 Xueping Wang
Xueping Wang Yi Su
Yi Su Hao Yan1,2,3
Hao Yan1,2,3 Weihua Yue
Weihua Yue- 1Peking University Sixth Hospital, Institute of Mental Health, Beijing, China
- 2NHC Key Laboratory of Mental Health, Peking University, Beijing, China
- 3National Clinical Research Center for Mental Disorders, Peking University Sixth Hospital, Beijing, China
- 4State Key Laboratory of Natural and Biomimetic Drugs, Department of Molecular and Cellular Pharmacology, School of Pharmaceutical Sciences, Peking University Health Science Center, Beijing, China
- 5Key Laboratory for Neuroscience, Ministry of Education, Beijing, China
- 6National Engineering Research Center for Software Engineering, Peking University, Beijing, China
Risperidone has been used to treat the symptoms of schizophrenia and to reduce its relapse. However, the responses to treatment show great variability among patients. The potassium channel has been reported as an effective target for antipsychotics. KCNH7, a member of the voltage-gated K+ channel Kv11 family, is primarily expressed in the brain. Here, we assessed the genetic association of KCNH7 with risperidone responses in 393 schizophrenia patients. The patients were treated with risperidone for 6 weeks. The reduction rates of Positive and Negative Syndrome Scale (PANSS) scores were determined to quantify drug response. We also examined the associations between six single-nucleotide polymorphisms (SNPs) of KCNH7 and the risperidone responses for a total of 6 weeks. The SNP rs77699177 (C > T) in the KCNH7 gene intron was significantly associated with the treatment response reflected by the PANSS reduction rate (CC, 55.8 ± 23.0; TC, 70.9 ± 20.3, P = 0.000110), indicating that patients with the TC genotype have better efficacy for antipsychotic therapy. The rs2241240 SNP also showed a significant association with treatment responses after 6 weeks of treatment (P = 0.00256). The findings indicate that the voltage-gated K+ channel KCNH7 is a potential functional marker for the identification of the response to risperidone treatment in schizophrenia patients.
Note: The study was registered under clinical trial number ChiCTR-RNC-09000522 (http://www.chictr.org/).
Introduction
Schizophrenia is a severe psychiatric disorder characterized by both positive and negative symptoms. Typical symptoms of schizophrenia include hallucinations, delusions, thought disorders, avolition, affective flattening, and social withdrawal. The condition affects about 1% of the population worldwide (1), and patients require treatment throughout their lives because of the chronic and debilitating course of this disease. Thus, schizophrenia imposes a heavy burden on both individual patients and the public health-care system (2).
Clinical management of schizophrenia involves different antipsychotic drugs, including typical/first-generation antipsychotics (FGAs) and atypical/second-generation antipsychotics (SGAs). These drugs are known to improve positive and negative symptoms in many patients. However, responses to drug treatment show wide variability (3). A previous review reported that approximately 75% of patients exhibit low drug compliance because of ineffectiveness or side effects of these drugs (4), consequently leading to relapse, clinical exacerbation, and/or repeated admissions. Therefore, understanding the individual responses to antipsychotics is imperative for the development of precision medicine, and it is critical to establish a personalized antipsychotic treatment system to improve the clinical outcome.
Studies have suggested that genetic factors play a significant role in personal responses to medication (5–7), and many genetic variants of the dopamine and serotonin receptor genes have been reported in the past decades (8–12). The Kv11 family, also called the HERG family, belongs to the subfamily H of the voltage-gated potassium (Kv) channels and plays an important role in the membrane potential regulation (13). The Kv11 family includes three members, which are KCNH2/Kv11.1, KCNH6/Kv11.2, and KCNH7/Kv11.3 (14). All Kv11 channels are expressed in the nervous system. KCNH7 and KCNH2 are expressed in the brain (15, 16), and KCNH6 is predominantly expressed in the superior mesenteric ganglia (15). These expression data indicate their functions in the nervous system. Several studies have reported KCNH2 as a risk gene for schizophrenia (16–19). One recent study revealed that schizophrenia patients with different KCNH2 genotypes showed different responses to risperidone, a common antipsychotic drug used in the clinic (20). The findings suggest that the Kv11 family plays an important role in the antipsychotic response.
The KCNH7 gene shows higher expression levels in the brain than in other tissues (15), while the KCNH2 gene has higher expression levels in the heart than in the brain (16). In this study, we hypothesized that KCNH7 contributes to the outcomes of risperidone treatment in schizophrenia patients. To this end, we performed a pharmacogenetics association study to identify the genetic function of KCNH7 and examined the associations between the KCNH7 genotype and risperidone efficacy in schizophrenia patients of Han Chinese ancestry over a 6-week acute treatment.
Methods
Study Design and Participants
A total of 452 patients were enrolled at our hospital and three other hospitals (from Hebei, Henan, and Liaoning provinces), and among these patients, 393 underwent single-nucleotide polymorphism (SNP) genotype assessment. Patients included in this study met the following inclusion criteria: (I) diagnosed with schizophrenia based on the Structured Clinical Interview of the Diagnostic and Statistical Manual of Mental Disorders, fourth edition, Text Revision (DSM-IV-TR); (II) aged 18–45 years; (III) a Han Chinese lineage; (IV) first-onset or chronic disease in the acute phase with severe symptoms; (V) total scores of more than 70 on the Positive and Negative Syndrome Scale (PANSS); and (VI) provided written informed consent.
The exclusion criteria were as follows: (I) pregnancy or breast-feeding; (II) contraindications to the recommended drugs; (III) severe or unstable physical diseases; (IV) presence of QTc > 450 ms in males or QTc > 470 ms in females, decompensated and congestive heart failure, or complete left bundle branch block delay; (V) a history of serious suicide attempts or severe excitement and agitation episodes; and (VI) requirement of long-acting injectable medication to maintain treatment adherence or regularly treated with clozapine for treatment over the past month.
We recruited patients and performed several routine baseline assessments at the start of the study, including DSM-IV-TR, inclusion/exclusion criteria selection, patient general information records, and Positive and Negative Syndrome Scale (PANSS). Within the first 2 weeks after enrollment, clinicians could adjust the drug dosages based on the treatment effectiveness (risperidone 2–6 mg/day), and the adjusted dosage of the antipsychotics remained unchanged throughout the rest of the study period. The patients visited the participating clinicians at weeks 2, 4, and 6, and their PANSS scores were recorded by the psychiatrists. All the patients and clinicians were blinded to the study design. Treatment was discontinued if the patient response was not adequate or if the patient decided to withdraw from the study; in that case, the last-observation-carried-forward procedure was used to show her or his treatment response. Patients with adequate responses continued treatment for up to 6 weeks. The objective and procedure of this study were explained to all participants, and all included participants provided written informed consent. The study was approved by the research ethics committees and institutional review boards of each local hospital. All procedures were conducted in accordance with the principles expressed in the Declaration of Helsinki. The study was registered under the clinical trial number ChiCTR-RNC-09000522 (http://www.chictr.org/).
A total of 59 individuals withdrew from this study, and the reasons were as follows: withdrawing the informed consent and refusing to continue treatment according to the study protocol (36 individuals); lost to follow-up (13 individuals); occurrence or recurrence of severe physical diseases during treatment (7 individuals); and changed treatment according to the doctors’ advice (3 individuals).
Phenotype Definition
Using the percentage changes in the PANSS scores to indicate treatment effects is a common strategy in statistical analyses. The PANSS is an interval scale with each item ranging from 1 to 7 and lacking a natural zero point. The 30-item PANSS yields scores of 30∼210. Categorizations using the PANSS may reduce the sensitivity and power of statistical tests. Therefore, continuous measures are suggested to cutoff-based dichotomous measures of treatment responses. We used percentage changes in the PANSS scores to evaluate acute responses to risperidone medications. The theoretical minimum (30 for the total score) was subtracted from the baseline score to avoid incorrect calculations, leading to a score range including zero (21). The PANSS change from baseline was defined as PANSS endpoint score − PANSS baseline score. The percentage changes of the PANSS reduction rate was defined as follows: the reduction rate of the total score of PANSS = (PANSS baseline score − PANSS endpoint score) ÷ (PANSS baseline score − 30) × 100%. The higher reduction rate indicates a better efficacy for antipsychotic therapy.
Genotyping
Genomic DNA was extracted using the QIAamp DNA Mini Kit (QIAGEN, Hilden, Germany). Sample genotype sequencing was conducted using the Sequenom MassARRAY system (Sequenom iPLEX assay). The DNA samples from 393 schizophrenia patients were amplified by multiplex polymerase chain reactions (PCRs), and the PCR products were then assessed for locus-specific single-base extension reactions. The primer sequences for PCR and extension are listed in Table S1. We selected six SNPs randomly across the gene, with more than 300-kb inter-SNP space, from different linkage disequilibrium (LD) blocks. The pairwise LD analysis was applied to detect the inter-marker relationship, using D′ values.
Statistical Analyses
After quality control, we used linear regression under an additive genetic model to evaluate the associations between the allele dosages and the PANSS percentage changes in PLINK v1.07 (22). We used percentage changes of PANSS to assess the treatment responses to antipsychotic medications in this study. The gender, age, and the first five principal components of the population structure were used as covariates in the analysis. The P values were adjusted by multiple comparisons (false discovery rate (FDR)).
The repeated-measures linear mixed model in SPSS (IBM SPSS Statistics 20) was used to analyze the assessments at different time points. The longitudinal PANSS percentage change was analyzed using linear mixed models in SPSS. We classified the subject as a “Subjects” variable and WEEK as a “Repeat” variable. The first-order autoregressive [AR(1)] covariance structure was selected, and we then defined the reduced PANSS percentage as a dependent variable, the genotype of KCNH7 as a categorical factor, and the WEEK as a continuous covariate. In the model, fixed effects involved WEEK, genotype, and the interaction between WEEK and genotype, and the Subject was defined as a random effect.
For the associated SNPs, we examined their genome-wide cis-e-quantitative trait loci (eQTL) effects in the Brain Expression Consortium (Braineac, GTEx, and Blood eQTL).
Results
PANSS Reduction Rate Indicates the Response to Risperidone Treatment
A total of 393 schizophrenia patients remained after quality control. All patients had Han Chinese ancestry and were unrelated to each other. We collected basic information of the included patients and performed baseline assessments at the start of the study. The mean age at study entry was 31.6 ± 8.0 years, and the baseline PANSS score was 89.3 ± 15.3, as shown in Table 1. The patients received risperidone monotherapy, and the PANSS scores were assessed after 2, 4, and 6 weeks to reflect the treatment effect. The endpoint PANSS score after the 6-week risperidone monotherapy was 55.4 ± 15.4, which indicated the general treatment effect of risperidone. The PANSS reduction rate was 57.2 ± 23.1, as shown in Table 1.
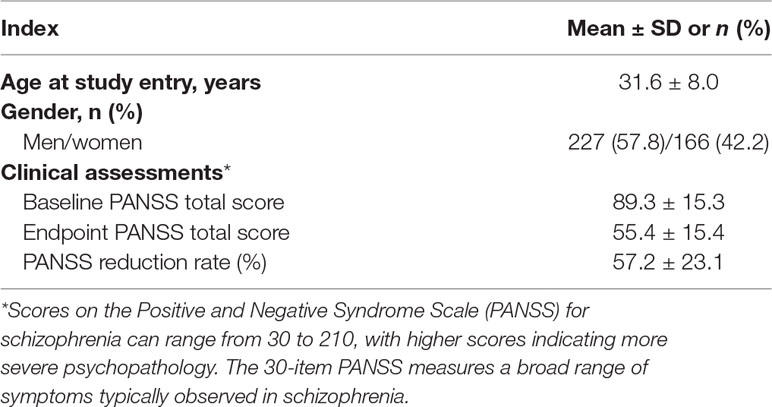
Table 1 Demographic and clinical characteristics of 393 patients following 6-week risperidone monotherapy.
KCNH7 Shows a Significant Association With Treatment Responses
We next analyzed the association between SNPs of KCNH7 and the PANSS reduction rate. Linear regression analysis revealed that SNPs showed differential associations with treatment responses (Table 2). The most significant association was observed between treatment responses and SNP rs77699177 at the intron regions of the KCNH7 gene. The PANSS reduction rates were statistically significant in patients with rs77699177 after treatment for 6 weeks (CC, 55.8 ± 23.0; TC, 70.9 ± 20.3, P = 0.000110), indicating that patients with the TC genotype have better efficacy for risperidone therapy. In addition, the PANSS reduction rates were also statistically significant in patients with rs2241240 (TT, 58.7 ± 22.3; CT + CC, 50.9 ± 25.2, P = 0.00256). However, other SNPs did not show significant associations with treatment responses.
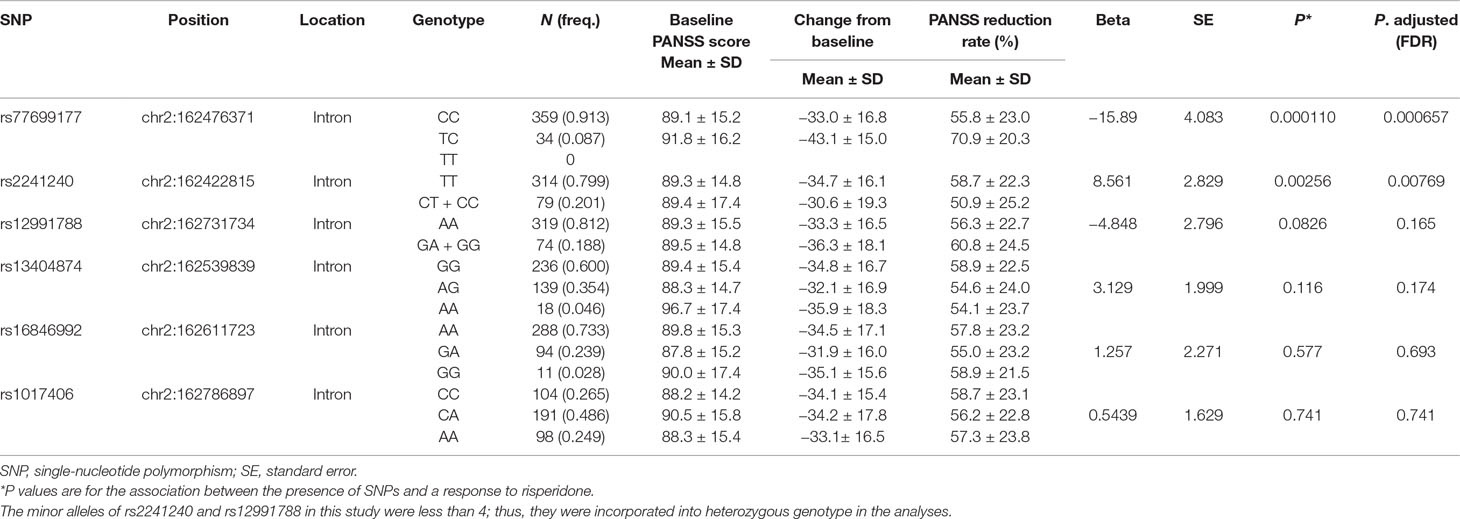
Table 2 The association of KCNH7 gene polymorphisms with risperidone treatment responses after 6 weeks.
In order to study the time course of responses in the treatment process, we assessed the PANSS reduction rates at 2, 4, and 6 weeks for each of the SNP genotypes. In this analysis, the PANSS reduction rates in patients with different genotypes (CC/TC) for rs77699177 showed significant differences (P ≤ 0.001) after 6 weeks of risperidone treatment (Figure 1A). The patients carrying the TC genotype showed better responses to the risperidone treatment for a total of 6 weeks. Significant differences in treatment responses were observed between the two sets of rs77699177 throughout the treatment process (P = 0.000569) (Figure 1B). In addition, significant PANSS percentage changes were found in patients with rs2241240 after the 6-week treatment (P ≤ 0.01) (Figure 1C), and the difference between the two sets during the 6-week treatment was noticeable (P = 0.0317) (Figure 1D). Therefore, SNP rs77699177 may serve as a genetic indicator for risperidone usage.
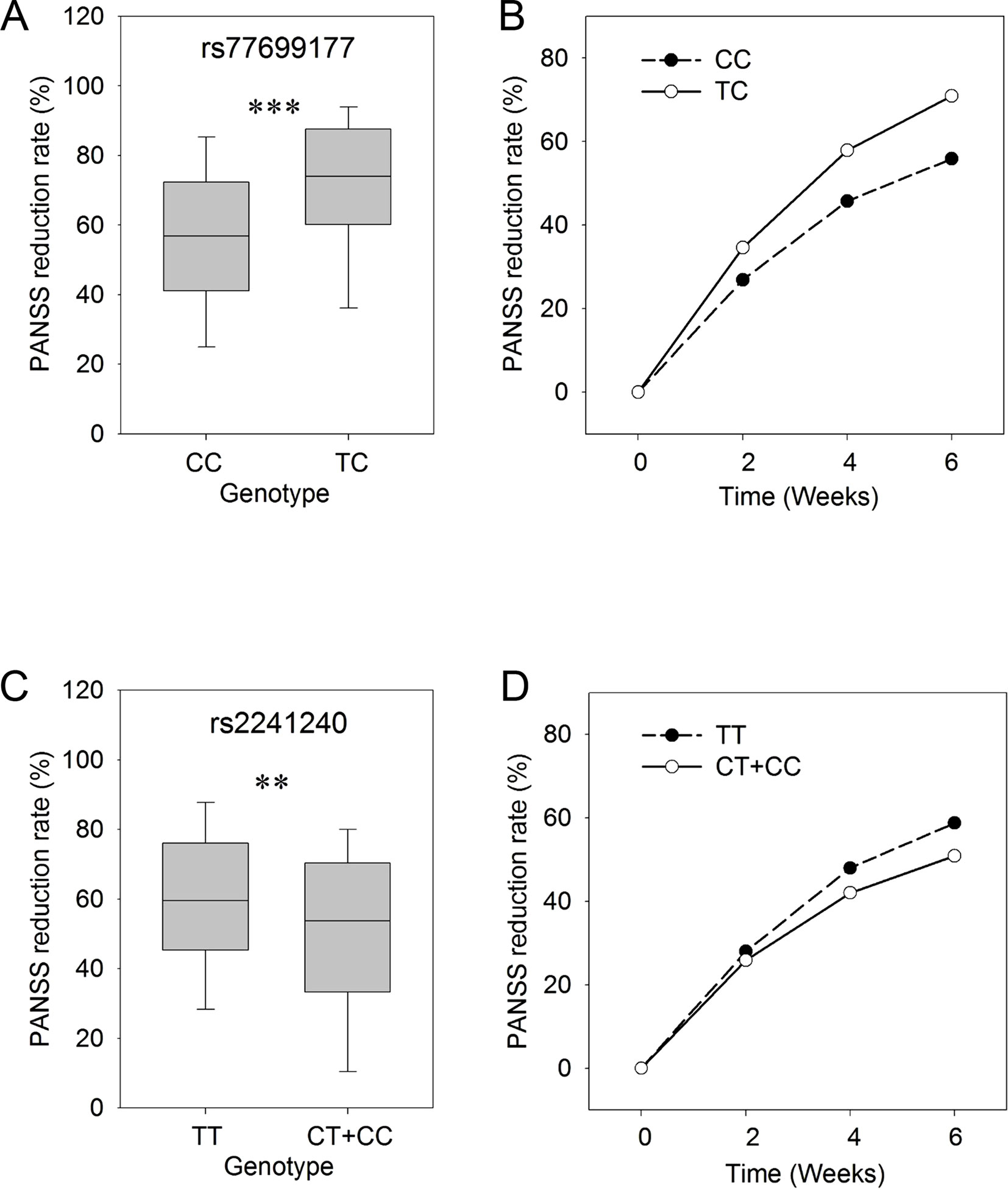
Figure 1 The genotypes of rs77699177 and rs2241240 associated with PANSS reduction rates following risperidone monotherapy. Box plot of the KCNH7 rs77699177 CC/TC allele and the PANSS reduction rates after 6 weeks (A). The PANSS reduction rates during the 6-week risperidone treatment (B). Plot of the rs2241240 TT/(CT + CC) allele after 6 weeks (C) and the change over 6 weeks (D). **P < 0.001; ***P < 0.0001.
Among the three members of the Kv11 family (KCNH2, KCNH6, and KCNH7) (14), the KCNH7 gene shows the highest expression level in the brain (Figure S1, https://www.ncbi.nlm.nih.gov/gene) and has a high expression during embryonic development (Figure S2, http://hbatlas.org/pages/hbtd). We attempted to assess the expression quantitative trait loci (eQTL) in several databases (Braineac, GTEx, and Blood eQTL). Unfortunately, the SNPs rs77699177 and rs2241240 analyzed in this study were not included in the eQTL databases. The eQTL for KCNH7 SNPs in several brain tissues is listed in Table S2, which was derived from the Braineac database (23). Both rs918927 and rs2216897 showed significant eQTL effects in the cerebellar cortex (Figure S3, http://www.braineac.org).
Discussion
Proper and optimal use of antipsychotic drugs personalized to each patient is important in psychiatric therapy. However, limited evidence is available to guide the use of antipsychotic drugs. Our findings showed that a SNP in KCNH7 (rs77699177; CC, 55.8 ± 23.0; TC, 70.9 ± 20.3, P = 0.000110) was associated with response to risperidone treatment, and patients with the TC genotype had better efficacy for risperidone therapy. KCNH7 encodes HERG3/Kv11.3, which is involved in potassium homeostasis and is essential for neuronal excitability. The present study indicates that rs77699177 in the KCNH7 gene could serve as a genetic factor in the identification of the treatment response to risperidone. These findings provide further evidence for the neuronal function of the Kv channel in the pathology of schizophrenia.
Risperidone is used to treat not only schizophrenia but also bipolar disorder and irritability associated with autism (24–26). Studies have demonstrated the association between KCNH7 and bipolar disorder and autism (27–29). A non-synonymous variant of KCNH7 (c.1181G > A, p.Arg394His), rs78247304, has a strong family-based association with bipolar spectrum disorder (27). Targeted massively parallel sequencing demonstrates a nominal gene-based rare variant association between autism spectrum disorder and KCNH7 (P = 0.043) (29). KCNH7 might be an important genetic factor in the efficacy of risperidone against mental disorders. Further studies are needed to investigate the pharmacogenetic relationships between KCNH7 and risperidone in bipolar disorder and autism, and the possible role of KCNH7 in the treatment efficacy of risperidone in bipolar disorder remains to be investigated.
Pharmacogenetic studies have investigated the genetic variants of neurotransmitter receptors, such as the dopamine and serotonin receptors (5). A number of clinical studies have examined the dopamine receptors (DRD2, DRD3, and DRD4), serotonin receptors (HTR2A, HTR2C, 5-HT6, and 5HTT), and genes related to their metabolic pathways (such as COMT andCYP2D6) (5). However, many genetic variants display variable results, probably because of the limited effect size. The K+ channel KCNH2 is now being assessed in pharmacogenetic studies. The primate-specific isoform KCNH2-3.1 affects cognition and neuronal repolarization and is associated with the risk of schizophrenia (16, 18, 19). Different antipsychotics have a distinct affinity for KCNH2 (30). One recent study found that risperidone caused greater block of KCNH2-3.1 isoform than that of full-length KCNH2 in electrophysiology recordings (20). The clinical observations suggest that patients with the KCNH2 risk genotype (higher KCNH2-3.1 expression levels) combined with slow metabolism status have better responses to risperidone treatment (20). The findings mentioned above further support the theory that the Kv11 channel could be the functional target for risperidone.
Two recent studies showed that potassium channel Kv2.1 and Kv7.2/7.3 could separately form complexes with dopamine transporter and affect dopamine transport and electrophysiological function (31, 32). A previous study found that the haloperidol treatment could block the K+-induced dopamine release in the caudate–putamen and nucleus accumbens and prevent K+-induced hyperlocomotion (33). However, whether KCNH7 and other members of Kv11 have similar roles remains unclear. In the Kv11 family, the three genes KCNH7, KCNH2, and KCNH6 show differentiated expression patterns (Figure S1, https://www.ncbi.nlm.nih.gov/gene). KCNH2 and KCNH7 show higher expression levels in the brain than KCNH6 (Figure S1), and KCNH7 has relatively high expression levels in neurons (Figure S4, http://www.alzdata.org), suggesting its role in the central nervous system. KCNH2 and KCNH7 might have overlapping functions in the central nervous system. In the study by Heide et al., risperidone showed greater blockade for KCNH2-3.1 than for KCNH2-1A in the electrophysiology assay (20); however, additional assessments were hindered because of the small sample size. Our study included 393 eligible patients in the association analysis, and the responder analysis for SNPs of KCNH7 gene is listed in Table S3. The SNPs (rs77699177 and rs2241240) found in our study are in the intron of the KCNH7 gene, and the specific functional study of the channel protein is thus limited. Moreover, information is lacking on these SNPs in the current public eQTL databases (Braineac, GTEx, and Blood eQTL Browser), probably because of their low minor allele frequencies in people of European descent. The other possible reason is the unchanged expression levels of KCNH7 between controls and schizophrenia patients (Figure S5, http://www.szdb.org/index.html). Thus, the SNPs in KCNH7 might affect the gene function in other ways that need to be studied further. Accordingly, the biological effect of KCNH7 on the efficacy of risperidone treatment in schizophrenia has been added to our study agenda.
This study has some limitations. First, the sample size was small. Therefore, further studies are needed to enroll more patients to confirm our present findings in this study. Second, the relatively sparse points selected in our study might have precluded the possibility of detecting some potential functional SNPs. Third, the biological function of rs77699177 was not sufficiently explored.
In this study, the SNPs in KCNH7 showed significant associations with the treatment response to risperidone in 393 patients of Han Chinese ancestry. The results indicate the essential role of KCNH7 in risperidone’s therapeutic efficacy. KCNH7 might serve not only as a new functional marker for predicting the response to risperidone but also as a new drug target in the discovery of new antipsychotics. KCNH2, more commonly known as hERG, is primarily expressed in the brain and plays an essential role in cardiac action potential, and its mutations cause long QT syndrome (34). The KCNH2-related effect of risperidone would be limited because of its potential side effect on QT interval prolongation; however, KCNH7 might not have similar side effects. Therefore, KCNH7 could be an ideal target for designing new antipsychotics. Further studies are needed to determine whether risperidone can bind to these potassium channels. Moreover, studies on the difference in regulation and effects between KCNH7 and KCNH2 will help understand the function of the Kv channel in schizophrenia.
Ethics Statement
The study was approved by the research ethical committees and institutional review boards of each local hospital. All procedures were conducted in accord with principles expressed in the Declaration of Helsinki. The study was registered for clinical trial number as ChiCTR-RNC-09000522 (http://www.chictr.org/).
Author Contributions
WY and YH designed the study and provided the funds. XW wrote the manuscript and analyzed the data with YS. HY and ZH supervised this study.
Funding
The study was funded by the National Key R&D Program of China (2016YFC1307000), National Natural Science Foundation of China (81571313 and 91432304), National High Technology Research and Development Program of China (2009AA022702), Beijing Nova Program Interdisciplinary Studies Cooperative Projects (Z161100004916038), Peking University Seed Fund for Medicine-Information Interdisciplinary Research Project (BMU20160597), the Fund for Fostering Young Scholars of Peking University Health Science Center (BMU2017PY030), Peking University Clinical Scientist Program (BMU2019LCKXJ012), supported by “the Fundamental Research Funds for the Central Universities”, and National Science and Technology Major Project for IND (investigational new drug) 2018ZX09201-014.
Conflict on Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank all subjects who participated in our study.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpsyt.2019.00633/full#supplementary-material
References
1. Thaker GK, Carpenter WT Jr. Advances in schizophrenia. Nat Med (2001) 7(6):667–71. doi: 10.1038/89040
2. Ilyas S, Moncrieff J. Trends in prescriptions and costs of drugs for mental disorders in England, 1998-2010. Br J Psychiatry (2012) 200(5):393–8. doi: 10.1192/bjp.bp.111.104257
3. Huber CG, Naber D, Lambert M. Incomplete remission and treatment resistance in first-episode psychosis: definition, prevalence and predictors. Expert Opin Pharmacother (2008) 9(12):2027–38. doi: 10.1517/14656566.9.12.2027
4. Lieberman JA. Effectiveness of antipsychotic drugs in patients with chronic schizophrenia: efficacy, safety and cost outcomes of CATIE and other trials. J Clin Psychiatry (2007) 68(2):e04. doi: 10.4088/JCP.0207e04
5. Zhang JP, Malhotra AK. Pharmacogenetics and antipsychotics: therapeutic efficacy and side effects prediction. Expert Opin Drug Metab Toxicol (2011) 7(1):9–37. doi: 10.1517/17425255.2011.532787
6. Hu X, Zhang J, Jin C, Mi W, Wang F, Ma W, et al. Association study of NRXN3 polymorphisms with schizophrenia and risperidone-induced bodyweight gain in Chinese Han population. Prog Neuropsychopharmacol Biol Psychiatry (2013) 43:197–202. doi: 10.1016/j.pnpbp.2012.12.007
7. McClay JL, Adkins DE, Aberg K, Stroup S, Perkins DO, Vladimirov VI, et al. Genome-wide pharmacogenomic analysis of response to treatment with antipsychotics. Mol Psychiatry (2011) 16(1):76–85. doi: 10.1038/mp.2009.89
8. Hill MJ, Reynolds GP. Functional consequences of two HTR2C polymorphisms associated with antipsychotic-induced weight gain. Pharmacogenomics (2011) 12(5):727–34. doi: 10.2217/pgs.11.16
9. Calarge CA, Ellingrod VL, Acion L, Miller DD, Moline J, Tansey MJ, et al. Variants of the dopamine D2 receptor gene and risperidone-induced hyperprolactinemia in children and adolescents. Pharmacogenet Genomics (2009) 19(5):373–82. doi: 10.1097/FPC.0b013e328329a60f
10. Weickert TW, Goldberg TE, Mishara A, Apud JA, Kolachana BS, Egan MF, et al. Catechol-O-methyltransferase val108/158met genotype predicts working memory response to antipsychotic medications. Biol Psychiatry (2004) 56(9):677–82. doi: 10.1016/j.biopsych.2004.08.012
11. Yamanouchi Y, Iwata N, Suzuki T, Kitajima T, Ikeda M, Ozaki N. Effect of DRD2, 5-HT2A, and COMT genes on antipsychotic response to risperidone. Pharmacogenomics J (2003) 3(6):356–61. doi: 10.1038/sj.tpj.6500211
12. Arranz MJ, Munro J, Sham P, Kirov G, Murray RM, Collier DA, et al. Meta-analysis of studies on genetic variation in 5-HT2A receptors and clozapine response. Schizophr Res (1998) 32(2):93–9. doi: 10.1016/S0920-9964(98)00032-2
13. Gutman GA, Chandy KG, Grissmer S, Lazdunski M, McKinnon D, Pardo LA, et al. International Union of Pharmacology. LIII. Nomenclature and molecular relationships of voltage-gated potassium channels. Pharmacol Rev (2005) 57(4):473–508. doi: 10.1124/pr.57.4.10
14. Einarsen K, Calloe K, Grunnet M, Olesen S-P, Schmitt N. Functional properties of human neuronal Kv11 channels. Pflügers Archiv - Eur J Physiol (2009) 458(4):689–700. doi: 10.1007/s00424-009-0651-5
15. Shi W, Wymore RS, Wang HS, Pan Z, Cohen IS, McKinnon D, et al. Identification of two nervous system-specific members of the erg potassium channel gene family. J Neurosci (1997) 17(24):9423–32. doi: 10.1523/JNEUROSCI.17-24-09423.1997
16. Huffaker SJ, Chen J, Nicodemus KK, Sambataro F, Yang F, Mattay V, et al. A primate-specific, brain isoform of KCNH2 affects cortical physiology, cognition, neuronal repolarization and risk of schizophrenia. Nat Med (2009) 15(5):509–18. doi: 10.1038/nm.1962
17. Atalar F, Acuner TT, Cine N, Oncu F, Yesilbursa D, Ozbek U, et al. Two four-marker haplotypes on 7q36.1 region indicate that the potassium channel gene HERG1 (KCNH2, Kv11.1) is related to schizophrenia: a case control study. Behav Brain Funct (2010) 6:27. doi: 10.1186/1744-9081-6-27
18. Hashimoto R, Ohi K, Yasuda Y, Fukumoto M, Yamamori H, Kamino K, et al. The KCNH2 gene is associated with neurocognition and the risk of schizophrenia. World J Biol Psychiatry (2013) 14(2):114–20. doi: 10.3109/15622975.2011.604350
19. Carr GV, Chen J, Yang F, Ren M, Yuan P, Tian Q, et al. KCNH2-3.1 expression impairs cognition and alters neuronal function in a model of molecular pathology associated with schizophrenia. Mol Psychiatry (2016) 21(11):1517–26. doi: 10.1038/mp.2015.219
20. Heide J, Zhang F, Bigos KL, Mann SA, Carr VJ, Shannon Weickert C, et al. Differential response to risperidone in schizophrenia patients by KCNH2 genotype and drug metabolizer status. Am J Psychiatry (2016) 173(1):53–9. doi: 10.1176/appi.ajp.2015.14050653
21. Obermeier M, Mayr A, Schennach-Wolff R, Seemüller F, Möller H-J, Riedel M. Should the PANSS be rescaled? Schizophr Bull (2010) 36(3):455–60. doi: 10.1093/schbul/sbp124
22. Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, Bender D, et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet (2007) 81(3):559–75. doi: 10.1086/519795
23. Ramasamy A, Trabzuni D, Guelfi S, Varghese V, Smith C, Walker R, et al. Genetic variability in the regulation of gene expression in ten regions of the human brain. Nat Neurosci (2014) 17(10):1418–28. doi: 10.1038/nn.3801
24. Lehman AF, Lieberman JA, Dixon LB, McGlashan TH, Miller AL, Perkins DO, et al. Practice guideline for the treatment of patients with schizophrenia, second edition. Am J Psychiatry (2004) 161(2 Suppl):1–56.
25. Malone RP, Maislin G, Choudhury MS, Gifford C, Delaney MA. Risperidone treatment in children and adolescents with autism: short- and long-term safety and effectiveness. J Am Acad Child Adolesc Psychiatry (2002) 41(2):140–7. doi: 10.1097/00004583-200202000-00007
26. American Psychiatric A. Practice guideline for the treatment of patients with bipolar disorder (revision). Am J Psychiatry (2002) 159(4 Suppl):1–50.
27. Strauss KA, Markx S, Georgi B, Paul SM, Jinks RN, Hoshi T, et al. A population-based study of KCNH7 p.Arg394His and bipolar spectrum disorder. Hum Mol Genet (2014) 23(23):6395–406. doi: 10.1093/hmg/ddu335
28. Kuo PH, Chuang LC, Liu JR, Liu CM, Huang MC, Lin SK, et al. Identification of novel loci for bipolar I disorder in a multi-stage genome-wide association study. Prog Neuropsychopharmacol Biol Psychiatry (2014) 51:58–64. doi: 10.1016/j.pnpbp.2014.01.003
29. Griswold AJ, Dueker ND, Van Booven D, Rantus JA, Jaworski JM, Slifer SH, et al. Targeted massively parallel sequencing of autism spectrum disorder-associated genes in a case control cohort reveals rare loss-of-function risk variants. Mol Autism (2015) 6:43. doi: 10.1186/s13229-015-0034-z
30. Kongsamut S, Kang J, Chen XL, Roehr J, Rampe D. A comparison of the receptor binding and HERG channel affinities for a series of antipsychotic drugs. Eur J Pharmacol (2002) 450(1):37–41. doi: 10.1016/S0014-2999(02)02074-5
31. Bartolomé-Martín D, Ibáñez I, Piniella D, Martínez-Blanco E, Pelaz SG, Zafra F. Identification of potassium channel proteins Kv7.2/7.3 as common partners of the dopamine and glutamate transporters DAT and GLT-1. Neuropharmacology (2019) doi: 10.1016/j.neuropharm.2019.03.011
32. Lebowitz JJ, Pino JA, Mackie PM, Lin M, Hurst C, Divita K, et al. Clustered Kv2.1 decreases dopamine transporter activity and internalization. J Biol Chem (2019) 294(17):6957–71. doi: 10.1074/jbc.RA119.007441
33. Amato D, Canneva F, Cumming P, Maschauer S, Groos D, Dahlmanns JK, et al. A dopaminergic mechanism of antipsychotic drug efficacy, failure, and failure reversal: the role of the dopamine transporter. Mol Psychiatry (2018) doi: 10.1038/s41380-018-0114-5
Keywords: schizophrenia, risperidone medications, KCNH7, treatment responses, pharmacogenomics
Citation: Wang X, Su Y, Yan H, Huang Z, Huang Y and Yue W (2019) Association Study of KCNH7 Polymorphisms and Individual Responses to Risperidone Treatment in Schizophrenia. Front. Psychiatry 10:633. doi: 10.3389/fpsyt.2019.00633
Received: 03 November 2018; Accepted: 06 August 2019;
Published: 30 August 2019.
Edited by:
Ming D. Li, Zhejiang University, ChinaReviewed by:
Bruce J. Kinon, Lundbeck, United StatesDavide Amato, Medical University of South Carolina, United States
Copyright © 2019 Wang, Su, Yan, Huang, Huang and Yue. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Weihua Yue, ZHJ5dWVAYmptdS5lZHUuY24=; Yu Huang, aHlAcGt1LmVkdS5jbg==
†These authors have contributed equally to this work