., et al. (2014). Neural progenitor cells orchestrate microglia migration and positioning into the developing cortex. Nat. Commun. 5:5611. doi: 10.1038/ncomms6611
Arrojo E Drigo, R., Lev-Ram, V., Tyagi, S., Ramachandra, R., Deerinck, T., Bushong, E., et al. (2019). Age mosaicism across multiple scales in adult tissues. Cell Metab. 30, 343–351.e3. doi: 10.1016/j.cmet.2019.05.010
Augusto-Oliveira, M., Arrifano, G. P., Lopes-Araújo, A., Santos-Sacramento, L., Takeda, P. Y., Anthony, D. C., et al. (2019). What do microglia really do in healthy adult brain? Cells 8:1293. doi: 10.3390/cells8101293
Bankhead, P., Loughrey, M. B., Fernández, J. A., Dombrowski, Y., McArt, D. G., Dunne, P. D., et al. (2017). QuPath: open source software for digital pathology image analysis. Sci. Rep. 7:16878. doi: 10.1038/s41598-017-17204-5
Bayerl, S. H., Ghori, A., Nieminen-Kelhä, M., Adage, T., Breitenbach, J., Vajkoczy, P., et al. (2019). In vitro and in vivo testing of a novel local nicardipine delivery system to the brain: a preclinical study. J. Neurosurg. 132, 465–472. doi: 10.3171/2018.9.JNS173085
Beaino, W., Janssen, B., Kooij, G., van der Pol, S. M. A., van Het Hof, B., van Horssen, J., et al. (2017). Purinergic receptors P2Y12R and P2X7R: potential targets for PET imaging of microglia phenotypes in multiple sclerosis. J. Neuroinflammation 14:259. doi: 10.1186/s12974-017-1034-z
Beckman, D., Ott, S., Donis-Cox, K., Janssen, W. G., Bliss-Moreau, E., Rudebeck, P. H., et al. (2019). Oligomeric Aβ in the monkey brain impacts synaptic integrity and induces accelerated cortical aging. Proc. Natl. Acad. Sci. U.S.A. 116, 26239–26246. doi: 10.1073/pnas.1902301116
Begemann, I., and Galic, M. (2016). Correlative light electron microscopy: connecting synaptic structure and function. Front. Synaptic Neurosci. 8:28. doi: 10.3389/fnsyn.2016.00028
Bellver-Landete, V., Bretheau, F., Mailhot, B., Vallières, N., Lessard, M., Janelle, M.-E., et al. (2019). Microglia are an essential component of the neuroprotective scar that forms after spinal cord injury. Nat. Commun. 10:518. doi: 10.1038/s41467-019-08446-0
Bennett, M. R. (2007). Synaptic P2X7 receptor regenerative-loop hypothesis for depression. Aust. N. Z. J. Psychiatry 41, 563–571. doi: 10.1080/00048670701399994
Berning, S., Willig, K. I., Steffens, H., Dibaj, P., and Hell, S. W. (2012). Nanoscopy in a living mouse brain. Science 335:551. doi: 10.1126/science.1215369
Bisht, K., El Hajj, H., Savage, J. C., Sánchez, M. G., and Tremblay, M. -È (2016a). Correlative light and electron microscopy to study microglial interactions with β-Amyloid plaques. J. Vis. Exp. JoVE 112:54060. doi: 10.3791/54060
Bisht, K., Sharma, K. P., Lecours, C., Gabriela Sánchez, M., El Hajj, H., Milior, G., et al. (2016b). Dark microglia: a new phenotype predominantly associated with pathological states. Glia 64, 826–839. doi: 10.1002/glia.22966
Bolasco, G., Weinhard, L., Boissonnet, T., Neujahr, R., and Gross, C. T. (2018). Three-dimensional nanostructure of an intact microglia cell. Front. Neuroanat. 12:105. doi: 10.3389/fnana.2018.00105
Bordeleau, M., Carrier, M., Luheshi, G. N., and Tremblay, M. -È (2019). Microglia along sex lines: from brain colonization, maturation and function, to implication in neurodevelopmental disorders. Semin. Cell Dev. Biol. 94, 152–163. doi: 10.1016/j.semcdb.2019.06.001
Briggman, K. L., and Bock, D. D. (2012). Volume electron microscopy for neuronal circuit reconstruction. Curr. Opin. Neurobiol. 22, 154–161. doi: 10.1016/j.conb.2011.10.022
Budde, M. D., and Annese, J. (2013). Quantification of anisotropy and fiber orientation in human brain histological sections. Front. Integr. Neurosci. 7:3. doi: 10.3389/fnint.2013.00003
Butovsky, O., Jedrychowski, M. P., Moore, C. S., Cialic, R., Lanser, A. J., Gabriely, G., et al. (2014). Identification of a unique TGF-β dependent molecular and functional signature in microglia. Nat. Neurosci. 17, 131–143. doi: 10.1038/nn.3599
Buttgereit, A., Lelios, I., Yu, X., Vrohlings, M., Krakoski, N. R., Gautier, E. L., et al. (2016). Sall1 is a transcriptional regulator defining microglia identity and function. Nat. Immunol. 17, 1397–1406. doi: 10.1038/ni.3585
Calì, C., Agus, M., Kare, K., Boges, D. J., Lehväslaiho, H., Hadwiger, M., et al. (2019). 3D cellular reconstruction of cortical glia and parenchymal morphometric analysis from serial block-face electron microscopy of juvenile rat. Prog. Neurobiol. 183:101696. doi: 10.1016/j.pneurobio.2019.101696
Cătălin, B., Stopper, L., Bălşeanu, T.-A., and Scheller, A. (2017). The in situ morphology of microglia is highly sensitive to the mode of tissue fixation. J. Chem. Neuroanat. 86, 59–66. doi: 10.1016/j.jchemneu.2017.08.007
Chaturvedi, R. K., and Flint Beal, M. (2013). Mitochondrial diseases of the brain. Free Radic. Biol. Med. 63, 1–29. doi: 10.1016/j.freeradbiomed.2013.03.018
Chen, Z., Shin, D., Chen, S., Mikhail, K., Hadass, O., Tomlison, B. N., et al. (2014). Histological quantitation of brain injury using whole slide imaging: a pilot validation study in mice. PLoS One 9:e92133. doi: 10.1371/journal.pone.0092133
Chéreau, R., Tønnesen, J., and Nägerl, U. V. (2015). STED microscopy for nanoscale imaging in living brain slices. Methods San Diego Calif. 88, 57–66. doi: 10.1016/j.ymeth.2015.06.006
Coda, A. R., Anzilotti, S., Boscia, F., Greco, A., Panico, M., Gargiulo, S., et al. (2020). In vivo imaging of CNS microglial activation/macrophage infiltration with combined [18F]DPA-714-PET and SPIO-MRI in a mouse model of relapsing remitting experimental autoimmune encephalomyelitis. Eur. J. Nucl. Med. Mol. Imaging doi: 10.1007/s00259-020-04842-7 [Epub ahead of print].
Colombelli, J., and Lorenzo, C. (2014). “Light sheet fluorescence microscopy applications for multicellular systems,” in Fluorescence Microscopy, ed. Academic Press (Amsterdam: Elsevier), 109–120. doi: 10.1016/B978-0-12-409513-7.00008-7
Daigle, T. L., Madisen, L., Hage, T. A., Valley, M. T., Knoblich, U., Larsen, R. S., et al. (2018). A suite of transgenic driver and reporter mouse lines with enhanced brain-cell-type targeting and functionality. Cell 174, 465–480.e22. doi: 10.1016/j.cell.2018.06.035
Davalos, D., Grutzendler, J., Yang, G., Kim, J. V., Zuo, Y., Jung, S., et al. (2005). ATP mediates rapid microglial response to local brain injury in vivo. Nat. Neurosci. 8, 752–758. doi: 10.1038/nn1472
Denk, W., and Horstmann, H. (2004). Serial block-face scanning electron microscopy to reconstruct three-dimensional tissue nanostructure. PLoS Biol. 2:e329. doi: 10.1371/journal.pbio.0020329
Desjardins, M., Kılıç, K., Thunemann, M., Mateo, C., Holland, D., Ferri, C. G. L., et al. (2019). Awake mouse imaging: from two-photon microscopy to blood oxygen level-dependent functional magnetic resonance imaging. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 4, 533–542. doi: 10.1016/j.bpsc.2018.12.002
Diaspro, A., Chirico, G., and Collini, M. (2005). Two-photon fluorescence excitation and related techniques in biological microscopy. Q. Rev. Biophys. 38, 97–166. doi: 10.1017/S0033583505004129
D’Mello, C., Le, T., and Swain, M. G. (2009). Cerebral microglia recruit monocytes into the brain in response to tumor necrosis Factorα signaling during peripheral organ inflammation. J. Neurosci. 29, 2089–2102. doi: 10.1523/JNEUROSCI.3567-08.2009
Dodt, H.-U., Leischner, U., Schierloh, A., Jährling, N., Mauch, C. P., Deininger, K., et al. (2007). Ultramicroscopy: three-dimensional visualization of neuronal networks in the whole mouse brain. Nat. Methods 4, 331–336. doi: 10.1038/nmeth1036
Earl, L. A., Falconieri, V., Milne, J. L., and Subramaniam, S. (2017). Cryo-EM: beyond the microscope. Curr. Opin. Struct. Biol. 46, 71–78. doi: 10.1016/j.sbi.2017.06.002
El Hajj, H., Savage, J. C., Bisht, K., Parent, M., Vallières, L., Rivest, S., et al. (2019). Ultrastructural evidence of microglial heterogeneity in Alzheimer’s disease amyloid pathology. J. Neuroinflammation 16:87. doi: 10.1186/s12974-019-1473-9
Elliott, A. D. (2020). Confocal microscopy: principles and modern practices. Curr. Protoc. Cytom. 92:e68. doi: 10.1002/cpcy.68
Evilsizor, M. N., Ray-Jones, H. F., Lifshitz, J., and Ziebell, J. (2015). Primer for immunohistochemistry on cryosectioned rat brain tissue: example staining for microglia and neurons. J. Vis. Exp. JoVE 99:e52293. doi: 10.3791/52293
Fang, T., Lu, X., Berger, D., Gmeiner, C., Cho, J., Schalek, R., et al. (2018). Nanobody immunostaining for correlated light and electron microscopy with preservation of ultrastructure. Nat. Methods 15, 1029–1032. doi: 10.1038/s41592-018-0177-x
Feng, G., Mellor, R. H., Bernstein, M., Keller-Peck, C., Nguyen, Q. T., Wallace, M., et al. (2000). Imaging neuronal subsets in transgenic mice expressing multiple spectral variants of GFP. Neuron 28, 41–51. doi: 10.1016/s0896-6273(00)00084-2
Fiala, J. C. (2005). Reconstruct: a free editor for serial section microscopy. J. Microsc. 218, 52–61. doi: 10.1111/j.1365-2818.2005.01466.x
Fiala, J. C., and Harris, K. M. (2001). Extending unbiased stereology of brain ultrastructure to three-dimensional volumes. J. Am. Med. Inform. Assoc. JAMIA 8, 1–16. doi: 10.1136/jamia.2001.0080001
Fiolka, R. (2019). Resolution upgrades for light-sheet microscopy. Nat. Methods 16, 813–814. doi: 10.1038/s41592-019-0542-4
Fleige, G., Nolte, C., Synowitz, M., Seeberger, F., Kettenmann, H., and Zimmer, C. (2001). Magnetic labeling of activated microglia in experimental gliomas. Neoplasia N.Y.N 3, 489–499. doi: 10.1038/sj.neo.7900176
Genoud, C., Quairiaux, C., Steiner, P., Hirling, H., Welker, E., and Knott, G. W. (2006). Plasticity of astrocytic coverage and glutamate transporter expression in adult mouse cortex. PLoS Biol. 4:e343. doi: 10.1371/journal.pbio.0040343
Ginhoux, F., Greter, M., Leboeuf, M., Nandi, S., See, P., Gokhan, S., et al. (2010). Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science 330, 841–845. doi: 10.1126/science.1194637
Ginhoux, F., Lim, S., Hoeffel, G., Low, D., and Huber, T. (2013). Origin and differentiation of microglia. Front. Cell. Neurosci. 7:45. doi: 10.3389/fncel.2013.00045
Glaser, A. K., Reder, N. P., Chen, Y., McCarty, E. F., Yin, C., Wei, L., et al. (2017). Light-sheet microscopy for slide-free non-destructive pathology of large clinical specimens. Nat. Biomed. Eng. 1:0084. doi: 10.1038/s41551-017-0084
Gomez Perdiguero, E., Klapproth, K., Schulz, C., Busch, K., Azzoni, E., Crozet, L., et al. (2015). Tissue-resident macrophages originate from yolk-sac-derived erythro-myeloid progenitors. Nature 518, 547–551. doi: 10.1038/nature13989
Gong, N., Liu, J., Reynolds, A. D., Gorantla, S., Mosley, R. L., and Gendelman, H. E. (2011). Brain ingress of regulatory T cells in a murine model of HIV-1 encephalitis. J. Neuroimmunol. 230, 33–41. doi: 10.1016/j.jneuroim.2010.08.014
González Ibanez, F., Picard, K., Bordelau, M., Sharma, K., Bisht, K., and Tremblay, M.-È. (2019). Immunofluorescence staining using IBA1 and TMEM119 for microglial density, morphology and peripheral myeloid cell infiltration analysis in mouse brain. J. Vis. Exp. JoVE. doi: 10.3791/60510
Graf, B. W., and Boppart, S. A. (2010). Imaging and analysis of three-dimensional cell culture models. Methods Mol. Biol. Clifton NJ 591, 211–227. doi: 10.1007/978-1-60761-404-3_13
Grewe, B. F., Voigt, F. F., van’t Hoff, M., and Helmchen, F. (2011). Fast two-layer two-photon imaging of neuronal cell populations using an electrically tunable lens. Biomed. Opt. Express 2, 2035–2046. doi: 10.1364/BOE.2.002035
Harrison, N. A., Cooper, E., Dowell, N. G., Keramida, G., Voon, V., Critchley, H. D., et al. (2015). Quantitative magnetization transfer imaging as a biomarker for effects of systemic inflammation on the brain. Biol. Psychiatry 78, 49–57. doi: 10.1016/j.biopsych.2014.09.023
Harrison, N. A., Doeller, C. F., Voon, V., Burgess, N., and Critchley, H. D. (2014). Peripheral inflammation acutely impairs human spatial memory via actions on medial temporal lobe glucose metabolism. Biol. Psychiatry 76, 585–593. doi: 10.1016/j.biopsych.2014.01.005
Hein, B., Willig, K. I., and Hell, S. W. (2008). Stimulated emission depletion (STED) nanoscopy of a fluorescent protein-labeled organelle inside a living cell. Proc. Natl. Acad. Sci. U.S.A. 105, 14271–14276. doi: 10.1073/pnas.0807705105
Hell, S. W., and Wichmann, J. (1994). Breaking the diffraction resolution limit by stimulated emission: stimulated-emission-depletion fluorescence microscopy. Opt. Lett. 19, 780–782. doi: 10.1364/ol.19.000780
Helmchen, F., and Denk, W. (2005). Deep tissue two-photon microscopy. Nat. Methods 2, 932–940. doi: 10.1038/nmeth818
Henstridge, C. M., Sideris, D. I., Carroll, E., Rotariu, S., Salomonsson, S., Tzioras, M., et al. (2018). Synapse loss in the prefrontal cortex is associated with cognitive decline in amyotrophic lateral sclerosis. Acta Neuropathol. (Berl.) 135, 213–226. doi: 10.1007/s00401-017-1797-4
Herz, J., Zinselmeyer, B. H., and McGavern, D. B. (2012). Two-photon imaging of microbial immunity in living tissues. Microsc. Microanal. Off. J. Microsc. Soc. Am. Microbeam Anal. Soc. Microsc. Soc. Can. 18, 730–741. doi: 10.1017/S1431927612000281
Heymann, J. A. W., Hayles, M., Gestmann, I., Giannuzzi, L. A., Lich, B., and Subramaniam, S. (2006). Site-specific 3D imaging of cells and tissues with a dual beam microscope. J. Struct. Biol. 155, 63–73. doi: 10.1016/j.jsb.2006.03.006
Jacobs, A. H., and Tavitian, B. INMiND consortium (2012). Noninvasive molecular imaging of neuroinflammation. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 32, 1393–1415. doi: 10.1038/jcbfm.2012.53
Jung, S., Aliberti, J., Graemmel, P., Sunshine, M. J., Kreutzberg, G. W., Sher, A., et al. (2000). Analysis of fractalkine receptor CX(3)CR1 function by targeted deletion and green fluorescent protein reporter gene insertion. Mol. Cell. Biol. 20, 4106–4114. doi: 10.1128/mcb.20.11.4106-4114.2000
Justicia, C., Ramos-Cabrer, P., and Hoehn, M. (2008). MRI detection of secondary damage after stroke: chronic iron accumulation in the thalamus of the rat brain. Stroke 39, 1541–1547. doi: 10.1161/STROKEAHA.107.503565
Katsumoto, A., Lu, H., Miranda, A. S., and Ransohoff, R. M. (2014). Ontogeny and functions of CNS macrophages. J. Immunol. Baltim. Md 1950, 2615–2621. doi: 10.4049/jimmunol.1400716
Keller, P. J., and Ahrens, M. B. (2015). Visualizing whole-brain activity and development at the single-cell level using light-sheet microscopy. Neuron 85, 462–483. doi: 10.1016/j.neuron.2014.12.039
Kenk, M., Selvanathan, T., Rao, N., Suridjan, I., Rusjan, P., Remington, G., et al. (2015). Imaging neuroinflammation in gray and white matter in schizophrenia: an in-vivo PET study with [18F]-FEPPA. Schizophr. Bull. 41, 85–93. doi: 10.1093/schbul/sbu157
Kipnis, J., Cohen, H., Cardon, M., Ziv, Y., and Schwartz, M. (2004). T cell deficiency leads to cognitive dysfunction: implications for therapeutic vaccination for schizophrenia and other psychiatric conditions. Proc. Natl. Acad. Sci. U.S.A. 101, 8180–8185. doi: 10.1073/pnas.0402268101
Kircher, M. F., Grimm, J., Swirski, F. K., Libby, P., Gerszten, R. E., Allport, J. R., et al. (2008). Noninvasive in vivo imaging of monocyte trafficking to atherosclerotic lesions. Circulation 117, 388–395. doi: 10.1161/CIRCULATIONAHA.107.719765
Klar, T. A., Engel, E., and Hell, S. W. (2001). Breaking Abbe’s diffraction resolution limit in fluorescence microscopy with stimulated emission depletion beams of various shapes. Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 64:066613. doi: 10.1103/PhysRevE.64.066613
Konishi, H., Kobayashi, M., Kunisawa, T., Imai, K., Sayo, A., Malissen, B., et al. (2017). Siglec-H is a microglia-specific marker that discriminates microglia from CNS-associated macrophages and CNS-infiltrating monocytes. Glia 65, 1927–1943. doi: 10.1002/glia.23204
Kyriazis, A. D., Noroozizadeh, S., Refaee, A., Choi, W., Chu, L.-T., Bashir, A., et al. (2019). An end-to-end system for automatic characterization of Iba1 immunopositive microglia in whole slide imaging. Neuroinformatics 17, 373–389. doi: 10.1007/s12021-018-9405-x
Ladd, M. E., Bachert, P., Meyerspeer, M., Moser, E., Nagel, A. M., Norris, D. G., et al. (2018). Pros and cons of ultra-high-field MRI/MRS for human application. Prog. Nucl. Magn. Reson. Spectrosc. 109, 1–50. doi: 10.1016/j.pnmrs.2018.06.001
Le Bihan, D., and Iima, M. (2015). Diffusion magnetic resonance imaging: what water tells us about biological tissues. PLoS Biol. 13:e1002203. doi: 10.1371/journal.pbio.1002203
Lecours, C., Bordeleau, M., Cantin, L., Parent, M., Paolo, T. D., and Tremblay, M. -È (2018). Microglial implication in Parkinson’s disease: loss of beneficial physiological roles or gain of inflammatory functions? Front. Cell. Neurosci. 12:282. doi: 10.3389/fncel.2018.00282
Leone, C., Le Pavec, G., Même, W., Porcheray, F., Samah, B., Dormont, D., et al. (2006). Characterization of human monocyte-derived microglia-like cells. Glia 54, 183–192. doi: 10.1002/glia.20372
Lewitus, G. M., and Schwartz, M. (2009). Behavioral immunization: immunity to self-antigens contributes to psychological stress resilience. Mol. Psychiatry 14, 532–536. doi: 10.1038/mp.2008.103
Liebmann, T., Renier, N., Bettayeb, K., Greengard, P., Tessier-Lavigne, M., and Flajolet, M. (2016). Three-dimensional study of Alzheimer’s disease Hallmarks using the iDISCO clearing method. Cell Rep. 16, 1138–1152. doi: 10.1016/j.celrep.2016.06.060
Lodygin, D., Hermann, M., Schweingruber, N., Flügel-Koch, C., Watanabe, T., Schlosser, C., et al. (2019). β-Synuclein-reactive T cells induce autoimmune CNS grey matter degeneration. Nature 566, 503–508. doi: 10.1038/s41586-019-0964-2
Lu, C.-H., Tang, W.-C., Liu, Y.-T., Chang, S.-W., Wu, F. C. M., Chen, C.-Y., et al. (2019). Lightsheet localization microscopy enables fast, large-scale, and three-dimensional super-resolution imaging. Commun. Biol. 2:177. doi: 10.1038/s42003-019-0403-9
Lu, Y., Yoo, B.-K., Ng, A. H. C., Kim, J., Yeom, S., Tang, J., et al. (2019). 4D electron microscopy of T cell activation. Proc. Natl. Acad. Sci. U.S.A. 116, 22014–22019. doi: 10.1073/pnas.1914078116
Madan, I., Vanacore, G. M., Pomarico, E., Berruto, G., Lamb, R. J., McGrouther, D., et al. (2019). Holographic imaging of electromagnetic fields via electron-light quantum interference. Sci. Adv. 5:eaav8358. doi: 10.1126/sciadv.aav8358
Marker, D. F., Tremblay, M.-E., Lu, S.-M., Majewska, A. K., and Gelbard, H. A. (2010). A thin-skull window technique for chronic two-photon in vivo imaging of murine microglia in models of neuroinflammation. J. Vis. Exp. JoVE 43, 2059. doi: 10.3791/2059
Martell, J. D., Deerinck, T. J., Lam, S. S., Ellisman, M. H., and Ting, A. Y. (2017). Electron microscopy using the genetically encoded APEX2 tag in cultured mammalian cells. Nat. Protoc. 12, 1792–1816. doi: 10.1038/nprot.2017.065
Masuda, T., Amann, L., Sankowski, R., Staszewski, O., Lenz, M., and Errico, D. (2020). Novel Hexb-based tools for studying microglia in the CNS. Nat. Immunol. 21, 802–815. doi: 10.1038/s41590-020-0707-4
Menard, C., Pfau, M. L., Hodes, G. E., Kana, V., Wang, V. X., Bouchard, S., et al. (2017). Social stress induces neurovascular pathology promoting depression. Nat. Neurosci. 20, 1752–1760. doi: 10.1038/s41593-017-0010-3
Mikula, S., Trotts, I., Stone, J. M., and Jones, E. G. (2007). Internet-enabled high-resolution brain mapping and virtual microscopy. NeuroImage 35, 9–15. doi: 10.1016/j.neuroimage.2006.11.053
Mittal, K., Eremenko, E., Berner, O., Elyahu, Y., Strominger, I., Apelblat, D., et al. (2019). CD4 T cells induce a subset of MHCII-expressing microglia that attenuates alzheimer pathology. iScience 16, 298–311. doi: 10.1016/j.isci.2019.05.039
Mondal, P. P. (2014). Temporal resolution in fluorescence imaging. Front. Mol. Biosci. 1:11. doi: 10.3389/fmolb.2014.00011
Montagne, A., Toga, A. W., and Zlokovic, B. V. (2016). Blood-brain barrier permeability and gadolinium: benefits and potential pitfalls in research. JAMA Neurol. 73, 13–14. doi: 10.1001/jamaneurol.2015.2960
Morimoto, K., and Nakajima, K. (2019). Role of the immune system in the development of the central nervous system. Front. Neurosci. 13:916. doi: 10.3389/fnins.2019.00916
Neuwelt, E. A., Hamilton, B. E., Varallyay, C. G., Rooney, W. R., Edelman, R. D., Jacobs, P. M., et al. (2009). Ultrasmall superparamagnetic iron oxides (USPIOs): a future alternative magnetic resonance (MR) contrast agent for patients at risk for nephrogenic systemic fibrosis (NSF)? Kidney Int. 75, 465–474. doi: 10.1038/ki.2008.496
Nimmerjahn, A., Kirchhoff, F., and Helmchen, F. (2005). Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 308, 1314–1318. doi: 10.1126/science.1110647
Niraula, A., Wang, Y., Godbout, J. P., and Sheridan, J. F. (2018). Corticosterone production during repeated social defeat causes monocyte mobilization from the bone marrow, glucocorticoid resistance, and neurovascular adhesion molecule expression. J. Neurosci. Off. J. Soc. Neurosci. 38, 2328–2340. doi: 10.1523/JNEUROSCI.2568-17.2018
Nnah, I. C., and Wessling-Resnick, M. (2018). Brain iron homeostasis: a focus on microglial iron. Pharm. Basel Switz. 11:129. doi: 10.3390/ph11040129
Nutma, E., Willison, H., Martino, G., and Amor, S. (2019). Neuroimmunology – the past, present and future. Clin. Exp. Immunol. 197, 278–293. doi: 10.1111/cei.13279
Olmedillas del Moral, M., Asavapanumas, N., Uzcátegui, N., and Garaschuk, O. (2019). Healthy brain aging modifies microglial calcium signaling in vivo. Int. J. Mol. Sci. 20:589. doi: 10.3390/ijms20030589
Oreopoulos, J., Berman, R., and Browne, M. (2014). Spinning-disk confocal microscopy: present technology and future trends. Methods Cell Biol. 123, 153–175. doi: 10.1016/B978-0-12-420138-5.00009-4
Otxoa-de-Amezaga, A., Miró-Mur, F., Pedragosa, J., Gallizioli, M., Justicia, C., Gaja-Capdevila, N., et al. (2019). Microglial cell loss after ischemic stroke favors brain neutrophil accumulation. Acta Neuropathol. (Berl.) 137, 321–341. doi: 10.1007/s00401-018-1954-4
Owen, D. R., Yeo, A. J., Gunn, R. N., Song, K., Wadsworth, G., Lewis, A., et al. (2012). An 18-kDa translocator protein (TSPO) polymorphism explains differences in binding affinity of the PET radioligand PBR28. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 32, 1–5. doi: 10.1038/jcbfm.2011.147
Paolicelli, R. C., Bolasco, G., Pagani, F., Maggi, L., Scianni, M., Panzanelli, P., et al. (2011). Synaptic pruning by microglia is necessary for normal brain development. Science 333, 1456–1458. doi: 10.1126/science.1202529
Park, Y.-G., Sohn, C. H., Chen, R., McCue, M., Yun, D. H., Drummond, G. T., et al. (2018). Protection of tissue physicochemical properties using polyfunctional crosslinkers. Nat. Biotechnol. doi: 10.1038/nbt.4281 [Epub ahead of print].
Parkhurst, C. N., Yang, G., Ninan, I., Savas, J. N., Yates, J. R., Lafaille, J. J., et al. (2013). Microglia promote learning-dependent synapse formation through BDNF. Cell 155, 1596–1609. doi: 10.1016/j.cell.2013.11.030
Pasternak, O., Westin, C.-F., Dahlben, B., Bouix, S., and Kubicki, M. (2015). The extent of diffusion MRI markers of neuroinflammation and white matter deterioration in chronic schizophrenia. Schizophr. Res. 161, 113–118. doi: 10.1016/j.schres.2014.07.031
Peddie, C. J., and Collinson, L. M. (2014). Exploring the third dimension: volume electron microscopy comes of age. Micron Oxf. Engl. 1993, 9–19. doi: 10.1016/j.micron.2014.01.009
Perego, C., Fumagalli, S., and De Simoni, M.-G. (2013). Three-dimensional confocal analysis of microglia/macrophage markers of polarization in experimental brain injury. J. Vis. Exp. JoVE 79:50605. doi: 10.3791/50605
Pérez-Alvarez, A., Araque, A., and Martín, E. D. (2013). Confocal microscopy for astrocyte in vivo imaging: Recycle and reuse in microscopy. Front. Cell. Neurosci. 7:51. doi: 10.3389/fncel.2013.00051
Pfeiffer, T., Poll, S., Bancelin, S., Angibaud, J., Inavalli, V. K., Keppler, K., et al. (2018). Chronic 2P-STED imaging reveals high turnover of dendritic spines in the hippocampus in vivo. eLife 7:e34700. doi: 10.7554/eLife.34700
Plemel, J. R., Stratton, J. A., Michaels, N. J., Rawji, K. S., Zhang, E., Sinha, S., et al. (2020). Microglia response following acute demyelination is heterogeneous and limits infiltrating macrophage dispersion. Sci. Adv. 6:eaay6324. doi: 10.1126/sciadv.aay6324
Power, R. M., and Huisken, J. (2017). A guide to light-sheet fluorescence microscopy for multiscale imaging. Nat. Methods 14, 360–373. doi: 10.1038/nmeth.4224
Prinz, M., and Priller, J. (2014). Microglia and brain macrophages in the molecular age: from origin to neuropsychiatric disease. Nat. Rev. Neurosci. 15, 300–312. doi: 10.1038/nrn3722
Prinz, M., and Priller, J. (2017). The role of peripheral immune cells in the CNS in steady state and disease. Nat. Neurosci. 20, 136–144. doi: 10.1038/nn.4475
Qi, Y., Yu, T., Xu, J., Wan, P., Ma, Y., Zhu, J., et al. (2019). FDISCO: advanced solvent-based clearing method for imaging whole organs. Sci. Adv. 5:eaau8355. doi: 10.1126/sciadv.aau8355
Quarantelli, M. (2015). MRI/MRS in neuroinflammation: methodology and applications. Clin. Transl. Imaging 3, 475–489. doi: 10.1007/s40336-015-0142-y
Rangachari, M., Kerfoot, S. M., Arbour, N., and Alvarez, J. I. (2017). Editorial: lymphocytes in MS and EAE: more than just a CD4+ World. Front. Immunol. 8:133. doi: 10.3389/fimmu.2017.00133
Renier, N., Wu, Z., Simon, D. J., Yang, J., Ariel, P., and Tessier-Lavigne, M. (2014). iDISCO: a simple, rapid method to immunolabel large tissue samples for volume imaging. Cell 159, 896–910. doi: 10.1016/j.cell.2014.10.010
Roetzer, T., Leskovar, K., Peter, N., Furtner, J., Muck, M., Augustin, M., et al. (2019). Evaluating cellularity and structural connectivity on whole brain slides using a custom-made digital pathology pipeline. J. Neurosci. Methods 311, 215–221. doi: 10.1016/j.jneumeth.2018.10.029
Rollins, C. P. E., Gallino, D., Kong, V., Ayranci, G., Devenyi, G. A., Germann, J., et al. (2018). Contributions of a high-fat diet to Alzheimer’s disease-related decline: a longitudinal behavioural and structural neuroimaging study in mouse models. Neuroimage Clin. 21:101606. doi: 10.1016/j.nicl.2018.11.016
Rust, M. J., Bates, M., and Zhuang, X. (2006). Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy (STORM). Nat. Methods 3, 793–795. doi: 10.1038/nmeth929
Sandiego, C. M., Gallezot, J.-D., Pittman, B., Nabulsi, N., Lim, K., Lin, S.-F., et al. (2015). Imaging robust microglial activation after lipopolysaccharide administration in humans with PET. Proc. Natl. Acad. Sci. U.S.A. 112, 12468–12473. doi: 10.1073/pnas.1511003112
Santi, P. A. (2011). Light sheet fluorescence microscopy: a review. J. Histochem. Cytochem. Off. J. Histochem. Soc. 59, 129–138. doi: 10.1369/0022155410394857
Savage, J. C., Carrier, M., and Tremblay, M.-E. (2019a). Morphology of microglia across contexts of health and disease. Methods Mol. Biol. Clifton NJ 2034, 13–26. doi: 10.1007/978-1-4939-9658-2_2
Savage, J. C., Novak, S. W., and Tremblay, M.-E. (2019b). Imaging and reconstructing microglia in 3 dimensions using FIB-SEM. Microsc. Microanal. 25, 1316–1317. doi: 10.1017/S1431927619007311
Savage, J. C., Picard, K., González-Ibáñez, F., and Tremblay, M. -È (2018). A brief history of microglial ultrastructure: distinctive features, phenotypes, and functions discovered over the past 60years by electron microscopy. Front. Immunol. 9:803. doi: 10.3389/fimmu.2018.00803
Savage, J. C., St-Pierre, M.-K., Carrier, M., El Hajj, H., Novak, S. W., Sanchez, M. G., et al. (2020). Microglial physiological properties and interactions with synapses are altered at presymptomatic stages in a mouse model of Huntington’s disease pathology. J. Neuroinflammation 17:98. doi: 10.1186/s12974-020-01782-9
Savage, J. C., St-Pierre, M.-K., Hui, C. W., and Tremblay, M.-E. (2019c). Microglial ultrastructure in the hippocampus of a lipopolysaccharide-induced sickness mouse model. Front. Neurosci. 13:1340. doi: 10.3389/fnins.2019.01340
Schafer, D. P., Lehrman, E. K., Kautzman, A. G., Koyama, R., Mardinly, A. R., Yamasaki, R., et al. (2012). Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron 74, 691–705. doi: 10.1016/j.neuron.2012.03.026
Schafer, D. P., Lehrman, E. K., and Stevens, B. (2013). The “quad-partite” synapse: microglia-synapse interactions in the developing and mature CNS. Glia 61, 24–36. doi: 10.1002/glia.22389
Schermelleh, L., Carlton, P. M., Haase, S., Shao, L., Winoto, L., Kner, P., et al. (2008). Subdiffraction multicolor imaging of the nuclear periphery with 3D structured illumination microscopy. Science 320, 1332–1336. doi: 10.1126/science.1156947
Schneider, C. A., Velez, D. X. F., Azevedo, R., Hoover, E. M., Tran, C. J., Lo, C., et al. (2019). Imaging the dynamic recruitment of monocytes to the blood–brain barrier and specific brain regions during Toxoplasma gondii infection. Proc. Natl. Acad. Sci. U.S.A. 116, 24796–24807. doi: 10.1073/pnas.1915778116
Sierra, A., Encinas, J. M., Deudero, J. J. P., Chancey, J. H., Enikolopov, G., Overstreet-Wadiche, L. S., et al. (2010). Microglia shape adult hippocampal neurogenesis through apoptosis-coupled phagocytosis. Cell Stem Cell 7, 483–495. doi: 10.1016/j.stem.2010.08.014
Sierra, A., and Tremblay, M. -È (2014). Microglia in Health and Disease, eds M. -È Tremblay and A. Sierra (New York, NY: Springer New York), 3–6. doi: 10.1007/978-1-4939-1429-6_1
Soeller, C., and Cannell, M. B. (1996). Construction of a two-photon microscope and optimisation of illumination pulse duration. Pflugers Arch. 432, 555–561. doi: 10.1007/s004240050169
Sofroniew, M. V. (2015). Astrocyte barriers to neurotoxic inflammation. Nat. Rev. Neurosci. 16, 249–263. doi: 10.1038/nrn3898
St-Pierre, M.-K., Šimončičová, E., Bögi, E., and Tremblay, M. -È (2020). Shedding light on the dark side of the microglia. ASN Neuro 12:1759091420925335. doi: 10.1177/1759091420925335
Stratoulias, V., Venero, J. L., Tremblay, M., and Joseph, B. (2019). Microglial subtypes: diversity within the microglial community. EMBO J. 38:e101997. doi: 10.15252/embj.2019101997
Sucksdorff, M., Tuisku, J., Matilainen, M., Vuorimaa, A., Smith, S., Keitilä, J., et al. (2019). Natalizumab treatment reduces microglial activation in the white matter of the MS brain. Neurol. Neuroimmunol. Neuroinflammation 6:e574. doi: 10.1212/NXI.0000000000000574
Susaki, E. A., Tainaka, K., Perrin, D., Kishino, F., Tawara, T., Watanabe, T. M., et al. (2014). Whole-brain imaging with single-cell resolution using chemical cocktails and computational analysis. Cell 157, 726–739. doi: 10.1016/j.cell.2014.03.042
Svoboda, K., and Yasuda, R. (2006). Principles of two-photon excitation microscopy and its applications to neuroscience. Neuron 50, 823–839. doi: 10.1016/j.neuron.2006.05.019
Tang, T., Valenzuela, A., Petit, F., Chow, S., Leung, K., Gorin, F., et al. (2018). In vivo MRI of functionalized iron oxide nanoparticles for brain inflammation. Contrast Media Mol. Imaging 2018:3476476. doi: 10.1155/2018/3476476
Tardif, C., Nadeau, G., Labrecque, S., Côté, D., Lavoie-Cardinal, F., and De Koninck, P. (2019). Fluorescence lifetime imaging nanoscopy for measuring Förster resonance energy transfer in cellular nanodomains. Neurophotonics 6:015002. doi: 10.1117/1.NPh.6.1.015002
Tay, T. L., Carrier, M., and Tremblay, M. -È (2019). Physiology of microglia. Adv. Exp. Med. Biol. 1175, 129–148. doi: 10.1007/978-981-13-9913-8_6
Tay, T. L., Savage, J. C., Hui, C. W., Bisht, K., and Tremblay, M. -È (2017). Microglia across the lifespan: from origin to function in brain development, plasticity and cognition. J. Physiol. 595, 1929–1945. doi: 10.1113/JP272134
Ter Veer, M. J. T., Pfeiffer, T., and Nägerl, U. V. (2017). Two-photon STED microscopy for nanoscale imaging of neural morphology in vivo. Methods Mol. Biol. Clifton NJ 1663, 45–64. doi: 10.1007/978-1-4939-7265-4_5
Theret, M., Mounier, R., and Rossi, F. (2019). The origins and non-canonical functions of macrophages in development and regeneration. Dev. Camb. Engl. 146:dev156000. doi: 10.1242/dev.156000
Tremblay, M. -È, Lowery, R. L., and Majewska, A. K. (2010). Microglial interactions with synapses are modulated by visual experience. PLoS Biol. 8:e1000527. doi: 10.1371/journal.pbio.1000527
Tremblay, M. -È, Paolicelli, R., Stevens, B., Wake, H., and Bessis, A. (2014). “Chapter 9. Developing and mature synapses,” in Microglia in Health and Disease, eds M. È Tremblay and A. Sierra (New York, NY: Springer).
Tremblay, M. -È, Stevens, B., Sierra, A., Wake, H., Bessis, A., and Nimmerjahn, A. (2011). The role of microglia in the healthy brain. J. Neurosci. Off. J. Soc. Neurosci 31, 16064–16069. doi: 10.1523/JNEUROSCI.4158-11.2011
Tronel, C., Largeau, B., Santiago Ribeiro, M. J., Guilloteau, D., Dupont, A.-C., and Arlicot, N. (2017). Molecular targets for PET imaging of activated microglia: the current situation and future expectations. Int. J. Mol. Sci. 18:802. doi: 10.3390/ijms18040802
Ueda, H. R., Ertürk, A., Chung, K., Gradinaru, V., Chédotal, A., Tomancak, P., et al. (2020). Tissue clearing and its applications in neuroscience. Nat. Rev. Neurosci. 21, 61–79. doi: 10.1038/s41583-019-0250-1
Van Dyken, P., and Lacoste, B. (2018). Impact of metabolic syndrome on neuroinflammation and the blood-brain barrier. Front. Neurosci. 12:930. doi: 10.3389/fnins.2018.00930
VanRyzin, J. W., Marquardt, A. E., Argue, K. J., Vecchiarelli, H. A., Ashton, S. E., Arambula, S. E., et al. (2019). Microglial phagocytosis of newborn cells is induced by endocannabinoids and sculpts sex differences in juvenile rat social play. Neuron 102, 435–449.e6. doi: 10.1016/j.neuron.2019.02.006
Varvel, N. H., Grathwohl, S. A., Baumann, F., Liebig, C., Bosch, A., Brawek, B., et al. (2012). Microglial repopulation model reveals a robust homeostatic process for replacing CNS myeloid cells. Proc. Natl. Acad. Sci. U.S.A. 109, 18150–18155. doi: 10.1073/pnas.1210150109
Varvel, N. H., Neher, J. J., Bosch, A., Wang, W., Ransohoff, R. M., Miller, R. J., et al. (2016). Infiltrating monocytes promote brain inflammation and exacerbate neuronal damage after status epilepticus. Proc. Natl. Acad. Sci. U.S.A. 113, E5665–E5674. doi: 10.1073/pnas.1604263113
Verkhratsky, A., Parpura, V., Vardjan, N., and Zorec, R. (2019). “Physiology of astroglia,” in Neuroglia in Neurodegenerative Diseases Advances in Experimental Medicine and Biology, eds A. Verkhratsky, M. S. Ho, R. Zorec, and V. Parpura (Singapore: Springer Singapore), 45–91. doi: 10.1007/978-981-13-9913-8_3
Villa, A., Gelosa, P., Castiglioni, L., Cimino, M., Rizzi, N., Pepe, G., et al. (2018). Sex-specific features of microglia from adult mice. Cell Rep. 23, 3501–3511. doi: 10.1016/j.celrep.2018.05.048
Villaseñor, R., and Collin, L. (2017). High-resolution confocal imaging of the blood-brain barrier: imaging, 3D reconstruction, and quantification of transcytosis. J. Vis. Exp. JoVE. doi: 10.3791/56407
Wang, F., Wan, H., Ma, Z., Zhong, Y., Sun, Q., Tian, Y., et al. (2019). Light-sheet microscopy in the near-infrared II window. Nat. Methods 16, 545–552. doi: 10.1038/s41592-019-0398-7
Wang, T., Ouzounov, D. G., Wu, C., Horton, N. G., Zhang, B., Wu, C.-H., et al. (2018). Three-photon imaging of mouse brain structure and function through the intact skull. Nat. Methods 15, 789–792. doi: 10.1038/s41592-018-0115-y
Weinhard, L., di Bartolomei, G., Bolasco, G., Machado, P., Schieber, N. L., Neniskyte, U., et al. (2018). Microglia remodel synapses by presynaptic trogocytosis and spine head filopodia induction. Nat. Commun. 9:1228. doi: 10.1038/s41467-018-03566-5
Westphal, V., Rizzoli, S. O., Lauterbach, M. A., Kamin, D., Jahn, R., and Hell, S. W. (2008). Video-rate far-field optical nanoscopy dissects synaptic vesicle movement. Science 320, 246–249. doi: 10.1126/science.1154228
Wohleb, E. S., McKim, D. B., Sheridan, J. F., and Godbout, J. P. (2015). Monocyte trafficking to the brain with stress and inflammation: a novel axis of immune-to-brain communication that influences mood and behavior. Front. Neurosci. 8:447. doi: 10.3389/fnins.2014.00447
Yamasaki, R., Lu, H., Butovsky, O., Ohno, N., Rietsch, A. M., Cialic, R., et al. (2014). Differential roles of microglia and monocytes in the inflamed central nervous system. J. Exp. Med. 211, 1533–1549. doi: 10.1084/jem.20132477
Yang, W., and Yuste, R. (2017). In vivo imaging of neural activity. Nat. Methods 14, 349–359. doi: 10.1038/nmeth.4230
Yi, S. Y., Barnett, B. R., Torres-Velázquez, M., Zhang, Y., Hurley, S. A., Rowley, P. A., et al. (2019). Detecting microglial density with quantitative multi-compartment diffusion MRI. Front. Neurosci. 13:81. doi: 10.3389/fnins.2019.00081
Zheng, Y., Chen, J., Shi, X., Zhu, X., Wang, J., Huang, L., et al. (2019). Two-photon focal modulation microscopy for high-resolution imaging in deep tissue. J. Biophotonics 12:e201800247. doi: 10.1002/jbio.201800247
Keywords: microscopy, microglia, fluorescence microscopy, electron microscopy, positron emission tomography, magnetic resonance imaging
Citation: Robert M-È, González Ibáñez F, Desjardins M and Tremblay M-È (2020) Imaging the Neuroimmune Dynamics Across Space and Time. Front. Neurosci. 14:903. doi: 10.3389/fnins.2020.00903
Received: 09 June 2020; Accepted: 04 August 2020;
Published: 23 September 2020.
Edited by:
João O. Malva, University of Coimbra, PortugalReviewed by:
Alejandra Sierra, University of Eastern Finland, FinlandRaquel Ferreira, University of Beira Interior, Portugal
Copyright © 2020 Carrier, Robert, González Ibáñez, Desjardins and Tremblay. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marie-Ève Tremblay, ZXZldHJlbWJsYXlAdXZpYy5jYQ==