 Bei Huang1†Djeda Belharazem2†Li Li2,3Susanne Kneitz1Philipp A. Schnabel4Ralf J. Rieker4Daniel Körner5
Bei Huang1†Djeda Belharazem2†Li Li2,3Susanne Kneitz1Philipp A. Schnabel4Ralf J. Rieker4Daniel Körner5 Wilfred Nix6Berthold Schalke7Hans Konrad Müller-Hermelink1German Ott8Andreas Rosenwald1Philipp Ströbel2,3
Wilfred Nix6Berthold Schalke7Hans Konrad Müller-Hermelink1German Ott8Andreas Rosenwald1Philipp Ströbel2,3 Alexander Marx2*
Alexander Marx2*- 1Pathologisches Institut der Universität Würzburg, Würzburg, Germany
- 2Pathologisches Institut und Zentrum für Medizinische Forschung (ZMF), Universitätsmedizin Mannheim der Universität Heidelberg, Mannheim, Germany
- 3Pathologisches Institut der Universitätsmedizin Göttingen, Göttingen, Germany
- 4Pathologisches Institut, Universität Heidelberg, Heidelberg, Germany
- 5Abteilung Thoraxchirurgie, Thoraxklinik Rohrbach, Universität Heidelberg, Heidelberg, Germany
- 6Neurologische Universitätsklinik Mainz, Mainz, Germany
- 7Neurologische Universitätsklink Regensburg, Regensburg, Germany
- 8Pathologisches Institut, Robert-Bosch-Krankenhaus, Stuttgart, Germany
The molecular pathogenesis of thymomas and thymic carcinomas (TCs) is poorly understood and results of adjuvant therapy are unsatisfactory in case of metastatic disease and tumor recurrence. For these clinical settings, novel therapeutic strategies are urgently needed. Recently, limited sequencing efforts revealed that a broad spectrum of genes that play key roles in various common cancers are rarely affected in thymomas and TCs, suggesting that other oncogenic principles might be important. This made us re-analyze historic expression data obtained in a spectrum of thymomas and thymic squamous cell carcinomas (TSCCs) with a custom-made cDNA microarray. By cluster analysis, different anti-apoptotic signatures were detected in type B3 thymoma and TSCC, including overexpression of BIRC3 in TSCCs. This was confirmed by qRT-PCR in the original and an independent validation set of tumors. In contrast to several other cancer cell lines, the BIRC3-positive TSCC cell line, 1889c showed spontaneous apoptosis after BIRC3 knock-down. Targeting apoptosis genes is worth testing as therapeutic principle in TSCC.
Introduction
Thymomas comprise a spectrum of unique thymic epithelial tumors that generally show intratumoral thymopoiesis. They are subdivided into WHO type A, AB, B1, B2, and B3 thymomas (1, 2), but this classification has been challenged by some authors (3, 4). Thymic carcinoma also show a spectrum subtypes that resemble analogously called extrathymic carcinomas (TCs) (2, 5). However, there is strong evidence (6–13) that thymic squamous cell carcinomas (TSCCs) and other squamous cell carcinomas are different entities. Nevertheless, treatments tailored to the unique biology of thymomas and TCs are missing (14–16) since there are almost no therapeutic targets (9–13, 17, 18), with few exceptions (e.g., Kit mutations) (12, 13). Even whole genome sequencing of a stage IVa B3 thymoma (19) and sequencing of 46 cancer genes in a TSCC (20) discovered no druggable mutations, suggesting that pathways other than in more common cancers might be operative (19). Considering this information and reports about the expression of apoptosis-related proteins in thymic tumors (21) we analyzed our unpublished gene expression data of thymuses and thymic tumors that were obtained with a custom-made cDNA microarray (representing 4606 genes). We found differential expression of anti-apoptotic genes in B3 thymomas and TSCC and could induce apoptosis by BIRC3 blockade in the thymic carcinoma cell line, 1889c (22).
Patient Characteristics and Methods
Patients and Tissues
Characteristics of the historic tumors (resected before 2004) stemmed from our data base (23) (Table 1). For the validation set of recent tumors obtained after 2006 see Table 2. Thymoma classification and staging followed the WHO and modified Masaoka system, respectively (2). All carcinomas were TSCCs without prior chemotherapy. “Combined thymomas” with >10% separable components were excluded. Thymocytes from a normal thymus were purified by Ficoll density gradient centrifugation. Ethical approval was obtained.
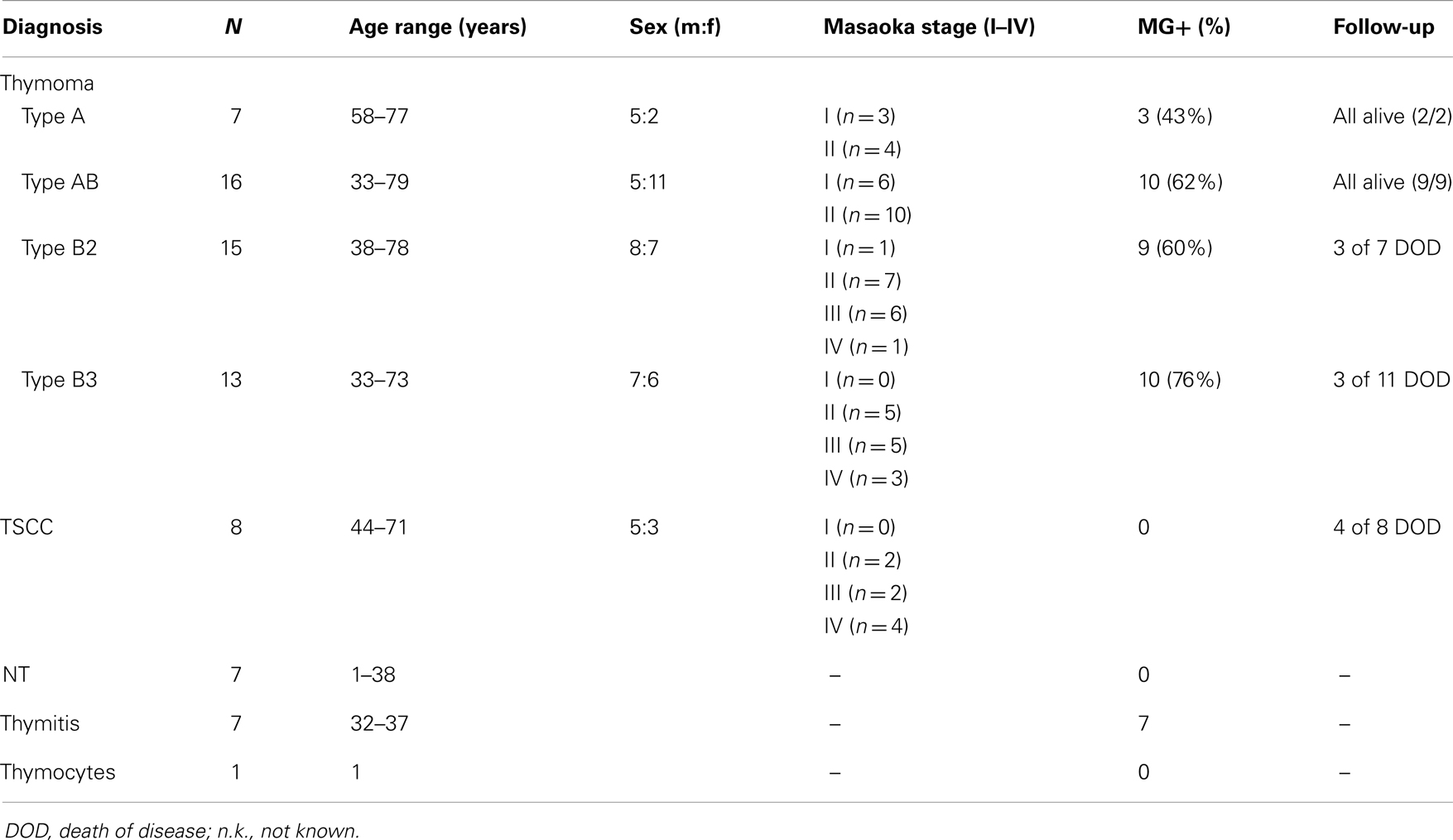
Table 1. Characteristics of patients and tissues: WHO type A, AB, B2, B3 thymomas; TSCC, thymic squamous cell carcinoma; NT, normal thymus; thymitis, non-neoplastic thymus with lymphofollicular hyperplasia (LFH) from early-onset Myasthenia Gravis (MG) patients; thymocytes, purified from NT; MG+ (%), %-age of patients with MG.
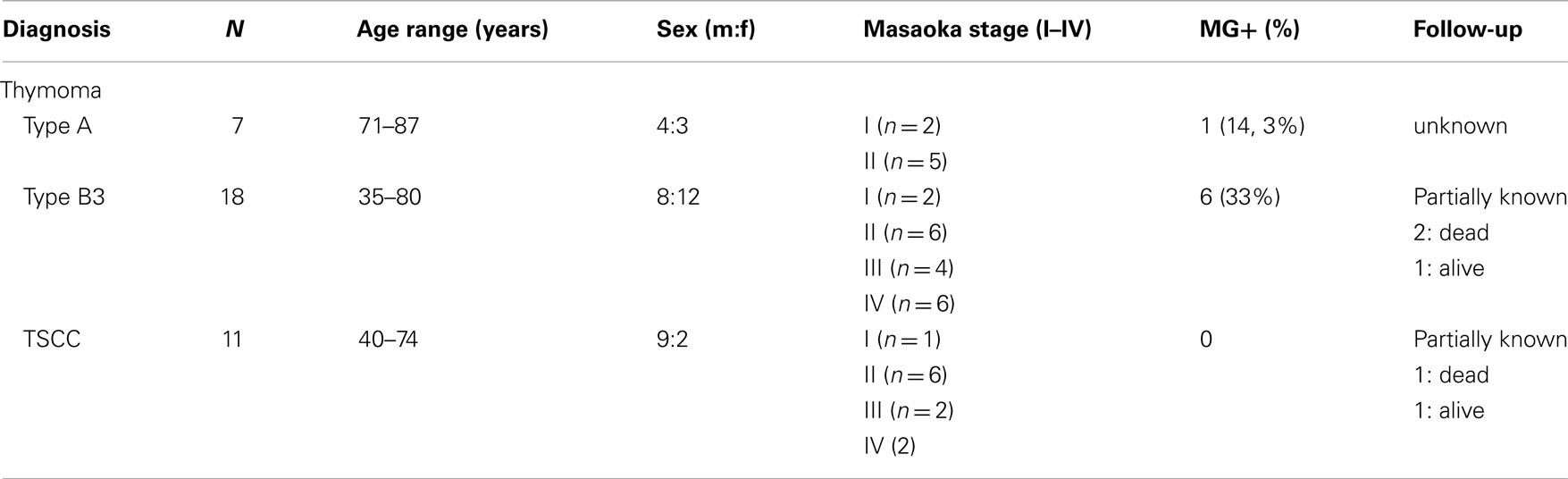
Table 2. Characteristics of 36 recent WHO type A and B3 thymomas and TSCCs that served as validation set for cases from Table 1.
Gene Expression Profiling
Gene expression profiling was achieved with a custom cDNA microarray representing 4606 genes with known relationship to cancers. To study reproducibility, 11 of 74 biopsies were studied in duplicate. Differential gene expression was analyzed by ANOVA (JMP Genomics, version 4; SAS, Cary, NC, USA).
RT-PCR
Array-based gene expression in the historic tumors was re-evaluated by qRT-PCR (TaqMan; FAST SYBR Green; Applied Biosystems) (Table S1 in Supplementary Material). Relative quantification was calculated using the ΔΔCt method with GAPDH as standard (Figure S2 in Supplementary Material). For primers and condition see Table S1 in Supplementary Material. Confirmed genes were checked by qRT-PCR in the validation set of 36 tumors.
Pathway Analysis
Pathways were studied using Gene Set Enrichment Analysis (GSEA) (24). Statistical significance (nominal p-value, NP) of enrichment scores (ES) was estimated using 1000 rounds of permutations per pathway. To adjust for multiple hypothesis testing, ES were normalized (“normalized enrichment scores,” NES) taking pathway size into account. The proportion of false positives was controlled by calculating the false discovery rate (FDR) for each NES.
Cell Lines and Transfection with si-BIRC3
BIRC3+ human cell lines studied here: 1889c (thymic carcinoma) (22); HaCat (keratinocytes); PC3 (prostate carcinoma); TE167 (rhabdomyosarcoma); A549 (lung carcinoma); MCF7 (breast carcinoma); HeLa (cervical adenocarcinoma). Culture conditions: RPMI (1889c) or DMEM (all others); 10% fetal bovine serum, 2 mM l-Glutamine and penicillin/streptomycin; 37°C; 5% CO2. About 105 cells transfected with 2 μM BIRC3 siRNA FlexTube: si00022827 (Qiagen, Hilden, Germany) or scramble siRNA using High-Perfect transfection reagent (Qiagen; fast forward transfection protocol) were harvested after 48 h.
Western Blot, Immunocytochemistry, and Apoptosis Detection
Cell lines and snap frozen tissues were used for Western blot (25). Methanol-fixed si-BIRC3 transfected cells and scramble siRNA transfected controls were stained with mouse anti-human cleaved caspase 3 (1:100), pH 6, Abcam for 1 h (25).
About 105 transfected cells including floating cells were harvested, stained at 20°C for 10 min with annexin V-FITC and Propidium Iodide PI (BD Biosciences), and evaluated for apoptosis by flow cytometry.
Results
Distinct Gene Expression Profiles Correlate with WHO-Defined Thymoma Subtypes
Gene expression profiling of eight cohorts, i.e., type A, AB, B2, B3 thymomas, TSCC, normal thymus, thymuses with thymitis, and thymocytes revealed 53 differentially expressed gens (p < 10−10). Based on these genes the cohorts were grouped into two major clusters: an epithelial-rich cluster with type A and B3 thymomas, and TSCCs, and a lymphocyte-rich cluster with AB, B2 thymomas, thymuses, and thymocytes (Figure 1). Clustering was closer between type A and B3 thymomas than between TSCC and B3 thymomas (Figure 1), despite the similar malignant potential of B3 thymomas and TSCC (22, 26, 27). Thymuses clustered closer to B2 than to AB thymomas, which may echo the more physiological thympoiesis in B2 thymomas (28, 29). GSEA (24)1 of genes over-expressed throughout the lymphocyte-rich cohorts showed that 13 out of 19 annotated genes (Frame in Figure 1) play a role in T cell biology (p < 10−12).
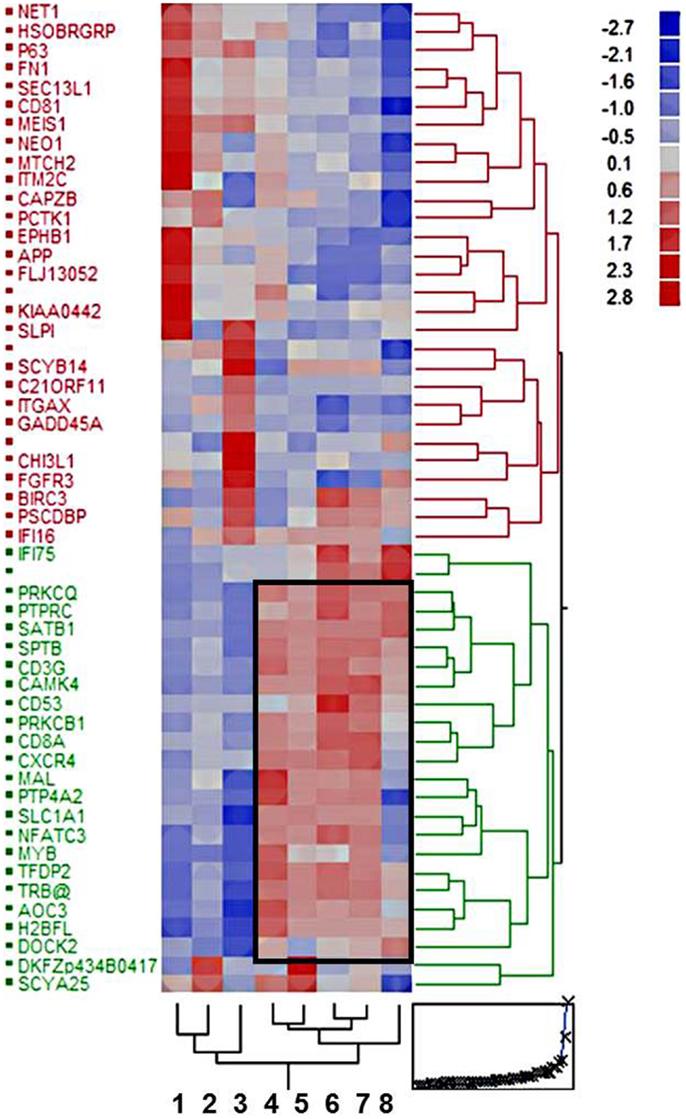
Figure 1. Hierarchical clustering of eight tissue cohorts: type A, AB, B2, and B3 thymomas; thymic squamous cell carcinomas (TSCCs); non-neoplastic thymuses (normal and MG-associated thymitis); and Ficoll gradient-purified thymocytes based on 53 differentially expressed genes (p < 10−10). The frame delineates 19 genes with coordinate up-regulation in the lymphocyte-rich cohorts, most of which (n = 13) play a role in lymphocyte biology (for details see text).
Functional Pathways Enriched in Epithelial-Rich Thymic Tumors
To reduce the impact of confounding thymocytes, we focused GSEA on type A and B3 thymomas and TSCCs. Using NES of >1.6 and p < 0.05 between at least two comparisons (24), we retrieved 11 functional pathways (Table 3). All three tumor cohorts were differentially enriched by all pair-wise comparisons for pathways comprising DNA damage response genes. Since enrichment increased from TSCCs to type A thymomas, this may reflect the highest genetic instability in TSCCs and lowest in type A thymomas (9, 10). Type A thymomas were enriched for three pathways: among the “cell migration genes,” over-expressed KAl1 has anti-metastatic function (30); IL8RB reinforces p53-dependent senescence (31); and SAA1 expression that has not been noted in thymomas before, can have pro- or anti-apoptotic functions in context-dependent manner (32–36). Among the “transcriptional corepressor genes” the tumor suppressor gene ID4 was over-expressed in type A thymomas; its reduction – as in type B3 thymomas and TSCCs – is typical of various cancers (37). These findings seemingly reflect the lower malignant potential of type A thymomas (23, 27).
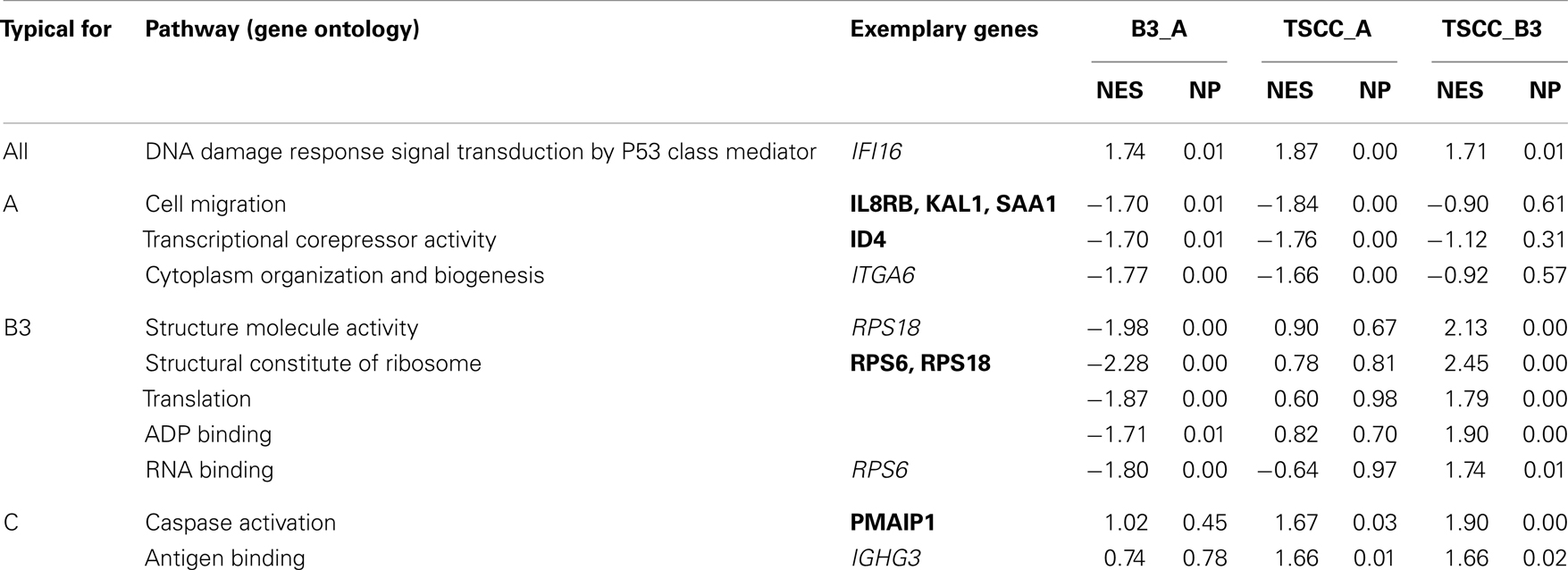
Table 3. Significantly enriched functional pathways in type A and B3 thymomas and TSCC using normalized enrichment scores (NES) of >1.6 and p < 0.05 between at least two comparisons; NP, normalized p-values (24). The “exemplary genes” are the genes with the strongest (>1.3-fold) significant (p < 0.05) differential expression in the indicated Gene Ontology pathway. Genes given in bold are discussed in the text.
In B3 thymomas, most of the enriched pathways concerned translation, including down-regulation of the ribosomal protein genes RPS6 and RPS18, which is rare in other cancers (38). Since RPS6 is an effector of the AKT/mTOR pathway, we studied expression of its negative regulators PTEN and PIK3R1 but found them up-regulated only slightly (p < 10−3) in B3 thymomas. Vice versa, up-regulation of RPS6 and down-regulation of PTEN and PIK3R1 in TSCCs is typical of aggressive cancers (39, 40).
For the role of caspase activity/apoptosis-related genes in TSCCs see next paragraph.
Apoptosis-Related Signature in TSCC
Nineteen genes were differentially expressed between type A and B3 thymomas and TSCC (>1.5-fold and p < 0.0001) and with low variation (var < 0.07) in at least one tumor subtype (Figure 2, Table S1 in Supplementary Material). By hierarchical clustering, type A thymomas and TSCCs formed distinct groups, while 6 out of 17 B3 thymomas were clustered with the type A thymomas and 11 with the TSCCs (Figure 2). The B3 thymoma subsets were not different with respect to age, sex, and tumor stage (not shown) but 9 of 10 myasthenia-associated B3 thymomas belonged to the “TSCC-like” subset. Figure 2 also depicts apoptosis-related genes that are co-up-regulated in TSCCs, two with anti-apoptotic (BIRC3; SCYA20) and two with pro-apoptotic function (PMAIP1; MYC). Importantly, the pro-apoptotic MTCH2 gene (41, 42) was down-regulated in TSCCs as also confirmed by qRT-PCR (Figure 3, left column; Table S1 in Supplementary Material) and validated in more recently obtained tumors (Figure 3, right column).
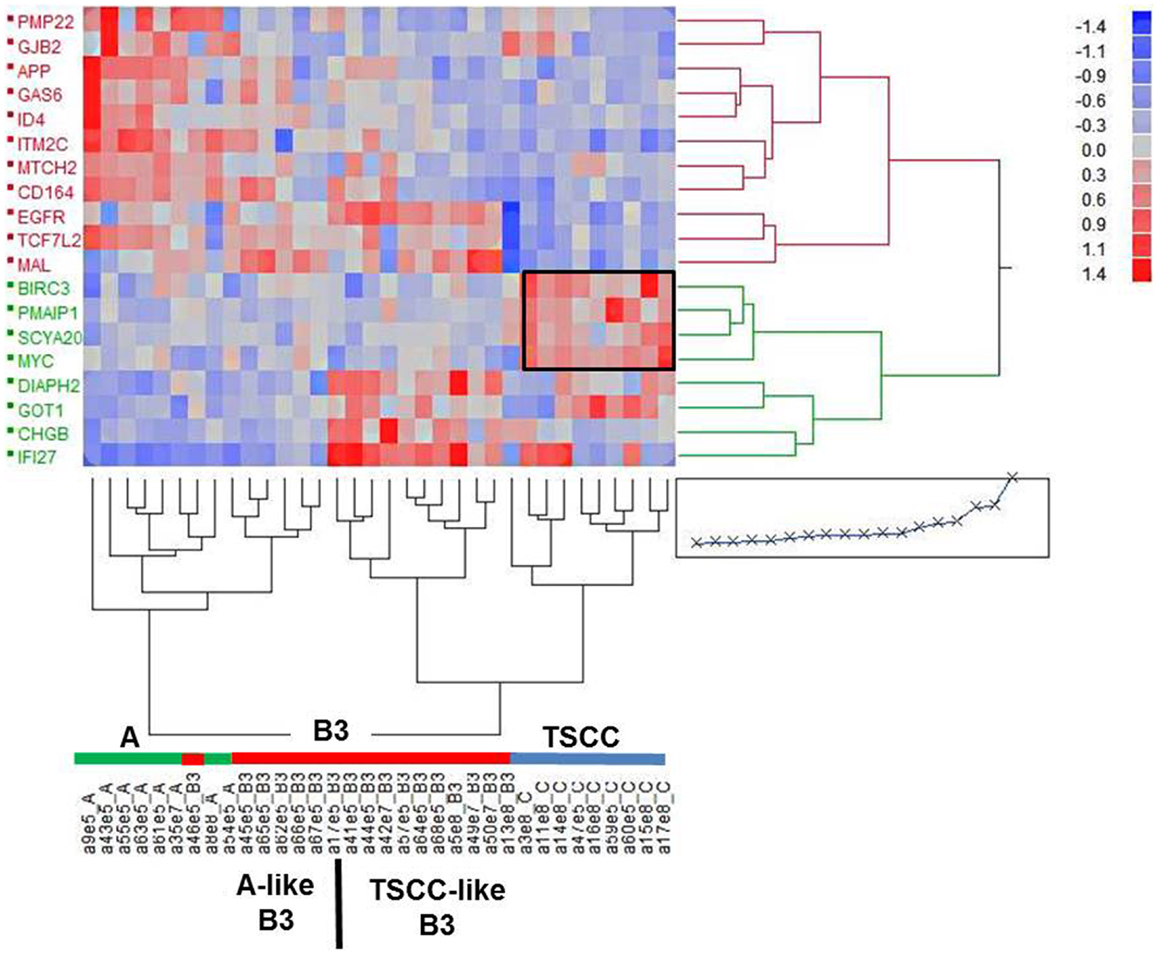
Figure 2. Cluster analysis of 19 expressed genes in individual epithelial-predominant type A and B3 thymomas and TSCC based on a fold-change >1.5, p < 10−4 and low variance (var < 0.07) in at least one of the cohorts of tumors. One of the two main clusters (left) harbors all of the type A thymomas and six of the (“type A-like”) B3 thymomas. The other cluster harbors all the TSCC and 11 of the (TSCC-like) B3 thymomas. The frame highlights a cluster of apoptosis-related genes (BIRC3, SCYA20, PMAIP1, MYC, MTCH2).
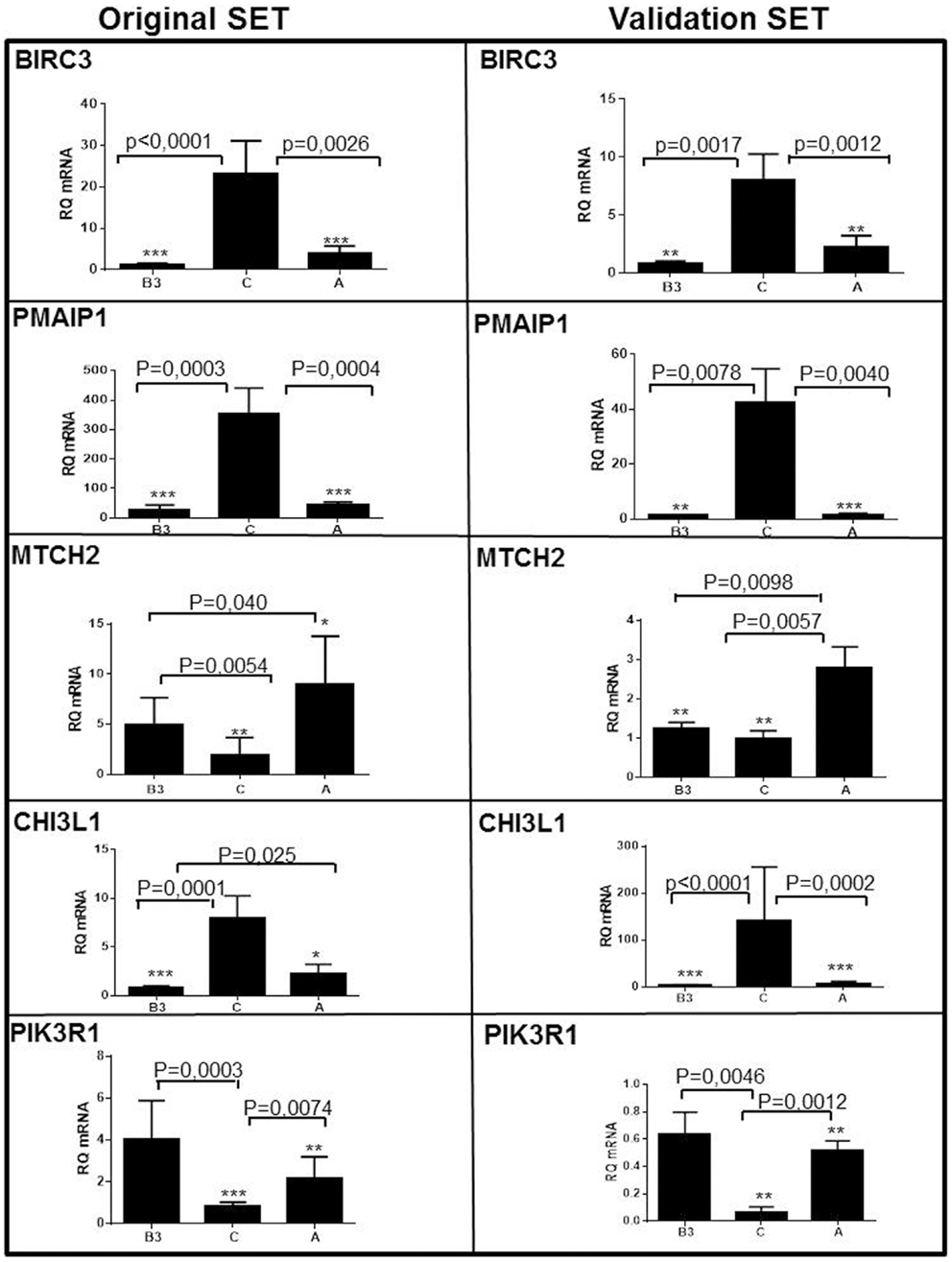
Figure 3. Validation of five differentially expressed genes in type A and type B3 thymomas and thymic squamous cell carcinomas (TSCC, C) in the original set of tumors (left column; see Table 1) and the validation set (right column; see Table 2). BIRC3, PMAIP1/NOXA, and Chitinase3-like1 genes are over-expressed in TSCC. The pro-apoptotic MTCH2 is expressed in type A thymomas (and largely missing in TSCC). PIK3R1 is most significantly expressed in type B3 thymomas.
Knock-Down of BIRC3 Induces Apoptosis in Thymic Carcinoma Cell Line 1889c
Among differentially expressed genes, we selected BIRC3 for functional analysis, because (i) it is a member of the inhibitors of apoptosis (IAPs) gene family (43) that serves key oncogenic roles in many cancers (44, 45); (ii) BIRC3 protein was over-expressed in parallel with mRNA in most TSCC biopsies compared to thymomas (Figure 4); and, (iii) the thymic carcinoma cell line, 1889c (22) with strong BIRC3 expression was available (Figure S2 in Supplementary Material). siRNA-mediated BIRC3 knock-down (Figures 5A,B) induced spontaneous apoptosis in 50% of 1889c cells within 48 h as shown by cytology, expression of activated caspase 3 and annexin 5 (Figures 5C–E). 1889c cells and HaCat keratinocytes were more sensitive toward BIRC3 knock-down than all other cancer cell lines tested (Figures 5A–E; Figure S3 in Supplementary Material), despite similar BIRC3 mRNA levels (Figure S2 in Supplementary Material).
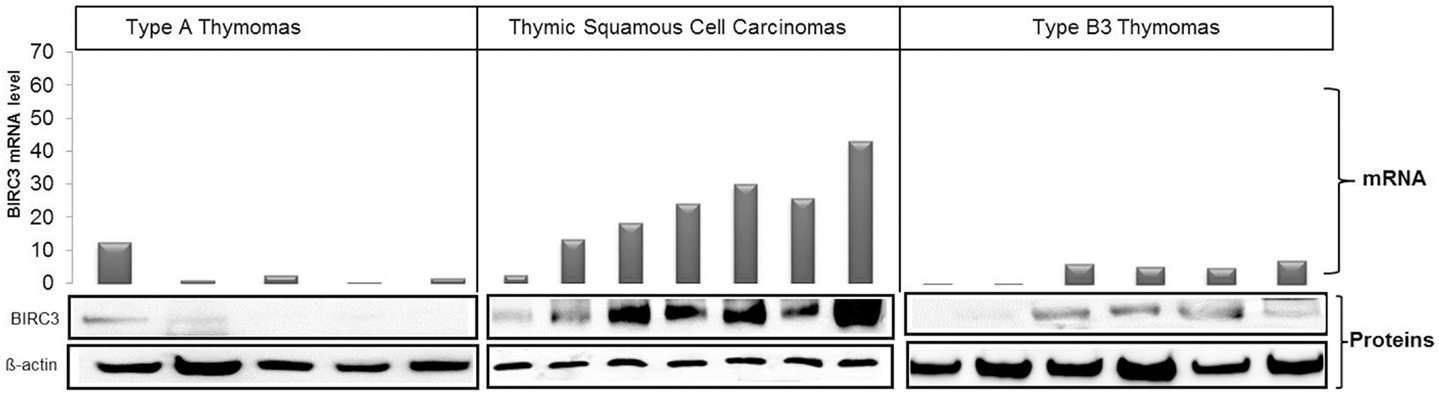
Figure 4. Correlation between BIRC3 mRNA and protein levels in type A (five cases) and type B3 (six cases) thymomas and TSCC (seven cases). Levels of mRNA and protein were higher in TSCCs than in thymomas. mRNA was quantified using real time PCR with GPDH as reference, β-actin was used as loading control in western blots.
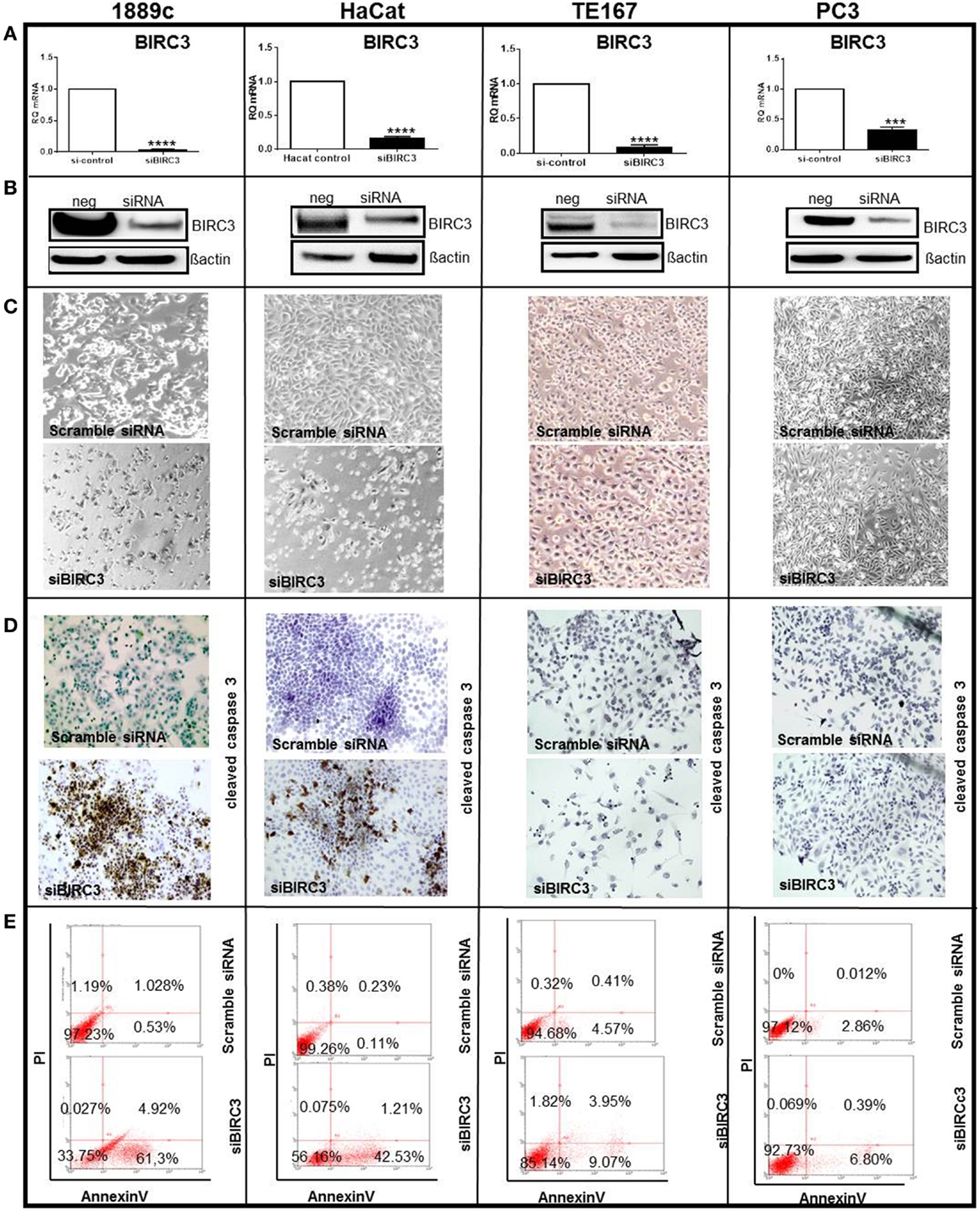
Figure 5. Several cell lines (1889c, HaCat, TE167, and PC3) were transfected using si-BIRC3 for 48h, which significantly repressed expression of (A) Birc3 mRNA ****p < 0.0001 in 1889c, HaCat and TE167 cells and ***p = 0.0002 in PC3) and (B) Birc3 protein. After transfection. Changes of the cytology of transfected cells is shown in (C). Cells were stained with an antibody to cleaved caspase 3 for apoptosis detection (immunoperoxidase ×20) (D). FACS analysis to quantify apoptosis using Annexin V-FITCS/PI (E). All experiments were done in triplicate and each measurement in duplicate. Similar low levels of apoptosis following si-BIRC3 transfection were observed in A549 (lung carcinoma); MCF7 (breast carcinoma); and HeLa (cervical adenocarcinoma) cells.
Discussion
The major finding here is the so far unreported (13, 19, 46) observation of different expression profiles of apoptosis-related genes in TSCCs and B3 thymomas in two independent sets of thymic epithelial tumors. Significantly, anti-apoptotic BIRC3 – a member of the “IAPs” gene family that mainly blocks the extrinsic apoptosis pathway – was over-expressed at the mRNA and protein level in most TSCCs compared to type A and B3 thymomas; and BIRC3-positive 1889c thymic carcinoma cells showed stronger spontaneous apoptosis on BIRC3 knock-down in vitro than all other cell lines tested. In addition, other than the thymomas, the TSCCs showed lower mRNA expression of the MTCH2 gene that induces apoptosis by cooperation with tBID to facilitate BAX-mediated mitochondrial cytochrome c release (47). These findings suggest that TSCCs suffer from attenuation of both the extrinsic and intrinsic apoptosis pathway. This is complemented in TSCCs by down-regulation of PIK3R1 (Figure 3) that normally interferes with various pro-survival cascades, including Akt signaling (40, 41). By contrast, the anti-apoptotic make-up of B3 thymomas in the current study was due to the down-regulation of the pro-apoptotic PMAIP1/NOXA gene (Figure 3) that plays a role in the mitochondrial apoptosis pathway and drug resistance (48).
A caveat here is the limited coverage of the transcriptome by our applied microarray. While key apoptosis-related gens like BCL2, MCL1, BIRC2, BIRC5, BCL2L1 were represented but did not show differential expression between TSCC and B3 thymomas (not shown), other important genes (e.g., BIRC4/XIAP, BIRC6-8, BAX, BBC3/PUMA, and BIM) were missing and warrant further analysis. Despite these limitations, our findings could have a translational perspective, since advanced tumor stage, limited resectability, and common relapses (13–15, 23, 26, 49–51) are frequent indications in TSCCs for (neo-)adjuvant therapies that may be critically attenuated by apoptosis resistance of the target tumor cells (52). Indeed, therapeutic options are already available (e.g., #NCT00977067; #NCT01078649)2 or up-coming (53), including drugs against BIRC3 (54) and strategies to induce PMAIP1 (48). RT-PCR-based expression profiles of anti-apoptotic genes are worth testing as “biomarkers” in clinical trials aiming to break treatment resistance in thymic cancers.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Deutsche Krebshilfe (Alexander Marx; Hans Konrad Müller-Hermelink; Andreas Rosenwald; Philipp Ströbel; Project 10-1740) and BMBF (Philipp Ströbel; Alexander Marx, Project 01DL12027).
Supplementary Material
The Supplementary Material for this article can be found online at http://www.frontiersin.org/Journal/10.3389/fonc.2013.00316/abstract
Table S1. Characteristics of the 19 genes with significantly different expression in epithelial-rich thymic epithelial tumors (Figure 2): type A and B3 thymomas and TSCCs. Functions are: AD, adhesion; AP, apoptosis; Diff, differentiation; IR, immune response; M, migration; P, proliferation; TD, T cell development; IF, inflammation; TR, tissue remodeling. The differential expression of all genes in the original set of tumors was confirmed by qRT-PCR (see also Figure 3).
Table S2. Primer sequences and annealing temperatures used to confirm and validate by qRT-PCR the expression of those genes that were found to be differentially expressed on microarray analysis and are depicted in Figure 3 and Figure S1 in Supplementary Material.
Figure S1. Confirmation and validation of cKIT gene expression using qRT-PCR in TSCC.
Figure S2. Semi-quantitative determination of BIRC3 mRNA levels in several cell lines using standard PCR with 10 ng cDNA as template for each sample. BIRC3 expression was detectable in all cell lines except the rhabdomyosarcoma cell lines RH30 and CRL2061. GAPDH was used as control.
Figure S3. Evaluation of annexin V-FITC/PI labeled cells using FACS 48 h after BIRC3 knock-down. The transfected 1889c thymic carcinoma cells showed the highest level of apoptosis (50%) compared to HaCat (40%), TE167 (10%, **p = 0.0097), A459 (20%, *p = 0.0432), PC3 (<10%, **p = 0.0082), MCF7 (<20%, *p = 0.0154), and HeLa (<10%, **p = 0.0079). The results represent the mean of three independent experiments, each with duplicate measurements.
Footnotes
References
2. Müller-Hermelink HK, Möller P, Engel P, Menestrina F, Kuo TT, Shimosato Y, et al. Tumours of the thymus. In: Travis WD, Brambilla E, Müller-Hermelink HK, Harris CC, editors. Tumours of the Lung, Thymus, and Heart Pathology and Genetics. Lyon: IARC Press (2004). p. 145–51.
3. Marchevsky AM, Gupta R, McKenna RJ, Wick M, Moran C, Zakowski MF, et al. Evidence-based pathology and the pathologic evaluation of thymomas: the World Health Organization classification can be simplified into only 3 categories other than thymic carcinoma. Cancer (2008) 112:2780–8. doi: 10.1002/cncr.23492
4. Verghese ET, den Bakker MA, Campbell A, Hussein A, Nicholson AG, Rice A, et al. Interobserver variation in the classification of thymic tumours – a multicentre study using the WHO classification system. Histopathology (2008) 53:218–23. doi:10.1111/j.1365-2559.2008.03088.x
5. Suster S, Rosai J. Thymic carcinoma. A clinicopathologic study of 60 cases. Cancer (1991) 67:1025–32. doi:10.1002/1097-0142(19910215)67:43.0.CO;2-F
6. Hishima T, Fukayama M, Fujisawa M, Hayashi Y, Arai K, Funata N, et al. CD5 expression in thymic carcinoma. Am J Pathol (1994) 145:268–75.
7. Henley JD, Cummings OW, Loehrer PJ Sr. Tyrosine kinase receptor expression in thymomas. J Cancer Res Clin Oncol (2004) 130:222–4. doi:10.1007/s00432-004-0545-z
8. Pan CC, Chen PC, Chiang H. KIT (CD117) is frequently overexpressed in thymic carcinomas but is absent in thymomas. J Pathol (2004) 202:375–81. doi:10.1002/path.1514
9. Zettl A, Ströbel P, Wagner K, Katzenberger T, Ott G, Rosenwald A, et al. Recurrent genetic aberrations in thymoma and thymic carcinoma. Am J Pathol (2000) 157:257–66. doi:10.1016/S0002-9440(10)64536-1
10. Inoue M, Starostik P, Zettl A, Ströbel P, Schwarz S, Scaravilli F, et al. Correlating genetic aberrations with WHO-defined histology and stage across the spectrum of thymomas. Cancer Res (2003) 63:3708–15.
11. Penzel R, Hoegel J, Schmitz W, Blaeker H, Morresi-Hauf A, Aulmann S, et al. Clusters of chromosomal imbalances in thymic epithelial tumours are associated with the WHO classification and the staging system according to Masaoka. Int J Cancer (2003) 105:494–8. doi:10.1002/ijc.11101
12. Ströbel P, Hartmann M, Jakob A, Mikesch K, Brink I, Dirnhofer S, et al. Thymic carcinoma with over-expression of mutated KIT: response to imatinib. N Engl J Med (2004) 350:2625–6. doi:10.1056/NEJM200406173502523
13. Girard N, Shen R, Guo T, Zakowski MF, Heguy A, Riely GJ, et al. Comprehensive genomic analysis reveals clinically relevant molecular distinctions between thymic carcinomas and thymomas. Clin Cancer Res (2009) 15:6790–9. doi:10.1158/1078-0432.CCR-09-0644
14. Rajan A, Giaccone G. Chemotherapy for thymic tumors: induction, consolidation, palliation. Thorac Surg Clin (2011) 21:107–14. doi:10.1016/j.thorsurg.2010.08.003
15. Kelly RJ, Petrini I, Rajan A, Wang Y, GiacconeG J. Thymic malignancies: from clinical management to targeted therapies. Clin Oncol (2011) 29:4820–7. doi:10.1200/JCO.2011.36.0487
16. Girard N. Chemotherapy and targeted agents for thymic malignancies. Expert Rev Anticancer Ther (2012) 12:685–95. doi:10.1586/era.12.29
17. Sasaki H, Ide N, Fukai I, Kiriyama M, Yamakawa Y, Fujii Y. Gene expression analysis of human thymoma correlates with tumor stage. Int J Cancer (2002) 101:342–7. doi:10.1002/ijc.10624
18. Sasaki H, Ide N, Sendo F, Takeda Y, Adachi M, Fukai I, et al. Glycosylphosphatidylinositol-anchored protein (GPI-80) gene expression is correlated with human thymoma stage. Cancer Sci (2003) 94:809–13. doi:10.1111/j.1349-7006.2003.tb01523.x
19. Petrini I, Rajan A, Pham T, Voeller D, Davis S, Gao J, et al. Whole genome and transcriptome sequencing of a b3 thymoma. PLoS One (2013) 8(4):e60572. doi:10.1371/journal.pone.0060572
20. Hu Z, Wang J, Yao T, Hong RL, Zhang K, Gao H, et al. Identification of novel mutations of TP53, ALK and RET gene in metastatic thymic squamous cell carcinoma and its therapeutic implication. Lung Cancer (2013) 81:27–31. doi:10.1016/j.lungcan.2013.04.006
21. Pan CC, Chen PC, Wang LS, Lee JY, Chiang H. Expression of apoptosis-related markers and HER-2/neu in thymic epithelial tumours. Histopathology (2003) 43:165–72. doi:10.1046/j.1365-2559.2003.01663.x
22. Ehemann V, Kern MA, Breinig M, Schnabel PA, Gunawan B, Schulten HJ, et al. Establishment, characterization and drug sensitivity testing in primary cultures of human thymoma and thymic carcinoma. Int J Cancer (2008) 122(12):2719–25. doi:10.1002/ijc.23335
23. Ströbel P, Bauer A, Puppe B, Kraushaar T, Krein A, Toyka K, et al. Tumor recurrence and survival in patients treated for thymomas and thymic squamous cell carcinomas: a retrospective analysis. J Clin Oncol (2004) 22:1501–9. doi:10.1200/JCO.2004.10.113
24. Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A (2005) 102:15545–50. doi:10.1073/pnas.0506580102
25. Simon-Keller K, Paschen A, Hombach AA, Ströbel P, Coindre JM, Eichmüller SB, et al. Survivin blockade sensitizes rhabdomyosarcoma cells for lysis by fetal acetylcholine receptor-redirected T cells. Am J Pathol (2013) 182(6):2121–31. doi:10.1016/j.ajpath.2013.02.017
26. Chen G, Marx A, Chen WH, Yong J, Puppe B, Stroebel P, et al. New WHO histologic classification predicts prognosis of thymic epithelial tumors: a clinicopathologic study of 200 thymoma cases from China. Cancer (2002) 95:420–9. doi:10.1002/cncr.10665
27. Jain RK, Mehta RJ, Henley JD, Kesler KA, Loehrer PJ, Badve S. WHO types A and AB thymomas: not always benign. Mod Pathol (2010) 23:1641–9. doi:10.1038/modpathol.2010.172
28. Nenninger R, Schultz A, Hoffacker V, Helmreich M, Wilisch A, Vandekerckhove B, et al. Abnormal thymocyte development and generation of autoreactive T cells in mixed and cortical thymomas. Lab Invest (1998) 78:743–53.
29. Ströbel P, Helmreich M, Menioudakis G, Lewin SR, Rüdiger T, Bauer A, et al. Paraneoplastic myasthenia gravis correlates with generation of mature naive CD4+ T cells in thymomas. Blood (2002) 100:159–66. doi:10.1182/blood.V100.1.159
30. Liu FS, Chen JT, Dong JT, Hsieh YT, Lin AJ, Ho ES, et al. KAI1 metastasis suppressor gene is frequently down-regulated in cervical carcinoma. Am J Pathol (2001) 159:1629–34. doi:10.1016/S0002-9440(10)63009-X
31. Acosta JC, O’Loghlen A, Banito A, Guijarro MV, Augert A, Raguz S, et al. Chemokine signaling via the CXCR2 receptor reinforces senescence. Cell (2008) 133:1006–18. doi:10.1016/j.cell.2008.03.038
32. Gutfeld O, Prus D, Ackerman Z, Dishon S, Linke RP, Levin M, et al. Expression of serum amyloid A, in normal, dysplastic, and neoplastic human colonic mucosa: implication for a role in colonic tumorigenesis. J Histochem Cytochem (2006) 54:63–73. doi:10.1369/jhc.5A6645.2005
33. Shinriki S, Ueda M, Ota K, Nakamura M, Kudo M, Ibusuki M, et al. Aberrant expression of serum amyloid A in head and neck squamous cell carcinoma. J Oral Pathol Med (2010) 39:41–7. doi:10.1111/j.1600-0714.2009.00777.x
34. Knebel FH, Albuquerque RC, Massaro RR, Maria-Engler SS, Campa A. Dual effect of serum amyloid A on the invasiveness of glioma cells. Mediators Inflamm (2013) 2013:509089. doi:10.1155/2013/509089
35. Kho Y, Kim S, Yoon BS, Moon JH, Kim B, Kwak S, et al. Induction of serum amyloid A genes is associated with growth and apoptosis of HC11 mammary epithelial cells. Biosci Biotechnol Biochem (2008) 72(1):70–81. doi:10.1271/bbb.70374
36. Yoon S, Woo SU, Kang JH, Kim K, Shin HJ, Gwak HS, et al. NF-κB and STAT3 cooperatively induce IL6 in starved cancer cells. Oncogene (2012) 31(29):3467–81. doi:10.1038/onc.2011.517
37. Umetani N, Takeuchi H, Fujimoto A, Shinozaki M, Bilchik AJ, Hoon DS. Epigenetic inactivation of ID4 in colorectal carcinomas correlates with poor differentiation and unfavorable prognosis. Clin Cancer Res (2004) 10:7475–83. doi:10.1158/1078-0432.CCR-04-0689
38. Hagner PR, Mazan-Mamczarz K, Dai B, Balzer EM, Corl S, Martin SS, et al. Ribosomal protein S6 is highly expressed in non-Hodgkin lymphoma and associates with mRNA containing a 5’ terminal oligopyrimidine tract. Oncogene (2011) 30:1531–41. doi:10.1038/onc.2010.533
39. Urick ME, Rudd ML, Godwin AK, Sgroi D, Merino M, Bell DW. PIK3R1 (p85{alpha}) is somatically mutated at high frequency in primary endometrial cancer. Cancer Res (2011) 71:4061–7. doi:10.1158/0008-5472.CAN-11-0549
40. Taniguchi CM, Winnay J, Kondo T, Bronson RT, Guimaraes AR, Alemán JO, et al. The phosphoinositide 3-kinase regulatory subunit p85alpha can exert tumor suppressor properties through negative regulation of growth factor signaling. Cancer Res (2010) 70:5305–15. doi:10.1158/0008-5472.CAN-09-3399
41. Yu K, Ganesan K, Tan LK, Laban M, Wu J, Zhao XD, et al. A precisely regulated gene expression cassette potently modulates metastasis and survival in multiple solid cancers. PLoS Genet (2008) 4:e1000129. doi:10.1371/journal.pgen.1000129
42. Zaltsman Y, Shachnai L, Yivgi-Ohana N, Schwarz M, Maryanovich M, Houtkooper RH, et al. MTCH2/MIMP is a major facilitator of tBID recruitment to mitochondria. Nat Cell Biol (2010) 12:553–62. doi:10.1038/ncb2057
43. Wu H, Tschopp J, Lin SC. Smac mimetics and TNFalpha: a dangerous liaison? Cell (2007) 131(4):655–8. doi:10.1016/j.cell.2007.10.042
44. Darding M, Feltham R, Tenev T, Bianchi K, Benetatos C, Silke J, et al. Molecular determinants of Smac mimetic induced degradation of cIAP1 and cIAP2. Cell Death Differ (2011) 18(8):1376–86. doi:10.1038/cdd.2011.10
45. Miura K, Karasawa H, Sasaki I. IAP2 as a therapeutic target in colorectal cancer and other malignancies. Expert Opin Ther Targets (2009) 13(11):1333–45. doi:10.1517/14728220903277256
46. Badve S, Goswami C, Gökmen-Polar Y, Nelson RP Jr, Henley J, Miller N, et al. Molecular analysis of thymoma. PLoS One (2012) 7(8):e42669. doi:10.1371/journal.pone.0042669
47. Katz C, Zaltsman-Amir Y, Mostizky Y, Kollet N, Gross A, Friedler A. Molecular basis of the interaction between proapoptotic truncated BID (tBID) protein and mitochondrial carrier homologue 2 (MTCH2) protein: key players in mitochondrial death pathway. J Biol Chem (2012) 287(18):15016–23. doi:10.1074/jbc.M111.328377
48. Zhang LN, Li JY, Xu W. A review of the role of Puma, Noxa and Bim in the tumorigenesis, therapy and drug resistance of chronic lymphocytic leukemia. Cancer Gene Ther (2013) 20:1–7. doi:10.1038/cgt.2012.84
49. Marulli G, Lucchi M, Margaritora S, Cardillo G, Mussi A, Cusumano G, et al. Surgical treatment of stage III thymic tumors: a multi-institutional review from four Italian centers. Eur J Cardiothorac Surg (2011) 39(3):e1–7. doi:10.1016/j.ejcts.2010.11.026
50. Rena O, Mineo TC, Casadio C. Multimodal treatment for stage IVA thymoma: a proposable strategy. Lung Cancer (2012) 76(1):89–92. doi:10.1016/j.lungcan.2011.10.004
51. Okereke IC, Kesler KA, Freeman RK, Rieger KM, Birdas TJ, Ascioti AJ, et al. Thymic carcinoma: outcomes after surgical resection. Ann Thorac Surg (2012) 93(5):1668–72. doi:10.1016/j.athoracsur.2012.01.014
52. Reed JC. Apoptosis mechanisms: implications for cancer drug discovery. Oncology (Williston Park) (2004) 18(13 Suppl 10):11–20.
53. Roberts D, Clinthorne G, Vasilevskaya I, Johnson SW, O’Dwyer PJ. Molecular pharmacology of a novel set of IAP antagonists. ASCO Poster, C87 (2010). Philadelphia, PA.
Keywords: thymoma, thymic carcinoma, thymus, apoptosis, gene expression, myasthenia gravis, MTCH2, targeted therapy
Citation: Huang B, Belharazem D, Li L, Kneitz S, Schnabel PA, Rieker RJ, Körner D, Nix W, Schalke B, Müller-Hermelink HK, Ott G, Rosenwald A, Ströbel P and Marx A (2013) Anti-apoptotic signature in thymic squamous cell carcinomas – functional relevance of anti-apoptotic BIRC3 expression in the thymic carcinoma cell line 1889c. Front. Oncol. 3:316. doi: 10.3389/fonc.2013.00316
Received: 07 October 2013; Accepted: 11 December 2013;
Published online: 31 December 2013.
Edited by:
Giuseppe Giaccone, Georgetown University, USAReviewed by:
Lorenzo Spaggiari, European Institute of Oncology, ItalyShahab Babakoohi, Medstar Good Samaritan Hospital, USA
Copyright: © 2013 Huang, Belharazem, Li, Kneitz, Schnabel, Rieker, Körner, Nix, Schalke, Müller-Hermelink, Ott, Rosenwald, Ströbel and Marx. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexander Marx, Pathologisches Institut, Universitätsklinikum Mannheim, Theodor-Kutzer-Ufer 1-3, Mannheim D-68135, Germany e-mail:YWxleGFuZGVyLm1hcnhAdW1tLmRl
†Shared first authorship