Abstract
Tumor microenvironment is considered nowadays as one of the main players in cancer development and progression. Tumor microenvironment is highly complex and consists of non-tumor cells (i.e., cancer-associated fibroblast, endothelial cells, or infiltrating leukocytes) and a large list of extracellular matrix proteins and soluble factors. The way that microenvironment components interact among them and with the tumor cells is very complex and only partially understood. However, it is now clear that these interactions govern and modulate many of the cancer hallmarks such as cell proliferation, the resistance to death, the differentiation state of tumor cells, their ability to migrate and metastasize, and the immune response against tumor cells. One of the microenvironment components that have emerged in the last years with strength is a heterogeneous group of multifaceted proteins grouped under the name of matricellular proteins. Matricellular proteins are a family of non-structural matrix proteins that regulate a variety of biological processes in normal and pathological situations. Many components of this family such as periostin (POSTN), osteopontin (SPP1), or the CNN family of proteins have been shown to regulate key aspect of tumor biology, including proliferation, invasion, matrix remodeling, and dissemination to pre-metastatic niches in distant organs. Matricellular proteins can be produced by tumor cells themselves or by tumor-associated cells, and their synthesis can be affected by intrinsic and/or extrinsic tumor cell factors. In this review, we will focus on the role of POSTN in the development and progression of cancer. We will describe their functions in normal tissues and the mechanisms involved in their regulation. We will analyze the tumors in which their expression is altered and their usefulness as a biomarker of tumor progression. Finally, we will speculate about future directions for research and therapeutic approaches targeting POSTN.
Introduction
Hanahan and Weinberg published in the year 2000 an article titled “The hallmarks of cancers” where they summarized by the first time the list of biological capabilities that should be acquired during the development of cancer. The list of such hallmarks included six fundamental biological functions: sustained proliferation signaling, evasion of growth suppression, resistance to cell death, replicative immortality, induction of angiogenesis, and activation of invasion and metastasis (1). Eleven years later, they revisited the concept of cancer hallmarks and included two new enabling characteristics, namely genome instability and mutation and tumor-promoting inflammation, and two emerging hallmarks, namely deregulation of cellular energetics and evasion of immune system (2). In addition, it was also anticipated the important role of tumor microenvironment in the acquisition and modulation of the hallmarks of cancers.
In addition to genetic and epigenetic alterations that occur in cancer cells, and which are in the origin of the hallmarks of cancer, in the last years, there have been accumulated numerous evidences supporting a main role of tumor microenvironment in the modulation, and even induction, of cancer hallmarks (3). As a proof of the interest generated by tumor microenvironment as a key player in cancer development, articles addressing this topic have increased a 400% from year 2000.
Tumor microenvironment can be defined as everything that surrounds tumor cells that are not the tumor cells themselves. Thus, tumor microenvironment is composed of stroma cells (for example, cancer-associated fibroblasts), immune cells that infiltrate the tumor, extracellular matrix (ECM) proteins, and soluble components such as hormones, growth factor, or cytokines. Tumor cells are able to alter the microenvironment and change the properties of the host tissue and vice versa, the composition of the tumor microenvironment influences the behavior of cancer cells (4). In consequence, these cross-talks between cancer cells and the different components of the tumor microenvironment are nowadays considered an essential player in the process that conducts to transformation and cancer.
As mentioned above, one of the main components of the tumor microenvironment is the extracellular cell matrix, which is composed by structural (e.g., collagens) and non-structural proteins that have diverse functions. Within the group of non-structural proteins, there is a subgroup of heterogeneous proteins particularly interesting, named matricellular proteins. The term matricellular proteins was initially proposed by Bornstein et al. (5, 6) to define a group of non-structural ECM proteins that exert several functions by binding to other ECM proteins, growth factor, and cytokines and cells through cell specific receptors in the cell membrane. Matricellular proteins play a central role in the homeostasis of normal tissues regulating cell proliferation and differentiation. These proteins are generally expressed at low levels in most adult tissues, but are highly expressed during inflammation, tissue repair, wound healing, and malignant transformation. Some of the proteins that belong this group of proteins are osteopontin, thrombospondins, members of the CCN family (Cyr61, CCN2, CCN3), tenascins, SPARC, fibulins, or periostin (POSTN) (7).
In this article, we will review the role of the matricellular protein POSTN in cancer development and progression in light of the hallmarks of cancer. First, we will review briefly its molecular characteristics and its functions in normal tissue homeostasis. Next, we described that we know about POSTN expression in solid tumors and their usefulness as a prognostic biomarker. As we will see in detail, POSTN expression is frequently overexpressed by the stromal component of solid tumors and is frequently associated to poor prognosis and metastasis in many cancers. Next, we will review the studies analyzing the effect of POSTN on tumor cells. POSTN interacts with tumor cells through integrin receptors and elicits a plethora of signaling pathways in tumors cells related to cell proliferation, cell survival, or cell migration. Finally, we will speculate about future directions for research and therapeutic approaches targeting POSTN.
POSTN Structure
Periostin was identified in 1993 as a putative cell adhesion protein produced by a mouse osteoblastic cell line (8). The periostin gene (POSTN) in human is located in the long arm of chromosomal 13 (13q13.3) (9) and spans approximately 36 kb. It has 23 exons and encodes for a protein of 836 amino acids with a molecular weight of 93 kDa (8, 10). Alternative splicing of the C-terminal sequence including exons 17–21 gives rise to four POSTN isoforms ranging from 83 to 93 kDa (11). These isoforms have been characterized from different tissues (9, 12–14) and have been shown to be differentially expressed during embryogenesis and bone development (9).
Structurally, POSTN is a multimodular protein composed of a signal peptide, which is necessary for secretion, a small cysteine-rich module (EMI domain) probably involved in the formation of multimers through cysteine disulfide bonds (15), four fasciclin-like domains (FAS1) that interact with integrins (αvβ3, αvβ5, α6β4) (16), and a hydrophilic C-terminal region known to interact with other ECM proteins such as collagens, fibronectin, tenascin C, or heparin (15, 17). Interesting, the FAS1 domains of human POSTN contain vitamin K-dependent γ-carboxyglutamic acid (Gla) residues (18), which are found in a small group of proteins called Gla-containing proteins. This group of particular proteins includes many coagulation and anticoagulation factors as well as bone-associated proteins such as osteocalcin (bone gamma-carboxyglutamate protein) and matrix-gla protein (MGP) (19). These proteins suffer posttranslational modifications by γ-glutamyl carboxylase (a vitamin K-dependent enzyme) on specific glutamyl amino acid residues (Glu) to produce γ-carboxyglutamic amino acid residues (Gla) (20, 21). The main known function of γ-carboxyglutamic acid residues is their ability to bind divalent cations such as calcium. Hence, Gla-containing proteins play important roles in the regulation of coagulation cascade and bone homeostasis (22, 23).
Expression and Function of POSTN in Normal Tissues
Periostin is preferentially expressed in the periosteum, hence its name. The periosteum is a specialized membrane, which covers the outer surface of bones and is responsible for growth in diameter of bone and for the cortical thickness. The activity of the periosteum is particularly elevated during the phases of embryonic development and body growth, although during the adult life, it also contributes to determinate bone diameter and, subsequently, bone strength (24, 25). Interestingly, POSTN is also expressed in other connective tissues rich in collagens undergone to mechanical stress such as the periodontal ligament (a specialized structure of the teeth) (26), heart valves (27), and tendons (28). During cardiac development, POSTN is highly expressed by embryonic fibroblasts (29) and pericardial cells that cover the embryonic heart (30), but not by cells of the cardiomyocyte lineages (31). In the bone, POSTN mRNA has been detected by in situ hybridization in pre-osteoblast cells (9).
As described above, POSTN is able to interact with cells through its FAS1 domains and ECM proteins through its N-terminal EMI domain and C-terminal region. These properties make POSTN a key player in the regulation of cell behavior and organization of the ECM. POSTN has been shown to bind integrins αvβ3 and αvβ5 in osteoblasts and several types of normal and cancer cells where it elicits activation of FAK, PI3-Kinase, and AKT signaling pathways (32–34). These findings suggest that POSTN can act as a prosurvival protein in many cellular contexts.
Periostin plays an important role in ECM structure and organization and particularly in collagen assembly. Collagen cross-linking is a natural process essential to provide stability to collagen-rich connective tissues. Two key elements in this process are BMP-1 and lysyl oxidase (LOX) (35). Briefly, BMP-1 cleavages the inactive form of LOX to produce the active LOX enzyme, which in turn catalyzes the covalent cross-linking of collagen molecules (35). Interestingly, POSTN binds BMP-1 and collagen I through its FAS1 domains and N-terminal EMI domain, respectively, and thus act as a key player in this process, serving as a scaffold for BMP-1 and collagens to accelerate collagen cross-linking (35). The importance of POSTN in collagen cross-linking is also supported by POSTN knockout animal models. Thus, POSTN null mice exhibit aberrant collagen fibrillogenesis in the periosteum and a decrease in collagen cross-linking observed in skin, tendons, and heart (36).
The function of the Gla residues is, however, much less known. Remarkably, POSTN has 28 glutamyl amino acid residues (Glu) that could be posttranslationally modified to generate γ-carboxyglutamic amino acid residues (Gla) (18). The high number of potential Gla residues present in POSTN contrasts with the number of Gla residues contained in others Gla proteins of the bone such as osteocalcin and matrix Gla protein, which have 3 and 5 Gla residues, respectively. Coutu et al. studied the form of POSTN (carboxylated vs uncarboxylated) that was secreted by adipocytes, chondrocytes, and osteoblasts differentiated from mesenchymal stem cells. They found that undifferentiated human mesenchymal cells and also differentiated adipocytes and osteoblasts synthetized carboxylated POSTN while no POSTN was detected in cells undergone chondrogenic differentiation. Interestingly, carboxylated POSTN was detected in the conditioned medium of undifferentiated human mesenchymal cells and differentiated adipocytes but not in the conditioned medium derived from differentiated osteoblasts. In the latter case, POSTN was found to be abundantly deposited in bone nodules produced in vitro, indicating that osteoblasts are able to produce carboxylated POSTN that is efficiently sequestered within the ECM in a calcium-dependent manner (18).
Regulation of POSTN Expression
Periostin expression is transcriptionally regulated by several transcription factors that are themselves involved in the commitment of pluripotent mesenchymal cells to cells of the osteoblastic lineage. Several studies indicate that the transcription factors Twist-1 and -2 are involved directly in the regulation of POSTN expression. For example, Oshima et al. detected a “Twist box” response element in the POSTN promoter and demonstrated that Twist can bind this Twist-box sequence and activate the POSTN promoter in a reporter assay (37). A direct demonstration about the involving of the Twist family of transcription factors in the regulation of POSTN expression comes from observations done in a rare autosomal recessive disease named Setleis syndrome (OMIM 227260) characterized by abnormal facial development. Franco et al. demonstrated that patients with this disease harbor mutations in the bHLH transcription factor TWIST2 and that expression of POSTN was severely downregulated in fibroblasts derived from patients with this disease. Reporter gene assays and ChIP assays demonstrate that wild-type TWIST2 was able to induce POSTN promoter, while the mutant TWIST2 found in the patients was not, providing a direct link between POSTN expression and TWIST2 (38). Other studies also suggest a relationship between Twist and POSTN. Thus, Hu et al. re-analyzed public data obtained from The Cancer Genome Atlas dataset and observed that POSTN expression levels correlated with Twist and Snail expression in lung cancer specimens (39). In addition, a Twist shRNA was also shown to be able to inhibit POSTN expression in prostate cancer cell lines (40).
c-Fos/c-Jun (AP-1) are other transcriptional factors that can be involved in the regulation of POSTN expression. Kashima et al. analyzed the expression of POSTN in bones from patients with fibrous dysplasia, a benign bone disease characterized by high expression of transcriptional factors such as c-Fos/c-Jun. Immunohistochemistry and in situ hybridization studies revealed that POSTN was expressed in the fibrous component of fibrous dysplasia lesions correlating with c-Fos expression. These authors also analyzed POSTN levels in the sclerotic lesions developed in transgenic mice overexpressing c-fos, which are similar to those observed in fibrous dysplasia. In these lesions, all transformed osteoblasts expressed high levels of POSTN whereas normal osteoblasts did not, providing a relationship between c-fos overexpression and POSTN expression (41). Other transcription factor that has been shown to regulate POSTN expression is p73. Landre et al. showed, using ChIP assays, that the transcription factor p73 binds POSTN promoter. They demonstrated that p73 conferred an invasive phenotype to glioblastoma cells, which was mediated by activation of POSTN. In fact, POSTN overexpression was sufficient to rescue the invasive phenotype of glioblastoma cells after p73 knockdown (42). Overexpression of the transcription factors Slug and Sox9 induced tenascin-C and POSTN expression.
There are several hormones and growth factors that have been shown to regulate POSTN expression, which are also involved in bone homeostasis. For example, several studies indicate that parathyroid hormone affects POSTN expression (43–46) and vice versa (45). Estrogens are other type of hormones that have been shown to regulate POSTN expression. So, estradiol induces POSTN mRNA expression in primary human periodontal ligament cells through stimulation of estrogen receptor B (47). Finally, some cytokines and growth factors also regulate the levels of POSTN. For instance, TGF-β induces POSTN expression in osteoblasts (26), human periodontal ligament cells (48–50), gingival fibroblasts (51, 52), and mesangial cells of the kidney (53). BMP-2 induces POSTN expression in a murine mesenchymal progenitor cell line (54) and in atrioventricular mesenchymal cells (55). Angiotensin II induces POSTN expression in rat cardiac fibroblasts (56) and vascular smooth muscle cells (57). Finally, several studies indicate that other growth factors and cytokines such as PDGF, bFGF, TNFα, IL-4, IL-13, or Oncostatin M have been shown to induce POSTN expression in determined cells and/or animal models (57–61).
Tumor POSTN as a Biomarker
Periostin expression is deregulated in several pathologies such as inflammation, tissue repair, and malignant transformation (6). Next, we will review the current knowledge about deregulation of POSTN expression in cancer. In general, high POSTN levels are usually associated with a more aggressive tumor behavior, advanced stage or poor prognosis, suggesting that POSTN levels could be a useful prognostic biomarker (Table 1).
Table 1
| Cancer | High expression in | Is associated with (1) | Reference |
|---|---|---|---|
| Prostate cancer | Stroma | OS (poor prognosis) | (62) |
| Stroma | PFS (poor prognosis) | (63) | |
| Stroma | High gleason score | (63–65) | |
| Stroma | Advanced stage | (63) | |
|
|
|||
| Lung cancer | Stroma | OS (poor prognosis) | (66) |
| Stroma | OS (poor prognosis) | (67) | |
|
|
|||
| Pancreatic cancer | Stroma and cancer epithelial cells | OS and PFS (poor prognosis) | (68) |
| Stroma produced by pancreatic stellate cells | Histological grade (poor prognosis) | (69) | |
|
|
|||
| Ovarian cancer | Stroma and cancer epithelial cells | OS and PFS (poor prognosis) | (70) |
| Stroma and cancer epithelial cells | Advanced stages and cancer recurrence | (71) | |
|
|
|||
| Breast cancer | Stroma and cancer epithelial cells | OS and PFS (poor prognosis) | (72, 73) |
| Cancer-associated fibroblasts | OS (poor prognosis) | (74) | |
|
|
|||
| Colorectal cancer | Stroma | OS and PFS (poor prognosis) | (75) |
| Cancer epithelial cells | Advanced stage and metastasis | (32, 76) | |
|
|
|||
| Hepatocellular carcinoma | Cancer epithelial cells | OS and PFS (poor prognosis) | (77) |
| Stroma and cancer epithelial cells | Tumor grade | (77) | |
| Stroma | Increased microvascular invasion (poor prognosis) | (78) | |
|
|
|||
| Bladder cancer | Stroma | Not studied | (14) |
| Extracellular vesicles | Tumor stage (poor prognosis) | (79) | |
|
|
|||
| Osteosarcoma | Tumor | OS and PFS (poor prognosis) | (80) |
Correlation between periostin expression levels and clinical parameters in solid tumors.
(1) OS, overall survival; PFS, progression-free survival.
Prostate Cancer
Several studies have demonstrated that POSTN expression is altered in prostate cancer. Nuzzo et al. focused their attention on the distribution of POSTN in tumor tissues. They showed that POSTN could be detected in the tumor stroma, cancer epithelial cells, as well as in peritumoral areas. High expression of POSTN in the stromal component was associated with shorter survival while lower POSTN expression in cancer epithelial cells was significantly correlated with shorter progression-free survival, suggesting that patients with high levels of POSTN in the stroma component and low levels in cancer epithelial cells had worse prognosis (62). Tischler et al. also showed that the strong POSTN expression observed in tumor stroma was associated with a shorter progression-free survival (63). Other studies have shown that high expression of POSTN in tumor stroma is related to Gleason score (63, 65), the stage of the tumor (63), and the degree of malignancy (64). High POSTN expression was associated with high fibronectin expression and low expression of integrin α4 in metastatic castration-resistant prostate cancer patients (81).
Lung Cancer
In non-small cell lung cancer (NSCLC), POSTN has been found in mesenchymal areas in tumor stroma, but not in the cancer epithelial cells themselves (66). Studies carried out in patients have revealed that high expression of POSTN in tumor stroma was significantly associated with shorter overall survival rates (66). A study accomplished by Nitsche et al. showed that POSTN expression was an independent prognostic factor in a multivariate analysis that included POSTN expression, histological tumor subtype, tumor stage, lymph node involvement, and resection status (67).
Pancreatic Cancer
In pancreatic ductal adenocarcinoma (PDAC), POSTN has been detected in cancer epithelial cells, pancreatic stellate cells, and tumor stroma. Ben et al. showed that high POSTN expression in stroma and cancer epithelial cells compared with adjacent tissue were indicative of poor prognosis (68). In other study, it was shown that high POSTN expression in tumor stroma correlated with pathological grade and with survival rate in PDAC patients (69).
Ovarian Cancer
Periostin is expressed a high levels in tumor stroma and in cancer epithelial cells in ovarian cancer. In a study, high POSTN expression in stroma was associated significantly with lower overall survival and progression-free survival (70). By contrast, high POSTN levels in cancer epithelial cells did not show significant prognostic value compared with patients with lower POSTN expression in the cancer cells. However, in other study, patients with high POSTN expression in both, stroma and cancer epithelial cells had the shortest overall survival and progression-free survival (70). High levels of POSTN have been also correlated with advanced clinical late stages (III/IV) and cancer recurrence (71).
Breast Cancer
Breast cancer is characterized by high POSTN expression in cancer epithelial cells when compared with normal tissue. The elevated expression of POSTN has been associated with poor progression free and overall survival (73). Lambert et al. studied the relationship between the expression of POSTN in cancer stem cells and the prognosis of the patients. Their studies revealed that high POSTN expression in these cells was associated with reduced relapse-free survival in basal-like type but not in breast cancers of the luminal type (72). In addition to cancer cells, POSTN is also present in cancer-associated fibroblasts (CAFs). In this case, the higher levels of POSTN detected in CAF are associated with the malignancy grade of tumors, suggesting that POSTN secreted by CAFs could be a key element in breast cancer progression (74).
Colorectal Carcinoma
In colorectal carcinoma, POSTN expression is increased compared with normal tissues. According to immunohistochemical analysis, POSTN is expressed by CAFs and secreted to the stroma (75). Several independent studies have shown a correlation between POSTN expression and disease aggressiveness. For example, Bao et al. showed that expression levels of POSTN mRNA were higher in liver metastasis than in the primary tumor derived from the same patients (32). A multivariate analysis described in Wu et al. showed that the presence of liver metastasis correlated with histological type, lymph node metastasis, TNM stage, and POSTN levels (76). Moreover, Li et al. described a positive correlation between high POSTN expression and tumor size, grade of cell differentiation, lymph node metastasis, serosal invasion, clinical stage, and 5-year survival rates (82). More recently, Xu et al. described a study carried out in two independent cohorts of patients showing that medium- and high-stromal POSTN expression predicted poor prognosis in patients with colorectal carcinoma (83). Finally, a multivariate analysis performed in patients with colorectal carcinoma showed that high expression of stromal POSTN was an independent prognostic biomarker of poor overall survival and progression-free survival (75). Taken together, these findings strongly support an important role for POSTN in the development, progression, and metastatic dissemination of colorectal cancer.
Liver Cancer
A strong POSTN expression has been observed in cancer epithelial cells and tumor stroma in hepatocellular carcinoma. Riener et al. showed that POSTN expression in cancer epithelial cells was associated with reduced overall survival and correlated with tumor grade (77). In other study, POSTN expression was associated with tumor nodules, microvascular invasion, Edmodson grade, TNM stage, and higher levels of vascular endothelial growth factor (VEGF) expression. Besides, the expression of POSTN was found to be an independent prognostic factor for predicting overall and disease-free survival in a multivariate analysis (84). More recently, Jang et al. showed that high POSTN expression in hepatocellular carcinoma was correlated with microvascular invasion, and advanced stage disease and patients with high perisotin expression had significantly lower overall survival rates suggesting a relationship between POSTN expression and poor prognosis (78). Utispan et al. showed that POSTN is over-expressed by CAFs in intrahepatic cholangiocarcinoma but not by cancer cells or immune infiltrating cells. Patients with higher levels of POSTN had shorter survival rates than those with lower levels (85).
Bladder Cancer
Unlike most tumors, POSTN expression appears to be downregulated in bladder cancer compared with normal tissue. Immunohistochemically, analysis demonstrated a strong staining of POSTN in the stroma of normal bladder, while it was mostly attenuated in bladder cancer tissues. Strikingly, POSTN downregulation correlated inversely with tumor grade. Thus, while POSTN expression was detected in 100% normal bladder tissues, only a 33% of grade 3 bladder cancer expressed POSTN (86). Normal bladder tissues expressed the canonical isoform of POSTN as well as others alternatively spliced mRNAs. By contrast, in bladder cancer tissues, the canonical POSTN isoform is not expressed and only some alternatively spliced isoforms can be detected. Interestingly, the alternative isoforms expressed in cancer tissues did not show tumor suppressor activity as the canonical isoform did (14, 86). These findings suggest that loss of canonical POSTN can be related with the development of bladder cancer.
However, other studies suggest an opposite role for POSTN in the development of bladder cancer. In muscle-invasive bladder cancer, an aggressive malignancy with high mortality, high POSTN expression correlated with worse prognosis. These tumors produce extracellular vesicles, which have been shown to promote cancer progression. Extracellular vesicles isolated from urine of patients had markedly higher levels of POSTN than controls, indicating that POSTN could be a potential urinary biomarker of cancer progression (79).
Osteosarcoma
Hu et al. analyzed the expression of POSTN and VEGF in patients with osteosarcoma. The histopathological and immunohistochemically analysis showed that POSTN expression was higher in osteosarcoma than in osteochondroma and correlated with VEGF expression, histological subtype, and tumor size (80). Furthermore, patients who showed higher levels of POSTN had a poorer prognosis than those with lower POSTN expression. Overall survival was twofold higher in patients with lower POSTN compared with patients with high POSTN expression (80).
Other Cancers
In esophageal squamous cell carcinoma, high POSTN expression correlated with poor prognosis and shorter overall survival. In a multivariate analysis including tumor differentiation, venous invasion, TNM stage, and other parameters, POSTN expression level was identified as an independent prognostic factor (87).
Morra et al. analyzed the levels of POSTN in renal cell carcinoma and showed that POSTN was expressed in mesenchymal cells, cancer epithelial cells, and tumor stroma. Higher levels of POSTN in cancer epithelial cells correlated with the presence of sarcomatoid differentiation, higher tumor stage, lymph node metastases, and poor overall survival. The authors suggest that POSTN expression in cancer epithelial cells may contribute to sarcomatoid differentiation and a more aggressive behavior of renal cell carcinoma (88).
Serum POSTN Levels as a Biomarker of Poor Prognosis
Since POSTN levels in tumor tissues show relationship with prognosis in many cancers, several studies have analyzed if POSTN levels in serum correlate with cancer prognosis. In fact, increased POSTN levels have been detected in serum of patients with some malignant tumors suggesting that it could be an effective biomarker, both for diagnosis and survival prediction (summarized in Table 2).
Table 2
| Cancer | Diagnostic/prognostic value | Reference |
|---|---|---|
| Lung cancer | Associated to bone metastasis | (89) |
| Associated to bone metastasis | (90) | |
| Response to chemotherapy | ||
| Associated to poor survival | ||
| Associated to poor survival (independent prognostic factor) | (91) | |
|
|
||
| Breast cancer | Associated to poor survival (in some subgroups) | (92) |
| Associated to bone metastasis | (93) | |
|
|
||
| Colorectal cancer | Associated to poor prognosis | (94) |
| Associated to metastasis | ||
|
|
||
| Hepatocellular carcinoma | Associated to poor prognosis | (95) |
|
|
||
| Cholangiocarcinoma | Associated to poor prognosis (independent prognostic factor) | (96) |
| Diagnostic value | (97) | |
Serum periostin levels as a prognostic factor.
Lung Cancer
Xu et al. analyzed the levels of serum POSTN in a large cohort of 296 patients with NSCLC. The levels of serum POSTN in patients with cancer were significantly elevated compared to healthy controls and patients with benign lung disease. Patients with the higher serum POSTN levels had a poor progression-free survival and overall survival than patients with lower levels. POSTN levels were shown to be an independent prognostic factor (91). In other similar study, Zhang et al. showed that serum POSTN levels were also significantly higher in NSCLC compared to patients with benign lung disease and healthy controls (90). It was also shown that serum POSTN levels were significantly associated with the presence of bone metastasis. Interestingly, POSTN levels before and after chemotherapy correlated significantly with prognosis suggesting that POSTN levels could be able to predict chemotherapy response and survival in patients with advanced NSCLC (90). Che et al. studied the levels of serum POSTN in patients with lung cancer with or without bone metastasis. Interestingly, patients with bone metastasis presented higher levels of POSTN compared to patients without bone metastasis and healthy controls (89). Other study, however, found no relationship between serum POSTN levels with clinic and pathologic parameters, although serum POSTN in the lung cancer patients were elevated compared with healthy controls (98).
Breast Cancer
The relationship between serum POSTN levels and prognosis in breast cancer has also been studied. Sasaki et al. found increased levels of serum POSTN in breast cancer patients with bone metastasis compared to patients without metastasis, although no correlation with other prognostic factors such as clinical stage and lymph node metastasis was found (93). More recently, Nuzzo et al. analyzed the serum POSTN levels in patients with early breast cancer who had received curative surgery and found some association between POSTN and prognosis only in some subgroups of patients. Specifically, they found a correlation between higher POSTN serum levels and mortality in patients with node-negative disease and in patients who did not receive adjuvant therapy (92).
Colorectal Cancer
Ben et al. showed that serum POSTN levels were significantly elevated in colorectal cancer patients compared to healthy controls or patients with benign colorectal polyps or adenomas. POSTN levels in serum correlated with mRNA expression levels in the tumor and also with tumor size, since serum POSTN levels undergone a significant decrease after surgery. Serum POSTN levels correlated significantly with distant metastasis, advanced-stage disease, and poor prognosis, suggesting that circulating POSTN may be of clinical value in identifying patients who could be at risk of metastasis and progression (94). By contrast, a recent study comparing serum POSTN levels between patients with and without bone metastasis not found significant differences between both groups although, in this study, the number of patients with colorectal cancer was low (99).
Liver Cancer
Serum POSTN levels have been shown to be increased in patients with hepatocellular carcinoma compared to healthy controls or patients with no cancerous liver diseases. Elevated serum POSTN were associated with poor overall survival and release-free survival. In multivariate analysis, POSTN was considered an independent prognostic marker for predicting survival (95). Fujimoto et al. analyzed the levels of serum POSTN in intrahepatic cholangiocarcinoma patients and observed that these were significantly higher in these patients compared to normal controls and patients with liver cirrhosis, hepatocellular carcinoma, and other malignancies, suggesting that serum POSTN levels could be useful to distinguish between cholangiocarcinoma and other hepatic malignancies (97). In a recent study, similar results were observed. Thus, serum POSTN levels in patients with cholangiocarcinoma were significantly increased compared with that in healthy controls, patients with benign liver diseases, and even patients with breast cancer, indicating that serum POSTN levels could be used as a diagnostic biomarker (96). Furthermore, high levels of serum POSTN, as well as tissue POSTN levels, were significantly associated with reduced survival rate. Serum POSTN was identified as an independent prognostic factor suggesting that it could be used as a marker of poor prognosis in patients with cholangiocarcinoma (96).
POSTN and Resistance to Treatment
Interestingly, several studies have showed that POSTN could play an important role in the resistance to treatment in some types of solid tumors (summarized in Table 3).
Table 3
| Cancer (cell lines) | Drug | Evidences | Pathways involved | Reference |
|---|---|---|---|---|
| Pancreatic cancer (SW1990, Panc-1) | Gemcitabine | In vitro, in vivo (xenograft) | Erk, Akt | (100) |
|
|
||||
| Non-small lung cancer (A549) | Cisplatin | In vitro, in vivo (xenograft) | Akt, Stat3, Survivin | (101) |
|
|
||||
| Ovarian cancer (A2780) | Cisplatin | In vitro, patients | Akt | (70) |
|
|
||||
| Ovarian cancer (ES-2) | Carboplatin Paclitaxel | In vitro, patients | Not studied | (102) |
|
|
||||
| Glioma (GSC272, GSC11) | Bevacizumab | In vitro, in vivo (xenograft) | TGFβ, hypoxia-inducible factor-1α, Stat3 | (103) |
|
|
||||
| Colon cancer (SW480, HT-29) | Oxaliplatin | In vitro | Akt, survivin | (104) |
| 5-fluorouracil | ||||
|
|
||||
| Gastric cancer (SGC-7901) | Cisplatin | In vitro | Bax, p53, Bcl-2, Akt | (105) |
| 5-fluorouracil | ||||
Periostin and resistance to chemotherapy.
For instance, Liu et al. showed that POSTN exerts a significant role in the resistance of pancreatic cancer cells to gemcitabine. In this study, it was shown that silencing of POSTN gene in pancreatic cancer cells sensitize gemcitabine-resistant cells to gemcitabine treatment both in vitro and in vivo, suggesting that POSTN targeting could be a new therapeutic approach to overcome gemcitabine resistance in pancreatic cancer (100).
Hu et al. studied the effect of POSTN on cisplatin resistance in the A549 NSCLC cells. It was shown that POSTN overexpression makes these cells more resistant to cisplatin-induced apoptosis, while POSTN knockdown sensitive these cells to cisplatin. These results indicate that POSTN induces cisplatin-resistance in non-small lung cancer and thus it could represent a new target for overcoming cisplatin-resistance (101).
Sung et al. also found a relationship between cisplatin resistance and POSTN in patients with ovarian carcinoma. In this case, patients with high levels of stromal POSTN showed cisplatin resistance compared to those with lower levels. In vitro, POSTN treatment of A2780 ovarian adenocarcinoma cells induced cisplatin-resistance and activation of the Akt pathway, which was reverted by incubation of the cells with an Akt inhibitor (70). Similar results were also observed by Ryner et al. in a large cohort of ovarian cancer patients (102). In this work, gene expression profile identified a specific gene signature associated to chemoresistant tumors, where POSTN was one of the three highest ranked genes. Furthermore, recombinant POSTN was able to promote resistance to carboplatin and paclitaxel in chemosensitive ovarian cells in vitro (102).
Park et al. analyzed the role of POSTN in the resistance to antiangiogenic therapy in glioma stem cells. Using mouse xenograft models of human glioma, they demonstrated that animals treated with bevacizumab expressed higher levels of POSTN than tumors derived from untreated animals. Interestingly, POSTN knockdown increased the median survival of the animals and the effect of POSTN knockdown in combination with bevacizumab was synergic (103). POSTN expression was associated with increased expression of TGFβ1 and hypoxia-inducible factor-1α in glioma stem cells, which explain the effect of POSTN on the resistance to antiangiogenic therapy in these cells.
Treatment of SW480 and HT-29 colon cancer cells with oxaliplatin or 5-fluorouracil increased the levels of POSTN mRNA and protein. POSTN knockdown increased significantly the sensibility of these cells to drug-induced apoptosis, which correlated with a reduction of survivin levels, linking POSTN to the antiapoptotic protein survivin. Conversely, POSTN overexpression increased survivin levels and Akt activation. All together, these results suggest that POSTN induces chemoresistance in colon cancer cells through activation of the PI3K/Akt/survivin pathway (104).
Similar results were also observed in gastric cancer cell lines. In this case, overexpression of POSTN in SGC-7901 cells rendered these cells more resistant to cisplatin or 5-fluorouracil-induced apoptosis. Cells overexpressing POSTN showed a reduction in the levels of Bax and p53 proteins and an increase of Bcl-2 proteins and Akt phosphorylation. Restoration of p53 expression or treatment of the cells with an inhibitor of Akt restored drug sensibility (105).
The molecular mechanism underlying the POSTN-mediated resistance to chemotherapy is largely unknown. Although activation of survival pathways seems to play an important role in the resistance to cancer drugs, other mechanisms, such as those mediated for the family of ATP-binding cassette proteins (ABC family) involved in the acquisition of the multiple drug resistance phenotype in cancer must be taken into consideration. In this sense, POSTN expression has been shown to correlate with some members of the ABC family in some cancer cells (106). Clearly, more studies are necessary to confirm if there is a relationship between POSTN expression and ABC proteins and to determine its contribution to chemotherapy resistance.
POSTN and the Hallmarks of Cancer
The deregulation of POSTN expression in many cancers suggests that it plays an important role in cancer development and progression. In addition, there are strong experimental evidences that involve POSTN in the acquisition of many hallmarks of cancer, such as proliferation, invasion and metastasis, angiogenesis, or cell survival (Figure 1; Table 4).
Figure 1
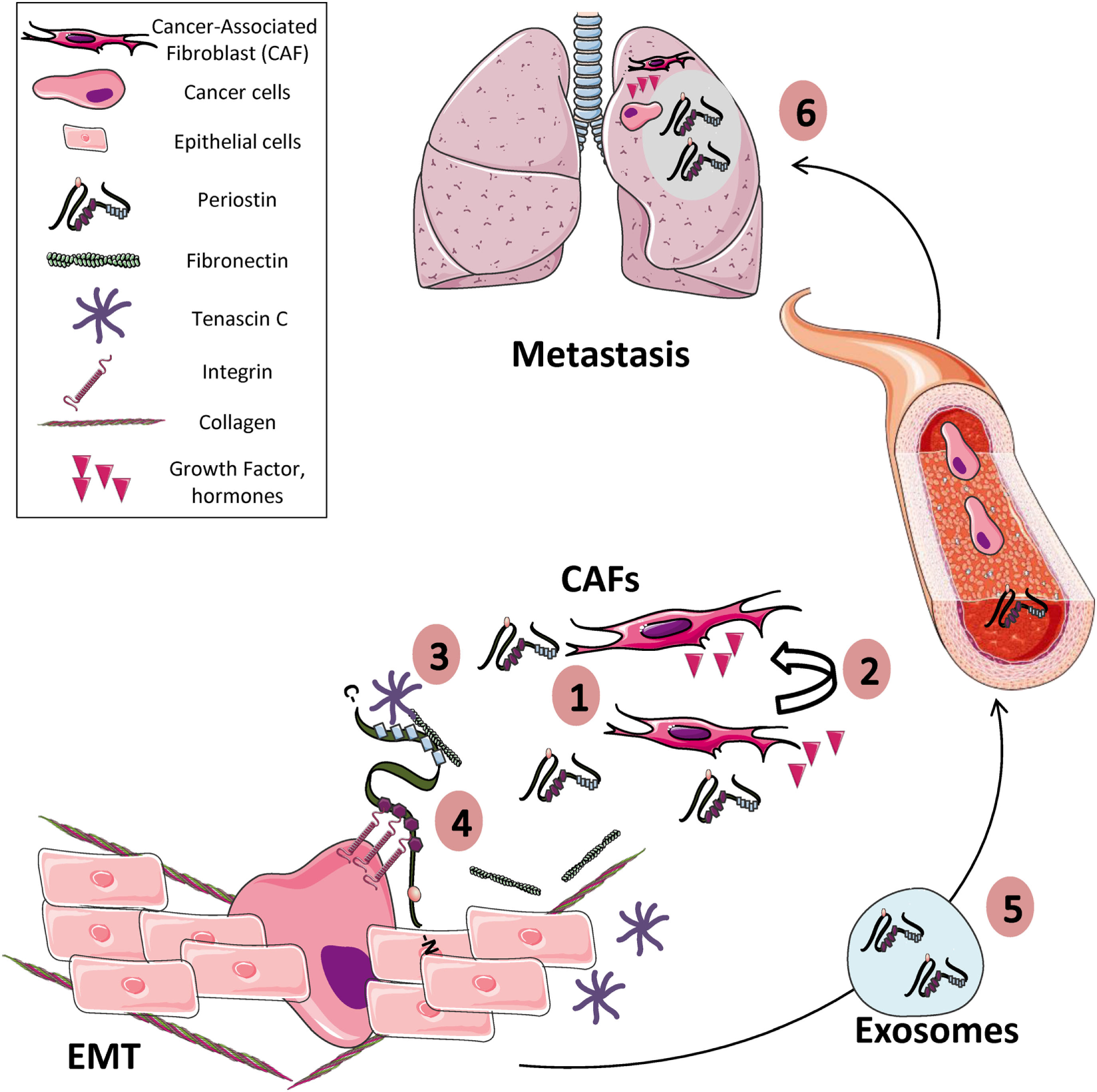
Periostin (POSTN) and the hallmarks of cancer. (1) POSTN is overexpressed in many solid tumors, mainly by stromal cells such as cancer-associated fibroblasts and to a lesser extent by tumor cells themselves. (2) Both paracrine and autocrine signals can stimulate the expression of POSTN by stromal and tumor cells. (3) Extracellularly secreted POSTN interacts with others extracellular matrix (ECM) proteins (e.g., collagens, tenascin C, fibronectin) contributing to produce a tumor-receptive ECM, by modulating, for example, collagen cross-linking. (4) In addition, POSTN interacts with integrin receptors present in the membrane of cancer cells, promoting cell proliferation, cell survival, epithelial–mesenchymal transition (EMT), and migration. (5) POSTN can be transported into exosomes produced from stromal and tumor cells to distant sites in other tissues (e.g., lung, liver, or bone) or produced by CAFs (6) where it contributes to prepare the metastatic niche before the arrival of tumor cells.
Table 4
| Cancer | Cell line | Effect of POSTN | Reference |
|---|---|---|---|
| Clear renal cancer | A498, 786-0 | Induces proliferation | (107) |
| 786-O, ACHN | Induces migration and invasion | (108) | |
|
|
|||
| Lung cancer | A549 | Induces proliferation and migration | (98) |
| CL1, CL1-5 | Induces epithelial–mesenchymal transition (EMT) | (80) | |
|
|
|||
| Gastric cancer | OCUM-2MLN | Induces proliferation | (109) |
| OCUM-12 | |||
|
|
|||
| Melanoma | NHDF | Induces proliferation | (110) |
| SP2/O, SKMEL-28 | Induces angiogenesis | (111) | |
| B6-BL6, B16 | Induces metastasis | (112) | |
| MC3T3-C1 | |||
|
|
|||
| Prostate cancer | RWPE-1, TA2 | Induces EMT | (64) |
| PC3, DU145 | Induces proliferation, invasion, and migration | (40) | |
|
|
|||
| Bladder cancer | SBT31A, T24 | Suppresses EMT and invasion | (113) |
|
|
|||
| Ovarian cancer | HOSE, CSOC, SK-OV-3 | Induces adhesion and migration | (13) |
| A2780 | Induces proliferation | (71). | |
|
|
|||
| Colorectal cancer | CX-1NS | Induces metastasis (preventing stress-induced apoptosis) | (32) |
|
|
|||
| Esophageal cancer | EPC2 | Induces invasion | (114–116) |
|
|
|||
| Glioblastoma | U87, T98G, and A172 | Induces proliferation, migration, and invasion | (117) |
| GSC and NSTCs | Promotes malignant growth | (118) | |
|
|
|||
| Breast cancer | MCF7 | Induces angiogenesis | (119) |
| MCF7, MDA-MB-231, 67NR, 4T1 | Induces metastasis | (120) | |
|
|
|||
| Head and neck cancer | HN4, HN6, HN13, HN30 | Induces metastasis | (121) |
| HNSCC | Induces lymphoangiogenesis | (122) | |
|
|
|||
| Oral squamous cell carcinoma | Rca-B, Rca-T | Induces metastasis | (121) |
|
|
|||
| Hepatocellular carcinoma | SMMC7721, Hep3B | Resistance to hipoxia | (123) |
|
|
|||
| Pancreatic cancer | AsPC-1, SW-1990, BxPC-3, Panc-1 | Induces proliferation and invasion | (68) |
Effect of periostin (POSTN) on tumor cells.
Sustaining Proliferative Signals
Periostin has been shown to induce proliferation in several tumor cell lines. For example, overexpression of POSTN in the A549 lung cancer cell line promoted cell proliferation and migration by inducing vimentin and N-cadherin expression and downregulating E-cadherin expression (98). Kikuchi et al. showed that POSTN increased proliferation of gastric cancer cell lines, concomitantly with the activation of ERK pathway. Interestingly, orthotopic xenograft tumors carried out with one gastric cancer cell line grew more slowly in Postn (−/−) mice than in wild-type mice, suggesting that POSTN produced by CAFs favor the establishment of a growth-supportive microenvironment for gastric cancer (109). In addition, Kotobuki et al. showed that recombinant POSTN and POSTN derived from normal human dermal fibroblasts induced melanoma cell proliferation by activating the MAPK signaling pathway (109, 110).
Activating Invasion and Metastasis
Periostin is involved in invasion and metastasis by regulating key processes such as epithelial–mesenchymal transition (EMT). Briefly, EMT is a physiological process, first discovered in embryonic development, and later observed also during tumor progression, consisting in the transition of epithelial cells to cells with mesenchymal phenotype. This process is particularly relevant in epithelial cancers, in which normal epithelial cells, which are polarized along their apical–basal axis and are tightly connected to each other, undergone a phenotypic transformation characterized by loss of apical–basal polarity and acquisition of migratory characteristic that increase invasion and mobility. EMT program is regulated by EMT-transcription factors (i.e., Snail, Twist, and Zeb families) and comprises the coordinated upregulation and downregulation of so-called EMT genes among which there are several ECM and cell–cell adhesion proteins (124–128).
Periostin has been shown to promote EMT in several types of cancer cells. For example, Hu et al. showed that recombinant POSTN induced EMT in A549 and CL1-0 lung cancer cells. POSTN induced upregulation of mesenchymal genes such as N-cadherin, Vimentin, Twist, and Snail and downregulation of E-cadherin. The effects of POSTN were mediated by activation of ERK and p38 pathways and by downregulation of miR-381 expression, a miRNA targeting Twist and Snail mRNAs (39). POSTN has been also shown to induce EMT in prostate cancer cells. In this case, POSTN overexpression in PC3 and DU145 prostate cancer cells promoted cell proliferation, invasion, and migration. POSTN increased the expression of EMT-associated factors and activated Akt and GSK-3β pathways. Interestingly, TGF-β increased POSTN and Twist expression, suggesting that POSTN is a mediator of TGF-β-induced EMT (40). Curiously, POSTN has been also shown to inhibit EMT in others cancer cells. Thus, Kim et al. studied the effect of POSTN on EMT and cell invasiveness in bladder and prostate cancer cell lines and found opposite effects. Particularly, POSTN upregulated E-cadherin expression and suppressed cell invasiveness in bladder cancer cells while the opposite effects were observed in prostate cancer cells, suggesting that POSTN effects regarding EMT are tumor cell-dependent (113).
Several studies suggest that POSTN can act facilitating the interaction between cancer cells and the tumor niche to promote cell migration. These interactions are mainly mediated by interactions with receptors of the integrin family. For example, POSTN has been shown to support adhesion and migration of ovarian epithelial cancer cells by interacting with αvβ3 and αvβ5 integrins and thus cells cultivated on POSTN showed more motility than cells cultivated on fibronectin (13, 111). Orecchia et al. demonstrated that proliferation of a melanoma cell in vivo was inhibited by the addition of antibodies directed against an epitope of POSTN involved in the interaction with αvβ3 and αvβ5 integrins, indicating that this interaction is critical for tumor growth (111). In addition, ectopic expression of POSTN in the non-metastatic 293T cells increased cell migration, invasion, and adhesion and required signaling through αvβ5 integrins and EGFR (129). Interestingly, inhibition of EGFR signaling was also shown to attenuate POSTN-mediated cell migration and invasion in esopahageal squamous cell carcinoma (114) supporting a relationship between POSTN and EGFR pathway.
Activating invasion is other critical step in the progression of malignant cancers. In this step, metalloproteinases play an important role by degrading different components of the extracellular matrix favoring the invasion of cancer cells. In this sense, POSTN has been shown to increase the activity of matrix metalloproteinase-2 and metalloproteinase-9 and promote migration and invasion in renal cell carcinoma cells (108, 109).
Finally, several studies indicate that POSTN can play a pivotal role in conditioning pre-metastatic niche and the formation of metastasis themselves. Fukuda et al. showed that the subcutaneous injection of osteoblasts that secrete large amounts of POSTN in vivo attracted remotely transplanted melanoma cells to the site of injection. By contrasts, osteoblasts in which POSTN expression was suppressed by shRNA showed a greatly reduced ability to promote metastasis (112). Malanchi et al. showed that infiltrating tumor cells are able to induce POSTN expression in the secondary target organ and that this ability is crucial for metastasis formation. Thus, the number of lung metastasis in an animal model was reduced in POSTN-null mice compared to wild-type control mice, suggesting that POSTN was a main mediator of metastasis (115). In addition, CAF-derived POSTN, induced by TGF-β3, was able to accelerate the migration and invasion of head and neck cancer cells (121). Taken together, these studies suggest that POSTN can act as a chemoattractant for cancer cells and that it is a key factor in metastatic colonization by conditioning of the premetastatic niche. Interestingly, POSTN could also be sent to the secondary metastatic sites by exosomes and thereby promote metastasis by conditioning the microenvironment of the target tissue before the arrival of tumor cells (120). In this sense, Vardaki et al. showed that the protein content of exosomes derived from metastatic human breast cancer cell lines was markedly different from that produced by non-metastatic cell lines. Specifically, exosomes secreted by metastatic cell lines showed a proteomic profile characterized by a high concentration of adhesion proteins such as β-Catenin, N-Cadherin, integrin-α2, integrin-β1, and POSTN. Remarkably, the characterization of exosomes isolated from the plasma of patients with metastatic or non-metastatic breast cancer showed similar characteristics to those observed in the metastatic and non-metastatic cell lines. These findings suggest that POSTN could serve as a biomarker for metastatic disease (120).
Inducing Angiogenesis
Periostin has been shown to act as a potent pro-angiogenic factor. Breast cancer cell lines overexpressing POSTN promote tumor angiogenesis in vivo by upregulating the VEGF receptor Flk-1/KDR in endothelial cells through activation FAK signaling via integrin-αvβ3 (119). In addition, POSTN expression in head and neck cancer has been shown to correlate with VEGF-C expression, both in tumor tissue and serum. POSTN promoted tube formation of lymphatic endothelial cells, which was mediated by Src and Akt pathway (122).
Resisting Cell Death
As previously mentioned above, POSTN acts as a prosurvival factor in several cellular contexts. POSTN has been shown to counteract hypoxia-induced apoptosis in cancer cells. For example, cancer cells that overexpressed POSTN were resistant to deferoxamine treatment, which mimic hypoxia (32). Aukkarasongsup et al. showed that periodontal cells in which POSTN expression was silenced using siRNAs were more susceptible to hipoxia-induced apopotosis than cells overexpressing perisotin. These effects were mediated by regulation of HIF-1α levels under hipoxic conditions (130). Furthermore, Liu et al. showed that POSTN expression was significantly increased in hepatocellular carcinoma cells under hypoxia. The regulation of POSTN was mediated by HIF-1α in such a way that HIF-1α depletion blocked the upregulation of POSTN induced by hipoxia. Interestingly, hypoxia-induced POSTN make the cells more resistant to arsenic trioxide treatment, and conversely, POSTN downregulation enhanced the anticancer effect of arsenic trioxide (123). In conclusion, POSTN could play an important role in the resistance to apoptosis under hypoxic conditions.
Avoiding Immune Destruction
Modulation of immune system is a key element in cancer progression. In glioblastoma, glioma stem cells secret POSTN that contribute to recruit tumor-associated macrophages (M2). Thus, Zhou et al. showed that the disruption of POSTN in glioma stem cells in vivo was able to reduce the recruitment of tumor-associated macrophages and inhibit tumor growth. Therefore, the interplay between glioma stem cells and tumor-associated macrophages can be modulated by POSTN and may be critical to promote glioblastoma tumor growth (118).
POSTN as a Therapeutic Target
Since POSTN plays multiple roles in cancer progression, targeting POSTN can be an attractive therapeutic approach. Several studies carried out in preclinical models suggest that blocking POSTN could be an attractive strategy to treat cancer. For example, Lee et al. used benzyl-d(U)TP-modified DNA aptamers (PNDAs) directed against human POSTN to treat breast cancer cell tumors in a xenograft mouse model. This strategy efficiently blocked tumor growth and cell dissemination to other organs (131). On the other hand, Kyutoku et al. used an anti-POSTN antibody (PN1-Ab), targeting the conserved exon 17 of POSTN, to treat mice of a lung metastasis model. Administration of this antibody significantly inhibited the growth of the primary tumors as well as the number of lung metastasis (132). Similar results were obtained by Zhu et al. using a monoclonal antibody (MZ-1) directed against POSTN in an in vivo model of ovarian cancer. In this case, in vivo administration of the antibody also produced a reduction in the number of metastasis (133).
Concluding Remarks
The accumulated knowledge about the relationship between POSTN and cancer in the last two decades indicate that this matricellular protein play an important role in cancer development and progression, beyond its implications in the homeostasis of specialized tissues (7). The fact that POSTN can modulate the hallmarks of cancer suggests that this multifaceted protein can have implications for cancer diagnosis and prognosis and, more importantly, for the development of therapies targeting POSTN function. Future research should be directed to consolidate some findings and explore new ones, particularly in the field of prognostic markers and therapy.
While it seems clear that POSTN is overexpressed in many tumors and that this overexpression is frequently associated with poor prognosis, data are not probably enough conclusive to stratify different patients groups in terms of clinical response. In this sense, quantification of tumor POSTN levels in large cohort of patients treated homogenously in the context of well-controlled clinical trials will be necessary to confirm if tumor POSTN could be used as a reliable prognostic marker. Standardization of the immunohistochemistry techniques used to detect POSTN in the tumor tissues (for instance, antibodies and methods of quantification) will also be necessary to reach solid conclusions. Several laboratories have studied the levels of POSTN in serum and its correlation with tumor burden and prognosis. In this case, the results are more complex and there are not clear conclusions about the usefulness of POSTN levels in serum as a prognostic marker. Again, studies in larger cohorts could be necessary to clarify this question.
In addition to its ability to predict outcome, and perhaps for this, POSTN has been shown to induce resistance to some chemotherapeutic drugs in determined cancer cell types. If this is true in the clinical setting must still be determined, again in the context of observational clinical trials. This association between POSTN and resistance to anticancer drugs suggests that could be necessary design new combinational therapies in determined types of cancer to precisely circumvent the effect of the POSTN. Evidently, more basic and translational research, specifically in clinical relevant animal models (i.e., patient-derived xenografts) must be performed before translating it to the clinic. Furthermore, the relationship between POSTN and the proteins involved in the multidrug resistant phenotype in cancer (i.e., ABC family of transporters) should be also studied in more detail.
One of the most important finding is the implication of POSTN in the metastatic process, including different aspects such as the EMT, cell migration, and “education” of the metastatic niche. The idea that tumor cells or tumor stromal cells can remotely send a protein such as POSTN to prepare the metastatic niche is really exciting. Interestingly, the current knowledge strongly supports this possibility. Thus, several studies performed in animal models have demonstrated that POSTN can be a key determinant player in the “education” of the metastatic niche. Since the consequences of this finding could have a great clinical impact, more research is necessary about this topic. One of the unsolved questions is to know how POSTN reach the metastatic niche. One interesting possibility is that POSTN produced by tumor stromal cells or in some cases by the tumor cells themselves was sent to metastatic niche encapsulated in circulating exosomes. In this case, interference with the transport process could be of therapeutic interest. Comprehensive characterization of these exosomes at the molecular level will be necessary to determine if these exosomes have some characteristic potentially targetable. In addition, detection of POSTN levels into circulating exosomes could predict the risk of metastasis in clinically localized tumors, which could be exploited to direct patient stratification.
Finally, we know few about the relationship between POSTN and immune system. Since POSTN seems to play a role in maintaining the hematopoietic niche, it is probable that it could also play a role in modulating immune cell activities in tumors. For example, it is unknown if POSTN expression is associated to “cold” or “hot” tumors from an immunological point of view.
In summary, POSTN is a multifunctional protein deregulated in many solid tumors, which modulates tumor cell behavior in many ways. Next years will provide us with new findings about the interesting role of this protein in tumor development and progression and its relationship with new therapeutic approaches.
Statements
Author contributions
LG-G and JA wrote the manuscript and designed the figures and tables. JA corrected and supervised the manuscript.
Acknowledgments
We sincerely apologize to all colleagues whose work has not been cited adequately in this review, either due to space limitations or unintentional omission. LG-G and JA are supported by Asociación Pablo Ugarte, ASION-La Hucha de Tomás, Fundación “Sonrisa de Alex,” and Instituto de Salud Carlos III (PI16CIII/00026). Some elements used to draw image 1 are attributed to Smart Servier Medical Art (smart.servier.com), which are licensed under a Creative Commons Attribution 3.0 Unported License.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationship that could be construed as a potential conflict of interest.
References
1
Hanahan D Weinberg RA . The hallmarks of cancer. Cell (2000) 100(1):57–70.10.1016/S0092-8674(00)81683-9
2
Hanahan D Weinberg RA . Hallmarks of cancer: the next generation. Cell (2011) 144(5):646–74.10.1016/j.cell.2011.02.013
3
Pickup MW Mouw JK Weaver VM . The extracellular matrix modulates the hallmarks of cancer. EMBO Rep (2014) 15(12):1243–53.10.15252/embr.201439246
4
Lu P Weaver VM Werb Z . The extracellular matrix: a dynamic niche in cancer progression. J Cell Biol (2012) 196(4):395–406.10.1083/jcb.201102147
5
Bornstein P Mcpherson JM Sage H Bornstein P Mcpherson J Sage H et al Synthesis and secretion of structural macromolecules by endothelial cells in culture. Pathobiol Endothelial Cell (1982) 6:215–28.10.1016/B978-0-12-521980-8.50020-1
6
Bornstein P Sage EH . Matricellular proteins: extracellular modulators of cell function. Curr Opin Cell Biol (2002) 14(5):608–16.10.1016/S0955-0674(02)00361-7
7
Murphy-Ullrich JE Sage EH . Revisiting the matricellular concept. Matrix Biol (2014) 37:1–14.10.1016/j.matbio.2014.07.005
8
Takeshita S Kikuno R Tezuka K Amann E . Osteoblast-specific factor 2: cloning of a putative bone adhesion protein with homology with the insect protein fasciclin I. Biochem J (1993) 294(Pt 1):271–8.10.1042/bj2940271
9
Litvin J Selim A-H Montgomery MO Lehmann K Rico MC Devlin H et al Expression and function of periostin-isoforms in bone. J Cell Biochem (2004) 92(5):1044–61.10.1002/jcb.20115
10
Nuzzo PV Buzzatti G Ricci F Rubagotti A Argellati F Zinoli L et al Periostin: a novel prognostic and therapeutic target for genitourinary cancer? Clin Genitourin Cancer (2014) 12(5):301–11.10.1016/j.clgc.2014.02.005
11
Kudo A . Introductory review: periostin-gene and protein structure. Cell Mol Life Sci (2017) 74(23):4259–68.10.1007/s00018-017-2643-5
12
Sugiura T Takamatsu H Kudo A Amann E . Expression and characterization of murine osteoblast-specific factor 2 (OSF-2) in a baculovirus expression system. Protein Expr Purif (1995) 6(3):305–11.10.1006/prep.1995.1040
13
Gillan L Matei D Fishman DA Gerbin CS Karlan BY Chang DD . Periostin secreted by epithelial ovarian carcinoma is a ligand for alpha(V)beta(3) and alpha(V)beta(5) integrins and promotes cell motility. Cancer Res (2002) 62(18):5358–64.
14
Kim CJ Isono T Tambe Y Chano T Okabe H Okada Y et al Role of alternative splicing of periostin in human bladder carcinogenesis. Int J Oncol (2008) 32(1):161–9.10.3892/ijo.32.1.161
15
Kii I Nishiyama T Li M Matsumoto K Saito M Amizuka N et al Incorporation of Tenascin-C into the extracellular matrix by periostin underlies an extracellular meshwork architecture. J Biol Chem (2010) 285(3):2028–39.10.1074/jbc.M109.051961
16
Hoersch S Andrade-Navarro MA . Periostin shows increased evolutionary plasticity in its alternatively spliced region. BMC Evol Biol (2010) 10(1):30.10.1186/1471-2148-10-30
17
Norris RA Moreno-Rodriguez RA Sugi Y Hoffman S Amos J Hart MM et al Periostin regulates atrioventricular valve maturation. Dev Biol (2008) 316(2):200–13.10.1016/j.ydbio.2008.01.003
18
Coutu DL Wu JH Monette A Rivard GE Blostein MD Galipeau J . Periostin, a member of a novel family of vitamin K-dependent proteins, is expressed by mesenchymal stromal cells. J Biol Chem (2008) 283(26):17991–8001.10.1074/jbc.M708029200
19
Ulrich MM Furie B Jacobs MR Vermeer C Furie BC . Vitamin K-dependent carboxylation. A synthetic peptide based upon the gamma-carboxylation recognition site sequence of the prothrombin propeptide is an active substrate for the carboxylase in vitro. J Biol Chem (1988) 263(20):9697–702.
20
Berkner KL . Vitamin K-dependent carboxylation. Vitam Horm (2008) 78:131–56.10.1016/S0083-6729(07)00007-6
21
Kidd PM . Vitamins D and K as pleiotropic nutrients: clinical importance to the skeletal and cardiovascular systems and preliminary evidence for synergy. Altern Med Rev (2010) 15(3):199–222.
22
Knapen MH Nieuwenhuijzen Kruseman AC Wouters RS Vermeer C . Correlation of serum osteocalcin fractions with bone mineral density in women during the first 10 years after menopause. Calcif Tissue Int (1998) 63(5):375–9.10.1007/s002239900543
23
Oldenburg J Marinova M Muller-Reible C Watzka M . The vitamin K cycle. Vitam Horm (2008) 78:35–62.10.1016/S0083-6729(07)00003-9
24
Merle B Garnero P . The multiple facets of periostin in bone metabolism. Osteoporos Int (2012) 23(4):1199–212.10.1007/s00198-011-1892-7
25
Bonnet N Garnero P Ferrari S . Periostin action in bone. Mol Cell Endocrinol (2016) 432:75–82.10.1016/j.mce.2015.12.014
26
Horiuchi K Amizuka N Takeshita S Takamatsu H Katsuura M Ozawa H et al Identification and characterization of a novel protein, periostin, with restricted expression to periosteum and periodontal ligament and increased expression by transforming growth factor beta. J Bone Miner Res (1999) 14(7):1239–49.10.1359/jbmr.1999.14.7.1239
27
Kruzynska-Frejtag A Machnicki M Rogers R Markwald RR Conway SJ . Periostin (an osteoblast-specific factor) is expressed within the embryonic mouse heart during valve formation. Mech Dev (2001) 103(1–2):183–8.10.1016/S0925-4773(01)00356-2
28
Yoshiba N Yoshiba K Hosoya A Saito M Yokoi T Okiji T et al Association of TIMP-2 with extracellular matrix exposed to mechanical stress and its co-distribution with periostin during mouse mandible development. Cell Tissue Res (2007) 330(1):133–45.10.1007/s00441-007-0439-x
29
Ieda M Tsuchihashi T Ivey KN Ross RS Hong TT Shaw RM et al Cardiac fibroblasts regulate myocardial proliferation through β1 integrin signaling. Dev Cell (2009) 16(2):233–44.10.1016/j.devcel.2008.12.007
30
Snider P Hinton RB Moreno-Rodriguez RA Wang J Rogers R Lindsley A et al Periostin is required for maturation and extracellular matrix stabilization of noncardiomyocyte lineages of the heart. Circ Res (2008) 102(7):752–60.10.1161/CIRCRESAHA.107.159517
31
Snider P Standley KN Wang J Azhar M Doetschman T Conway SJ . Origin of cardiac fibroblasts and the role of periostin. Circ Res (2009) 105(10):934–47.10.1161/CIRCRESAHA.109.201400
32
Bao S Ouyang G Bai X Huang Z Ma C Liu M et al Periostin potently promotes metastatic growth of colon cancer by augmenting cell survival via the Akt/PKB pathway. Cancer Cell (2004) 5(4):329–39.10.1016/S1535-6108(04)00081-9
33
Butcher JT Norris RA Hoffman S Mjaatvedt CH Markwald RR . Periostin promotes atrioventricular mesenchyme matrix invasion and remodeling mediated by integrin signaling through Rho/PI 3-kinase. Dev Biol (2007) 302(1):256–66.10.1016/j.ydbio.2006.09.048
34
Li G Jin R Norris RA Zhang L Yu S Wu F et al Periostin mediates vascular smooth muscle cell migration through the integrins alphavbeta3 and alphavbeta5 and focal adhesion kinase (FAK) pathway. Atherosclerosis (2010) 208(2):358–65.10.1016/j.atherosclerosis.2009.07.046
35
Maruhashi T . Interaction between periostin and BMP-1 promotes proteolytic activation of lysyl oxidase. J Biol Chem (2010) 285(17):13294–303.
36
Canty EG Kadler KE . Procollagen trafficking, processing and fibrillogenesis. J Cell Sci (2005) 118(Pt 7):1341–53.10.1242/jcs.01731
37
Oshima A Tanabe H Yan T Lowe GN Glackin CA Kudo A . A novel mechanism for the regulation of osteoblast differentiation: transcription of periostin, a member of the fasciclin I family, is regulated by the bHLH transcription factor, twist. J Cell Biochem (2002) 86(4):792–804.10.1002/jcb.10272
38
Franco HL Casasnovas JJ Leon RG Friesel R Ge Y Desnick RJ et al Nonsense mutations of the bHLH transcription factor TWIST2 found in Setleis syndrome patients cause dysregulation of periostin. Int J Biochem Cell Biol (2011) 43(10):1523–31.10.1016/j.biocel.2011.07.003
39
Hu WW Chen PC Chen JM Wu YM Liu PY Lu CH et al Periostin promotes epithelial-mesenchymal transition via the MAPK/miR-381 axis in lung cancer. Oncotarget (2017) 8(37):62248–60.10.18632/oncotarget.19273
40
Hu Q Tong S Zhao X Ding W Gou Y Xu K et al Periostin mediates TGF-beta-induced epithelial mesenchymal transition in prostate cancer cells. Cell Physiol Biochem (2015) 36(2):799–809.10.1159/000430139
41
Kashima TG Nishiyama T Shimazu K Shimazaki M Kii I Grigoriadis AE et al Periostin, a novel marker of intramembranous ossification, is expressed in fibrous dysplasia and in c-Fos-overexpressing bone lesions. Hum Pathol (2009) 40(2):226–37.10.1016/j.humpath.2008.07.008
42
Landre V Antonov A Knight R Melino G . p73 promotes glioblastoma cell invasion by directly activating POSTN (periostin) expression. Oncotarget (2016) 7(11):11785–802.10.18632/oncotarget.7600
43
Onyia JE Helvering LM Gelbert L Wei T Huang S Chen P et al Molecular profile of catabolic versus anabolic treatment regimens of parathyroid hormone (PTH) in rat bone: an analysis by DNA microarray. J Cell Biochem (2005) 95(2):403–18.10.1002/jcb.20438
44
Li X Liu H Qin L Tamasi J Bergenstock M Shapses S et al Determination of dual effects of parathyroid hormone on skeletal gene expression in vivo by microarray and network analysis. J Biol Chem (2007) 282(45):33086–97.10.1074/jbc.M705194200
45
Fortunati D Reppe S Fjeldheim AK Nielsen M Gautvik VT Gautvik KM . Periostin is a collagen associated bone matrix protein regulated by parathyroid hormone. Matrix Biol (2010) 29(7):594–601.10.1016/j.matbio.2010.07.001
46
Bonnet N Conway SJ Ferrari SL . Regulation of beta catenin signaling and parathyroid hormone anabolic effects in bone by the matricellular protein periostin. Proc Natl Acad Sci U S A (2012) 109(37):15048–53.10.1073/pnas.1203085109
47
Mamalis A Markopoulou C Lagou A Vrotsos I . Oestrogen regulates proliferation, osteoblastic differentiation, collagen synthesis and periostin gene expression in human periodontal ligament cells through oestrogen receptor beta. Arch Oral Biol (2011) 56(5):446–55.10.1016/j.archoralbio.2010.11.001
48
Wen W Chau E Jackson-Boeters L Elliott C Daley TD Hamilton DW . TGF-ss1 and FAK regulate periostin expression in PDL fibroblasts. J Dent Res (2010) 89(12):1439–43.10.1177/0022034510378684
49
Watanabe T Yasue A Tanaka E . Hypoxia-inducible factor-1alpha is required for transforming growth factor-beta1-induced type I collagen, periostin and alpha-smooth muscle actin expression in human periodontal ligament cells. Arch Oral Biol (2014) 59(6):595–600.10.1016/j.archoralbio.2014.03.003
50
Hasegawa D Wada N Maeda H Yoshida S Mitarai H Tomokiyo A et al Wnt5a induces collagen production by human periodontal ligament cells through TGFbeta1-mediated upregulation of periostin expression. J Cell Physiol (2015) 230(11):2647–60.10.1002/jcp.24950
51
Arancibia R Oyarzun A Silva D Tobar N Martinez J Smith PC . Tumor necrosis factor-alpha inhibits transforming growth factor-beta-stimulated myofibroblastic differentiation and extracellular matrix production in human gingival fibroblasts. J Periodontol (2013) 84(5):683–93.10.1902/jop.2012.120225
52
Kim SS Jackson-Boeters L Darling MR Rieder MJ Hamilton DW . Nifedipine induces periostin expression in gingival fibroblasts through TGF-beta. J Dent Res (2013) 92(11):1022–8.10.1177/0022034513503659
53
Sen K Lindenmeyer MT Gaspert A Eichinger F Neusser MA Kretzler M et al Periostin is induced in glomerular injury and expressed de novo in interstitial renal fibrosis. Am J Pathol (2011) 179(4):1756–67.10.1016/j.ajpath.2011.06.002
54
Noack S Seiffart V Willbold E Laggies S Winkel A Shahab-Osterloh S et al Periostin secreted by mesenchymal stem cells supports tendon formation in an ectopic mouse model. Stem Cells Dev (2014) 23(16):1844–57.10.1089/scd.2014.0124
55
Inai K Norris RA Hoffman S Markwald RR Sugi Y . BMP-2 induces cell migration and periostin expression during atrioventricular valvulogenesis. Dev Biol (2008) 315(2):383–96.10.1016/j.ydbio.2007.12.028
56
Li L Fan D Wang C Wang JY Cui XB Wu D et al Angiotensin II increases periostin expression via Ras/p38 MAPK/CREB and ERK1/2/TGF-beta1 pathways in cardiac fibroblasts. Cardiovasc Res (2011) 91(1):80–9.10.1093/cvr/cvr067
57
Li G Oparil S Sanders JM Zhang L Dai M Chen LB et al Phosphatidylinositol-3 kinase signaling mediates vascular smooth muscle cell expression of periostin in vivo and in vitro. Atherosclerosis (2006) 188(2):292–300.10.1016/j.atherosclerosis.2005.11.002
58
Takayama G Arima K Kanaji T Toda S Tanaka H Shoji S et al Periostin: a novel component of subepithelial fibrosis of bronchial asthma downstream of IL-4 and IL-13 signals. J Allergy Clin Immunol (2006) 118(1):98–104.10.1016/j.jaci.2006.02.046
59
Ouyang G Liu M Ruan K Song G Mao Y Bao S . Upregulated expression of periostin by hypoxia in non-small-cell lung cancer cells promotes cell survival via the Akt/PKB pathway. Cancer Lett (2009) 281(2):213–9.10.1016/j.canlet.2009.02.030
60
Rani S Barbe MF Barr AE Litivn J . Role of TNF alpha and PLF in bone remodeling in a rat model of repetitive reaching and grasping. J Cell Physiol (2010) 225(1):152–67.10.1002/jcp.22208
61
Lee MJ Heo SC Shin SH Kwon YW Do EK Suh DS et al Oncostatin M promotes mesenchymal stem cell-stimulated tumor growth through a paracrine mechanism involving periostin and TGFBI. Int J Biochem Cell Biol (2013) 45(8):1869–77.10.1016/j.biocel.2013.05.027
62
Nuzzo PV Rubagotti A Zinoli L Ricci F Salvi S Boccardo S et al Prognostic value of stromal and epithelial periostin expression in human prostate cancer: correlation with clinical pathological features and the risk of biochemical relapse or death. BMC Cancer (2012) 12:625.10.1186/1471-2407-12-625
63
Tischler V Fritzsche FR Wild PJ Stephan C Seifert HH Riener MO et al Periostin is up-regulated in high grade and high stage prostate cancer. BMC Cancer (2010) 10:273.10.1186/1471-2407-10-273
64
Tsunoda T Furusato B Takashima Y Ravulapalli S Dobi A Srivastava S et al The increased expression of periostin during early stages of prostate cancer and advanced stages of cancer stroma. Prostate (2009) 69(13):1398–403.10.1002/pros.20988
65
Tian Y Choi CH Li QK Rahmatpanah FB Chen X Kim SR et al Overexpression of periostin in stroma positively associated with aggressive prostate cancer. PLoS One (2015) 10(3):e0121502.10.1371/journal.pone.0121502
66
Hong LZ Wei XW Chen JF Shi Y . Overexpression of periostin predicts poor prognosis in non-small cell lung cancer. Oncol Lett (2013) 6(6):1595–603.10.3892/ol.2013.1590
67
Nitsche U Stangel D Pan Z Schlitter AM Esposito I Regel I et al Periostin and tumor-stroma interactions in non-small cell lung cancer. Oncol Lett (2016) 12(5):3804–10.10.3892/ol.2016.5132
68
Ben QW Jin XL Liu J Cai X Yuan F Yuan YZ . Periostin, a matrix specific protein, is associated with proliferation and invasion of pancreatic cancer. Oncol Rep (2011) 25(3):709–16.10.3892/or.2011.1140
69
Liu Y Du L . Role of pancreatic stellate cells and periostin in pancreatic cancer progression. Tumour Biol (2015) 36(5):3171–7.10.1007/s13277-015-3386-2
70
Sung PL Jan YH Lin SC Huang CC Lin H Wen KC et al Periostin in tumor microenvironment is associated with poor prognosis and platinum resistance in epithelial ovarian carcinoma. Oncotarget (2016) 7(4):4036–47.10.18632/oncotarget.6700
71
Zhu M Fejzo MS Anderson L Dering J Ginther C Ramos L et al Periostin promotes ovarian cancer angiogenesis and metastasis. Gynecol Oncol (2010) 119(2):337–44.10.1016/j.ygyno.2010.07.008
72
Lambert AW Wong CK Ozturk S Papageorgis P Raghunathan R Alekseyev Y et al Tumor cell-derived periostin regulates cytokines that maintain breast cancer stem cells. Mol Cancer Res (2016) 14(1):103–13.10.1158/1541-7786.MCR-15-0079
73
Kim GE Lee JS Park MH Yoon JH . Epithelial periostin expression is correlated with poor survival in patients with invasive breast carcinoma. PLoS One (2017) 12(11):e0187635.10.1371/journal.pone.0187635
74
Ratajczak-Wielgomas K Grzegrzolka J Piotrowska A Gomulkiewicz A Witkiewicz W Dziegiel P . Periostin expression in cancer-associated fibroblasts of invasive ductal breast carcinoma. Oncol Rep (2016) 36(5):2745–54.10.3892/or.2016.5095
75
Oh HJ Bae JM Wen XY Cho NY Kim JH Kang GH . Overexpression of POSTN in tumor stroma is a poor prognostic indicator of colorectal cancer. J Pathol Transl Med (2017) 51(3):306–13.10.4132/jptm.2017.01.19
76
Wu G Wang X Zhang X . Clinical implications of periostin in the liver metastasis of colorectal cancer. Cancer Biother Radiopharm (2013) 28(4):298–302.10.1089/cbr.2012.1374
77
Riener MO Fritzsche FR Soll C Pestalozzi BC Probst-Hensch N Clavien PA et al Expression of the extracellular matrix protein periostin in liver tumours and bile duct carcinomas. Histopathology (2010) 56(5):600–6.10.1111/j.1365-2559.2010.03527.x
78
Jang SY Park SY Lee HW Choi YK Park KG Yoon GS et al The combination of periostin overexpression and microvascular invasion is related to a poor prognosis for hepatocellular carcinoma. Gut Liver (2016) 10(6):948–54.10.5009/gnl15481
79
Silvers CR Liu YR Wu CH Miyamoto H Messing EM Lee YF . Identification of extracellular vesicle-borne periostin as a feature of muscle-invasive bladder cancer. Oncotarget (2016) 7(17):23335–45.10.18632/oncotarget.8024
80
Hu F Shang XF Wang W Jiang W Fang C Tan D et al High-level expression of periostin is significantly correlated with tumour angiogenesis and poor prognosis in osteosarcoma. Int J Exp Pathol (2016) 97(1):86–92.10.1111/iep.12171
81
Konac E Kiliccioglu I Sogutdelen E Dikmen AU Albayrak G Bilen CY . Do the expressions of epithelial-mesenchymal transition proteins, periostin, integrin-alpha4 and fibronectin correlate with clinico-pathological features and prognosis of metastatic castration-resistant prostate cancer?Exp Biol Med (Maywood) (2017) 242(18):1795–801.10.1177/1535370217728499
82
Li Z Zhang X Yang Y Yang S Dong Z Du L et al Periostin expression and its prognostic value for colorectal cancer. Int J Mol Sci (2015) 16(6):12108–18.10.3390/ijms160612108
83
Xu X Chang W Yuan J Han X Tan X Ding Y et al Periostin expression in intra-tumoral stromal cells is prognostic and predictive for colorectal carcinoma via creating a cancer-supportive niche. Oncotarget (2016) 7(1):798–813.10.18632/oncotarget.5985
84
Lv Y Wang W Jia WD Sun QK Li JS Ma JL et al High-level expression of periostin is closely related to metastatic potential and poor prognosis of hepatocellular carcinoma. Med Oncol (2013) 30(1):385.10.1007/s12032-012-0385-7
85
Utispan K Thuwajit P Abiko Y Charngkaew K Paupairoj A Chau-in S et al Gene expression profiling of cholangiocarcinoma-derived fibroblast reveals alterations related to tumor progression and indicates periostin as a poor prognostic marker. Mol Cancer (2010) 9:13.10.1186/1476-4598-9-13
86
Kim CJ Yoshioka N Tambe Y Kushima R Okada Y Inoue H . Periostin is down-regulated in high grade human bladder cancers and suppresses in vitro cell invasiveness and in vivo metastasis of cancer cells. Int J Cancer (2005) 117(1):51–8.10.1002/ijc.21120
87
Wang W Sun QK He YF Ma DC Xie MR Ji CS et al Overexpression of periostin is significantly correlated to the tumor angiogenesis and poor prognosis in patients with esophageal squamous cell carcinoma. Int J Clin Exp Pathol (2014) 7(2):593–601.
88
Morra L Rechsteiner M Casagrande S Duc Luu V Santimaria R Diener PA et al Relevance of periostin splice variants in renal cell carcinoma. Am J Pathol (2011) 179(3):1513–21.10.1016/j.ajpath.2011.05.035
89
Che J Shen WZ Deng Y Dai YH Liao YD Yuan XL et al Effects of lentivirus-mediated silencing of periostin on tumor microenvironment and bone metastasis via the integrin-signaling pathway in lung cancer. Life Sci (2017) 182:10–21.10.1016/j.lfs.2017.05.030
90
Zhang Y Yuan D Yao Y Sun W Shi Y Su X . Predictive and prognostic value of serum periostin in advanced non-small cell lung cancer patients receiving chemotherapy. Tumour Biol (2017) 39(5):1010428317698367.10.1177/1010428317698367
91
Xu CH Wang W Lin Y Qian LH Zhang XW Wang QB et al Diagnostic and prognostic value of serum periostin in patients with non-small cell lung cancer. Oncotarget (2017) 8(12):18746–53.10.18632/oncotarget.13004
92
Nuzzo PV Rubagotti A Argellati F Di Meglio A Zanardi E Zinoli L et al Prognostic value of preoperative serum levels of periostin (PN) in early breast cancer (BCa). Int J Mol Sci (2015) 16(8):17181–92.10.3390/ijms160817181
93
Sasaki H Yu CY Dai M Tam C Loda M Auclair D et al Elevated serum periostin levels in patients with bone metastases from breast but not lung cancer. Breast Cancer Res Treat (2003) 77(3):245–52.10.1023/A:1021899904332
94
Ben QW Zhao Z Ge SF Zhou J Yuan F Yuan YZ . Circulating levels of periostin may help identify patients with more aggressive colorectal cancer. Int J Oncol (2009) 34(3):821–8.10.3892/ijo_00000208
95
Lv Y Wang W Jia WD Sun QK Huang M Zhou HC et al High preoparative levels of serum periostin are associated with poor prognosis in patients with hepatocellular carcinoma after hepatectomy. Eur J Surg Oncol (2013) 39(10):1129–35.10.1016/j.ejso.2013.06.023
96
Thuwajit C Thuwajit P Jamjantra P Pairojkul C Wongkham S Bhudhisawasdi V et al Clustering of patients with intrahepatic cholangiocarcinoma based on serum periostin may be predictive of prognosis. Oncol Lett (2017) 14(1):623–34.10.3892/ol.2017.6250
97
Fujimoto K Kawaguchi T Nakashima O Ono J Ohta S Kawaguchi A et al Periostin, a matrix protein, has potential as a novel serodiagnostic marker for cholangiocarcinoma. Oncol Rep (2011) 25(5):1211–6.10.3892/or.2011.1194
98
Hong L Sun H Lv X Yang D Zhang J Shi Y . Expression of periostin in the serum of NSCLC and its function on proliferation and migration of human lung adenocarcinoma cell line (A549) in vitro. Mol Biol Rep (2010) 37(5):2285–93.10.1007/s11033-009-9721-1
99
Windrichova J Fuchsova R Kucera R Topolcan O Fiala O Finek J et al Testing of a novel cancer metastatic multiplex panel for the detection of bone-metastatic disease – a pilot study. Anticancer Res (2016) 36(4):1973–8.
100
Liu Y Li F Gao F Xing L Qin P Liang X et al Periostin promotes the chemotherapy resistance to gemcitabine in pancreatic cancer. Tumour Biol (2016) 37(11):15283–91.10.1007/s13277-016-5321-6
101
Hu W Jin P Liu W . Periostin contributes to cisplatin resistance in human non-small cell lung cancer A549 cells via activation of Stat3 and Akt and upregulation of survivin. Cell Physiol Biochem (2016) 38(3):1199–208.10.1159/000443068
102
Ryner L Guan Y Firestein R Xiao Y Choi Y Rabe C et al Upregulation of periostin and reactive stroma is associated with primary chemoresistance and predicts clinical outcomes in epithelial ovarian cancer. Clin Cancer Res (2015) 21(13):2941–51.10.1158/1078-0432.CCR-14-3111
103
Park SY Piao Y Jeong KJ Dong J de Groot JF . Periostin (POSTN) regulates tumor resistance to antiangiogenic therapy in glioma models. Mol Cancer Ther (2016) 15(9):2187–97.10.1158/1535-7163.MCT-15-0427
104
Xiao ZM Wang XY Wang AM . Periostin induces chemoresistance in colon cancer cells through activation of the PI3K/Akt/survivin pathway. Biotechnol Appl Biochem (2015) 62(3):401–6.10.1002/bab.1193
105
Li B Wang L Chi B . Upregulation of periostin prevents P53-mediated apoptosis in SGC-7901 gastric cancer cells. Mol Biol Rep (2013) 40(2):1677–83.10.1007/s11033-012-2218-3
106
Gonzalez HE Gujrati M Frederick M Henderson Y Arumugam J Spring PW et al Identification of 9 genes differentially expressed in head and neck squamous cell carcinoma. Arch Otolaryngol Head Neck Surg (2003) 129(7):754–9.10.1001/archotol.129.7.754
107
Bakhtyar N Wong N Kapoor A Cutz JC Hill B Ghert M et al Clear cell renal cell carcinoma induces fibroblast-mediated production of stromal periostin. Eur J Cancer (2013) 49(16):3537–46.10.1016/j.ejca.2013.06.032
108
Chuanyu S Yuqing Z Chong X Guowei X Xiaojun Z . Periostin promotes migration and invasion of renal cell carcinoma through the integrin/focal adhesion kinase/c-Jun N-terminal kinase pathway. Tumour Biol (2017) 39(4):1010428317694549.10.1177/1010428317694549
109
Kikuchi Y Kunita A Iwata C Komura D Nishiyama T Shimazu K et al The niche component periostin is produced by cancer-associated fibroblasts, supporting growth of gastric cancer through ERK activation. Am J Pathol (2014) 184(3):859–70.10.1016/j.ajpath.2013.11.012
110
Kotobuki Y Yang L Serada S Tanemura A Yang F Nomura S et al Periostin accelerates human malignant melanoma progression by modifying the melanoma microenvironment. Pigment Cell Melanoma Res (2014) 27(4):630–9.10.1111/pcmr.12245
111
Orecchia P Conte R Balza E Castellani P Borsi L Zardi L et al Identification of a novel cell binding site of periostin involved in tumour growth. Eur J Cancer (2011) 47(14):2221–9.10.1016/j.ejca.2011.04.026
112
Fukuda K Sugihara E Ohta S Izuhara K Funakoshi T Amagai M et al Periostin is a key niche component for wound metastasis of melanoma. PLoS One (2015) 10(6):e0129704.10.1371/journal.pone.0129704
113
Kim CJ Sakamoto K Tambe Y Inoue H . Opposite regulation of epithelial-to-mesenchymal transition and cell invasiveness by periostin between prostate and bladder cancer cells. Int J Oncol (2011) 38(6):1759–66.10.3892/ijo.2011.997
114
Michaylira CZ Wong GS Miller CG Gutierrez CM Nakagawa H Hammond R et al Periostin, a cell adhesion molecule, facilitates invasion in the tumor microenvironment and annotates a novel tumor-invasive signature in esophageal cancer. Cancer Res (2010) 70(13):5281–92.10.1158/0008-5472.CAN-10-0704
115
Malanchi I Santamaria-Martínez A Susanto E Peng H Lehr H-A Delaloye J-F et al Interactions between cancer stem cells and their niche govern metastatic colonization. Nature (2011) 481:85.10.1038/nature10694
116
Wong GS Lee JS Park YY Klein-Szanto AJ Waldron TJ Cukierman E et al Periostin cooperates with mutant p53 to mediate invasion through the induction of STAT1 signaling in the esophageal tumor microenvironment. Oncogenesis (2013) 2:e59.10.1038/oncsis.2013.17
117
Yan S Han X Xue H Zhang P Guo X Li T et al Let-7f inhibits glioma cell proliferation, migration, and invasion by targeting periostin. J Cell Biochem (2015) 116(8):1680–92.10.1002/jcb.25128
118
Zhou W Ke SQ Huang Z Flavahan W Fang X Paul J et al Periostin secreted by glioblastoma stem cells recruits M2 tumour-associated macrophages and promotes malignant growth. Nat Cell Biol (2015) 17(2):170–82.10.1038/ncb3090
119
Shao R Bao S Bai X Blanchette C Anderson RM Dang T et al Acquired expression of periostin by human breast cancers promotes tumor angiogenesis through up-regulation of vascular endothelial growth factor receptor 2 expression. Mol Cell Biol (2004) 24(9):3992–4003.10.1128/MCB.24.9.3992-4003.2004
120
Vardaki I Ceder S Rutishauser D Baltatzis G Foukakis T Panaretakis T . Periostin is identified as a putative metastatic marker in breast cancer-derived exosomes. Oncotarget (2016) 7(46):74966–78.10.18632/oncotarget.11663
121
Qin X Yan M Zhang J Wang X Shen Z Lv Z et al TGFbeta3-mediated induction of periostin facilitates head and neck cancer growth and is associated with metastasis. Sci Rep (2016) 6:20587.10.1038/srep20587
122
Kudo Y Iizuka S Yoshida M Nguyen PT Siriwardena SB Tsunematsu T et al Periostin directly and indirectly promotes tumor lymphangiogenesis of head and neck cancer. PLoS One (2012) 7(8):e44488.10.1371/journal.pone.0044488
123
Liu Y Gao F Song W . Periostin contributes to arsenic trioxide resistance in hepatocellular carcinoma cells under hypoxia. Biomed Pharmacother (2017) 88:342–8.10.1016/j.biopha.2017.01.052
124
Tse JC Kalluri R . Mechanisms of metastasis: epithelial-to-mesenchymal transition and contribution of tumor microenvironment. J Cell Biochem (2007) 101(4):816–29.10.1002/jcb.21215
125
Ye X Weinberg RA . Epithelial-mesenchymal plasticity: a central regulator of cancer progression. Trends Cell Biol (2015) 25(11):675–86.10.1016/j.tcb.2015.07.012
126
Kim DH Xing T Yang Z Dudek R Lu Q Chen YH . Epithelial mesenchymal transition in embryonic development, tissue repair and cancer: a comprehensive overview. J Clin Med (2017) 7(1):1–25.10.3390/jcm7010001
127
Wang L Yang F Jia LT Yang AG . Missing links in epithelial-mesenchymal transition: long non-coding RNAs enter the Arena. Cell Physiol Biochem (2017) 44(4):1665–80.10.1159/000485766
128
Brabletz T Kalluri R Nieto MA Weinberg RA . EMT in cancer. Nat Rev Cancer (2018) 18(2):128–34.10.1038/nrc.2017.118
129
Yan W Shao R . Transduction of a mesenchyme-specific gene periostin into 293T cells induces cell invasive activity through epithelial-mesenchymal transformation. J Biol Chem (2006) 281(28):19700–8.10.1074/jbc.M601856200
130
Aukkarasongsup P Haruyama N Matsumoto T Shiga M Moriyama K . Periostin inhibits hypoxia-induced apoptosis in human periodontal ligament cells via TGF-beta signaling. Biochem Biophys Res Commun (2013) 441(1):126–32.10.1016/j.bbrc.2013.10.027
131
Lee YJ Kim IS Park SA Kim Y Lee JE Noh DY et al Periostin-binding DNA aptamer inhibits breast cancer growth and metastasis. Mol Ther (2013) 21(5):1004–13.10.1038/mt.2013.30
132
Kyutoku M Taniyama Y Katsuragi N Shimizu H Kunugiza Y Iekushi K et al Role of periostin in cancer progression and metastasis: inhibition of breast cancer progression and metastasis by anti-periostin antibody in a murine model. Int J Mol Med (2011) 28(2):181–6.10.3892/ijmm.2011.712
133
Zhu M Saxton RE Ramos L Chang DD Karlan BY Gasson JC et al Neutralizing monoclonal antibody to periostin inhibits ovarian tumor growth and metastasis. Mol Cancer Ther (2011) 10(8):1500–8.10.1158/1535-7163.MCT-11-0046
Summary
Keywords
periostin, matricellular proteins, cancer, microenvironment, biomarker, cancer hallmarks
Citation
González-González L and Alonso J (2018) Periostin: A Matricellular Protein With Multiple Functions in Cancer Development and Progression. Front. Oncol. 8:225. doi: 10.3389/fonc.2018.00225
Received
14 February 2018
Accepted
30 May 2018
Published
12 June 2018
Volume
8 - 2018
Edited by
Gabriel Leprivier, Heinrich Heine Universität Düsseldorf, Germany
Reviewed by
Leonardo Freire-de-Lima, Universidade Federal do Rio de Janeiro, Brazil; Edmund Ui-Hang Sim, Universiti Malaysia Sarawak, Malaysia
Updates
Copyright
© 2018 González-González and Alonso.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Javier Alonso, fjalonso@isciii.es
Specialty section: This article was submitted to Molecular and Cellular Oncology, a section of the journal Frontiers in Oncology
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.