 Ming Xia
Ming Xia Di Liu
Di Liu Hong Liu
Hong Liu- Department of Nephrology, The Second Xiangya Hospital, Central South University, Hunan Key Laboratory of Kidney Disease and Blood Purification, Changsha, China
Artemisinin (ARS) and its derivatives (ARSs) are recommended as the first-line antimalarial drugs for the treatment of malaria. Besides antimalarial function, its potent anti-inflammatory and immunoregulatory properties, as well as the ability to regulate oxidative stress have brought them to a prominent position. As researchers around the world are continually exploring the unknown biological activities of ARS derivatives, experimental studies have shown much progress in renal therapy. This review aims to give a brief overview of the current research on ARSs applications for kidney treatment with the evaluation of therapeutic properties and potential molecular mechanisms.
Introduction
The imbalance between the molecular mechanisms that govern oxidative stress, inflammation, immunity, and cell death are important causes of acute kidney injury (AKI) and chronic kidney diseases (CKD) (Sureshbabu et al., 2015). Both AKI and CKD can lead to diminished kidney function and are associated with high mortality and morbidity. Accumulated evidence demonstrated that natural products are alternative sources for treating renal diseases on account of the conventional experience and multi-target characteristics (Chen et al., 2018).
Artemisinin (ARS) is an effective constituent with a molecular weight of 282 originally extracted from traditional Chinese medicine Artemisia annua L, which was first discovered by Chinese scientists in 1972. Its chemical structure-sesquiterpene lactone with a peroxide bridge has been demonstrated to exert an excellent antimalarial effect (White et al., 2015; Chang, 2016). In the presence of heme or free iron, the production of reactive oxygen species and carbon-centered free radicals generated by the cleavage of the endoperoxide bridge can directly poison the parasites (Vennerstrom et al., 2004). ARS selectively kills plasmodium-infected red blood cells without destroying healthy cells, making it the recommended drug for the treatment of malaria (Lalloo et al., 2016) and more clinically effective than other antimalarial drugs such as hydroxychloroquine (HCQ) and chloroquine (CQ) (Golenser et al., 2006; Efferth and Kaina, 2010). ARS has a rapid onset of action and can be rapidly absorbed by the gastrointestinal tract after oral administration, with half-live ranging from 2 to 5 h. It is mainly distributed in the liver, kidney, and bile, and approximately 80% of the drug was excreted through the urine and feces within 24 h of administration (German and Aweeka, 2008; Li, 2012). Currently, a series of ARS derivatives (ARSs) with improved pharmacological features are used in clinical treatment including artemether (ARM), artesunate (ART), β-aminoarteether maleate (SM934), and dihydroartemisinin (DHA) (chemical structures were shown in Figure 1). The half-lives of ARM (2–4 h), ART (< 1 h), DHA (∼1 h) are shorter (Krishna et al., 2004; German and Aweeka, 2008) and oral intake represents a relatively safe route in the clinic.
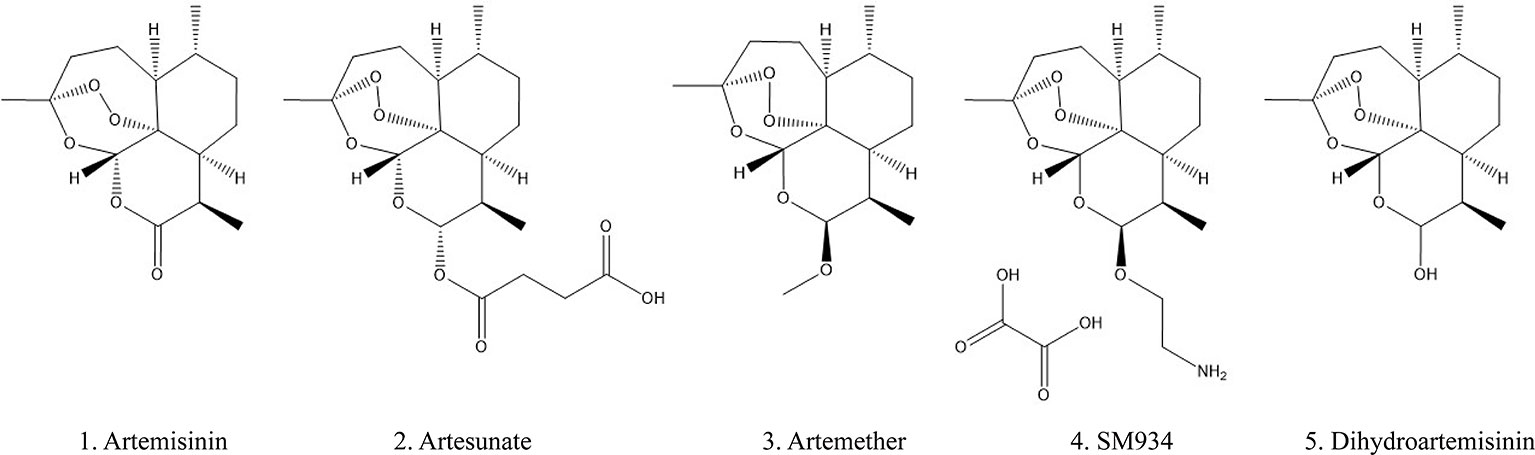
Figure 1 Chemical structures of artemisinin and its derivatives.
In addition to decades of remarkable progress against malaria, studies have demonstrated a variety of other pharmacological effects beyond antimalarial, such as anti-virus, anti-neoplastic, anti-inflammation as well as immunosuppressive effects (An J. et al., 2017). The properties of ARSs have been intensively reviewed in systemic lupus erythematosus, rheumatoid, arthritis, multiple sclerosis, etc (Shi et al., 2015; Mu and Wang, 2018), while the treatment of kidney disease has not been summarized. This review will focus on the proposed therapeutic properties and mechanisms of ARSs in kidney disease, and discuss the potential application of ARSs as novel agents for future treatment.
Search Strategy
Comprehensive literature searches for candidate studies were undertaken in two English and three Chinese biomedical databases from inception through February 2020. These databases included PubMed, Springer, Chinese National Knowledge Infrastructure, WanFang Med Online, and Chinese Biomedical Databases. Searches were limited to studies in English and Chinese. The following terms were used in the search: “artemisinin,” “artemisinins,” “artesunate,” “dihydroartemisinin,” “artemether,” “SM934,” “β-aminoarteether maleate,” “kidney,” “renal,” “nephro,” “nephritis,” nephropathy.”
Overview of Researches on ARSs in Renal Disease
The effects of ARSs were mainly studied on animal models and cells, with two clinical studies targeting lupus nephritis (LN). In a randomized, 5-year follow-up clinical trial for LN (Lu, 2002), the treatment group (ARS 0.6 g/d and cordyceps 3–4 g/d) was reported to improve 24 h urine protein, creatinine clearance rate, level of C3, and was more effective than the control group (tripterygium wilfordii polyglycosides tablets 1 mg/kg, three times a day and/or baoshenkang tablets 150 mg, three times a day). A 2-month randomized trial had similar findings (Li et al., 2011), treatment with ART (50 mg, twice a day) was reported to improve systemic symptoms and reduce the immunological activity index than either before treatment or the control group with tripterygium wilfordii polyglycosides tablets (10 mg, three times a day and/or prednisone 0.5 mg/kg/d). However, these two studies were not blinded and placebo-controlled, which may result in information bias, and observations of larger samples are still lacking.
Table 1 summarized the characteristics of animal studies with ARSs treatment, covering LN, adriamycin nephropathy (AN), subtotal nephrectomy (SNx), IgA nephropathy (IgAN), diabetic nephropathy (DN), AKI, unilateral ureteral obstruction (UUO), pristine or lipopolysaccharide (LPS)-induced nephritis, nephrotic syndrome (NS), and Heymann nephritis (HN). In vitro cell models and in vivo animal models investigations for ARSs efficacy on kidney disease involve various aspects including oxidative stress, inflammation action, and immune response, we will describe the effects of ARSs in sections below.

Table 1 Study characteristics of animal experiments in kidney disease.
Mechanism of Arss in Kidney
Oxidative Stress Regulation of Artemisinins
Oxidative stress is an important mediator in the development and progression of CKD and AKI and its complications due to increased production of reactive oxygen species (ROS) and diminished antioxidant capacity (Ruiz et al., 2013). In the condition of a surplus of ROS, ARSs were reported to exhibit an antioxidant effect (Kim et al., 2014; Yang et al., 2018; Liu et al., 2019). In addition, according to the characteristics of ARSs, tumor cells are more vulnerable due to higher levels of iron (Robert et al., 2005) and are more susceptible to further ROS insults induced by ARSs (Hamacher-Brady et al., 2011; Efferth, 2017). Accordingly, ARSs have the potential to treat kidney cancer, and the emergence of new technologies such as ARS-based smart nanomedicine offers more possibilities (Luo et al., 2019).
In Vitro
Receptor-interacting protein kinase 1 (RIP1) is verified to modulate mitochondrial ROS production via excessive generation of mitochondrial superoxide and depletion of GSH (Zhou et al., 2017). ART was reported to induce ROS production and cell death in human renal carcinoma cells, while pretreatment with RIP1 inhibitor or knockdown of RIP1 reverted ART-induced cytotoxicity (Chauhan et al., 2017).
In Vivo
Pretreated DHA or ARS could ameliorate oxidative stress in AKI mice by restoring malonyl dialdehyde (MDA), nitric oxide (NO), glutathione peroxidase (GSH), catalase (CAT), and superoxide dismutase (SOD) activity in the kidney (An Y. et al., 2017; Liu et al., 2019). In addition, ARM was shown to reduce the serum H2O2 level and elevated renal cortical PGC-1α expression, but it did not exert obvious effects on CAT and SOD expression in the renal cortex in DN (Han et al., 2019). In normal rats without oxidative stress, orally taken artemether-lumefantrine (1.14/6.86 mg/kg/d, twice a day) or artesunate-amodiaquine (2.86/8.58 mg/kg/d, twice a day) for 7 d did not apparently alter renal antioxidant status compared with the control. Although there was no significant alteration in kidney, liver, lung, and brain weights, the artesunate-amodiaquine group showed cardiotoxicity (decreased heart weight by 27.2% compared with control) (Otuechere et al., 2012).
To date, ARSs could trigger cell death by inducing oxidative stress, and could also resist oxidation to reduce cell damage. Detailed understanding of the molecular mechanisms and the events by which ARSs regulate oxidative stress to control cellular processes in different cells remain to be explored.
Anti-Inflammation Effect of ARSs
Inflammation plays a pivotal role in the pathophysiological processes of kidney diseases and associated with renal injury (Ernandez and Mayadas, 2016). The anti-inflammatory effects of ARS have been widely recognized, including repression of nuclear factor-κB (NF-κB), toll-like receptors (TLRs), signal transducer and activator of transcription (STAT), and phosphatidylinositol-3-kinase (PI3K)/protein kinase B (AKT) activity (Aldieri et al., 2003; Ho et al., 2014; Shi et al., 2015), which are key factors mediating immune-inflammatory response and are associated with kidney disease progression (Ruiz-Andres et al., 2016).
In Vitro
ART ameliorated high glucose-induced injury by suppressing TLR4/NF-κB/nod-like receptor protein 3 (NLRP3) inflammasome pathway in rat glomerular mesangial cell (Sun et al., 2018).
In Vivo
For LN mice, it has been reported that treatment with ARSs could decrease interferon-gamma (IFN-γ), tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6) in serum, and blocked intercellular adhesion molecule-1 (ICAM-1), fractalkine, NF-κB signaling pathway in renal (Dong et al., 2003; Li et al., 2006; Wang et al., 2010; Wu X. et al., 2010; You et al., 2014; Liang et al., 2018). A similar effect was observed in the nephritis and septic AKI models (Wu P. et al., 2011; Wan and Li, 2017; Liu et al., 2019). Also, ARSs were reported to inhibit disease progression via downregulating renal monocyte chemoattractant protein 1 (MCP-1) expression in LN, IgAN, and septic AKI models (Mi et al., 2009; Jin et al., 2009; Ma et al., 2009; Liu et al., 2019). In addition, ARSs were proved to alleviate the tubule-interstitial inflammation and fibrosis by inhibiting NF‐κB and mothers against decapentaplegic homolog (Smad) signaling pathway in SNx rats, Heymann nephritis rats and UUO models (Ma et al., 2010; Li et al., 2015; Wen et al., 2019). ARSs were also reported to reduce diabetic kidney damage by inhibiting TLR4, IL-8, and the DNA-binding activity of NF−κB in renal (Zhou et al., 2014a; Nie et al., 2015).
These data indicate that ARSs act as anti-inflammatory drugs at multiple components of inflammation signals and have a potential therapeutic effect on disease activity.
Immunoregulatory Effect of ARSs
Under physiological conditions, the kidney contributes to immune homeostasis, assist in the removal of metabolic wastes and toxins, and maintain peripheral tolerance. The disruption of immune homeostasis an autoimmune response, such as the occurrence of LN and IgAN, resulting in the loss of renal function (Tecklenborg et al., 2018). T cells, B cells, and macrophages, as well as cytokines, are involved in immune regulation and are activated to varying degrees depending on disease pattern. The activated pathogenic cells are more likely to lead the breakdown of the peroxide bridge structure of ARS (Shi et al., 2015).
In Vitro
Chemokine ligand 2 (CCL2) and single immunoglobulin IL-1-related receptor (SIGIRR) are involved in the inflammatory pathogenesis of LN, DHA was reported to inhibit CCL2 secretion and increase SIGIRR expression and protect LPS-induced HK-2 inflammation (Huang et al., 2015).
In Vivo
ARS combined with prednisone was reported to increase the sensitivity of glucocorticoid compared to the group administrated glucocorticoid only in LN (Wu X.L et al., 2011), which may offer a possibility of alleviating the common side effects of existing glucocorticoids or immunosuppressants. SM934 was shown to protect LN mice by inhibiting both Th1 cells and Th17 cells responses (Hou et al., 2011) and reduce the number of activated B cells by inhibiting the expression of TLR7/9 (Gui et al., 2019). SIGIRR, as an inhibitor of TLR signal transduction, could be elevated by DHA. This might be a negative immune-modulating way for DHA to slow the progression of LN (Huang et al., 2015). In addition, ARS combined with HCQ was shown to improve IgAN rats immunity (Lin et al., 2016), possibly via inhibiting the differentiation of Th2 and Th17 cells while promoting Th1 and Treg cells differential (Bai et al., 2019).
All these studies suggest that ARS family drugs are able to perform immunosuppressive functions primarily through suppressing the activation of pathogenic immune cells and have a regulatory effect on autoimmune diseases.
Other Effects
Anti-Fibrosis
Myofibroblasts can be differentiated by the epithelial-mesenchymal transformation (EMT) process, and are primarily responsible for excessive extracellular matrix production. TGF-β1, smooth muscle actin (α-SMA) and connective tissue growth factor (CTGF), metalloprotease (MMP), bone morphogenetic protein (BMP) are all considered to be major regulators of EMT and renal fibrosis (Liu et al., 2018). For UUO, both in vitro and in vivo study showed anti-fibrosis effect of ARSs related to the inhibition of EMT, fibroblast proliferation, and collagen synthesis (Zhang et al., 2017). And upregulating BMP-7 and downregulating BMP antagonist-uterine sensitization-associated gene-1 (USAG-1) (Cao et al., 2016), or mitigating CTGF, α-SMA (Mi et al., 2007), or PI3K/AKT pathway (Zhang et al., 2019) are all possible mechanisms.
Anti-Proliferation
Glomerular mesangial cell proliferation is a common pathological change of glomerular disease, effective control of mesangial cell proliferation is of great clinical significance. ARSs were reported to exert an inhibitory effect on the proliferation of rat mesangial cells (MA et al., 2007a; Ma et al., 2007b), possibly by inducing apoptosis and downregulating inflammatory cytokines TNF-α and IL-6 (Wang et al., 2016) or enhance caspase-3 activity (Wu X.L. et al., 2010). Our team recently demonstrated that DHA could inhibit the proliferation of aIgA1-induced human mesangial cells through the mTOR signaling pathway in vitro (Xia et al., 2020). In addition, ARSs were found to inhibit renal carcinoma cell proliferation by inhibiting the expression of fascin (Zhang et al., 2018), meanwhile inhibiting colony formation, migration, invasion, and tumorigenesis (Yu et al., 2019).
Regulate Glomerular Filtration
Glomerular permeability is regulated by the glomerular filtration barrier (GFB), which composed of glomerular endothelium, the glomerular basement membrane, and the podocyte layer. The dysfunction of intercellular adhesion and connection will result in the loss of the structural and functional integrity of GFB and the occurrence of proteinuria (Mehta and Malik, 2006). ART was proved to reduce glomerular permeability and improve proteinuria in LN mice by inhibiting vascular endothelial growth factor (VEGF) (Jin et al., 2007). Studies also showed that DHA ameliorated the hyperpermeability of GFB by inhibiting TNF-α and maintaining occludin expression (Cheng et al., 2018) or elevation of vascular endothelial (VE)-cadherin expression in endothelial cells (Li et al., 2018). In addition, ARS was observed to attenuate podocyte effacement and fusion via nephrin and podocin regulation in adriamycin-induced nephropathy (Wu et al., 2014), and reduce the shedding of podocyte and excretion of nephrin and podocin in Heymann nephritis (Liu et al., 2017).
Anti-Virus
ARS was shown to be effective in inhibiting polyomavirus BK replication in primary human kidney cells (Sharma et al., 2014).
Renal-Protective.
ART was reported to ameliorate proteinuria and suppress the progression of NS (Razavi et al., 2007). Studies also showed that ARS could relieve renal lesions in DN rats, through inhibiting platelet-derived growth factor-B (PDGF-B) expression (Zhang et al., 2014a), metalloproteinase tissue inhibitor-2 (TIMP-2) (Zhang et al., 2014b), spatiotemporal dynamics activation of protein kinase c (PKC) (Zhang et al., 2014c) and its downstream c-fos and c-jun (Zhou et al., 2014b), and their heterodimer activator protein (AP-1) (Zhou et al., 2014c). The results from the high-throughput sequence from DN rats treated with ARS may identifying promising targets for future treatment (Xiang et al., 2019). In addition, kidney function was found to be improved in cases of malarial nephropathy after treatment with ARSs (Ezzedine et al., 2007; Calitri et al., 2014; Gleeson et al., 2019).
Interaction, Safety, and Side Effects
The toxicity of ARSs in cell culture, animals (mice, rats, rabbits, dogs, monkeys), and human clinical trials were well described (Efferth and Kaina, 2010). Large clinical studies and meta-analyses did not show serious side effects, despite mild and self-limited effects including mild nausea, vomiting, and diarrhea (Mssusa et al., 2016). Individual patients may appear transient transaminase elevated and mild rash. Non-hematological side effects include mild hepatitis, neurological, renal, cutaneous, and cardiac manifestations were uncommon (Roussel et al., 2017). Rare severe adverse events include prolongation of the QTc interval and cardiac arrhythmias (in LiverTox, 2012).
In addition, animal studies showed that artesunate can reduce glomerular filtration rate, increase renal blood flow, and has certain organ toxicity (Campos et al., 2001; Otuechere et al., 2012), while in a clinical study, liver function, kidney function, and routine blood tests remained normal in most patients treated with artesunate (von Hagens et al., 2017). A systematic review and meta-analysis showed that the use of ARS-based combination therapy in adults, children, and pregnant women in the 2nd or 3rd trimester was relatively safe (Kovacs et al., 2016). The drug interactions of ARSs are relatively unknown, more rigorous and comprehensive studies of interaction mechanisms are needed, as well as monitoring the safety of ARSs, especially concerning the genotoxicity and embryotoxicity (Amorim et al., 2013).
Conclusion and Future Directions
Much knowledge has been gained about the antimalarial drugs in recent years, and more attention has been paid to ARSs application for renal damages. Many years of laboratory applications and research proved that ARSs have excellent anti-inflammatory and immunoregulatory functions. It is also a good regulator of the balance between oxidation and oxidation resistance. The regulation of the glomerular barrier highlights a unique aspect of the use of ARSs in kidney disease.
Despite accumulating evidence on the use of ARSs, the literature on its potential as a treatment for renal disease is still insufficient due to the lack of randomized controlled clinical trials. The additive effects of ARSs in combined administration with immunosuppressants and the structure-activity relationship need to be further clarified. Investigation of the improved properties of ARSs analogs also facilitates the discovery of novel drug targets for kidney disease (Santos et al., 2015; Zuma et al., 2016; de Lange et al., 2018).
Author Contributions
MX provided direction, collected related literature, and drafted the manuscript. DL and YL made significant revisions to the manuscript and directed the review to be more focused. HL gave the final approval for the article to be published. All authors have read and approved the final manuscript.
Funding
This work was supported by a research grant (81770714), (81470947), and (81570622) from the National Natural Science Foundation of China. It also funded by the postgraduate innovation project of Central South University in China (Project no.2018zzts920).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Aldieri, E., Atragene, D., Bergandi, L., Riganti, C., Costamagna, C., Bosia, A., et al. (2003). Artemisinin inhibits inducible nitric oxide synthase and nuclear factor NF-kB activation. FEBS Lett. 552 (2-3), 141–144. doi: 10.1016/S0014-5793(03)00905-0
Amorim, M. H., Gil da Costa, R. M., Lopes, C., Bastos, M. M. (2013). Sesquiterpene lactones: adverse health effects and toxicity mechanisms. Crit. Rev. Toxicol. 43 (7), 559–579. doi: 10.3109/10408444.2013.813905
An, J., Minie, M., Sasaki, T., Woodward, J. J., Elkon, K. B. (2017). Antimalarial Drugs as Immune Modulators: New Mechanisms for Old Drugs. Annu. Rev. Med. 68, 317–330. doi: 10.1146/annurev-med-043015-123453
An, Y., Li, Q., Sui, C. H., Wang, C. (2017). Effects of artemisinin intragastric administration on acute kidney injury induced by cisplatin in mice. Shandong Med. J. 57 (11), 36–39. doi: 10.3969/j.issn.1002-266X.2017.11.011
Bai, L., Li, H., Li, J., Song, J., Zhou, Y., Liu, B., et al. (2019). Immunosuppressive effect of artemisinin and hydroxychloroquine combination therapy on IgA nephropathy via regulating the differentiation of CD4+ T cell subsets in rats. Int. Immunopharmacol. 70, 313–323. doi: 10.1016/j.intimp.2019.02.056
Calitri, C., Garazzino, S., Camilla, R., Peruzzi, L., Amore, A., Tovo, P. A., et al. (2014). “Collateral” effect of artemether in an atypical kidney involvement by Plasmodium falciparum malaria: a case report. J. Vector Borne Dis. 51 (2), 147–148.
Campos, S. B., Rouch, L. H., Seguro, A. C. (2001). Effects of sodium artesunate, a new antimalarial drug, on renal function. Kidney Int. 59 (3), 1044–1051. doi: 10.1046/j.1523-1755.2001.0590031044.x
Cao, J., Wang, W., Li, Y., Xia, J., Peng, Y., Zhang, Y., et al. (2016). Artesunate attenuates unilateral ureteral obstruction-induced renal fibrosis by regulating the expressions of bone morphogenetic protein-7 and uterine sensitization-associated gene-1 in rats. Int. Urol Nephrol. 48 (4), 619–629. doi: 10.1007/s11255-016-1232-0
Chang, Z. (2016). The discovery of Qinghaosu (artemisinin) as an effective anti-malaria drug: A unique China story. Sci. China Life Sci. 59 (1), 81–88. doi: 10.1007/s11427-015-4988-z
Chauhan, A. K., Min, K. J., Kwon, T. K. (2017). RIP1-dependent reactive oxygen species production executes artesunate-induced cell death in renal carcinoma Caki cells. Mol. Cell Biochem. 435 (1-2), 15–24. doi: 10.1007/s11010-017-3052-7
Chen, D. Q., Hu, H. H., Wang, Y. N., Feng, Y. L., Cao, G., Zhao, Y. Y., et al. (2018). Natural products for the prevention and treatment of kidney disease. Phytomedicine 50, 50–60. doi: 10.1016/j.phymed.2018.09.182
Cheng, Z., Qi, R., Li, L., Liu, Q., Zhang, W., Zhou, X., et al. (2018). Dihydroartemisinin ameliorates sepsis-induced hyperpermeability of glomerular endothelium via up-regulation of occludin expression. BioMed. Pharmacother. 99, 313–318. doi: 10.1016/j.biopha.2018.01.078
de Lange, C., Coertzen, D., Smit, F. J., Wentzel, J. F., Wong, H. N., Birkholtz, L. M., et al. (2018). Synthesis, antimalarial activities and cytotoxicities of amino-artemisinin-1,2-disubstituted ferrocene hybrids. Bioorg Med. Chem. Lett. 28 (19), 3161–3163. doi: 10.1016/j.bmcl.2018.08.037
Dong, Y. J., Li, W. D., Tu, Y. Y., Zhang, H. N., Zou, W. Z., Yan, L., et al. (2003). The effects of DQHS on the pathologic changes in BXSB mice lupus nephritis and the effect mechanism. Chin. Pharmacol. Bull. 19 (10), 1125–1128. doi: 10.3321/j.issn:1001-1978.2003.10.011
Efferth, T., Kaina, B. (2010). Toxicity of the antimalarial artemisinin and its dervatives. Crit. Rev. Toxicol. 40 (5), 405–421. doi: 10.3109/10408441003610571
Efferth, T. (2017). From ancient herb to modern drug: Artemisia annua and artemisinin for cancer therapy. Semin. Cancer Biol. 46, 65–83. doi: 10.1016/j.semcancer.2017.02.009
Ernandez, T., Mayadas, T. N. (2016). The Changing Landscape of Renal Inflammation. Trends Mol. Med. 22 (2), 151–163. doi: 10.1016/j.molmed.2015.12.002
Ezzedine, K., Pistone, T., Receveur, M. C., Cressend, T., Dieval, C., Malvy, D., et al. (2007). Haemoglobinuria in a 38-year-old French expatriate man living in Cameroon following artemisinin-based antimalarial treatment. Travel Med. Infect. Dis. 5 (4), 256–258. doi: 10.1016/j.tmaid.2007.01.003
German, I., Aweeka, F. T. (2008). Clinical pharmacology of artemisinin-based combination therapies. Clin. Pharmacokinet. 47 (2), 91–102. doi: 10.2165/00003088-200847020-00002
Gleeson, P. J., O'Regan, J. A., McHale, T., Tuite, H., Giblin, L., Reddan, D., et al. (2019). Acute interstitial nephritis with podocyte foot-process effacement complicating Plasmodium falciparum infection. Malar J. 18 (1), 58. doi: 10.1186/s12936-019-2674-5
Golenser, J., Waknine, J. H., Krugliak, M., Hunt, N. H., Grau, G. E. (2006). Current perspectives on the mechanism of action of artemisinins. Int. J. Parasitol. 36 (14), 1427–1441. doi: 10.1016/j.ijpara.2006.07.011
Gui, J. X., Cao, L., Wang, B., Zhang, Y., Li, Y. Y., Xiong, A. L., et al. (2019). Effects of artemisinin derivatives SM934 inhibiting TLR7/9 signaling pathway on B cells in systemic lupus erythematosus mice. Chin J. Clin. Pharmacol. 35 (19), 2343–2346. doi: 10.13699/j.cnki.1001-6821.2019.19.041
Hamacher-Brady, A., Stein, H. A., Turschner, S., Toegel, I., Mora, R., Jennewein, N., et al. (2011). Artesunate activates mitochondrial apoptosis in breast cancer cells via iron-catalyzed lysosomal reactive oxygen species production. J. Biol. Chem. 286 (8), 6587–6601. doi: 10.1074/jbc.M110.210047
Han, P., Wang, Y., Zhan, H., Weng, W., Yu, X., Ge, N., et al. (2019). Artemether ameliorates type 2 diabetic kidney disease by increasing mitochondrial pyruvate carrier content in db/db mice. Am. J. Transl. Res. 11 (3), 1389–1402.
Ho, W. E., Peh, H. Y., Chan, T. K., Wong, W. S. (2014). Artemisinins: pharmacological actions beyond anti-malarial. Pharmacol. Ther. 142 (1), 126–139. doi: 10.1016/j.pharmthera.2013.12.001
Hou, L. F., He, S. J., Li, X., Yang, Y., He, P. L., Zhou, Y., et al. (2011). Oral administration of artemisinin analog SM934 ameliorates lupus syndromes in MRL/lpr mice by inhibiting Th1 and Th17 cell responses. Arthritis Rheum 63 (8), 2445–2455. doi: 10.1002/art.30392
Huang, M., Jin, X. K., Cai, Q. C., Li, M., Lin, Z. B., Li, W. D., et al. (2015). Therapeutic effect of DHA on lupus nephritis and its relationship with SIGIRR inducing immune negative regulation. Chin. J. Immunol. 31 (12), 1637–1647. doi: 10.3969/j.issn.1000-484X.2015.12.012
in LiverTox (2012). Clinical and Research Information on Drug-Induced Liver Injury. (Bethesda (MD): National Institute of Diabetes and Digestive and Kidney Diseases).
Jin, O., Zhang, H., Xu, T., Zhao, S., Zhou, K., Sun, L., et al. (2007). Pathological Change and Mechanism Of Artesunate Treatment for Lupus Nephritis in MRL/lpr Mice. J. Clin. Med. Pract. 11 (4), 5–9. doi: 10.3969/j.issn.1672-2353.2007.07.003
Jin, O., Zhang, H., Gu, Z., Zhao, S., Xu, T., Zhou, K., et al. (2009). A pilot study of the therapeutic efficacy and mechanism of artesunate in the MRL/lpr murine model of systemic lupus erythematosus. Cell Mol. Immunol. 6 (6), 461–467. doi: 10.1038/cmi.2009.58
Kim, M. H., Seo, J. Y., Liu, K. H., Kim, J. S. (2014). Protective effect of Artemisia annua L. extract against galactose-induced oxidative stress in mice. PloS One 9 (7), e101486. doi: 10.1371/journal.pone.0101486
Kovacs, S. D., van Eijk, A. M., Sevene, E., Dellicour, S., Weiss, N. S., Emerson, S., et al. (2016). The Safety of Artemisinin Derivatives for the Treatment of Malaria in the 2nd or 3rd Trimester of Pregnancy: A Systematic Review and Meta-Analysis. PloS One 11 (11), e0164963. doi: 10.1371/journal.pone.0164963
Krishna, S., Uhlemann, A. C., Haynes, R. K. (2004). Artemisinins: mechanisms of action and potential for resistance. Drug Resist. Update 7 (4-5), 233–244. doi: 10.1016/j.drup.2004.07.001
Lalloo, D. G., Shingadia, D., Bell, D. J., Beeching, N. J., Whitty, C. J. M., Chiodini, P. L., et al. (2016). UK malaria treatment guidelines 2016. J. Infect. 72 (6), 635–649. doi: 10.1016/j.jinf.2016.02.001
Li, W. D., Dong, Y. J., Tu, Y. Y., Lin, Z. B. (2006). Dihydroarteannuin ameliorates lupus symptom of BXSB mice by inhibiting production of TNF-alpha and blocking the signaling pathway NF-kappa B translocation. Int. Immunopharmacol. 6 (8), 1243–1250. doi: 10.1016/j.intimp.2006.03.004
Li, X., Wei, Y., Zhang, P. L., Deng, Z. Y. (2011). Clinical study on immune function regulation of artesunate in patients with lupus nephritis. Lishizhen Med. Materia Med. Res. 22 (7), 1673–1674. doi: 10.3969/j.issn.1008-0805.2011.07.057
Li, T. T., Zhang, X. H., Jing, J. F., Li, X., Yang, X. Q., Zhu, F. H., et al. (2015). Artemisinin analogue SM934 ameliorates the proteinuria and renal fibrosis in rat experimental membranous nephropathy. Acta Pharmacol. Sin. 36 (2), 188–199. doi: 10.1038/aps.2014.134
Li, L., Chen, X., Dong, F., Liu, Q., Zhang, C., Xu, D., et al. (2018). Dihydroartemisinin up-regulates VE-cadherin expression in human renal glomerular endothelial cells. J. Cell Mol. Med. 22 (3), 2028–2032. doi: 10.1111/jcmm.13448
Li, Y. (2012). Qinghaosu (artemisinin): chemistry and pharmacology. Acta Pharmacol. Sin. 33 (9), 1141–1146. doi: 10.1038/aps.2012.104
Liang, N., Zhong, Y., Zhou, J., Liu, B., Lu, R., Guan, Y., et al. (2018). Immunosuppressive effects of hydroxychloroquine and artemisinin combination therapy via the nuclear factor-kappaB signaling pathway in lupus nephritis mice. Exp. Ther. Med. 15 (3), 2436–2442. doi:10.3892/etm.2018.5708
Lin, X. Q., Yuan, J., Song, J. P., Guan, Y. Z., Fu, L., Zhou, J. Y., et al. (2016). Experimental Study on Artemisinin Combined with Hydroxychloroquine in Treating Rats with IgA Nephropathy. Traditional Chin. Drug Res. & Clin. Pharmacol. 27 (5), 608–613. doi: 10.19378/j.issn.1003-9783.2016.05.003
Liu, L. Z., Zhang, M. X., Wang, S. (2017). Effects of artemisinin on excretion of nephrin, podocin mRNA and urinary podocyte cells in Heymann nephritis rats. Chin. Traditional Patent Med. 39 (10), 2176–2178. doi: 10.3969/j.issn.1001-1528.2017.10.041
Liu, J. H., He, L., Zou, Z. M., Ding, Z. C., Zhang, X., Wang, H., et al. (2018). A Novel Inhibitor of Homodimerization Targeting MyD88 Ameliorates Renal Interstitial Fibrosis by Counteracting TGF-beta1-Induced EMT in Vivo and in Vitro. Kidney Blood Press Res. 43 (5), 1677–1687. doi: 10.1159/000494745
Liu, X., Lu, J., Liao, Y., Liu, S., Chen, Y., He, R., et al. (2019). Dihydroartemisinin attenuates lipopolysaccharide-induced acute kidney injury by inhibiting inflammation and oxidative stress. BioMed. Pharmacother. 117, 109070. doi: 10.1016/j.biopha.2019.109070
Lu, L. (2002). Study on effect of Cordyceps sinensis and artemisinin in preventing recurrence of lupus nephritis. Chin. J. Integrated Traditional Western Med. 22 (3), 169–171. doi: 10.3321/j.issn:1003-5370.2002.03.003
Luo, Y., et al. (2019). Artemisinin-Based Smart Nanomedicines with Self-Supply of Ferrous Ion to Enhance Oxidative Stress for Specific and Efficient Cancer Treatment. ACS Appl. Mater Interfaces 11 (33), 29490–29497. doi: 10.1021/acsami.9b07390
Ma, X. Y., Yang, M., Mi, X. H., Wu, W. H., Zeng, Y., Ma, Y. X., et al. (2007a). Effects of artesunate on the proliferation of rats’ glomerular mesangial cells. Modern Preventive Med. 34 (23), 4415–4416, 4439. doi: 10.3969/j.issn.1003-8507.2007.23.007
MA, X. Y., Yang, M., Wu, W. H., Zeng, Y., Mi, X. H. (2007b). Effects of artesunate on the cell cycle of rat glomerular mesangial cells. Sichuan Med. J. 28 (12), 1308. doi: 10.3969/j.issn.1004-0501.2007.12.001
Ma, Y. X., Luan, S., Ma, X. Y., Zeng, Y., Wu, W. H., Mi, X. H., et al. (2009). The effect of artemisia on IL-2 and IL-6 in IgA nephropathy in rat. Med. J. West China 21 (5), 716–718. doi: 10.3969/j.issn.1672-3511.2009.05.005
Ma, X. Y., Wu, W. H., Zeng, Y., Ma, Y. X., Mi, X. H. (2010). The anti-inflammatory effect and its mechanism of artemisia on the renal of UUO rats. Chongqing Med. 39 (12), 1514–1516. doi: 10.3969/j.issn.1671-8348.2010.12.013
Mehta, D., Malik, A. B. (2006). Signaling mechanisms regulating endothelial permeability. Physiol. Rev. 86 (1), 279–367. doi: 10.1152/physrev.00012.2005
Mi, X. H., Luan, S., Zeng, Y., Shu, B., Wu, W. H., Ma, Y. X., et al. (2007). Effects of Artesunate on the expression of CTGF and α-SMA in renal tissue of unilateral ureteral obstructed rat model. Med. J. West China 19 (3), 332–338. doi: 10.3969/j.issn.1672-3511.2007.03.002
Mi, X. H., Luan, S., Ma, X. Y., Ma, Y. X., Zeng, Y., Wu, W. H., et al. (2009). Effects of Artesunate on MCP-1 and MCP-1 mRNA expression of renal tissue in the rat IgA nephropathy model. J. Sichuan Univ (Med Sci. Edi) 40 (5), 821–825. doi: 10.3969/j.issn.1672-173X.2009.05.013
Mssusa, A. K., Fimbo, A. M., Nkayamba, A. F., Irunde, H. F., Sillo, H. B., Shewiyo, D. H., et al. (2016). Safety Profile of Artemether-Lumefantrine: A Cohort Event Monitoring Study in Public Health Facilities in Tanzania. Clin. Drug Invest. 36 (5), 401–411. doi: 10.1007/s40261-016-0385-z
Mu, X., Wang, C. (2018). Artemisinins-a Promising New Treatment for Systemic Lupus Erythematosus: a Descriptive Review. Curr. Rheumatol Rep. 20 (9), 55. doi: 10.1007/s11926-018-0764-y
Nie, H., Su, K., Long, Y., Jang, S. S., Sun, Q. (2015). Effect of artesunate on expressions of Toll-like receptor 4 and interleukin-8 in renaI tissues of diabetic nephropathy rat. Tianjin Med. J. 43 (4), 356–359. doi: 10.11958/j.issn.0253-9896.2015.04.006
Otuechere, C. A., Edewor, G., Kale, O. E., Ekor, M. (2012). Subacute therapeutic dosing of artemether-lumefantrine and artesunate-amodiaquine combination preserves plasma cholesterol, renal antioxidant status, and organ weights in rats. Malar Res. Treat 2012, 257986. doi: 10.1155/2012/257986
Razavi, A., Nouri, H. R., Mehrabian, F., Mirshafiey, A. (2007). Treatment of experimental nephrotic syndrome with artesunate. Int. J. Toxicol. 26 (4), 373–380. doi: 10.1080/10915810701493293
Robert, A., Benoit-Vical, F., Claparols, C., Meunier, B. (2005). The antimalarial drug artemisinin alkylates heme in infected mice. Proc. Natl. Acad. Sci. U S A 102 (38), 13676–13680. doi: 10.1073/pnas.0500972102
Roussel, C., Caumes, E., Thellier, M., Ndour, P. A., Buffet, P. A., Jaureguiberry, S., et al. (2017). Artesunate to treat severe malaria in travellers: review of efficacy and safety and practical implications. J. Travel Med. 24 (2), 1–9. doi: 10.1093/jtm/taw093
Ruiz, S., Pergola, P. E., Zager, R. A., Vaziri, N. D. (2013). Targeting the transcription factor Nrf2 to ameliorate oxidative stress and inflammation in chronic kidney disease. Kidney Int. 83 (6), 1029–1041. doi: 10.1038/ki.2012.439
Ruiz-Andres, O., Sanchez-Niño, M. D., Moreno, J. A., Ruiz-Ortega, M., Ramos, A. M., Sanz, A. B., et al. (2016). Downregulation of kidney protective factors by inflammation: role of transcription factors and epigenetic mechanisms. Am. J. Physiology-Renal Physiol. 311 (6), F1329–F1340. doi: 10.1152/ajprenal.00487.2016
Santos, C. B., Lobato, C. C., Braga, F. S., Costa Jda, S., Favacho, H. A., Carvalho, J. C., et al. (2015). Rational Design of Antimalarial Drugs Using Molecular Modeling and Statistical Analysis. Curr. Pharm. Des. 21 (28), 4112–4127. doi: 10.2174/1381612821666150528121423
Sharma, B. N., Marschall, M., Henriksen, S., Rinaldo, C. H. (2014). Antiviral effects of artesunate on polyomavirus BK replication in primary human kidney cells. Antimicrob Agents Chemother. 58 (1), 279–289. doi: 10.1128/AAC.01800-13
Shi, C., Li, H., Yang, Y., Hou, L. (2015). Anti-inflammatory and immunoregulatory functions of artemisinin and its derivatives. Mediators Inflammation 2015, 435713. doi: 10.1155/2015/435713
Sun, Z., Ma, Y., Chen, F., Wang, S., Chen, B., Shi, J., et al. (2018). Artesunate ameliorates high glucose-induced rat glomerular mesangial cell injury by suppressing the TLR4/NF-kappaB/NLRP3 inflammasome pathway. Chem. Biol. Interact. 293, 11–19. doi: 10.1016/j.cbi.2018.07.011
Sureshbabu, A., Ryter, S. W., Choi, M. E. (2015). Oxidative stress and autophagy: crucial modulators of kidney injury. Redox Biol. 4, 208–214. doi: 10.1016/j.redox.2015.01.001
Tecklenborg, J., Clayton, D., Siebert, S., Coley, S. M. (2018). The role of the immune system in kidney disease. Clin. Exp. Immunol. 192 (2), 142–150. doi: 10.1111/cei.13119
Vennerstrom, J. L., Arbe-Barnes, S., Brun, R., Charman, S. A., Chiu, F. C., Chollet, J., et al. (2004). Identification of an antimalarial synthetic trioxolane drug development candidate. Nature 430 (7002), 900–904. doi: 10.1038/nature02779
von Hagens, C., Walter-Sack, I., Goeckenjan, M., Osburg, J., Storch-Hagenlocher, B., Sertel, S., et al. (2017). Prospective open uncontrolled phase I study to define a well-tolerated dose of oral artesunate as add-on therapy in patients with metastatic breast cancer (ARTIC M33/2). Breast Cancer Res. Treat 164 (2), 359–369. doi: 10.1007/s10549-017-4261-1
Wan, R. J., Li, Y. H. (2017). Effects of Artesunate prevent nephritis via the Tolllike receptor 4/nuclear factorkappaB signaling pathway in rats. Mol. Med. Rep. 16 (5), 6389–6395. doi: 10.3892/mmr.2017.7362
Wang, H., Wang, H., Jiang, B., Zhang, H. Y., Liu, B. J., Sun, L. Y., et al. (2010). Artesunate relieves lupus nephritis by inhibiting the expression of ICAM-1. J. Clin. Med. Pract. 14 (17), 1–3. doi: 10.3969/j.issn.1672-2353.2010.17.001
Wang, H. X., Su, K., Xun, J. Q., Huang, X. J., Wang, J. G., et al. (2016). Effects of artesunate on proliferation, apoptosis and expression of TNF-α and IL-6 in rat mesangial cells. China Health Care Nutr. 26, 208.
Wen, Y., Pan, M. M., Lv, L. L., Tang, T. T., Zhou, L. T., Wang, B., et al. (2019). Artemisinin attenuates tubulointerstitial inflammation and fibrosis via the NF-kappaB/NLRP3 pathway in rats with 5/6 subtotal nephrectomy. J. Cell Biochem. 120 (3), 4291–4300. doi: 10.1002/jcb.27714
White, N. J., Hien, T. T., Nosten, F. H. (2015). A Brief History of Qinghaosu. Trends Parasitol. 31 (12), 607–610. doi: 10.1016/j.pt.2015.10.010
Wu, X., Wang, X., Zhang, W., Shi, X., An, P., Sun, W., Wang, Z., et al. (2010). Therapeutic effect of artemisinin on lupus nephritis mice and its mechanisms. Acta Biochim. Biophys. Sin. (Shanghai) 42 (12), 916–923. doi: 10.1093/abbs/gmq101
Wu, X. L., Zhang, W. G., Shi, X. M., An, P., Sun, W. S., Qiao, C. L., et al. (2010). Effects of Artemisinin on Proliferation, Apoptosis and Caspase-3 Expression of Rat Mesangial Cell. J. Chin. Med. Mater. 33 (3), 407–410.
Wu, P., Li, Q. J., Xia, Z. L., Zhang, F. Q., Xia, Q. J. (2011). Establishment of Mice Nephritis Models and Observation of the Effects of Dihydroartemisinin on the Release of Inflammatory Cytokines. West China Med. J. 26 (7), 1028–1031.
Wu, X. L., Shi, X. M., An, P., Sun, W. S., Qiao, C. L., et al. (2011). Effect of artemisinin combined with glucocorticoid on the expressions of glucocorticoid receptor alpha mRNA, glucocorticoid receptor beta mRNA and P300/CBP protein in lupus nephritis mice. Chin J. Integr. Med. 17 (4), 277–282. doi: 10.1007/s11655-011-0693-3
Wu, X., An, P., Ye, B., Shi, X., Dang, H., Fu, R., et al. (2014). Artemisinin ameliorated proteinuria in rats with adriamycin-induced nephropathy through regulating nephrin and podocin expressions. J. Tradit. Chin Med. 34 (1), 63–68. doi: 10.1016/S0254-6272(14)60056-X
Xia, M., Liu, D., Tang, X., Liu, Y., Liu, H., Liu, Y., et al. (2020). Dihydroartemisinin inhibits the proliferation of IgAN mesangial cells through the mTOR signaling pathway. Int. Immunopharmacol. 80, 106125. doi: 10.1016/j.intimp.2019.106125
Xiang, M., Chen, Z., He, L., Xiong, G., Lu, J. (2019). Transcription profiling of artemisinin-treated diabetic nephropathy rats using high-throughput sequencing. Life Sci. 219, 353–363. doi: 10.1016/j.lfs.2019.01.032
Yang, D. X., Qiu, J., Zhou, H. H., Yu, Y., Zhou, D. L., Xu, Y., et al. (2018). Dihydroartemisinin alleviates oxidative stress in bleomycin-induced pulmonary fibrosis. Life Sci. 205, 176–183. doi: 10.1016/j.lfs.2018.05.022
You, Y. W., Liao, P. H., Yang, F. F., Lin, X. (2014). Regulating effect of dihydroartemisinin on fractalkine expression in renal cortex of lupus-prone MRL/Ipr mice. Immunological J. 30 (7), 617–622. doi: 10.13431/j.cnki.immunol.j.20140135
Yu, C., Sun, P., Zhou, Y., Shen, B., Zhou, M., Wu, L., et al. (2019). Inhibition of AKT enhances the anti-cancer effects of Artemisinin in clear cell renal cell carcinoma. BioMed. Pharmacother. 118, 109383. doi: 10.1016/j.biopha.2019.109383
Zhang, L. K., Su, Y. J., Zhou, F. J., Zhang, J. Y., An, J. X. (2014a). Effect of Artemisinin on the upregulation of PDGF-B protein expression in the kidney of experimental diabetic rats. Modern J. Integrated Traditional Chin. Western Med. 23 (13), 1392–1396. doi: 10.3969/j.issn.1008-8849.2014.13.009
Zhang, L. K., Zhang, J. Y., Zhou, F. J., Su, Y. J., Zhang, J. Y, An, J. X. (2014b). Study on the mechanism of renoprotective effects of artemisinin in diabetic rats. Modern J. Integrated Traditional Chin. Western Med. 23 (26), 2862–2864. doi: 10.3969/j.issn.1008-8849.2014.26.005
Zhang, L. K., Zhang, J. Y., Zhou, F. J., Su, Y. J., An, J. X., et al. (2014c). Inhibitory effect of artemisinin on the spatiotemporal dynamics activation of protein kinase C in the kidney of experimental diabetic rats. Modern J. Integrated Traditional Chin. Western Med. 23 (18), 1964–1966.
Zhang, Y., Li, H., Zhu, J., Wei, T., Peng, Y., Li, R., et al. (2017). Role of artesunate in TGF-β1-induced renal tubular epithelial-mesenchymal transdifferentiation in NRK-52E cells. Mol. Med. Rep. 16 (6), 8891–8899. doi: 10.3892/mmr.2017.7728
Zhang, J., Cha, W. Q., Chen, X. H., Jiang, L., Jiang, Y. X. (2018). Effect of Artemisinin on Cell Proliferation and Fascin Expression of Human Renal Cell Carcinoma Cell Line 786-0. Med. Innovation China 15 (24), 29–31. doi: 10.3969/j.issn.1674-4985.2018.24.008
Zhang, B., Liu, P., Zhou, Y., Chen, Z., He, Y., Mo, M., et al. (2019). Dihydroartemisinin attenuates renal fibrosis through regulation of fibroblast proliferation and differentiation. Life Sci. 223, 29–37. doi: 10.1016/j.lfs.2019.03.020
Zhou, F. J., Su, Y. J., Zhang, L. K., Zhang, J. Y., An, J. X. (2014a). Inhibitory effect of artemisinin on the upregulation of DNA-binding activity of NF-κB in kidney tissue of diabetic rats. Modern J. Integrated Traditional Chin. Western Med. 23 (19), 2075–2079. doi: 10.3969/j.issn.1008-8849.2014.19.009
Zhou, F. J., Zhang, L. K., Su, Y. J., Zhang, J. Y., An, J. X. (2014b). Inhibitory effect of artemisinin on the upregulation of c-fos and c-jun gene expression in kidney tissue of diabetic rats. Modern J. Integrated Traditional Chin. Western Med. 23 (21), 2294–2298. doi: 10.3969/j.issn.1008-8849.2014.21.005
Zhou, F. J., Zhang, J. Y., Zhang, L. K., Su, Y. J., An, J. X. (2014c). Inhibitory effect of artemisinin on the upregulation of the DNA-binding activity of AP-1 in the kidney tissue of experimental diabetic rats. Modern J. Integrated Traditional Chin. Western Med. 23 (24), 2648–2652. doi: 10.3969/j.issn.1008-8849.2014.24.008
Zhou, Z., Lu, B., Wang, C., Wang, Z., Luo, T., Piao, M., et al. (2017). RIP1 and RIP3 contribute to shikonin-induced DNA double-strand breaks in glioma cells via increase of intracellular reactive oxygen species. Cancer Lett. 390, 77–90. doi: 10.1016/j.canlet.2017.01.004
Keywords: artemisinin derivatives (artemisinins), kidney disease, inflammation, immunity, oxidative stress
Citation: Xia M, Liu D, Liu Y and Liu H (2020) The Therapeutic Effect of Artemisinin and Its Derivatives in Kidney Disease. Front. Pharmacol. 11:380. doi: 10.3389/fphar.2020.00380
Received: 19 September 2019; Accepted: 12 March 2020;
Published: 31 March 2020.
Edited by:
Norberto Perico, Istituto Di Ricerche Farmacologiche Mario Negri, ItalyReviewed by:
Ilse Sofia Daehn, Icahn School of Medicine at Mount Sinai, United StatesBente Jespersen, Aarhus University Hospital, Denmark
Copyright © 2020 Xia, Liu, Liu and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hong Liu, bGl1aG9uZzYxOEBjc3UuZWR1LmNu