 Céline S. Reinbold1,2†
Céline S. Reinbold1,2† Andreas J. Forstner1,2,3,4,5†
Andreas J. Forstner1,2,3,4,5† Julian Hecker6
Julian Hecker6 Janice M. Fullerton7,8
Janice M. Fullerton7,8 Per Hoffmann1,2,3,4Liping Hou9
Per Hoffmann1,2,3,4Liping Hou9 Urs Heilbronner10
Urs Heilbronner10 Franziska Degenhardt3,4
Franziska Degenhardt3,4 Mazda Adli11
Mazda Adli11 Kazufumi Akiyama12Nirmala Akula9Raffaella Ardau13Bárbara Arias14Lena Backlund15Antonio Benabarre16Susanne Bengesser17Abesh K. Bhattacharjee18
Kazufumi Akiyama12Nirmala Akula9Raffaella Ardau13Bárbara Arias14Lena Backlund15Antonio Benabarre16Susanne Bengesser17Abesh K. Bhattacharjee18 Joanna M. Biernacka19,20Armin Birner17
Joanna M. Biernacka19,20Armin Birner17 Cynthia Marie-Claire21,22Pablo Cervantes23Guo-Bo Chen24
Cynthia Marie-Claire21,22Pablo Cervantes23Guo-Bo Chen24 Hsi-Chung Chen25Caterina Chillotti13Scott R. Clark26
Hsi-Chung Chen25Caterina Chillotti13Scott R. Clark26 Francesc Colom27David A. Cousins28Cristiana Cruceanu29
Francesc Colom27David A. Cousins28Cristiana Cruceanu29 Piotr M. Czerski30
Piotr M. Czerski30 Alexandre Dayer31
Alexandre Dayer31 Bruno Étain21,22,32
Bruno Étain21,22,32 Peter Falkai33Louise Frisén15Sébastien Gard34Julie S. Garnham35
Peter Falkai33Louise Frisén15Sébastien Gard34Julie S. Garnham35 Fernando S. Goes36Paul Grof37Oliver Gruber38
Fernando S. Goes36Paul Grof37Oliver Gruber38 Ryota Hashimoto39Joanna Hauser30Stefan Herms1,2,3,4
Ryota Hashimoto39Joanna Hauser30Stefan Herms1,2,3,4 Stéphane Jamain40Esther Jiménez16Jean-Pierre Kahn41Layla Kassem9
Stéphane Jamain40Esther Jiménez16Jean-Pierre Kahn41Layla Kassem9 Sarah Kittel-Schneider42Sebastian Kliwicki43Barbara König44
Sarah Kittel-Schneider42Sebastian Kliwicki43Barbara König44 Ichiro Kusumi45Nina Lackner17Gonzalo Laje9Mikael Landén46,47Catharina Lavebratt15Marion Leboyer48Susan G. Leckband49Carlos A. López Jaramillo50Glenda MacQueen51
Ichiro Kusumi45Nina Lackner17Gonzalo Laje9Mikael Landén46,47Catharina Lavebratt15Marion Leboyer48Susan G. Leckband49Carlos A. López Jaramillo50Glenda MacQueen51 Mirko Manchia52,53Lina Martinsson54
Mirko Manchia52,53Lina Martinsson54 Manuel Mattheisen55
Manuel Mattheisen55 Michael J. McCarthy56
Michael J. McCarthy56 Susan L. McElroy57
Susan L. McElroy57 Marina Mitjans14Francis M. Mondimore36Palmiero Monteleone58,59
Marina Mitjans14Francis M. Mondimore36Palmiero Monteleone58,59 Caroline M. Nievergelt18Urban Ösby60Norio Ozaki61Roy H. Perlis62Andrea Pfennig
Caroline M. Nievergelt18Urban Ösby60Norio Ozaki61Roy H. Perlis62Andrea Pfennig 63Daniela Reich-Erkelenz10
63Daniela Reich-Erkelenz10 Guy A. Rouleau64
Guy A. Rouleau64 Peter R. Schofield8,65K. Oliver Schubert26Barbara W. Schweizer36Florian Seemüller33
Peter R. Schofield8,65K. Oliver Schubert26Barbara W. Schweizer36Florian Seemüller33 Giovanni Severino66Tatyana Shekhtman18Paul D. Shilling18Kazutaka Shimoda67
Giovanni Severino66Tatyana Shekhtman18Paul D. Shilling18Kazutaka Shimoda67 Christian Simhandl68,69Claire M. Slaney35
Christian Simhandl68,69Claire M. Slaney35 Jordan W. Smoller62
Jordan W. Smoller62 Alessio Squassina66Thomas J. Stamm11,70Pavla Stopkova71 Sarah K. Tighe72
Alessio Squassina66Thomas J. Stamm11,70Pavla Stopkova71 Sarah K. Tighe72 Alfonso Tortorella59
Alfonso Tortorella59 Gustavo Turecki29Julia Volkert42Stephanie H. Witt73Adam J. Wright74L. Trevor Young75
Gustavo Turecki29Julia Volkert42Stephanie H. Witt73Adam J. Wright74L. Trevor Young75 Peter P. Zandi76James B. Potash72J. Raymond DePaulo36Michael Bauer63
Peter P. Zandi76James B. Potash72J. Raymond DePaulo36Michael Bauer63 Eva Reininghaus17
Eva Reininghaus17 Tomáš Novák71Jean-Michel Aubry31Mario Maj59
Tomáš Novák71Jean-Michel Aubry31Mario Maj59 Bernhard T. Baune26
Bernhard T. Baune26 Philip B. Mitchell74Eduard Vieta16
Philip B. Mitchell74Eduard Vieta16 Mark A. Frye20
Mark A. Frye20 Janusz K. Rybakowski43
Janusz K. Rybakowski43 Po-Hsiu Kuo77
Po-Hsiu Kuo77 Tadafumi Kato78
Tadafumi Kato78 Maria Grigoroiu-Serbanescu79Andreas Reif42Maria Del Zompo66Frank Bellivier21,22,32Martin Schalling15
Maria Grigoroiu-Serbanescu79Andreas Reif42Maria Del Zompo66Frank Bellivier21,22,32Martin Schalling15 Naomi R. Wray24
Naomi R. Wray24 John R. Kelsoe18
John R. Kelsoe18 Martin Alda35,71
Martin Alda35,71 Francis J. McMahon9
Francis J. McMahon9 Thomas G. Schulze9,10,36,38,73
Thomas G. Schulze9,10,36,38,73 Marcella Rietschel73
Marcella Rietschel73 Markus M. Nöthen3,4
Markus M. Nöthen3,4 Sven Cichon1,2,3,4,80*
Sven Cichon1,2,3,4,80*- 1Human Genomics Research Group, Department of Biomedicine, University of Basel, Basel, Switzerland
- 2Institute of Medical Genetics and Pathology, University Hospital Basel, Basel, Switzerland
- 3Institute of Human Genetics, University of Bonn, Bonn, Germany
- 4Department of Genomics, Life & Brain Center, University of Bonn, Bonn, Germany
- 5Department of Psychiatry (UPK), University of Basel, Basel, Switzerland
- 6Department of Biostatistics, Harvard T.H. Chan School of Public Health, Boston, MA, United States
- 7Neuroscience Research Australia, Sydney, NSW, Australia
- 8School of Medical Sciences, University of New South Wales, Sydney, NSW, Australia
- 9Intramural Research Program, US Department of Health & Human Services, National Institute of Mental Health, National Institutes of Health, Bethesda, MD, United States
- 10Department Psychiatry and Psychotherapy, Institute of Psychiatric Phenomics and Genomics, University Hospital, LMU Munich, Munich, Germany
- 11Department of Psychiatry and Psychotherapy, Charité Universitätsmedizin, Berlin, Germany
- 12Department of Biological Psychiatry and Neuroscience, Dokkyo Medical University School of Medicine, Mibu, Japan
- 13Unit of Clinical Pharmacology, University Hospital of Cagliari, Cagliari, Italy
- 14Zoology and Biological Anthropology Section (Department of Evolutive Biology, Ecology and Environmental Sciences), Facultat de Biologia and Institut de Biomedicina, CIBERSAM, Universitat de Barcelona, Barcelona, Spain
- 15Department of Molecular Medicine and Surgery, Karolinska Institutet and Center for Molecular Medicine, Karolinska University Hospital, Stockholm, Sweden
- 16Bipolar Disorder Program, Institute of Neuroscience, Hospital Clinic, CIBERSAM, IDIBAPS, University of Barcelona, Barcelona, Spain
- 17Special Outpatient Center for Bipolar Affective Disorder, Medical University of Graz, Graz, Austria
- 18Health Sciences Research, Mayo Clinic, Rochester, MN, United States
- 19Department of Psychiatry and Psychology, Mayo Clinic, Rochester, MN, United States
- 20Institut National de la Santé et de la Recherche Médicale, U1144, Paris, France
- 21Université Paris Diderot, Sorbonne Paris Cité, UMR-S 1144, Paris, France
- 22Department of Psychiatry, University of California, San Diego, San Diego, CA, United States
- 23The Neuromodulation Unit, McGill University Health Centre, Montreal, QC, Canada
- 24Queensland Brain Institute, The University of Queensland, Brisbane, QLD, Australia
- 25Department of Psychiatry & Center of Sleep Disorders, National Taiwan University Hospital, Taipei, Taiwan
- 26Discipline of Psychiatry, University of Adelaide, Adelaide, SA, Australia
- 27Mental Health Research Group, IMIM-Hospital del Mar, Barcelona, Spain
- 28Campus for Ageing and Vitality, Newcastle University, Newcastle upon Tyne, United Kingdom
- 29Douglas Mental Health University Institute, McGill University, Montreal, QC, Canada
- 30Psychiatric Genetic Unit, Poznan University of Medical Sciences, Poznan, Poland
- 31Mood Disorders Unit, HUG - Geneva University Hospitals, Geneva, Switzerland
- 32AP-HP, GH Saint-Louis – Lariboisière – F. Widal, Département de Psychiatrie et de Médecine Addictologique, Paris, France
- 33Department of Psychiatry and Psychotherapy, Ludwig-Maximilians-University Munich, Munich, Germany
- 34Service de Psychiatrie, Hôpital Charles Perrens, Bordeaux, France
- 35Department of Psychiatry, Dalhousie University, Halifax, NS, Canada
- 36Department of Psychiatry and Behavioral Sciences, Johns Hopkins University, Baltimore, MD, United States
- 37Mood Disorders Center of Ottawa, Ottawa, ON, Canada
- 38Department of Psychiatry and Psychotherapy, University Medical Center, Georg-August University Göttingen, Göttingen, Germany
- 39Molecular Research Center for Children's Mental Development, United Graduate School of Child Development, Osaka University, Osaka, Japan
- 40Institut National de la Santé et de la Recherche Médicale U955, Psychiatrie Translationnelle, Créteil, France
- 41Service de Psychiatrie et Psychologie Clinique, Centre Psychothérapique de Nancy - Université de Lorraine, Nancy, France
- 42Department of Psychiatry, Psychosomatic Medicine and Psychotherapy, University Hospital Frankfurt, Frankfurt, Germany
- 43Department of Adult Psychiatry, Poznan University of Medical Sciences, Poznan, Poland
- 44Department of Psychiatry and Psychotherapeuthic Medicine, Landesklinikum Neunkirchen, Neunkirchen, Austria
- 45Department of Psychiatry, Hokkaido University Graduate School of Medicine, Sapporo, Japan
- 46Institute of Neuroscience and Physiology, The Sahlgrenska Academy at the Gothenburg University, Gothenburg, Sweden
- 47Department of Medical Epidemiology and Biostatistics, Karolinska Institutet, Stockholm, Sweden
- 48Assistance Publique-Hôpitaux de Paris, Hôpital Albert Chenevier - Henri Mondor, Pôle de Psychiatrie, Créteil, France
- 49Department of Pharmacy, VA San Diego Healthcare System, San Diego, CA, United States
- 50Department of Psychiatry, University of Antioquia, Medellín, Medellín, Colombia
- 51Department of Psychiatry, University of Calgary, Calgary, AB, Canada
- 52Section of Psychiatry, Department of Medical Sciences and Public Health, University of Cagliari, Cagliari, Italy
- 53Department of Pharmacology, Dalhousie University, Halifax, NS, Canada
- 54Department of Clinical Neurosciences, Karolinska Institutet, Stockholm, Sweden
- 55Department of Biomedicine, Aarhus University, Aarhus, Denmark
- 56Department of Psychiatry, VA San Diego Healthcare System, San Diego, CA, United States
- 57Department of Psychiatry, Lindner Center of Hope/University of Cincinnati, Mason, OH, United States
- 58Neurosciences Section, Department of Medicine and Surgery, University of Salerno, Salerno, Italy
- 59Department of Psychiatry, University of Campania “Luigi Vanvitelli”, Naples, Italy
- 60Department of Neurobiology, Care Sciences, and Society, Karolinska Institutet and Center for Molecular Medicine, Karolinska University Hospital, Stockholm, Sweden
- 61Department of Psychiatry, Nagoya University Graduate School of Medicine, Nagoya, Japan
- 62Department of Psychiatry, Massachusetts General Hospital and Harvard Medical School, Boston, MA, United States
- 63Department of Psychiatry and Psychotherapy, Medical Faculty, University Hospital Carl Gustav Carus, Technische Universität Dresden, Dresden, Germany
- 64Department of Neurology and Neurosurgery, Montreal Neurological Institute and Hospital, McGill University, Montreal, QC, Canada
- 65Mental Illness, Neuroscience Research Australia, Sydney, NSW, Australia
- 66Department of Biomedical Sciences, University of Cagliari, Cagliari, Italy
- 67Department of Psychiatry, Dokkyo Medical University School of Medicine, Mibu, Japan
- 68Medical school, Sigmund Freud University, Vienna, Austria
- 69Bipolar Center Wiener Neustadt, Vienna, Austria
- 70Department of Psychiatry, Psychotherapy and Psychosomatics, Medical School Brandenburg, Neuruppin, Germany
- 71Department of Psychiatry, National Institute of Mental Health, Klecany, Czechia
- 72Department of Psychiatry, University of Iowa, Iowa, IA, United States
- 73Department of Genetic Epidemiology in Psychiatry, Medical Faculty Mannheim, Central Institute of Mental Health, University of Heidelberg, Mannheim, Germany
- 74School of Psychiatry, University of New South Wales, and Black Dog Institute, Sydney, NSW, Australia
- 75Department of Psychiatry, University of British Columbia, Vancouver, BC, Canada
- 76Department of Mental Health, Johns Hopkins Bloomberg School of Public Health, Baltimore, MD, United States
- 77Department of Public Health and Institute of Epidemiology and Preventive Medicine, National Taiwan University, Taipei, Taiwan
- 78Laboratory for Molecular Dynamics of Mental Disorders, RIKEN Brain Science Institute, Saitama, Japan
- 79Biometric Psychiatric Genetics Research Unit, Alexandru Obregia Clinical Psychiatric Hospital, Bucharest, Romania
- 80Research Centre Jülich, Institute of Neuroscience and Medicine (INM-1), Jülich, Germany
Bipolar disorder (BD) is a common, highly heritable neuropsychiatric disease characterized by recurrent episodes of mania and depression. Lithium is the best-established long-term treatment for BD, even though individual response is highly variable. Evidence suggests that some of this variability has a genetic basis. This is supported by the largest genome-wide association study (GWAS) of lithium response to date conducted by the International Consortium on Lithium Genetics (ConLiGen). Recently, we performed the first genome-wide analysis of the involvement of miRNAs in BD and identified nine BD-associated miRNAs. However, it is unknown whether these miRNAs are also associated with lithium response in BD. In the present study, we therefore tested whether common variants at these nine candidate miRNAs contribute to the variance in lithium response in BD. Furthermore, we systematically analyzed whether any other miRNA in the genome is implicated in the response to lithium. For this purpose, we performed gene-based tests for all known miRNA coding genes in the ConLiGen GWAS dataset (n = 2,563 patients) using a set-based testing approach adapted from the versatile gene-based test for GWAS (VEGAS2). In the candidate approach, miR-499a showed a nominally significant association with lithium response, providing some evidence for involvement in both development and treatment of BD. In the genome-wide miRNA analysis, 71 miRNAs showed nominally significant associations with the dichotomous phenotype and 106 with the continuous trait for treatment response. A total of 15 miRNAs revealed nominal significance in both phenotypes with miR-633 showing the strongest association with the continuous trait (p = 9.80E-04) and miR-607 with the dichotomous phenotype (p = 5.79E-04). No association between miRNAs and treatment response to lithium in BD in either of the tested conditions withstood multiple testing correction. Given the limited power of our study, the investigation of miRNAs in larger GWAS samples of BD and lithium response is warranted.
Introduction
Bipolar disorder (BD) is a severe neuropsychiatric condition categorized by recurrent episodes of depression and mania. BD is common, with a lifetime prevalence of around 1% in the general population (1). The elevated morbidity and mortality, the typically early age at onset in young adulthood and the chronic course of BD make it a major public health problem, and BD is classified as one of the top 25 leading causes of the global burden of disease (2). Epidemiological and molecular genetic data strongly suggest that BD is a complex disorder (3) which means that both genetic and environmental factors influence illness risk. Based on twin studies the overall heritability of BD has been estimated to be over 70% (4, 5), suggesting a substantial involvement of genetic factors in the development of the disease.
Mood stabilizers are used as the first-line mode of medication in the treatment of BD (6). Amongst these drugs, lithium is used as a preventive agent for manic and depressive episodes (7), suicide attempts, and death by suicide, and shows the greatest support for long-term relapse prevention (8, 9). Consequently, lithium is endorsed as a first-line and best-established long-term treatment for BD, even though individual response is highly variable (6, 8, 10). Evidence suggests that some of the variability in lithium response has a genetic basis (11, 12). This hypothesis is supported by the largest genome-wide association study (GWAS) of lithium response to date, which was conducted by the International Consortium on Lithium Genetics (ConLiGen) (13, 14). The study investigated genomic data of 2,563 BD patients, identifying a genome-wide significant locus on chromosome 21, which contains two long, non-coding RNA genes (lncRNAs) (14).
Non-coding RNAs (ncRNAs) are transcribed from DNA but do not encode protein, and are involved in complex mechanisms of gene regulation, particularly in fine regulation of the timing and level of expression of their target genes. Another class of ncRNAs whose role in the pathophysiology of psychiatric disorders is emerging, is that of microRNAs (miRNAs). miRNAs are short RNA molecules, which in the mature processed form are 21 to 25-nucleotides in length, that work as post-transcriptional regulators of gene expression (15). To create a mature miRNA, a primary miRNA (pri-miRNA, typically >1,000 nucleotides in length) is first transcribed, and forms a secondary structure through self-base pairing (16, 17). This is cleaved by the Drosha-DiGeorge syndrome critical region gene 8 (Drosha-DGCR8) complex to create a pre-miRNA of around 70 nucleotides (16). This double stranded RNA is exported from the nucleus, cleaved by Dicer-transactivation-responsive RNA-binding protein (Dicer-TRBP) to form the mature miRNA (16), which can then target complementary messenger RNA (mRNA) transcripts through the RNA-induced silencing complex (RISC) to regulate expression (e.g., via mRNA degradation or translational repression) (18). Several studies have reported that miRNAs are potential predictors of treatment response in complex genetic disorders (19–21) including lithium response in BD (22). Furthermore, miRNAs are implicated in biological pathways that regulate brain development and synaptic plasticity (23, 24). Indeed, miR-137 has emerged as a key risk gene in schizophrenia, and is known to regulate the expression of several genes that are independently associated with schizophrenia (25, 26). This implies the potential involvement of miRNAs in the pathogenesis of psychiatric disorders including BD. This hypothesis is further supported by the results of a large GWAS of BD (27) where a single-nucleotide polymorphism (SNP) flanking miR-2113 was amongst the strongest findings.
Our group performed the first genome-wide analysis of the involvement of miRNAs in the development of BD, in a sample of 9,747 patients and 14,278 controls (28) in which we identified nine BD-associated miRNAs that withstood stringent Bonferroni-correction for multiple testing. However, it is largely unknown whether these miRNAs are also associated with lithium response in BD.
Therefore, the aim of the present study was to determine whether common variants at any of the nine BD-associated miRNAs contribute to the variance in lithium response in BD. Furthermore, we systematically analyzed whether any other miRNA is implicated in the response to lithium. For this purpose, we performed window-based association testing for all known miRNA coding genes in the largest GWAS dataset of lithium response so far.
Materials and Methods
Sample Description
Analyses were performed using summary statistics from the previously published GWAS of lithium response in BD patients (n = 2,563 patients) (14). These GWAS datasets were collected by ConLiGen and combine imputed genotype data from 22 contributing sites from four continents (Europe, America, Asia, and Australia). The study was approved by the respective local ethics committees. Written informed consent was obtained from all participants prior to inclusion. The Alda scale was used to create a dichotomous (good vs. poor response to lithium) and a continuous measure (range 0–10) for the evaluation of long-term treatment response to lithium. Briefly, the Alda scale measures symptom improvement in the course of lithium treatment (A score, range 0–10), which is then weighted against five criteria (B score) that assess confounding factors, each scored 0, 1, or 2. The total score is calculated by subtracting the total B score from the A score. Negative scores are set to 0 by default leading to a total score range from 0 to 10. For the purpose of the present analysis, subjects with a total score of 7 or higher were defined as showing “good response” to lithium treatment in the dichotomous phenotype. As continuous measurement, we used the A score, but excluded all individuals with a total B score greater than 4, as continuous measure (14, 29).
Definition of Candidate and Genome-Wide miRNAs
Information on the nine BD-associated miRNAs was obtained from our previously published genome-wide analysis of miRNAs in BD (28). The chromosomal positions of the miRNAs were obtained from the miRBase database (release 21) (30).
For the genome-wide miRNA association analysis chromosomal positions for all 1,871 remaining miRNAs were obtained from miRBase (release 21). miRNAs which were not located on autosomal chromosomes (n = 120) were removed from further analysis. Only miRNA genes which were covered in the summary statistics of lithium response were included, resulting in 1,692 miRNAs which were tested in the genome-wide analysis. For each gene, the entire preprocessed transcript ± 20 kilobase (kb) flanking sequence were analyzed, which would include the majority of the regulatory regions (17).
miRNA-Based Association Tests
For the gene-based tests, we applied a set-based testing approach adapted from the versatile gene-based test for GWAS (VEGAS2) (31, 32) with a minor correction for the top-0.1-test option (33). This algorithm is obtainable upon request. The top-0.1-test was used since it showed the highest sensitivity with less than 1% false positives across a variety of investigated gene-level methods (34). The applied statistical algorithm is described in more detail in the article by Mishra and Macgregor (32). Briefly, we grouped SNPs within the miRNA loci ± 20 kb flanking sequence together and calculated a set-based test statistic as the sum of the χ2 one degree of freedom association P-values within the miRNA. The observed test statistic was compared with simulated test statistics from the multivariate normal distribution with correlation equal to the corresponding LD structure as derived from the 1,000 Genomes phase 3 European population genotypes (35, 36). We calculated an empirical miRNA-based P-value as the proportion of simulated test statistics above the observed test statistic. For the purposes of the present study, we used the top-0.1-test option which summarizes the 10% most significant SNPs for each miRNA.
Using the two summary statistics, miRNA-based P-values were calculated for all miRNAs. The calculated miRNA-based P-values were corrected for multiple testing according to Benjamini-Hochberg.
Enrichment Tests
To test whether nominally significant SNPs were enriched within miRNAs and their flanking regions, we conducted the Fisher's Exact Test for each summary statistic separately. Additionally, we tested whether the number of cis-miR-eQTL SNPs identified by Huan et al. (37) with a p-value of < 0.05 was higher than expected using the Fisher's Exact Test.
Results
Of the nine tested BD-associated miRNAs, miR-499a showed nominally significant P-values in both datasets (dichotomous and continuous treatment response, Table 1). Of the remaining 1,692 miRNAs tested for the genome-wide miRNA analysis, 71 miRNAs showed nominally significant associations with the dichotomous and 106 with the continuous treatment response. Fifteen miRNAs revealed nominal significance with both phenotypes. miR-633 showed the strongest association with the continuous phenotype (p = 9.80E-04). Regarding the dichotomous phenotype, miR-607 showed the strongest association (p = 5.79E-04). No association between miRNAs and treatment response to lithium in BD in either of the tested conditions withstood multiple testing correction (Tables 1, 2).
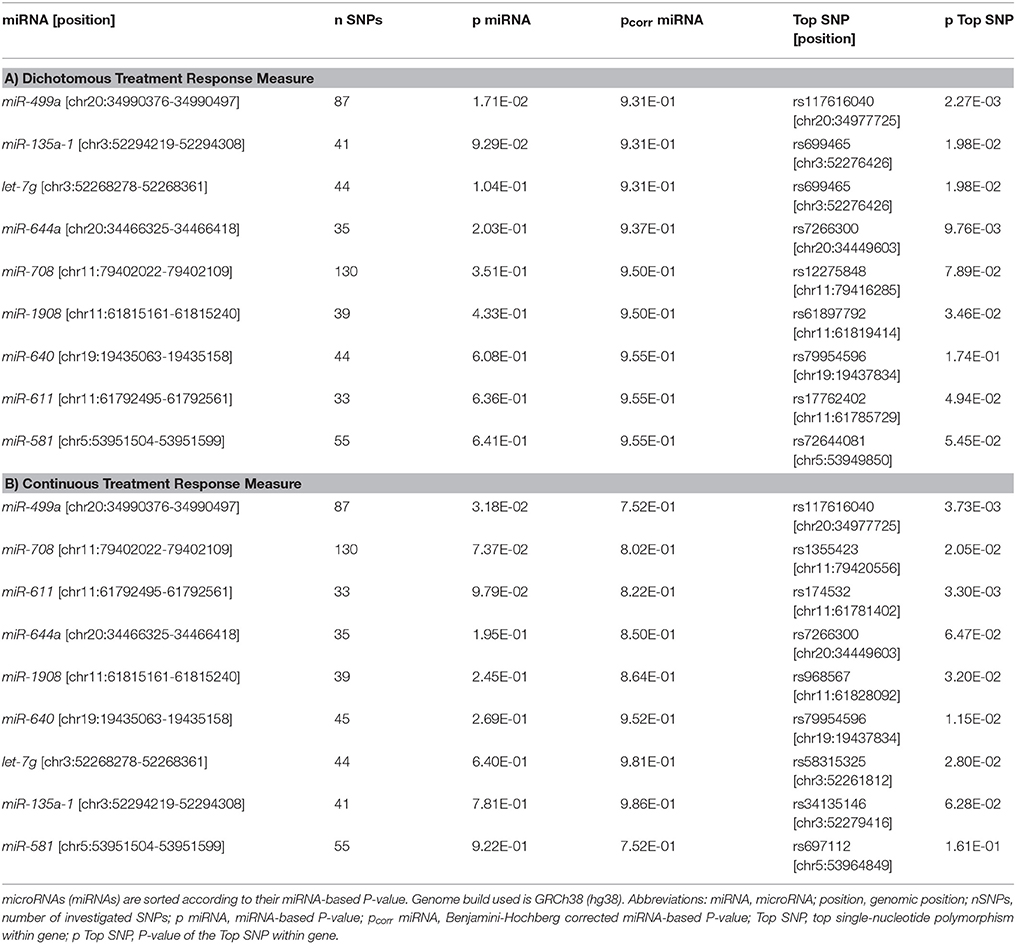
Table 1. Results of the window-based tests for the nine BD-associated miRNAs.
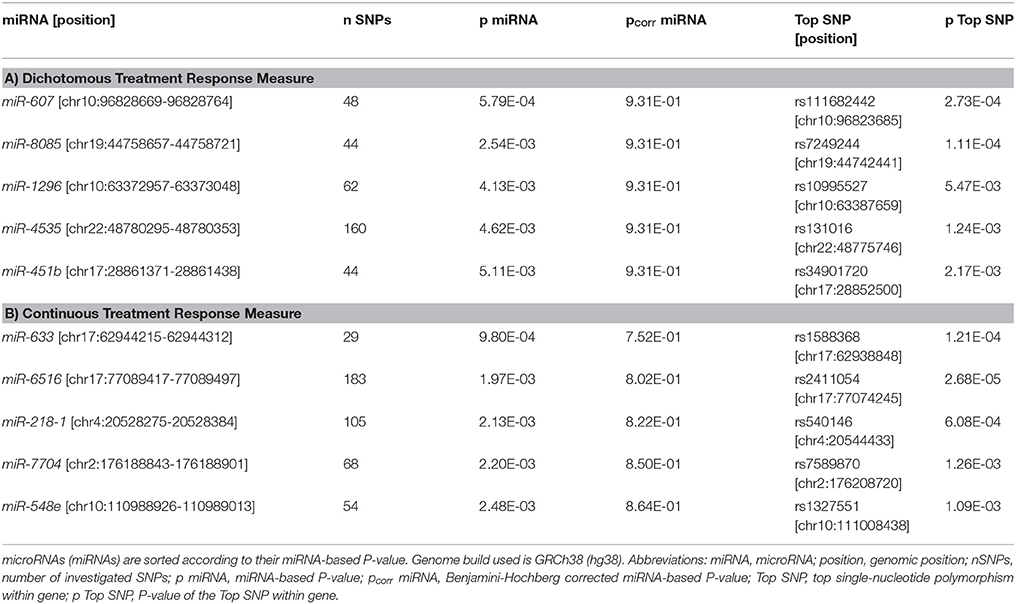
Table 2. Results of the window-based tests for the top five genome-wide miRNAs.
The number of nominally significant SNPs in both of our GWAS of lithium response located at miRNA loci (n = 6,321 and n = 5,742 for continuous and dichotomous measurement, respectively) was not significantly higher than expected (p = 9.96E-01 and p = 1 for continuous and dichotomous measurement, respectively, Fisher's Exact Test).
The number of cis-miR-eQTL SNPs [identified by Huan et al. (37) in our summary statistics (n = 341 and n = 318 for continuous and dichotomous measurement, respectively) were significantly higher than expected (p = 3.31E-05 and p = 6.23E-03 for continuous and dichotomous measurement, respectively, Fisher's Exact Test).
Discussion
The current study investigated whether common variants at BD-associated miRNA, or any other miRNA loci, contribute to the differences in lithium response in BD patients.
miR-499a showed a nominally significant association with lithium response in the candidate approach. Although the association did not withstand correction for multiple testing, this result provides some evidence that miR-499a might be involved in both development and treatment of BD. A previous study has shown an upregulation of this miRNA in the prefrontal cortex of patients with depression (38). In another study, miR-499a was differentially expressed in the postmortem brains of BD patients compared with controls (39). Furthermore, a recent study by Banach et al. (40) reported lower expression levels of miR-499 in the peripheral blood of BD patients during depressive episodes in comparison to remission, suggesting miR-499 as a potential new biomarker of illness state in BD.
Overall, the results of our candidate approach do not suggest that individual BD-associated miRNAs might have a strong influence on differential responses to lithium treatment in BD as no association withstood multiple testing correction. On the one hand, this might at least in part reflect that the power to detect associations between common variants and lithium response was limited in the present study, even though the ConLiGen GWAS comprised several thousand individuals (41). On the other hand, it might also indicate that the genetic factors that contribute to BD etiology are different from those contributing to treatment response or illness course. That there are such effects in multifactorial diseases is supported by a study in ulcerative colitis in which no SNPs from 163 inflammatory bowel disease susceptibility loci (42) were found to be associated with the disease course (43).
In our systematic, genome-wide analysis of miRNAs, 106 miRNAs revealed nominally significant associations with the continuous and 71 with the dichotomous lithium treatment response.
The intergenic miR-633 located on chromosome 17 showed the strongest association with the continuous phenotype (p = 9.80E-04). To date, few published studies have investigated the function of miR-633. Interestingly, one study reported that miR-633 was differentially regulated in the cerebrospinal fluid of patients with multiple sclerosis compared to patients with other neurologic diseases. In addition, miR-633 differentiated relapsing-remitting from secondary progressive multiple sclerosis courses suggesting this miRNA as a potential biomarker for disease course in multiple sclerosis (44).
miR-607, an intergenic miRNA located upstream of the ligand dependent nuclear receptor corepressor (LCOR) gene on chromosome 10, displayed the strongest association with the dichotomous treatment response measure (p = 5.79E-04). The function of this miRNA has been poorly characterized so far, so that we cannot currently speculate about possible disease- and treatment-relevant biological processes. Further research is needed to elucidate the potential role of miR-607 in health or disease.
No association between miRNAs and BD treatment response to lithium in either of the tested conditions withstood multiple testing correction. In addition, we did not observe a significant enrichment for SNPs at all microRNA loci in the present study. Given the limited power of our study, future investigation of miRNAs in larger GWAS samples of BD and lithium response is warranted as better understanding of genetic factors contributing to disease etiology and treatment response might enable the individualization of treatment as well as the identification of novel therapeutic targets (45).
In the present study, we investigated all currently known miRNAs regardless of their tissue or developmental expression patterns. Approximately 70% of ncRNAs are thought to be brain expressed (23) and are dynamically regulated during development and over the lifespan. While the exact mechanisms by which lithium exerts its therapeutic effects remain unclear, pharmacokinetics and pharmacodynamics highlight the importance of specific tissues (e.g., brain and kidney) in treatment responsiveness (46, 47). Therefore, an analysis including miRNAs expressed specifically in these tissues would seem to be a rational follow-up step to reduce the multiple testing burden and to narrow-down the miRNAs to those that a priori may have a greater chance to be involved in lithium response. Unfortunately, a systematic enrichment analyses for miRNAs in particular tissues would be premature, since there are currently no comprehensive expression databases derived from normal tissue covering all known miRNAs investigated in the present study. Data on miRNA expression at various developmental stages is also still limited, as non-polyadenylated transcripts are typically not captured with standard library preparation for RNA sequencing. Furthermore, some miRNAs may only be expressed during early developmental stages but can still have an influence on lithium response later on in life, particularly if expression is induced by pharmaceutical treatment. Nevertheless, these aspects remain important and should be considered in future analyses as soon as more comprehensive data on miRNA expression become available.
Using the present approach, we were not able to investigate SNPs with trans-expression quantitative trait loci (eQTL) effects on miRNAs. Previous studies suggest that a substantial proportion of the identified miR-eQTLs are trans-eQTLs (48). Therefore, future investigations into the molecular interactions underlying the association between miRNA trans-eQTLs and treatment response to lithium in BD are also warranted. Huan et al. (37) conducted a genome-wide miR-eQTL mapping study and found consistent evidence for 5,269 cis-miR-eQTLs for 76 mature microRNAs. The significant enrichment for cis-miR-eQTL SNPs found in our summary statistics provides some evidence for the importance of cis-miR-eQTLs in lithium response, although we were not able to identify cis-miR-eQTL SNPs in our top findings since those miRNAs were not among the 76 mature microRNAs reported by Huan et al. (37).
Moreover, miRNAs only represent one class of non-coding RNAs. In the ConLiGen GWAS a genome-wide significant locus containing two lncRNAs was identified (14). Further analyses on the contribution of lncRNAs to lithium response are therefore warranted. This was beyond the scope of the present analysis as the current understanding of the predicted structure of lncRNA molecules and their biological functions remains limited (49).
In conclusion, our analyses do not provide strong evidence that miRNAs are involved in individual response to lithium treatment in BD, as no association between miRNAs and lithium response withstood multiple testing correction. Our data should still be interesting for follow-up of independent studies, particularly when sufficient data is available to accurately define the tissue and temporal expression profile of all human miRNAs, which would allow a more targeted analysis of brain-expressed miRNAs, thereby reducing the search space to miRNAs with relevant expression profiles. We did not find any strong effect that could be useful in terms of a personalized treatment for individual patients. This does not exclude a possible (small) effect of miRNAs on lithium response, and further independent and even larger studies should be envisaged to clarify this question. In parallel, the investigation of other biological mechanisms possibly contributing to lithium treatment response may provide insights for individualizing future pharmacotherapy in BD.
Author Contributions
CR, AF, MN, and SC contributed to the conception and design of the study. AF, LH, UH, FD, MA, KA, NA, RA, BA, LB, AB, SB, AKB, JB, ABi, CM-C, PC, G-BC, H-CC, CC, SRC, FC, DC, CCr, PMC, AD, BE, PF, LF, JF, SG, JG, FG, PG, OG, RH, JHa, SH, SJ, EJ, J-PK, LK, SK-S, SK, BK, IK, NL, GL, ML, CL, MLe, SL, CAL, GM, MM, LM, MMa, MJM, SM, MMi, FM, PM, CN, UÖ, NO, RP, AP, DR-E, GR, PRS, KOS, BS, FS, GS, TS, PDS, KS, CS, CMS, JS, AS, TSt, PS, ST, AT, GT, JV, SW, AW, LY, PZ, JP, JD, MB, ER, TN, J-MA, MMaj, BB, PM, EV, MF, JR, P-HK, TK, MG-S, AR, MD, FB, MS, NW, JK, MAl, FM, TGS, MR, and MN recruited the patients and contributed genotype data. CR and JH performed the statistical analysis. CR, AF, MR, MN, and SC prepared the manuscript, with feedback from the other authors. All authors contributed to manuscript revision, read and approved the submitted version.
Funding
The study was supported by the German Federal Ministry of Education and Research (BMBF) through the Integrated Network IntegraMent (Integrated Understanding of Causes and Mechanisms in Mental Disorders), under the auspices of the e:Med Programme (grant 01ZX1314A/01ZX1614A to MN and SC, grant 01ZX1314G/01ZX1614G to MR). The study was also supported by the German Research Foundation (DFG; grant FOR2107; RI908/11-1 to MR; NO246/10-1 to MN), and the Swiss National Science Foundation (SNSF, grant 310030_156791 to SC). MMN is a member of the DFG-funded Excellence-Cluster ImmunoSensation. AF received support from the BONFOR program of the Medical Faculty of the University of Bonn. The collection of the Geneva sample was supported by the Swiss National Foundation (grants Synapsy 51NF40-158776 and 32003B-125469). The Australian sample contribution was supported by the Australian National Health and Medical Research Council (grants 1037196, 1063960) and the Janette Mary O'Neil Research Fellowship to JF.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to all patients and control subjects for their participation. We thank the International Consortium on Lithium Genetics (ConLiGen) for providing access to the relevant data.
References
1. Craddock N, Sklar P. Genetics of bipolar disorder. Lancet (2013) 381:1654–62. doi: 10.1016/S0140-6736(13)60855-7
2. GBD 2016 Disease and Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990-2016: a systematic analysis for the Global Burden of Disease Study 2016. Lancet (2017) 390:1211–59. doi: 10.1016/s0140-6736(17)32154-2
3. Lichtenstein P, Yip BH, Björk C, Pawitan Y, Cannon TD, Sullivan PF, et al. Common genetic determinants of schizophrenia and bipolar disorder in Swedish families: a population-based study. Lancet (2009) 373:234–9. doi: 10.1016/S0140-6736(09)60072-6
4. McGuffin P, Rijsdijk F, Andrew M, Sham P, Katz R, Cardno A. The heritability of bipolar affective disorder and the genetic relationship to unipolar depression. Arch Gen Psychiatry (2003) 60:497–502. doi: 10.1001/archpsyc.60.5.497
5. Edvardsen J, Torgersen S, Røysamb E, Lygren S, Skre I, Onstad S, et al. Heritability of bipolar spectrum disorders. Unity or heterogeneity? J Affect Disord. (2008) 106:229–40. doi: 10.1016/j.jad.2007.07.001
6. Geddes JR, Miklowitz DJ. Treatment of bipolar disorder. Lancet (2013) 381:1672–82. doi: 10.1016/S0140-6736(13)60857-0
7. Geddes JR, Burgess S, Hawton K, Jamison K, Goodwin GM. Long-term lithium therapy for bipolar disorder: systematic review and meta-analysis of randomized controlled trials. Am. J. Psychiatry (2004) 161:217–22. doi: 10.1176/appi.ajp.161.2.217
8. Baldessarini RJ, Tondo L, Hennen J. Lithium treatment and suicide risk in major affective disorders: update and new findings. J Clin Psychiatry (2003) 64:(Suppl. 5), 44–52.
9. Cipriani A, Pretty H, Hawton K, Geddes JR. Lithium in the prevention of suicidal behavior and all-cause mortality in patients with mood disorders: a systematic review of randomized trials. Am J Psychiatry (2005) 162:1805–19. doi: 10.1176/appi.ajp.162.10.1805
10. Garnham J, Munro A, Slaney C, MacDougall M, Passmore M, Duffy A, et al. Prophylactic treatment response in bipolar disorder: results of a naturalistic observation study. J Affect Disord. (2007) 104:185–90. doi: 10.1016/j.jad.2007.03.003
11. Grof P, Duffy A, Cavazzoni P, Grof E, Garnham J, MacDougall M, et al. Is response to prophylactic lithium a familial trait? J Clin Psychiatry (2002) 63:942–7. doi: 10.4088/JCP.v63n1013
12. Kleindienst N, Engel R, Greil W. Which clinical factors predict response to prophylactic lithium? A systematic review for bipolar disorders. Bipolar Disord. (2005) 7:404–17. doi: 10.1111/j.1399-5618.2005.00244.x
13. Schulze TG, Alda M, Adli M, Akula N, Ardau R, Bui ET, et al. The International Consortium on Lithium Genetics (ConLiGen): an initiative by the NIMH and IGSLI to study the genetic basis of response to lithium treatment. Neuropsychobiology (2010) 62:72–8. doi: 10.1159/000314708
14. Hou L, Heilbronner U, Degenhardt F, Adli M, Akiyama K, Akula N, et al. Genetic variants associated with response to lithium treatment in bipolar disorder: a genome-wide association study. Lancet (2016) 387:1085–93. doi: 10.1016/S0140-6736(16)00143-4
15. Godnic I, Zorc M, Jevsinek Skok D, Calin GA, Horvat S, Dovc P, et al. Genome-wide and species-wide in silico screening for intragenic MicroRNAs in human, mouse and chicken. PLoS ONE (2013) 8:e65165. doi: 10.1371/journal.pone.0065165
16. Krol J, Loedige I, Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nat Rev Genet. (2010) 11:597–610. doi: 10.1038/nrg2843
17. Wang G, Wang Y, Shen C, Huang Y-W, Huang K, Huang TH, et al. RNA polymerase II binding patterns reveal genomic regions involved in MicroRNA gene regulation. PLoS ONE (2010) 5:e13798. doi: 10.1371/journal.pone.0013798
18. Gregory RI, Chendrimada TP, Cooch N, Shiekhattar R. Human RISC couples microRNA biogenesis and posttranscriptional gene silencing. Cell (2005) 123:631–40. doi: 10.1016/j.cell.2005.10.022
19. Mahdian-Shakib A, Dorostkar R, Tat M, Hashemzadeh MS, Saidi N. Differential role of microRNAs in prognosis, diagnosis, and therapy of ovarian cancer. Biomed Pharmacother. (2016) 84:592–600. doi: 10.1016/j.biopha.2016.09.087
20. Campos-Parra AD, Mitznahuatl GC, Pedroza-Torres A, Romo RV, Reyes FIP, Lopez-Urrutia E, et al. Micro-RNAs as potential predictors of response to breast cancer systemic therapy: future clinical implications. Int J Mol Sci. (2017) 18:E1182. doi: 10.3390/ijms18061182
21. Liu Q, Wu DH, Han L, Deng JW, Zhou L, He R, et al. Roles of microRNAs in psoriasis: immunological functions and potential biomarkers. Exp Dermatol. (2017) 26:359–67. doi: 10.1111/exd.13249
22. Hunsberger JG, Chibane FL, Elkahloun AG, Henderson R, Singh R, Lawson J, et al. Novel integrative genomic tool for interrogating lithium response in bipolar disorder. Transl Psychiatry (2015) 5:e504. doi: 10.1038/tp.2014.139
23. Fineberg SK, Kosik KS, Davidson BL. MicroRNAs potentiate neural development. Neuron (2009) 64:303–9. doi: 10.1016/j.neuron.2009.10.020
25. The Schizophrenia Psychiatric Genome-Wide Association Study (GWAS) Consortium. Genome-wide association study identifies five new schizophrenia loci. Nat Genet. (2011) 43:969. doi: 10.1038/ng.940
26. Schizophrenia Working Group of the Psychiatric Genomics Consortium. Biological insights from 108 schizophrenia-associated genetic loci. Nature (2014) 511:421–7. doi: 10.1038/nature13595
27. Mühleisen TW, Leber M, Schulze TG, Strohmaier J, Degenhardt F, Treutlein J, et al. Genome-wide association study reveals two new risk loci for bipolar disorder. Nat Commun. (2014) 5:3339. doi: 10.1038/ncomms4339
28. Forstner AJ, Hofmann A, Maaser A, Sumer S, Khudayberdiev S, Mühleisen TW, et al. Genome-wide analysis implicates microRNAs and their target genes in the development of bipolar disorder. Transl Psychiatry (2015) 5:e678. doi: 10.1038/tp.2015.159
29. Manchia M, Adli M, Akula N, Ardau R, Aubry J.-M, Backlund L, et al. Assessment of response to lithium maintenance treatment in bipolar disorder: a Consortium on Lithium Genetics (ConLiGen) Report. PLoS ONE (2013) 8:e65636. doi: 10.1371/journal.pone.0065636
30. Kozomara A, Griffiths-Jones S. miRBase: annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. (2014) 42:D68–73. doi: 10.1093/nar/gkt1181
31. Liu JZ, McRae AF, Nyholt DR, Medland SE, Wray NR, Brown KM, et al. A versatile gene-based test for genome-wide association studies. Am J Hum Genet. (2010) 87:139–45. doi: 10.1016/j.ajhg.2010.06.009
32. Mishra A, Macgregor S. VEGAS2: Software for more flexible gene-based testing. Twin Res Hum Genet. (2015) 18:86–91. doi: 10.1017/thg.2014.79
33. Hecker J, Maaser A, Prokopenko D, Fier HL, Lange C. Reporting correct p values in VEGAS analyses. Twin Res Hum Genet. (2017) 20:257–9. doi: 10.1017/thg.2017.16
34. Wojcik GL, Kao WHL, Duggal P. Relative performance of gene- and pathway-level methods as secondary analyses for genome-wide association studies. BMC Genet. (2015) 16:34. doi: 10.1186/s12863-015-0191-2
35. Auton A, Brooks LD, Durbin RM, Garrison EP, Kang HM, Korbel JO, et al. A global reference for human genetic variation. Nature (2015) 526:68–74. doi: 10.1038/nature15393
36. Sudmant PH, Rausch T, Gardner EJ, Handsaker RE, Abyzov A, Huddleston J, et al. An integrated map of structural variation in 2,504 human genomes. Nature (2015) 526:75. doi: 10.1038/nature15394
37. Huan T, Rong J, Liu C, Zhang X, Tanriverdi K, Joehanes R, et al. Genome-wide identification of microRNA expression quantitative trait loci. Nat Commun. (2015) 6:6601. doi: 10.1038/ncomms7601
38. Smalheiser NR, Lugli G, Zhang H, Rizavi H, Cook EH, Dwivedi Y. Expression of microRNAs and other small RNAs in prefrontal cortex in schizophrenia, bipolar disorder and depressed subjects. PLoS ONE (2014) 9:e86469. doi: 10.1371/journal.pone.0086469
39. Banigan MG, Kao PF, Kozubek JA, Winslow AR, Medina J, Costa J, et al. Differential expression of exosomal microRNAs in prefrontal cortices of schizophrenia and bipolar disorder patients. PLoS ONE (2013) 8:e48814. doi: 10.1371/journal.pone.0048814
40. Banach E, Dmitrzak-Weglarz M, Pawlak J, Kapelski P, Szczepankiewicz A, Rajewska-Rager A, et al.(2017). Dysregulation of miR-499, miR-708 and miR-1908 during a depression episode in bipolar disorders. Neurosci Lett. 654(Suppl. C):117–9. doi: 10.1016/j.neulet.2017.06.019
41. Sullivan PF, Daly MJ, O'Donovan M. Genetic architectures of psychiatric disorders: the emerging picture and its implications. Nat Rev Genet. (2012) 13:537–51. doi: 10.1038/nrg3240
42. Jostins L, Ripke S, Weersma RK, Duerr RH, McGovern DP, Hui KY, et al. Host-microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature (2012) 491:119–24. doi: 10.1038/nature11582
43. Kopylov U, Boucher G, Waterman M, Rivers CR, Patel M, Cho JH, et al. Genetic predictors of benign course of ulcerative colitis-a north American inflammatory bowel disease genetics Consortium Study. Inflamm Bowel Dis. (2016) 22:2311–6. doi: 10.1097/mib.0000000000000913
44. Haghikia A, Haghikia A, Hellwig K, Baraniskin A, Holzmann A, Decard BF, et al. Regulated microRNAs in the CSF of patients with multiple sclerosis: a case-control study. Neurology (2012) 79:2166–70. doi: 10.1212/WNL.0b013e3182759621
45. Bevilacqua L, Goldman D. Genes and addictions. Clin Pharmacol. Ther. (2009) 85:359–61. doi: 10.1038/clpt.2009.6
46. Mason RW, McQueen EG, Keary PJ, James NM. Pharmacokinetics of lithium: elimination half-time, renal clearance and apparent volume of distribution in schizophrenia. Clin Pharmacokinet. (1978) 3:241–6.
47. Brown KM, Tracy DK. Lithium: the pharmacodynamic actions of the amazing ion. Ther Adv. Psychopharmacol. (2013) 3:163–76. doi: 10.1177/2045125312471963
48. Borel C, Deutsch S, Letourneau A, Migliavacca E, Montgomery SB, Dimas AS, et al. Identification of cis- and trans-regulatory variation modulating microRNA expression levels in human fibroblasts. Genome Res. (2011) 21:68–73. doi: 10.1101/gr.109371.110
Keywords: bipolar disorder, lithium response, microRNA, common variants, genome-wide association study
Citation: Citation: Reinbold CS, Forstner AJ, Hecker J, Fullerton JM, Hoffmann P, Hou L, Heilbronner U, Degenhardt F, Adli M, Akiyama K, Akula N, Ardau R, Arias B, Backlund L, Benabarre A, Bengesser S, Bhattacharjee AK, Biernacka JM, Birner A, Marie-Claire C, Cervantes P, Chen G-B, Chen H-C, Chillotti C, Clark SR, Colom F, Cousins DA, Cruceanu C, Czerski PM, Dayer A, Étain B, Falkai P, Frisén L, Gard S, Garnham JS, Goes FS, Grof P, Gruber O, Hashimoto R, Hauser J, Herms S, Jamain S, Jiménez E, Kahn J-P, Kassem L, Kittel-Schneider S, Kliwicki S, König B, Kusumi I, Lackner N, Laje G, Landén M, Lavebratt C, Leboyer M, Leckband SG, López Jaramillo CA, MacQueen G, Manchia M, Martinsson L, Mattheisen M, McCarthy MJ, McElroy SL, Mitjans M, Mondimore FM, Monteleone P, Nievergelt CM, Ösby U, Ozaki N, Perlis RH, Pfennig A, Reich- Erkelenz D, Rouleau GA, Schofield PR, Schubert KO, Schweizer BW, Seemüller F, Severino G, Shekhtman T, Shilling PD, Shimoda K, Simhandl C, Slaney CM, Smoller JW, Squassina A, Stamm TJ, Stopkova P, Tighe SK, Tortorella A, Turecki G, Volkert J,Witt SH,Wright AJ, Young LT, Zandi PP, Potash JB, DePaulo JR, BauerM, Reininghaus E, Novák T, Aubry J-M, Maj M, Baune BT, Mitchell PB, Vieta E, Frye MA, Rybakowski JK, Kuo P-H, Kato T, Grigoroiu-Serbanescu M, Reif A, Del Zompo M, Bellivier F, Schalling M, Wray NR, Kelsoe JR, Alda M, McMahon FJ, Schulze TG, Rietschel M, Nöthen MM and Cichon S (2018) Analysis of the Influence of microRNAs in Lithium Response in Bipolar Disorder. Front. Psychiatry 9:207. doi: 10.3389/fpsyt.2018.00207
Received: 09 February 2018; Accepted: 03 May 2018;
Published: 31 May 2018.
Edited by:
Elizabeth A. Thomas, The Scripps Research Institute, United StatesReviewed by:
Janine Adele Lamb, University of Manchester, United KingdomEmma Sprooten, Donders Institute for Brain, Cognition and Behaviour, Radboud University, Netherlands
Copyright © 2018 Reinbold, Forstner, Hecker, Fullerton, Hoffmann, Hou, Heilbronner, Degenhardt, Adli, Akiyama, Akula, Ardau, Arias, Backlund, Benabarre, Bengesser, Bhattacharjee, Biernacka, Birner, Marie-Claire, Cervantes, Chen, Chen, Chillotti, Clark, Colom, Cousins, Cruceanu, Czerski, Dayer, Étain, Falkai, Frisén, Gard, Garnham, Goes, Grof, Gruber, Hashimoto, Hauser, Herms, Jamain, Jiménez, Kahn, Kassem, Kittel-Schneider, Kliwicki, König, Kusumi, Lackner, Laje, Landén, Lavebratt, Leboyer, Leckband, López Jaramillo, MacQueen, Manchia, Martinsson, Mattheisen, McCarthy, McElroy, Mitjans, Mondimore, Monteleone, Nievergelt, Ösby, Ozaki, Perlis, Pfennig, Reich-Erkelenz, Rouleau, Schofield, Schubert, Schweizer, Seemüller, Severino, Shekhtman, Shilling, Shimoda, Simhandl, Slaney, Smoller, Squassina, Stamm, Stopkova, Tighe, Tortorella, Turecki, Volkert, Witt, Wright, Young, Zandi, Potash, DePaulo, Bauer, Reininghaus, Novák, Aubry, Maj, Baune, Mitchell, Vieta, Frye, Rybakowski, Kuo, Kato, Grigoroiu-Serbanescu, Reif, Del Zompo, Bellivier, Schalling, Wray, Kelsoe, Alda, McMahon, Schulze, Rietschel, Nöthen and Cichon. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sven Cichon, c3Zlbi5jaWNob25AdW5pYmFzLmNo
†These authors have contributed equally to this work.