 Kaifee Arman1,2*
Kaifee Arman1,2* Tarik Möröy1,2,3*
Tarik Möröy1,2,3*- 1Institut de recherches cliniques de Montréal, Montreal, QC, Canada
- 2Division of Experimental Medicine, McGill University, Montreal, QC, Canada
- 3Département de microbiologie, infectiologie et immunologie, Université de Montréal, Montreal, QC, Canada
The human genome project revealed the existence of many thousands of long non-coding RNAs (lncRNAs). These transcripts that are over 200 nucleotides long were soon recognized for their importance in regulating gene expression. However, their poor conservation among species and their still controversial annotation has limited their study to some extent. Moreover, a generally lower expression of lncRNAs as compared to protein coding genes and their enigmatic biochemical mechanisms have impeded progress in the understanding of their biological roles. It is, however, known that lncRNAs engage in various kinds of interactions and can form complexes with other RNAs, with genomic DNA or proteins rendering their functional regulatory network quite complex. It has emerged from recent studies that lncRNAs exert important roles in gene expression that affect many cellular processes underlying development, cellular differentiation, but also the pathogenesis of blood cancers like leukemia and lymphoma. A number of lncRNAs have been found to be regulated by several well-known transcription factors including Myelocytomatosis viral oncogene homolog (MYC). The c-MYC gene is known to be one of the most frequently deregulated oncogenes and a driver for many human cancers. The c-MYC gene is very frequently activated by chromosomal translocations in hematopoietic cancers most prominently in B- or T-cell lymphoma or leukemia and much is already known about its role as a DNA binding transcriptional regulator. Although the understanding of MYC’s regulatory role controlling lncRNA expression and how MYC itself is controlled by lncRNA in blood cancers is still at the beginning, an intriguing picture emerges indicating that c-MYC may execute part of its oncogenic function through lncRNAs. Several studies have identified lncRNAs regulating c-MYC expression and c-MYC regulated lncRNAs in different blood cancers and have unveiled new mechanisms how these RNA molecules act. In this review, we give an overview of lncRNAs that have been recognized as critical in the context of activated c-MYC in leukemia and lymphoma, describe their mechanism of action and their effect on transcriptional reprogramming in cancer cells. Finally, we discuss possible ways how an interference with their molecular function could be exploited for new cancer therapies.
Introduction
The knowledge gained by exploring the human genome is expanding every day thanks to high-throughput RNA sequencing (RNA-Seq) and other next generation sequencing technologies. One outcome was the discovery of thousands of non-coding transcripts (1), which were once considered transcriptional noise or “junk” (2, 3). It has now been recognized that only 2% of all transcribed regions of the human genome represent protein coding genes while the rest are non-protein coding transcripts with long non-coding RNAs (lncRNAs) being in the majority (4–7). LncRNAs are defined as transcripts longer than 200 nucleotides in length that lack open reading frames with the capacity to encode more than 100 amino acids (8, 9). Most if not all of these lncRNAs are transcribed by RNA polymerase II, are 5′ capped and polyadenylated at their 3′ ends (10). LncRNAs are classified according to their transcriptional origin and can be associated with enhancers or promoters. An additional classification is made according to the position of their genes with respect to neighboring protein-coding genes, which groups them into intronic lncRNAs, exonic/sense lncRNAs, antisense lncRNAs, intergenic lncRNAs, and bidirectional or divergent lncRNAs (11). Recent studies have shown that lncRNAs are implicated in almost all types of cancer, functioning either as tumor suppressors or oncogenes [reviewed in (12)], but in most cases a precise molecular mechanism remains yet to be discovered. Nevertheless, the relevance of lncRNAs in malignant transformation and several other important biological processes has been recognized and has placed these molecules in the center of attention (13, 14).
It is now known that lncRNAs can interact with other molecules and form RNA-RNA, RNA-DNA, or RNA-protein complexes, which renders their functional network quite complex. Although many questions remain, a number of molecular mechanisms have been unveiled that can explain how lncRNAs function. LncRNAs show interactions with chromatin modifiers such as the BRG1/BAF or the SWI/SNF complexes [reviewed in (15)] or act as modulators of protein activity or as enzyme cofactors [reviewed in (16)]. They also function as super enhancer RNAs (eRNAs) affecting multiple genes in trans (17) or by competing with miRNAs for binding to their targets (competing endogenous RNAs or ceRNAs) (18). Moreover, lncRNAs are implicated in transcriptional and post transcriptional regulation of gene expression (19) and may have direct interactions with R-loops and triple helices (20). LncRNAs often exhibit tissue- and cell-type specificity (8, 21), a feature which makes them excellent candidates for biomarkers of selected human cancers (22–25). The localization of lncRNAs within the cell closely correlates with their mode of action. LncRNAs localized in the nucleus can act in cis by controlling the expression of neighboring genes or in trans by regulating gene expression on other chromosomes or remote loci. Cytoplasmic lncRNAs have a direct regulatory function on gene expression post-transcriptionally and affect for instance mRNA stability and mRNA translation or the sequestering of proteins or miRNAs (26).
Long non-coding RNAs can directly or indirectly regulate different pathways in cancer (27), and their relevance for hematological malignancies in particular has been established at a rapid pace (28–37). Well known transcription factors and or proto-oncoproteins such as c- Myelocytomatosis viral oncogene homolog (MYC), Notch1, beta-catenin, or RAS regulate lncRNAs or are being regulated by lncRNAs in different hematological disorders (38–43), [reviewed in (29, 30, 44)]. One of the most frequently activated proto-oncogenes in human cancers is the c-MYC gene (MYC henceforth). MYC is a helix-loop-helix, leucine zipper (HLH-LZ) transcription factor, which dimerizes with MAX, a smaller HLH-LZ protein and MYC/MAX heterodimers bind to cognate sites containing so called E-box sequences in gene promoters and enhancers. The MYC/MAX complex has been reported to activate gene expression, albeit more recent reports also indicate that MYC can have a function in gene repression (45–47). MYC plays a significant role in many human cancers by regulating several cellular processes including cell proliferation, cell differentiation, metabolism, apoptosis, angiogenesis, and genomic stability (48–50). One of the first direct links between MYC and human cancers was the discovery of its role in Burkitt type B cell lymphoma. Here, a t(8;14) chromosomal translocation juxtaposes the coding part of the MYC gene to immunoglobulin μ heavy chain locus and places it under the control of the Eμ enhancer, which leads to a constitutive, transcriptional activation of MYC (51, 52). Besides translocations, several other mechanisms like gene amplifications and epigenetic alterations of MYC and posttranslational regulation of the MYC protein, in particular its phosphorylation, lead to its constitutive activation in many human cancers including hematological neoplasms (50, 53–61). MYC is known to target many protein-coding and lncRNA genes in different cancers (62–69), but a number studies have also pointed out that the reverse is also true and that lncRNAs regulate MYC activity at the post-translational level (70, 71).
It is quite pellucid that the interrelation between MYC and lncRNAs plays a critical role in blood cancers such as leukemia and lymphoma. Although much is known about MYC on one hand and lncRNA on the other in cancers, their significance of their regulatory interaction with each other in hematological malignancies is still largely unknown. In this review, we focus on the known functional interactions between MYC and lncRNAs in order to illustrate the intensive crosstalk that occurs between both in many hematological malignancies (Table 1). Given the importance of MYC for human cancers, we discuss the implications that these interactions have for new therapeutic strategies against human blood cancers.
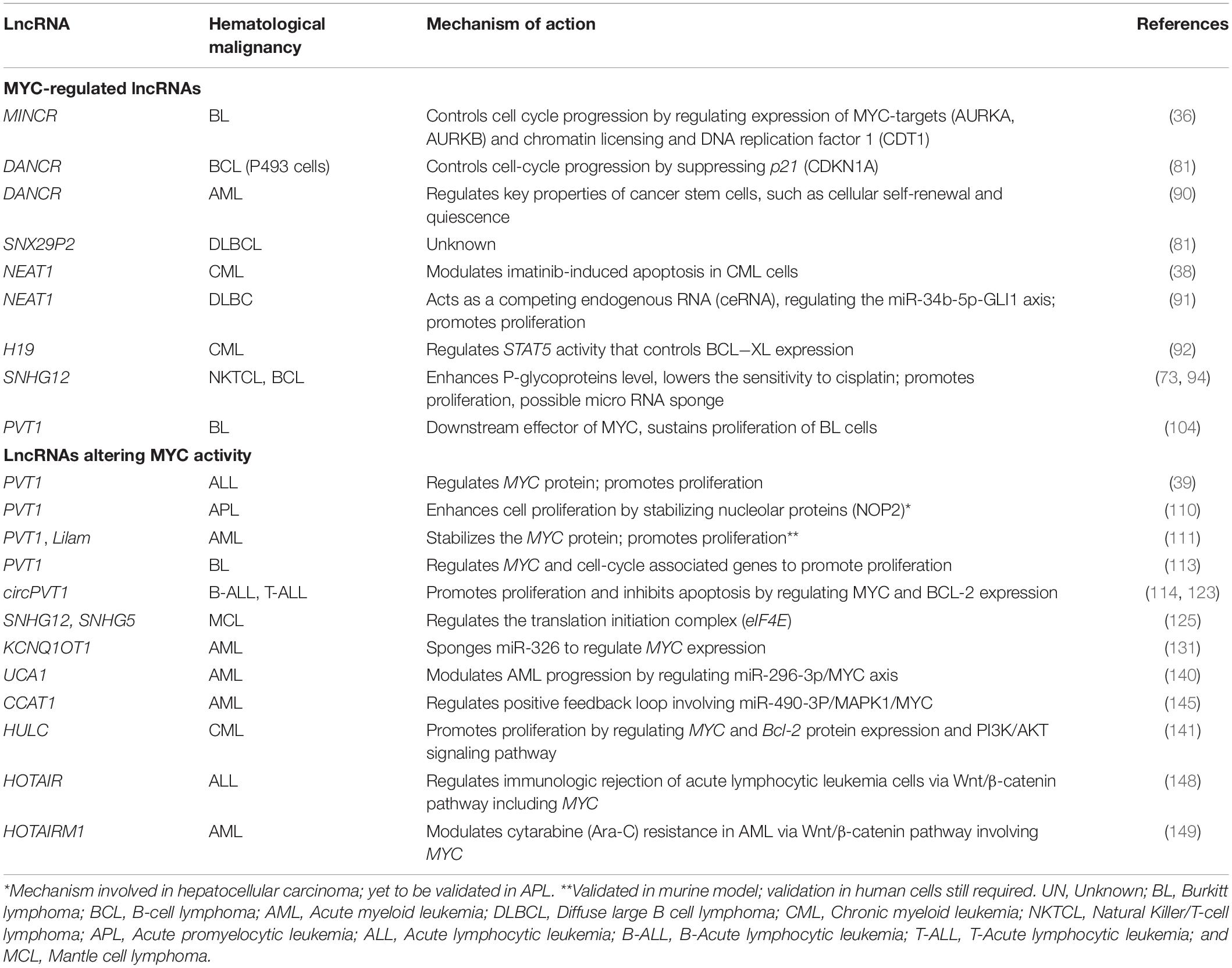
Table 1. Summary of lncRNAs and their crosstalk with MYC involved in different hematological malignancies.
MYC Regulated lncRNAs
LncRNAs as Direct MYC Targets Are Found in Many Hematopoietic Cancers
Kluiver and his group (72) used the immortalized B cell line P493-6, which was engineered to express MYC in a tetracycline-repressible manner, and primary B-cell lymphoma samples from patients to show that lncRNAs are a main component of the MYC-regulated transcriptional program comparable to regular mRNAs and micro RNAs (miRNA). This study demonstrated also that both MYC-induced and MYC-repressed lncRNA genes are significantly enriched for MYC binding sites at their promoters, suggesting that lncRNA genes are indeed direct MYC targets. Expression profiling of lncRNAs using a custom-designed microarray allowed the identification of 1244 lncRNA loci that were candidates to be regulated by MYC at least in this cellular model. The authors validated MYC induced upregulation of 10 lncRNAs. Furthermore, the analysis of lncRNA expression in P493-6 B-cell lymphoma cells with either high or low MYC levels led to the identification of 498 MYC-regulated lncRNAs. These lncRNAs were responsive to the inactivation of MYC in BL cell lines, further supporting their relevance for MYC-driven B-cell lymphomas (72).
Further analysis by RNA-sequencing of P493-6 B-cells (73) revealed 534 MYC regulated lncRNAs, 296 upregulated and 238 downregulated, in response to MYC overexpression. Publicly available ChIP-seq data of P493-6 cells demonstrated that MYC-responsive lncRNA genes were occupied by MYC near their transcription start sites (TSS) further supporting the notion that these lncRNAs are direct MYC targets. Nuclear run-on (NRO) analysis on a few selected lncRNAs also provided evidence for their direct transcriptional regulation by MYC. Some of these lncRNA genes (Snhg12, Snhg17, and Mir17hg) have murine homologues and the occupation of their promoters by MYC can be confirmed by interrogating publicly available ChIP-seq data (74) from normal, pre-lymphomatous, and fully malignant B cells from Eμ-Myc transgenic animals (Figures 1A–D). These animals carry a human MYC transgene, which is under the control of the strong immunoglobulin heavy chain enhancer (Eμ) and therefore express high levels of MYC in B cells (Figures 1A,B). The constitutive MYC expression drives an expansion of B cells in the bone marrow which subsequently turns to an overt, aggressive B-cell lymphoma (Figure 1A). Binding of MYC near the promoters of Snhg12, Mir17hg is readily detected and increases in B cells from the pre-lymphomatous and the fully malignant stage (lymphoma; Figures 1C,D). Other studies have confirmed this (73), which also suggests that the regulation of lncRNAs by MYC is conserved between species and is not restricted to human cells.
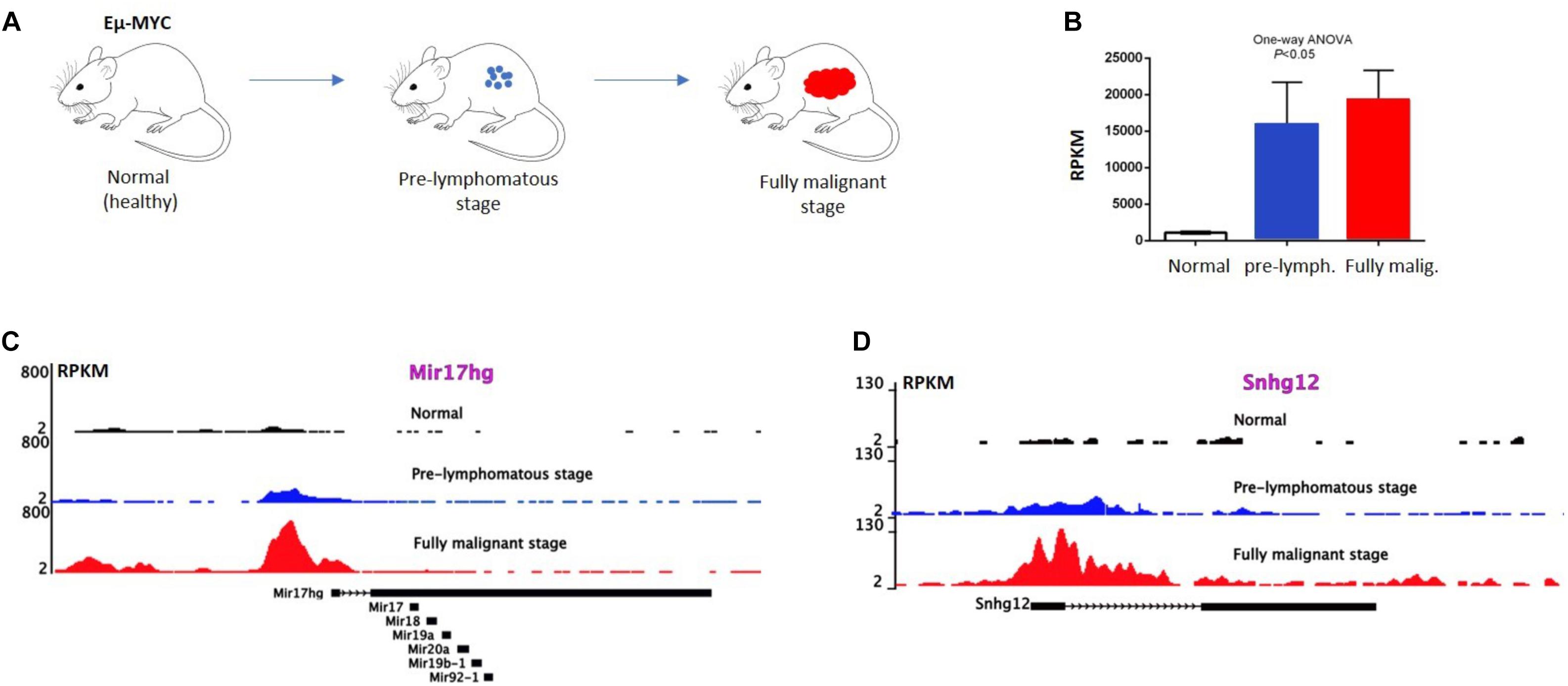
Figure 1. MYC occupy lncRNA gene promoters in Eμ-MYC transgenic B-cells. (A) Eμ MYC transgenic model, in which the tumor progression takes place. Here, a pre-lymphomatous stage exists prior to the manifestation of a fully malignant lymphoma. (B) Expression of MYC in the indicated B cells revealed by RNA-seq data. MYC expression in B cells positively correlates with the tumor progression at different stages of B-cell lymphoma development. Error bars, SD. P value < 0.05; One-way Anova analysis. (C,D) ChIP-seq analysis showing MYC binding to the promoter of Snhg12 and Mir17hg, respectively, in normal (wild type) B-cells, in B-cells from a pre-lymphomatous animal and in B-lymphoma cells from Eμ MYC mice. Correlation of MYC binding to the promoters of lncRNA genes and MYC expression level in Eμ MYC transgenic mice (data extracted from GEO GSE51011).
The Case of MINCR
To further investigate the lncRNAs that are being regulated by MYC in blood cancers, several studies have chosen different strategies. The group of Ingram Iaccarino (36) analyzed RNA-seq data of 91 patients representing several B-cell lymphoma subgroups including Burkitt lymphoma, Diffuse large B-cell lymphoma (DLBCL) and Follicular lymphoma, and compared them with data from controls such as normal germinal center B cells. They also used RNA-seq data from two different MYC-inducible cell lines, namely hT-RPE-MycER cells, which are epithelial cells expressing a MycER fusion protein (75, 76) and P493-6 cells, with the tetracycline-repressible MYC allele (77, 78). This led to the discovery of 13 lncRNAs that were differentially expressed in Burkitt lymphoma carrying the t(8:14) Ig/MYC translocation. These lncRNAs were concordantly regulated by MYC in the model cell lines. Out of these 13 lncRNAs, MINCR (MYC-induced long non-coding RNA) showed a significant correlation with MYC expression in MYC-positive lymphomas. An siRNA mediated knockdown of MINCR in hT-RPE-MycER cells led to a reduction in cellular proliferation which was MYC independent indicating that MINCR acts downstream of MYC. RNA-seq followed by differential gene expression analysis revealed a significant enrichment of genes encoding regulators of cell cycle progression among genes that were downregulated after an RNAi mediated MINCR knockdown, which was in agreement with the observed defect in cell cycle progression.
While these results seemed to demonstrate that MINCR is a direct effector of MYC, several known MYC target genes such as Aurora kinases A and B (AURKA, AURKB), Cyclin dependent kinase 2 (CDK2), and Non-SMC condensing I complex subunit D2 (NCAPD2) were found downregulated upon MINCR knockdown, suggesting that MINCR can regulate MYC targets. Moreover, the promoters of 125 cell cycle associated genes that were down-regulated upon MINCR knockdown, showed a significant enrichment of E-boxes, i.e., sites where MYC/Max complexes contact DNA. ChIP-qPCR experiments indeed showed that MYC binding was reduced at all those promoter regions when the expression of MINCR was reduced indicating that silencing of MINCR might dampen MYC’s ability to engage a transcriptionally active complex at target gene promoters. However, given the complex nature of this regulatory interaction, it is presently a matter of debate whether MINCR is a direct MYC-induced lncRNA or whether MINCR can act back on MYC regulated genes; or whether both situations occur depending on cellular context (36, 79, 80). Indeed, another study has put into question whether the regulatory link between MINCR and MYC is universal, since it was not reproducible using data generated from an independent set of Burkitt lymphoma cells (79). In reply to this criticism, Doose et al. defended MINCR to be a MYC-induced lncRNA by highlighting several lines of evidence: MINCR is up-regulated by MYC in cells that contain MYC-inducible constructs; MYC occupies the promoter of MINCR and expression of MINCR and MYC correlates in a number of different MYC positive B-cell lymphomas (80). The same authors further stated that the heterogeneity typical for most cancers and cell lines, in terms of patient age, ethnicity, and effect of chemotherapy might be the reason for the lack of a general correlation between MINCR and MYC in cancer cell lines that are established in culture since long period of time (80). Along this line, the expression of MYC and MINCR did not always correlate in normal tissues and was not observed in ovarian carcinomas, chronic lymphocytic leukemia or pancreatic endocrine neoplasm (80) further supporting that the link between MYC and MINCR is context dependent and that a full clarification of the mechanistic relationship between both genes requires further experimentation.
MYC Regulated lncRNAs as New Targets for Anti-MYC Cancer Therapies
DANCR and Acute Myeloid Leukemia
In a similar recent study by Lu et al. (81), 545 MYC regulated lncRNAs were identified via RNA-seq on P493-6 cells; many of them (238 out of 545 lncRNAs) were encoded by intergenic sequences and were found upregulated at very high levels in the malignant hematopoietic cells. Here, the lncRNA ENSG00000198106 (SNX29P2) was found to be specifically expressed in DLBCL. Four out of 238 MYC regulated intergenic lncRNAs were highly expressed in nearly all CCLE (Cancer Cell Line Encyclopedia) cell lines and were significantly positively correlated with MYC expression levels. One of which, the lncRNA ENSG00000226950, that has attracted the attention of several groups had previously been identified as a non-coding RNA associated with enhancer of zeste homolog 2 and was found to repress the runt related transcription factor 2 gene (81) and was called DANCR (Differentiation Antagonizing Non-Protein Coding RNA). The DANCR gene is bound by MYC when MYC is induced in the P493-6 cells in a time-dependent manner which is consistent with the findings of Hart et al. (73) and other studies that followed DANCR expression in several tumors (82–89). Of interest here is the finding that DANCR plays a role in Acute Myeloid Leukemia (AML) (90). AML is a heterogeneous leukemia with a hierarchical cellular organization, driven by a population of so-called leukemia stem cells (LSCs) that have an abnormally robust self-renewal capacity and increased chemotherapy resistance. RNA-seq on cytogenetically normal AML patients (CN-AML, i.e., lacking chromosomal aberrations) demonstrated that DANCR was specifically expressed in LSCs and the knockdown of DANCR resulted in the downregulation of MYC expression (90). These observations and the finding that disrupting DANCR in a murine AML mouse model prolonged the survival of the animals after serial transplantation owing to a lower self-renewal capacity and dormancy of leukemic cells made a strong case for an important functional role of this lncRNA in the maintenance or progression of AML (90). The features of DANCR in hematological malignancies makes this lncRNA a good candidate for a potential therapeutical application. How to target DANCR in AML will depend, however, on more insight into its precise molecular function in hematological disorders.
NEAT and H19 in CML
The lncRNA NEAT1 (Nuclear Enriched Abundant Transcript 1) serves as a scaffold for so called paraspeckles, that are found in the nucleus at regions not occupied by chromatin. The function of paraspeckles in not well known, but their formation depends on the presence of RNA Polymerase II and is therefore linked to transcription. It was shown that NEAT1 expression is regulated by MYC in DLBCL (91) and Qian and colleagues demonstrated that NEAT1 can drive B cell proliferation and -lymphomagenesis through the miR-34b-5p-GLI1 axis and silencing of NEAT1 dampened cell proliferation and facilitated apoptosis (91). A report from Zeng et al., indicated that NEAT1 expression is downregulated in primary CML cells, but that its expression was rescued in K562 CML cell line when the expression or activity of the BCR-ABL kinase, that typically is present in CML cells as a consequence of the t(9:22) translocation, is blocked (38). In addition, the findings suggested that NEAT1 which is regulated by MYC modulates imatinib-induced apoptosis in CML cells (38). A model was proposed in which pathways activated by BCR-ABL can increase MYC expression which leads to repression of NEAT1 transcription. MYC knockdown upregulated NEAT1 expression, and imatinib treatment of the MYC knockdown cells increased NEAT1 expression further (38). Although CML can be well treated with Imatinib, resistance to this drug occurs and represents a major problem in the management of this leukemia. Of interest in this context is the observation that knockdown of NEAT1 renders K562 CML cells more sensitive to Imatinib and proteasome inhibitors, since combinatorial treatment leads to accelerated apoptosis of K562 cells. Understanding the link between BCR-ABL, NEAT1, MYC, and the sensitivity of CML cells to Imatinib will provide further insight into the mechanisms of Imatinib resistance and ultimately lead to a way to circumvent it.
Similar to NEAT1, H19, a maternal lncRNA, is expressed in CML cells with the t(9:22) translocation and its expression depends on the presence of the BCR-ABL fusion protein, since a knockdown of BCR−ABL expression led to a downregulation of H19 expression in K562 cells (92). Treatment with imatinib also decreased the level of H19 in K562 cells, further confirming that the expression of H19 is BCR−ABL kinase dependent. Again, as for NEAT1, knockdown of H19 significantly increased the apoptosis in K562 cells following imatinib treatment pointing to the possibility of a way to enhance the drug’s effectiveness. Further studies into the underlying mechanisms demonstrated that H19 may have an impact on leukemic cell survival by regulating STAT5 activity, an anti-apoptotic protein that stimulates expression of pro-survival factors such as BCL-XL. In vivo experiments involving subcutaneous injection of K562 cells engineered with a stable knockdown of H19 into mice demonstrated the tumorigenic role of H19, since a significant decrease of tumor growth was observed (92). Moreover, the expression of H19 was found to be MYC dependent in K562 leukemic cells. Treatment with imatinib resulted in a decreased level of MYC mRNA similar to H19. A knockdown of MYC downregulated H19 expression while the ectopic over-expression of MYC upregulated H19 levels in K562 cells, regardless whether they were treated with imatinib or not, supporting the notion that the expression of H19 is MYC-dependent (92). It is therefore thought that the disruption of H19 expression along with imatinib treatment would be a promising therapeutic strategy to combat CML, as the combination of these two could trigger leukemic cells to undergo apoptosis. However, as for the action of NEAT1 further studies on H19 are required to better understand this combination strategy in order to have continuous positive effect with minimal imatinib resistance in CML.
SNHG12 in Natural Killer/T-Cell Lymphoma
The lncRNA SNHG12 (small nucleolar host gene 12), also known as LINC00100/ASLNC04080 is located on chromosome 1 in humans and is suspected to play a pivotal role in a number of cancers including blood cancer [reviewed in (93)]. It was noticed that the expression of SNHG12 was higher in Natural Killer/T-cell lymphoma (NKTCL) tissues as compared to controls, in this case reactive hyperplasia of lymph node (RHLN) tissues. Moreover, SNHG12 showed more expression in NKTCL tissues of advanced stages (3 and 4) compared to earlier stages 1 and 2, indicating its positive correlation with clinical grading (stage in 3/4) of NKTCL. MYC was shown to act directly on the SNHG12 gene and the expression of MYC positively correlated with the expression of SNHG12 in NKTCL (94). Both MYC and SNHG12 acted in a similar fashion in NKTCL by promoting proliferation, enhancing the level of P-glycoproteins (P-gp) that are linked to multi drug resistance (MDR) proteins and inhibiting the sensitivity to cisplatin (CCDP) (94). Besides, MYC was found to bind directly to the promoter of SNHG12 and to upregulate its expression. Rescue experiments further demonstrated that the overexpression of SNHG12 was able to partially rescue the negative effects of MYC knockdown on drug resistance and on cell proliferation (94). The results found by Zhu et al. indicated that the regulation of SNHG12 by MYC might be an important factor for the chemotherapy resistance for NKTCL (94). Therefore, loss of function of SNHG12 could possibly overcome or defer tolerance of NKTCL to therapy, which would establish the lncRNA SNHG12 as a therapeutic target for controlling MDR in NKTCL. RNA-seq data from a study using with P493-6 revealed that SNHG12 can also be upregulated by MYC in human B cells confirming the findings in NKTCL (73). A positive correlation between MYC and Snhg12 was also seen in the Eμ-MYC transgenic lymphoma model (see Figure 1) adding support to the notion that Snhg12 might have important roles in other hematopoietic malignancies besides NKTCL. An overview of all the above described MYC-regulated lncRNAs in different hematopoietic malignancies is shown in Figure 2.
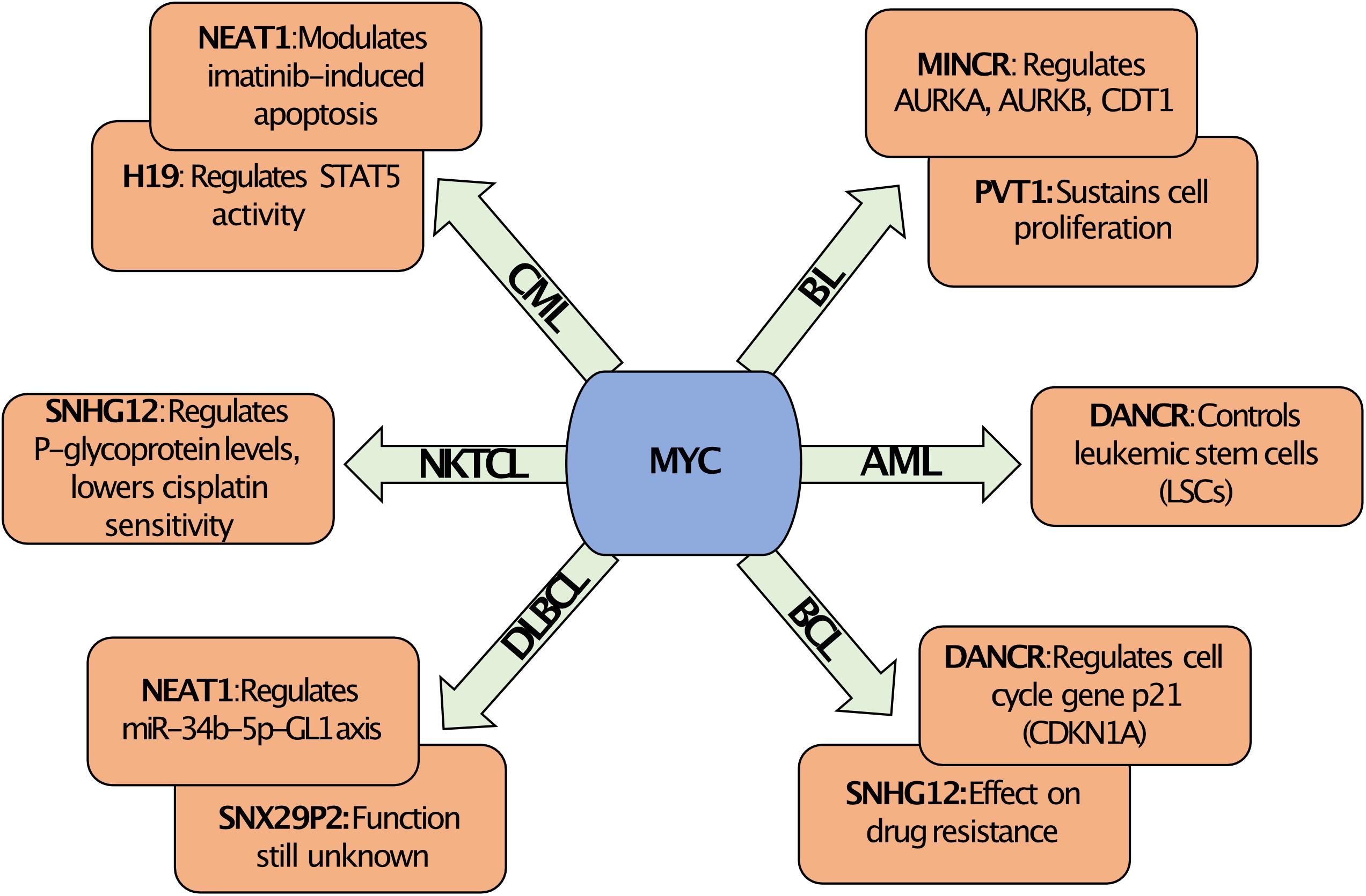
Figure 2. MYC regulated lncRNAs. Overview of MYC regulated lncRNAs along with their known biological function in different hematological malignancies. The regulation of the expression of specific lncRNAs by MYC is context dependent and is specific for the hematological malignancy. For example, MYC regulates DANCR in AML, which affects leukemic stem cells, but in BCL the regulation of DANCR by MYC targets the function of the negative cell cycle regulator p21 (CDKN1A). Similarly, MYC can regulate the expression of NEAT1 in CML or DLBCL, but this affects Imatinib response or the miR-34b-5p-GL1 axis, respectively. In contrast, MYC can regulate SNHG12 in BCL or in NKTCL, which in both cases can alter drug resistance. Abbreviations: BL, Burkitt lymphoma; BCL, B-cell lymphoma; AML, Acute myeloid leukemia; DLBCL, Diffuse large B cell lymphoma; CML, Chronic myeloid leukemia; and NKTCL, Natural Killer/T-cell lymphoma.
LncRNAs That Alter MYC Activity
Since a constitutive activation or overexpression of MYC plays an exceptionally important role in many, if not all, hematological malignancies, a tight regulation of this proto-oncoprotein in normal cells is essential to avoid a derailment of physiological processes such as cell cycle progression, cell death, or metabolic pathways (49, 95). To achieve this, the expression and the activity of MYC is controlled by multiple mechanisms beyond the control of MYC gene expression at the transcriptional level. The multitude of regulatory mechanisms include post-translational modifications that alter MYC’s stability and half-life and the interaction with a large number of partner proteins that modify MYC’s activity and also its capacity to bind DNA (96, 97). Genes for several non-coding RNAs including both miRNA and lncRNAs are located in the vicinity of the MYC gene locus where they play important roles in the regulation of MYC (98–100) in both normal cells and in hematological malignancies. Here, we discuss in more detail those lncRNAs that regulate MYC activity in hematological malignancies. One of the best studied lncRNAs that exerts such activities is PVT1.
LncRNAs Regulating MYC as New Targets for Blood Cancer Therapies
The Link Between MYC and PVT1
Plasmacytoma Variant Translocation 1 (PVT1) is a lncRNA that is known since a long time and was linked to MYC shortly after its discovery. The gene encoding PVT1 is located on the long arm of chromosome 8q24 in a region about 55 kb distal to the MYC gene (Figure 3) and, very similar to MYC, is frequently involved in translocations occurring in Burkitt lymphoma (101, 102). PVT1 functions in close coordination with MYC and the intricate relationship between both genes was recently explained in a new study (70), which showed a PVT1 dependence for those cancers that have a copy-number increase of MYC. It is known that MYC is consistently co-gained with the adjacent genes PVT1, CCDC26, and GSDMC that lie within a region of about 2 Mb (70). To explore whether a low copy-number gain of these genes promotes tumorigenesis or not, three strains of transgenic mice were created; one with single copy of MYC alone, another one with the region containing PVT1, CDC26, and GSDMC and third line that carried the entire 2 Mb syntenic region including MYC, PVT1, CDC26, and GSDMC. All three lines showed normal phenotypes with no developmental abnormalities. However, a single copy of MYC was insufficient to accelerate MMTV-Neu driven tumorigenesis and a combination of both MYC and PVT1 was required (70) indicating a collaborative relationship between both genes (Figure 3).
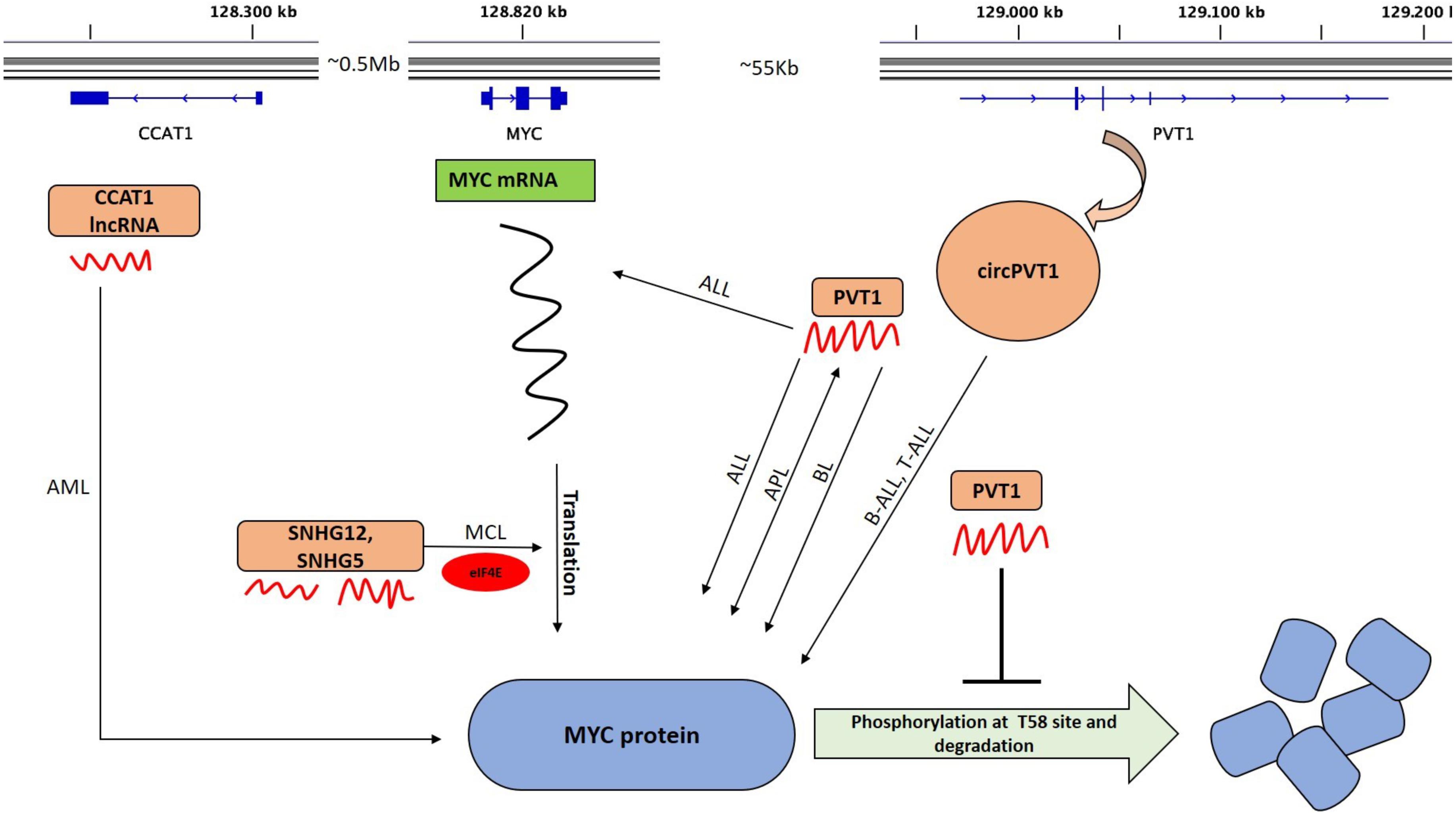
Figure 3. LncRNAs that alter MYC activity. The upper part shows the graphical representation of the genomic location of MYC, PVT1, and CCAT1. The transcribed lncRNAs (curved red structures) regulate MYC at mRNA or protein level and some at both. Those that regulate MYC mRNA translation in different hematological malignancies are indicated. Positive feedback interaction between lncRNA and MYC is also shown (double headed arrow). circPVT1 is transcribed from exon 2 (E2) of PVT1. PVT1 maintains a high MYC protein level and prevents the degradation of MYC by blocking the phosphorylation of Thr58 in MYC. SNHG5 and SNHG12 modulate MYC translation in MCL cells via translation machinery through eIF4E. LncRNA CCAT1 interacts with MYC in AML. BL, Burkitt lymphoma; AML, Acute myeloid leukemia; APL, Acute promyelocytic leukemia; ALL, Acute lymphocytic leukemia; B-ALL, Acute lymphocytic leukemia; and MCL, Mantle cell lymphoma.
Analysis of data from The Cancer Genome Atlas (TCGA) database revealed that a co-gain of both MYC and PVT1 in almost all (98%) tumors showing 8q24 copy-number increases, whereas very few (0.15%) tumors showed an increase in copy number of MYC alone without PVT1. CRISPR Cas9-mediated disruption of PVT1 in MYC dependent HCT116 colon cancer cells significantly reduced its tumorigenic potency suggesting that PVT1 regulates the activity of MYC as an oncogene (70) or even act as an oncogene independently of MYC (103). Several studies provided data supporting a direct regulation of MYC by PVT1, which is discussed further below. However, Carramusa et al. (104) proposed that, in contrast, MYC transcriptionally activates PVT1 and PVT1 is a downstream effector of MYC in Burkitt lymphoma. Their finding that the PVT1 gene locus bears specific E-boxes allowing binding of MYC to bind to the PVT1 promoter region supported this view (104).
The MYC – PVT1 feedback loop mechanism
Since PVT1 also acts as a cis regulatory lncRNA (70, 105–108), the interaction between MYC and PVT1 in different hematological malignancies has to take place via a positive feedback loop (109, 110). To exert its oncogenic activity via MYC, PVT1 prevents the degradation of MYC by hindering its phosphorylation at threonine 58 resulting in increased MYC stability (Figure 3), possibly by a direct binding or interaction with this amino acid residue (70, 107). This causes a feed forward loop since the stabilization of MYC enhances the expression of MYC target genes among them also PVT1, which leads to even more active MYC. Support for this comes from studies with acute promyelocytic leukemia (APL) cell lines, where knockdown of PVT1 resulted in the downregulation of MYC whereas MYC silencing also reduced PVT1 levels, causing reduced proliferation (110). PVT1 also regulates MYC in acute lymphoblastic leukemia (ALL) where PVT1 can act as an oncogene and drives both the development and progression of this type of blood cancer (39). Experiments in Jurkat cells (a human acute T cell lymphoblastic leukemia cell line) showed downregulation of MYC both at mRNA and protein levels upon siRNA mediated knockdown of PVT1 resulting in accelerated apoptosis, slower proliferation rates and a G0/G1 cell cycle arrest. These effects were attributed to the degradation of MYC and BCL2, which increased levels of caspase-3 and the negative cell cycle regulators and tumor suppressor genes CDKN2A and -2B (p16INK4a and p15INK4b) (39). All these data suggest that PVT1 is required for the development of ALL (Figure 3) and successful interference with PVT1 could offer new possibilities for a targeted therapy against ALL (39). Since MYC depends on PVT1 in its ability to drive malignant transformation and is itself difficult to target because it lacks enzymatic activity, several groups have speculated about the validity and feasibility to target PVT1 to weaken or inhibit MYC in tumor cells.
In murine MLL-ENL AML models, depletion of Pvt1 together with two other lncRNAs, called Lilam (leukemia-induced lncRNA affecting Myc) and Pilna (progenitor-induced lncRNA neighboring Ak3), activated a myeloid differentiation program which was reversed by Myc overexpression showing an epistatic relationship between these three lncRNAs and Myc (111). In addition, the PVT1 promoter also has tumor suppressor activity independently of the PVT1 lncRNA. In fact, both PVT1 and MYC promoters compete for four intragenic enhancers in the PVT1 locus. Here, the PVT1 promoter functions as a DNA boundary element, inhibiting MYC from accessing its downstream enhancers and finally hindering the transcription of MYC and impeding its oncogenic functions (112). In another study, knockdown of PVT1 inhibited the proliferation of Burkitt lymphoma cells by arresting the cells in G0/G1 phase which was associated with a reduction of MYC expression and alterations in the expression of cell cycle-associated genes. A cell cycle PCR array showed that 54 genes including CCNG2 (Cyclin G2), RBL2 (Retinoblastoma-like 2), CDKN1A (Cyclin-dependent kinase inhibitor 1A) and others were upregulated while 26 genes such as CCNE1 (Cyclin E1), CCND1 (Cyclin D1) and CDC20 (Cell division cycle 20) were downregulated. The authors concluded that the silencing of PVT1 retarded the proliferation of Raji cells though a downregulation of MYC expression and subsequent alterations in the expression levels of cell cycle-associated genes (113).
Another intriguing property of the PVT1 gene locus is the fact that it also encodes a circular RNA called circPVT1 (Figure 3) with 26 different isoforms (114). The most common isoform which has the highest level of expression in blood cells includes the whole exon 2 of PVT1, which forms a closed loop-like structure (114–116). Circular RNAs (circRNAs) have been identified as a new class of non-coding RNAs which are well conserved, widespread, abundant and are regulated independently of their cognate linear isoform in the eukaryotic transcriptome (115, 117, 118). These are generally formed by back-splicing of pre-mRNA and form a closed loop structure in which their 3′ and 5′ ends are covalently linked conferring them an increased stability and resistance (115, 119, 120). LncRNA PVT1 and circPVT1 regulate their expression independently of each other as they arise from separate individual promoters and have different genome localizations (114, 121, 122). The role of circPVT1 and PVT1 in different hematological malignancies with downstream deregulation of MYC has been very well highlighted in a recent review by Ghetti et al. (114). circPVT1 is well studied in ALL in which the authors demonstrate that circPVT1 (but not PVT1) was specifically highly expressed in human patient samples of ALL but not AML samples as compared to healthy control groups (123). Knockdown of circPVT1 had no effect on its mother gene PVT1 but significantly decreased the protein levels of neighboring genes, MYC and BCL2 suggesting that circPVT1 might enhance BCL2 expression to inhibit MYC mediated apoptosis (123). The authors speculate that circPVT1 might be acting as competing endogenous RNAs (ceRNAs) of miR-let-7 and miR-125 as the expression of these miRNAs were reduced in the ALL patients through the sponging effect of circPVT1 (123). These two miRNAs also target MYC and BCL2 and their lower expression reduces their inhibitory effect on MYC and BCL2 resulting in expansion of ALL (123). The above cited examples indicate that the specific functions of PVT1 and circPVT1 in hematological malignancies are with some certainty caused by the downstream deregulation of MYC. Interference with PVT1 RNAs would therefore have a direct effect on MYC expression, which makes them potential therapeutic targets.
Colon cancer associated transcript1 (CCAT1) is a another lncRNA present along in the same genomic region as PVT1 and MYC (Figure 3). CCAT1 is implicated in multiple solid tumors and is generally viewed as a dominant oncogene. High CCAT1 expression seems to induce a block of myeloid differentiation; a situation which can be seen as a prerogative of myeloid transformation and the emergence of an AML. Furthermore, both CCAT1 and PVT1 expression was found to be over 5 fold higher in t(8;21) positive versus t(8;21) negative AML samples and was associated with clinical parameter indicating higher risk and lower overall survival of AML patients positive for this translocation (124). Similar to PVT1, the promoter of CCAT1 is occupied by MYC and MYC can upregulate CCAT1 expression. It therefore can be inferred that part of MYC’s oncogenic potential is mediated indirectly through the action of CCAT1 and PVT1. These findings support a role of CCAT1 and PVT1 as dominant oncogenes in AML and suggest that interference with the function of these lncRNAs could represent a new therapeutic tool against MYC driven hematological malignancies (124).
LncRNA Acting on the Translation of MYC mRNA
The translation of mRNAs and, in particular, ribosomal entry is regulated in part by a group of proteins called “eukaryotic translation initiation factors” or eIFs. In lymphoma cells, lncRNAs exist that can regulate the translation of the MYC specific mRNA in cooperation with the eukaryotic translation initiation factor 4E (eIF4E). Recently, Mamta and her group identified several other translation-machinery associated lncRNAs in mantle cell lymphoma (MCL) (125). To find lncRNAs associated with the translation initiation complex, RNA Immunoprecipitation and sequencing (RIP-seq) followed by RNA-IP using an anti-eIF4E (translation initiation factor-4E) antibody was performed using samples from MCL patients, normal controls and MCL cell lines (125). The lncRNA SNHG12 which was also shown to be regulated by MYC (see above) and lncRNA SNHG5 turned out to be highly enriched in the precipitates. Knockdown of SNHG12 or SNHG5 significantly increased the protein levels of MYC indicating that these lncRNAs can modulate MYC expression level by altering its translation in MCL (125). It is therefore be concluded that among the bona fide lncRNAs that interact with the translation machinery through eIF4E, i.e., both SNHG5 and SNHG12, can modulate MYC translation in MCL cells (125) and thus strongly suggesting that these lncRNAs regulate the translation of MYC (Figure 3).
LncRNAs and MYC: The Micro RNA Connection
Among the most interesting observations made by studying non-protein coding RNAs was that lncRNAs and miRNAs can compete for shared binding sequences and that this competition affects the expression of specific target genes. In this case, lncRNAs that function as competitive endogenous RNAs are designated “ceRNAs”; they sequester miRNAs which directly affects the miRNA target gene. The ceRNA networks involving lncRNAs, miRNAs and protein coding mRNAs affect a wide spectrum of biological processes. Relevant for this review is the lncRNA/miRNA/MYC axis, because it is involved in hematological malignancies (Figure 4). An example is the ubiquitously expressed lncRNA KCNQ1OT1, which is transcribed in an antisense direction from intron 11 of the KCNQ1 gene and is localized in the nucleus (126). KCNQ1OT1 is involved in epigenetic gene silencing (127), controls maternal CDKN1C (p57Kip2) expression in muscle cells by promoting accumulation of H3K27me3 at its promoter, which is catalyzed by the histone-methyl-transferase EZH2 (128). Knockdown of KCNQ1OT1 led to the upregulation of the maternal and functional p57Kip2 allele during muscle differentiation (128). Previous evidence had already indicated that this lncRNA can act as an oncogene in AML via different mechanisms (129, 130). In one recent study, KCNQ1OT1 was shown to regulate MYC expression by sponging miR-326 in AML (131). Knockdown of KCNQ1OT1 significantly decreased MYC protein expression in AML cell lines which was rescued by miR-326 abrogation. These findings indicated that KCNQ1OT1 is a so called “competing endogenous” RNA (ceRNA) for miR-326 to regulate MYC expression in AML cells. Other loss of function studies showed that KCNQ1OT1 is required for cell proliferation and survival and that loss of KCNQ1OT1 promotes cell differentiation in AML cells, a feature which makes it attractive as a therapeutic target for this type of leukemia (131).
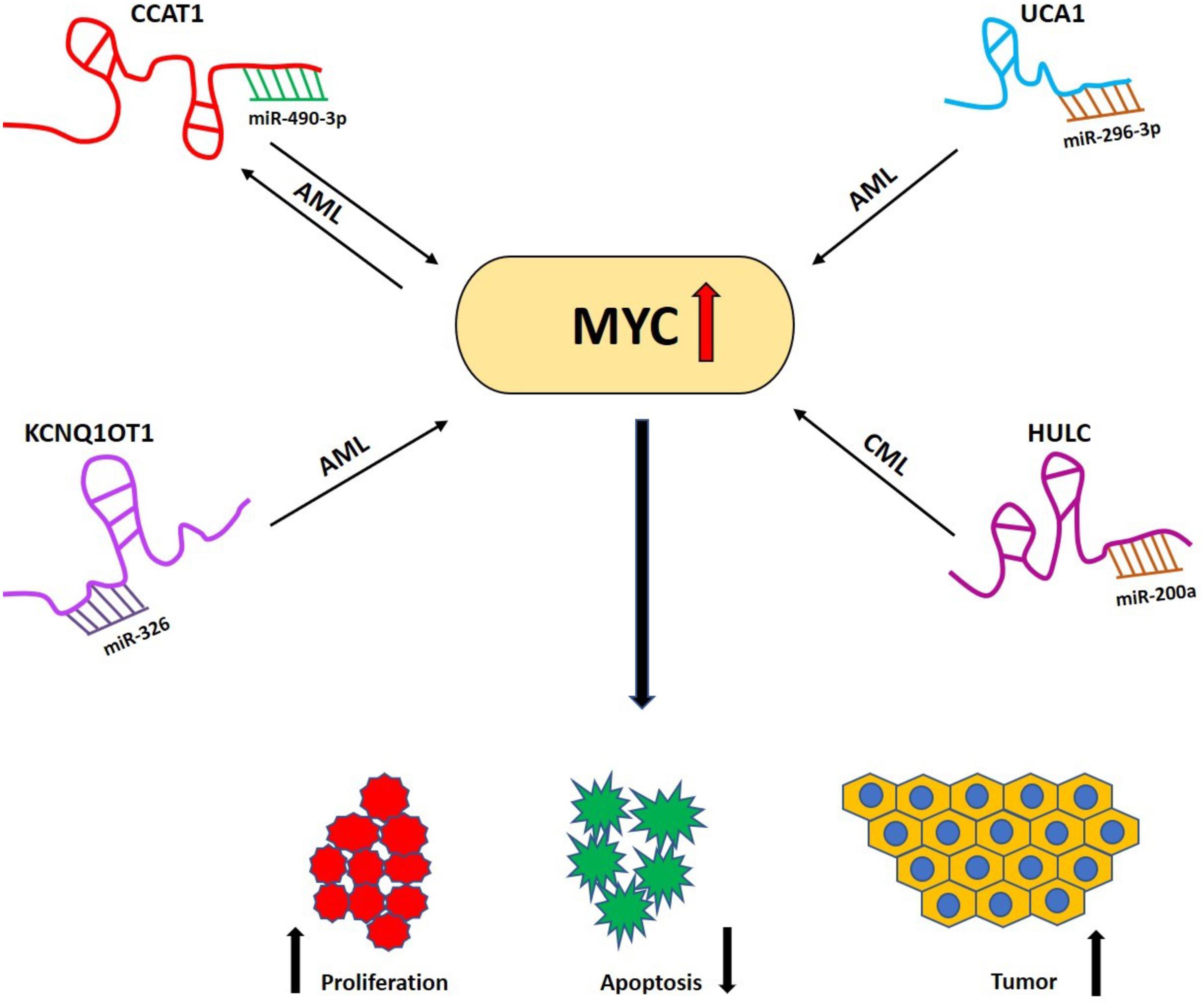
Figure 4. LncRNA/miRNA/MYC axis in different hematological malignancies. Different lncRNAs acting as competing endogenous RNAs (ceRNAs) sequestering miRNAs, which results in the elevation of MYC expression and eventually enhances its tumorigenic effect (decreased apoptosis, increased proliferation and tumor progression) in different hematological malignancies. LncRNA CCAT1 acts as a sponge for miR-490-3p and generates a feedback loop mechanism in AML. Other lncRNAs such as UCA1, KCNQ1OT1 sequester miR-296-3p and miR-326, respectively, in AML while lncRNA HULC sponges miR-200a in CML. AML, Acute myeloid leukemia; CML, Chronic myeloid leukemia.
The lncRNA urothelial carcinoma associated 1 (UCA1) was discovered by Wang et al. as a specific and possibly unique biomarker for bladder carcinoma (132). However, it was also found to be overexpressed in different other malignancies (133–135) and mounting evidence shows its carcinogenic role in AML (136–139). Li et al. discovered the existence of a miR-296-3p/MYC axis in AML being regulated by UCA1 (140). Here, UCA1 acted as a ceRNA of miR-296-3p by binding to miR-296-3p. MYC is a target of miR-296-3p and UCA1 positively regulates MYC expression. The study also carried out in vivo experiments in which HL-60 cells with stable UCA1 knockdown were injected subcutaneously into the armpit of NOD/SCID mice (140). It was found that UCA1 knockdown led to significant reduction in tumor volume. The expression analysis in the tumors showed miR-296-3p to be significantly upregulated while MYC was downregulated both at mRNA and protein levels that complements their findings with in vitro assay (140).
Work from the Wang group established the lncRNA HULC as a novel candidate that regulates MYC in CML (141). HULC is well conserved in primates; its gene is located on chromosome 6p24.3. The HULC lncRNA is spliced, polyadenylated and is located in the cytoplasm (142, 143). In bone marrow samples from CML patients, the expression levels of HULC correlated positively with MYC and knockdown of HULC retarded the proliferation of CML leukemic cells and induced apoptosis by repressing MYC, Bcl-2 and by upregulating the PI3K/AKT signaling pathway (141). Moreover, HULC can function as a ceRNA for miR-200a to modulate MYC and Bcl-2 expression in CML cells; however, whether HULC is a new target for CML therapy needs still to be validated in particular using animal studies.
CCAT1/miR-490-3p/MAPK1/MYC axis involving a feedback loop mechanism in AML
Colon cancer−associated transcript−1 has been studied and shown to play important roles in AML (124, 144). Recently Wang et al. revealed the existence of a feedback loop mechanism involving CCAT1/miR-490-3p/MAPK1/MYC in AML (145). They demonstrated that CCAT1 acted as an oncogene promoting the proliferation and inhibiting the apoptosis while miR-490-3p acted as tumor-suppressor with opposite phenotypes in AML cells. CCAT1 acted as sponge for miR-490-3p to elevate MAPK1 and MYC expression (Figure 4). Overexpression of MYC upregulated the expression of CCAT1 and p-MAPK1 while downregulated the expression of miR-490-3p. Upon silencing CCAT1, expression of p-MAPK1 and MYC was significantly decreased indicating that a positive feedback loop exists among CCAT1/miR-490-3P/MAPK1/MYC in AML cells and therefore provided new candidates for future research and treatment of AML (145).
LncRNAs in HOX Gene Clusters: HOTAIR and HOTAIRM1
HOX genes are found in clusters on different chromosomes and encode several important lncRNAs most notably, HOTAIR, HOXA11-AS, HOTTIP, and HOTAIR myeloid 1 (HOTAIRM1) (146). Among these, HOTAIR is one of the most well-studied lncRNAs arising from the HOXC gene cluster on chromosome 12 (Figure 5). This lncRNA is upregulated in the majority of cancers and acts as an oncogene [reviewed in (147)]. To study the role of Hotair in the immunologic rejection of leukemic cells, DBA/2 mice were injected intravenously with cells from the L1210 lymphocytic leukemia cell line (148). Leukemic mice were then treated with Hotair mimics as well as small interfering RNA against Hotair. Interestingly, the Wnt/β-catenin pathway was activated and the expression of Myc, cyclinD1, and GSK−3β were increased in the bone marrow of these mice induced by overexpressing Hotair while knockdown of Hotair had the opposite effects. One of the major conclusions of this study was that Hotair inhibits the immunologic rejection of acute lymphocytic leukemia cells in mice by activating the Wnt/β-catenin pathway including Myc as one of its effectors (148).
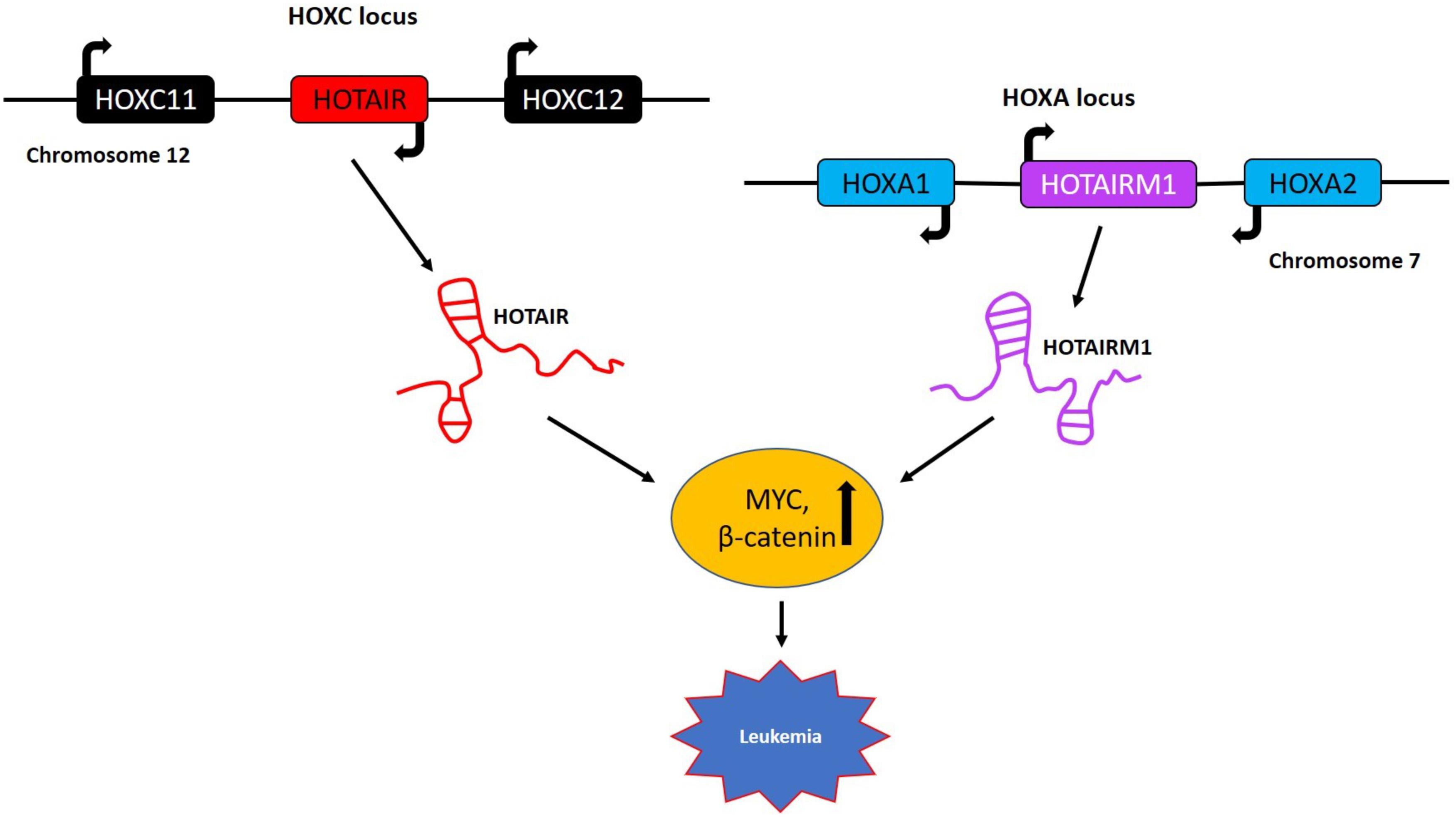
Figure 5. LncRNAs in HOX gene clusters with role in leukemia. HOTAIR is generated by transcription of the antisense strand of the HOXC gene, which is specifically located between HOXC11 and HOXC12 on chromosome 12. The lncRNA HOTAIRM1 is transcribed from the antisense strand of the HOXA gene cluster and is located between the HOXA1 and HOXA2 genes on chromosome 7. These two lncRNAs primarily regulate the expression of MYC and β-catenin and are implicated in lymphocytic leukemia and acute myeloid leukemia, respectively.
HOTAIR myeloid 1 is another lncRNA belonging to the HOXA gene cluster with a specific role in the maturation of myeloid cells and possibly in AML (Figure 5). Most recently, a potential role of HOTAIRM1 as a therapeutic target for overcoming the cytarabine (Ara-C) resistance in AML was reported (149). These studies showed that knockdown of HOTAIRM1 in the HL60 and THP-1 leukemia lines inactivated the Wnt/β-catenin pathway by downregulating the expression of β-catenin, PFKP (platelet-type phosphofructokinase) and MYC. In addition, HOTAIRM1 knockdown enhanced cytarabine cytotoxicity via the Wnt/β-catenin pathway also involving MYC in HL60 cells (149). The HOX gene clusters harbor several lncRNAs regulating MYC expression and even protein coding genes from HOXA and HOXC gene clusters like HOXA9, HOXA10, and HOXC6 have been shown to play crucial roles in AML and ALL by regulating MYC (150, 151). The above examples indicate the involvement of a wide spectrum of genes from the HOX gene clusters in the regulation of MYC in different hematological malignancies offering a series of alternative targets for future therapies.
Conclusion and Future Perspectives
The relationship between the proto-oncoprotein MYC, the MYC gene and lncRNAs is of high complexity since they form an interactive network in which all molecules involved can be regulators or targets connected by linear dependencies and feed-back or feed-forward loops. This network between MYC and lncRNAs clearly plays a crucial role in the regulation of gene expression in different hematological malignancies and a number of ongoing studies will reveal additional lncRNAs that regulate or are controlled by MYC. Since MYC is activated in a majority of human cancers and in many malignancies of the hematopoietic system, targeting this transcription factor would be a powerful approach for an effective tumor therapy. However, despite many efforts direct targeting of MYC remains challenging because of its “undruggable” protein structure, lack of enzymatic activity and its nuclear localization, rendering it less accessible as for example membrane receptors or cytoplasmic kinases. Therefore, alternative possibilities have to be explored such as targeting pathways that are involved in the regulation of MYC expression, or the activity or stability of the MYC protein. First studies have now appeared describing how to achieve effective targeting of lncRNAs and several different methods have emerged. One possibility remains the direct transcriptional inhibition of lncRNAs through Crispr/Cas9 mediated disruption or silencing via siRNAs. However, because lncRNAs interact with proteins, an alternative strategy would be to interrupt this interaction with small molecules, which could then be developed into drug candidates (152). The future will show which method will prevail, but given the importance and critical role of lncRNAs that regulate MYC or act as MYC effectors, their role as therapeutic targets cannot be underestimated. It is well possible that they represent a new way to control MYC activity indirectly and lead to successful strategies to neutralize MYC and offer promising as well as intriguing new therapeutic approaches where previous concepts have not been successful.
Author Contributions
KA and TM have generated the text and the figures. Both authors contributed to the article and approved the submitted version.
Funding
Work in the laboratory of the authors was supported by a CIHR Foundation grant (FDN – 148372), a Tier 1 Canadian Research Chair and a grant from AmorChem Inc. to TM and a Jean Coutu post-doctoral fellowship from the IRCM to KA. The authors declare that this study received funding from AmorChem, Inc. The funder was not involved in the study design, collection, analysis, interpretation of data, the writing of this article or the decision to submit it for publication.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Iyer MK, Niknafs YS, Malik R, Singhal U, Sahu A, Hosono Y, et al. The landscape of long non-coding RNAs in the human transcriptome. Nat Genet. (2015) 47:199–208. doi: 10.1038/ng.3192
2. Doolittle WF. Is junk DNA bunk? A critique of ENCODE. Proc Natl Acad Sci USA. (2013) 110:5294–300. doi: 10.1073/pnas.1221376110
3. Palazzo AF, Lee ES. Non-coding RNA: what is functional and what is junk? Front Genet. (2015) 6:2. doi: 10.3389/fgene.2015.00002
4. Djebali S, Davis CA, Merkel A, Dobin A, Lassmann T, Mortazavi A, et al. Landscape of transcription in human cells. Nature. (2012) 489:101–8. doi: 10.1038/nature11233
5. Carninci P, Kasukawa T, Katayama S, Gough J, Frith MC, Maeda N, et al. The transcriptional landscape of the mammalian genome. Science. (2005) 309:1559–63. doi: 10.1126/science.1112014
6. Cabili MN, Trapnell C, Goff L, Koziol M, Tazon-Vega B, Regev A, et al. Integrative annotation of human large intergenic non-coding RNAs reveals global properties and specific subclasses. Genes Dev. (2011) 25:1915–27. doi: 10.1101/gad.17446611
7. Esteller M. Non-coding RNAs in human disease. Nat Rev Genet. (2011) 12:861–74. doi: 10.1038/nrg3074
8. Agirre X, Meydan C, Jiang Y, Garate L, Doane AS, Li Z, et al. Long non-coding RNAs discriminate the stages and gene regulatory states of human humoral immune response. Nat Commun. (2019) 10:821. doi: 10.1038/s41467-019-08679-z
9. Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. (2009) 10:155–9. doi: 10.1038/nrg2521
10. Guttman M, Amit I, Garber M, French C, Lin MF, Feldser D, et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature. (2009) 458:223–7. doi: 10.1038/nature07672
11. Dahariya S, Paddibhatla I, Kumar S, Raghuwanshi S, Pallepati A, Gutti RK. Long non-coding RNA: classification, biogenesis and functions in blood cells. Mol Immunol. (2019) 112:82–92. doi: 10.1016/j.molimm.2019.04.011
12. Gutschner T, Diederichs S. The hallmarks of cancer: a long non-coding RNA point of view. RNA Biol. (2012) 9:703–19. doi: 10.4161/rna.20481
13. Liu SJ, Horlbeck MA, Cho SW, Birk HS, Malatesta M, He D, et al. CRISPRi-based genome-scale identification of functional long non-coding RNA loci in human cells. Science. (2017) 17:355. doi: 10.1126/science.aah7111
14. Kopp F, Mendell JT. Functional classification and experimental dissection of long non-coding RNAs. Cell. (2018) 172:393–407. doi: 10.1016/j.cell.2018.01.011
15. Han P, Chang CP. Long non-coding RNA and chromatin remodeling. RNA Biol. (2015) 12:1094–8. doi: 10.1080/15476286.2015.1063770
16. Marchese FP, Raimondi I, Huarte M. The multidimensional mechanisms of long non-coding RNA function. Genome Biol. (2017) 18:206. doi: 10.1186/s13059-017-1348-2
17. Wu M, Shen J. From super-enhancer non-coding RNA to immune checkpoint: frameworks to functions. Front Oncol. (2019) 9:1307. doi: 10.3389/fonc.2019.01307
18. Kartha RV, Subramanian S. Competing endogenous RNAs (ceRNAs): new entrants to the intricacies of gene regulation. Front Genet. (2014) 5:8. doi: 10.3389/fgene.2014.00008
19. Dykes IM, Emanueli C. Transcriptional and Post-transcriptional gene regulation by long non-coding RNA. Genom Proteom Bioinform. (2017) 15:177–86. doi: 10.1016/j.gpb.2016.12.005
20. Senturk Cetin N, Kuo CC, Ribarska T, Li R, Costa IG, Grummt I. Isolation and genome-wide characterization of cellular DNA:RNA triplex structures. Nucleic Acids Res. (2019) 47:2306–21. doi: 10.1093/nar/gky1305
21. Huarte M. The emerging role of lncRNAs in cancer. Nat. Med. (2015) 21:1253–61. doi: 10.1038/nm.3981
22. Sun J, Cheng L, Shi H, Zhang Z, Zhao H, Wang Z, et al. A potential panel of six-long non-coding RNA signature to improve survival prediction of diffuse large-B-cell lymphoma. Sci Rep. (2016) 6:27842. doi: 10.1038/srep27842
23. Zhou M, Zhao H, Xu W, Bao S, Cheng L, Sun J. Discovery and validation of immune-associated long non-coding RNA biomarkers associated with clinically molecular subtype and prognosis in diffuse large B cell lymphoma. Mol Cancer. (2017) 16:16. doi: 10.1186/s12943-017-0580-4
24. Liu YR, Jiang YZ, Xu XE, Yu KD, Jin X, Hu X, et al. Comprehensive transcriptome analysis identifies novel molecular subtypes and subtype-specific RNAs of triple-negative breast cancer. Breast Cancer Res. (2016) 18:33. doi: 10.1186/s13058-016-0690-8
25. Brazao TF, Johnson JS, Muller J, Heger A, Ponting CP, Tybulewicz VL. Long non-coding RNAs in B-cell development and activation. Blood. (2016) 128:e10–9. doi: 10.1182/blood-2015-11-680843
26. Garitano-Trojaola A, Agirre X, Prosper F, Fortes P. Long non-coding RNAs in haematological malignancies. Int J Mol Sci. (2013) 14:15386–422. doi: 10.3390/ijms140815386
27. Li Y, Li W, Liang B, Li L, Wang L, Huang H, et al. Identification of cancer risk lncRNAs and cancer risk pathways regulated by cancer risk lncRNAs based on genome sequencing data in human cancers. Sci Rep. (2016) 6:39294. doi: 10.1038/srep39294
28. James AR, Schroeder MP, Neumann M, Bastian L, Eckert C, Gokbuget N, et al. Long non-coding RNAs defining major subtypes of B cell precursor acute lymphoblastic leukemia. J Hematol Oncol. (2019) 12:8. doi: 10.1186/s13045-018-0692-3
29. Ng M, Heckl D, Klusmann JH. The regulatory roles of long non-coding RNAs in acute myeloid leukemia. Front Oncol. (2019) 9:570. doi: 10.3389/fonc.2019.00570
30. Cruz-Miranda GM, Hidalgo-Miranda A, Barcenas-Lopez DA, Nunez-Enriquez JC, Ramirez-Bello J, Mejia-Arangure JM, et al. Long non-coding RNA and acute leukemia. Int J Mol Sci. (2019) 20:735. doi: 10.3390/ijms20030735
31. Sehgal L, Mathur R, Braun FK, Wise JF, Berkova Z, Neelapu S, et al. FAS-antisense 1 lncRNA and production of soluble versus membrane Fas in B-cell lymphoma. Leukemia. (2014) 28:2376–87. doi: 10.1038/leu.2014.126
32. Wang P, Xue Y, Han Y, Lin L, Wu C, Xu S, et al. The STAT3-binding long noncoding RNA lnc-DC controls human dendritic cell differentiation. Science. (2014) 344:310–3. doi: 10.1126/science.1251456
33. Atianand MK, Hu W, Satpathy AT, Shen Y, Ricci EP, Alvarez-Dominguez JR, et al. A long noncoding RNA lincRNA-EPS Acts as a transcriptional brake to restrain inflammation. Cell. (2016) 165:1672–85. doi: 10.1016/j.cell.2016.05.075
34. Alvarez-Dominguez JR, Knoll M, Gromatzky AA, Lodish HF. The super-enhancer-derived alncRNA-EC7/Bloodlinc potentiates red blood cell development in trans. Cell Rep. (2017) 19:2503–14. doi: 10.1016/j.celrep.2017.05.082
35. Schwarzer A, Emmrich S, Schmidt F, Beck D, Ng M, Reimer C, et al. The non-coding RNA landscape of human hematopoiesis and leukemia. Nat Commun. (2017) 8:218. doi: 10.1038/s41467-017-00212-4
36. Doose G, Haake A, Bernhart SH, Lopez C, Duggimpudi S, Wojciech F, et al. MINCR is a MYC-induced lncRNA able to modulate MYC’s transcriptional network in Burkitt lymphoma cells. Proc Natl Acad Sci USA. (2015) 112:E5261–70. doi: 10.1073/pnas.1505753112
37. Lee S, Luo W, Shah T, Yin C, O’Connell T, Chung TH, et al. The effects of DLEU1 gene expression in Burkitt lymphoma (BL): potential mechanism of chemoimmunotherapy resistance in BL. Oncotarget. (2017) 8:27839–53. doi: 10.18632/oncotarget.15711
38. Zeng C, Liu S, Lu S, Yu X, Lai J, Wu Y, et al. The c-Myc-regulated lncRNA NEAT1 and paraspeckles modulate imatinib-induced apoptosis in CML cells. Mol Cancer. (2018) 17:130. doi: 10.1186/s12943-018-0884-z
39. Yazdi N, Houshmand M, Atashi A, Kazemi A, Najmedini AA, Zarif MN. Long noncoding RNA PVT1: potential oncogene in the development of acute lymphoblastic leukemia. Turk J Biol. (2018) 42:405–13. doi: 10.3906/biy-1801-46
40. Wang Y, Wu P, Lin R, Rong L, Xue Y, Fang Y. LncRNA NALT interaction with NOTCH1 promoted cell proliferation in pediatric T cell acute lymphoblastic leukemia. Sci Rep. (2015) 5:13749. doi: 10.1038/srep13749
41. Trimarchi T, Bilal E, Ntziachristos P, Fabbri G, Dalla-Favera R, Tsirigos A, et al. Genome-wide mapping and characterization of Notch-regulated long noncoding RNAs in acute leukemia. Cell. (2014) 158:593–606. doi: 10.1016/j.cell.2014.05.049
42. Wang QM, Lian GY, Song Y, Huang YF, Gong Y. LncRNA MALAT1 promotes tumorigenesis and immune escape of diffuse large B cell lymphoma by sponging miR-195. Life Sci. (2019) 231:116335. doi: 10.1016/j.lfs.2019.03.040
43. Zhao CC, Jiao Y, Zhang YY, Ning J, Zhang YR, Xu J, et al. Lnc SMAD5-AS1 as ceRNA inhibit proliferation of diffuse large B cell lymphoma via Wnt/beta-catenin pathway by sponging miR-135b-5p to elevate expression of APC. Cell Death Dis. (2019) 10:252. doi: 10.1038/s41419-019-1479-3
44. Wurm AA, Pina C. Long Non-coding RNAs as functional and structural chromatin modulators in acute myeloid leukemia. Front Oncol. (2019) 9:899. doi: 10.3389/fonc.2019.00899
45. Lin CY, Loven J, Rahl PB, Paranal RM, Burge CB, Bradner JE, et al. Transcriptional amplification in tumor cells with elevated c-Myc. Cell. (2012) 151:56–67. doi: 10.1016/j.cell.2012.08.026
46. Nie Z, Hu G, Wei G, Cui K, Yamane A, Resch W, et al. c-Myc is a universal amplifier of expressed genes in lymphocytes and embryonic stem cells. Cell. (2012) 151:68–79. doi: 10.1016/j.cell.2012.08.033
47. Blackwood EM, Eisenman RN. Max: a helix-loop-helix zipper protein that forms a sequence-specific DNA-binding complex with Myc. Science. (1991) 251:1211–7. doi: 10.1126/science.2006410
48. Meyer N, Penn LZ. Reflecting on 25 years with MYC. Nat Rev Cancer. (2008) 8:976–90. doi: 10.1038/nrc2231
50. Soucek L, Whitfield J, Martins CP, Finch AJ, Murphy DJ, Sodir NM, et al. Modelling Myc inhibition as a cancer therapy. Nature. (2008) 455:679–83. doi: 10.1038/nature07260
51. Dalla-Favera R, Bregni M, Erikson J, Patterson D, Gallo RC, Croce CM. Human c-myc onc gene is located on the region of chromosome 8 that is translocated in Burkitt lymphoma cells. Proc Natl Acad Sci USA. (1982) 79:7824–7. doi: 10.1073/pnas.79.24.7824
52. Molyneux EM, Rochford R, Griffin B, Newton R, Jackson G, Menon G, et al. Burkitt’s lymphoma. Lancet. (2012) 379:1234–44. doi: 10.1016/S0140-6736(11)61177-X
53. Albihn A, Johnsen JI, Henriksson MA. MYC in oncogenesis and as a target for cancer therapies. Adv Cancer Res. (2010) 107:163–224. doi: 10.1016/S0065-230X(10)07006-5
54. Kalkat M, De Melo J, Hickman KA, Lourenco C, Redel C, Resetca D, et al. MYC deregulation in primary human cancers. Genes (Basel). (2017) 8:151. doi: 10.3390/genes8060151
55. Cerami E, Gao J, Dogrusoz U, Gross BE, Sumer SO, Aksoy BA, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov. (2012) 2:401–4. doi: 10.1158/2159-8290.CD-12-0095
56. Court EL, Smith MA, Avent ND, Hancock JT, Morgan LM, Gray AG, et al. DNA microarray screening of differential gene expression in bone marrow samples from AML, non-AML patients and AML cell lines. Leukemia Res. (2004) 28:743–53. doi: 10.1016/j.leukres.2003.11.011
57. Diaz-Blanco E, Bruns I, Neumann F, Fischer JC, Graef T, Rosskopf M, et al. Molecular signature of CD34(+) hematopoietic stem and progenitor cells of patients with CML in chronic phase. Leukemia. (2007) 21:494–504. doi: 10.1038/sj.leu.2404549
58. Shi J, Vogt PK. Posttranslational regulation of Myc by promyelocytic leukemia zinc finger protein. Int J Cancer J Int Cancer. (2009) 125:1558–65. doi: 10.1002/ijc.24449
59. Wall M, Poortinga G, Hannan KM, Pearson RB, Hannan RD, McArthur GA. Translational control of c-MYC by rapamycin promotes terminal myeloid differentiation. Blood. (2008) 112:2305–17. doi: 10.1182/blood-2007-09-111856
60. De Barrios O, Meler A, Parra M. MYC’s Fine line between b cell development and malignancy. Cells. (2020) 9:523. doi: 10.3390/cells9020523
61. Delgado MD, Leon J. Myc roles in hematopoiesis and leukemia. Genes Cancer. (2010) 1:605–16. doi: 10.1177/1947601910377495
62. Fernandez PC, Frank SR, Wang L, Schroeder M, Liu S, Greene J, et al. Genomic targets of the human c-Myc protein. Genes Dev. (2003) 17:1115–29. doi: 10.1101/gad.1067003
63. Li Z, Van Calcar S, Qu C, Cavenee WK, Zhang MQ, Ren BA. global transcriptional regulatory role for c-Myc in Burkitt’s lymphoma cells. Proc Natl Acad Sci USA. (2003) 100:8164–9. doi: 10.1073/pnas.1332764100
64. Zhang L, Gao L, Shao M, Sun GY. A MYC target long non-coding RNA GATA2-AS1 regulates non-small cell lung cancer growth. Neoplasma. (2019) 66:954–62. doi: 10.4149/neo_2019_190210N112
65. Chen S, Gu T, Lu Z, Qiu L, Xiao G, Zhu X, et al. Roles of MYC-targeting long non-coding RNA MINCR in cell cycle regulation and apoptosis in non-small cell lung Cancer. Respir Res. (2019) 20:202. doi: 10.1186/s12931-019-1174-z
66. Zhang J, Chen C, Wu L, Wang Q, Chen J, Zhang S, et al. C-myc contributes to the release of Muller cells-derived proinflammatory cytokines by regulating lncRNA MIAT/XNIP pathway. Int J Biochem Cell Biol. (2019) 114:105574. doi: 10.1016/j.biocel.2019.105574
67. Tokgun PE, Tokgun O, Kurt S, Tomatir AG, Akca H. MYC driven regulation of long non-coding RNA profiles in breast cancer cells. Gene. (2019) 714:143955. doi: 10.1016/j.gene.2019.143955
68. Xiang S, Gu H, Jin L, Thorne RF, Zhang XD, Wu M. LncRNA IDH1-AS1 links the functions of c-Myc and HIF1alpha via IDH1 to regulate the Warburg effect. Proc Natl Acad Sci USA. (2018) 115:E1465–74. doi: 10.1073/pnas.1711257115
69. Li CM, Margolin AA, Salas M, Memeo L, Mansukhani M, Hibshoosh H, et al. PEG10 is a c-MYC target gene in cancer cells. Cancer Res. (2006) 66:665–72. doi: 10.1158/0008-5472.CAN-05-1553
70. Tseng YY, Moriarity BS, Gong W, Akiyama R, Tiwari A, Kawakami H, et al. PVT1 dependence in cancer with MYC copy-number increase. Nature. (2014) 512:82–6. doi: 10.1038/nature13311
71. Hung CL, Wang LY, Yu YL, Chen HW, Srivastava S, Petrovics G, et al. A long noncoding RNA connects c-Myc to tumor metabolism. Proc Natl Acad Sci USA. (2014) 111:18697–702. doi: 10.1073/pnas.1415669112
72. Winkle M, van den Berg A, Tayari M, Sietzema J, Terpstra M, Kortman G, et al. Long noncoding RNAs as a novel component of the Myc transcriptional network. FASEB J. (2015) 29:2338–46. doi: 10.1096/fj.14-263889
73. Hart JR, Roberts TC, Weinberg MS, Morris KV, Vogt PK. MYC regulates the non-coding transcriptome. Oncotarget. (2014) 5:12543–54. doi: 10.18632/oncotarget.3033
74. Sabo A, Kress TR, Pelizzola M, de Pretis S, Gorski MM, Tesi A, et al. Selective transcriptional regulation by Myc in cellular growth control and lymphomagenesis. Nature. (2014) 511:488–92. doi: 10.1038/nature13537
75. Alfano D, Votta G, Schulze A, Downward J, Caputi M, Stoppelli MP, et al. Modulation of cellular migration and survival by c-Myc through the downregulation of urokinase (uPA) and uPA receptor. Mol Cell Biol. (2010) 30:1838–51. doi: 10.1128/MCB.01442-09
76. Pocsfalvi G, Votta G, De Vincenzo A, Fiume I, Raj DA, Marra G, et al. Analysis of secretome changes uncovers an autocrine/paracrine component in the modulation of cell proliferation and motility by c-Myc. J Prot Res. (2011) 10:5326–37. doi: 10.1021/pr200584y
77. Yustein JT, Liu YC, Gao P, Jie C, Le A, Vuica-Ross M, et al. Induction of ectopic Myc target gene JAG2 augments hypoxic growth and tumorigenesis in a human B-cell model. Proc Natl Acad Sci USA. (2010) 107:3534–9. doi: 10.1073/pnas.0901230107
78. Schuhmacher M, Staege MS, Pajic A, Polack A, Weidle UH, Bornkamm GW, et al. Control of cell growth by c-Myc in the absence of cell division. Curr Biol. (1999) 9:1255–8. doi: 10.1016/s0960-9822(99)80507-7
79. Hart JR, Weinberg MS, Morris KV, Roberts TC, Janda KD, Garner AL, et al. MINCR is not a MYC-induced lncRNA. Proc Natl Acad Sci USA. (2016) 113:E496–7. doi: 10.1073/pnas.1519903113
80. Doose G, Hoffmann S, Iaccarino I. Reply to Hart et al.: MINCR and MYC: More than expression correlation. Proc Natl Acad Sci USA. (2016) 113:E498. doi: 10.1073/pnas.1520413113
81. Lu Y, Hu Z, Mangala LS, Stine ZE, Hu X, Jiang D, et al. MYC targeted long noncoding RNA DANCR promotes cancer in Part by reducing p21 levels. Cancer Res. (2018) 78:64–74. doi: 10.1158/0008-5472.CAN-17-0815
82. Jiang N, Wang X, Xie X, Liao Y, Liu N, Liu J, et al. lncRNA DANCR promotes tumor progression and cancer stemness features in osteosarcoma by upregulating AXL via miR-33a-5p inhibition. Cancer Lett. (2017) 405:46–55. doi: 10.1016/j.canlet.2017.06.009
83. Liang H, Zhang C, Guan H, Liu J, Cui Y. LncRNA DANCR promotes cervical cancer progression by upregulating ROCK1 via sponging miR-335-5p. J Cell Physiol. (2019) 234:7266–78. doi: 10.1002/jcp.27484
84. Wang S, Jiang M. The long non-coding RNA-DANCR exerts oncogenic functions in non-small cell lung cancer via miR-758-3p. Biomed Pharmacother Biomed Pharmacother. (2018) 103:94–100. doi: 10.1016/j.biopha.2018.03.053
85. Li J, Zhou L. Overexpression of lncRNA DANCR positively affects progression of glioma via activating Wnt/beta-catenin signaling. Biomed Pharmacother Biomed Pharmacother. (2018) 102:602–7. doi: 10.1016/j.biopha.2018.03.116
86. Lu QC, Rui ZH, Guo ZL, Xie W, Shan S, Ren T. LncRNA-DANCR contributes to lung adenocarcinoma progression by sponging miR-496 to modulate mTOR expression. J Cell Mol Med. (2018) 22:1527–37. doi: 10.1111/jcmm.13420
87. Li Z, Hou P, Fan D, Dong M, Ma M, Li H, et al. The degradation of EZH2 mediated by lncRNA ANCR attenuated the invasion and metastasis of breast cancer. Cell Death Different. (2017) 24:59–71. doi: 10.1038/cdd.2016.95
88. Tang J, Zhong G, Zhang H, Yu B, Wei F, Luo L, et al. LncRNA DANCR upregulates PI3K/AKT signaling through activating serine phosphorylation of RXRA. Cell Death Dis. (2018) 9:1167. doi: 10.1038/s41419-018-1220-7
89. Wang N, Zhang C, Wang W, Liu J, Yu Y, Li Y, et al. Long noncoding RNA DANCR regulates proliferation and migration by epigenetically silencing FBP1 in tumorigenesis of cholangiocarcinoma. Cell Death Dis. (2019) 10:585. doi: 10.1038/s41419-019-1810-z
90. Bill M, Papaioannou D, Karunasiri M, Kohlschmidt J, Pepe F, Walker CJ, et al. Expression and functional relevance of long non-coding RNAs in acute myeloid leukemia stem cells. Leukemia. (2019) 33:2169–82. doi: 10.1038/s41375-019-0429-5
91. Qian CS, Li LJ, Huang HW, Yang HF, Wu DP. MYC-regulated lncRNA NEAT1 promotes B cell proliferation and lymphomagenesis via the miR-34b-5p-GLI1 pathway in diffuse large B-cell lymphoma. Cancer Cell Int. (2020) 20:87. doi: 10.1186/s12935-020-1158-6
92. Guo G, Kang Q, Chen Q, Chen Z, Wang J, Tan L, et al. High expression of long non-coding RNA H19 is required for efficient tumorigenesis induced by Bcr-Abl oncogene. FEBS Lett. (2014) 588:1780–6. doi: 10.1016/j.febslet.2014.03.038
93. Tamang S, Acharya V, Roy D, Sharma R, Aryaa A, Sharma U, et al. SNHG12: An LncRNA as a potential therapeutic target and biomarker for human cancer. Front Oncol. (2019) 9:901. doi: 10.3389/fonc.2019.00901
94. Zhu L, Zhang X, Fu X, Li Z, Sun Z, Wu J, et al. c-Myc mediated upregulation of long noncoding RNA SNHG12 regulates proliferation and drug sensitivity in natural killer/T-cell lymphoma. J Cell Biochem. (2019) 120:12628–37. doi: 10.1002/jcb.28529
95. Kress TR, Sabo A, Amati B. MYC connecting selective transcriptional control to global RNA production. Nat Rev Cancer. (2015) 15:593–607. doi: 10.1038/nrc3984
96. Wierstra I, Alves J. The c-myc promoter: still MysterY and challenge. Adv Cancer Res. (2008) 99:113–333. doi: 10.1016/S0065-230X(07)99004-1
97. Shi Y, Frost PJ, Hoang BQ, Benavides A, Sharma S, Gera JF, et al. IL-6-induced stimulation of c-myc translation in multiple myeloma cells is mediated by myc internal ribosome entry site function and the RNA-binding protein, hnRNP A1. Cancer Res. (2008) 68:10215–22. doi: 10.1158/0008-5472.CAN-08-1066
98. Jackstadt R, Hermeking H. MicroRNAs as regulators and mediators of c-MYC function. Biochim Biophys Acta. (2015) 1849:544–53. doi: 10.1016/j.bbagrm.2014.04.003
99. Iaccarino I. lncRNAs and MYC: an intricate relationship. Int J Mol Sci. (2017) 18:18071497. doi: 10.3390/ijms18071497
100. Kajino T, Shimamura T, Gong S, Yanagisawa K, Ida L, Nakatochi M, et al. Divergent lncRNA MYMLR regulates MYC by eliciting DNA looping and promoter-enhancer interaction. EMBO J. (2019) 38:e98441. doi: 10.15252/embj.201798441
101. Shtivelman E, Bishop JM. Effects of translocations on transcription from PVT. Mol Cell Biol. (1990) 10:1835–9. doi: 10.1128/mcb.10.4.1835
102. Huppi K, Siwarski D. Chimeric transcripts with an open reading frame are generated as a result of translocation to the Pvt-1 region in mouse B-cell tumors. Int J Cancer J Int Cancer. (1994) 59:848–51. doi: 10.1002/ijc.2910590623
103. Guan Y, Kuo WL, Stilwell JL, Takano H, Lapuk AV, Fridlyand J, et al. Amplification of PVT1 contributes to the pathophysiology of ovarian and breast cancer. Clin Cancer Res. (2007) 13:5745–55. doi: 10.1158/1078-0432.CCR-06-2882
104. Carramusa L, Contino F, Ferro A, Minafra L, Perconti G, Giallongo A, et al. The PVT-1 oncogene is a Myc protein target that is overexpressed in transformed cells. J Cell Physiol. (2007) 213:511–8. doi: 10.1002/jcp.21133
105. Tang J, Li Y, Sang Y, Yu B, Lv D, Zhang W, et al. LncRNA PVT1 regulates triple-negative breast cancer through KLF5/beta-catenin signaling. Oncogene. (2018) 37:4723–34. doi: 10.1038/s41388-018-0310-4
106. Cui M, You L, Ren X, Zhao W, Liao Q, Zhao Y. Long non-coding RNA PVT1 and cancer. Biochem Biophys Res Commun. (2016) 471:10–4. doi: 10.1016/j.bbrc.2015.12.101
107. Jin K, Wang S, Zhang Y, Xia M, Mo Y, Li X, et al. Long non-coding RNA PVT1 interacts with MYC and its downstream molecules to synergistically promote tumorigenesis. Cell Mol Life Sci. (2019) 76:4275–89. doi: 10.1007/s00018-019-03222-1
108. Riquelme E, Suraokar MB, Rodriguez J, Mino B, Lin HY, Rice DC, et al. Frequent coamplification and cooperation between C-MYC and PVT1 oncogenes promote malignant pleural mesothelioma. J Thorac Oncol. (2014) 9:998–1007. doi: 10.1097/JTO.0000000000000202
109. Colombo T, Farina L, Macino G, Paci P. PVT1: a rising star among oncogenic long noncoding RNAs. BioMed Res Int. (2015) 2015:304208. doi: 10.1155/2015/304208
110. Zeng C, Yu X, Lai J, Yang L, Chen S, Li Y. Overexpression of the long non-coding RNA PVT1 is correlated with leukemic cell proliferation in acute promyelocytic leukemia. J Hematol Oncol. (2015) 8:126. doi: 10.1186/s13045-015-0223-4
111. Delas MJ, Sabin LR, Dolzhenko E, Knott SR, Munera ME, Jackson BT, et al. lncRNA requirements for mouse acute myeloid leukemia and normal differentiation. Elife. (2017) 6:25607. doi: 10.7554/eLife.25607
112. Cho SW, Xu J, Sun R, Mumbach MR, Carter AC, Chen YG, et al. Promoter of lncRNA Gene PVT1 is a tumor-suppressor DNA boundary element. Cell. (2018) 173:1398–1412.e1322. doi: 10.1016/j.cell.2018.03.068
113. Zheng C, Xiao Y, Li Y, He D. Knockdown of long non-coding RNA PVT1 inhibits the proliferation of Raji cells through cell cycle regulation. Oncol Lett. (2019) 18:1225–34. doi: 10.3892/ol.2019.10450
114. Ghetti M, Vannini I, Storlazzi CT, Martinelli G, Simonetti G. Linear and circular PVT1 in hematological malignancies and immune response: two faces of the same coin. Mol Cancer. (2020) 19:69. doi: 10.1186/s12943-020-01187-5
115. Salzman J, Gawad C, Wang PL, Lacayo N, Brown PO. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS One. (2012) 7:e30733. doi: 10.1371/journal.pone.0030733
116. Panda AC, Grammatikakis I, Kim KM, De S, Martindale JL, Munk R, et al. Identification of senescence-associated circular RNAs (SAC-RNAs) reveals senescence suppressor CircPVT1. Nucleic Acids Res. (2017) 45:4021–35. doi: 10.1093/nar/gkw1201
117. Wang PL, Bao Y, Yee MC, Barrett SP, Hogan GJ, Olsen MN, et al. Circular RNA is expressed across the eukaryotic tree of life. PLoS One. (2014) 9:e90859. doi: 10.1371/journal.pone.0090859
118. Memczak S, Jens M, Elefsinioti A, Torti F, Krueger J, Rybak A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. (2013) 495:333–8. doi: 10.1038/nature11928
119. Chen J, Li Y, Zheng Q, Bao C, He J, Chen B, et al. Circular RNA profile identifies circPVT1 as a proliferative factor and prognostic marker in gastric cancer. Cancer Lett. (2017) 388:208–19. doi: 10.1016/j.canlet.2016.12.006
120. Ebbesen KK, Hansen TB, Kjems J. Insights into circular RNA biology. RNA Biol. (2017) 14:1035–45. doi: 10.1080/15476286.2016.1271524
121. Verduci L, Ferraiuolo M, Sacconi A, Ganci F, Vitale J, Colombo T, et al. The oncogenic role of circPVT1 in head and neck squamous cell carcinoma is mediated through the mutant p53/YAP/TEAD transcription-competent complex. Genome Biol. (2017) 18:237. doi: 10.1186/s13059-017-1368-y
122. Panda AC, De S, Grammatikakis I, Munk R, Yang X, Piao Y, et al. High-purity circular RNA isolation method (RPAD) reveals vast collection of intronic circRNAs. Nucleic Acids Res. (2017) 45:e116. doi: 10.1093/nar/gkx297
123. Hu J, Han Q, Gu Y, Ma J, McGrath M, Qiao F, et al. Circular RNA PVT1 expression and its roles in acute lymphoblastic leukemia. Epigenomics. (2018) 10:723–32. doi: 10.2217/epi-2017-0142
124. El-Khazragy N, Elayat W, Matbouly S, Seliman S, Sami A, Safwat G, et al. The prognostic significance of the long non-coding RNAs “CCAT1, PVT1” in t(8;21) associated Acute Myeloid Leukemia. Gene. (2019) 707:172–7. doi: 10.1016/j.gene.2019.03.055
125. Hu G, Zhang Y, Gupta M. RIP sequencing in mantle cell lymphoma identifies functional long non-coding RNAs associated with translation machinery. Blood Cancer J. (2019) 9:55. doi: 10.1038/s41408-019-0216-6
126. Korostowski L, Sedlak N, Engel N. The Kcnq1ot1 long non-coding RNA affects chromatin conformation and expression of Kcnq1, but does not regulate its imprinting in the developing heart. PLoS Genetics. (2012) 8:e1002956. doi: 10.1371/journal.pgen.1002956
127. Redrup L, Branco MR, Perdeaux ER, Krueger C, Lewis A, Santos F, et al. The long noncoding RNA Kcnq1ot1 organises a lineage-specific nuclear domain for epigenetic gene silencing. Development. (2009) 136:525–30. doi: 10.1242/dev.031328
128. Andresini O, Rossi MN, Matteini F, Petrai S, Santini T, Maione R. The long non-coding RNA Kcnq1ot1 controls maternal p57 expression in muscle cells by promoting H3K27me3 accumulation to an intragenic MyoD-binding region. Epigenet Chromatin. (2019) 12:8. doi: 10.1186/s13072-019-0253-1
129. Sun H, Sun Y, Chen Q, Xu Z. LncRNA KCNQ1OT1 contributes to the progression and chemoresistance in acute myeloid leukemia by modulating Tspan3 through suppressing miR-193a-3p. Life Sci. (2020) 241:117161. doi: 10.1016/j.lfs.2019.117161
130. Jia ZW, Li Y, Cui GR, Zhao HB, Li PY, Luo JM. [Expression and clinical significance of LncRNA KCNQ1OT1 in patients with acute myeloid leukemia]. Zhongguo Shi Yan Xue Ye Xue Za Zhi. (2018) 26:653–7. doi: 10.7534/j.issn.1009-2137.2018.03.004
131. Cheng P, Lu P, Guan J, Zhou Y, Zou L, Yi X, et al. LncRNA KCNQ1OT1 controls cell proliferation, differentiation and apoptosis by sponging miR-326 to regulate c-Myc expression in acute myeloid leukemia. Neoplasma. (2020) 67:238–48. doi: 10.4149/neo_2018_181215N972
132. Wang XS, Zhang Z, Wang HC, Cai JL, Xu QW, Li MQ, et al. Rapid identification of UCA1 as a very sensitive and specific unique marker for human bladder carcinoma. Clin Cancer Res. (2006) 12:4851–8. doi: 10.1158/1078-0432.CCR-06-0134
133. Xue M, Chen W, Li X. Urothelial cancer associated 1: a long noncoding RNA with a crucial role in cancer. J Cancer Res Clin Oncol. (2016) 142:1407–19. doi: 10.1007/s00432-015-2042-y
134. Zhang Z, Li JZ, Wei ZW, Li F, Li HM, Xiao Y, et al. Correlation between expression levels of lncRNA UCA1 and miR-18a with prognosis of hepatocellular cancer. Eur Rev Med Pharmacol Sci. (2020) 24:3586–91. doi: 10.26355/eurrev_202004_20820
135. Wei H, Qiu YQ, Zeng QS, Wang SF, Yi CJ. LncRNA UCA1 regulates proliferation, migration and invasion of cervical cancer cells by targeting miR-145. Eur Rev Med Pharmacol Sci. (2020) 24:3555–64. doi: 10.26355/eurrev_202004_20816
136. Zhang Y, Liu Y, Xu X. Knockdown of LncRNA-UCA1 suppresses chemoresistance of pediatric AML by inhibiting glycolysis through the microRNA-125a/hexokinase 2 pathway. J Cell Biochem. (2018) 119:6296–308. doi: 10.1002/jcb.26899
137. Sun MD, Zheng YQ, Wang LP, Zhao HT, Yang S. Long noncoding RNA UCA1 promotes cell proliferation, migration and invasion of human leukemia cells via sponging miR-126. Eur Rev Med Pharmacol Sci. (2018) 22:2233–45. doi: 10.26355/eurrev_201804_14809
138. Li JJ, Chen XF, Wang M, Zhang PP, Zhang F, Zhang JJ. Long non-coding RNA UCA1 promotes autophagy by targeting miR-96-5p in acute myeloid leukaemia. Clin Exp Pharmacol Physiol. (2020) 47:877–85. doi: 10.1111/1440-1681.13259
139. Liang Y, Li E, Zhang H, Zhang L, Tang Y, Wanyan Y. Silencing of lncRNA UCA1 curbs proliferation and accelerates apoptosis by repressing SIRT1 signals by targeting miR-204 in pediatric AML. J Biochem Mol Toxicol. (2020) 34:e22435. doi: 10.1002/jbt.22435
140. Li J, Wang M, Chen X. Long non-coding RNA UCA1 modulates cell proliferation and apoptosis by regulating miR-296-3p/Myc axis in acute myeloid leukemia. Cell Cycle. (2020) 20:1–12. doi: 10.1080/15384101.2020.1750814
141. Lu Y, Li Y, Chai X, Kang Q, Zhao P, Xiong J, et al. Long noncoding RNA HULC promotes cell proliferation by regulating PI3K/AKT signaling pathway in chronic myeloid leukemia. Gene. (2017) 607:41–6. doi: 10.1016/j.gene.2017.01.004
142. Wang J, Liu X, Wu H, Ni P, Gu Z, Qiao Y, et al. CREB up-regulates long non-coding RNA, HULC expression through interaction with microRNA-372 in liver cancer. Nucleic Acids Res. (2010) 38:5366–83. doi: 10.1093/nar/gkq285
143. Panzitt K, Tschernatsch MM, Guelly C, Moustafa T, Stradner M, Strohmaier HM, et al. Characterization of HULC, a novel gene with striking up-regulation in hepatocellular carcinoma, as noncoding RNA. Gastroenterology. (2007) 132:330–42. doi: 10.1053/j.gastro.2006.08.026
144. Chen L, Wang W, Cao L, Li Z, Wang X. Long non-coding RNA CCAT1 acts as a competing endogenous RNA to regulate cell growth and differentiation in acute myeloid leukemia. Mol Cells. (2016) 39:330–6. doi: 10.14348/molcells.2016.2308
145. Wang C, Chen F, Fan Z, Yao C, Xiao L. lncRNA CCAT1/miR-490-3p/MAPK1/c-Myc positive feedback loop drives progression of acute myeloid leukaemia. J Biochem. (2020) 167:379–88. doi: 10.1093/jb/mvz107
146. Li L, Wang Y, Song G, Zhang X, Gao S, Liu H. HOX cluster-embedded antisense long non-coding RNAs in lung cancer. Cancer Lett. (2019) 450:14–21. doi: 10.1016/j.canlet.2019.02.036
147. Zhang J, Zhang P, Wang L, Piao HL, Ma L. Long non-coding RNA HOTAIR in carcinogenesis and metastasis. Acta Biochim Biophys Sinica. (2014) 46:1–5. doi: 10.1093/abbs/gmt117
148. Li GJ, Ding H, Miao D. Long-noncoding RNA HOTAIR inhibits immunologic rejection of mouse leukemia cells through activating the Wnt/beta-catenin signaling pathway in a mouse model of leukemia. J Cell Physiol. (2019) 234:10386–96. doi: 10.1002/jcp.27705
149. Chen L, Hu N, Wang C, Zhao H. HOTAIRM1 knockdown enhances cytarabine-induced cytotoxicity by suppression of glycolysis through the Wnt/beta-catenin/PFKP pathway in acute myeloid leukemia cells. Arch Biochem Biophys. (2020) 680:108244. doi: 10.1016/j.abb.2019.108244
150. Zhong X, Prinz A, Steger J, Garcia-Cuellar MP, Radsak M, Bentaher A, et al. HoxA9 transforms murine myeloid cells by a feedback loop driving expression of key oncogenes and cell cycle control genes. Blood Adv. (2018) 2:3137–48. doi: 10.1182/bloodadvances.2018025866
151. Ferrando AA, Armstrong SA, Neuberg DS, Sallan SE, Silverman LB, Korsmeyer SJ, et al. Gene expression signatures in MLL-rearranged T-lineage and B-precursor acute leukemias: dominance of HOX dysregulation. Blood. (2003) 102:262–8. doi: 10.1182/blood-2002-10-3221
Keywords: MYC, long non-coding RNAs, leukemia, lymphoma, hematological malignancies
Citation: Arman K and Möröy T (2020) Crosstalk Between MYC and lncRNAs in Hematological Malignancies. Front. Oncol. 10:579940. doi: 10.3389/fonc.2020.579940
Received: 03 July 2020; Accepted: 14 September 2020;
Published: 08 October 2020.
Edited by:
Alessandro Isidori, AORMN Hospital, ItalyReviewed by:
Guru Prasad Maiti, Oklahoma Medical Research Foundation, United StatesFrancesca Lovat, The Ohio State University, United States
Copyright © 2020 Arman and Möröy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kaifee Arman, bW9oYW1tYWRrYWlmZWUuYXJtYW5AaXJjbS5xYy5jYQ==; Tarik Möröy, VGFyaWsuTW9yb3lAaXJjbS5xYy5jYQ==