 Fernanda F. Peres1,2
Fernanda F. Peres1,2 Raquel Levin1,2
Raquel Levin1,2 Valéria Almeida1,2
Valéria Almeida1,2 Antonio W. Zuardi3,4
Antonio W. Zuardi3,4 Jaime E. Hallak3,4
Jaime E. Hallak3,4 José A. Crippa3,4
José A. Crippa3,4 Vanessa C. Abilio1,2*
Vanessa C. Abilio1,2*- 1Interdisciplinary Laboratory of Clinical Neurosciences, Department of Psychiatry, Escola Paulista De Medicina, Federal University of São Paulo, São Paulo, Brazil
- 2Department of Pharmacology, Escola Paulista De Medicina, Federal University of São Paulo, São Paulo, Brazil
- 3Department of Neuroscience and Behavior, University of São Paulo, Ribeirão Preto, Brazil
- 4National Institute for Translational Medicine (INCT-TM, CNPq), Ribeirão Preto, Brazil
Schizophrenia is a severe psychiatric disorder that involves positive, negative and cognitive symptoms. Prepulse inhibition of startle reflex (PPI) is a paradigm that assesses the sensorimotor gating functioning and is impaired in schizophrenia patients as well as in animal models of this disorder. Recent data point to the participation of the endocannabinoid system in the pathophysiology and pharmacotherapy of schizophrenia. Here, we focus on the effects of cannabinoid drugs on the PPI deficit of animal models of schizophrenia, with greater focus on the SHR (Spontaneously Hypertensive Rats) strain, and on the future prospects resulting from these findings.
Schizophrenia is a debilitating neuropsychiatric disorder that affects 0.7% of world's population (MacDonald and Schulz, 2009) and involves positive (i.e., delusions and hallucinations), negative (e.g., anhedonia, social withdrawal, affective flattening), and cognitive symptoms (such as impaired processing of information and deficits in working memory) (van Os and Kapur, 2009). Currently, schizophrenia's pharmacotherapy is mainly limited to the positive symptoms and associated with severe side effects and high rates of treatment resistance (Briles et al., 2012; Hasan et al., 2012; Abi-Dargham, 2014).
KEY CONCEPT 1. Schizophrenia
Debilitating psychiatric disorder that affects 0.7% of world's population and presents an onset between late adolescence and early adulthood. Schizophrenia's symptomatology includes positive, negative and cognitive symptoms.
Sensorimotor gating is a physiological process that filters sensory information as it is transmitted to motor output systems, preventing information overload and cognitive fragmentation (Cryan and Reif, 2012). This process is impaired especially in schizophrenia (Braff et al., 2001), but also in other neuropsychiatric disorders such as obsessive-compulsive disorder (Ahmari et al., 2012), Tourette's syndrome (Swerdlow et al., 2001), Huntington's disease (Swerdlow et al., 1995), and bipolar disorder (Perry et al., 2001). Prepulse inhibition of startle reflex (PPI) is considered an operational measure of sensorimotor gating and is extensively used in translational studies of schizophrenia, since it is seen in both rodents and humans (Braff et al., 2001). PPI is defined as a reduction of acoustic startle reflex to an intense stimulus (pulse) when immediately preceded by a low intensity stimulus (prepulse).
KEY CONCEPT 2. Sensorimotor gating
Physiological process that filters sensory information as it is transmitted to motor output systems, preventing information overload and cognitive fragmentation. It is impaired in schizophrenia and in other neuropsychiatric disorders.
KEY CONCEPT 3. Prepulse inhibition of startle reflex (PPI)
Reduction of acoustic startle reflex to an intense stimulus (pulse) when immediately preceded by a low intensity stimulus (prepulse). PPI is an operational measure of sensorimotor gating and is seen in both rodents and humans, being extensively used in translational studies.
Prepulse inhibition of startle reflex (PPI) is disrupted in schizophrenia patients and evidence show that PPI deficits are positively correlated to thought disorder (Perry and Braff, 1994; Perry et al., 1999), and associated with impaired functional status and with the presence of auditory hallucinations (Swerdlow et al., 2006; Kumari et al., 2008). PPI deficits are improved by treatment with antipsychotic drugs (Kumari et al., 1999; Weike et al., 2000; Leumann et al., 2002; Oranje et al., 2002; Minassian et al., 2007; Wynn et al., 2007; Martinez-Gras et al., 2009), and this improvement is associated with treatment-related amelioration of schizophrenia symptoms (Minassian et al., 2007). PPI is also disrupted in several animal models of this disorder (Swerdlow et al., 2008), being a useful paradigm to investigate the neurobiology and pharmacotherapy of information processing abnormalities in schizophrenia.
KEY CONCEPT 4. Antipsychotic drugs
Drugs used primarily to treat psychotic states, in particular schizophrenia and bipolar disorder. The antipsychotic drugs are classified in typical and atypical compounds: the typical antipsychotic drugs are associated with motor side effects, and the atypical are linked to metabolic disturbances.
Recent data point to the involvement of the Endocannabinoid system in the pathophysiology of schizophrenia. The endocannabinoid system was described subsequent to the identification of the molecular target of Δ9-tetrahydrocannabinol (Δ9-THC), the main psychoactive compound of Cannabis sativa, and comprises the classical cannabinoid-1 and 2 receptors (CB1 and CB2), their endogenous ligands known as endocannabinoids (e.g., anandamide and 2-arachidonoylglycerol), and the enzymes involved in the endocannabinoid's synthesis and degradation.
KEY CONCEPT 5. Endocannabinoid system
System described subsequent to the identification of the molecular target of Δ9-THC. It comprises the cannabinoid-1 and 2 receptors (CB1 and CB2), their endogenous ligands (named endocannabinoids) and the enzymes involved in the endocannabinoid's metabolism.
The contribution of the endocannabinoid system in schizophrenia is suggested based on some compelling evidence. The exposure to Δ9-THC may induce a transient psychotic condition in healthy subjects (D'Souza et al., 2004; Morrison et al., 2009), whereas in schizophrenia patients, cannabis consumption provokes more and earlier psychotic relapses, even among those under antipsychotic treatment (Linszen et al., 1994; D'Souza et al., 2005; Grech et al., 2005). Moreover, cannabis use has been proved to be a risk factor for psychotic outcomes (Matheson et al., 2011). In accordance, several alterations in the endocannabinoid system are seen in schizophrenia: (1) levels of anandamide are increased in patients' cerebrospinal fluid and peripheral blood (Leweke et al., 1999, 2007; De Marchi et al., 2003; Giuffrida et al., 2004); (2) post-mortem studies show increased CB1 density in patients' dorsolateral prefrontal, anterior cingulate and posterior cingulate cortices (Dean et al., 2001; Zavitsanou et al., 2004; Newell et al., 2006; Dalton et al., 2011); (3) in vivo studies using positron emission tomography (PET) reveal increased density of CB1 in the brain in both medicated and non-medicated schizophrenia patients (Wong et al., 2010; Ceccarini et al., 2013); (4) polymorphisms of the genes that code the cannabinoid receptors CB1 and CB2 are associated to some schizophrenia phenotypes (Ujike et al., 2002; Chavarría-Siles et al., 2008; Ishiguro et al., 2010). In addition, pre-clinical studies show that cannabinoid drugs are able to modulate schizophrenia-like behaviors, including PPI.
In the work that generated this focused review, we evaluated the effects of four cannabinoid drugs on the PPI deficit displayed by the Spontaneously Hypertensive Rat (SHR) strain an animal model characterized by our group to study several aspects of schizophrenia (Levin et al., 2014; Table 1). Here, we will focus on the effects of these four cannabinoid drugs—WIN 55212,2 (WIN–cannabinoid agonist), rimonabant (CB1 antagonist/inverse agonist), AM404 (anandamide uptake inhibitor) and cannabidiol (CB1 antagonist, anandamide uptake inhibitor, among other effects)—on PPI of animal models, mainly the SHR strain, and on the future prospects resulting from these findings.
KEY CONCEPT 6. Spontaneously Hypertensive Rat (SHR) strain
Inbred strain developed by brother-sister mating rats with hypertensive phenotype of the outbred strain Wistar. SHRs display spontaneous hypertension and also several behavioral abnormalities that fit the behavioral phenotype associated with schizophrenia.

Table 1. Cannabinoid drugs as potential agents to treat prepulse inhibition of startle (PPI) deficits in schizophrenia.
SHR strain was developed by selecting rats from Wistar strain with hypertensive phenotype and brother-sister mating (Okamoto and Aoki, 1963). In addition to hypertension, the inbreeding selected some behavioral abnormalities—such as impulsivity, impaired sustained attention and hyperactivity—leading to the proposal of the SHR strain (mainly young animals) as an animal model for attention deficit/hyperactivity disorder (ADHD). It is noteworthy that most of the studies using SHR as ADHD model were performed using as control the Wistar-Kyoto strain (developed by inbreeding Wistar rats without hypertension). Wistar-Kyoto rats may be inappropriate as a control strain since they display inactivity and depressive-like behavior when compared to Wistar rats (WRs) (Overstreet, 2012), and do not show genetic similarities when compared to SHRs (Johnson et al., 1992; St Lezin et al., 1992). In addition, SHR's predictive validity as ADHD model is inconsistent: several studies describe that the administration of psychostimulants (drugs used to treat ADHD) does not attenuate SHRs' behavioral abnormalities (Amini et al., 2004; Yang et al., 2006; van den Bergh et al., 2006; Bizot et al., 2007; Barron et al., 2009; Calzavara et al., 2009), and may even potentiate them (Amini et al., 2004; Yang et al., 2006; Barron et al., 2009; Calzavara et al., 2009).
Conversely, we reported that SHRs, when compared to WRs, display schizophrenia-like behavioral abnormalities. SHRs display increased locomotion (a model for the positive symptoms of schizophrenia—Lipska and Weinberger, 2000), decreased social interaction (that mimics the negative symptoms—File and Seth, 2003), and deficits in the contextual fear conditioning (associated with impairments in emotional memory seen in schizophrenia—Maren et al., 2013) and PPI. These abnormalities are reversed by antipsychotic drugs (with varied effects in WRs, depending on the drug, the dose, and the behavior), but not by psychostimulant drugs, mood stabilizers, dopaminergic antagonists without antipsychotic activity or drugs that modulate anxiety (Calzavara et al., 2009, 2011; Levin et al., 2011). In addition, psychotomimetic manipulations, such as sleep deprivation and administration of psychostimulants, potentiate the behavioral abnormalities displayed by SHRs and induce a schizophrenia-like behavioral phenotype in WRs (Calzavara et al., 2009, 2011; Levin et al., 2011)—as seen in other animal models of the disorder, as well as in patients (Laruelle et al., 1999; Jones et al., 2011). As a result, the SHR strain has been used to investigate genetic alterations related to schizophrenia as well as novel therapeutic strategies for this disorder, including cannabinoid drugs (Levin et al., 2012, 2014; Almeida et al., 2013, 2014; Diana et al., 2015).
WIN is an agonist of CB1 and CB2 receptors. The acute administration of WIN attenuates the SHRs' PPI deficit, and does not modify the PPI of WRs—although a trend to reduce the WRs' PPI is seen with the lowest dose (Levin et al., 2014). Accordingly, other studies demonstrate that WIN reverses the PPI impairment displayed by psychosocially stressed mice (Brzózka et al., 2011) and induced by chronic administration phencyclidine (Spano et al., 2010), two other animal models of schizophrenia. The beneficial effect of WIN on the PPI of psychosocially stressed mice is prevented by pretreatment with the CB1 antagonist rimonabant (Brzózka et al., 2011), suggesting an involvement of these receptors on the WIN's actions on sensorimotor gating functioning.
In rodents without PPI deficits some authors describe absence of WIN effect on PPI (Bortolato et al., 2005; Brzózka et al., 2011), but others show that WIN disrupts PPI when administered systemically (Schneider and Koch, 2002; Wegener et al., 2008; Brosda et al., 2011), intra-prefrontal cortex, or intra-ventral hippocampus (Wegener et al., 2008). In addition, repeated administration of WIN during puberty induces PPI deficits that last until adulthood (Schneider and Koch, 2003; Schneider et al., 2005; Wegener and Koch, 2009; Klein et al., 2013). Therefore, WIN seems to present a psychotomimetic profile (supported also by its effects on other schizophrenia-like symptoms—Schneider and Koch, 2002; Pamplona and Takahashi, 2006; Wegener et al., 2008; Spano et al., 2010; Almeida et al., 2014), being unsuitable for schizophrenia patients.
The administration of the CB1 antagonist rimonabant worsens SHRs' PPI deficit, and does not alter PPI levels in WRs (Levin et al., 2014). The absence of rimonabant effects in WRs is corroborated by previous studies with animals without PPI impairments (Martin et al., 2003; Malone et al., 2004; Malone and Taylor, 2006; Ballmaier et al., 2007). In animal models of schizophrenia, while some studies show that rimonabant is able to counteract the PPI impairments (Malone et al., 2004; Nagai et al., 2006; Ballmaier et al., 2007), others show no effect (Martin et al., 2003; Malone and Taylor, 2006). In addition, clinical trials show that rimonabant induces significant psychiatric side effects, such as symptoms of depression and anxiety, and increases suicide-related adverse events (Christensen et al., 2007; Food and Drug Administration Advisory Committee, 2007; Topol et al., 2010). These data, thus, argue against the clinical use of this compound.
It should be noted that both a CB1/CB2 agonist and a CB1 antagonist provoke differential effects in WRs and SHRs (Levin et al., 2014). This information suggests that these rat strains display distinct endocannabinoid system functioning. Indeed, our group observed that SHRs present higher CB1 density in the prefrontal and anterior cingulate cortices when compared to WRs (Almeida et al., submitted), which is in accordance with data from schizophrenia patients (Dean et al., 2001; Zavitsanou et al., 2004; Newell et al., 2006; Wong et al., 2010; Dalton et al., 2011; Ceccarini et al., 2013).
AM 404 is a competitive and selective inhibitor of anandamide transportation, and therefore increases its extracellular levels. Anandamide is an endocannabinoid that acts as an agonist of CB1 and CB2 receptors and of vanilloid receptor 1 (TRPV1). When administered to WRs and SHRs, AM 404 did not modify their PPI levels (Levin et al., 2014). No other study has investigated the effects of AM 404 on PPI of animal models of schizophrenia. In control animals, one study shows absence of AM 404 effects on PPI in Sprague-Dawley rats (Bortolato et al., 2006), but another reveals that AM 404, either injected acutely or chronically, disrupts PPI in Swiss mice (Fernandez-Espejo and Galan-Rodriguez, 2004). Evidence, therefore, do not support the use of AM 404 as a strategy to treat sensorimotor processing deficits associated to schizophrenia.
It is worth mentioning that the doses of AM 404 used in our previous study have been shown to increase anandamide levels in plasma and brain regions of rats (Giuffrida et al., 2000; Bortolato et al., 2006). Clinical findings show that the levels of anandamide in the cerebrospinal fluid of non-medicated schizophrenia patients are negatively correlated to their psychotic symptoms (De Marchi et al., 2003; Giuffrida et al., 2004), and that prodromal individuals with lower levels of anandamide display a higher risk for transitting to psychosis earlier (Koethe et al., 2009). These results led some authors to suggest that anandamide plays a protective role in schizophrenia. The absence of AM 404 effects on the PPI of SHRs suggest that increasing anandamide levels is not sufficient to restore SHRs' PPI impairment.
Cannabidiol is one of the major constituent of cannabis, being the most abundant after Δ9-THC. Cannabidiol acts as an antagonist of the cannabinoid receptors CB1 and CB2 agonists and inhibits the reuptake and degradation of anandamide. It is also an antagonist of the orphan receptor GPR55 and an agonist of the serotonin receptor 5-HT1A and of the vanilloid receptors TRPV1 and TRPV2 (Izzo et al., 2009). When administered to SHRs, cannabidiol restores the PPI deficits (Levin et al., 2014). In fact, this compound has been shown to restore the PPI impairments provoked by MK-801 (Long et al., 2006), and by systemic or intra-accumbens administration of amphetamine (Pedrazzi et al., 2015). In addition, when administered repeatedly, cannabidiol prevents the PPI disruption induced by chronic administration of MK-801 (Gomes et al., 2015). Acute or chronic administration of psychostimulant drugs such as MK-801 and amphetamine are used to model schizophrenia. The results, thus, point to an antipsychotic profile of cannabidiol and to the use of this compound on the treatment of sensorimotor gating impairments seen in schizophrenia.
KEY CONCEPT 7. Cannabidiol
One of the over 60 compounds of Cannabis sativa, being the most abundant after Δ9-THC. Unlike Δ9-THC, cannabidiol is a non-psychotomimetic drug, and presents antipsychotic, anxiolytic, anti-inflammatory and neuroprotective actions.
In animals without PPI impairments, some authors describe that cannabidiol does not modify the PPI levels when administered acutely or chronically (Long et al., 2006; Gomes et al., 2015; Pedrazzi et al., 2015), while one study shows that it is able to disrupt PPI (Gururajan et al., 2011). Nonetheless, in our previous work, administration of cannabidiol increased PPI in WRs (Levin et al., 2014). Interestingly, typical and atypical antipsychotic drugs have been shown to increase PPI in WRs and Sprague-Dawley rats (Hoffman et al., 1993; Swerdlow and Geyer, 1993; Johansson et al., 1995; Depoortere et al., 1997a,b; Levin et al., 2011). This effect is mainly seen when PPI levels are below 50%, which is our case. Therefore, the effects of cannabidiol on PPI of WRs are in accordance with the antipsychotic profile suggested for this drug.
The mechanisms whereby cannabidiol modifies PPI are still to be elucidated. Our data suggest that the increase in PPI promoted by this drug is not only due to an increase in anandamide levels, since the anandamide uptake inhibitor AM 404 does not modify PPI (Levin et al., 2014). Likewise, it is unlikely that the cannabidiol's antagonism of CB1 receptors is the mechanism responsible for its effect on the PPI deficit of SHRs, given that the CB1 antagonist rimonabant promoted an opposite outcome (Levin et al., 2014). Regarding cannabidiol's action on CB1 receptors, it is noteworthy that although this compound is able to antagonize cannabinoid CB1/CB2 receptor agonists-induced stimulation in brain membranes, this effect is observed with KB values in the nanomolar range, way below the Ki for displacement of [3H]CP55940 from CB1 and CB2 (Pertwee, 2008). In addition, the effectiveness of cannabidiol in inhibiting [35S]GTPγS binding to brain membranes of wild-type mouse is not different from CB1−/− mouse (Pertwee, 2008). On the other hand, Long et al. (2006) described that cannabidiol's ability of restoring the PPI deficit induced by MK-801 is prevented by pretreatment with capsazepine, a TRPV1 antagonist. Thus, although other mechanisms of action cannot be disregarded, cannabidiol's effects on PPI seem to be at least partially related to its action on the vanilloid system.
Other animal studies, investigating additional schizophrenia-like behavioral paradigms, support cannabidiol's antipsychotic properties (Table 3). The administration of this compound is able to diminish the stereotyped behavior and the hyperlocomotion—behaviors that model the positive symptoms of schizophrenia—induced by psychotomimetic drugs (Zuardi et al., 1991; Moreira and Guimaraes, 2005; Long et al., 2010; Gururajan et al., 2012). Cannabidiol also restores drug-induced impairments in social interaction (Malone et al., 2009; Gururajan et al., 2012), and counteracts the deficit in contextual fear conditioning displayed by the SHRs (Levin et al., 2012).
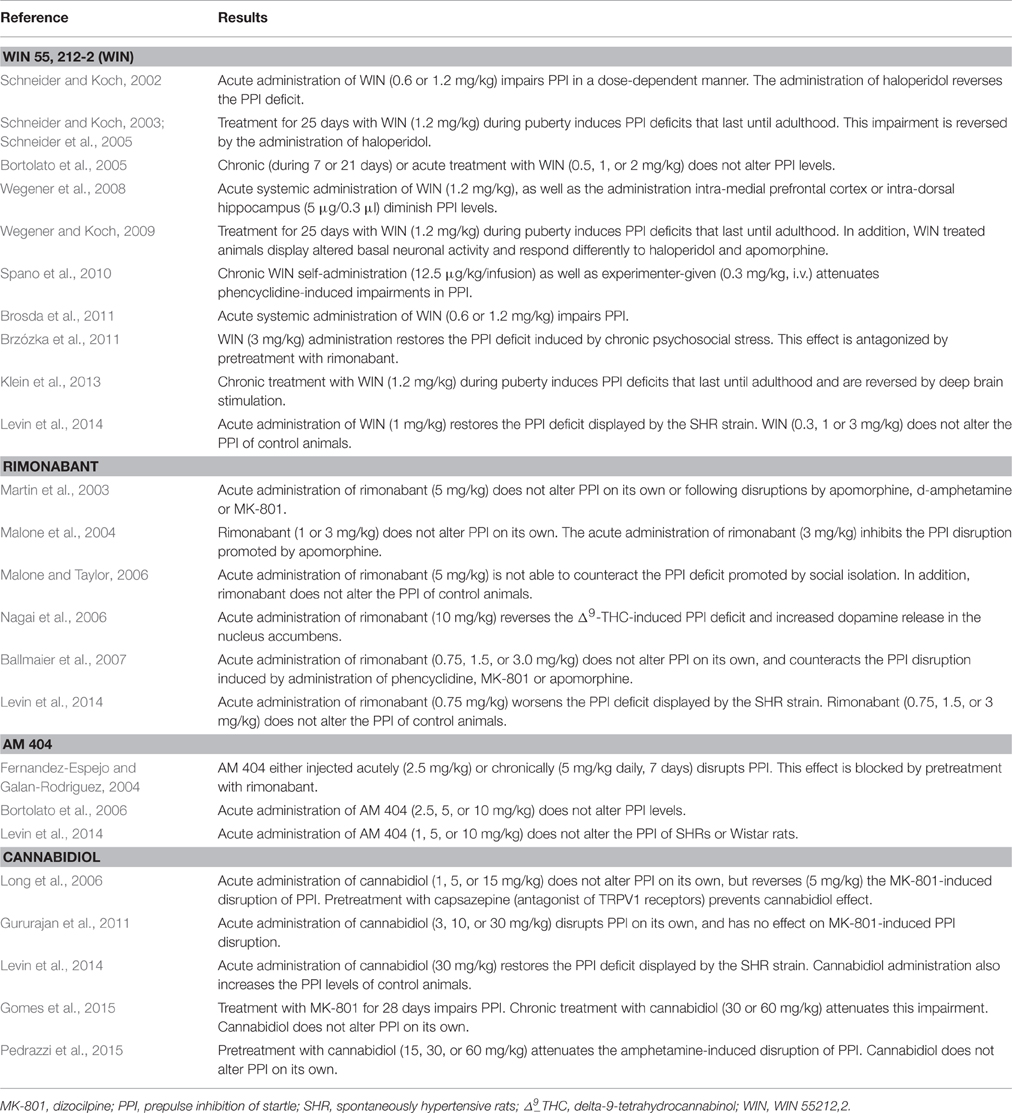
Table 2. Summary of the studies investigating the effects of cannabinoid drugs on the PPI of animal models.

Table 3. Antipsychotic effects of cannabidiol in psychiatric patients and in schizophrenia-like behaviors in animal models (for PPI, see Table 2).
The promising aforementioned pre-clinical data led to clinical studies (Table 3). In healthy volunteers, cannabidiol is able to attenuate the psychotic symptoms induced by the administration of psychotomimetic drugs (Karniol et al., 1974; Zuardi et al., 1982; Bhattacharyya et al., 2010). In a case-study, this cannabinoid significantly reduced schizophrenia symptoms in the Brief Psychiatric Rating Scale (BPRS), in a way superior to the typical antipsychotic drug haloperidol (Zuardi et al., 1995). Moreover, an open-label study with Parkinson's disease patients showed that cannabidiol administration, in addition to their usual treatment, decreases psychotic symptoms—evaluated by BPRS and by the Parkinson Psychosis Questionnaire—without worsening their motor function (Zuardi et al., 2009). Also, several studies suggest that cannabidiol is safe in humans and animals, and does not alter physiological parameters (blood pressure, heart rate and body temperature) or induce motor and psychological side effects (Bergamaschi et al., 2011).
Thereafter, a double-blind controlled clinical trial with schizophrenia patients was performed: treatment with cannabidiol, for 4 weeks, decreased patients symptoms—evaluated by BPRS and Positive and Negative Syndrome Scale (PANSS)—in a way non-inferior to amisulpride, one of the most effective antipsychotic drugs currently in use. Interestingly, the cannabinoid induced fewer side effects (weight gain, changes in prolactin levels and extrapyramidal symptoms) than amisulpride, and treatment with cannabidiol, but not with amisulpride, led to an increase in the levels of anandamide in serum that was associated with the decrease of psychotic symptoms (Leweke et al., 2012).
All these encouraging clinical and pre-clinical data led to a recent proof-of-concept study by GW Pharmaceuticals. The phase IIa included 88 schizophrenia patients only partially responsive to standard antipsychotic treatment, who received cannabidiol or placebo in addition to their antipsychotic medication for 6 weeks. Cannabidiol was consistently superior to placebo in attenuating the schizophrenia symptoms, and did not induce serious adverse events (GW Pharmaceuticals, 2015).
Taken as a whole, data regarding the effects of cannabinoid drugs on PPI reinforce the involvement of the endocannabinoid system in the sensorimotor gating functioning and in the pathophysiology of schizophrenia. Among the drugs that act on the endocannabinoid system, pre-clinical and the subsequent clinical data point to cannabidiol as the most promising compound for treating schizophrenia symptoms without inducing significant side effects. Nevertheless, most of the clinical evidence that suggests cannabidiol as a new antipsychotic agent or adjunctive treatment does not investigate specifically sensorimotor gating/ PPI deficits. Accordingly, data from pre-clinical studies using animal models, like our previous one (Levin et al., 2014), are fundamental to support future clinical studies focused on these deficits in schizophrenia patients.
Author Contributions
All authors listed, have made substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
This work was supported by grants of “Fundação de Amparo à Pesquisa do Estado de São Paulo–FAPESP” and of “Conselho Nacional do Desenvolvimento Científico e Tecnológico–CAPES.”
Conflict of Interest Statement
JH, AZ, and JC are co-inventors of the patent “Fluorinated CBD compounds, compositions and uses thereof. Pub. No.: WO/2014/108899. International Application No.: PCT/IL2014/050023.”
The Other authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Author Biography

References
Abi-Dargham, A. (2014). Schizophrenia: overview and dopamine dysfunction. J. Clin. Psychiatry 75:e31. doi: 10.4088/JCP.13078tx2c
Ahmari, S. E., Risbrough, V. B., Geyer, M. A., and Simpson, H. B. (2012). Impaired sensorimotor gating in unmedicated adults with obsessive-compulsive disorder. Neuropsychopharmacology 37, 1216–1223. doi: 10.1038/npp.2011.308
Almeida, V., Levin, R., Peres, F. F., Niigaki, S. T., Calzavara, M. B., Zuardi, A. W., et al. (2013). Cannabidiol exhibits anxiolytic but not antipsychotic property evaluated in the social interaction test. Prog. Neuropsychopharmacol. Biol. Psychiatry 41, 30–35. doi: 10.1016/j.pnpbp.2012.10.024
Almeida, V., Peres, F. F., Levin, R., Suiama, M. A., Calzavara, M. B., Zuardi, A. W., et al. (2014). Effects of cannabinoid and vanilloid drugs on positive and negative-like symptoms on an animal model of schizophrenia: the SHR strain. Schizophr. Res. 153, 150–159. doi: 10.1016/j.schres.2014.01.039
Amini, B., Yang, P. B., Swann, A. C., and Dafny, N. (2004). Differential locomotor responses in male rats from three strains to acute methylphenidate. Int. J. Neurosci. 114, 1063–1084. doi: 10.1080/00207450490475526
Ballmaier, M., Bortolato, M., Rizzetti, C., Zoli, M., Gessa, G., Heinz, A., et al. (2007). Cannabinoid receptor antagonists counteract sensorimotor gating deficits in the phencyclidine model of psychosis. Neuropsychopharmacology 32, 2098–2107. doi: 10.1038/sj.npp.1301344
Barron, E., Yang, P. B., Swann, A. C., and Dafny, N. (2009). Adolescent and adult male spontaneous hyperactive rats (SHR) respond differently to acute and chronic methylphenidate (Ritalin). Int. J. Neurosci. 119, 40–58. doi: 10.1080/00207450802330546
Bergamaschi, M. M., Queiroz, R. H., Zuardi, A. W., and Crippa, J. A. (2011). Safety and side effects of cannabidiol, a Cannabis sativa constituent. Curr. Drug Saf. 6, 237–249. doi: 10.2174/157488611798280924
Bhattacharyya, S., Morrison, P. D., Fusar-Poli, P., Martin-Santos, R., Borgwardt, S., Winton-Brown, T., et al. (2010). Opposite effects of delta-9-tetrahydrocannabinol and cannabidiol on human brain function and psychopathology. Neuropsychopharmacology 35, 764–774. doi: 10.1038/npp.2009.184
Bizot, J. C., Chenault, N., Houze, B., Herpin, A., David, S., Pothion, S., et al. (2007). Methylphenidate reduces impulsive behaviour in juvenile Wistar rats, but not in adult Wistar, SHR and WKY rats. Psychopharmacology (Berl.) 193, 215–223. doi: 10.1007/s00213-007-0781-4
Bortolato, M., Aru, G. N., Frau, R., Orrù, M., Luckey, G. C., Boi, G., et al. (2005). The CB receptor agonist WIN 55,212-2 fails to elicit disruption of prepulse inhibition of the startle in Sprague-Dawley rats. Psychopharmacology (Berl.) 177, 264–271. doi: 10.1007/s00213-004-1941-4
Bortolato, M., Campolongo, P., Mangieri, R. A., Scattoni, M. L., Frau, R., Trezza, V., et al. (2006). Anxiolytic-like properties of the anandamide transport inhibitor AM404. Neuropsychopharmacology 31, 2652–2659. doi: 10.1038/sj.npp.1301061
Braff, D. L., Geyer, M. A., and Swerdlow, N. R. (2001). Human studies of prepulse inhibition of startle: normal subjects, patient groups, and pharmacological studies. Psychopharmacology (Berl.) 156, 234–258. doi: 10.1007/s002130100810
Briles, J. J., Rosenberg, D. R., Brooks, B. A., Roberts, M. W., and Diwadkar, V. A. (2012). Review of the safety of second-generation antipsychotics: are they really “atypically” safe for youth and adults? Prim. Care Companion CNS Disord. 14:11r01298. doi: 10.4088/PCC.11r01298
Brosda, J., Hayn, L., Klein, C., Koch, M., Meyer, C., Schallhorn, R., et al. (2011). Pharmacological and parametrical investigation of prepulse inhibition of startle and prepulse elicited reactions in Wistar rats. Pharmacol. Biochem. Behav. 99, 22–28. doi: 10.1016/j.pbb.2011.03.017
Brzózka, M. M., Fischer, A., Falkai, P., and Havemann-Reinecke, U. (2011). Acute treatment with cannabinoid receptor agonist WIN55212.2 improves prepulse inhibition in psychosocially stressed mice. Behav. Brain Res. 218, 280–287. doi: 10.1016/j.bbr.2010.11.003
Calzavara, M. B., Levin, R., Medrano, W. A., Almeida, V., Sampaio, A. P., Barone, L. C., et al. (2011). Effects of antipsychotics and amphetamine on social behaviors in spontaneously hypertensive rats. Behav. Brain Res. 225, 15–22. doi: 10.1016/j.bbr.2011.06.026
Calzavara, M. B., Medrano, W. A., Levin, R., Kameda, S. R., Andersen, M. L., Tufik, S., et al. (2009). Neuroleptic drugs revert the contextual fear conditioning deficit presented by spontaneously hypertensive rats: a potential animal model of emotional context processing in schizophrenia? Schizophr. Bull. 35, 748–759. doi: 10.1093/schbul/sbn006
Ceccarini, J., De Hert, M., Van Winkel, R., Peuskens, J., Bormans, G., Kranaster, L., et al. (2013). Increased ventral striatal CB1 receptor binding is related to negative symptoms in drug-free patients with schizophrenia. Neuroimage 79, 304–312. doi: 10.1016/j.neuroimage.2013.04.052
Chavarría-Siles, I., Contreras-Rojas, J., Hare, E., Walss-Bass, C., Quezada, P., Dassori, A., et al. (2008). Cannabinoid receptor 1 gene (CNR1) and susceptibility to a quantitative phenotype for hebephrenic schizophrenia. Am. J. Med. Genet. B Neuropsychiatr. Genet. 147, 279–284. doi: 10.1002/ajmg.b.30592
Christensen, R., Kristensen, P. K., Bartels, E. M., Bliddal, H., and Astrup, A. (2007). Efficacy and safety of the weight-loss drug rimonabant: a meta-analysis of randomised trials. Lancet 370, 1706–1713. doi: 10.1016/S0140-673661721-8
Dalton, V. S., Long, L. E., Weickert, C. S., and Zavitsanou, K. (2011). Paranoid schizophrenia is characterized by increased CB1 receptor binding in the dorsolateral prefrontal cortex. Neuropsychopharmacology 36, 1620–1630. doi: 10.1038/npp.2011.43
Dean, B., Sundram, S., Bradbury, R., Scarr, E., and Copolov, D. (2001). Studies on [3H]CP-55940 binding in the human central nervous system: regional specific changes in density of cannabinoid-1 receptors associated with schizophrenia and cannabis use. Neuroscience 103, 9–15. doi: 10.1016/S0306-4522(00)00552-2
De Marchi, N., De Petrocellis, L., Orlando, P., Daniele, F., Fezza, F., and Di Marzo, V. (2003). Endocannabinoid signalling in the blood of patients with schizophrenia. Lipids Health Dis. 2:5. doi: 10.1186/1476-511X-2-5
Depoortere, R., Perrault, G., and Sanger, D. J. (1997a). Potentiation of prepulse inhibition of the startle reflex in rats: pharmacological evaluation of the procedure as a model for detecting antipsychotic activity. Psychopharmacology (Berl.) 132, 366–374.
Depoortere, R., Perrault, G., and Sanger, D. J. (1997b). Some, but not all, antipsychotic drugs potentiate a low level of prepulse inhibition shown by rats of the Wistar strain. Behav. Pharmacol. 8, 364–372.
Diana, M. C., Santoro, M. L., Xavier, G., Santos, C. M., Spindola, L. N., Moretti, P. N., et al. (2015). Low expression of Gria1 and Grin1 glutamate receptors in the nucleus accumbens of Spontaneously Hypertensive Rats (SHR). Psychiatry Res. 229, 690–694. doi: 10.1016/j.psychres.2015.08.021
D'Souza, D. C., Abi-Saab, W. M., Madonick, S., Forselius-Bielen, K., Doersch, A., Braley, G., et al. (2005). Delta-9-tetrahydrocannabinol effects in schizophrenia: implications for cognition, psychosis, and addiction. Biol. Psychiatry 57, 594–608. doi: 10.1016/j.biopsych.2004.12.006
D'Souza, D. C., Perry, E., MacDougall, L., Ammerman, Y., Cooper, T., Wu, Y. T., et al. (2004). The psychotomimetic effects of intravenous delta-9-tetrahydrocannabinol in healthy individuals: implications for psychosis. Neuropsychopharmacology 29, 1558–1572. doi: 10.1038/sj.npp.1300496
Fernandez-Espejo, E., and Galan-Rodriguez, B. (2004). Sensorimotor gating in mice is disrupted after AM404, an anandamide reuptake and degradation inhibitor. Psychopharmacology (Berl.) 175, 220–224. doi: 10.1007/s00213-004-1851-5
File, S. E., and Seth, P. (2003). A review of 25 years of the social interaction test. Eur. J. Pharmacol. 463, 35–53. doi: 10.1016/S0014-2999(03)01273-1
Food Drug Administration Advisory Committee, U. (2007). FDA Briefing Document: Zimulti (Rimonabant) Tablets, 20 mg. Rockville, MD: FSA.
Giuffrida, A., Leweke, F. M., Gerth, C. W., Schreiber, D., Koethe, D., Faulhaber, J., et al. (2004). Cerebrospinal anandamide levels are elevated in acute schizophrenia and are inversely correlated with psychotic symptoms. Neuropsychopharmacology 29, 2108–2114. doi: 10.1038/sj.npp.1300558
Giuffrida, A., Rodriguez de Fonseca, F., Nava, F., Loubet-Lescoulíe, P., and Piomelli, D. (2000). Elevated circulating levels of anandamide after administration of the transport inhibitor, AM404. Eur. J. Pharmacol. 408, 161–168. doi: 10.1016/S0014-2999(00)00786-X
Gomes, F. V., Issy, A. C., Ferreira, F. R., Viveros, M. P., Del Bel, E. A., and Guimarães, F. S. (2015). Cannabidiol attenuates sensorimotor gating disruption and molecular changes induced by chronic antagonism of NMDA receptors in mice. Int. J. Neuropsychopharmacol. 18, 1–10. doi: 10.1093/ijnp/pyu041
Grech, A., Van Os, J., Jones, P. B., Lewis, S. W., and Murray, R. M. (2005). Cannabis use and outcome of recent onset psychosis. Eur. Psychiatry 20, 349–353. doi: 10.1016/j.eurpsy.2004.09.013
Gururajan, A., Taylor, D. A., and Malone, D. T. (2011). Effect of cannabidiol in a MK-801-rodent model of aspects of schizophrenia. Behav. Brain Res. 222, 299–308. doi: 10.1016/j.bbr.2011.03.053
Gururajan, A., Taylor, D. A., and Malone, D. T. (2012). Cannabidiol and clozapine reverse MK-801-induced deficits in social interaction and hyperactivity in Sprague-Dawley rats. J. Psychopharmacol. 26, 1317–1332. doi: 10.1177/0269881112441865
GW Pharmaceuticals (2015). GW PharmaceuticalsAnnounces Positive Proof of Concept Data in Schizophrenia. London: GWP Press Release.
Hasan, A., Falkai, P., Wobrock, T., Lieberman, J., Glenthoj, B., Gattaz, W. F., et al. (2012). World Federation of Societies of Biological Psychiatry (WFSBP) guidelines for biological treatment of schizophrenia, part 1: update 2012 on the acute treatment of schizophrenia and the management of treatment resistance. World J. Biol. Psychiatry 13, 318–378. doi: 10.3109/15622975.2012.696143
Hoffman, D. C., Donovan, H., and Cassella, J. V. (1993). The effects of haloperidol and clozapine on the disruption of sensorimotor gating induced by the noncompetitive glutamate antagonist MK-801. Psychopharmacology (Berl.) 111, 339–344.
Ishiguro, H., Horiuchi, Y., Ishikawa, M., Koga, M., Imai, K., Suzuki, Y., et al. (2010). Brain cannabinoid CB2 receptor in schizophrenia. Biol. Psychiatry 67, 974–982. doi: 10.1016/j.biopsych.2009.09.024
Izzo, A. A., Borrelli, F., Capasso, R., Di Marzo, V., and Mechoulam, R. (2009). Non-psychotropic plant cannabinoids: new therapeutic opportunities from an ancient herb. Trends Pharmacol. Sci. 30, 515–527. doi: 10.1016/j.tips.2009.07.006
Johansson, C., Jackson, D. M., Zhang, J., and Svensson, L. (1995). Prepulse inhibition of acoustic startle, a measure of sensorimotor gating: effects of antipsychotics and other agents in rats. Pharmacol. Biochem. Behav. 52, 649–654.
Johnson, M. L., Ely, D. L., and Turner, M. E. (1992). Genetic divergence between the Wistar-Kyoto rat and the spontaneously hypertensive rat. Hypertension 19, 425–427.
Jones, C. A., Watson, D. J., and Fone, K. C. (2011). Animal models of schizophrenia. Br. J. Pharmacol. 164, 1162–1194. doi: 10.1111/j.1476-5381.2011.01386.x
Karniol, I. G., Shirakawa, I., Kasinski, N., Pfeferman, A., and Carlini, E. A. (1974). Cannabidiol interferes with the effects of delta 9 - tetrahydrocannabinol in man. Eur. J. Pharmacol. 28, 172–177.
Klein, J., Hadar, R., Gotz, T., Manner, A., Eberhardt, C., Baldassarri, J., et al. (2013). Mapping brain regions in which deep brain stimulation affects schizophrenia-like behavior in two rat models of schizophrenia. Brain Stimul. 6, 490–499. doi: 10.1016/j.brs.2012.09.004
Koethe, D., Giuffrida, A., Schreiber, D., Hellmich, M., Schultze-Lutter, F., Ruhrmann, S., et al. (2009). Anandamide elevation in cerebrospinal fluid in initial prodromal states of psychosis. Br. J. Psychiatry 194, 371–372. doi: 10.1192/bjp.bp.108.053843
Kumari, V., Peters, E. R., Fannon, D., Premkumar, P., Aasen, I., Cooke, M. A., et al. (2008). Uncontrollable voices and their relationship to gating deficits in schizophrenia. Schizophr. Res. 101, 185–194. doi: 10.1016/j.schres.2007.12.481
Kumari, V., Soni, W., and Sharma, T. (1999). Normalization of information processing deficits in schizophrenia with clozapine. Am. J. Psychiatry 156, 1046–1051. doi: 10.1176/ajp.156.7.1046
Laruelle, M., Abi-Dargham, A., Gil, R., Kegeles, L., and Innis, R. (1999). Increased dopamine transmission in schizophrenia: relationship to illness phases. Biol. Psychiatry 46, 56–72.
Leumann, L., Feldon, J., Vollenweider, F. X., and Ludewig, K. (2002). Effects of typical and atypical antipsychotics on prepulse inhibition and latent inhibition in chronic schizophrenia. Biol. Psychiatry 52, 729–739. doi: 10.1016/S0006-3223(02)01344-6
Levin, R., Almeida, V., Peres, F. F., Calzavara, M. B., da Silva, N. D., Suiama, M. A., et al. (2012). Antipsychotic profile of cannabidiol and rimonabant in an animal model of emotional context processing in schizophrenia. Curr. Pharm. Des. 18, 4960–4965. doi: 10.2174/138161212802884735
Levin, R., Calzavara, M. B., Santos, C. M., Medrano, W. A., Niigaki, S. T., and Abílio, V. C. (2011). Spontaneously Hypertensive Rats (SHR) present deficits in prepulse inhibition of startle specifically reverted by clozapine. Prog. Neuropsychopharmacol. Biol. Psychiatry 35, 1748–1752. doi: 10.1016/j.pnpbp.2011.06.003
Levin, R., Peres, F. F., Almeida, V., Calzavara, M. B., Zuardi, A. W., Hallak, J. E., et al. (2014). Effects of cannabinoid drugs on the deficit of prepulse inhibition of startle in an animal model of schizophrenia: the SHR strain. Front. Pharmacol. 5:10. doi: 10.3389/fphar.2014.00010
Leweke, F. M., Giuffrida, A., Koethe, D., Schreiber, D., Nolden, B. M., Kranaster, L., et al. (2007). Anandamide levels in cerebrospinal fluid of first-episode schizophrenic patients: impact of cannabis use. Schizophr. Res. 94, 29–36. doi: 10.1016/j.schres.2007.04.025
Leweke, F. M., Giuffrida, A., Wurster, U., Emrich, H. M., and Piomelli, D. (1999). Elevated endogenous cannabinoids in schizophrenia. Neuroreport 10, 1665–1669.
Leweke, F. M., Piomelli, D., Pahlisch, F., Muhl, D., Gerth, C. W., Hoyer, C., et al. (2012). Cannabidiol enhances anandamide signaling and alleviates psychotic symptoms of schizophrenia. Transl. Psychiatry 2:e94. doi: 10.1038/tp.2012.15
Linszen, D. H., Dingemans, P. M., and Lenior, M. E. (1994). Cannabis abuse and the course of recent-onset schizophrenic disorders. Arch. Gen. Psychiatry 51, 273–279.
Lipska, B. K., and Weinberger, D. R. (2000). To model a psychiatric disorder in animals: schizophrenia as a reality test. Neuropsychopharmacology 23, 223–239. doi: 10.1016/S0893-133X00137-8
Long, L. E., Chesworth, R., Huang, X. F., McGregor, I. S., Arnold, J. C., and Karl, T. (2010). A behavioural comparison of acute and chronic Delta9-tetrahydrocannabinol and cannabidiol in C57BL/6JArc mice. Int. J. Neuropsychopharmacol. 13, 861–876. doi: 10.1017/S1461145709990605
Long, L. E., Malone, D. T., and Taylor, D. A. (2006). Cannabidiol reverses MK-801-induced disruption of prepulse inhibition in mice. Neuropsychopharmacology 31, 795–803. doi: 10.1038/sj.npp.1300838
MacDonald, A. W., and Schulz, S. C. (2009). What we know: findings that every theory of schizophrenia should explain. Schizophr. Bull. 35, 493–508. doi: 10.1093/schbul/sbp017
Malone, D. T., Jongejan, D., and Taylor, D. A. (2009). Cannabidiol reverses the reduction in social interaction produced by low dose Delta-tetrahydrocannabinol in rats. Pharmacol. Biochem. Behav. 93, 91–96. doi: 10.1016/j.pbb.2009.04.010
Malone, D. T., Long, L. E., and Taylor, D. A. (2004). The effect of SR 141716 and apomorphine on sensorimotor gating in Swiss mice. Pharmacol. Biochem. Behav. 77, 839–845. doi: 10.1016/j.pbb.2004.02.010
Malone, D. T., and Taylor, D. A. (2006). The effect of Delta9-tetrahydrocannabinol on sensorimotor gating in socially isolated rats. Behav. Brain Res. 166, 101–109. doi: 10.1016/j.bbr.2005.07.009
Maren, S., Phan, K. L., and Liberzon, I. (2013). The contextual brain: implications for fear conditioning, extinction and psychopathology. Nat. Rev. Neurosci. 14, 417–428. doi: 10.1038/nrn3492
Martin, R. S., Secchi, R. L., Sung, E., Lemaire, M., Bonhaus, D. W., Hedley, L. R., et al. (2003). Effects of cannabinoid receptor ligands on psychosis-relevant behavior models in the rat. Psychopharmacology (Berl.) 165, 128–135. doi: 10.1007/s00213-002-1240-x
Martinez-Gras, I., Rubio, G., del Manzano, B. A., Rodriguez-Jimenez, R., Garcia-Sanchez, F., Bagney, A., et al. (2009). The relationship between prepulse inhibition and general psychopathology in patients with schizophrenia treated with long-acting risperidone. Schizophr. Res. 115, 215–221. doi: 10.1016/j.schres.2009.09.035
Matheson, S. L., Shepherd, A. M., Laurens, K. R., and Carr, V. J. (2011). A systematic meta-review grading the evidence for non-genetic risk factors and putative antecedents of schizophrenia. Schizophr. Res. 133, 133–142. doi: 10.1016/j.schres.2011.09.020
Minassian, A., Feifel, D., and Perry, W. (2007). The relationship between sensorimotor gating and clinical improvement in acutely ill schizophrenia patients. Schizophr. Res. 89, 225–231. doi: 10.1016/j.schres.2006.08.006
Moreira, F. A., and Guimarães, F. S. (2005). Cannabidiol inhibits the hyperlocomotion induced by psychotomimetic drugs in mice. Eur. J. Pharmacol. 512, 199–205. doi: 10.1016/j.ejphar.2005.02.040
Morrison, P. D., Zois, V., McKeown, D. A., Lee, T. D., Holt, D. W., Powell, J. F., et al. (2009). The acute effects of synthetic intravenous Delta9-tetrahydrocannabinol on psychosis, mood and cognitive functioning. Psychol. Med. 39, 1607–1616. doi: 10.1017/S0033291709005522
Nagai, H., Egashira, N., Sano, K., Ogata, A., Mizuki, A., Mishima, K., et al. (2006). Antipsychotics improve Delta9-tetrahydrocannabinol-induced impairment of the prepulse inhibition of the startle reflex in mice. Pharmacol. Biochem. Behav. 84, 330–336. doi: 10.1016/j.pbb.2006.05.018
Newell, K. A., Deng, C., and Huang, X. F. (2006). Increased cannabinoid receptor density in the posterior cingulate cortex in schizophrenia. Exp. Brain Res. 172, 556–560. doi: 10.1007/s00221-006-0503-x
Okamoto, K., and Aoki, K. (1963). Development of a strain of spontaneously hypertensive rats. Jpn. Circ. J. 27, 282–293.
Oranje, B., Van Oel, C. J., Gispen-De Wied, C. C., Verbaten, M. N., and Kahn, R. S. (2002). Effects of typical and atypical antipsychotics on the prepulse inhibition of the startle reflex in patients with schizophrenia. J. Clin. Psychopharmacol. 22, 359–365. doi: 10.1097/00004714-200208000-00005
Overstreet, D. H. (2012). Modeling depression in animal models. Methods Mol. Biol. 829, 125–144. doi: 10.1007/978-1-61779-458-2_7
Pamplona, F. A., and Takahashi, R. N. (2006). WIN 55212-2 impairs contextual fear conditioning through the activation of CB1 cannabinoid receptors. Neurosci. Lett. 397, 88–92. doi: 10.1016/j.neulet.2005.12.026
Pedrazzi, J. F., Issy, A. C., Gomes, F. V., Guimarães, F. S., and Del-Bel, E. A. (2015). Cannabidiol effects in the prepulse inhibition disruption induced by amphetamine. Psychopharmacology (Berl.) 232, 3057–3065. doi: 10.1007/s00213-015-3945-7
Perry, W., and Braff, D. L. (1994). Information-processing deficits and thought disorder in schizophrenia. Am. J. Psychiatry 151, 363–367. doi: 10.1176/ajp.151.3.363
Perry, W., Geyer, M. A., and Braff, D. L. (1999). Sensorimotor gating and thought disturbance measured in close temporal proximity in schizophrenic patients. Arch. Gen. Psychiatry 56, 277–281.
Perry, W., Minassian, A., Feifel, D., and Braff, D. L. (2001). Sensorimotor gating deficits in bipolar disorder patients with acute psychotic mania. Biol. Psychiatry 50, 418–424. doi: 10.1016/S0006-3223(01)01184-2
Pertwee, R. G. (2008). The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: delta9-tetrahydrocannabinol, cannabidiol and delta9-tetrahydrocannabivarin. Br. J. Pharmacol. 153, 199–215. doi: 10.1038/sj.bjp.0707442
Schneider, M., Drews, E., and Koch, M. (2005). Behavioral effects in adult rats of chronic prepubertal treatment with the cannabinoid receptor agonist WIN 55,212-2. Behav. Pharmacol. 16, 447–454. doi: 10.1097/00008877-200509000-00018
Schneider, M., and Koch, M. (2002). The cannabinoid agonist WIN 55,212-2 reduces sensorimotor gating and recognition memory in rats. Behav. Pharmacol. 13, 29–37. doi: 10.1097/00008877-200202000-00003
Schneider, M., and Koch, M. (2003). Chronic pubertal, but not adult chronic cannabinoid treatment impairs sensorimotor gating, recognition memory, and the performance in a progressive ratio task in adult rats. Neuropsychopharmacology 28, 1760–1769. doi: 10.1038/sj.npp.1300225
Spano, M. S., Fadda, P., Frau, R., Fattore, L., and Fratta, W. (2010). Cannabinoid self-administration attenuates PCP-induced schizophrenia-like symptoms in adult rats. Eur. Neuropsychopharmacol. 20, 25–36. doi: 10.1016/j.euroneuro.2009.09.004
St Lezin, E., Simonet, L., Pravenec, M., and Kurtz, T. W. (1992). Hypertensive strains and normotensive ‘control’ strains. How closely are they related? Hypertension 19, 419–424.
Swerdlow, N. R., and Geyer, M. A. (1993). Clozapine and haloperidol in an animal model of sensorimotor gating deficits in schizophrenia. Pharmacol. Biochem. Behav. 44, 741–744.
Swerdlow, N. R., Karban, B., Ploum, Y., Sharp, R., Geyer, M. A., and Eastvold, A. (2001). Tactile prepuff inhibition of startle in children with Tourette's syndrome: in search of an “fMRI-friendly” startle paradigm. Biol. Psychiatry 50, 578–585. doi: 10.1016/S0006-3223(01)01164-7
Swerdlow, N. R., Light, G. A., Cadenhead, K. S., Sprock, J., Hsieh, M. H., and Braff, D. L. (2006). Startle gating deficits in a large cohort of patients with schizophrenia: relationship to medications, symptoms, neurocognition, and level of function. Arch. Gen. Psychiatry 63, 1325–1335. doi: 10.1001/archpsyc.63.12.1325
Swerdlow, N. R., Paulsen, J., Braff, D. L., Butters, N., Geyer, M. A., and Swenson, M. R. (1995). Impaired prepulse inhibition of acoustic and tactile startle response in patients with Huntington's disease. J. Neurol. Neurosurg. Psychiatr. 58, 192–200.
Swerdlow, N. R., Weber, M., Qu, Y., Light, G. A., and Braff, D. L. (2008). Realistic expectations of prepulse inhibition in translational models for schizophrenia research. Psychopharmacology (Berl.) 199, 331–388. doi: 10.1007/s00213-008-1072-4
Topol, E. J., Bousser, M. G., Fox, K. A., Creager, M. A., Despres, J. P., Easton, J. D., et al. (2010). Rimonabant for prevention of cardiovascular events (CRESCENDO): a randomised, multicentre, placebo-controlled trial. Lancet 376, 517–523. doi: 10.1016/S0140-673660935-X
Ujike, H., Takaki, M., Nakata, K., Tanaka, Y., Takeda, T., Kodama, M., et al. (2002). CNR1, central cannabinoid receptor gene, associated with susceptibility to hebephrenic schizophrenia. Mol. Psychiatry 7, 515–518. doi: 10.1038/sj.mp.4001029
van den Bergh, F. S., Bloemarts, E., Chan, J. S., Groenink, L., Olivier, B., and Oosting, R. S. (2006). Spontaneously hypertensive rats do not predict symptoms of attention-deficit hyperactivity disorder. Pharmacol. Biochem. Behav. 83, 380–390. doi: 10.1016/j.pbb.2006.02.018
van Os, J., and Kapur, S. (2009). Schizophrenia. Lancet 374, 635–645. doi: 10.1016/S0140-673660995-8
Wegener, N., and Koch, M. (2009). Behavioural disturbances and altered Fos protein expression in adult rats after chronic pubertal cannabinoid treatment. Brain Res. 1253, 81–91. doi: 10.1016/j.brainres.2008.11.081
Wegener, N., Kuhnert, S., Thüns, A., Roese, R., and Koch, M. (2008). Effects of acute systemic and intra-cerebral stimulation of cannabinoid receptors on sensorimotor gating, locomotion and spatial memory in rats. Psychopharmacology (Berl.) 198, 375–385. doi: 10.1007/s00213-008-1148-1
Weike, A. I., Bauer, U., and Hamm, A. O. (2000). Effective neuroleptic medication removes prepulse inhibition deficits in schizophrenia patients. Biol. Psychiatry 47, 61–70. doi: 10.1016/S0006-3223(99)00229-2
Wong, D. F., Kuwabara, H., Horti, A. G., Raymont, V., Brasic, J., Guevara, M., et al. (2010). Quantification of cerebral cannabinoid receptors subtype 1 (CB1) in healthy subjects and schizophrenia by the novel PET radioligand [11C]OMAR. Neuroimage 52, 1505–1513. doi: 10.1016/j.neuroimage.2010.04.034
Wynn, J. K., Green, M. F., Sprock, J., Light, G. A., Widmark, C., Reist, C., et al. (2007). Effects of olanzapine, risperidone and haloperidol on prepulse inhibition in schizophrenia patients: a double-blind, randomized controlled trial. Schizophr. Res. 95, 134–142. doi: 10.1016/j.schres.2007.05.039
Yang, P. B., Swann, A. C., and Dafny, N. (2006). Acute and chronic methylphenidate dose-response assessment on three adolescent male rat strains. Brain Res. Bull. 71, 301–310. doi: 10.1016/j.brainresbull.2006.09.019
Zavitsanou, K., Garrick, T., and Huang, X. F. (2004). Selective antagonist [3H]SR141716A binding to cannabinoid CB1 receptors is increased in the anterior cingulate cortex in schizophrenia. Prog. Neuropsychopharmacol. Biol. Psychiatry 28, 355–360. doi: 10.1016/j.pnpbp.2003.11.005
Zuardi, A. W., Crippa, J. A., Hallak, J. E., Pinto, J. P., Chagas, M. H., Rodrigues, G. G., et al. (2009). Cannabidiol for the treatment of psychosis in Parkinson's disease. J. Psychopharmacol. 23, 979–983. doi: 10.1177/0269881108096519
Zuardi, A. W., Morais, S. L., Guimarães, F. S., and Mechoulam, R. (1995). Antipsychotic effect of cannabidiol. J. Clin. Psychiatry 56, 485–486.
Zuardi, A. W., Rodrigues, J. A., and Cunha, J. M. (1991). Effects of cannabidiol in animal models predictive of antipsychotic activity. Psychopharmacology (Berl.) 104, 260–264.
Keywords: prepulse inhibition of startle reflex, animal models, schizophrenia, cannabidiol, endocannabinoid system, SHR strain
Citation: Peres FF, Levin R, Almeida V, Zuardi AW, Hallak JE, Crippa JA and Abilio VC (2016) Cannabidiol, among Other Cannabinoid Drugs, Modulates Prepulse Inhibition of Startle in the SHR Animal Model: Implications for Schizophrenia Pharmacotherapy. Front. Pharmacol. 7:303. doi: 10.3389/fphar.2016.00303
Received: 14 March 2016; Accepted: 26 August 2016;
Published: 09 September 2016.
Edited by:
Andrew C. McCreary, Janssen Prevention Center, NetherlandsCopyright © 2016 Peres, Levin, Almeida, Zuardi, Hallak, Crippa and Abilio. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: dmFuYWJpbGlvQGdtYWlsLmNvbQ==