 Linda Elowsson Rendin*
Linda Elowsson Rendin* Anna Löfdahl
Anna Löfdahl Måns KadeforsZackarias Söderlund
Måns KadeforsZackarias Söderlund Emil Tykesson
Emil Tykesson Sara Rolandsson EnesJenny Wigén
Sara Rolandsson EnesJenny Wigén Gunilla Westergren-Thorsson
Gunilla Westergren-Thorsson- Lung Biology Unit, Department of Experimental Medical Science, Lund University, Lund, Sweden
It is known that the cell environment such as biomechanical properties and extracellular matrix (ECM) composition dictate cell behaviour including migration, proliferation, and differentiation. Important constituents of the microenvironment, including ECM molecules such as proteoglycans and glycosaminoglycans (GAGs), determine events in both embryogenesis and repair of the adult lung. Mesenchymal stromal/stem cells (MSC) have been shown to have immunomodulatory properties and may be potent actors regulating tissue remodelling and regenerative cell responses upon lung injury. Using MSC in cell-based therapy holds promise for treatment of chronic lung diseases such as idiopathic pulmonary fibrosis (IPF) and chronic obstructive pulmonary disease (COPD). However, so far clinical trials with MSCs in COPD have not had a significant impact on disease amelioration nor on IPF, where low cell survival rate and pulmonary retention time are major hurdles to overcome. Research shows that the microenvironment has a profound impact on transplanted MSCs. In our studies on acellular lung tissue slices (lung scaffolds) from IPF patients versus healthy individuals, we see a profound effect on cellular activity, where healthy cells cultured in diseased lung scaffolds adapt and produce proteins further promoting a diseased environment, whereas cells on healthy scaffolds sustain a healthy proteomic profile. Therefore, modulating the environmental context for cell-based therapy may be a potent way to improve treatment using MSCs. In this review, we will describe the importance of the microenvironment for cell-based therapy in chronic lung diseases, how MSC-ECM interactions can affect therapeutic output and describe current progress in the field of cell-based therapy.
Introduction
Chronic lung diseases such as idiopathic pulmonary fibrosis (IPF) and chronic obstructive pulmonary disease (COPD) are life-threatening progressive lower respiratory diseases that are increasing world-wide. COPD alone is the 3rd cause of death, and complicated by co-morbidities such as cardiovascular disease and lung cancer (Sin et al., 2006). COPD patients are mainly treated with bronchodilators and inhaled corticosteroids to help reduce symptoms, however, disease progression is not halted (Yawn et al., 2021). The complex pathology exhibits a diverse spectrum of phenotypes with formation of fibrosis and/or emphysema and current treatment options involve only strategies to slow down the disease progression, and to improve quality of life (Negewo et al., 2015; Barrecheguren and Miravitlles, 2016). IPF, affecting 3 million people world-wide (Martinez et al., 2017) is also associated with cardiovascular comorbidities (Caminati et al., 2021) and share the risk factors of smoking and inhaled toxins with COPD. IPF, being a chronic fibrosing interstitial lung disease (ILD) with unknown etiology, is characterized by the histopathological pattern of usual interstitial pneumonia (UIP). The abnormal wound healing in response to epithelial distress results in exaggerated mesenchymal cell activities including build-up and turnover of ECM, causing the altered biomechanics and increased stiffness of the lung tissue. This ultimately disrupts gas exchange and lung function (Raghu et al., 2011). No treatment today affects the mortality associated with IPF to a great extent, but merely prolongs life, with patients still rarely exceeding 3–5 years after IPF diagnosis (Raghu et al., 2011). IPF is primarily treated with the antifibrotic drugs Nintedanib or Pirfenidone, which generate prolonged life-span, but treatment adherence is unfortunately low due to multiple side effects. Future treatment strategies include combining drugs to manage several activated cellular pathways that may differ between patients (Gao et al., 2021). Therefore stratifying patients in more detail may uncover novel therapeutic targets, which is reviewed in Trachalaki et al. (Trachalaki et al., 2021). The lack of curative treatments for both IPF and COPD therefore opts for new efficient strategies that not only affect disease severity but also inhibit progression or at best reverse the deterioration entirely and healing the lung. This is a priority for individual patients as well as for society (Hilberg et al., 2018), and the socioeconomic costs due to these diseases could be significantly reduced by minimizing prevalence and severity (Anees Ur and et al., 2020).
Common to both diseases is the structural remodeling of the lung tissue, resulting in impaired gas exchange that leads to decline in lung function. To date, the knowledge of the inherent capacity of the lung to repair is limited, but mesenchymal stromal cells (MSC) have been proposed to have a therapeutic potential due to their regenerative and immunomodulatory abilities in combination with their low immunogenicity attributed to low or absent human leukocyte antigen (HLA) expression (Lee et al., 2014). The structurally remodeled ECM may have a larger impact on cellular behavior in the local microenvironment than previously anticipated. An altered ECM actively affects the biomechanical properties of the lung tissue (Elowsson Rendin et al., 2019) and its function, creating an inherent vicious feedback loop that further propagate these diseases (Figure 1). Therefore, approaching the microenvironment of the lung to control local pathological cell responses that may also enhance therapeutic activities of local MSC, is an appealing therapeutic thought that can revolutionize treatment of serious chronic lung diseases such as IPF and COPD. This review elaborates on ways to modulate the pulmonary microenvironment of the therapeutic MSCs to improve and control efficacy of MSC treatment. This could mean providing an injectable cell-instructive artificial milieu (scaffold), or by directly affecting in situ environment of the injected MSC.
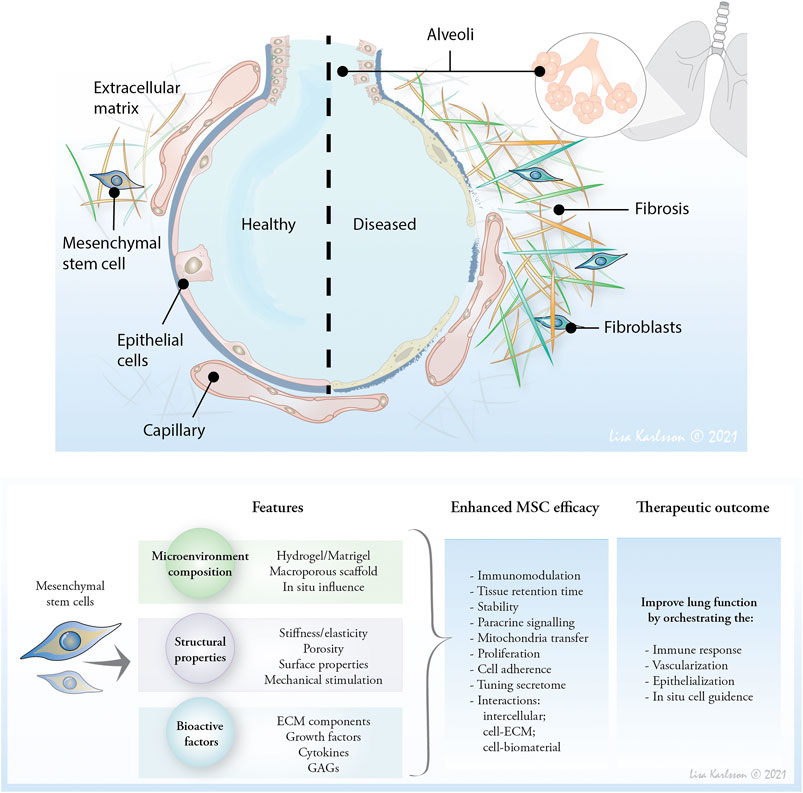
FIGURE 1. The composition and mechanoelastic properties of ECM affect cell response and activity and therefore, moderating the microenvironment of therapeutic mesenchymal stem cells (MSC) may provide a novel approach to affect cell retention time, and improve therapeutic properties. Affecting the ECM in situ or providing an artificial microenvironment to enhance MSC therapeutic potential are future lines of research to affect the distorted lung tissue and reduce lung inflammation, and optimally heal emphysematic and fibrotic lesions to restore oxygen uptake.
MSC Based Therapy in IPF and COPD
MSCbased therapy has been explored in clinical trials in both IPF and COPD using intravenous (i.v.) or intratracheal routes of administration of bone marrow, umbilical cord or adipose tissue derived MSC in a dosage range of 106–108 cells per dose, single dose or in some cases, up to 4 doses. Completed trials using primarily one dosage regimen show in COPD trials a safe profile and ameliorated clinical parameters in up to 50% of patients (Cruz and Rocco, 2020). In a small human trial in IPF, patients demonstrated that allogeneic MSC treatment is safe and tolerable upon applying a single injected dose of bone marrow derived MSC up to 200 x 106 cells (Glassberg et al., 2017). Similarly, low adverse effects were also shown when using placenta derived MSC (i.v. infusion) or adipose derived stromal cells from stromal vascular fraction (endobronchial infusion) in a phase 1b study in IPF patients (Tzouvelekis et al., 2013; Chambers et al., 2014). A recent study examined the efficacy of repeated i. v. administered MSC, showing improved lung function and 6°min walk test in patients with moderate to severe IPF(Averyanov et al., 2020). Collectively, pre-clinical studies demonstrate antifibrotic effects with MSC treatment, however, the long-term beneficial effects and feasibility of MSC as a therapeutic strategy in IPF patients are yet to be determined, where the few existing clinical trials demonstrate an acceptable safety profile. According to ClinicalTrials.gov, there are currently 5 clinical trials recruiting for stem cell treatment of pulmonary fibrosis. Of these, one is carried out at the University of North Carolina, Chapel Hill, United States, with the aim to evaluate safety and efficacy of infusing autologous lung spheroid stem cells derived from trans-bronchial biopsy specimens in IPF patients.
Several animal studies show promising results in treating pulmonary fibrosis with MSC or derivates thereof such as exosomes and extracellular vesicles (EVs), alleviating fibroblast activation and ECM deposition along with anti-inflammatory outcomes (Reddy et al., 2016; Mansouri et al., 2019; Gad et al., 2020; Wan et al., 2020). Pre-clinical studies on COPD demonstrate that MSCs reduce inflammation and improve histological lung structure (Cruz and Rocco, 2020), which have been translated into several human clinical trials for treatment of COPD(Ribeiro-Paes et al., 2011; Weiss et al., 2013; Stolk et al., 2016; de Oliveira et al., 2017; Armitage et al., 2018). These clinical trials all had safety as their primary endpoint and was not powered for detection of efficacy. Although the clinical trials failed to reproduce the promising pre-clinical results, they all demonstrated that MSC administration was well tolerated among COPD and emphysema patients (Ribeiro-Paes et al., 2011; Weiss et al., 2013; Stolk et al., 2016; de Oliveira et al., 2017; Armitage et al., 2018). Interestingly, Armitage et al. demonstrated that MSCs radiolabeled with indium-111 reached the lungs within 30 min and remained detectable for 24 h after administration (Armitage et al., 2018). This should mean that the effect of MSCs on damaged lung tissue and resident cells need to be rapid. Moreover, an increasing body of research suggests that the MSC therapeutic functions depend on the microenvironment encountered (Kusuma et al., 2017; Abreu et al., 2019; Islam et al., 2019; Abreu et al., 2020). Therefore, it is tempting to speculate that by modifying the microenvironment it would be possible to enhance the therapeutic time-frame for MSCs by increasing residency in the COPD lungs and thereby increasing the potency of the infused MSCs. Currently, there are three clinical trials recruiting patients to evaluate MSC for the treatment of COPD. A comprehensive overview of ongoing and completed human trials in IPF and COPD is searchable at ClinicalTrials.gov. These trials are encouraging and gives great hope to the scientific community to increase the therapeutic potential of MSC.
The Mechanism of MSC
Anti-Inflammatory Function
MSCs are extensively explored as cell therapy candidates and are known for their potential to differentiate into multiple cell lineages, however their major therapeutic mechanism is from interactions via paracrine signaling and cell-cell contact (Murphy et al., 2013; Gao et al., 2016). MSCs have demonstrated both anti-inflammatory and anti-fibrotic capacity, which could be of importance for tissue repair and regeneration in chronic lung diseases (Cruz and Rocco, 2020). The observed anti-inflammatory properties of MSCs include effects on several cells of the immune system, including T-cells, B-cells, dendritic cells, monocytes, macrophages, and natural killer (NK) cells. For example, MSCs can suppress T-cell proliferation via Indoleamine-2,3dioxygenase (IDO) secretion, which mediates tryptophan depletion causing cellular stress in T cells (Meisel et al., 2004; Laing et al., 2019), or by inducing a switch toward a regulatory T-cell phenotype (Bartholomew et al., 2002; Benvenuto et al., 2007; Casiraghi et al., 2008). In an inflammatory setting, MSCs are able to direct macrophage polarization from a proinflammatory (M1) to an anti-inflammatory phenotype (M2). This occurs through feedback-loop involving MSC secreted prostaglandin E2 (PGE2) (reviewed in) (Prockop, 2013). In addition, MSCs are able to affect resident macrophages to reduce their secretion of pro-inflammatory mediators through a different negative feedback-loop, by secreting tumor necrosis factor (TNF)-α stimulated gene 6 (TSG-6) (Prockop, 2013).
While the anti-inflammatory effect of MSCs has been extensively used as a rationale for MSC-based therapy, their anti-fibrotic qualities could further motivate the use of MSC-based therapy in chronic diseases. In animal models of lung fibrosis, it has been shown that MSC administration can reduce the extent of fibrosis (reviewed in (Srour and Thébaud, 2015). Increased levels of the hepatocyte growth factor (HGF) has been linked to the anti-fibrotic properties of MSCs. In vitro, HGF inhibited epithelial-mesenchymal-transition (EMT) through inhibition of transforming growth factor β (TGF-β) signaling (Mias et al., 2009; Shukla et al., 2009). Interestingly, administration of MSC has been associated with changes in levels of ECM modifying enzymes such as metalloproteases (MMPs) and tissue inhibitor of metalloproteases (TIMPs), and the balance of these enzymes is important in both fibrosis and emphysema development (Mias et al., 2009; Moodley et al., 2009; Semedo et al., 2009).
Operating Neighboring Cells: Extracellular Vesicles
In addition to the secretion of anti-inflammatory and anti-fibrotic mediators, MSCs are also known to communicate with other cells by secretion of EVs (reviewed in (Farhat et al., 2019)). EVs are a heterogenous group of small lipid vesicles with cargoes containing mediators for intercellular communication such as proteins, microRNAs, and lipids (Thery and et al., 2018; van Niel et al., 2018). An increasing body of literature suggests that MSC-derived EVs possess immune regulatory functions (Farhat et al., 2019). MSC-derived EVs have been demonstrated to mitigate allergic hyperresponsiveness (Cruz et al., 2015), enhance M1 to M2 macrophage phenotype polarization, macrophage bioenergetics, and phagocytosis (Phinney et al., 2015; Jackson et al., 2016; Morrison et al., 2017), as well as to have an inhibitory effect on T-cell proliferation (Blazquez et al., 2014; Pachler et al., 2017; Mardpour et al., 2019). Moreover, MSC-derived EVs have been shown to decrease the influx of inflammatory cells in an E. coli endotoxin-induced acute lung injury model. This therapeutic effect was at least partly mediated through keratinocyte growth factor (KGF/FGF7)(Zhu et al., 2014). Interestingly, in an E. coli-induced acute lung injury rat model, it was demonstrated that EVs isolated from MSCs pre-exposed to IFN-g were more effective in attenuating lung injury compared to EVs isolated from naïve, non-exposed MSCs (Varkouhi et al., 2019). These data suggest that the EV cargo released by MSCs can vary significantly based on the microenvironment the cells have encountered.
Restore Cellular Function: Mitochondrial Transfer
Another possibility of MSCs potential in cell therapy is to use their mitochondria. The first evidence that MSCs have the capacity to transfer mitochondria to other cells was published in 2006 (Spees et al., 2006). In this study MSCs were able to restore the aerobic respiration in epithelial cells lacking functional mitochondria (Spees et al., 2006). Since then, it has been shown that MSCs can transfer mitochondria via EVs, microtubules, and tunneling nanotubes (TNTs) (Spees et al., 2006; Phinney et al., 2015; Morrison et al., 2017). For example, mitochondria transferred from MSCs to macrophages partly via TNTs, modulated macrophage phagocytosis both in vitro and in vivo (Jackson et al., 2016). MSC mitochondrial transfer has also been reported to reduce lung injury in a cigarette smoke-induced emphysema model (Li et al., 2014). Furthermore, Islam et al. reported that MSCs could protect against acute lung injury through mitochondrial transfer via Cx43-dependent alveolar attachment (Islam et al., 2012). Interestingly, Mahrouf-Yorgov et al. observed that MSCs had the capacity to sense, and subsequently engulf and degrade mitochondria from damaged cells. This process enhanced mitochondrial biogenesis and mitochondrial transfer by the MSCs (Mahrouf-Yorgov et al., 2017). Taken together, future directives using MSC based therapy in chronic lung diseases needs to consider the engraftment in the lung for therapeutic efficacy (Behnke et al., 2020).
The Intrinsic Properties of the Lung-ECM
MSCs and other mesenchymal cells are the main producer of ECM. Interestingly, the role of the ECM is receiving increasing recognition as an important element in controlling cellular behavior during homeostasis and in disease progression (Deng et al., 2020). Apart from providing mechanical and structural support, the complex 3D structure of the ECM, consisting of glycoproteins, collagens, glycosaminoglycans (GAGs), proteoglycans, dictate cell migration, polarity, proliferation, differentiation, and survival, where the dynamics of the ECM molecules play a critical part (Wigén et al., 2019). Some proteins e.g., elastin fibers are stable over almost a lifetime (Shapiro et al., 1991), while certain collagens have a short turnaround, changing every day (Laurent, 1987). In the pulmonary interstitial layer, the major structural components are fibrillar collagens type I, III, IV, V, and XI (extensively reviewed by Laurent, 1987; Sherman 2015) that make up the core structure and contribute to the tensile strength, and elastin along with fibrillin contribute to the compliance of the tissue (Laurent, 1987; Sherman et al., 2015). This intricate meshwork of fibers is also composed of glycoproteins, GAGs, and proteoglycans providing the tissue with its viscoelastic properties. In the alveolar space, the epithelial and endothelial cells are anchored to a basement membrane primarily consisting of collagen type IV, laminins, and proteoglycans. Both in IPF and COPD the basement membrane and the interstitial layer are affected with altered structural properties affecting the biomechanics. In IPF, the alveolar basement membrane is fragmented and the interstitial layer is thickened, while in COPD, the ECM components are degraded and the structural properties are lost (Burgess et al., 2016)
Glycosaminoglycans and Growth Factors
While the most studied group of ECM molecules are the various collagens, lately however the family of proteoglycans have drawn large attention. Proteoglycans are proteins decorated with covalently attached linear polysaccharides, GAGs. The proteoglycans are heterogeneous molecules-both in terms of the core protein structure, as well as the chain length and chemical structure of the GAG side chains. These side chains strongly decide the physical properties of the proteoglycans. The main classes of proteoglycan-bound GAGs in the human lung are heparan sulfate/heparin (HS/hep) chondroitin/dermatan sulfate (CS/DS) and keratan sulfate (KS) (Papakonstantinou and Karakiulakis, 2009). A fourth type of GAG, hyaluronan (HA), is also prevalent in the lung, but is not bound to a core protein (Couchman and Pataki, 2012). All four types of GAGs contain disaccharide motifs, which are repeated throughout the polymer. For HS/hep, CS/DS and KS the disaccharide units can be modified by sulfation at various positions, giving rise to motifs/domains (Persson et al., 2020), which in turn are important for binding of specific mediators to be involved in chronic lung diseases such as TGF-b, HGF, and TNF-a (Esko et al., 2015). In this way the GAGs play an important role in interacting with growth factors, where the GAGs build gradients during embryonic development but also during tissue repair and delivering of growth factors to receptors. Interestingly, alterations in sulfation pattern, or induction of deacetylations and epimerizations in the GAG chains affect their role in tuning growth factor gradients, which alter cell activities (Wigén et al., 2019). In IPF, an increase in sulfation of both HS and CS/DS has been recognized in the border zone toward fibrotic lesions (Westergren-Thorsson et al., 2017). This may reflect an altered pulmonary landscape, navigating nearby cells toward regions of activated repair process, promoting a profibrotic phenotype.
ECM Bound Growth Factors
Arguably one of the most important interactions between cells is mediated through growth factors and cytokines. These factors travel large distances in the body to recruit and direct cells where they are needed. A largely overlooked process is how growth factors interact with the ECM via GAGs and how the ECM can increase or decrease the cell response to growth factors. Thus, the ECM serves as a reservoir for growth factors, which are slowly released over time. At the same time the growth factors bound to the ECM via the GAGs become protected from circulating proteases. Because of these properties, if the ECM is injured the release of growth factors will be instant, thus triggering an immediate cell response. For example, latent TGF-β (Saito et al., 2018) in the ECM become activated upon injury and is known to be one of the factors activating mesenchymal cells that in fibrosing diseases such as IPF lead to an over production of ECM. Another example of ECM bound factor is fibroblast growth factor (FGF)-2, which binds on a nanomolar level to HS (Ibrahimi et al., 2004). This GAG-growth factor complex increases the binding to the FGF receptor 1 and is crucial for adequate cell signaling (Yayon et al., 1991) and may be immunomodulatory factor in COPD (Tan et al., 2020). Therefore, the ECM and growth factor complexes works to fine-tune cell responses and control timing, making it an important target in the development of therapies in chronic lung diseases.
Cell-ECM Interaction
Even though lung cells derived from patients with chronic lung disease have shown to have inherent cellular features such as gene expression and proteomic profile (Hallgren et al., 2010; Woldhuis et al., 2020), the microenvironment of the tissue, however, appears to be the dominating factor in regulating cellular behavior, at least in vitro (Elowsson Rendin et al., 2019). It has been demonstrated that the pathological reconstructed ECM in IPF overrules intrinsic cellular characterizations, ultimately repr ogramming cells for fibrogenesis (Parker et al., 2014; Philp et al., 2018; Rodriguez et al., 2018). The wavy and aligned collagen structures of the ECM in distal IPF tissue alter migration patterns and cell morphology, creating elongated human lung fibroblasts with increased migratory speed (Tisler et al., 2020). Lung fibroblasts engrafted on fibrotic or emphysematous lung tissue readily adapt to surrounding structures and ECM niches of the lung with high morphological plasticity (Burgstaller et al., 2018). Non-cultured fibroblasts, isolated from lungs of IPF patients, demonstrated an altered genomic profile in comparison to healthy fibroblasts (Emblom-Callahan et al., 2010), as do epithelial cells when cultured on bronchial-ECM derived from COPD (Hedström et al., 2018). The positive feedback-loop created between cells and ECM is manifested quickly with marked differences in cellular activity, inducing a shift in the production of ECM proteins that steers the cellular response toward a continuous rebuild of fibrotic tissue with altered basement membrane structures (Elowsson Rendin et al., 2019). The constant changes in ECM composition during disease progression, resulting in pathological differences in lung architecture creates a disease specific proteomic change where the local microenvironment signals for continuous tissue reconstruction (Westergren-Thorsson et al., 2017; Åhrman et al., 2018).
The deformed lung tissue, caused by either excessive tissue repair or the lack thereof, also has an immense impact on the mechanical properties of the tissue, where mechanical forces sensed by surrounding cells activate mechanosensitive pathways (e.g., YAP/TAZ) and influence cell behavior (Asano et al., 2017; Haak et al., 2018). To maintain tissue homeostasis, the cells require anchoring to the ECM via integrin binding through which the cells sense the microenvironment and activate signaling transduction pathways, inducing mechanical and chemical intracellular signaling through the actin cytoskeleton (reviewed by Vogel, 2018 (Vogel, 2018)). Changes in the local mechanical properties create a mechanical gradient that affects cellular behavior such as migration, proliferation and differentiation. Integrins are also thought to be involved in growth factor signaling, acting synergistically with receptors for different growth factors. More specifically, the integrins are known to bind to Arg-Gly-Asp (RGD) motifs found predominantly on fibronectin (FN), but also in ECM proteins e.g., laminin, tenascin, and vitronectin (Widhe et al., 2016; Miron-Mendoza et al., 2017). Integrins are also the main receptors forming focal adhesions and are thought to be required for cell motility. The local microenvironment has been found to be decisive for the size and concentration of focal adhesions, which further impact on cell behavior (Wozniak et al., 2004; Costa et al., 2013).
Modulating the 3D Environment for Treatment
There are several in vitro studies showing how MSC are differently affected by the environment the cells encounter. Increasing research is focused on how to control the therapeutic effect of MSC, where one option being explored is by modulating the microenviroment in situ or by injectable biomaterials.
Hydrogel Encapsulation
Encapsulating MSC in injectable hydrogels is extensively explored to increase retention time and to modulate the cellular activity in situ (Farhat et al., 2019; Wong et al., 2020) by providing the cells with a 3D microenvironment that protects and promotes an anti-inflammatory and regenerative response, see outline in Figure 1. Techniques range from single cell encapsulation to larger scaffolds with macroporous structure (Kim et al., 2019). Efforts have been made to mimic the mechanical properties of the ECM to steer cellular behavior in health and disease (de Hilster et al., 2020; Pouliot et al., 2020). So far, hydrogels have been by far the most produced and studied substrate, where ECM-based hydrogels from murine, porcine and human tissues have been developed with different decellularization methods. In order to mimic the natural tissue when designing an artificial matrix, it is vital to maintain the components of the ECM, often measured as the GAG-DNA content ratio, as well as the viscoelastic properties (Uhl et al., 2020). Apart from hydrogels based on decellularized tissue, there are various natural, synthetic and hybrid (meaning combining natural, synthetic or ECM components) polymers that are being explored, including proteins, polysaccharides, recombinantly expressed peptides, and polyethylene glycol (Uhl et al., 2020). The advantage of hydrogels is that the structural properties can be easily modified through the crosslinking of the hydrophilic polymer chains. In addition, the biomaterial can be modulated by incorporating cell-instructive factors e.g. growth factors, RGD domains, and ECM proteins. Recombinantly produced biomaterials such as the elastin-like recombinamer polymer allows for the precise composition of structural and chemical properties, with gene technology introducing sequences for cell attachment and MMP degradation and at the same time exposing functional groups for binding of for example growth factors to promote or maintain the MSC therapeutic effect (Ibáñez-Fonseca et al., 2020). De Santis et al. recently demonstrated the functional outcome of a 3D bioprinted hybrid hydrogel combining decellularized lung tissue with alginate to form human airways containing primary human airway epithelial progenitor cells (De Santis et al., 2020). Transplanted in mice, the cells showed evidence of differentiation into mature epithelial cells. The hydrogel properties both had matched biomechanical properties and contained ECM instructive factors from the native lung tissue (De Santis et al., 2020). Hydrogels based on collagen type I, one of the major structural components of tissues, has been extensively explored in combination with other materials such as silk, a highly elastic protein, to improve the mechanical properties to mimic the viscoelastic properties of tissues (Sanz-Fraile et al., 2020). As discussed above, GAGs multiple functions in cell signaling and in tissue homeostasis and have been explored as biomaterials. CS that have modifiable functional groups for covalent and non-covalent bonding has been used for the development of injectable hydrogels to tune for different physical properties as well as release of therapeutic factors (Ornell et al., 2019).
Macroporous Scaffolds
In contrast to solid hydrogels, macroporous biomaterials or scaffolds introduces a more complex 3D environment, where the surface properties of the scaffold can be tuned to functionalize cell attachment, differentiation and migration, as well as to enhance cell diffusion to the scaffold.
In a recently published article, a collagen-based scaffold was developed with macroporous structures to mimic the distal lung tissue. The scaffold was functionalized with collagen binding HGF to enhance the regeneration of alveolar-like structures (Wang et al., 2020). When making a porous scaffold, pore size is often determined by changing the initial concentration of the reagents, which in turn changes the bulk stiffness. By adding an antifreezing agent (e.g., DMSO), pore size can be kept constant regardless of the initial concentration (Jiang et al., 2019). Jiang et al. could show that larger and stiffer pores induced an anti-inflammatory phenotype of macrophages (Jiang et al., 2019). Shamskhou et al. has shown that local delivery of IL-10 using a hyaluronan and heparin-based hydrogel system had an anti-inflammatory effect in bleomycin-treated mice (Shamskhou et al., 2019).
Pre-Conditioning of MSC to Enhance Therapeutic Effect
There are increasing evidence that MSC have a cellular memory, with sustained responses to preceding stimuli, which may influence the desired effect of MSC-based therapies (Yang et al., 2014). MSC cultured on substrates with different ECM molecules have been shown to affect the paracrine function of MSC. For example, De Lisio et al. showed that cell cultures on collagen reduced the gene expression of inflammatory factors in MSC compared to culturing on laminin (De Lisio et al., 2014). Preconditioning of the MSC prior to administration has also been investigated, where low doses of toxic or lethal factors or exposures to hypoxic or nutrient deprived environment triggered a beneficial effect of the cells with increased expression of immunomodulatory, anti-inflammatory and repair factors (Silva et al., 2018; Nonaka et al., 2020). In a recent study, lung-derived MSCs were biophysically preconditioned to modulate their paracrine signaling by culturing the cells in a natural microenvironment, on lung scaffolds, and subjecting them to cyclic stretch prior to being injected into a rat model for acute respiratory distress syndrome (ARDS). Compared to non-preconditioned MSCs, rats treated with biophysically preconditioned MSCs had an improvement in lung elastance and reduced amounts of inflammatory cytokines in the bronchoalveolar lavage fluid (Nonaka et al., 2020). In a recent study biophysical and biochemical cues were combined to explore the immunological effect of MSC. Bone marrow-derived MSC were encapsulated in alginate-based hydrogels with different stiffnesses and pre-cultured prior to exposure to the inflammatory cytokine TNF-α. MSC preconditioned in the softer hydrogel in combination with TNFα had an improved effect in controlling monocyte turnover (Wong et al., 2020). These studies demonstrate how intrinsic cues of the microenvironment impact cellular behavior that can be harnessed in improving the therapeutic potential of MSCs.
Future Directions
For MSC therapy to become an efficient alternative to current medicine in IPF and COPD, it is essential to ensure the functional outcome of the cells in the diseased tissue. One way is to control the microenvironment in which the cells are delivered. Recent advances have demonstrated that MSC actively interact and modify their pericellular environment, that there is an active interplay between the surrounding properties, both in terms of structural properties and chemical composition, thus will play a role in directing MSC function (Ferreira et al., 2018; Loebel et al., 2019). Several obstacles remain with current MSC therapy. Ensure long-term retention of the MSCs in the lung is one, although they do get trapped in pulmonary capillaries after systemic administration, they are considered cleared within 24 h (Wang et al., 2009). MSCs are also vulnerable to the inflammatory microenvironment, which affect their restorative potential (Silva et al., 2018). Therefore, we foresee a future, where we are able to increase the retention time in the organ and control the therapeutic effect of MSCs by providing a microenvironment that supports these properties. Research into the field of artificial scaffolds for medicinal purposes that can retain a particular therapeutic property is warranted to push forward this line of novel therapies for chronic lung diseases. Modifying the microenvironment in situ may hold difficulties in its approach in retaining tuned levels of cell mediators or in adapting specific ECM components in the location of transplanted MSC. Identifying key elements or biological properties of the ECM warrants further investigation as this would allow for more precise and directed manipulation of the pathological tissue in situ. The environmental responsiveness of the MSCs creates both challenges and opportunities in applying transplanted MSC as a cell-based therapy in lung diseases that are marked with massive structural and compositional tissue remodeling. Thus, pre-conditioning of MSC may hold a more transient approach in overcoming the powerful influential impact of surrounding tissue on cellular behavior, providing a more instant regenerative response.
The artificial scaffolds hold great promise in future therapeutic purposes as it can combine cell-instructive cues together with a supporting microenvironment, delicately designed in both structure and biomechanics, that together yield an optimal curative setting for MSC. The purpose of a controlled microenvironment is to create an interim support as well as in situ cell guidance over time as the scaffold is degraded and replaced with functional lung tissue. By providing structural support, we anticipate that both the MSC retention time will be greatly improved with remained activity and allowing endogenous cells to repopulate lost tissue area.
Altogether, harnessing the microenvironment holds promise to ameliorate future cell therapeutic regimens to be able to induce lung regeneration and healing of lung tissue in chronic lung diseases marked with massive structural remodeling like IPF and COPD.
Author Contributions
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
Funding
This work was funded by Swedish Research Council in Medicine and Health (Grant number: 2016-01190), the Swedish Heart-Lung Foundation (20190297), the Royal Physiographical Society in Lund (20191114), the Medical Faculty of Lund University, the Evy and Gunnar Sandberg Foundation, Greta and John Kock Foundation, the Alfred Österlund Foundation, the Crafoord Foundation, the Consul Thure Bergh Foundation, ALF Grants Region Skåne and the Swedish Foundation for Strategic Research.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abreu, S. C., Hampton, T. H., Hoffman, E., Dearborn, J., Ashare, A., Singh Sidhu, K., et al. (2020). Differential effects of the cystic fibrosis lung inflammatory environment on mesenchymal stromal cells. Am. J. Physiology-Lung Cell Mol. Physiol. 319, L908–L925. doi:10.1152/ajplung.00218.2020
Abreu, S. C., Rolandsson Enes, S., Dearborn, J., Goodwin, M., Coffey, A., Borg, Z. D., et al. (2019). Lung inflammatory environments differentially alter mesenchymal stromal cell behavior. Am. J. Physiology-Lung Cell Mol. Physiol. 317, L823–L831. doi:10.1152/ajplung.00263.2019
Åhrman, E., Hallgren, O., Malmström, L., Hedström, U., Malmström, A., Bjermer, L., et al. (2018). Quantitative proteomic characterization of the lung extracellular matrix in chronic obstructive pulmonary disease and idiopathic pulmonary fibrosis. J. Proteomics 189, 23. doi:10.1016/j.jprot.2018.02.027
Anees Ur, R., Ahmad Hassali, M. A., Muhammad, S. A., Shah, S., Abbas, S., Hyder Ali, I. A. B., et al. (2020). The economic burden of chronic obstructive pulmonary disease (COPD) in the USA, Europe, and Asia: results from a systematic review of the literature. Expert Rev. Pharmacoecon Outcomes Res. 20, 661–672. doi:10.1080/14737167.2020.1678385
Armitage, J., Tan, D. B. A., Troedson, R., Young, P., Lam, K.-v., Shaw, K., et al. (2018). Mesenchymal stromal cell infusion modulates systemic immunological responses in stable COPD patients: a phase I pilot study. Eur. Respir. J. 51, 1702369. doi:10.1183/13993003.02369-2017
Asano, S., Ito, S., Takahashi, K., Furuya, K., Kondo, M., Sokabe, M., et al. (2017). Matrix stiffness regulates migration of human lung fibroblasts. Physiol. Rep. 5, e13281. doi:10.14814/phy2.13281
Averyanov, A., Koroleva, I., Konoplyannikov, M., Revkova, V., Lesnyak, V., Kalsin, V., et al. (2020). First‐in‐human high‐cumulative‐dose stem cell therapy in idiopathic pulmonary fibrosis with rapid lung function decline. Stem Cell Transl Med 9, 6–16. doi:10.1002/sctm.19-0037
Barrecheguren, M., and Miravitlles, M. (2016). COPD heterogeneity: implications for management. Multidiscip Respir. Med. 11, 14. doi:10.1186/s40248-016-0053-4
Bartholomew, A., Sturgeon, C., Siatskas, M., Ferrer, K., McIntosh, K., Patil, S., et al. (2002). Mesenchymal stem cells suppress lymphocyte proliferation in vitro and prolong skin graft survival in vivo. Exp. Hematol. 30, 42–48. doi:10.1016/s0301-472x(01)00769-x
Behnke, J., Kremer, S., Shahzad, T., Chao, C.-M., Böttcher-Friebertshäuser, E., Morty, R. E., et al. (2020). MSC based therapies-new perspectives for the injured lung. Jcm 9, 682. doi:10.3390/jcm9030682
Benvenuto, F., Ferrari, S., Gerdoni, E., Gualandi, F., Frassoni, F., Pistoia, V., et al. (2007). Human mesenchymal stem cells promote survival of T cells in a quiescent state. Stem Cells 25, 1753–1760. doi:10.1634/stemcells.2007-0068
Blazquez, R., Sanchez-Margallo, F. M., de la Rosa, O., Dalemans, W., Ã lvarez, V., Tarazona, R., et al. (2014). Immunomodulatory potential of human adipose mesenchymal stem cells derived exosomes on in vitro stimulated T cells. Front. Immunol. 5, 556. doi:10.3389/fimmu.2014.00556
Burgess, J. K., Mauad, T., Tjin, G., Karlsson, J. C., and Westergren‐Thorsson, G. (2016). The extracellular matrix - the under‐recognized element in lung disease?. J. Pathol. 240, 397–409. doi:10.1002/path.4808
Burgstaller, G., Sengupta, A., Vierkotten, S., Preissler, G., Lindner, M., Behr, J., et al. (2018). Distinct niches within the extracellular matrix dictate fibroblast function in (cell free) 3D lung tissue cultures. Am. J. Physiology-Lung Cell Mol. Physiol. 314, L708–L723. doi:10.1152/ajplung.00408.2017
Caminati, A., Madotto, F., Conti, S., Cesana, G., Mantovani, L., and Harari, S. (2021). The natural history of idiopathic pulmonary fibrosis in a large European population: the role of age, sex and comorbidities. Intern. Emerg. Med. doi:10.1007/s11739-021-02651-w
Casiraghi, F., Azzollini, N., Cassis, P., Imberti, B., Morigi, M., Cugini, D., et al. (2008). Pretransplant infusion of mesenchymal stem cells prolongs the survival of a semiallogeneic heart transplant through the generation of regulatory T cells. J. Immunol. 181, 3933–3946. doi:10.4049/jimmunol.181.6.3933
Chambers, D. C., Enever, D., Ilic, N., Sparks, L., Whitelaw, K., Ayres, J., et al. (2014). A phase 1b study of placenta-derived mesenchymal stromal cells in patients with idiopathic pulmonary fibrosis. Respirology 19, 1013–1018. doi:10.1111/resp.12343
Costa, P., Scales, T. M. E., Ivaska, J., and Parsons, M. (2013). Integrin-specific control of focal adhesion kinase and RhoA regulates membrane protrusion and invasion. PLoS One 8, e74659. doi:10.1371/journal.pone.0074659
Couchman, J. R., and Pataki, C. A. (2012). An introduction to proteoglycans and their localization. J. Histochem. Cytochem. 60, 885–897. doi:10.1369/0022155412464638
Cruz, F. F., Borg, Z. D., Goodwin, M., Sokocevic, D., Wagner, D. E., Coffey, A., et al. (2015). Systemic administration of human bone marrow-derived mesenchymal stromal cell extracellular vesicles AmelioratesAspergillusHyphal extract-induced allergic airway inflammation in immunocompetent mice. Stem Cell Transl. Med. 4, 1302–1316. doi:10.5966/sctm.2014-0280
Cruz, F. F., and Rocco, P. R. M. (2020). The potential of mesenchymal stem cell therapy for chronic lung disease. Expert Rev. Respir. Med. 14, 31–39. doi:10.1080/17476348.2020.1679628
de Hilster, R. H. J., Sharma, P. K., Jonker, M. R., White, E. S., Gercama, E. A., Roobeek, M., et al. (2020). Human lung extracellular matrix hydrogels resemble the stiffness and viscoelasticity of native lung tissue. Am. J. Physiology-Lung Cell Mol. Physiol. 318, L698–L704. doi:10.1152/ajplung.00451.2019
De Lisio, M., Jensen, T., Sukiennik, R. A., Huntsman, H. D., and Boppart, M. D. (2014). Substrate and strain alter the muscle-derived mesenchymal stem cell secretome to promote myogenesis. Stem Cel Res. Ther. 5, 74. doi:10.1186/scrt463
de Oliveira, H. G., Cruz, F. F., Antunes, M. A., de Macedo Neto, A. V., Oliveira, G. A., Svartman, F. M., et al. (2017). Combined bone marrow-derived mesenchymal stromal cell therapy and one-way endobronchial valve placement in patients with pulmonary emphysema: a phase I clinical trial. STEM CELLS Translational Med. 6, 962–969. doi:10.1002/sctm.16-0315
De Santis, M. M., Alsafadi, H. N., Tas, S., Bölükbas, D. A., Prithiviraj, S., Da Silva, I. A. N., et al. (2020). Extracellular‐matrix‐reinforced bioinks for 3D bioprinting human tissue. Adv. Mater. 33, 2005476. doi:10.1002/adma.202005476
Deng, Z., Fear, M. W., Suk Choi, Y., Wood, F. M., Allahham, A., Mutsaers, S. E., et al. (2020). The extracellular matrix and mechanotransduction in pulmonary fibrosis. Int. J. Biochem. Cel Biol. 126, 105802. doi:10.1016/j.biocel.2020.105802
Elowsson Rendin, L., Löfdahl, A., Åhrman, E., Müller, C., Notermans, T., Michaliková, B., et al. (2019). Matrisome properties of scaffolds direct fibroblasts in idiopathic pulmonary fibrosis. Ijms 20, 4013. doi:10.3390/ijms20164013
Emblom-Callahan, M. C., Chhina, M. K., Shlobin, O. A., Ahmad, S., Reese, E. S., Iyer, E. P. R., et al. (2010). Genomic phenotype of non-cultured pulmonary fibroblasts in idiopathic pulmonary fibrosis. Genomics 96, 134–145. doi:10.1016/j.ygeno.2010.04.005
J. D. Esko, H. P. James, A. Varki, R. D. Cummings, P. Stanley, R. J. Linhardtet al. Editors (2015). in Essentials of glycobiology, 3rd edition. Cold Spring Harbor (NY): Cold Spring Harbor Laboratory Press, 493–502.
Farhat, W., Hasan, A., Lucia, L., Becquart, F., Ayoub, A., and Kobeissy, F. (2019). Hydrogels for advanced stem cell therapies: a biomimetic materials approach for enhancing natural tissue function. IEEE Rev. Biomed. Eng. 12, 333–351. doi:10.1109/RBME.2018.2824335
Ferreira, S. A., Motwani, M. S., Faull, P. A., Seymour, A. J., Yu, T. T. L., Enayati, M., et al. (2018). Bi-directional cell-pericellular matrix interactions direct stem cell fate. Nat. Commun. 9, 4049. doi:10.1038/s41467-018-06183-4
Gad, E. S., Salama, A. A. A., El-Shafie, M. F., Arafa, H. M. M., Abdelsalam, R. M., and Khattab, M. (2020). The anti-fibrotic and anti-inflammatory potential of bone marrow-derived mesenchymal stem cells and Nintedanib in bleomycin-induced lung fibrosis in rats. Inflammation 43, 123–134. doi:10.1007/s10753-019-01101-2
Gao, F., Chiu, S. M., Motan, D. A. L., Zhang, Z., Chen, L., Ji, H.-L., et al. (2016). Mesenchymal stem cells and immunomodulation: current status and future prospects. Cell Death Dis 7, e2062. doi:10.1038/cddis.2015.327
Gao, J., Kalafatis, D., Carlson, L., Pesonen, I. H. A., Li, C. X., Wheelock, Å., et al. (2021). Baseline characteristics and survival of patients of idiopathic pulmonary fibrosis: a longitudinal analysis of the Swedish IPF Registry. Respir. Res. 22, 40. doi:10.1186/s12931-021-01634-x
Glassberg, M. K., Minkiewicz, J., Toonkel, R. L., Simonet, E. S., Rubio, G. A., DiFede, D., et al. (2017). Allogeneic human mesenchymal stem cells in patients with idiopathic pulmonary fibrosis via intravenous delivery (AETHER). Chest 151, 971–981. doi:10.1016/j.chest.2016.10.061
Haak, A. J., Tan, Q., and Tschumperlin, D. J. (2018). Matrix biomechanics and dynamics in pulmonary fibrosis. Matrix Biol. 73, 64–76. doi:10.1016/j.matbio.2017.12.004
Hallgren, O., Nihlberg, K., Dahlbäck, M., Bjermer, L., Eriksson, L. T., Erjefält, J. S., et al. (2010). Altered fibroblast proteoglycan production in COPD. Respir. Res. 11, 55. doi:10.1186/1465-9921-11-55
Hedström, U., Hallgren, O., Öberg, L., DeMicco, A., Vaarala, O., Westergren-Thorsson, G., et al. (2018). Bronchial extracellular matrix from COPD patients induces altered gene expression in repopulated primary human bronchial epithelial cells. Sci. Rep. 8, 3502. doi:10.1038/s41598-018-21727-w
Hilberg, O., Bendstrup, E., Ibsen, R., Løkke, A., and Hyldgaard, C. (2018). Economic consequences of idiopathic pulmonary fibrosis in Denmark. ERJ Open Res. 4, 00045. doi:10.1183/23120541.00045-2017
Ibáñez-Fonseca, A., Santiago Maniega, S., Gorbenko del Blanco, D., Catalán Bernardos, B., Vega Castrillo, A., Álvarez Barcia, Á. J., et al. (2020). Elastin-like recombinamer hydrogels for improved skeletal muscle healing through modulation of macrophage polarization. Front. Bioeng. Biotechnol. 8, 413. doi:10.3389/fbioe.2020.00413
Ibrahimi, O. A., Zhang, F., Lang Hrstka, S. C., Mohammadi, M., and Linhardt, R. J. (2004). Kinetic model for FGF, FGFR, and proteoglycan signal transduction complex assembly†. Biochemistry 43, 4724–4730. doi:10.1021/bi0352320
Islam, D., Huang, Y., Fanelli, V., Delsedime, L., Wu, S., Khang, J., et al. (2019). Identification and modulation of microenvironment is crucial for effective mesenchymal stromal cell therapy in acute lung injury. Am. J. Respir. Crit. Care Med. 199, 1214–1224. doi:10.1164/rccm.201802-0356OC
Islam, M. N., Das, S. R., Emin, M. T., Wei, M., Sun, L., Westphalen, K., et al. (2012). Mitochondrial transfer from bone-marrow-derived stromal cells to pulmonary alveoli protects against acute lung injury. Nat. Med. 18, 759–765. doi:10.1038/nm.2736
Jackson, M. V., Morrison, T. J., Doherty, D. F., McAuley, D. F., Matthay, M. A., Kissenpfennig, A., et al. (2016). Mitochondrial transfer via tunneling nanotubes is an important mechanism by which mesenchymal stem cells enhance macrophage phagocytosis in the in vitro and in vivo models of ARDS. Stem Cells 34, 2210–2223. doi:10.1002/stem.2372
Jiang, S., Lyu, C., Zhao, P., Li, W., Kong, W., Huang, C., et al. (2019). Cryoprotectant enables structural control of porous scaffolds for exploration of cellular mechano-responsiveness in 3D. Nat. Commun. 10, 3491. doi:10.1038/s41467-019-11397-1
Kim, H., Bae, C., Kook, Y.-M., Koh, W.-G., Lee, K., and Park, M. H. (2019). Mesenchymal stem cell 3D encapsulation technologies for biomimetic microenvironment in tissue regeneration. Stem Cell Res. Ther. 10, 51. doi:10.1186/s13287-018-1130-8
Kusuma, G. D., Carthew, J., Lim, R., and Frith, J. E. (2017). Effect of the microenvironment on mesenchymal stem cell paracrine signaling: opportunities to engineer the therapeutic effect. Stem Cell Development 26, 617–631. doi:10.1089/scd.2016.0349
Laing, A. G., Fanelli, G., Ramirez-Valdez, A., Lechler, R. I., Lombardi, G., and Sharpe, P. T. (2019). Mesenchymal stem cells inhibit T-cell function through conserved induction of cellular stress. PLoS One 14, e0213170. doi:10.1371/journal.pone.0213170
Laurent, G. J. (1987). Dynamic state of collagen: pathways of collagen degradation in vivo and their possible role in regulation of collagen mass. Am. J. Physiology-Cell Physiol. 252, C1–C9. doi:10.1152/ajpcell.1987.252.1.C1
Lee, M., Jeong, S. Y., Ha, J., Kim, M., Jin, H. J., Kwon, S.-J., et al. (2014). Low immunogenicity of allogeneic human umbilical cord blood-derived mesenchymal stem cells in vitro and in vivo. Biochem. Biophysical Res. Commun. 446, 983–989. doi:10.1016/j.bbrc.2014.03.051
Li, X., Zhang, Y., Yeung, S. C., Liang, Y., Liang, X., Ding, Y., et al. (2014). Mitochondrial transfer of induced pluripotent stem cell-derived mesenchymal stem cells to airway epithelial cells attenuates cigarette smoke-induced damage. Am. J. Respir. Cel Mol. Biol. 51, 455–465. doi:10.1165/rcmb.2013-0529OC
Loebel, C., Mauck, R. L., and Burdick, J. A. (2019). Local nascent protein deposition and remodelling guide mesenchymal stromal cell mechanosensing and fate in three-dimensional hydrogels. Nat. Mater. 18, 883–891. doi:10.1038/s41563-019-0307-6
Mahrouf-Yorgov, M., Augeul, L., Da Silva, C. C., Jourdan, M., Rigolet, M., Manin, S., et al. (2017). Mesenchymal stem cells sense mitochondria released from damaged cells as danger signals to activate their rescue properties. Cell Death Differ 24, 1224–1238. doi:10.1038/cdd.2017.51
Mansouri, N., Willis, G. R., Fernandez-Gonzalez, A., Reis, M., Nassiri, S., Mitsialis, S. A., et al. (2019). Mesenchymal stromal cell exosomes prevent and revert experimental pulmonary fibrosis through modulation of monocyte phenotypes. JCI Insight 4, e128060. doi:10.1172/jci.insight.128060
Mardpour, S., Hamidieh, A. A., Taleahmad, S., Sharifzad, F., Taghikhani, A., and Baharvand, H. (2019). Interaction between mesenchymal stromal cell‐derived extracellular vesicles and immune cells by distinct protein content. J. Cel Physiol 234, 8249–8258. doi:10.1002/jcp.27669
Martinez, F. J., Collard, H. R., Pardo, A., Raghu, G., Richeldi, L., Selman, M., et al. (2017). Idiopathic pulmonary fibrosis. Nat. Rev. Dis. Primers 3, 17074. doi:10.1038/nrdp.2017.74
Meisel, R., Zibert, A., Laryea, M., Göbel, U., Däubener, W., and Dilloo, D. (2004). Human bone marrow stromal cells inhibit allogeneic T-cell responses by indoleamine 2,3-dioxygenase-mediated tryptophan degradation. Blood 103, 4619–4621. doi:10.1182/blood-2003-11-3909
Mias, C. l., Lairez, O., Trouche, E., Roncalli, J. r., Calise, D., Seguelas, M.-H. l. n., et al. (2009). Mesenchymal stem cells promote matrix metalloproteinase secretion by cardiac fibroblasts and reduce cardiac ventricular fibrosis after myocardial infarction. Stem Cells 27, 2734–2743. doi:10.1002/stem.169
Miron-Mendoza, M., Graham, E., Manohar, S., and Petroll, W. M. (2017). Fibroblast-fibronectin patterning and network formation in 3D fibrin matrices. Matrix Biol. 64, 69–80. doi:10.1016/j.matbio.2017.06.001
Moodley, Y., Atienza, D., Manuelpillai, U., Samuel, C. S., Tchongue, J., Ilancheran, S., et al. (2009). Human umbilical cord mesenchymal stem cells reduce fibrosis of bleomycin-induced lung injury. Am. J. Pathol. 175, 303–313. doi:10.2353/ajpath.2009.080629
Morrison, T. J., Jackson, M. V., Cunningham, E. K., Kissenpfennig, A., McAuley, D. F., O’Kane, C. M., et al. (2017). Mesenchymal stromal cells modulate macrophages in clinically relevant lung injury models by extracellular vesicle mitochondrial transfer. Am. J. Respir. Crit. Care Med. 196, 1275–1286. doi:10.1164/rccm.201701-0170OC
Murphy, M. B., Moncivais, K., and Caplan, A. I. (2013). Mesenchymal stem cells: environmentally responsive therapeutics for regenerative medicine. Exp. Mol. Med. 45, e54. doi:10.1038/emm.2013.94
Negewo, N. A., Gibson, P. G., and McDonald, V. M. (2015). COPD and its comorbidities: impact, measurement and mechanisms. Respirology 20, 1160–1171. doi:10.1111/resp.12642
Nonaka, P. N., Falcones, B., Farre, R., Artigas, A., Almendros, I., and Navajas, D. (2020). Biophysically preconditioning mesenchymal stem cells improves treatment of ventilator-induced lung injury. Archivos de Bronconeumología 56, 179–181. doi:10.1016/j.arbres.2019.08.014
Ornell, K. J., Lozada, D., Phan, N. V., and Coburn, J. M. (2019). Controlling methacryloyl substitution of chondroitin sulfate: injectable hydrogels with tunable long-term drug release profiles. J. Mater. Chem. B 7, 2151–2161. doi:10.1039/c8tb03020k
Pachler, K., Ketterl, N., Desgeorges, A., Dunai, Z., Laner-Plamberger, S., Streif, D., et al. (2017). An in vitro potency assay for monitoring the immunomodulatory potential of stromal cell-derived extracellular vesicles. Ijms 18, 1413. doi:10.3390/ijms18071413
Papakonstantinou, E., and Karakiulakis, G. (2009). The 'sweet' and 'bitter' involvement of glycosaminoglycans in lung diseases: pharmacotherapeutic relevance. Br. J. Pharmacol. 157, 1111–1127. doi:10.1111/j.1476-5381.2009.00279.x
Parker, M. W., Rossi, D., Peterson, M., Smith, K., Sikström, K., White, E. S., et al. (2014). Fibrotic extracellular matrix activates a profibrotic positive feedback loop. J. Clin. Invest. 124, 1622–1635. doi:10.1172/JCI71386
Persson, A., Vorontsov, E., Larson, G., and Nilsson, J. (2020). Glycosaminoglycan domain mapping of cellular chondroitin/dermatan sulfates. Sci. Rep. 10, 3506. doi:10.1038/s41598-020-60526-0
Philp, C. J., Siebeke, I., Clements, D., Miller, S., Habgood, A., John, A. E., et al. (2018). Extracellular matrix cross-linking enhances fibroblast growth and protects against matrix proteolysis in lung fibrosis. Am. J. Respir. Cel Mol. Biol. 58, 594–603. doi:10.1165/rcmb.2016-0379OC
Phinney, D. G., Di Giuseppe, M., Njah, J., Sala, E., Shiva, S., St Croix, C. M., et al. (2015). Mesenchymal stem cells use extracellular vesicles to outsource mitophagy and shuttle microRNAs. Nat. Commun. 6, 8472. doi:10.1038/ncomms9472
Pouliot, R. A., Young, B. M., Link, P. A., Park, H. E., Kahn, A. R., Shankar, K., et al. (2020). Porcine lung-derived extracellular matrix hydrogel properties are dependent on pepsin digestion time. Tissue Eng. C: Methods 26, 332–346. doi:10.1089/ten.TEC.2020.0042
Prockop, D. J. (2013). Concise review: two negative feedback loops place mesenchymal stem/stromal cells at the center of early regulators of inflammation. Stem Cells 31, 2042–2046. doi:10.1002/stem.1400
Raghu, G., Collard, H. R., Egan, J. J., Martinez, F. J., Behr, J., Brown, K. K., et al. (2011). An official ATS/ERS/JRS/ALAT statement: idiopathic pulmonary fibrosis: evidence-based guidelines for diagnosis and management. Am. J. Respir. Crit. Care Med. 183, 788–824. doi:10.1164/rccm.2009-040GL
Reddy, M., Fonseca, L., Gowda, S., Chougule, B., Hari, A., and Totey, S. (2016). Human adipose-derived mesenchymal stem cells attenuate early stage of bleomycin induced pulmonary fibrosis: comparison with Pirfenidone. Ijsc 9, 192–206. doi:10.15283/ijsc16041
Ribeiro-Paes, J. T., Bilaqui, fnm., Greco, O. T., Ruiz, M. A., Marcelino, fnm., Stessuk, fnm., et al. (2011). Unicentric study of cell therapy in chronic obstructive pulmonary disease/pulmonary emphysema. Copd 6, 63–71. doi:10.2147/COPD.S15292
Rodriguez, L. R., Emblom-Callahan, M., Chhina, M., Bui, S., Aljeburry, B., Tran, L. H., et al. (2018). Global gene expression analysis in an in vitro fibroblast model of idiopathic pulmonary fibrosis reveals potential role for CXCL14/CXCR4. Sci. Rep. 8, 3983. doi:10.1038/s41598-018-21889-7
Saito, A., Horie, M., and Nagase, T. (2018). TGF-β signaling in lung health and disease. Ijms 19, 2460. doi:10.3390/ijms19082460
Sanz-Fraile, H., Amoros, S., Mendizabal, I., Galvez-Monton, C., Prat-Vidal, C., Bayes-Genis, A., et al. (2020). Silk-reinforced collagen hydrogels with raised multiscale stiffness for mesenchymal cells 3D culture. Tissue Eng. A 26, 358–370. doi:10.1089/ten.TEA.2019.0199
Semedo, P., Correa-Costa, M., Cenedeze, M. A., Malheiros, D. M. A. C., Antonia dos Reis, M., Shimizu, M. H., et al. (2009). Mesenchymal stem cells attenuate renal fibrosis through immune modulation and remodeling properties in a rat remnant kidney model. Stem Cells 27, A–N. doi:10.1002/stem.214
Shamskhou, E. A., Kratochvil, M. J., Orcholski, M. E., Nagy, N., Kaber, G., Steen, E., et al. (2019). Hydrogel-based delivery of Il-10 improves treatment of bleomycin-induced lung fibrosis in mice. Biomaterials 203, 52–62. doi:10.1016/j.biomaterials.2019.02.017
Shapiro, S. D., Endicott, S. K., Province, M. A., Pierce, J. A., and Campbell, E. J. (1991). Marked longevity of human lung parenchymal elastic fibers deduced from prevalence of D-aspartate and nuclear weapons-related radiocarbon. J. Clin. Invest. 87, 1828–1834. doi:10.1172/JCI115204
Sherman, V. R., Yang, W., and Meyers, M. A. (2015). The materials science of collagen. J. Mech. Behav. Biomed. Mater. 52, 22–50. doi:10.1016/j.jmbbm.2015.05.023
Shukla, M. N., Rose, J. L., Ray, R., Lathrop, K. L., Ray, A., and Ray, P. (2009). Hepatocyte growth factor inhibits epithelial to myofibroblast transition in lung cells via Smad7. Am. J. Respir. Cell Mol Biol 40, 643–653. doi:10.1165/rcmb.2008-0217OC
Silva, L. H. A., Antunes, M. A., Dos Santos, C. C., Weiss, D. J., Cruz, F. F., and Rocco, P. R. M. (2018). Strategies to improve the therapeutic effects of mesenchymal stromal cells in respiratory diseases. Stem Cel Res Ther 9, 45. doi:10.1186/s13287-018-0802-8
Sin, D. D., Anthonisen, N. R., Soriano, J. B., and Agusti, A. G. (2006). Mortality in COPD: role of comorbidities. Eur. Respir. J. 28, 1245–1257. doi:10.1183/09031936.00133805
Spees, J. L., Olson, S. D., Whitney, M. J., and Prockop, D. J. (2006). Mitochondrial transfer between cells can rescue aerobic respiration. Proc. Natl. Acad. Sci. 103, 1283–1288. doi:10.1073/pnas.0510511103
Srour, N., and Thébaud, B. (2015). Mesenchymal stromal cells in animal bleomycin pulmonary fibrosis models: a systematic review. Stem Cell Transl Med 4, 1500–1510. doi:10.5966/sctm.2015-0121
Stolk, J., Broekman, W., Mauad, T., Zwaginga, J. J., Roelofs, H., Fibbe, W. E., et al. (2016). A phase I study for intravenous autologous mesenchymal stromal cell administration to patients with severe emphysema. QJM 109, 331–336. doi:10.1093/qjmed/hcw001
Tan, Y., Qiao, Y., Chen, Z., Liu, J., Guo, Y., Tran, T., et al. (2020). FGF2, an immunomodulatory factor in asthma and chronic obstructive pulmonary disease (COPD). Front. Cel Dev. Biol. 8, 223. doi:10.3389/fcell.2020.00223
Thery, C., Witwer, K. W., Aikawa, E., Alcaraz, M. J., Anderson, J. D., Andriantsitohaina, R., et al. (2018). Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell Vesicles 7, 1535750. doi:10.1080/20013078.2018.1535750
Tisler, M., Alkmin, S., Chang, H.-Y., Leet, J., Bernau, K., Sandbo, N., et al. (2020). Analysis of fibroblast migration dynamics in idiopathic pulmonary fibrosis using image-based scaffolds of the lung extracellular matrix. Am. J. Physiology-Lung Cell Mol. Physiol. 318, L276–L286. doi:10.1152/ajplung.00087.2019
Trachalaki, A., Irfan, M., and Wells, A. U. (2021). Pharmacological management of Idiopathic Pulmonary Fibrosis: current and emerging options. Expert Opin. Pharmacother. 22, 191–204. doi:10.1080/14656566.2020.1822326
Tzouvelekis, A., Paspaliaris, V., Koliakos, G., Ntolios, P., Bouros, E., Oikonomou, A., et al. (2013). A prospective, non-randomized, no placebo-controlled, phase Ib clinical trial to study the safety of the adipose derived stromal cells-stromal vascular fraction in idiopathic pulmonary fibrosis. J. Translational Med. 11, 171. doi:10.1186/1479-5876-11-171
Uhl, F. E., Zhang, F., Pouliot, R. A., Uriarte, J. J., Rolandsson Enes, S., Han, X., et al. (2020). Functional role of glycosaminoglycans in decellularized lung extracellular matrix. Acta Biomater. 102, 231–246. doi:10.1016/j.actbio.2019.11.029
van Niel, G., D'Angelo, G., and Raposo, G. (2018). Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cel Biol 19, 213–228. doi:10.1038/nrm.2017.125
Varkouhi, A. K., Jerkic, M., Ormesher, L., Gagnon, S., Goyal, S., Rabani, R., et al. (2019). Extracellular vesicles from interferon-γ-primed human umbilical cord mesenchymal stromal cells reduce Escherichia coli-induced acute lung injury in rats. Anesthesiology 130, 778–790. doi:10.1097/ALN.0000000000002655
Vogel, V. (2018). Unraveling the mechanobiology of extracellular matrix. Annu. Rev. Physiol. 80, 353–387. doi:10.1146/annurev-physiol-021317-121312
Wan, X., Chen, S., Fang, Y., Zuo, W., Cui, J., and Xie, S. (2020). Mesenchymal stem cell‐derived extracellular vesicles suppress the fibroblast proliferation by downregulating FZD6 expression in fibroblasts via micrRNA‐29b‐3p in idiopathic pulmonary fibrosis. J. Cel Physiol. 235, 8613–8625. doi:10.1002/jcp.29706
Wang, H., Cao, F., De, A., Cao, Y., Contag, C., Gambhir, S. S., et al. (2009). Trafficking mesenchymal stem cell engraftment and differentiation in tumor-bearing mice by bioluminescence imaging. Stem Cells 27, 1548–1558. doi:10.1002/stem.81
Wang, L., Zhao, Y., Yang, F., Feng, M., Zhao, Y., Chen, X., et al. (2020). Biomimetic collagen biomaterial induces in situ lung regeneration by forming functional alveolar. Biomaterials 236, 119825. doi:10.1016/j.biomaterials.2020.119825
Weiss, D. J., Casaburi, R., Flannery, R., LeRoux-Williams, M., and Tashkin, D. P. (2013). A placebo-controlled, randomized trial of mesenchymal stem cells in COPD. Chest 143, 1590–1598. doi:10.1378/chest.12-2094
Westergren-Thorsson, G., Hedström, U., Nybom, A., Tykesson, E., Åhrman, E., Hornfelt, M., et al. (2017). Increased deposition of glycosaminoglycans and altered structure of heparan sulfate in idiopathic pulmonary fibrosis. Int. J. Biochem. Cel Biol. 83, 27–38. doi:10.1016/j.biocel.2016.12.005
Widhe, M., Shalaly, N. D., and Hedhammar, M. (2016). A fibronectin mimetic motif improves integrin mediated cell biding to recombinant spider silk matrices. Biomaterials 74, 256–266. doi:10.1016/j.biomaterials.2015.10.013
Wigén, J., Elowsson-Rendin, L., Karlsson, L., Tykesson, E., and Westergren-Thorsson, G. (2019). Glycosaminoglycans: a link between development and regeneration in the lung. Stem Cell Development 28, 823–832. doi:10.1089/scd.2019.0009
Woldhuis, R. R., de Vries, M., Timens, W., van den Berge, M., Demaria, M., Oliver, B. G. G., et al. (2020). Link between increased cellular senescence and extracellular matrix changes in COPD. Am. J. Physiology-Lung Cell Mol. Physiol. 319, L48–L60. doi:10.1152/ajplung.00028.2020
Wong, S. W., Lenzini, S., Cooper, M. H., Mooney, D. J., and Shin, J.-W. (2020). Soft extracellular matrix enhances inflammatory activation of mesenchymal stromal cells to induce monocyte production and trafficking. Sci. Adv. 6, eaaw0158. doi:10.1126/sciadv.aaw0158
Wozniak, M. A., Modzelewska, K., Kwong, L., and Keely, P. J. (2004). Focal adhesion regulation of cell behavior. Biochim. Biophys. Acta (Bba) - Mol. Cel Res. 1692, 103–119. doi:10.1016/j.bbamcr.2004.04.007
Yang, C., Tibbitt, M. W., Basta, L., and Anseth, K. S. (2014). Mechanical memory and dosing influence stem cell fate. Nat. Mater 13, 645–652. doi:10.1038/nmat3889
Yawn, B. P., Mintz, M. L., and Doherty, D. E. (2021). GOLD in practice: chronic obstructive pulmonary disease treatment and management in the primary care setting. Copd 16, 289–299. doi:10.2147/COPD.S222664
Yayon, A., Klagsbrun, M., Esko, J. D., Leder, P., and Ornitz, D. M. (1991). Cell surface, heparin-like molecules are required for binding of basic fibroblast growth factor to its high affinity receptor. Cell 64, 841–848. doi:10.1016/0092-8674(91)90512-w
Keywords: chronic objective pulmonary disease, MSC, extracellular matrix, artificial lung scaffolds, idiopathic pulmonary fibrosis
Citation: Elowsson Rendin L, Löfdahl A, Kadefors M, Söderlund Z, Tykesson E, Rolandsson Enes S, Wigén J and Westergren-Thorsson G (2021) Harnessing the ECM Microenvironment to Ameliorate Mesenchymal Stromal Cell-Based Therapy in Chronic Lung Diseases. Front. Pharmacol. 12:645558. doi: 10.3389/fphar.2021.645558
Received: 23 December 2020; Accepted: 17 March 2021;
Published: 15 April 2021.
Edited by:
Miquéias Lopes-Pacheco, University of Lisbon, PortugalReviewed by:
Elga Bandeira, Karolinska Institutet (KI), SwedenVera Luiza Capelozzi, University of São Paulo, Brazil
Laura Russo, Università di Milano-Bicocca, Italy
Copyright © 2021 Elowsson Rendin, Löfdahl, Kadefors, Söderlund, Tykesson, Rolandsson Enes, Wigén and Westergren-Thorsson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Linda Elowsson Rendin, bGluZGEuZWxvd3Nzb25fcmVuZGluQG1lZC5sdS5zZQ==