 Shivankar Agrawal
Shivankar Agrawal Alok Adholeya
Alok Adholeya Sunil K. Deshmukh
Sunil K. Deshmukh- TERI–Deakin Nano Biotechnology Centre, The Energy and Resources Institute, New Delhi, India
Marine biodiversity is recognized by a wide and unique array of fascinating structures. The complex associations of marine microorganisms, especially with sponges, bryozoans, and tunicates, make it extremely difficult to define the biosynthetic source of marine natural products or to deduce their ecological significance. Marine sponges and tunicates are important source of novel compounds for drug discovery and development. Majority of these compounds are nitrogen containing and belong to non-ribosomal peptide (NRPs) or mixed polyketide–NRP natural products. Several of these peptides are currently under trial for developing new drugs against various disease areas, including inflammatory, cancer, neurodegenerative disorders, and infectious disease. This review features pharmacologically active NRPs from marine sponge and tunicates based on their biological activities.
Introduction
Nature provides a wide and structurally diverse array of active biomolecules that have proved vital for the development of novel pharmaceuticals. The marine world, covering more than 70% of the Earth's surface, is the home of tremendous biodiversity. Due to very diverse oceanic environments, marine organisms have developed the capacity to produce unique compounds (Steele, 1985; Mehbub et al., 2014). This rich and unprecedented chemo diversity of marine natural products provides an unlimited resource of novel biomolecules in the field of drug development. The importance of marine metabolites in current drug research is driven by the fact, that during 1981–2002, around half of US FDA-approved drugs consisted of either marine metabolites or their synthetic analogs (Vinothkumar and Parameswaran, 2013). Interestingly, the majority of these natural products involved in clinical or preclinical trials are produced by invertebrates, that is, sponges, tunicates, bryozoans, or molluscs. Sixty per cent of these natural products belong to non-ribosomal peptide (NRP) families, which are biosynthesized by poly-functional mega-synthases called NRP synthetases (NRPSs) (Finking and Marahiel, 2004; Mehbub et al., 2014). The excellent binding properties, low off-target toxicity, and high stability of NRPs make them a promising molecule for development of new therapeutics. Currently, only a handful of NRPs are used as drug (Table 1).
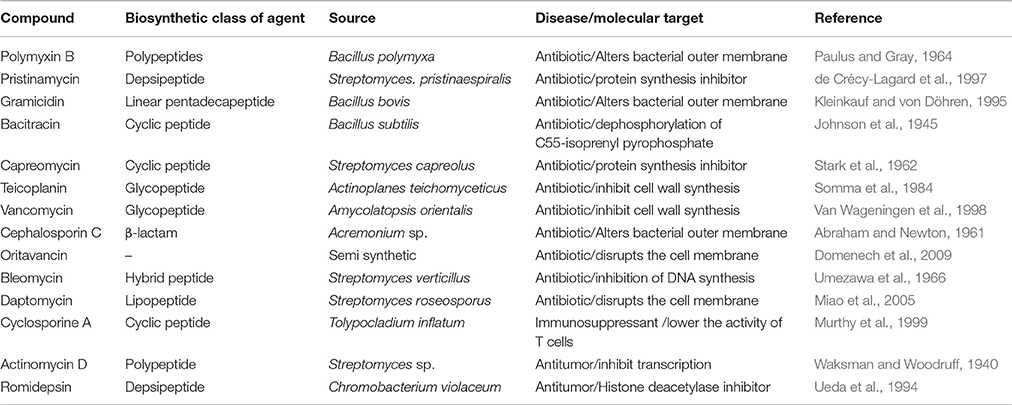
Table 1. NRPs-based drugs in market.
Marine sponges (Phylum porifera) represent the most primitive multicellular animals, with origins dating back to the Precambrian era (Hentschel et al., 2002). There are about 9000 reported species of sponges and (perhaps twice as many unreported species) available in the ocean (Brusca et al., 1990; Wörheide et al., 2005). These have been broadly categorized in 3 classes : Calcarea (5 orders and 24 families), Demospongiae (15 orders and 92 families), and Hexactinellida (6 orders and 20 families). Till date, more than 5300 different natural products have been isolated from marine sponges, and each year more than 200 additional new metabolites are being discovered (Laport et al., 2009; Mehbub et al., 2014). There are several sponge derived metabolites currently available in market and many in clinical studies (Table 2).
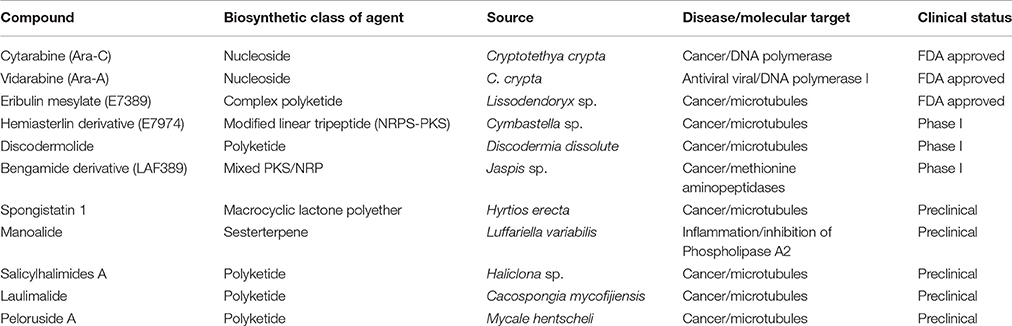
Table 2. Sponge secondary metabolites that are FDA-approved agents in clinical trial (Mayer et al., 2010; Newman and Cragg, 2016).
It is proposed that some of the bioactive compounds isolated from sponges are produced by functional enzyme clusters originated from the sponges and their associated microorganisms (Laport et al., 2009; Thomas et al., 2010). It has been observed that bacterial phyla such as Proteobacteria, Nitrospira, Cyanobacteria, Bacteriodetes, Actinobacteria, Chloroflexi, Planctomycetes, Acidobacteria, Poribacteria, and Verrucomicrobia besides members of the domain Archaea are most sponge-associated bacterial community (Hentschel et al., 2002; Olson and McCarthy, 2005). However, fungi and microalgae also symbiotically inhabit sponges. It has been recognized that one host sponge can possess diverse symbionts. For example, unicellular heterotrophic bacteria, unicellular cyanobacteria, and filamentous heterotrophic bacteria all grow together in sponge Theonella swinhoei (Bewley et al., 1996). Likewise, a sponge belonging to Aplysina includes heterogeneous bacteria Bacillus sp., Micrococcus sp., Arthrobacter sp., Vibrio sp., Pseudoalteromonas sp., and so on (Hentschel et al., 2001). Sponge Rhopaloeides odorabile has β-Proteobacteria, γ-Proteobacteria, Cytophaga, Actinobacteria, and green sulfur bacteria (Webster et al., 2001). Besides this, species-specific symbiotic relationship has also been observed. For example, sponge T. swinhoei and δ-proteobacteria have shown a specific association with each other (Schmidt et al., 2000). A species of α-proteobacteria dominates in sponge R. odorabile over various habitats but is not detected from seawater, which strongly suggests that the symbiont is species specific (Lee Y. K. et al., 2001). On the other hand, one symbiont occurs commonly in various sponges from different regions indicating its wide host range (Wilkinson et al., 1981). For example, cyanobacteria Aphanocapsa sp., Phormidium sp., or Oscillatoria spongeliae are found in numerous sponges (Wilkinson, 1978). Symbiotic associations between sponges and marine microorganisms might be involved in nutrient acquisition, stabilization of sponge skeleton, processing of metabolic waste, and secondary metabolite synthesis. It is assumed that symbiotic marine microorganisms harbored by sponges are the original producers of some of these bioactive compounds (Newman and Hill, 2006). For example, antibiotic polybrominated biphenyl ether isolated from the sponge Dysidea herbacea (Demospongiae) are actually produced by endosymbiotic cyanobacterium O. spongeliae (Unson et al., 1994). A symbiotic bacterium Micrococcus sp. produces diketopiperazines previously ascribed to the host sponge Tedania ignis (Stierle et al., 1988). Another symbiotic bacterium Vibrio sp. produces brominated biphenyl ethers formerly attributed to the host sponge Dysidea sp. (Elyakov et al., 1991). Symbiotic bacterium Vibrio sp. produces an anti-Bacillus peptide andrimid that was found in the sponge Hyatella sp. extract (Oclarit et al., 1994). Antimicrobial activity is detected in Micrococcus luteus isolated from the sponge Xestospongia sp. (Bultel-Poncé et al., 1998). Antimicrobial compounds such as quinolones and phosphatidyl glyceride are isolated from a Pseudomonas sp. collected at the surface of the sponge Homophymia sp. (Bultel-Poncé et al., 1999). However, the mutual mechanism between sponge and its microbial associate, in metabolite production, is not well-understood. Thus, it is extremely relevant to highlight the therapeutic potential of various secondary metabolites synthesized by the microbial flora inhabiting sponges. This is because they open up the possibility of providing a continuous supply of the biologically active compounds by laboratory cultivation of the producer (Thomas et al., 2010).
Tunicates include a wide variety of invertebrates that are classified within the Phylum chordata based on presence of a larval notochord during early development. Tunicates contains about 2150 described species that are divided into 4 classes: Ascidiacea (Aplousoobranchia, Phlebobranchia, Stolidobranchia) Thaliacea (Pyrosomida, Doliolida, Salpida), Appendicularia (Larvacea), and Sorberacea (Ruppert and Fox, 2004). Amongst these, Ascidacea (commonly known as the ascidians) are highly studied due to their biologically active metabolites that serve as antineoplastic agents. Geranyl hydroquinone was first ascidian metabolite isolated from Aplidium sp. which displayed chemo protective activity against some forms of leukemia, rous sarcoma, and mammary carcinoma in test animals (Fenical, 1976) (Menna, 2009). Since then, ascidians are known as the source of numerous marine natural products. The biologically active metabolites originated from tunicates which are approved by FDA or in clinical trials along with their biological properties are given in Table 3.

Table 3. Tunicate secondary metabolites that are FDA-approved agents or in clinical trial (Mayer et al., 2010; Newman and Cragg, 2016).
To date, significant biological activities, such as antimicrobial, anticancer, neurotoxic, antiprotozoal and their associated cellular targets have been reported for several NRPs from the marine sponges and tunicates. These NRPs have unique structures as compared with those from other sources. It is this attribute that makes marine sponge- and tunicate-derived NRPs highly attractive as potential drug and molecular probes. In this review, we survey the discoveries of NRPs derived from marine sponges and tunicates, which have shown in vivo efficacy or potent in vitro activity against various human diseases. Our objective is to highlight NRPs that have the greatest potential to be clinically useful. The details of sponge- and tunicate-derived NRPs along with biological properties is given Table 4.
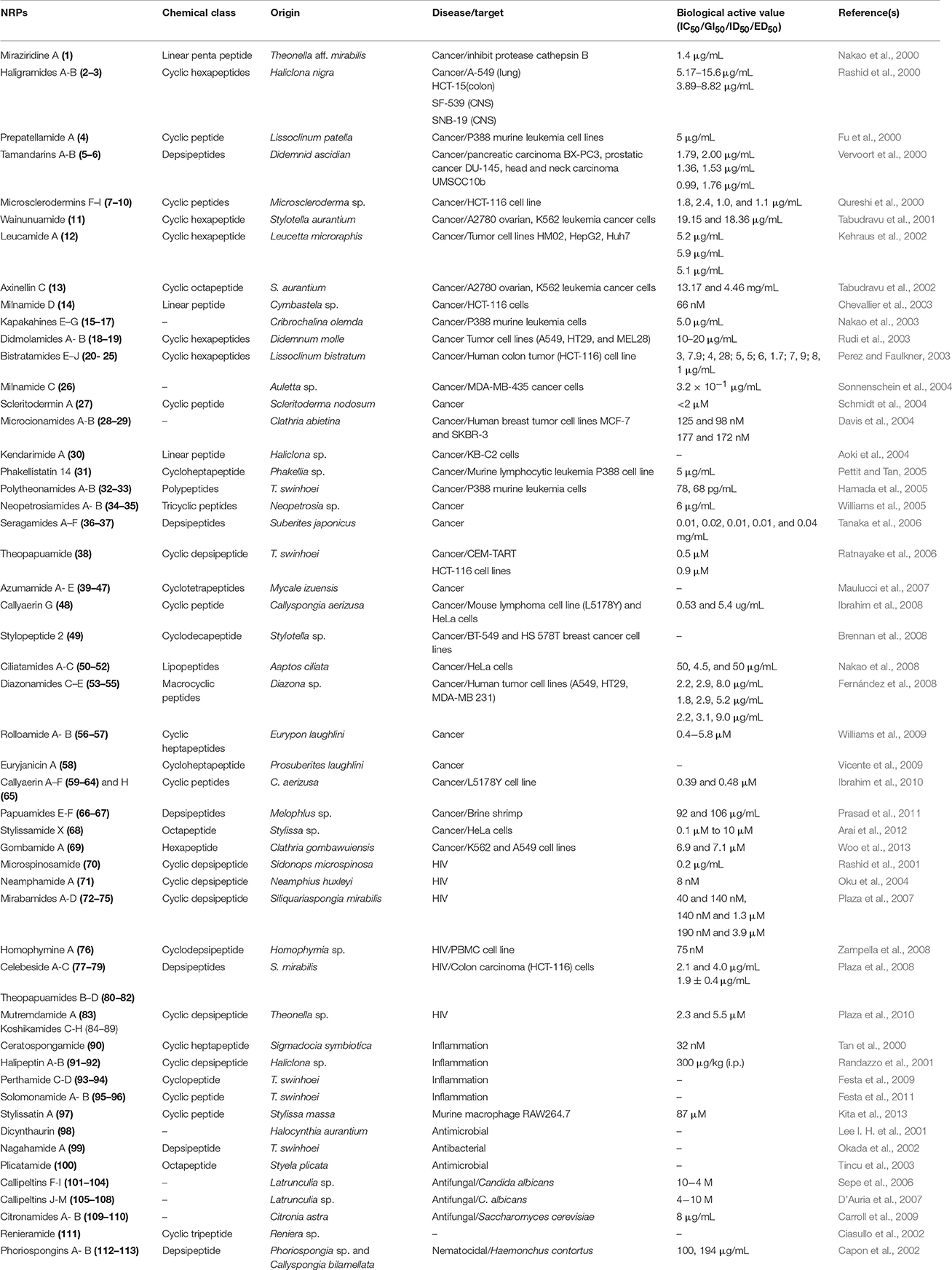
Table 4. Biological activities of NRPs isolated from marine sponges and tunicates.
Anticancer NRPs from Marine Sponges and Tunicates (Figures 1–6)
Sponge Theonella aff. mirabilis was the source of linear penta peptide Miraziridine A (1), which showed inhibitory activity on cathepsin B with an IC50 value of 1.4 μg/mL (Nakao et al., 2000). Two cyclic hexapeptides, Haligramides A (2) and B (3), were isolated from an aqueous extract of the sponge H. nigra. Both compounds exhibited cytotoxicity against various cell lines. Haligramide A exhibited cytotoxicity against A-549, HCT-15, SF-539, SNB-19 cell line with IC50 values of 5.17, 15.62, 9.00, and 9.08 μg/mL, respectively. Haligramide B, was found to be more active than Haligramide A against A-549, HCT-15, SF-539, SNB-19 cell line with IC50 values of 3.89, 8.82, 5.01, and 6.56 μg/mL, respectively (Rashid et al., 2000).
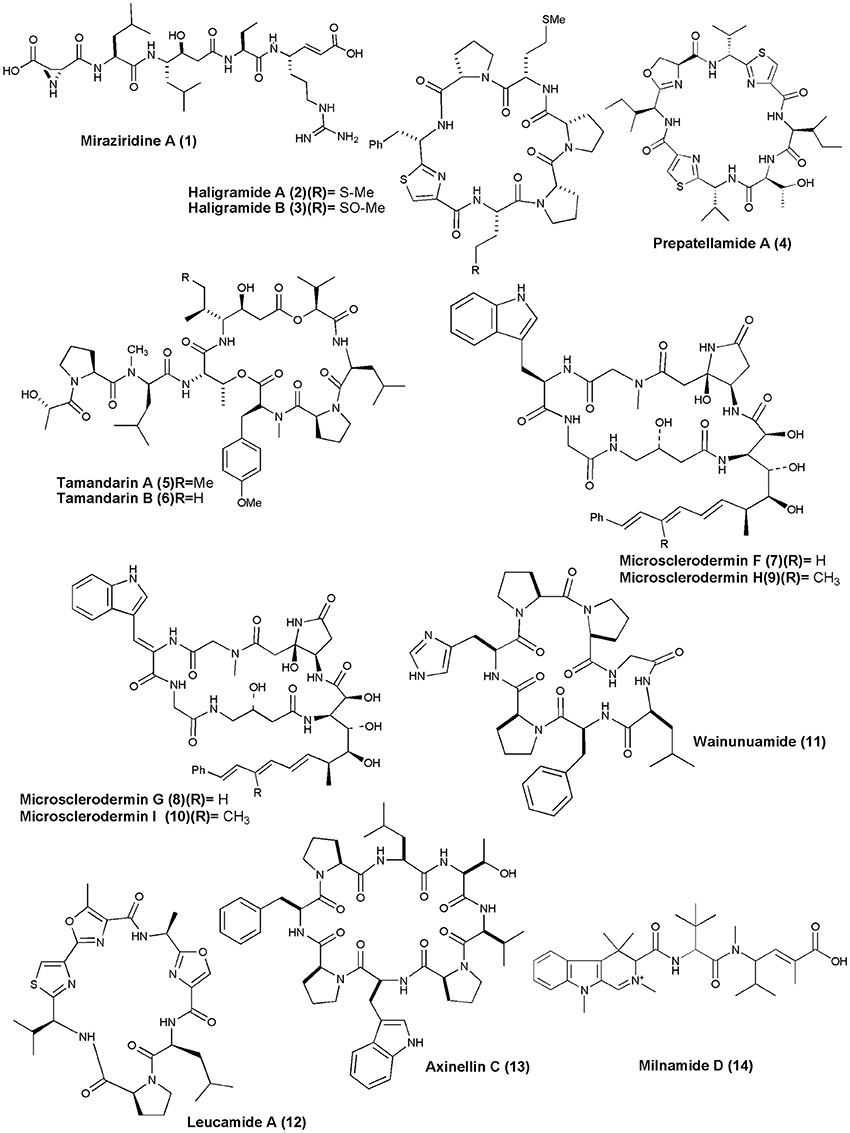
Figure 1. Structures of anticancer non-ribosomal peptides (1–14).
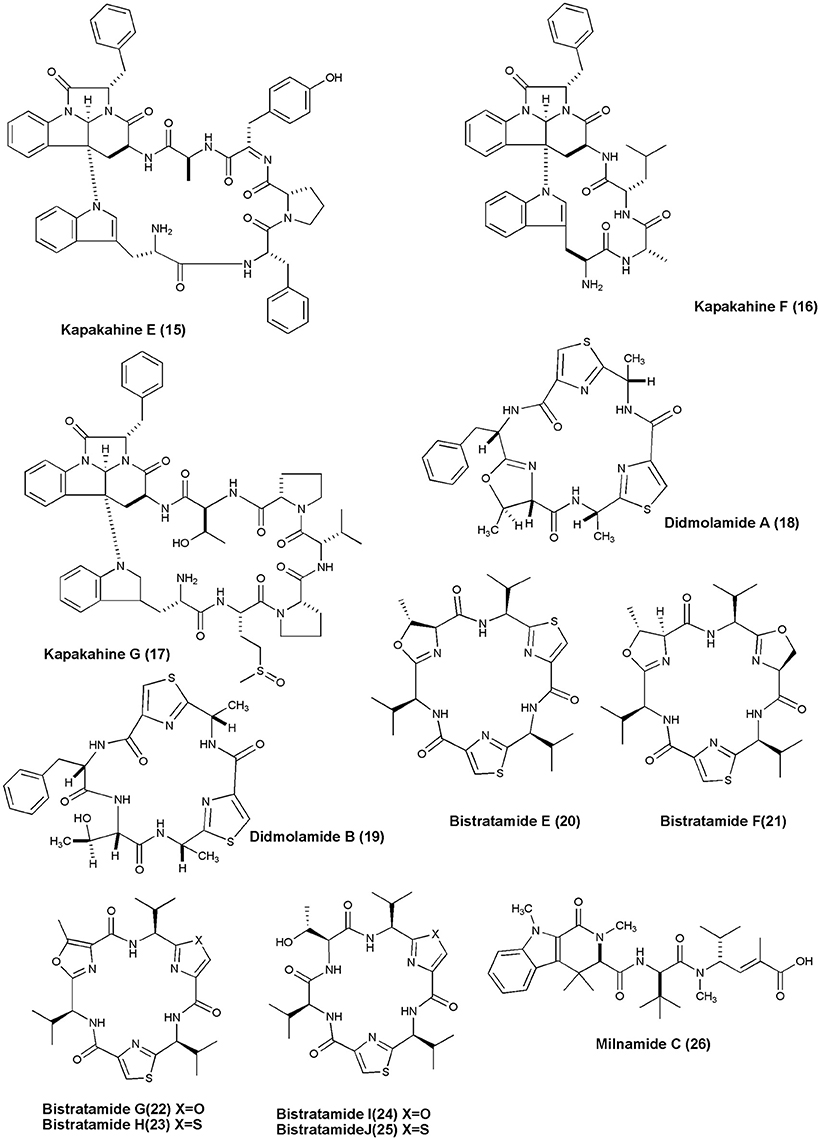
Figure 2. Structures of anticancer non-ribosomal peptides (15–26).
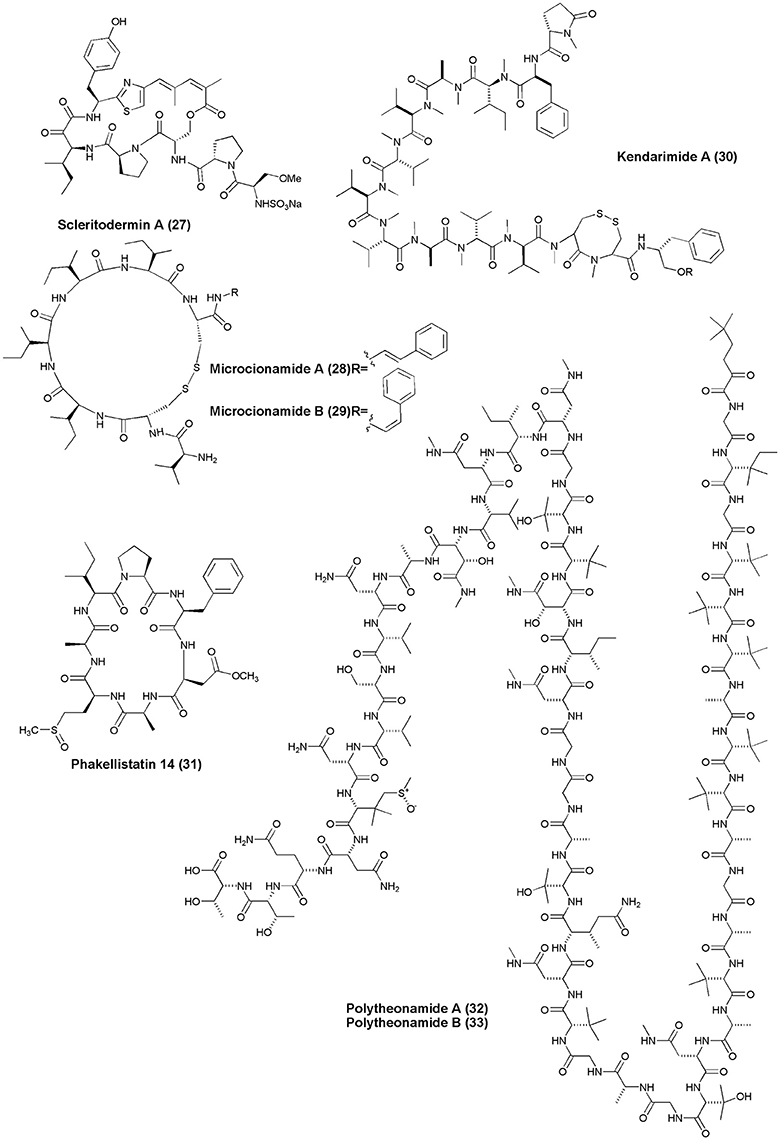
Figure 3. Structures of anticancer non-ribosomal peptides (27–33).
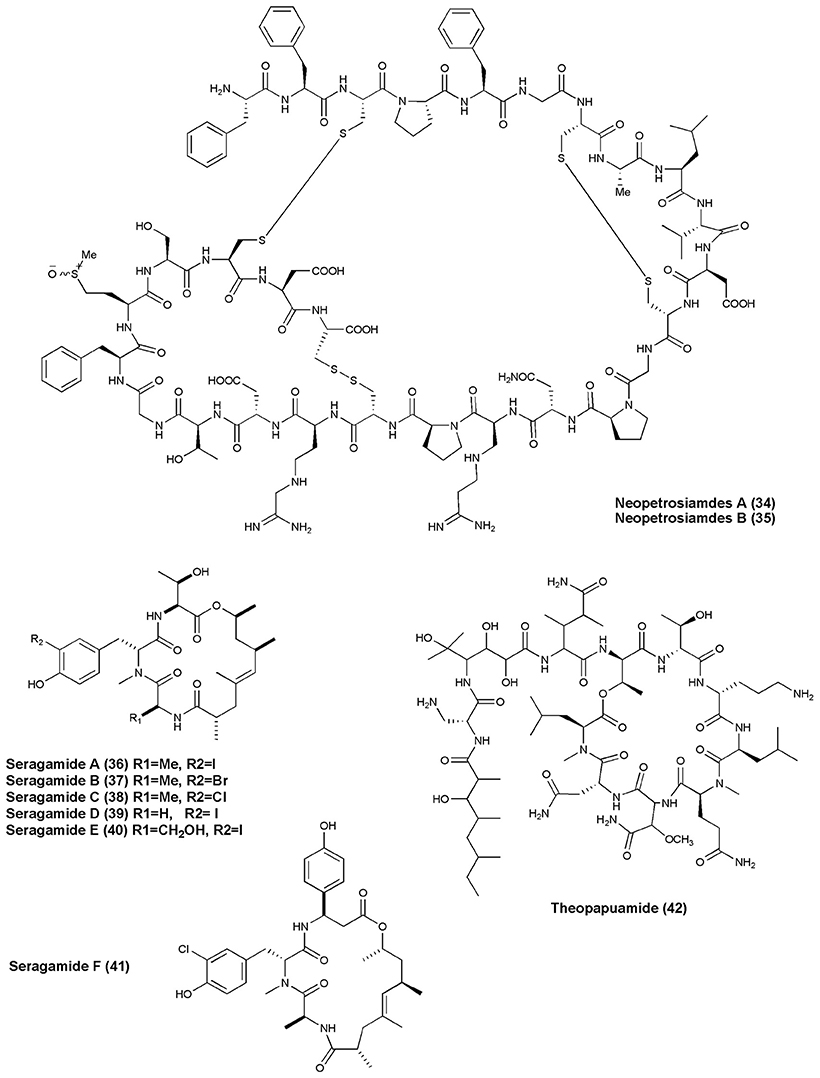
Figure 4. Structures of anticancer non-ribosomal peptides (34–42).
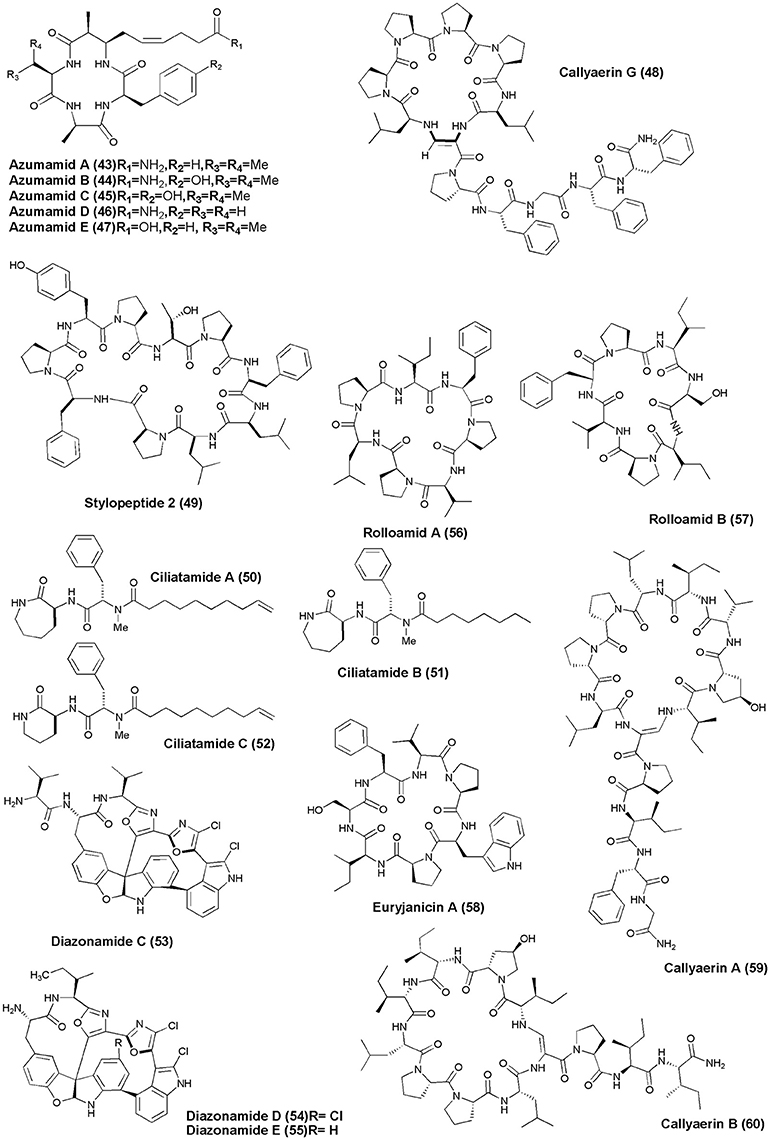
Figure 5. Structures of anticancer non-ribosomal peptides (43–60).
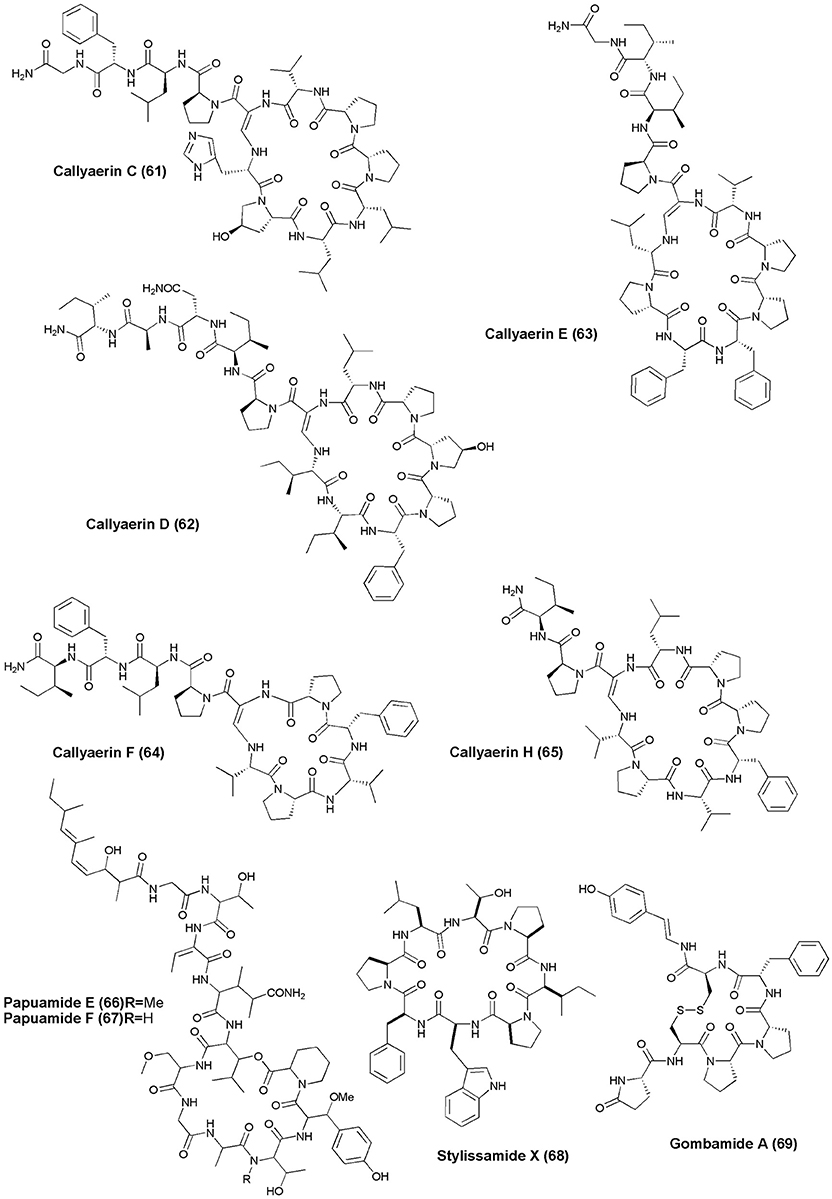
Figure 6. Structures of anticancer non-ribosomal peptides (61–69).
A cyclic peptide, Prepatellamide A (4) was isolated from the cytotoxic extracts of L. patella. The crude extract of this ascidian showed cytotoxicity against P388 murine leukemia cell lines with IC50 = ~5 μg/mL (Fu et al., 2000). Naturally occurring depsipeptides, Tamandarins A and B (5 and 6) were discovered from a Brazilian ascidian of the family Didemnidae and were cytotoxic against various human cancer cell lines including pancreatic carcinoma BX-PC3, IC50 = 1.79, 2.00 μg/mL, prostatic cancer DU-145, IC50 = 1.36, 1.53 μg /mL, head and neck carcinoma UMSCC10b, IC50 = 0.99, 1.7 6 μg/mL, respectively (Vervoort et al., 2000). Microscleroderma sp. from Palau gave 4 new cyclic peptides, Microsclerodermins F–I (7–10), all of which inhibited the growth of C. albicans with MIC value of 1.5, 3, 12, and 25 μg per disk, respectively, and also showed cytotoxicity against the HCT-116 cell line with IC50 value of 1.8, 2.4, 1.0, and 1.1 μg/mL, respectively (Qureshi et al., 2000).
A histidine-containing proline-rich cyclic heptapeptide, Wainunuamide (11), was isolated from the Fijian marine sponge S. aurantium. Compound (11) exhibited weak cytotoxic activity against A2780 ovarian tumor and K562 leukemia cancer cells with ID50 of 19.15 and 18.36 μg/mL, respectively (Tabudravu et al., 2001). The Australian marine sponge L. microraphis was the source of a bioactive cyclic heptapeptide, Leucamide A (12). Compound (12) inhibited the growth of the 3 tumor cell lines HM02 (gastric, GI50 = 5.2 μg/mL), HepG2 (liver, GI50 = 5.9 μg/mL), and Huh7 (liver, GI50 = 5.1 μg/mL) (Kehraus et al., 2002). The Fijian collection of marine sponge S. aurantium gave a proline-rich cyclic octapeptide, Axinellin C (13) (cyclo [Thr1-Val2-Pro3-Trp4-Pro5-Phe6-Pro7-Leu8]). Axinellin C displayed weak cytotoxicity against A2780 ovarian tumor and K562 leukemia cancer cells with ID50s of 13.17 and 4.46 mg/mL, respectively (Tabudravu et al., 2002). The crude extract of a marine sponge Cymbastela sp. (Papua New Guinea) gave a cytotoxic peptide Milnamide D (14). Milnamide D was found to exhibit cytotoxicity against HCT-116 at IC50 value of 66 nM and inhibition of tubulin polymerization at IC50 of 16 μM (Chevallier et al., 2003).
Investigation of marine sponge C. olemda yielded 3 new Kapakahines E−G (15–17). Only kapakahines E was found to display moderate cytotoxicity against P388 murine leukemia cells at IC50 of 5.0 μg/mL (Nakao et al., 2003). Two novel cyclic hexapeptides, Didmolamides A-B (18 and 19) were isolated from ascidian D. molle (Madagascar). Both peptides showed mild cytotoxicity against tumor cell lines (Lung—A549, Colon—HT29, and Skin—MEL28) with IC50 values of 10–20 μg/mL (Rudi et al., 2003). The Philippines ascidian L. bistratum was the source of 6 cyclic hexapeptides, Bistratamides E-J (20–25) which showed weak to moderate activity against the human colon tumor (HCT-116) cell line (IC50's: 3, 7.9; 4, 28; 5, 5; 6, 1.7; 7, 9; 8, 1 μg/mL, respectively) (Perez and Faulkner, 2003). Milnamide C (26) was isolated from Auletta sp., which showed significant activity against MDA-MB-435 breast cancer cells with IC50 values of 3.2 × 10−1 μg/mL (Sonnenschein et al., 2004).
Scleritodermin A (27), a new cyclic peptide, was isolated from the Lithistid Sponge S. nodosum. Scleritodermin A, inhibited tubulin polymerization and demonstrated significant in vitro cytotoxicity against a panel of human tumor cell lines (IC50 < 2 μM), including colon (HCT116), ovarian (A2780), and breast carcinoma cell lines SKBR3 (Schmidt et al., 2004). Microcionamides A (28) and B (29) were isolated from the Philippine marine sponge Clathria (Thalysias) abietina. Both compounds displayed significant cytotoxicity toward human breast tumor cell lines MCF-7 and SKBR-3 with IC50 of 125 and 98 nM for compound (28) and 177 and 172 nM for compound (29), respectively (Davis et al., 2004). Methanol extract of an Indonesian marine sponge Haliclona sp. gave a linear peptide Kendarimide A (30) which reversed P-glycoprotein-mediated multi-drug resistance in mammalian cells (Aoki et al., 2004).
Cycloheptapeptide, Phakellistatin 14 (31), was isolated from Phakellia sp., (Chuuk, Federated States of Micronesia). Compound (31) exhibited cytotoxicity against the murine lymphocytic leukemia P388 cell line at ED50 of 5 μg/mL (Pettit and Tan, 2005). The marine sponge T. swinhoei was found to produce highly cytotoxic polypeptides Polytheonamides A and B (32–33) with 48 amino acid residues. Both compounds were found to be cytotoxic against P388 murine leukemia cells with IC50 values of 78, 68 pg/mL, respectively (Hamada et al., 2005). Two diastereomeric tricyclic peptides Neopetrosiamdes A (34) and B (35) have been isolated from the marine sponge Neopetrosia sp. collected in Papua New Guinea. These peptides inhibited amoeboid invasion of human tumor cells at 6 μg/mL (Williams et al., 2005). Six new depsipeptides, Seragamides A–F (36–41) were isolated from sponge S. japonicas (Okinawan). Except seragamide F, all seragamides have showed multinuclei formation in NBT-T2 cells at 0.01, 0.02, 0.01, 0.01, and 0.04 mg/mL, respectively. Compound (36) also promotes the polymerization of G-actin and stabilizes F-actin filaments (Tanaka et al., 2006). Theonella swinhoei from Papua New Guinea gave a cyclic depsipeptide, Theopapuamide (42). This peptide contains several unusual amino acid residues such as β-methoxyasparagine, 4-amino-5-methyl-2, 3, 5-trihydroxy-hexanoic acid, and also contains an amide linked fatty acid moiety, 3-hydroxy-2, 4, 6-trimethyl-octanoic acid (Htoa) with cytotoxicity against CEM-TART (EC50 = 0.5 μM) and HCT-116 (EC50 = 0.9 μM) cell lines (Ratnayake et al., 2006).
Azumamide A-E (43–47) carboxylic acid containing histone deacetylase (HDAC) inhibitor cyclotetrapeptides were recovered from the sponge M. izuensis. Only compound (47) displayed human histone deacetylase inhibitory activity (Maulucci et al., 2007). An Indonesian sponge C. aerizusa gave a new cyclic peptide named Callyaerin G (48) with cytotoxicity against mouse lymphoma cell line (L5178Y) and HeLa cells with ED50(s) of 0.53 and 5.4 ug/mL, respectively (Ibrahim et al., 2008). The Papua New Guinea marine sponge Stylotella sp. was found to produce a new proline-rich cyclodecapeptide, Stylopeptide 2 (49) which inhibited the growth of BT-549 and HS 578T 2 breast cancer cell lines by 77 and 56%, respectively (Brennan et al., 2008). Bioactive lipopeptides Ciliatamides A-C (50–52) were isolated from the deep-sea sponge A. ciliate. Ciliatamides A-B have showed anti-leishmanial activity at 10 μg/mL with 50 and 45.5% growth inhibition, respectively. Ciliatamides A-C also inhibited growth of HeLa cells with IC50 values of 50, 4.5, and 50 μg/mL, respectively (Nakao et al., 2008).
The marine ascidian Diazona sp. (Indonesia) gave 3 new macrocyclic peptides, Diazonamides C–E (53–55). All the isolated peptides displayed moderate cytotoxicity against a panel of 3 human tumor cell lines (IC50's: A549 = 2.2, 2.9, 8.0 μg/mL; HT29 = 1.8, 2.9, 5.2 μg/mL; MDA-MB-231 = 2.2, 3.1, 9.0 μg/mL) (Fernández et al., 2008). Dominican marine sponge E. laughlini gave 2 cyclic heptapeptides, Rolloamides A (56) and B (57). Rolloamide A displayed significant growth suppression against several cancer cells (prostate, breast, ovarian, glioma, and renal) with IC50's of 0.4–5.8 μM (Williams et al., 2009). Proline-containing cycloheptapeptide, Euryjanicin A (58) was extracted from the marine sponge P. laughlini (Vicente et al., 2009). Bioassay guided extraction of the sponge C. aerizusa (Ambon, Indonesia) revealed 7 new cytotoxic cyclic peptides Callyaerins A–F (59–64) and H (65). All peptides have showed cytotoxicity, however, callyaerins E and H exhibited strong activity against the L5178Y lymphoma cell line with ED50 values of 0.39 and 0.48 μM, respectively (Ibrahim et al., 2010).
An undescribed sponge of the genus Melophlus sp. (Karumolum, Russell Is., Solomon Is.) yielded 2 depsipeptides, Papuamides E (66) and F (67), which were cytotoxic against brine shrimp with LD50 values between 92 and 106 μg/mL (Prasad et al., 2011). A proline-rich octapeptide Stylissamide X (68) isolated from an Indonesian marine sponge of Stylissa sp. inhibited HeLa cells in the concentration range 0.1–10 μM through wound-healing assay (Arai et al., 2012). The marine sponge C. gombawuiensis collected from Korean waters gave a disulphide-linked hexapeptide, Gombamide A (69). Gombamide A showed weak cytotoxic activity against the K562 and A549 cell lines with LC50 values of 6.9 and 7.1 μM, respectively, as well as moderate inhibitory activity against Na+/K+-ATPase with an LC50 value of 17.8 μM (Woo et al., 2013).
Anti-HIV Agents (Figures 7, 8)
The marine sponge S. microspinosa was the source of a cyclic depsipeptide Microspinosamide (70), inhibited HIV-1 infection with an EC50 value of approximately 0.2 μg/mL (Rashid et al., 2001). A Papua New Guinea collection of the marine sponge N. huxleyi has been shown to produce a new HIV-inhibitory cyclic depsipeptide, Neamphamide A (71). Neamphamide A displayed potent cytoprotective activity against HIV-1 infection with EC50 value ~28 nM (Oku et al., 2004). Four cyclic depsipeptides, Mirabamides A–D (72–75), were isolated from the marine sponge S. mirabilis. Mirabamides A, C and D inhibited HIV-1 fusion (Mirabamides A IC50 values between 40 and 140 nM: Mirabamides C IC50 values between 140 nM and 1.3 μM: and Mirabamides D IC50 values between 190 nM and 3.9 μM). Mirabamides A–C also inhibited the growth of B. subtilis and C. albicans at 1–5 μg/disk (Plaza et al., 2007).
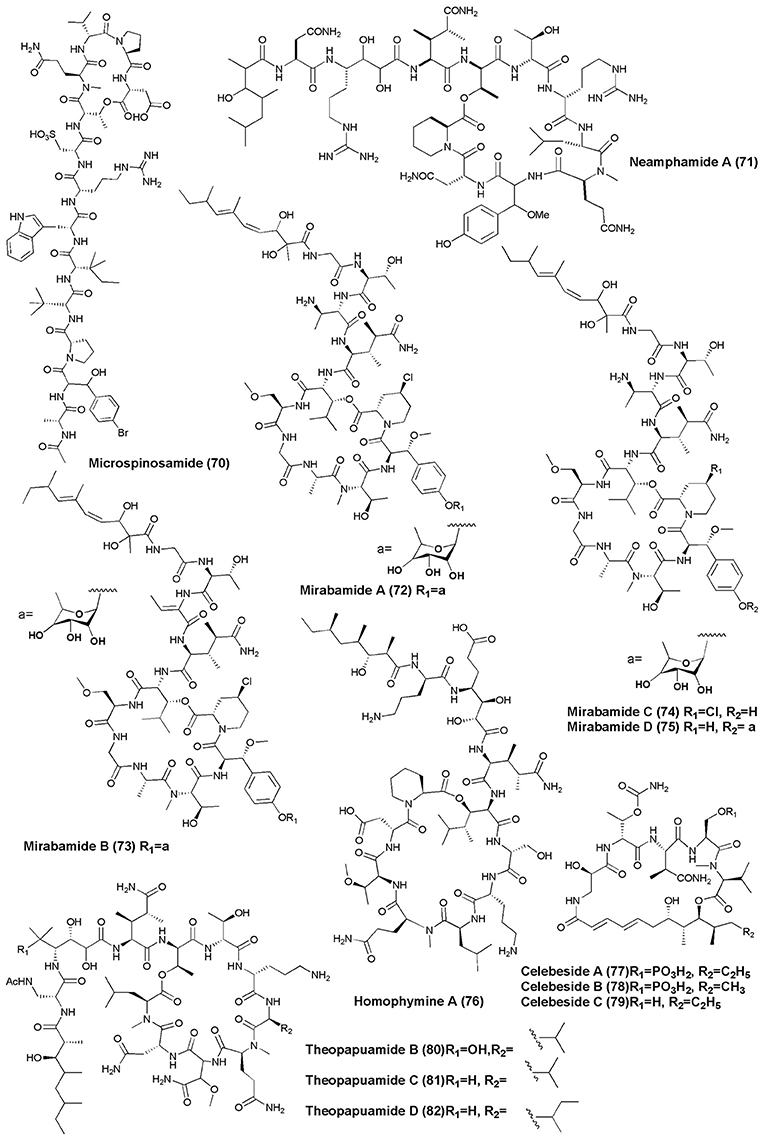
Figure 7. Structures of non-ribosomal peptides with anti-HIV activity (70–82).
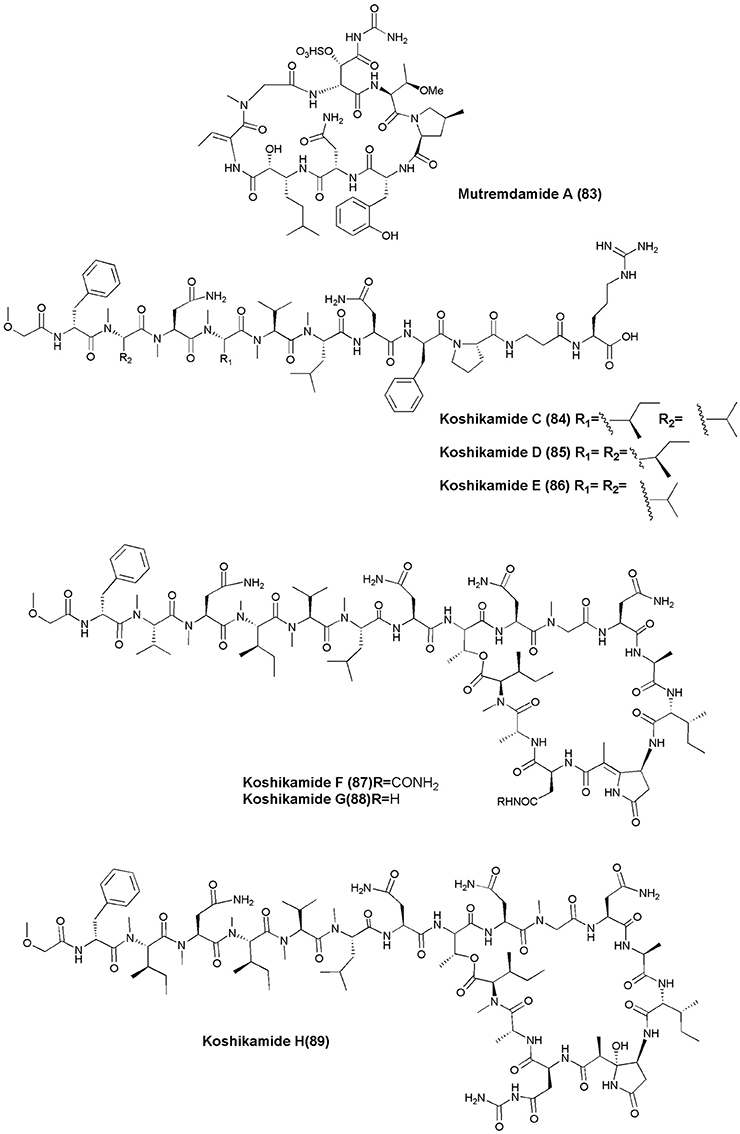
Figure 8. Structures of non-ribosomal peptides with anti-HIV activity (83–89).
The marine sponge Homophymia sp. was the source of an anti-HIV cyclodepsipeptide, Homophymine A (76). This peptide inhibited the infection of HIV-1 in PBMC cell line with an IC50 of 75 nM (Zampella et al., 2008). Six depsipeptides Celebesides A-C (77–79) and Theopapuamides B-D (80–82) were isolated from an Indonesian sponge S. mirabilis. Compound (77) neutralized HIV-1 with an IC50 value of 1.9 ± 0.4 μg/mL, while the non-phosphorylated analog Celebeside C was inactive at concentrations as high as 50 μg/mL. Theopapuamides A-C displayed cytotoxicity against human colon carcinoma (HCT-116) cells with IC50 values between 2.1 and 4.0 μg/mL, and antifungal activity against wild type and amphotericin B-resistant strains of C. albicans at 1–5 μg/disk (Plaza et al., 2008). The deep-water specimens of T. swinhoei and Theonella cupola (Mutremdiu Reef, Palau) gave sulphated cyclic depsipeptide, Mutremdamide A (83) and 6 N-methylated peptides Koshikamides C–H (84–89). Cyclic koshikamides F and H inhibited HIV-1 entry at IC50 values of 2.3 and 5.5 μM, respectively, while their linear counterparts were inactive (Plaza et al., 2010).
Anti-Inflammatory NRPs (Figure 9)
Marine sponge S. symbiotica collected from Biaro Island, Indonesia, alongwith its symbiont marine red alga (Rhodophyta) Ceratodictyon spongiosum gave thiazole-containing cyclic heptapeptide, Ceratospongamide (90). Compound (90) consists of two l-phenylalanine residues, one (l-isoleucine)-l-methyloxazoline residue, one l-proline residue, and one (l-proline) thiazole residue. The trans-isomer of ceratospongamide exhibits potent inhibition of sPLA2 expression in a cell-based model for anti-inflammation at ED50 32 nM (Tan et al., 2000). Two cyclic depsipeptides, Halipeptins A and B (91,92) were obtained from marine sponge Haliclona sp. Only halipeptins A displayed in vivo potent anti-inflammatory activity (mice at the dose of 300 μg/kg [i.p.]) (Randazzo et al., 2001). A Solomon Lithistid sponge T. swinhoei was the source of 2 new cyclopeptides Perthamides C and D with potent anti-inflammatory (93,94) (Festa et al., 2009). Cyclic peptides, Solomonamides A and B (95,96), were separated out from the marine sponge T. swinhoei; however, only compound (86) showed anti-inflammatory activity (Festa et al., 2011). The marine sponge S. massa produced a cyclic peptide Stylissatin A (97) that inhibited nitric oxide production in LPS-stimulated murine macrophage RAW264.7 cells with an IC50 value of 87 μM (Kita et al., 2013).
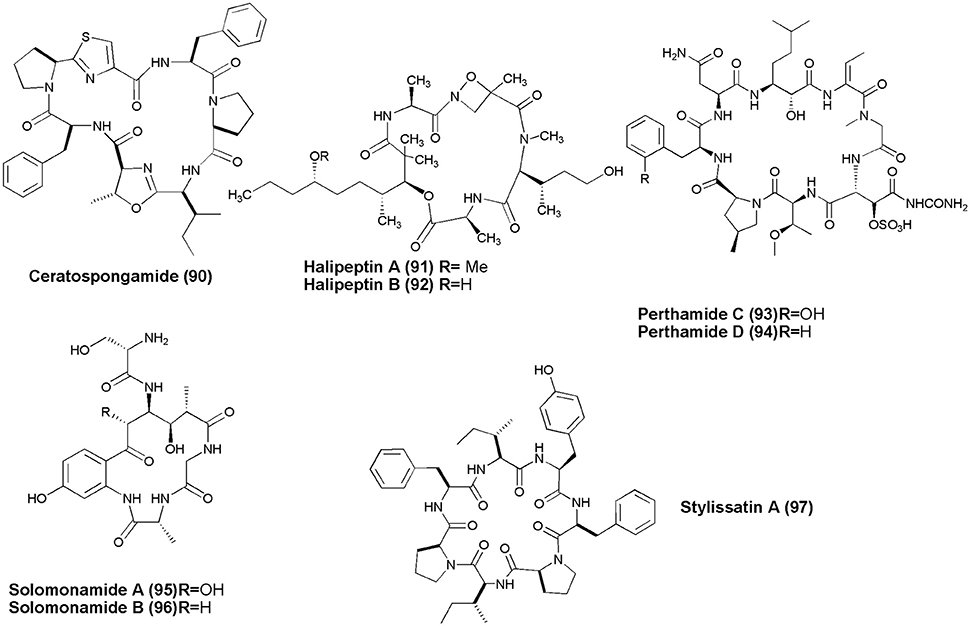
Figure 9. Structures of non-ribosomal peptides with anti-inflammatory activity (90–97).
Antimicrobial Agents (Figure 10)
The solitary tunicate, H. aurantium, was the source of a novel antimicrobial peptide Dicynthaurin (98) (Lee I. H. et al., 2001). An antibacterial depsipeptide, Nagahamide A (99), was discovered from the marine sponge T. swinhoei (Okada et al., 2002). An antimicrobial octapeptide Plicatamide (100) was isolated from S. plicata (Tincu et al., 2003). The marine sponge Latrunculia sp., (Vanuatu Islands) was the source of four new antifungal peptides, Callipeltins F–I (101–104). Callipeltins F–I inhibit the growth of C. albicans (ATCC24433) with a MIC value of 10−4 M (Sepe et al., 2006). Four new peptides, Callipeltins J–M (105–108), were isolated from the marine sponge Latrunculia sp. However, only Callipeltins J and K inhibited the growth of C. albicans with MIC values of ca. 4−10 M (D'Auria et al., 2007). Two new linear tetrapeptides, Citronamides A (109) and B (110), were isolated from the Australian sponge C. astra. Only citronamides A showed moderate antifungal activity against Saccharomyces cerevisiae at MIC value of 8 μg/mL (Carroll et al., 2009).
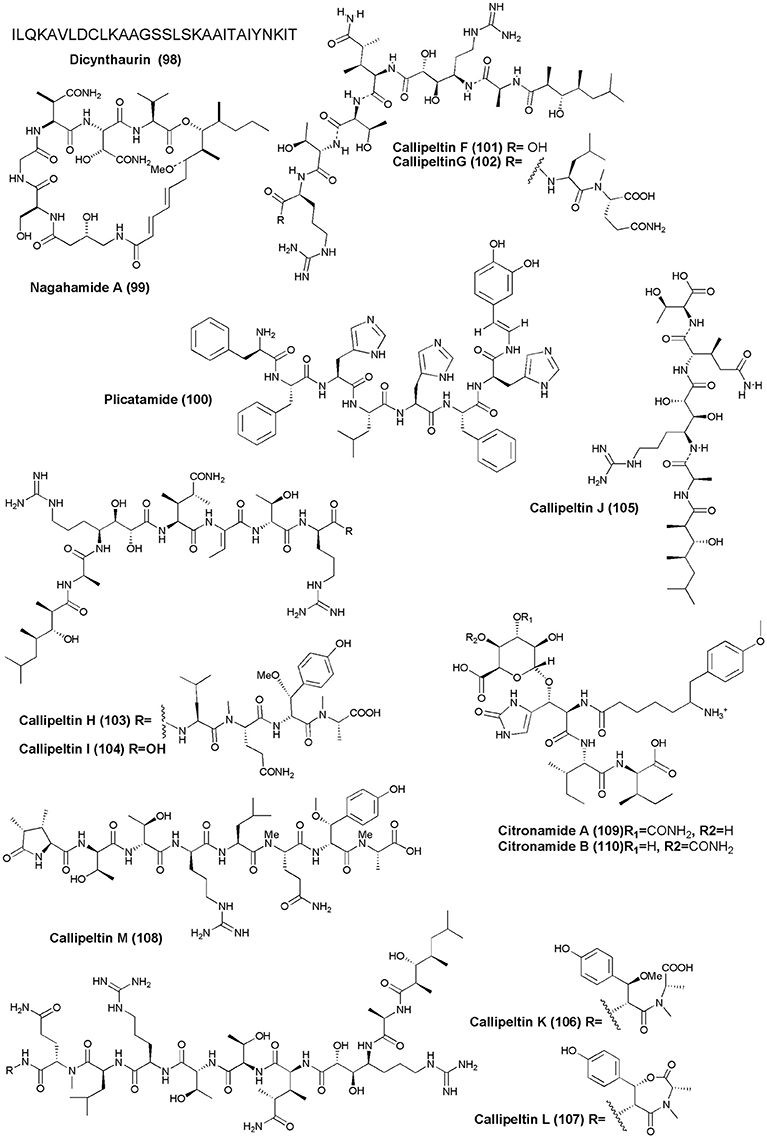
Figure 10. Structures of antimicrobial non-ribosomal peptides (98–110).
Miscellaneous (Figure 11)
A cyclic tripeptide Renieramide (111) was isolated from Vanuatu collection of sponge Reniera sp. that showed immunomodulating activity in preliminary tests (Ciasullo et al., 2002). Two nematocidal depsipeptides, Phoriospongin A and B (112 and 113), were isolated from Australian marine sponges Phoriospongia sp. and C. bilamellata. Both compounds have displayed significant nematocidal activity against H. contortus (LD99 = 100, 194 μg/mL, respectively) (Capon et al., 2002).
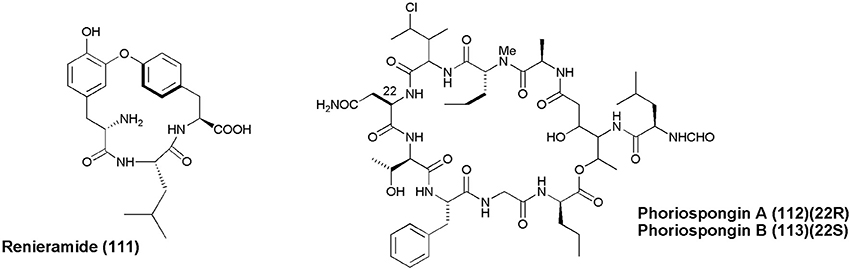
Figure 11. Structures of non-ribosomal peptides with (111–113).
Biological Aspects, Challenges, and Future Perspectives
Like their structural diversity, metabolites produced from marine sponges and tunicates bind to a variety of cellular targets to elicit their effects. Numerous articles published in recent years highlighting the significance of these metabolites in disease control, the details of their biological significance from molecular recognition perspective have been rather scarce. Although some promising leads have been obtained, the discovery of their cellular targets, molecular interactions, and adverse effects are lacking. In cases where the therapeutic potential has been reported, details of a proper screening approach to identify nucleic acid or protein targets are missing. However, some established metabolites from these sources (see Tables 1, 2) and their derivatives have been examined extensively and their molecular targets are varied. One of the earliest examples in this class is FDA-approved drug Ara-C (cytarabine), which is known to elicit anticancer properties by inhibiting the functions of DNA polymerase (Furth and Cohen, 1968), which ultimately results in stalling DNA synthesis. Another FDA approved related compound Ara-A(vidarabine), which is known to have antiviral properties (active against herpes simplex and varicella zoster viruses), targets viral DNA polymerase (Chadwick et al., 1978) by functioning as mimic of natural nucleotides. Both Ara-C and Ara-A resemble natural cytidine and adenine nucleosides where the structural differences are in the sugar components of the two (arabinose vs. deoxyribose). The natural nucleoside mimics Ara-A and Ara-C are easily phosphorylated as their triphosphate derivatives by kinases and act as terminators of DNA synthesis. Ara-A is also known to impede 3′-end processing of pre-mRNAs by inhibiting cleavage and polyadenylation (Ghoshal and Jacob, 1991; Rose and Jacob, 1978).
Several other molecules that are either FDA approved or in early stages of clinical trials have been identified as anticancer agents with microtubules as their primary molecular targets. The predominance of natural metabolites being microtubule binding agents has been hypothesized as evolutionary response to predation by plants and animals (Dumontet and Jordan, 2010). Some of these molecules, such as discodermolide, are among the first non-taxane stabilizers of microtubules (Mooberry et al., 2004). The microtubule stabilizers act by enhancing microtubule polymerization at high concentrations. Discodermolide has been known to bind to tubulin dimers in a stoichiometric ratio. Competitive binding experiments have shown that it blocks taxol binding and is a much stronger binder of microtubules than taxol (Kowalski et al., 1997). The microtubule binding of Tau proteins is interfered by discodermolide (Kar et al., 2003). Similarly, laulimalide showed properties very similar to paclitaxel where it helped in enhancing tubulin assembly (Gapud et al., 2004). However, laulimalide modulation of microtubule assembly in C. elegans is dose dependent where it stabilization effects were observed only at concentrations higher than 100 nM (Bajaj and Srayko, 2013).
The antiviral effect of homophymine A has been established by measuring the reverse transcriptase activity in HIV-infected primary peripheral blood mononuclear cells (Zampella et al., 2008). The reverse transcriptase activity is exhibited by 2 classes of molecules: one that directly competes with natural nucleotide triphosphates and the other that either directly blocks the catalytic reactions or by allosteric binding that leads to structural changes in the viral enzyme. Since homophymine A lacks structural features to act as mimics of natural nucleotide triphosphates, it is likely to impede the catalytic activity of the enzyme by direct binding.
A tunicate-derived metabolite trabectedin (ET-743) uses DNA binding to exert its anticancer properties. Trabectin binds to the GC rich regions in the B-DNA where it uses its carbolinamine moiety to form adduct with the exocylic amine (N-2) of guanine (Pommier et al., 1996) and covers 3 base pairs during this process (Marco et al., 2006). Unlike B-DNA minor groove binders, such as Hoechst 33258, which binds snugly along the minor groove curvature with high-affinity (Haq et al., 1997), trabectedin only uses part of its structure to make necessary contacts for the antitumor action (D'Incalci and Galmarini, 2010).
Despite these advances in determining the mode of their binding, a large number of recently discovered metabolites are still not explored to assess it functional capabilities. In the past, well-known anti-retroviral drug zidovudine, which was initially thought to be functionally inert, turned out as excellent therapeutic agent. Such discoveries are possible only when a rational screening design is aimed to asses it full potential as a drug. For example, compounds that have structural regions favorable for protein binding should be screened against all potential protein targets. Similarly, compounds that show preference toward nucleic acid binding should be screened using assays such as competition dialysis that establish a preferential nucleic acid target. Such approaches not only determine the best target for a particular compound but also shed light to its secondary targets, which may be helpful in dealing with toxicity issues. Current target design of marine and tunicate metabolites clearly need to take these approaches.
Some of the metabolites that have weaker binding to a target or have poor bioavailability can be improved by nano-encapsulation techniques. Additionally, DNA binding metabolites can be chemically modified to enhance their affinity using multi-recognition of the target (Willis and Arya, 2010), which has led to remarkable enhancement in the affinity of double,(Arya et al., 2003), triple (Arya and Willis, 2003), and four-stranded DNA helical structures (Ranjan et al., 2013).
Conclusion
Extreme environment of the ocean plays a vital role in exploring and studying marine bio-resources and their bio-actives. The large biodiversity of the sea serves as a huge resource for developing potential drugs with promising pharmacological activities. The significance of marine-derived secondary metabolites has recently been highlighted by introduction of Prialt and Yondelis to the market. In the past three decades, numerous NRPs with unique chemical structures and varied biological activities have been discovered from marine sponges and tunicates as described in this. Some of these exhibit strong potential to be developed as a new drug. However, none of the NRPs highlighted in this review have been successfully marketed as therapeutics. To translate bioactivity of these important metabolites into therapeutically significant outcomes, it is crucial to further unravel their modes of action and measure their toxicity. Since the majority of these studies have been focused on in vitro bioassays and elucidation of the chemical structures only, a complete examination of their biological target selectivity is required. Nevertheless, large-scale production of these NRPs for clinical use is a real challenge. Therefore, environmentally sound and economically feasible alternatives are required. To counter these challenges, many strategies have been established.
Chemical synthesis of NRPs is among the first strategies to be used. However, the structural complexity limits its chemical synthesis and has resulted in only a few successful achievements (e.g., analgesic drug ziconotide; Olivera, 2000). A second strategy uses screening the pharmacological significance of NRPs and subsequently attempting to define the critical pharmacophore that can result in practical drugs based on a marine prototype via chemical synthesis, degradation, modification, or a combination of these. Aquaculture of the source organisms has also been used to secure a sustainable supply of active compounds. However, in most cases, the biomass currently generated is still far from the requirement from an industrial perspective (Mendola, 2000). Identification and large-scale culturing of true producers that are known to thrive within the tissues of marine invertebrates (sponge or tunicate) is an intriguing strategy. However, to date only 5% or less of the symbiotic microbes present in marine specimens can be cultivated under standard conditions. Consequently, molecular approaches such as transfer of biosynthetic gene clusters to a vector suitable for large-scale fermentation could be used to avoid obstacles in culturing symbiotic bacteria. Enzyme technology and solid-phase peptide synthesis offer particularly promising alternatives to generate variety of unique peptides using native peptide as a template. Besides, combinations of chemical synthesis and biosynthetic technologies have potential to accelerate the discovery of novel drugs derived from sponge and their microbial association in future.
Author Contributions
SD reviewed the collected information critically. SA collected the relevant information from various sources including databases like Scifider. AA gave the concept of the work.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abraham, E., and Newton, G. (1961). The structure of cephalosporin C. Biochem. J. 79:377. doi: 10.1042/bj0790377
Aoki, S., Cao, L., Matsui, K., Rachmat, R., Akiyama, S.-I., and Kobayashi, M. (2004). Kendarimide A, a novel peptide reversing P-glycoprotein-mediated multidrug resistance in tumor cells, from a marine sponge of Haliclona sp. Tetrahedron 60, 7053–7059. doi: 10.1016/j.tet.2003.07.020
Arai, M., Yamano, Y., Fujita, M., Setiawan, A., and Kobayashi, M. (2012). Stylissamide X, a new proline-rich cyclic octapeptide as an inhibitor of cell migration, from an Indonesian marine sponge of Stylissa sp. Bioorg. Med. Chem. Lett. 22, 1818–1821. doi: 10.1016/j.bmcl.2011.10.023
Arya, D. P., and Willis, B. (2003). Reaching into the major groove of B-DNA: synthesis and nucleic acid binding of a Neomycin-Hoechst 33258 conjugate. J. Am. Chem. Soc. 125, 12398–12399. doi: 10.1021/ja036742k
Arya, D. P., Xue, L., and Tennant, P. (2003). Combining the best in triplex recognition: synthesis and nucleic acid binding of a BQQ-Neomycin conjugate. J. Am. Chem. Soc. 125, 8070–8071. doi: 10.1021/ja034241t
Bajaj, M., and Srayko, M. (2013). Laulimalide induces dose-dependent modulation of microtubule behaviour in the C. elegans embryo. PLoS ONE 8:e71889. doi: 10.1371/journal.pone.0071889
Bewley, C., Holland, N., and Faulkner, D. (1996). Two classes of metabolites from Theonella swinhoei are localized in distinct populations of bacterial symbionts. Experientia 52, 716–722. doi: 10.1007/BF01925581
Brennan, M. R., Costello, C. E., Maleknia, S. D., Pettit, G. R., and Erickson, K. L. (2008). Stylopeptide 2, a proline-rich cyclodecapeptide from the sponge Stylotella sp. J. Nat. Prod. 71, 453–456. doi: 10.1021/np0704856
Brusca, R. C., Brusca, G. J., and Haver, N. J. (1990). Invertebrates. Sunderland, MA: Sinauer Associates.
Bultel-Poncé, V., Berge, J.-P., Debitus, C., Nicolas, J.-L., and Guyot, M. (1999). Metabolites from the sponge-associated bacterium Pseudomonas species. Mar. Biotechnol. 1, 384–390. doi: 10.1007/PL00011792
Bultel-Poncé, V., Debitus, C., Berge, J.-P., Cerceau, C., and Guyot, M. (1998). Metabolites from the sponge-associated bacterium Micrococcus luteus. J. Mar. Biotechnol. 6, 233–236.
Capon, R. J., Ford, J., Lacey, E., Gill, J. H., Heiland, K., and Friedel, T. (2002). Phoriospongin A and B: two new nematocidal depsipeptides from the Australian marine sponges Phoriospongia sp. and Callyspongia bilamellata. J. Nat. Prod. 65, 358–363. doi: 10.1021/np010329d
Carroll, A. R., Duffy, S., and Avery, V. M. (2009). Citronamides A and B, tetrapeptides from the australian sponge Citronia astra. J. Nat. Prod. 72, 764–768. doi: 10.1021/np800832w
Chadwick, R. G., Bassendine, M. F., Crawford, E. M., Thomas, H. C., and Sherlock, S. (1978). Hbsag-positive chronic liver disease: inhibition of DNA polymerase activity by vidarabine. Br. Med. J. 2, 531–533. doi: 10.1136/bmj.2.6136.531
Chevallier, C., Richardson, A. D., Edler, M. C., Hamel, E., Harper, M. K., and Ireland, C. M. (2003). A new cytotoxic and tubulin-interactive milnamide derivative from a marine sponge Cymbastela sp. Org. Lett. 5, 3737–3739. doi: 10.1021/ol035476c
Ciasullo, L., Casapullo, A., Cutignano, A., Bifulco, G., Debitus, C., Hooper, J., et al. (2002). Renieramide, a cyclic tripeptide from the Vanuatu sponge Reniera n. sp. J. Nat. Prod. 65, 407–410. doi: 10.1021/np010383u
D'Auria, M. V., Sepe, V., D'Orsi, R., Bellotta, F., Debitus, C., and Zampella, A. (2007). Isolation and structural elucidation of callipeltins J–M: antifungal peptides from the marine sponge Latrunculia sp. Tetrahedron 63, 131–140. doi: 10.1016/j.tet.2006.10.032
Davis, R. A., Mangalindan, G. C., Bojo, Z. P., Antemano, R. R., Rodriguez, N. O., Concepcion, G. P., et al. (2004). Microcionamides A and B, bioactive peptides from the Philippine sponge Clathria (Thalysias) abietina. J. Org. Chem. 69, 4170–4176. doi: 10.1021/jo040129h
de Crécy-Lagard, V., Blanc, V., Gil, P., Naudin, L., Lorenzon, S., Famechon, A., et al. (1997). Pristinamycin I biosynthesis in Streptomyces pristinaespiralis: molecular characterization of the first two structural peptide synthetase genes. J. Bacteriol. 179, 705–713.
D'Incalci, M., and Galmarini, C. M. (2010). A review of trabectedin (ET-743): a unique mechanism of action. Mol. Cancer Therap. 9, 2157–2163. doi: 10.1158/1535-7163.MCT-10-0263
Domenech, O., Francius, G., Tulkens, P. M., Van Bambeke, F., Dufrêne, Y., and Mingeot-Leclercq, M.-P. (2009). Interactions of oritavancin, a new lipoglycopeptide derived from vancomycin, with phospholipid bilayers: effect on membrane permeability and nanoscale lipid membrane organization. Biochim. Biophys. Acta 1788, 1832–1840. doi: 10.1016/j.bbamem.2009.05.003
Dumontet, C., and Jordan, M. A. (2010). Microtubule-binding agents: a dynamic field of cancer therapeutics. Nat. Rev. Drug Discov. 9, 790–803. doi: 10.1038/nrd3253
Elyakov, G., Kuznetsova, T., Mikhailov, V., Maltsev, I., Voinov, V., and Fedoreyev, S. (1991). Brominated diphenyl ethers from a marine bacterium associated with the sponge Dysidea sp. Experientia 47, 632–633. doi: 10.1007/BF01949894
Fenical, W. (1976). Geranyl hydroquinone, a cancer-protective agent from the tunicate Aplidium species. Food Drugs Sea Proc. 4, 388–394.
Fernández, R., Martín, M. J., Rodríguez-Acebes, R., Reyes, F., Francesch, A., and Cuevas, C. (2008). Diazonamides C–E, new cytotoxic metabolites from the ascidian Diazona sp. Tetrahedron Lett. 49, 2283–2285. doi: 10.1016/j.tetlet.2008.02.012
Festa, C., De Marino, S., Sepe, V., D'Auria, M. V., Bifulco, G., Débitus, C., et al. (2011). Solomonamides A and B, new anti-inflammatory peptides from Theonella swinhoei. Org. Lett. 13, 1532–1535. doi: 10.1021/ol200221n
Festa, C., De Marino, S., Sepe, V., Monti, M. C., Luciano, P., D'auria, M. V., et al. (2009). Perthamides C and D, two new potent anti-inflammatory cyclopeptides from a Solomon Lithistid sponge Theonella swinhoei. Tetrahedron 65, 10424–10429. doi: 10.1016/j.tet.2009.10.026
Finking, R., and Marahiel, M. A. (2004). Biosynthesis of nonribosomal peptides1. Annu. Rev. Microbiol. 58, 453–488. doi: 10.1146/annurev.micro.58.030603.123615
Fu, X., Su, J., and Zeng, L. (2000). Prepatellamide A, a new cyclic peptide from the ascidian Lissoclinum patella. Sci. China Ser. B Chem. 43, 643–648. doi: 10.1007/BF02969512
Furth, J. J., and Cohen, S. S. (1968). Inhibition of mammalian DNA polymerase by the 5′-triphosphate of 1-β-d-arabinofuranosylcytosine and the 5′-triphosphate of 9-β-d-arabinofuranosyladenine. Cancer Res. 28, 2061–2067.
Gapud, E. J., Bai, R., Ghosh, A. K., and Hamel, E. (2004). Laulimalide and paclitaxel: a comparison of their effects on tubulin assembly and their synergistic action when present simultaneously. Mol. Pharmacol. 66, 113–121. doi: 10.1124/mol.66.1.113
Ghoshal, K., and Jacob, S. T. (1991). Ara-ATP impairs 3′-end processing of pre-mRNAs by inhibiting both cleavage and polyadenylation. Nucleic Acids Res. 19, 5871–5875. doi: 10.1093/nar/19.21.5871
Hamada, T., Matsunaga, S., Yano, G., and Fusetani, N. (2005). Polytheonamides A and B, highly cytotoxic, linear polypeptides with unprecedented structural features, from the marine sponge, Theonella swinhoei. J. Am. Chem. Soc. 127, 110–118. doi: 10.1021/ja045749e
Haq, I., Ladbury, J. E., Chowdhry, B. Z., Jenkins, T. C., and Chaires, J. B. (1997). Specific binding of Hoechst 33258 to the d(CGCAAATTTGCG)2 duplex: calorimetric and spectroscopic studies1. J. Mol. Biol. 271, 244–257.
Hentschel, U., Hopke, J., Horn, M., Friedrich, A. B., Wagner, M., Hacker, J., et al. (2002). Molecular evidence for a uniform microbial community in sponges from different oceans. Appl. Environ. Microbiol. 68, 4431–4440. doi: 10.1128/AEM.68.9.4431-4440.2002
Hentschel, U., Schmid, M., Wagner, M., Fieseler, L., Gernert, C., and Hacker, J. (2001). Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiol. Ecol. 35, 305–312. doi: 10.1111/j.1574-6941.2001.tb00816.x
Ibrahim, S. R., Edrada-Ebel, R., Mohamed, G. A., Youssef, D. T., Wray, V., and Proksch, P. (2008). Callyaerin G, a new cytotoxic cyclic peptide from the marine sponge Callyspongia aerizusa. ARKIVOC 12, 164–171. doi: 10.3998/ark.5550190.0009.c18
Ibrahim, S. R., Min, C. C., Teuscher, F., Ebel, R., Kakoschke, C., Lin, W., et al. (2010). Callyaerins A–F and H, new cytotoxic cyclic peptides from the Indonesian marine sponge Callyspongia aerizusa. Bioorg. Med. Chem. 18, 4947–4956. doi: 10.1016/j.bmc.2010.06.012
Johnson, B. A., Anker, H., and and, Meleney, F. L. (1945). Bacitracin: a new antibiotic produced by a member of the B. subtilis group. Science 102, 376–377. doi: 10.1126/science.102.2650.376
Kar, S., Florence, G. J., Paterson, I., and Amos, L. A. (2003). Discodermolide interferes with the binding of tau protein to microtubules. FEBS Lett. 539, 34–36. doi: 10.1016/S0014-5793(03)00181-9
Kehraus, S., König, G. M., Wright, A. D., and Woerheide, G. (2002). Leucamide A: a new cytotoxic heptapeptide from the Australian sponge Leucetta microraphis. J. Org. Chem. 67, 4989–4992. doi: 10.1021/jo020058r
Kita, M., Gise, B., Kawamura, A., and Kigoshi, H. (2013). Stylissatin A, a cyclic peptide that inhibits nitric oxide production from the marine sponge Stylissa massa. Tetrahedron Lett. 54, 6826–6828. doi: 10.1016/j.tetlet.2013.10.003
Kleinkauf, H., and von Döhren, H. (1995). The nonribosomal peptide biosynthetic system—on the origins of structural diversity of peptides, cyclopeptides and related compounds. Antonie Van Leeuwenhoek 67, 229–242. doi: 10.1007/BF00873687
Kowalski, R. J., Giannakakou, P., Gunasekera, S. P., Longley, R. E., Day, B. W., and Hamel, E. (1997). The microtubule-stabilizing agent discodermolide competitively inhibits the binding of paclitaxel (taxol) to tubulin polymers, enhances tubulin nucleation reactions more potently than paclitaxel, and inhibits the growth of paclitaxel-resistant cells. Mol. Pharmacol. 52, 613–622.
Laport, M., Santos, O., and Muricy, G. (2009). Marine sponges: potential sources of new antimicrobial drugs. Curr. Pharm. Biotechnol. 10, 86–105. doi: 10.2174/138920109787048625
Lee, I. H., Lee, Y. S., Kim, C. H., Kim, C. R., Hong, T., Menzel, L., et al. (2001). Dicynthaurin: an antimicrobial peptide from hemocytes of the solitary tunicate, Halocynthia aurantium. Biochim. Biophys. Acta 1527, 141–148. doi: 10.1016/S0304-4165(01)00156-8
Lee, Y. K., Lee, J.-H., and Lee, H. K. (2001). Microbial symbiosis in marine sponges. J. Microbiol. 39, 254–264.
Marco, E., David-Cordonnier, M.-H., Bailly, C., Cuevas, C., and Gago, F. (2006). Further insight into the DNA recognition mechanism of trabectedin from the differential affinity of its demethylated analogue ecteinascidin ET729 for the triplet DNA binding site CGA. J. Med. Chem. 49, 6925–6929. doi: 10.1021/jm060640y
Maulucci, N., Chini, M. G., Di Micco, S., Izzo, I., Cafaro, E., Russo, A., et al. (2007). Molecular insights into azumamide E histone deacetylases inhibitory activity. J. Am. Chem. Soc. 129, 3007–3012. doi: 10.1021/ja0686256
Mayer, A. M., Glaser, K. B., Cuevas, C., Jacobs, R. S., Kem, W., Little, R. D., et al. (2010). The odyssey of marine pharmaceuticals: a current pipeline perspective. Trends Pharmacol. Sci. 31, 255–265. doi: 10.1016/j.tips.2010.02.005
Mehbub, M. F., Lei, J., Franco, C., and Zhang, W. (2014). Marine sponge derived natural products between 2001 and 2010: trends and opportunities for discovery of bioactives. Mar. Drugs 12, 4539–4577. doi: 10.3390/md12084539
Mendola, D. (2000). “Aquacultural production of bryostatin 1 and ecteinascidin 743,” in Drugs from the Sea, ed N. Fusetani (Basel: Karger), 120–133. doi: 10.1159/000062482
Menna, M. (2009). Antitumor potential of natural products from Mediterranean ascidians. Phytochem. Rev. 8, 461–472. doi: 10.1007/s11101-009-9131-y
Miao, V., Coeffet-Legal, M.-F., Brian, P., Brost, R., Penn, J., Whiting, A., et al. (2005). Daptomycin biosynthesis in Streptomyces roseosporus: cloning and analysis of the gene cluster and revision of peptide stereochemistry. Microbiology 151, 1507–1523. doi: 10.1099/mic.0.27757-0
Mooberry, S. L., Randall-Hlubek, D. A., Leal, R. M., Hegde, S. G., Hubbard, R. D., Zhang, L., et al. (2004). Microtubule-stabilizing agents based on designed laulimalide analogues. Proc. Natl. Acad. Sci. U.S.A. 101, 8803–8808. doi: 10.1073/pnas.0402759101
Murthy, M. R., Mohan, E., and Sadhukhan, A. (1999). Cyclosporin-A production by Tolypocladium inflatum using solid state fermentation. Process Biochem. 34, 269–280. doi: 10.1016/S0032-9592(98)00095-8
Nakao, Y., Fujita, M., Warabi, K., Matsunaga, S., and Fusetani, N. (2000). Miraziridine A, a novel cysteine protease inhibitor from the marine sponge Theonella aff. mirabilis 1. J. Am. Chem. Soc. 122, 10462–10463. doi: 10.1021/ja001859j
Nakao, Y., Kawatsu, S., Okamoto, C., Okamoto, M., Matsumoto, Y., Matsunaga, S., et al. (2008). Ciliatamides A−C, bioactive lipopeptides from the deep-sea sponge Aaptos ciliata. J. Nat. Prod. 71, 469–472. doi: 10.1021/np8000317
Nakao, Y., Kuo, J., Yoshida, W. Y., Kelly, M., and Scheuer, P. J. (2003). More kapakahines from the marine sponge Cribrochalina olemda. Org. Lett. 5, 1387–1390. doi: 10.1021/ol026830u
Newman, D. J., and Cragg, G. M. (2016). Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 79, 629–661. doi: 10.1021/acs.jnatprod.5b01055
Newman, D. J., and Hill, R. T. (2006). New drugs from marine microbes: the tide is turning. J. Ind. Microbiol. Biotechnol. 33, 539–544. doi: 10.1007/s10295-006-0115-2
Oclarit, J., Okada, H., Ohta, S., Kaminura, K., Yamaoka, Y., Iizuka, T., et al. (1994). Anti-bacillus substance in the marine sponge, Hyatella species, produced by an associated Vibrio species bacterium. Microbios 78, 7.
Okada, Y., Matsunaga, S., van Soest, R. W., and Fusetani, N. (2002). Nagahamide A, an antibacterial depsipeptide from the marine sponge Theonella swinhoei 1. Org. Lett. 4, 3039–3042. doi: 10.1021/ol0262791
Oku, N., Gustafson, K. R., Cartner, L. K., Wilson, J. A., Shigematsu, N., Hess, S., et al. (2004). Neamphamide A, a new HIV-inhibitory depsipeptide from the Papua New Guinea marine sponge Neamphius huxleyi. J. Nat. Prod. 67, 1407–1411. doi: 10.1021/np040003f
Olivera, B. M. (2000). “ω- Conotoxin MVIIA: from marine snail venom to analgesic drug,” in Drugs from the Sea, ed N. Fusetani (Basel: Karger), 74–85. doi: 10.1159/000062483
Olson, J. B., and McCarthy, P. J. (2005). Associated bacterial communities of two deep-water sponges. Aquat. Microb. Ecol. 39, 47–55. doi: 10.3354/ame039047
Paulus, H., and Gray, E. (1964). The biosynthesis of polymyxin B by growing cultures of Bacillus polymyxa. J. Biol. Chem. 239, 865–871.
Perez, L. J., and Faulkner, D. J. (2003). Bistratamides EJ, modified cyclic hexapeptides from the Philippines ascidian Lissoclinum bistratum. J. Nat. Prod. 66, 247–250. doi: 10.1021/np0204601
Pettit, G. R., and Tan, R. (2005). Isolation and structure of Phakellistatin 14 from the Western Pacific marine sponge Phakellia sp., 1. J. Nat. Prod. 68, 60–63. doi: 10.1021/np040092w
Plaza, A., Bifulco, G., Keffer, J. L., Lloyd, J. R., Baker, H. L., and Bewley, C. A. (2008). Celebesides A−C and theopapuamides B−D, depsipeptides from an Indonesian sponge that inhibit HIV-1 entry. J. Org. Chem. 74, 504–512. doi: 10.1021/jo802232u
Plaza, A., Bifulco, G., Masullo, M., Lloyd, J. R., Keffer, J. L., Colin, P. L., et al. (2010). Mutremdamide A and koshikamides C−H, peptide inhibitors of HIV-1 entry from different Theonella species. J. Org. Chem. 75, 4344–4355. doi: 10.1021/jo100076g
Plaza, A., Gustchina, E., Baker, H. L., Kelly, M., and Bewley, C. A. (2007). Mirabamides A–D, depsipeptides from the sponge Siliquariaspongia mirabilis that inhibit HIV-1 fusion. J. Nat. Prod. 70, 1753–1760. doi: 10.1021/np070306k
Pommier, Y., Kohlhagen, G., Bailly, C., Waring, M., Mazumder, A., and Kohn, K. W. (1996). DNA Sequence- and structure-selective alkylation of guanine N2 in the DNA minor groove by ecteinascidin 743, a potent antitumor compound from the caribbean tunicate Ecteinascidia turbinata. Biochemistry 35, 13303–13309. doi: 10.1021/bi960306b
Prasad, P., Aalbersberg, W., Feussner, K.-D., and Van Wagoner, R. M. (2011). Papuamides E and F, cytotoxic depsipeptides from the marine sponge Melophlus sp. Tetrahedron 67, 8529–8531. doi: 10.1016/j.tet.2011.08.100
Qureshi, A., Colin, P. L., and Faulkner, D. J. (2000). Microsclerodermins F–I, antitumor and antifungal cyclic peptides from the lithistid sponge Microscleroderma sp. Tetrahedron 56, 3679–3685. doi: 10.1016/S0040-4020(00)00286-6
Randazzo, A., Bifulco, G., Giannini, C., Bucci, M., Debitus, C., Cirino, G., et al. (2001). Halipeptins A and B: two novel potent anti-inflammatory cyclic depsipeptides from the Vanuatu marine sponge Haliclona species. J. Am. Chem. Soc. 123, 10870–10876. doi: 10.1021/ja010015c
Ranjan, N., Davis, E., Xue, L., and Arya, D. P. (2013). Dual recognition of the human telomeric G-quadruplex by a Neomycin-anthraquinone conjugate. Chem. Commun. 49, 5796–5798. doi: 10.1039/c3cc42721h
Rashid, M. A., Gustafson, K. R., Boswell, J. L., and Boyd, M. R. (2000). Haligramides A and B, two new cytotoxic hexapeptides from the marine sponge Haliclona nigra. J. Nat. Prod. 63, 956–959. doi: 10.1021/np000051+
Rashid, M. A., Gustafson, K. R., Cartner, L. K., Shigematsu, N., Pannell, L. K., and Boyd, M. R. (2001). Microspinosamide, a new HIV-inhibitory cyclic depsipeptide from the marine sponge Sidonops microspinosa 1. J. Nat. Prod. 64, 117–121. doi: 10.1021/np0002379
Ratnayake, A. S., Bugni, T. S., Feng, X., Harper, M. K., Skalicky, J. J., Mohammed, K. A., et al. (2006). Theopapuamide, a cyclic depsipeptide from a Papua New Guinea lithistid sponge Theonella swinhoei. J. Nat. Prod. 69, 1582–1586. doi: 10.1021/np060229d
Rose, K. M., and Jacob, S. T. (1978). Selective inhibition of RNA polyadenylation by Ara-ATP in vitro: a possible mechanism for antiviral action of Ara-A. Biochem. Biophys. Res. Commun. 81, 1418–1424. doi: 10.1016/0006-291X(78)91294-9
Rudi, A., Chill, L., Aknin, M., and Kashman, Y. (2003). Didmolamide A and B, two new cyclic hexapeptides from the marine ascidian Didemnum molle. J. Nat. Prod. 66, 575–577. doi: 10.1021/np020531w
Ruppert, E., and Fox, R. (2004). Invertebrate Zoology: A Functional Evolutionary Approach (of RD Barnes' Invertebrate Zoology). Belmont, CA: Brooks/Cole.
Schmidt, E. W., Obraztsova, A., Davidson, S., Faulkner, D., and Haygood, M. (2000). Identification of the antifungal peptide-containing symbiont of the marine sponge Theonella swinhoei as a novel δ-proteobacterium,“Candidatus Entotheonella palauensis”. Mar. Biol. 136, 969–977. doi: 10.1007/s002270000273
Schmidt, E. W., Raventos-Suarez, C., Bifano, M., Menendez, A. T., Fairchild, C. R., and Faulkner, D. J. (2004). Scleritodermin A, a cytotoxic cyclic peptide from the lithistid sponge Scleritoderma nodosum. J. Nat. Prod. 67, 475–478. doi: 10.1021/np034035z
Sepe, V., D'Orsi, R., Borbone, N., D'auria, M. V., Bifulco, G., Monti, M. C., et al. (2006). Callipeltins F–I: new antifungal peptides from the marine sponge Latrunculia sp. Tetrahedron 62, 833–840. doi: 10.1016/j.tet.2005.10.058
Somma, S., Gastaldo, L., and Corti, A. (1984). Teicoplanin, a new antibiotic from Actinoplanes teichomyceticus nov. sp. Antimicrob. Agents Chemother. 26, 917–923. doi: 10.1128/AAC.26.6.917
Sonnenschein, R. N., Farias, J. J., Tenney, K., Mooberry, S. L., Lobkovsky, E., Clardy, J., et al. (2004). A further study of the cytotoxic constituents of a milnamide-producing sponge. Org. Lett. 6, 779–782. doi: 10.1021/ol036446c
Stark, W., Higgens, C., Wolfe, R., Hoehn, M., and McGuire, J. (1962). Capreomycin, a new antimycobacterial agent produced by Streptomyces capreolus sp. n. Antimicrob. Agents Chemother. 1962, 596–606.
Steele, J. H. (1985). A comparison of terrestrial and marine ecological systems. Nature 313, 355–358. doi: 10.1038/313355a0
Stierle, A., Cardellina, I. I. J., and Singleton, F. (1988). A marine Micrococcus produces metabolites ascribed to the sponge Tedania ignis. Experientia 44, 1021–1021. doi: 10.1007/BF01939910
Tabudravu, J., Morris, L. A., Kettenes-Van Den Bosch, J. J., and Jaspars, M. (2001). Wainunuamide, a histidine-containing proline-rich cyclic heptapeptide isolated from the Fijian marine sponge Stylotella aurantium. Tetrahedron Lett. 42, 9273–9276. doi: 10.1016/S0040-4039(01)01993-1
Tabudravu, J. N., Morris, L. A., Kettenes-van den Bosch, J. J., and Jaspars, M. (2002). Axinellin C, a proline-rich cyclic octapeptide isolated from the Fijian marine sponge Stylotella aurantium. Tetrahedron 58, 7863–7868. doi: 10.1016/S0040-4020(02)00898-0
Tan, L. T., Williamson, R. T., Gerwick, W. H., Watts, K. S., McGough, K., and Jacobs, R. (2000). cis, cis-and trans, trans-Ceratospongamide, new bioactive cyclic heptapeptides from the Indonesian red alga Ceratodictyon spongiosum and symbiotic sponge Sigmadocia symbiotica. J. Org. Chem. 65, 419–425. doi: 10.1021/jo991165x
Tanaka, C., Tanaka, J., Bolland, R. F., Marriott, G., and Higa, T. (2006). Seragamides A–F, new actin-targeting depsipeptides from the sponge Suberites japonicus Thiele. Tetrahedron 62, 3536–3542. doi: 10.1016/j.tet.2006.01.099
Thomas, T. R. A., Kavlekar, D. P., and Lokabharathi, P. A. (2010). Marine drugs from sponge-microbe association—a review. Mar. Drugs 8, 1417–1468. doi: 10.3390/md8041417
Tincu, J. A., Menzel, L. P., Azimov, R., Sands, J., Hong, T., Waring, A. J., et al. (2003). Plicatamide, an antimicrobial octapeptide from Styela plicata hemocytes. J. Biol. Chem. 278, 13546–13553. doi: 10.1074/jbc.M211332200
Ueda, H., Nakajima, H., Hori, Y., Fujita, T., Nishimura, M., Goto, T., et al. (1994). FR901228, a novel antitumor bicyclic depsipeptide produced by Chromobacterium violaceum No. 968. I. Taxonomy, fermentation, isolation, physico-chemical and biological properties, and antitumor activity. J. Antibiot. 47, 301–310. doi: 10.7164/antibiotics.47.301
Umezawa, H., Maeda, K., Takeuchi, T., and Okami, Y. (1966). New antibiotics, bleomycin A and B. J. Antibiot. 19, 200.
Unson, M., Holland, N., and Faulkner, D. (1994). A brominated secondary metabolite synthesized by the cyanobacterial symbiont of a marine sponge and accumulation of the crystalline metabolite in the sponge tissue. Mar. Biol. 119, 1–11. doi: 10.1007/BF00350100
Van Wageningen, A. A., Kirkpatrick, P. N., Williams, D. H., Harris, B. R., Kershaw, J. K., Lennard, N. J., et al. (1998). Sequencing and analysis of genes involved in the biosynthesis of a vancomycin group antibiotic. Chem. Biol. 5, 155–162. doi: 10.1016/S1074-5521(98)90060-6
Vervoort, H., Fenical, W., and Epifanio, R. D. A. (2000). Tamandarins A and B: new cytotoxic depsipeptides from a Brazilian ascidian of the family Didemnidae. J. Org. Chem. 65, 782–792. doi: 10.1021/jo991425a
Vicente, J., Vera, B., Rodríguez, A. D., Rodríguez-Escudero, I., and Raptis, R. G. (2009). Euryjanicin A: a new cycloheptapeptide from the Caribbean marine sponge Prosuberites laughlini. Tetrahedron Lett. 50, 4571–4574. doi: 10.1016/j.tetlet.2009.05.067
Vinothkumar, S., and Parameswaran, P. (2013). Recent advances in marine drug research. Biotechnol. Adv. 31, 1826–1845. doi: 10.1016/j.biotechadv.2013.02.006
Waksman, S. A., and Woodruff, H. B. (1940). Bacteriostatic and bactericidal substances produced by a soil actinomyces. Exp. Biol. Med. 45, 609–614. doi: 10.3181/00379727-45-11768
Webster, N. S., Wilson, K. J., Blackall, L. L., and Hill, R. T. (2001). Phylogenetic diversity of bacteria associated with the marine sponge Rhopaloeides odorabile. Appl. Environ. Microbiol. 67, 434–444. doi: 10.1128/AEM.67.1.434-444.2001
Wilkinson, C. (1978). Microbial associations in sponges. III. Ultrastructure of the in situ associations in coral reef sponges. Mar. Biol. 49, 177–185. doi: 10.1007/BF00387117
Wilkinson, C. R., Nowak, M., Austin, B., and Colwell, R. R. (1981). Specificity of bacterial symbionts in Mediterranean and Great Barrier Reef sponges. Microb. Ecol. 7, 13–21. doi: 10.1007/BF02010474
Williams, D. E., Austin, P., Diaz-Marrero, A. R., Soest, R. V., Matainaho, T., Roskelley, C. D., et al. (2005). Neopetrosiamides, peptides from the marine sponge Neopetrosia sp. that inhibit amoeboid invasion by human tumor cells. Org. Lett. 7, 4173–4176. doi: 10.1021/ol051524c
Williams, D. E., Yu, K., Behrisch, H. W., Van Soest, R., and Andersen, R. J. (2009). Rolloamides A and B, cytotoxic cyclic heptapeptides isolated from the Caribbean marine sponge Eurypon laughlini. J. Nat. Prod. 72, 1253–1257. doi: 10.1021/np900121m
Willis, B., and Arya, D. P. (2010). Triple recognition of B-DNA by a Neomycin−Hoechst 33258−Pyrene conjugate. Biochemistry 49, 452–469. doi: 10.1021/bi9016796
Woo, J.-K., Jeon, J.-E., Kim, C.-K., Sim, C. J., Oh, D.-C., Oh, K.-B., et al. (2013). Gombamide A, a cyclic thiopeptide from the sponge Clathria gombawuiensis. J. Nat. Prod. 76, 1380–1383. doi: 10.1021/np4003367
Wörheide, G., Solé-Cava, A. M., and Hooper, J. N. (2005). Biodiversity, molecular ecology and phylogeography of marine sponges: patterns, implications and outlooks. Integr. Comp. Biol. 45, 377–385. doi: 10.1093/icb/45.2.377
Keywords: marine ecosystem, sponge, tunicates, marine natural products, non-ribosomal peptides, pharmacology
Citation: Agrawal S, Adholeya A and Deshmukh SK (2016) The Pharmacological Potential of Non-ribosomal Peptides from Marine Sponge and Tunicates. Front. Pharmacol. 7:333. doi: 10.3389/fphar.2016.00333
Received: 23 March 2016; Accepted: 07 September 2016;
Published: 25 October 2016.
Edited by:
George Tegos, Massachusetts General Hospital, USAReviewed by:
Donatella De Pascale, National Research Council (CNR), ItalyPietro Tedesco, National Research Council (CNR), Italy
Copyright © 2016 Agrawal, Adholeya and Deshmukh. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sunil K. Deshmukh, c3VuaWwuZGVzaG11a2hAdGVyaS5yZXMuaW4=