 Ping Song
Ping Song Zohaib Shafiq
Zohaib Shafiq Hongxiang Jiang
Hongxiang Jiang Qiang Cai
Qiang Cai- Department of Neurosurgery, Renmin Hospital of Wuhan University, Wuhan, China
Stroke ranks among the top global causes of death and disability, with hemorrhagic stroke accounting for 9%–27% of cases. In China, intracerebral hemorrhage (ICH) outpaces Western rates, driving adult mortality and disability. Aging populations and urbanization diversify stroke risk factors, yielding a 28-day ICH mortality rate up to 47% (Greenberg et al., 2022)—sharply higher than ischemic stroke’s 3%—with ∼75% of survivors facing permanent neurological deficits. ICH imposes heavy burdens on individuals, families, and society. Current medical and surgical treatments struggle to enhance long-term outcomes, prompting exploration of neural stem cell (NSC) transplantation. This approach replaces lost neurons via differentiation while offering anti-inflammatory, anti-apoptotic, and neuroprotective benefits. This systematic review evaluates NSC sources, delivery routes, ICH models, therapeutic mechanisms, and early clinical trials, outlining recent progress and future directions for treating hemorrhagic stroke with NSC transplantation.
1 Methods
This systematic review was conducted in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines. We searched PubMed, Embase, Web of Science, ClinicalTrials.gov, and the Chinese Clinical Trial Registry (ChiCTR.org.cn) from January 1990 to August 2025 using keywords such as “neural stem cells,” “NSC,” “stem cell transplantation,” “intracerebral hemorrhage,” “ICH,” “hemorrhagic stroke,” and combinations thereof. Inclusion criteria were: English-language studies (preclinical or clinical) on NSC application in ICH, published peer-reviewed articles, and full-text availability. Exclusion criteria included case reports, reviews, non-ICH stroke studies, and non-NSC stem cell types. Two independent reviewers screened titles/abstracts and full texts, resolving disagreements by consensus. Data extraction included study design, NSC source/route/dose, outcomes, and mechanisms. Quality assessment used SYRCLE’s risk of bias tool for preclinical studies and Cochrane for clinical trials. A PRISMA flow diagram (not shown here; to be included in final submission) identified 1,250 records, with 68 studies included after duplicates and screening. No meta-analysis was performed due to heterogeneity.
2 Introduction
ICH is spontaneous bleeding within the brain, often triggered by hypertension, cerebral amyloid angiopathy, or vascular anomalies such as aneurysms, arteriovenous malformations, Moyamoya disease, or vasculitis (Kim et al., 2022). This devastating condition carries an early mortality rate of 30%–47% (Greenberg et al., 2022) and leaves about two-thirds of patients either dead or permanently disabled (Chen et al., 2020). Current ICH treatments aim to prevent re-bleeding and reduce secondary brain injury, but the nervous system’s limited regenerative capacity results in persistently poor prognoses. Repairing damaged neural tissue or stimulating neurogenesis offers a promising strategy to improve outcomes for ICH patients.
Standard treatments for primary and secondary brain injuries following ICH include surgery, intracranial pressure management, symptomatic care, and rehabilitation. However, these approaches have shown only modest success, highlighting the need for innovative therapies. Stem cell therapy has emerged as a compelling option, gaining attention from researchers (Tuazon et al., 2019; Gao et al., 2018). Stem cells are unique in their ability to proliferate, self-renew, and differentiate into various cell types under specific conditions. They are broadly classified into embryonic stem cells (ESCs) and adult stem cells (ASCs)—also termed somatic stem cells (SSCs)—found in tissues like the brain (neural stem cells, NSCs), blood (hematopoietic stem cells, HSCs), and bone marrow (mesenchymal stem cells, MSCs). NSCs, located in the nervous system, can divide symmetrically or asymmetrically to produce central nervous system (CNS) cell types, including neurons, astrocytes, and oligodendrocytes. First identified by Reynolds and Weiss in 1992 through isolation from adult mouse striatum (Reynolds et al., 1992), NSCs exhibit proliferative and multipotent potential. They can remain quiescent for extended periods, activating in response to injury to support tissue regeneration and cell replacement (Gao et al., 2018). Recent work by Petrelli et al. (2023) underscores the role of mitochondrial metabolism—specifically the mitochondrial pyruvate carrier (MPC)—in regulating adult neural stem/progenitor cell (NSPC) activity, revealing how metabolic cues influence whether NSPCsremain dormant or activate for neuroregeneration.
NSC transplantation holds significant therapeutic potential due to its ability to suppress inflammation, differentiate into neurons, and promote neuroprotection and neurogenesis key features that make it a promising treatment for ICH. This review synthesizes current research on NSCs in ICH therapy, focusing on their sources, administration routes, and clinical implications.
3 Sources of neural stem cells
NSCs used for injury repair can be classified into endogenous and exogenous sources. Current research indicates that both activating dormant endogenous NSCs and transplanting exogenous NSCs can secrete neurotrophic factors to improve their growth microenvironment, suppress inflammation, enhance recovery, and differentiate into neurons to restore lost neurological function in damaged areas.
3.1 Endogenous source of neural stem cells
The traditional view that NSCs exist only during embryonic development and lack regenerative capacity after neural injury has been overturned by accumulating evidence. In 1992, Reynolds and Weiss isolated self-renewing, multipotent NSCs from the striatum of adult mice (Reynolds et al., 1992), sparking further discoveries of NSCs in mammals, birds, and humans, where they exhibit directed migration, proliferation, and differentiation (Lendahl, 2022). These cells predominantly reside in the hippocampal dentate gyrus and the brain’s subventricular zone key regions for NSC research (Gao et al., 2018; Martinez-Galdamez et al., 2019; Mitrecic et al., 2022). Typically quiescent in healthy states, endogenous NSCs activate and migrate to injury sites under pathological conditions. For instance, Gao et al. stimulated endogenous NSCs in ICH rat models to support neuronal repair, with signs of neurogenesis observed in tissues near hematomas in human ICH patients (Gao et al., 2018). Similarly, repeated transcranial magnetic stimulation has been shown to boost NSC proliferation and differentiation via the MAPK signaling pathway post-ICH (Cui et al., 2019). These responsive NSCs, termed endogenous stem cells, offer a safe therapeutic avenue due to their lack of immune rejection and tumorigenic risk (Rodriguez-Frutos et al., 2016). Yu et al. found that upregulating the hypoxia-inducible factor-1α (HIF-1α) gene enhances NSC proliferation, migration, and differentiation, aiding neural recovery in ICH patients (Yu et al., 2013).Physical exercise has even been shown to enhance NSC survival and migration following ICH.These findings highlight NSCs’ neuroplasticity and regenerative potential, supported by their low immunogenicity and compatibility with brain tissue. However, the specific factors triggering NSC differentiation remain unclear (Zhao et al., 2022; Zhou et al., 2022), and their limited numbers and migration challenges restrict their use as a primary ICH treatment strategy.
3.2 Exogenous sources of neural stem cells
Exogenous NSCs are derived from primary tissue cultures, pluripotent stem cell differentiation, or somatic cell reprogramming (Tuazon et al., 2019; Gao et al., 2018). Advances in microdissection have simplified NSC isolation from embryonic tissues, with Krutika et al. successfully extracting NSCs from whole mouse brains (Deshpande et al., 2019) and Zhao Quanjun et al. refining primary culture techniques to obtain NSCs from discarded brain tissue of patients with acute traumatic brain injury, later inducing neuronal differentiation (Quanjun et al., 2019). While feasible, primary tissue culture faces challenges such as low cell yield, limited differentiation efficiency, and the need to maintain stable NSC passages without mutations or senescence. Emerging techniques such as growth factor use, 3D cultures, and microRNA-induced differentiation are improving scalability and safety for clinical applications (Tang et al., 2017). Pluripotent stem cells, though unable to form complete organisms, can differentiate into NSCs and other cell types (Wu et al., 2017). These cells are typically sourced from embryonic tissues or somatic cell nuclear transfer, but ethical concerns, immune rejection risks, and potential tumorigenicity make embryonic sources less ideal (Zhao et al., 2022). Induced pluripotent stem cells (iPSCs), pioneered by Yamanaka through somatic cell reprogramming (Takahashi and Yamanaka, 2006), mimic embryonic stem cells’ pluripotency and serve as a key NSC source. However, producing high-purity NSCs from iPSCs is time-intensive and poses safety risks (Yang et al., 2017).
Viral vectors, the primary method for delivering transgenes, raise concerns about tumorigenesis, complicating clinical translation (Mollinari and Merlo, 2021). Transdifferentiation, the direct conversion of one differentiated cell type into another via selective gene expression (Erharter et al., 2019), offers an alternative approach. Vierbuchen et al. transformed mouse fibroblasts into neurons using transcription factors Ascl1, Brn2, and Myt1l (Vierbuchen et al., 2010), while Hermann et al. employed a two-step process to convert mesenchymal stem cells (MSCs) into NSCs and then neurons (Hermann et al., 2006). Direct transdifferentiation to neurons is simpler but also yields NSCs as a byproduct.
4 Neural stem cell transplantation routes
Currently, there are several pathways for NSC transplantation, including intracerebral transplantation, intrathecal administration, intravascular injection, and intranasal administration. The most commonly used method is intracerebral transplantation, through intracerebral transplantation, intravascular injection, and intracerebral transplantation via the cerebrospinal fluid (CSF) route (Tuazon et al., 2019; Zhao et al., 2022). However, each method has its own advantages and disadvantages, with no optimal transplantation route identified at present (Figure 1). (See Table 1 for a summary of key translational parameters, including routes, time windows, and doses from preclinical and clinical studies.).
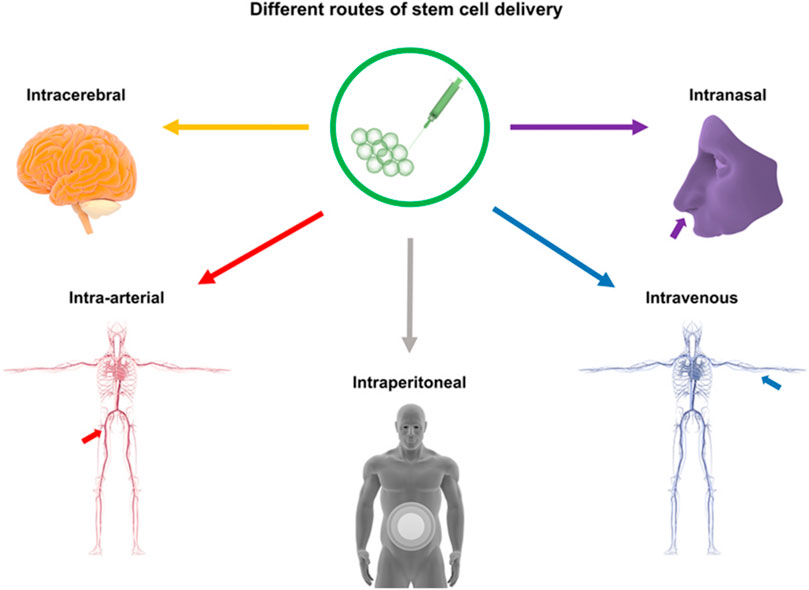
Figure 1. Routes of Stem Cell Transplantation: This figure illustrates stem cell delivery pathways for stroke treatment, including intracerebral (stereotactic injection), intravascular (venous/arterial), intraperitoneal, intraventricular, intravenous, intranasal, and intrathecal methods.
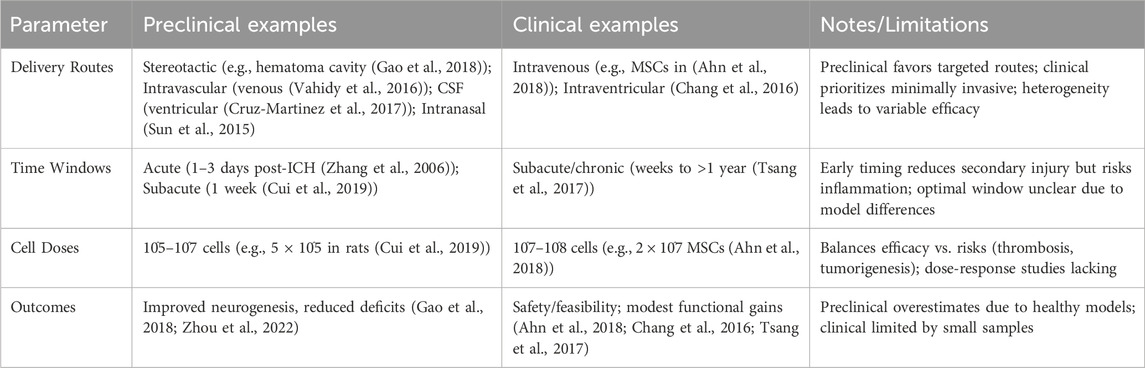
Table 1. Summary of key translational parameters in NSC studies for ICH.
4.1 Stereotactic injection
Stereotactic injection, or intracerebral transplantation, delivers NSC suspensions directly into the hematoma cavity, offering the most targeted approach among transplantation methods. However, blood degradation products and reactive astrocytes in the cavity can trigger NSC death. While NSCs differentiate effectively in vitro, in vivo outcomes are less promising, possibly due to high local cell density impeding differentiation. To improve survival, researchers have developed genetically modified, traceable, anti-apoptotic NSCs (Patent: CN202111510957.7, March 1, 2022). Others suggest co-administering neurotrophic factors during injection to enhance NSC viability, though these strategies await further validation (Gao et al., 2018; Zhou et al., 2022).
4.2 Intracerebrospinal fluid route transplantation
Transplantation via the CSF route includes lumbar puncture, ventricular puncture, and cisterna magna injection. Lumbar puncture delivers NSCs into the subarachnoid space, enabling circulation throughout the brain via CSF. Ventricular puncture injects NSC suspensions into the lateral ventricle, allowing rapid distribution across ventricles and brain tissue while potentially stimulating endogenous NSC proliferation through secreted factors (Cruz-Martinez et al., 2017). Cisterna magna injection, limited to animal models due to its risk of brainstem damage, has not been reported in clinical use.
4.3 Intravascular injection
Intravascular injection of NSCs can be venous or arterial. Venous delivery sends NSCs through systemic circulation, leading to substantial cell loss, with many trapped in the liver and lungs (Vahidy et al., 2016), though multiple injections could theoretically boost survival rates. Arterial injection (typically via the carotid artery) provides direct brain access but poses technical challenges and risks thrombosis due to NSC aggregation (Kim et al., 2022; Gao et al., 2018). Both intravascular and CSF routes face challenges: poor NSC migration, the blood-brain barrier (BBB), and lack of targeted guidance limit cell accumulation in the hematoma cavity. Solutions like targeting molecules or nanomaterial carriers could enhance delivery precision. As NSC transplantation strategies diversify, their impact on neural repair varies, and while experimental progress continues, identifying the safest, most effective routes remains a research priority. Advances in genetics, neurophysiology, and novel therapeutics promise standardized clinical applications soon (Tuazon et al., 2019; Zhao et al., 2022).
Alternative NSC transplantation routes include intranasal administration, which minimizes secondary damage. Sun et al. (Sun et al., 2015) detected hypoxia-preconditioned mesenchymal stem cells (MSCs) in ICH brain tissue following intranasal delivery. Other options, such as abdominal delivery and nanomaterial-enhanced combined routes, aim to improve transplantation efficiency (Tuazon et al., 2019; Gao et al., 2018) (Figure 1).
5 ICH animal model studies
Preclinical research aims to enhance clinical efficacy, prognosis, and quality of life, focusing heavily on constructing animal models to simulate ICH and evaluate SC interventions. These models strive to replicate key human ICH pathophysiological processes, such as hematoma expansion and recurrence. While insights into animal ICH mechanisms grow, future efforts will likely prioritize translating findings to human studies. Given ICH’s complex primary and secondary injury mechanisms, experimental models are vital for elucidating pathophysiology and developing treatments. Established models include autologous blood injection, collagenase injection, intracerebral balloon inflation, and cerebrovascular injury, alongside less common approaches. Rodents (rats and mice) and pigs dominate studies, with rats used in 63% of cases and the autologous blood injection model leading at 51% (Paiva et al., 2023). These models enhance understanding of intracranial pressure, neuroinflammation, immunity, and cerebral hemodynamics, informing treatment strategies and boosting clinical diagnostic and therapeutic capabilities. Table 2 summarizes the main models’ advantages and disadvantages.
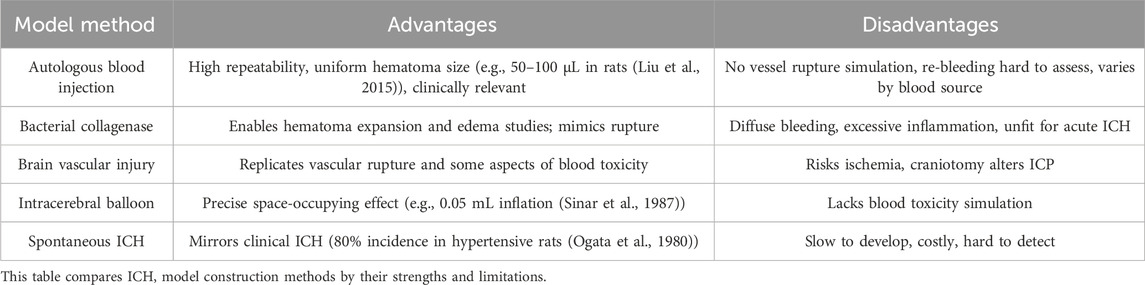
Table 2. Comparison of major ICH models.
5.1 Autologous blood intracerebral injection model
Since the 1960s, the autologous blood intracerebral injection model has offered a straightforward, effective method to create brain parenchymal hematomas by injecting blood into the frontal lobe or basal ganglia. Initially developed in larger animals (e.g., cats, dogs, pigs, sheep, monkeys) (Cao et al., 2016), it enables assessment of hemolysis-induced toxicity, immune-inflammatory responses, and intervention efficacy. Its primary advantage is producing uniform hematoma sizes, mimicking rapid blood accumulation in acute clinical ICH (Liu et al., 2015). However, it fails to replicate spontaneous vascular rupture and often results in blood reflux along the needle track (Yang et al., 1994). By adjusting injected volume, it simulates intracranial hypertension’s biochemical and pathophysiological effects, creating hematomas with varying mass effects (Manaenko et al., 2011). Typically a single injection, the model was refined in 1996 by Deinsberger’s “two-injection method” (Deinsberger et al., 1996): a small initial injection (e.g., into the basal ganglia) forms a clot, followed by a second injection to prevent backflow, a technique widely adopted in small animal studies. Limitations include its inability to model spontaneous bleeding, vascular rupture, or rebleeding (Gao et al., 2018; Yang et al., 1994).
5.2 ICH collagenase model
Bacterial collagenase, a metalloproteinase, degrades collagen in the extracellular matrix and basement membranes around brain capillaries, weakening vessels and inducing hemorrhage (Manaenko et al., 2011). Introduced in mice in the early 1990s (Rosenberg et al., 1990), this model excels in studying hematoma expansion, vasogenic edema, anticoagulation effects, axonal degeneration, iron-mediated apoptosis, endothelial dysfunction, and BBB damage. Compared to the autologous blood injection model, it produces more pronounced pathological effects despite sharing similar injury mechanisms (Paiva et al., 2023). Bleeding is diffuse with a gradual hematoma onset, differing from acute clinical hematomas. Typically, researchers inject bacterial type IV collagenase though types VI, VII S, or XI are also used over 2–16 min (Paiva et al., 2023). Preferred for small animals like rats and mice, it effectively mimics hematoma expansion and edema, aiding investigations into ICH-related brain edema, neurological recovery, drug efficacy, and homeostasis-modulating therapies (Jing et al., 2019). It also simulates vascular rupture, supporting long-term outcome studies. However, its early and prolonged inflammatory response may exaggerate inflammation, complicating mechanistic studies, while excessive bleeding risks unintended ischemic damage, potentially worsening neurological deficits and recovery (Zhou et al., 2013).
5.3 ICH cerebrovascular injury model
The cerebrovascular injury model induces cortical bleeding in rats by exposing cortical veins via craniotomy and puncturing them with a bent needle (Xue and Del Bigio, 2003). Compared to blood infusion and collagenase models, it shows distinct inflammatory, molecular, and cellular responses (Xue and Del Bigio, 2003). Peak DNA damage in neurons and CD8-reactive lymphocytes occurs at 3 days post-injury, with sustained microglia/macrophage activation from 3 to 7 days, though only minor neuronal death persists by 21–28 days. Neutrophil counts are notably lower than in the other models (Xue and Del Bigio, 2003). Alternatives include laser-induced microbleeds to study coagulation (Lauer et al., 2011) and ultrasound-guided middle cerebral artery puncture in dogs, achieving high success (Zhou et al., 2013). Limited to open craniotomy, this model tends to cause less severe brain damage.
5.4 ICH intracerebral balloon model
This rat ICH model uses a microsphere catheter method (Sinar et al., 1987). A 25 Fr microballoon, mounted on a needle, is inserted into the right caudate nucleus via a skull burr hole, inflated to 0.05 mL in 20 s, and held for 10 min before deflation. Post-study analyses of brain histology, intracranial pressure (ICP), and cerebral blood flow reveal reduced flow and elevated ICP, confirming effective injury. The model’s key strength is mimicking the space-occupying effect of acute human ICH (Alharbi et al., 2016), making it ideal for studying mechanical brain injury. However, limitations include: (1) no replication of blood toxicity, reducing ischemia compared to blood injection models; (2) no blood-brain barrier disruption; and (3) no edema formation (Paiva et al., 2023).
5.5 Spontaneous ICH model
Hypertension, a leading cause of human cerebrovascular disease, underpins this model, where genetically manipulated hypertensive rats develop spontaneous vascular rupture resembling clinical ICH. Inbreeding achieves an 80% ICH incidence (Ogata et al., 1980). Its strength is replicating human spontaneous ICH and hypertension-related stroke pathophysiology. Drawbacks include lengthy breeding times, high costs, and risks of inbreeding-related genetic variability, restricting its use.
5.6 Other ICH models
5.6.1 High salt method
High sodium intake elevates blood pressure, disrupting cerebral vessel tight junctions and causing small, scattered hematomas similar to clinical cases (Appel, 2014). IIscts drawback is the prolonged time to results, delaying research.
5.6.2 Ischemia induction method
Occluding the middle cerebral artery induces ischemia, followed by CO2 and anticoagulants to trigger hemorrhage in the ischemic zone, ideal for studying post-stroke rebleeding.
5.6.3 Trauma-induced ICH method
Violent trauma creates intracerebral hematomas but is hard to control, with high variability, poor reproducibility, and frequent animal mortality, limiting it to traumatic ICH simulation.
5.6.4 Composite and other methods
Combining methods (e.g., with heparin) enhances ICH models, while ultrasound or angiography-guided vessel punctures offer additional approaches (Zhou et al., 2013).
5.7 Limitations of animal models and translational relevance
Although healthy animal models provide valuable insights, they do not fully replicate clinical ICH pathophysiology. For example, the collagenase model exaggerates acute inflammation (e.g., higher IL-1β/TNF-α levels than in humans) and diffuse bleeding, while autologous blood models fail to capture spontaneous vascular rupture or rebleeding, leading to overestimation of NSC efficacy in controlled environments (Paiva et al., 2023; Zhou et al., 2013). Human ICH often involves comorbidities like hypertension, diabetes, or aging, absent in young rodent models, which may reduce generalizability—e.g., comorbidities could impair NSC migration or survival. These discrepancies highlight the need for advanced models (e.g., aged/comorbid animals) to bridge the translational gap and better predict clinical outcomes.
6 Impact of ICH pathophysiology on NSCs
ICH pathophysiology arises from hematoma compression and ongoing damage by blood and its breakdown products, categorized as primary brain injury (PBI) and secondary brain injury (SBI). In experimental animals, prognosis hinges on: (1) the hematoma’s space-occupying effect and resulting ischemia-hypoxia; (2) thrombin toxicity and plasminogen activation with fibrinolysis; (3) SBI from red blood cell lysis and metabolic byproducts; and (4) tissue damage from clots and inflammatory/complement cascades. These factors highlight key considerations for NSC transplantation:
6.1 Cell quantity
The optimal NSC dose for ICH remains unclear, varying by cell type and delivery route. Most studies use millions of cells, balancing efficacy against risks like thrombosis or tumorigenesis from excessive doses (Gao et al., 2018).
6.2 Timing
Early transplantation is favored (Auriat et al., 2011), reducing SBI (e.g., inflammation, apoptosis) within the first week post-ICH to aid recovery. Zhang et al. found MSC transplantation on day 3 outperformed days 1, 5, or 7, linking early hematoma clearance to better cell survival (Zhang et al., 2006).
6.3 Immunomodulation
While some studies report no acute rejection in autologous settings, evidence of immune rejection exists in allogeneic NSC transplantation (e.g., T-cell mediated responses), necessitating further research on immunosuppressive therapy (Pluchino et al., 2005).
7 NSCs neurorepair mechanisms
7.1 Replacement effect
Experiments demonstrate that NSC transplantation improves neurological deficits by differentiating into neurons and glial cells to replace lost neurons. These new neurons form functional synaptic connections with existing ones a process termed the replacement effect (Ceto et al., 2020). However, subsequent studies suggest this mechanism alone cannot fully account for functional improvement, as NSC-derived neurons exhibit limited synaptic integration, and their signal transmission efficacy remains unconfirmed (Wen et al., 2014).
7.2 Bystander effect
NSCs reduce host cell death by secreting neurotrophic factors, boosting endogenous repair, promoting vascular regeneration, and suppressing inflammation a mechanism known as the bystander effect (Reynolds et al., 1992; Huang and Zhang, 2019). Specific pathways include downregulation of pro-inflammatory cytokines (e.g., IL-1β, IL-6, TNF-α) via NF-κB inhibition. NSC-derived exosomes, small vesicles containing RNA and proteins, may drive this effect, enhancing autophagy, reducing neuronal apoptosis, and curbing inflammation with their neurotrophic cargo (Rong et al., 2019). In a mouse ICH model, gene-modified NSCs overexpressing BDNF, GDNF, Akt1, and VEGF enhanced neurological recovery, likely by improving vascular regeneration, repairing the blood-brain barrier (BBB), and reducing edema (Gao et al., 2018) (Figure 2), though the precise mechanisms await further exploration (Pasi et al., 2021).
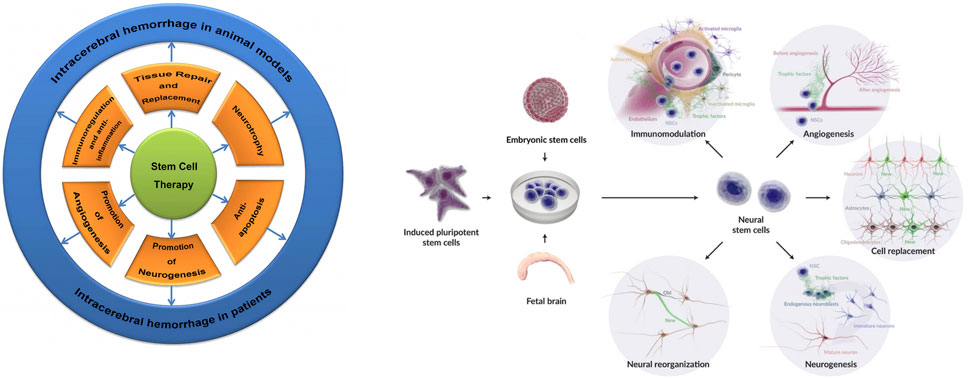
Figure 2. Stem cell and NSC therapeutic mechanisms in ICH: Illustrates protective roles including repair, replacement, neurotrophic support, anti-apoptosis, anti-inflammation, vascular regeneration, neurogenesis, immunomodulation, neural repair, differentiation into astrocytes/neurons/oligodendrocytes, and host tissue integration—culminating in enhanced tissue repair and neurological recovery.
The treatment of ICH shows limited effectiveness in reducing damage and improving the recovery of neurological function, often resulting in persistent severe neurological deficits. Given the complex molecular mechanisms underlying this pathology, exploring appropriate and effective treatment methods is of significant importance (Li et al., 2022). In animal models and patients, the therapeutic mechanisms of Stem Cells (SCs) in ICH involve tissue repair and replacement, neural nutrition, anti-apoptotic effects, anti-inflammatory properties, promotion of vascular regeneration, and neurogenesis (Gao et al., 2018). Transplantation of NSCs derived from ESCs, iPSCs, or fetal brain into veins, parenchyma, or brain ventricles has shown various therapeutic functions (Baker et al., 2019).
8 Basic research on NSCs therapy for hemorrhagic stroke
8.1 Immunomodulation and inflammatory response regulation by NSCs
Early studies linked post-ICH recovery from neural NSC transplantation to neuron replacement and network formation. Recent data emphasize inflammation’s pivotal role in stroke-induced brain damage, tying neuronal injury severity to immune activation (Lee et al., 2007). NSC-driven recovery may hinge more on immune modulation than cell replacement. In experimental autoimmune encephalomyelitis models, transplanted NSCs bolstered neuroprotection by curbing T-cell activity and triggering T-cell apoptosis (Pluchino et al., 2005). Post-stroke, activated microglia—brain-resident immune cells—release pro-inflammatory cytokines (e.g., IL-1β, IL-6, TNF-α) within minutes, amplifying damage by breaching the blood-brain barrier (BBB). NSCs likely dampen this cascade through a bystander effect, with studies affirming their anti-inflammatory, neuroprotective effects (Baker et al., 2019). This involves specific mediators like suppression of NF-κB and JAK-STAT pathways.
Genetically modified human NSCs (hNSCs) overexpressing BDNF, GDNF, Akt1, and VEGF markedly enhance neurological recovery in ICH mice (Zhou et al., 2022). Boosting post-stroke endogenous neurogenesis also shows promise. In ICH rats, repeated transcranial magnetic stimulation directed NSC differentiation toward neurons, minimizing glial-like outcomes and aiding functional recovery (Cui et al., 2019). NSC transplantation amplifies this by spurring endogenous NSC proliferation in the subventricular zone (SVZ) and dentate gyrus (DG), driving neural progenitor migration to damaged areas for neuronal differentiation. Though exact mechanisms remain elusive, neurotrophic and regenerative growth factors likely underpin tissue repair and immune-inflammatory suppression (Eriksson et al., 1998).
8.2 NSCs’ role in replacement and neurotrophic functions
NSCs differentiate into mature neurons and integrate into host brain tissue, functioning as cell replacement therapy while also delivering neurotrophic support. Transplanted NSCs express the neuroblast marker Doublecortin (DCX) within 2 months, with levels declining as they become neurons or glial cells expressing markers like NeuN, HuD, MAP2, and βIII-tubulin (Tornero et al., 2013). Their replacement role’s timing and scope remain uncertain (Baker et al., 2019). Animal studies show NSCs reduce secondary damage via anti-inflammatory effects, enhance endogenous neurogenesis and synaptic remodeling, and serve as cell substitutes (Baker et al., 2019). However, rapid spontaneous recovery in rodent models complicates distinguishing neurotrophic benefits from integration effects 2–3 months post-transplantation (Tornero et al., 2013). Whether NSC differentiation is driven by intrinsic cues, host environment, or occurs spontaneously requires further investigation.
8.3 NSCs’ promotion of angiogenesis
NSCs promote angiogenesis, driving neural repair through synaptic reorganization and neurogenesis. Post-stroke, nutrient and growth factor exchange between endothelial cells and migrating endogenous NSCs supports vascular regeneration. Rodent studies show transplanted NSCs boost microvessel density and angiogenic receptor expression in the ischemic penumbra, enhancing angiogenesis (Baker et al., 2019). This process hinges on VEGF signaling produced by NSCs or amplified by host tissue closely tied to neurological recovery (Ryu et al., 2016). Embryonic NSCs also express Ang1, increasing microvessel counts post-transplantation (Hicks et al., 2013). Effective vascular networks in the penumbra depend on neurovascular units (NVUs), which NSC therapy strengthens by upregulating tight junction proteins (claudin, occludin, ZO1) and dystroglycan, while reducing BBB leakage (Horie et al., 2011). However, while NSC therapy enhances angiogenesis via VEGF signaling, it may transiently exacerbate BBB leakage in the acute phase, potentially worsening edema. Studies suggest that balanced upregulation of tight junction proteins mitigates this risk (Horie et al., 2011), but further research is needed to optimize timing and dosing to prevent adverse effects.
9 Clinical trials
SC transplantation holds significant promise for stroke treatment, yet research predominantly targets ischemic stroke, with ICH receiving less attention and limited clinical trial validation. SC therapies for neuroregeneration are classified into three generations:
9.1 First generation
Utilizes adult stem cells hematopoietic stem cells (HSCs), mesenchymal stem cells (MSCs), and fetal-derived NSCs offering immunomodulation, anti-inflammatory, angiogenic, anti-apoptotic, and trophic benefits. These are primarily studied for autoimmune diseases, neurodegeneration, heart failure, skeletal disorders, and gastrointestinal conditions (Ratcliffe et al., 2013).
9.2 Second generation
Employs embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) for their unlimited self-renewal and pluripotency, with trials focusing on spinal cord injuries, retinal degeneration, type 1 diabetes, and ischemic heart failure (Kim et al., 2022).
9.3 Third generation
Enhances efficacy by engineering first- and second-generation SCs as drug carriers to optimize outcomes (Kim et al., 2022).
Only six SC transplantation trials for hemorrhagic stroke are registered on ClinicalTrials.gov: two completed, two ongoing, and two with unreported results. A South Korean phase I trial demonstrated that MSC transplantation for severe intraventricular hemorrhage in preterm infants reduced periventricular damage in some cases, with no deaths or severe adverse effects, suggesting safety and feasibility (Ahn et al., 2018). Chang et al. found that bone marrow or umbilical cord MSC transplantation in patients with moderate-to-severe ICH deficits (over 60 months) outperformed surgical hematoma evacuation alone in functional outcomes (Chang et al., 2016). Tsang et al.’s phase I/II randomized controlled trials showed autologous bone marrow MSC transplantation improved neurological recovery compared to placebo (Tsang et al., 2017). Conversely, the Chinese Clinical Trial Registry (www.chictr.org.cn) lists 16 NSC trials, mostly for ischemic stroke, including exosome therapy, cerebral palsy, stroke sequelae, and spinal cord injuries, conducted as single- or multi-center studies. SCs, often sourced from accessible umbilical cord or bone marrow, are typically administered late (weeks to over a year post ICH) rather than acutely, reflecting the priority of life-saving interventions in the initial phase. (See Table 3 for a summary of key clinical trials.).
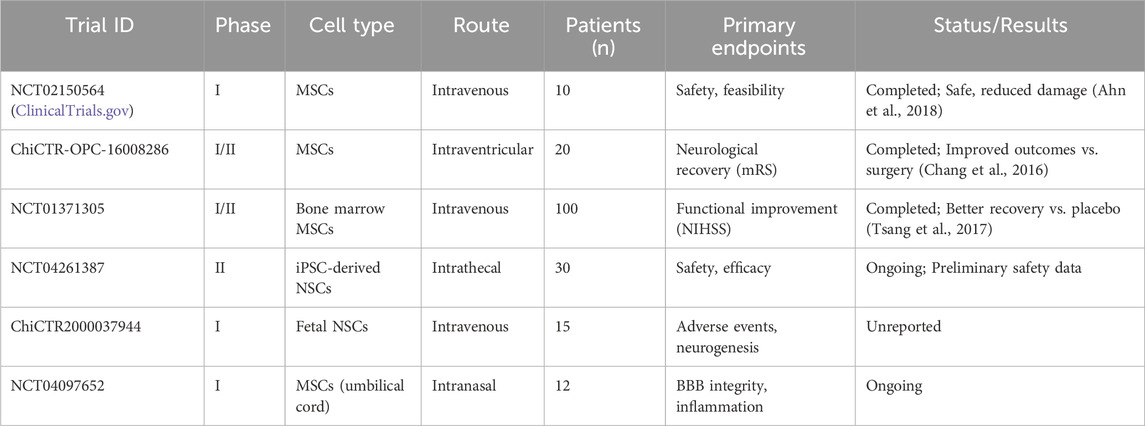
Table 3. Summary of key clinical trials on stem cell therapy for ICH.
ESCs and iPSCs can differentiate into specific cell types, but ESCs face ethical and immunogenicity challenges due to their embryonic origin (Kimbrel and Lanza, 2015), while iPSCs, developed to bypass these issues, require safety validation due to genetic modifications via viral vectors like lentiviruses and adenoviruses (Ratcliffe et al., 2013). NSCs excel in treating neurological diseases, differentiating into functional cells and releasing nerve growth factors to stimulate endogenous repair, though their deep brain location complicates harvesting. MSCs, widely studied for their ease of isolation from adult tissues, support regeneration through anti-inflammatory, angiogenic, and growth factor secretion (Ji et al., 2021). Advancing SC therapy for ICH requires integrating biomaterials, bioengineering, gene overexpression, and pretreatment strategies to boost endogenous regeneration (Kim et al., 2022). Despite promising efficacy, large-scale ICH trials and robust safety data are lacking (Kim et al., 2022) (Figure 3). Future research should optimize delivery methods, dosages, timing, and SC mass production feasibility.

Figure 3. Stem cell Types for ICH Treatment: This diagram displays stem cell types for treating ICH (parenchymal hemorrhage, intraventricular hemorrhage [IVH], and subarachnoid hemorrhage [SAH]), including NSCs, mesenchymal stem cells (MSCs), induced pluripotent stem cells (iPSCs), and embryonic stem cells (ESCs).
10 The promise and challenge of NSCs treatment
10.1 The application prospects of neural stem cells
Spontaneous intracerebral hemorrhage (SICH) poses a major threat to human health, with current medical and surgical treatments debated for their ability to improve long-term outcomes due to extensive neuronal damage that remains difficult to repair with existing technology. Preclinical studies, including animal and cell-based research, demonstrate that NSC transplantation offers promising therapeutic benefits for ICH. NSCs replace damaged neurons, reduce inflammation, and inhibit apoptosis, positioning this emerging therapy as a potentially effective solution for post ICH recovery (Zhou et al., 2022).
Endogenous NSCs provide a safe repair option, but their limited numbers and inefficient migration and differentiation necessitate future research into effective stimulants (Zhao et al., 2022). Intrinsic repair alone struggles to regenerate lost neurons, making exogenous NSC transplantation the dominant approach. Combining epidermal growth factor (EGF) and basic fibroblast growth factor (bFGF) in serum-free cultures rapidly produces abundant NSCs, ensuring a reliable supply (Schmidt et al., 2021). Studies show that pairing hematoma clearance with NSC transplantation outperforms standalone interventions, suggesting new treatment pathways (Zhang et al., 2015). While preclinical success drives expanding clinical trials—mostly for ischemic stroke—further trials are needed to confirm NSC efficacy in ICH (Kalladka et al., 2016).
10.2 The application challenges
NSC neuroprotective mechanisms, likely involving inflammation modulation and apoptosis suppression, remain incompletely understood and require deeper study. Current animal and in vitro models, often using healthy subjects, fail to fully replicate clinical ICH pathophysiology, highlighting the need for improved models. Challenges such as transplantation timing, methods, differentiation cues, immune responses, and ethical issues persist. Despite these uncertainties, NSC transplantation represents the most promising strategy for addressing ICH-related challenges.
10.3 Immunological and safety challenges
A critical barrier to NSC translation is immunological rejection and safety risks. While autologous NSCs minimize rejection, allogeneic transplants can trigger T-cell mediated immune responses, as reported in several studies (Pluchino et al., 2005). Tumorigenicity is a major concern, particularly with iPSC- or ESC-derived NSCs, due to potential teratoma formation from undifferentiated cells (Yang et al., 2017; Mollinari and Merlo, 2021). Mitigation strategies include CRISPR-based gene editing for immune evasion, pretreatment with immunosuppressants, and rigorous purity checks. Ethical issues (e.g., embryonic sourcing) and manufacturing scalability (e.g., GMP-compliant production) further complicate clinical adoption, requiring regulatory oversight from bodies like the FDA.
11 Conclusion and future perspectives
Post-ICH interventions, whether surgical or pharmacological, have reduced mortality, disability, and improved quality of life to some degree, yet persistent neurological deficits remain difficult to fully resolve. NSC transplantation is being explored as a potential solution, though it remains largely in the preclinical animal stage, with clinical use still distant. The complex mechanisms of neuroprotection, neurogenesis, and repair following ICH drive ongoing research, with preventing neuronal death via ferroptosis and necroptosis emerging as a key focus. NSCs’ strong immunotolerance and versatile differentiation capacity make them a promising therapy for hemorrhagic stroke. However, current NSC subtypes, mostly rat-derived, differ from human cells, and debates over transplantation timing, dosage, and mechanisms of induction, differentiation, and migration require further study to optimize strategies.
While NSC transplantation appears safe and feasible in many studies, challenges persist, including ethical concerns, efficacy, complications, immune rejection, and the critical risk of tumorigenesis. Long-term safety remains contentious, with uncertainties in controlling proliferation and differentiation. Clinical research lags behind preclinical promise, with trials limited to phases I and II and no large-scale phase III studies. Extensive future trials are essential to confirm the safety and efficacy of NSC transplantation for hemorrhagic stroke. The heterogeneity in parameters (e.g., doses of 10^5–10^8 cells, acute vs. chronic timing) contributes to inconsistent results, underscoring the need for standardized protocols to close the translational gap.
Author contributions
PS: Methodology, Writing – review and editing, Resources, Data curation, Writing – original draft. ZS: Methodology, Data curation, Conceptualization, Writing – original draft. HJ: Data curation, Formal Analysis, Writing – review and editing. QC: Project administration, Conceptualization, Resources, Writing – review and editing, Supervision, Funding acquisition.
Funding
The authors declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ahn, S. Y., Chang, Y. S., Sung, S. I., and Park, W. S. (2018). Mesenchymal stem cells for severe intraventricular hemorrhage in preterm infants: phase I dose-escalation clinical trial. Stem Cells Transl. Med. 7 (12), 847–856. doi:10.1002/sctm.17-0219
Alharbi, B. M., Tso, M. K., and Macdonald, R. L. (2016). Animal models of spontaneous intracerebral hemorrhage. Neurol. Res. 38 (5), 448–455. doi:10.1080/01616412.2016.1144671
Appel, L. J. (2014). Reducing sodium intake to prevent stroke: time for action, not hesitation. Stroke 45 (3), 909–911. doi:10.1161/STROKEAHA.113.002870
Auriat, A. M., Rosenblum, S., Smith, T. N., and Guzman, R. (2011). Intravascular stem cell transplantation for stroke. Transl. Stroke Res. 2 (3), 250–265. doi:10.1007/s12975-011-0093-1
Baker, E. W., Kinder, H. A., and West, F. D. (2019). Neural stem cell therapy for stroke: a multimechanistic approach to restoring neurological function. Brain Behav. 9 (3), e01214. doi:10.1002/brb3.1214
Cao, S., Zheng, M., Hua, Y., Chen, G., Keep, R. F., and Xi, G. (2016). Hematoma changes during clot resolution after experimental intracerebral hemorrhage. Stroke 47 (6), 1626–1631. doi:10.1161/STROKEAHA.116.013146
Ceto, S., Sekiguchi, K. J., Takashima, Y., Nimmerjahn, A., and Tuszynski, M. H. (2020). Neural stem cell grafts form extensive synaptic networks that integrate with host circuits after spinal cord injury. Cell Stem Cell 27 (3), 430–440.e5. doi:10.1016/j.stem.2020.07.007
Chang, Z., Mao, G., Sun, L., Ao, Q., Gu, Y., and Liu, Y. (2016). Cell therapy for cerebral hemorrhage: five year follow-up report. Exp. Ther. Med. 12 (6), 3535–3540. doi:10.3892/etm.2016.3811
Chen, Y., Wright, N., Guo, Y., Turnbull, I., Kartsonaki, C., Yang, L., et al. (2020). Mortality and recurrent vascular events after first incident stroke: a 9-year community-based study of 0.5 million Chinese adults. Lancet Glob. Health 8 (4), e580–e590. doi:10.1016/S2214-109X(20)30069-3
Cruz-Martinez, P., Gonzalez-Granero, S., Molina-Navarro, M. M., Pacheco-Torres, J., García-Verdugo, J. M., Geijo-Barrientos, E., et al. (2017). Corrigendum: intraventricular injections of mesenchymal stem cells activate endogenous functional remyelination in a chronic demyelinating murine model. Cell Death Dis. 8 (5), e2809. doi:10.1038/cddis.2016.340
Cui, M., Ge, H., Zeng, H., Yan, H., Zhang, L., Feng, H., et al. (2019). Repetitive transcranial magnetic stimulation promotes neural stem cell proliferation and differentiation after intracerebral hemorrhage in mice. Cell Transpl. 28 (5), 568–584. doi:10.1177/0963689719834870
Deinsberger, W., Vogel, J., Kuschinsky, W., Auer, L. M., and Böker, D. K. (1996). Experimental intracerebral hemorrhage: description of a double injection model in rats. Neurol. Res. 18 (5), 475–477. doi:10.1080/01616412.1996.11740456
Deshpande, K., Saatian, B., Martirosian, V., Lin, M., Julian, A., and Neman, J. (2019). Isolation of neural stem cells from whole brain tissues of adult mice. Curr. Protoc. Stem Cell Biol. 49 (1), e80. doi:10.1002/cpsc.80
Erharter, A., Rizzi, S., Mertens, J., and Edenhofer, F. (2019). Take the shortcut - direct conversion of somatic cells into induced neural stem cells and their biomedical applications. FEBS Lett. 593 (23), 3353–3369. doi:10.1002/1873-3468.13656
Eriksson, P. S., Perfilieva, E., Bjork-Eriksson, T., Alborn, A. M., Nordborg, C., Peterson, D. A., et al. (1998). Neurogenesis in the adult human hippocampus. Nat. Med. 4 (11), 1313–1317. doi:10.1038/3305
Gao, L., Xu, W., Li, T., Chen, J., Shao, A., Yan, F., et al. (2018). Stem cell therapy: a promising therapeutic method for intracerebral hemorrhage. Cell Transpl. 27 (12), 1809–1824. doi:10.1177/0963689718773363
Greenberg, S. M., Ziai, W. C., Cordonnier, C., Dowlatshahi, D., Francis, B., Goldstein, J. N., et al. (2022). 2022 guideline for the management of patients with spontaneous intracerebral hemorrhage: a guideline from the American heart association/american stroke Association. Stroke 53 (7), e282–e361. doi:10.1161/STR.0000000000000407
Hermann, A., Liebau, S., Gastl, R., Fickert, S., Habisch, H. J., Fiedler, J., et al. (2006). Comparative analysis of neuroectodermal differentiation capacity of human bone marrow stromal cells using various conversion protocols. J. Neurosci. Res. 83 (8), 1502–1514. doi:10.1002/jnr.20840
Hicks, C., Stevanato, L., Stroemer, R. P., Tang, E., Richardson, S., and Sinden, J. D. (2013). In vivo and in vitro characterization of the angiogenic effect of CTX0E03 human neural stem cells. Cell Transpl. 22 (9), 1541–1552. doi:10.3727/096368912X657936
Horie, N., Pereira, M. P., Niizuma, K., Sun, G., Keren-Gill, H., Encarnacion, A., et al. (2011). Transplanted stem cell-secreted vascular endothelial growth factor effects poststroke recovery, inflammation, and vascular repair. Stem Cells 29 (2), 274–285. doi:10.1002/stem.584
Huang, L., and Zhang, L. (2019). Neural stem cell therapies and hypoxic-ischemic brain injury. Prog. Neurobiol. 173, 1–17. doi:10.1016/j.pneurobio.2018.05.004
Ji, X. L., Ma, L., Zhou, W. H., and Xiong, M. (2021). Narrative review of stem cell therapy for ischemic brain injury. Transl. Pediatr. 10 (2), 435–445. doi:10.21037/tp-20-262
Jing, C., Bian, L., Wang, M., Keep, R. F., Xi, G., and Hua, Y. (2019). Enhancement of Hematoma clearance with CD47 blocking antibody in experimental intracerebral hemorrhage. Stroke 50 (6), 1539–1547. doi:10.1161/STROKEAHA.118.024578
Kalladka, D., Sinden, J., Pollock, K., Haig, C., McLean, J., Smith, W., et al. (2016). Human neural stem cells in patients with chronic ischaemic stroke (PISCES): a phase 1, first-in-man study. Lancet 388 (10046), 787–796. doi:10.1016/S0140-6736(16)30513-X
Kim, J. T., Youn, D. H., Kim, B. J., Rhim, J. K., and Jeon, J. P. (2022). Recent stem cell research on hemorrhagic stroke: an update. J. Korean Neurosurg. Soc. 65, 161–172. doi:10.3340/jkns.2021.0126
Kimbrel, E. A., and Lanza, R. (2015). Current status of pluripotent stem cells: moving the first therapies to the clinic. Nat. Rev. Drug Discov. 14 (10), 681–692. doi:10.1038/nrd4738
Lauer, A., Cianchetti, F. A., Van Cott, E. M., Schlunk, F., Schulz, E., Pfeilschifter, W., et al. (2011). Anticoagulation with the oral direct thrombin inhibitor dabigatran does not enlarge hematoma volume in experimental intracerebral hemorrhage. Circulation 124 (15), 1654–1662. doi:10.1161/CIRCULATIONAHA.111.035972
Lee, H. J., Kim, K. S., Kim, E. J., Choi, H. B., Lee, K. H., Park, I. H., et al. (2007). Brain transplantation of immortalized human neural stem cells promotes functional recovery in mouse intracerebral hemorrhage stroke model. Stem Cells 25 (5), 1204–1212. doi:10.1634/stemcells.2006-0409
Lendahl, U. (2022). 100 plus years of stem cell research-20 years of ISSCR. Stem Cell Rep. 17 (6), 1248–1267. doi:10.1016/j.stemcr.2022.04.004
Li, Z., Khan, S., Liu, Y., Wei, R., Yong, V. W., and Xue, M. (2022). Therapeutic strategies for intracerebral hemorrhage. Front. Neurol. 13, 1032343. doi:10.3389/fneur.2022.1032343
Liu, L., Wang, S., Xu, R., Zheng, J., Tang, J., Tang, X., et al. (2015). Experimental intracerebral haemorrhage: description of a semi-coagulated autologous blood model in rats. Neurol. Res. 37 (10), 874–879. doi:10.1179/1743132815Y.0000000067
Manaenko, A., Chen, H., Zhang, J. H., and Tang, J. (2011). Comparison of different preclinical models of intracerebral hemorrhage. Acta Neurochir. Suppl. 111, 9–14. doi:10.1007/978-3-7091-0693-8_2
Martinez-Galdamez, M., Biondi, A., Kalousek, V., Pereira, V. M., Ianucci, G., Gentric, J. C., et al. (2019). Periprocedural safety and technical outcomes of the new silk vista baby flow diverter for the treatment of intracranial aneurysms: results from a multicenter experience. J. Neurointerv Surg. 11 (7), 723–727. doi:10.1136/neurintsurg-2019-014770
Mitrecic, D., Hribljan, V., Jagecic, D., Isaković, J., Lamberto, F., Horánszky, A., et al. (2022). Regenerative neurology and regenerative cardiology: shared hurdles and achievements. Int. J. Mol. Sci. 23 (2), 855. doi:10.3390/ijms23020855
Mollinari, C., and Merlo, D. (2021). Direct reprogramming of somatic cells to neurons: pros and cons of chemical approach. Neurochem. Res. 46 (6), 1330–1336. doi:10.1007/s11064-021-03282-5
Ogata, J., Fujishima, M., Tamaki, K., Nakatomi, Y., Ishitsuka, T., and Omae, T. (1980). Stroke-prone spontaneously hypertensive rats as an experimental model of malignant hypertension. I. A light- and electron-microscopic study of the brain. Acta Neuropathol. 51 (3), 179–184. doi:10.1007/BF00687384
Paiva, W. S., Zippo, E., Miranda, C., Brasil, S., Godoy, D. A., De Andrade, A. F., et al. (2023). Animal models for the study of intracranial hematomas (review). Exp. Ther. Med. 25 (1), 20. doi:10.3892/etm.2022.11719
Pasi, M., Sugita, L., Xiong, L., Charidimou, A., Boulouis, G., Pongpitakmetha, T., et al. (2021). Association of cerebral small vessel disease and cognitive decline after intracerebral hemorrhage. Neurology 96 (2), e182–e192. doi:10.1212/WNL.0000000000011050
Petrelli, F., Scandella, V., Montessuit, S., Zamboni, N., Martinou, J. C., and Knobloch, M. (2023). Mitochondrial pyruvate metabolism regulates the activation of quiescent adult neural stem cells. Sci. Adv. 9 (9), eadd5220. doi:10.1126/sciadv.add5220
Pluchino, S., Zanotti, L., Rossi, B., Brambilla, E., Ottoboni, L., Salani, G., et al. (2005). Neurosphere-derived multipotent precursors promote neuroprotection by an immunomodulatory mechanism. Nature 436 (7048), 266–271. doi:10.1038/nature03889
Quanjun, Z., Jia, W., and Sun, Y. (2019). Culture and identification of autologous neural stem cells from abandoned brain tissue in patients during craniocerebral operation. Chin. J. Anat. 42 (02), 142–147. doi:10.3969/j.issn.1001-1633.2019.02.008
Ratcliffe, E., Glen, K. E., Naing, M. W., and Williams, D. J. (2013). Current status and perspectives on stem cell-based therapies undergoing clinical trials for regenerative medicine: case studies. Br. Med. Bull. 108, 73–94. doi:10.1093/bmb/ldt034
Reynolds, B. A., Tetzlaff, W., and Weiss, S. (1992). A multipotent EGF-responsive striatal embryonic progenitor cell produces neurons and astrocytes. J. Neurosci. 12 (11), 4565–4574. doi:10.1523/JNEUROSCI.12-11-04565.1992
Rodriguez-Frutos, B., Otero-Ortega, L., Gutierrez-Fernandez, M., Fuentes, B., Ramos-Cejudo, J., and Díez-Tejedor, E. (2016). Stem cell therapy and administration routes after stroke. Transl. Stroke Res. 7 (5), 378–387. doi:10.1007/s12975-016-0482-6
Rong, Y., Liu, W., Wang, J., Fan, J., Luo, Y., Li, L., et al. (2019). Neural stem cell-derived small extracellular vesicles attenuate apoptosis and neuroinflammation after traumatic spinal cord injury by activating autophagy. Cell Death Dis. 10 (5), 340. doi:10.1038/s41419-019-1571-8
Rosenberg, G. A., Mun-Bryce, S., Wesley, M., and Kornfeld, M. (1990). Collagenase-induced intracerebral hemorrhage in rats. Stroke 21 (5), 801–807. doi:10.1161/01.str.21.5.801
Ryu, S., Lee, S. H., Kim, S. U., and Yoon, B. W. (2016). Human neural stem cells promote proliferation of endogenous neural stem cells and enhance angiogenesis in ischemic rat brain. Neural Regen. Res. 11 (2), 298–304. doi:10.4103/1673-5374.177739
Schmidt, S. I., Bogetofte, H., Ritter, L., Agergaard, J. B., Hammerich, D., Kabiljagic, A. A., et al. (2021). Microglia-secreted factors enhance dopaminergic differentiation of Tissue- and iPSC-Derived human neural stem cells. Stem Cell Rep. 16 (2), 281–294. doi:10.1016/j.stemcr.2020.12.011
Sinar, E. J., Mendelow, A. D., Graham, D. I., and Teasdale, G. M. (1987). Experimental intracerebral hemorrhage: effects of a temporary mass lesion. J. Neurosurg. 66 (4), 568–576. doi:10.3171/jns.1987.66.4.0568
Sun, J., Wei, Z. Z., Gu, X., Zhang, J. Y., Zhang, Y., Li, J., et al. (2015). Intranasal delivery of hypoxia-preconditioned bone marrow-derived mesenchymal stem cells enhanced regenerative effects after intracerebral hemorrhagic stroke in mice. Exp. Neurol. 272, 78–87. doi:10.1016/j.expneurol.2015.03.011
Takahashi, K., and Yamanaka, S. (2006). Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126 (4), 663–676. doi:10.1016/j.cell.2006.07.024
Tang, Y., Yu, P., and Cheng, L. (2017). Current progress in the derivation and therapeutic application of neural stem cells. Cell Death Dis. 8 (10), e3108. doi:10.1038/cddis.2017.504
Tornero, D., Wattananit, S., Gronning Madsen, M., Koch, P., Wood, J., Tatarishvili, J., et al. (2013). Human induced pluripotent stem cell-derived cortical neurons integrate in stroke-injured cortex and improve functional recovery. Brain 136 (Pt 12), 3561–3577. doi:10.1093/brain/awt278
Tsang, K. S., Ng, C. P. S., Zhu, X. L., Wong, G. K. C., Lu, G., Ahuja, A. T., et al. (2017). Phase I/II randomized controlled trial of autologous bone marrow-derived mesenchymal stem cell therapy for chronic stroke. World J. Stem Cells 9 (8), 133–143. doi:10.4252/wjsc.v9.i8.133
Tuazon, J. P., Castelli, V., and Borlongan, C. V. (2019). Drug-like delivery methods of stem cells as biologics for stroke. Expert Opin. Drug Deliv. 16 (8), 823–833. doi:10.1080/17425247.2019.1645116
Vahidy, F. S., Rahbar, M. H., Zhu, H., Rowan, P. J., Bambhroliya, A. B., and Savitz, S. I. (2016). Systematic review and meta-analysis of bone marrow-derived mononuclear cells in animal models of ischemic stroke. Stroke 47 (6), 1632–1639. doi:10.1161/STROKEAHA.116.012701
Vierbuchen, T., Ostermeier, A., Pang, Z. P., Kokubu, Y., Südhof, T. C., and Wernig, M. (2010). Direct conversion of fibroblasts to functional neurons by defined factors. Nature 463 (7284), 1035–1041. doi:10.1038/nature08797
Wen, X., Wang, Y., Zhang, F., Zhang, X., Lu, L., Shuai, X., et al. (2014). In vivo monitoring of neural stem cells after transplantation in acute cerebral infarction with dual-modal MR imaging and optical imaging. Biomaterials 35 (16), 4627–4635. doi:10.1016/j.biomaterials.2014.02.042
Wu, H., Zhao, J., Fu, B., Yin, S., Song, C., Zhang, J., et al. (2017). Retinoic acid-induced upregulation of miR-219 promotes the differentiation of embryonic stem cells into neural cells. Cell Death Dis. 8 (7), e2953. doi:10.1038/cddis.2017.336
Xue, M., and Del Bigio, M. R. (2003). Comparison of brain cell death and inflammatory reaction in three models of intracerebral hemorrhage in adult rats. J. Stroke Cerebrovasc. Dis. 12 (3), 152–159. doi:10.1016/S1052-3057(03)00036-3
Yang, G. Y., Betz, A. L., Chenevert, T. L., Brunberg, J. A., and Hoff, J. T. (1994). Experimental intracerebral hemorrhage: relationship between brain edema, blood flow, and blood-brain barrier permeability in rats. J. Neurosurg. 81 (1), 93–102. doi:10.3171/jns.1994.81.1.0093
Yang, H., Zhang, L., An, J., Zhang, Q., Liu, C., He, B., et al. (2017). MicroRNA-Mediated reprogramming of somatic cells into neural stem cells or neurons. Mol. Neurobiol. 54 (2), 1587–1600. doi:10.1007/s12035-016-0115-9
Yu, Z., Chen, L. F., Tang, L., and Hu, C. L. (2013). Effects of recombinant adenovirus-mediated hypoxia-inducible factor-1alpha gene on proliferation and differentiation of endogenous neural stem cells in rats following intracerebral hemorrhage. Asian Pac J. Trop. Med. 6 (10), 762–767. doi:10.1016/S1995-7645(13)60134-0
Zhang, Q., Shang, X., Hao, M., Zheng, M., Li, Y., Liang, Z., et al. (2015). Effects of human umbilical cord mesenchymal stem cell transplantation combined with minimally invasive hematoma aspiration on intracerebral hemorrhage in rats. Am. J. Transl. Res. 7 (11), 2176–2186.
Zhang, H., Huang, Z., Xu, Y., and Zhang, S. (2006). Differentiation and neurological benefit of the mesenchymal stem cells transplanted into the rat brain following intracerebral hemorrhage. Neurol. Res. 28 (1), 104–112. doi:10.1179/016164106X91960
Zhao, T., Zhu, T., Xie, L., Li, Y., Xie, R., Xu, F., et al. (2022). Neural stem cells therapy for ischemic stroke: progress and challenges. Transl. Stroke Res. 13 (5), 665–675. doi:10.1007/s12975-022-00984-y
Zhou, X., Chen, L., Feng, C., Tang, J., Liu, A., et al. (2013). Establishing an animal model of intracerebral hemorrhage under the guidance of ultrasound. Ultrasound Med. Biol. 39 (11), 2116–2122. doi:10.1016/j.ultrasmedbio.2013.05.021
Keywords: intracerebral hemorrhage, neural stem cells, transplantation, neural repair, treatment
Citation: Song P, Shafiq Z, Jiang H and Cai Q (2025) Application of neural stem cells in the treatment of intracerebral hemorrhage: a systematic review. Front. Pharmacol. 16:1660614. doi: 10.3389/fphar.2025.1660614
Received: 06 July 2025; Accepted: 20 November 2025;
Published: 04 December 2025.
Edited by:
Hansen Chen, Stanford University, United StatesReviewed by:
Danpeng Shen, Shanghai Jiao Tong University, ChinaHongjian Pu, University of Pittsburgh, United States
Qiaoshi Jiang, Stanford University, United States
Copyright © 2025 Song, Shafiq, Jiang and Cai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Qiang Cai, Y3Fub0BzaW5hLmNvbQ==
†These authors share first authorship