 Sandra Genís1
Sandra Genís1 Ronaldo L. A. Cerri2
Ronaldo L. A. Cerri2 Àlex Bach1,3Bruna F. Silper2Matheus Baylão2José Denis-Robichaud4Anna Arís1*
Àlex Bach1,3Bruna F. Silper2Matheus Baylão2José Denis-Robichaud4Anna Arís1*- 1Department of Ruminant Production, Institut de Recerca i Tecnologia Agroalimentàries, Barcelona, Spain
- 2Applied Animal Biology, Faculty of Land and Food Systems, University of British Columbia, Vancouver, BC, Canada
- 3Institució Catalana de Recerca i Estudis Avançats, Barcelona, Spain
- 4Department of Population Medicine, Ontario Veterinary College, University of Guelph, Guelph, ON, Canada
Metritis affects up to 40% of dairy cows and it is usually treated with antibiotics. In spite of their advantages, there is an increased concern about antibiotic resistance leading to the research of alternative methods. The aim of this study was to evaluate the effects of a combination of lactic acid bacteria (LAB) on the prevalence of metritis and modulation of endometrial and neutrophil inflammatory markers in dairy cows. One hundred and thirty-five cows were enrolled 3 week before calving and randomly assigned to three treatments. Treatment groups were: (1) two intravaginal doses of LAB/wk during 3 week pre-calving (vaginal, n = 45); (2) an intra-uterine dose, once 1 d after calving (uterine, n = 44); and (3) no intervention (CTRL, n = 45). Metritis was defined as body temperature > 39.5°C and purulent vaginal discharge (> 50% pus), and diagnosed 6 d after calving. Blood samples were taken at d −14, −10, −7, −4, +1, +3, +6, and +14 relative to calving for non-esterified fatty acids (NEFA) analysis. At d −10, +1, +3, and +6 neutrophils were isolated from blood for gene expression analysis by RT-qPCR. Endometrium biopsies were taken from 30 cows, 15 from CTRL and 15 from the uterine group at d +1, +3, and +6 after calving for pro-inflammatory markers analysis by NanoString®. Vaginal treatment reduced metritis prevalence (6/45) up to 58% compared with CTRL group (14/45), but there was no difference between the uterine and CTRL group. Uterine and vaginal treatments reduced blood neutrophil gene expression. Expression of pro-inflammatory markers in the endometrium did not differ between uterine and CTRL cows. Metritic cows expressed more C-X-C motif chemokine ligand 8 (CXCL8) and interleukin 1 beta (IL1B) at d 3 than healthy cows, whereas healthy cows expressed more CXCL8 at d 1 relative to calving in the endometrium. This study shows a promising potential of LAB probiotics as a preventive treatment against metritis in dairy cows.
Introduction
Acute metritis is an inflammation of the uterus due to bacterial infection occurring during the first 21 days after calving [with greater prevalence during the first 7 days in milk (DIM)] (1). The main characteristics are systemic signs of sickness, including fever over 39.5°C, red-brown watery foul-smelling vaginal discharge, dullness, inappetence, elevated heart rate, and low milk production (2). Damage to the uterine tissue during parturition leads to inflammation that likely contributes to the systemic condition (3). This inflammation in the uterus comes from the need for substantial tissue repair as part of post-partum involution characterized by the recruiting of immune cells (4). When microorganisms reach the uterus, the development of disease depends on the balance between the host defense systems, the virulence of the microbes, and the environmental bacterial load (5).
Traditional antimicrobial treatments against metritis are not very effective as only 67–77% of treated cows decrease fever after 5–10 days of treatment and there is not always resolution of the fetid odor (1). Furthermore, an improvement of the reproductive performance of the treated animals is not always found, especially in cases of sustained inflammation (1). The recommendations from the World Health Organization (6) to reduce antibiotic treatments leads to investigate alternative therapies.
The use of probiotic strains has been proposed as an alternative to prevent postpartum uterine infections and inflammation. A Canadian group (7) demonstrated that some lactic acid bacteria (LAB) strains were able to reduce uterine disease prevalence in dairy cows. Our recent work has also demonstrated that other LAB strains are able to modulate uterine inflammation and decrease Escherichia coli infection in vitro and ex vivo (8, 9). Interestingly, when LAB were administered intra-vaginally they altered the vaginal environment and tended to restrict the amount of bacteria reaching the uterus despite the fact that LAB did not reach the endometrium (10).
Periparturient dairy cows have nutritional requirements that exceed their dietary intake potential. This leads to a state of negative energy balance (NEB), during which body tissue reserves are mobilized to provide energy, hampering the resolution of peripartum diseases (11). Cows with a greater degree of NEB have higher concentrations of non-esterified fatty acids (NEFA) in blood, which are also associated with decreased feed intake (12, 13).
Parturition in dairy cattle is associated with impairment of neutrophil phagocytosis and oxidative burst activity (14), which leads to a decreased ability to fight bacterial infections. A study (11) demonstrated in vitro that an increase of NEFA in blood tended to desensitize neutrophil oxidative burst, which could compromise uterine health because specific phagocytosis by neutrophils is the most important component of uterine defense after bacterial infections (15). During inflammation, L-selectin (SELL), an adhesion molecule expressed on the neutrophil surface, promotes neutrophil infiltration into the infected tissue (16). When the neutrophils are in the infected tissue they recognize and bind pathogens and proceed to phagocytose and destroy them. One of the mechanisms they use is the respiratory burst which some of their components are the neutrophil cytosolic factor 1 (NCF1), a component of the NAPDH oxidase enzyme complex, and the superoxide dismutase (SOD) (17).
The idea behind this treatment was to study not only its effect on metritis prevalence but also to study the difference between an intra-uterine and intra-vaginal application and whether a more direct application of LAB to the endometrium would be able to down-regulate post-calving inflammation. In consequence, the aim of this study was to test two different approaches to apply a LAB combination in dairy cows (vaginal or uterine) to reduce metritis prevalence and to study the influence of LAB on NEFA concentrations, and gene expression of pro-inflammatory cytokines in blood neutrophils and endometrium.
Materials and Methods
Ethics Statement
This experiment was conducted in the facilities of the Dairy Research and Educational Centre from the University of British Columbia (UBC) in Agassiz, Canada. All experimental procedures were approved by the UBC Animal Care Committee.
Animals and Experimental Design
One hundred and thirty-five cows (45 primiparous and 90 multiparous) were enrolled 3 week before calving and randomly assigned to treatments using frequency matching to ensure similar frequencies for parity and previous illness in all treatment groups. The treatment groups were: (1) two intravaginal doses of lactic acid bacteria (LAB) per week during 3 week pre-calving (vaginal, n = 45); (2) one intra-uterine dose 1 day after calving (uterine, n = 45); and (3) no intervention (CTRL, n = 45). The average number of doses that the cows belonging to the vaginal treatment received was 5. The LAB treatment dose was a mixture of Lactobacillus rhamnosus CECT 278 (Colección Española de Cultivos Tipo, Valencia, Spain), Pediococcus acidilactici CECT 5,915, and Lactobacillus reuteri DSM 20016 (German collection of microorganisms and cell cultures, Leibniz, Germany) with a final cell count of 4.5 × 1010 CFU/dose and a proportion of 25/25/2, respectively. The LAB treatment was made fresh every day. Intravaginal LAB treatments were applied with a plastic cannula, previously cleaned with water and 70% ethanol, placed in the vagina just before the cervix. For the uterine treatment, 12 mL of LAB was applied with a sterile syringe directly inside the uterus after cleaning the exterior of the vagina with iodine. Aseptic procedures were maintained during LAB administration. A small sample from the endometrium was collected through a non-surgical procedure (1, 3, and 6 d after calving) from 30 cows randomly chosen (15 cows from the uterine treatment and 15 cows from the CTRL group). No cows from the vaginal treatment were biopsied because it had been previously shown that LAB treatment applied in the vagina does not reach the endometrium and does not modulate gene expression of pro-inflammatory cytokines (10). An epidural anesthesia was done using 100 mg of lidocaine (Lidocaine HCl 2%, Vetoquinol, Lavaltrie, QC, Canada). The vulva was cleaned, and a disinfected guarded biopsy instrument (crocodile-type biopsy forceps, made by Aries Surgical, Davis, CA, USA) was introduced via the cervix in the body of the uterus by vaginal (d 1) or trans-rectal (d 3, and 6) manipulations. Tissue collected was submerged in 500 μL of RNAlater (Thermo Fiser Scientific, Cramlington, UK) and kept overnight at 4°C. Then RNAlater was removed and the tissue stored at −80°C until further analysis.
Clinical Observations and Measurements
All cows were monitored clinically at 1, 3, and 6 DIM for metritis. Rectal temperature was measured twice and fever was defined as a temperature greater than 39.5°C. Retained placenta was diagnosed if a cow did not expel the placenta within 24 h after parturition. Cows with retained placenta were removed from the study as they were treated with antibiotics following the farm's standard procedures. A case of metritis was defined as a cow with reddish brown vaginal discharge, fetid odor, at least 50% pus and fever at 6 DIM. The Metricheck device, a soft rubber hemisphere connected to a stainless steel rod, was inserted into the vagina to assess the vaginal discharge. Vaginal discharge was evaluated after retracting the device caudally (18).
Sampling and Laboratory Analysis
Blood samples were collected from the coccygeal vein into 10 mL vacutainer tubes with ethylenediaminetetraacetic acid (EDTA, BD Vacutainer Systems, Plymuth, UK) on −14, −10, −7, −4, +1, +3, +6, and +14 d relative to calving. Blood samples were processed for neutrophil RNA isolation or centrifuged at 1,573 × g at 5°C for 15 min to separate plasma. Two aliquots were stored at −20°C until further analysis. Concentrations of NEFA were measured in plasma of d −10, +1, +3, and +6 using a commercially available kit (Wako Diagnostics, Mountain View, USA) within 2 months after sampling.
Bacterial Strains and Culture Conditions
Bacterial cultures were grown inoculating 1 mL of a glycerinate in 45 mL of Man, Rogosa, and Sharpe medium (MRS, Scharlau, Sentmenat, Spain) for L. rhamnosus, and P. acidilactici; and in 20 mL of MRS medium for L. reuteri at 37°C in static conditions for 2 days. Then bacteria were centrifuged at 6,000 × g for 15 min and re-suspended with sterile NaCl with a final concentration of 4.5 × 1010 CFU/dose. Each dose had a ratio of 25/25/2 for L. rhamnosus, P. acidilactici, and L. reuteri, respectively.
Neutrophil Isolation
Neutrophils were isolated from 20 mL of blood based on the procedures described by Carlson and Kaneko (19) and Farinacci et al. (20) from blood extracted at 10 d before calving, and at d +1, +3, +6 after calving. Specifically, blood was centrifuged at 1,000 × g for 45 min at 4°C, and then plasma, buffy coat, and two-thirds of the red blood pack were removed. Red blood cells in the remaining sample were lysed by adding 12 mL of hypotonic solution (10.6 mM Na2HPO4, and 2.7 mM NaH2PO4) and mixing for 90 s. Then, 6 mL of 4°C hypertonic solution (10.6 mM Na2HPO4, 2.7 mM Na2PO4, and 430 mM NaCl) were added, and tubes were mixed to restore isotonicity. Samples were centrifuged at 800 × g for 5 min at 4°C. The supernatant was discarded and the pellet was washed with 10 mL of Hank's Balanced Salt Solution (HBSS, Sigma-Aldrich, St. Louis, USA). Then samples were centrifuged again at 800 × g for 5 min at 4°C and the pellet was washed with 10 mL of HBSS. Samples were centrifuged at the same conditions and the pellet resuspended with 1 mL of HBSS and poured into 1.5 mL microcentrifuge tubes. The tubes were centrifuged at 9,600 × g for 15 min at 4°C, the supernatant was pipetted off, and 500 μL of TRIzol reagent (5Prima, Gaithesburg, USA) was added. Samples were stored at −20°C until RNA extraction.
Reverse Transcription and Quantitative PCR (RT-qPCR)
Total RNA was extracted from neutrophil isolations using TRIzol Reagent (5Prima, Gaithesburg, USA) following manufacturer's instructions and quantified using a Nanodrop instrument (Thermo Fisher Scientific, Cramlington, UK). The RNA was reverse transcribed to DNA using iScript cDna Synthesis kit (Bio-Rad, CA, USA) using 2 μL with a concentration of 50 ng/μL. Reactions of RT-qPCR were performed using CFX384 Touch Real-Time PCR Detection System (Bio-Rad, CA, USA) and RT-qPCR conditions for each set of primers were optimized (Table 1). The specificity of the amplification was evaluated by the single band identification at the expected molecular weight in DNA agarose gel and a single peak in the RT-qPCR melting curves. The efficiency was calculated by amplifying serial 1/10 dilutions of each gene amplicon. A standard curve at Ct vs. log was plotted to obtain the efficiency, which was calculated using the formula 101/slope, with an acceptable range of 1.8–2.05 (21). A total reaction volume of 20 μL was used, 10 μL of SYBER Green Fluoresecent (Bio-rad, California, USA), and the optimal primer concentration for each gene (Table 1). The RT-qPCR reactions were cycled as follows: an initial denaturalizing step of 10 min at 95°C, followed by 40 cycles of 10 s at 95°C, 15 s at optimized annealing temperature for each gene, 30 s at 72°C and a final extension of 10 min at 72°C. Relative gene expression was calculated using the Pfaffl method (22) with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and ribosomal protein S9 (RPS9) as reference gene controls. All RT-qPCR reactions were performed in triplicate. GAPDH and RPS9 were chosen because they are stably-expressed in a variety of cells, including uterine tissues (23, 24).
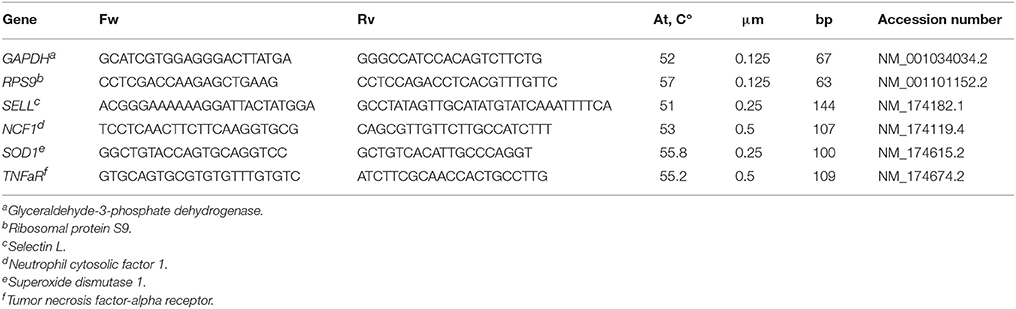
Table 1. Sequence, annealing temperatures (At), concentration (μM), amplicon size (bp) of forward (Fw), and reverse (Rv) bovine primers used for RT-qPCR and accession number for neutrophil gene expression (17).
Nanostring® nCounter Assay
Gene expression from biopsy samples was measured on the NanoString® nCounter Analysis System (NanoString® Technologies, Washington, USA). The system measures the relative abundance of each mRNA transcript of interest using a multiplex hybridization assay as digital readouts of fluorescent barcode probes that are hybridized to each transcript. An nCounter CodeSet (NanoString® Technologies, Washington, USA) containing a biotinylated capture prove for 4 genes and four reference genes (Table S1), and reporter proves attached to a color barcode tags according to the nCounter™ code-set design was hybridized in solution to 200 ng of total RNA for 18 h at 67°C according to the manufacturer's instructions.
Bioinformatics
To obtain gene expression data from the NanoString® nCounter assay, filtering of samples using quality control (QC) criteria was performed according to the manufacturer's recommendations. Row counts of QC-passed samples were normalized using four reference genes as internal controls: GAPDH, actin beta (ACTB), RPL19, and phosphoglycerate kinase 1 (PGK1).
Statistical Analysis of Results
All data were analyzed using SAS 9.2 software (SAS Institute Inc., Cary, NC). The number of animals that needed to be enrolled was determined with a power analysis. Metritis prevalence rate in the experimental farm was 33%, thus based on that, we hypothesized that the vaginal and/or the uterine treatment would be able to reduce the incidence rate by 15% (half). A power analysis using a nominal power of 85% at a 0.05% two-sided significant level indicated a minimum sample size of 45 (45 cows in each group) to detect treatment differences. One hundred fifty-eight cows were enrolled into this study, 7 cows were discarded because of retained placenta, 5 due to abortion, and 12 were culled resulting in 134 cows included in the statistical analysis.
Prior to statistical analysis, data were either log- or square root-transformed when necessary to achieve a normal distribution of the residuals. SELL, SOD1, NCF1, TNFaR, CXCL8, IL1B, IL6, and NEFA data was log-transformed while TNFa data was square root-transformed. Results herein are reported as the means of non-transformed data ± SEM obtained with normalized data (except otherwise indicated). Neutrophils' gene expression, blood NEFA concentration, and NanoString® data were analyzed using an ANOVA, considering treatment, parity, and day as fixed effects, and animal as a random effect. Three-way interaction between treatment, metritis, and day were tested on the NanoString® data. Stratified analysis was performed to evaluate differences in gene expression between metritic and healthy cows with NanoString® results, thus removing all data from treated cows (uterine treatment) and only considering those belonging to the CTRL group. Interaction between neutrophil gene expression and NEFA concentration were tested. Differences were considered significant when P < 0.05. Metritis prevalence was analyzed using a Chi-square comparing each treatment against CTRL cows.
Results
Metritis Prevalence
Cows in the vaginal treatment group (6/45, metritic cows/total) had a lower prevalence of metritis compared with cows belonging to the CTRL group (14/45, P = 0.04), whereas the uterine treatment (13/44) did not differ from the CTRL group (P = 0.87). Vaginal treatment lowered the prevalence of metritis by 58% compared with the CTRL group, resulting in a prevalence of 13.3%; whereas the CTRL group had a prevalence of metritis of 31.1%. Primiparous cows (16/44, metritic cows/total) had a greater (P = 0.02) metritis prevalence than multiparous cows (16/88).
Neutrophil Relative Gene Expression
The expression of SELL, NCF1, SOD1, and TNFaR genes was used as a marker of neutrophil potential killing activity (17). Treatment affected the expression of SELL, NCF1, SOD1, and TNFaR (P = 0.002, P = 0.014, P = 0.014 and P = 0.035, respectively; Table 2). The uterine treatment reduced 2.26-fold SELL, 2.08-fold NCF1, 1.62-fold SOD1, and 1.69-fold TNFaR gene expression compared with the CTRL, whereas vaginal treatment reduced SELL and TNFaR gene expression (1.72-fold, 1.22-fold, respectively), and had a tendency to reduce NCF1 (P = 0.056). When we analyzed the expression of each gene by treatment per day we observed that the uterine treatment tended to decrease SELL gene expression at day 1 and reduced it at day 3 and 6 compared with CTRL treatment (Figure 1A, P = 0.053, 0.035, and 0.007, respectively) and that the vaginal treatment tended to reduce SELL gene expression at day 6 (P = 0.092). NCF1 gene expression tended to decrease with the uterine and the vaginal treatments at day 1 (P = 0.057 and 0.098, respectively), were downregulated at day 3 (P = 0.007 and 0.004, respectively), and again, there was a tendency to decrease NCF1 expression at day 6 compared with CTRL treatment (Figure 1B, P = 0.093). The expression of SOD1 was reduced in the uterine treatment at days 1, and 6 and tended to be decreased at day 3 compared with CTRL and also compared with the vaginal treatment at day 6 (Figure 1C, P = 0.028, 0.096, 0.004, and 0.028, respectively). Uterine treatment reduced and the vaginal treatment tended to decrease the expression of TNFaR gene compared with the CTRL treatment (Figure 1D, P = 0.013 and 0.088, respectively).
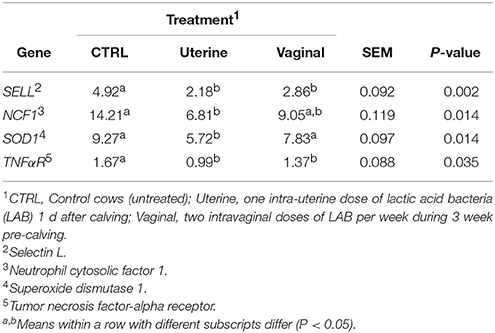
Table 2. Neutrophil gene expression by treatment on dairy cows (relative units of gene expression).
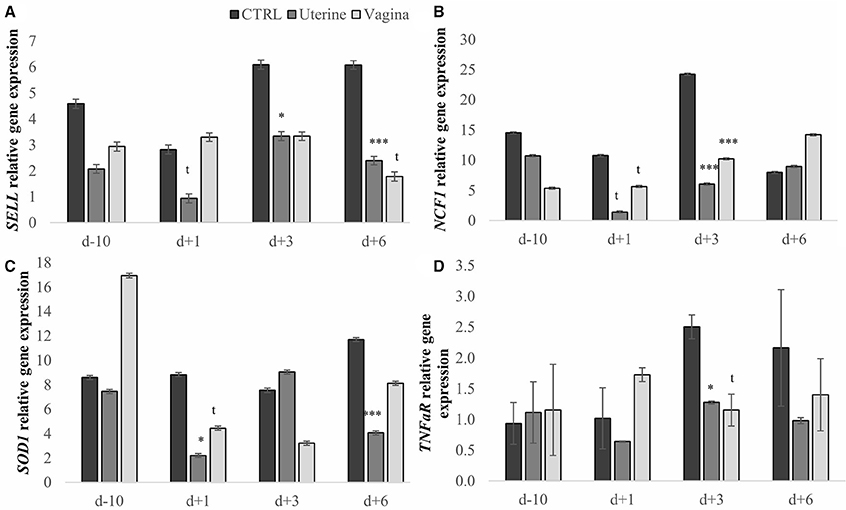
Figure 1. Neutrophil relative gene expression by treatment per day in dairy cows. Expression of selectin L (SELL, A), expression of neutrophil cytosolic factor 1 (NCF1, B), expression of superoxide dismutase 1 (SOD1, C), and expression of tumor necrosis factor-alpha receptor (TNFaR, D). CTRL, Control cows (untreated); Uterine, one intra-uterine dose of lactic acid bacteria (LAB) 1 day after calving; Vaginal, two intravaginal doses of LAB per week during 3 week pre-calving. Bars represent means ± SEM for different conditions. Bars with t (P < 0.1), *P < 0.05 or ***P < 0.005 differ from CTRL treatment. D-10, 10 days before calving; d1, day 1 after calving; d3, day 3 after calving; d6, day 6 after calving.
Metritis also affected the expression of TNFaR (P = 0.041, Table 3). Cows with metritis expressed 1.65-fold more TNFaR, and tended to express more SELL (P = 0.079) than healthy cows. Furthermore, day of sampling tended to affect the expression of NCF1 (P = 0.070, data not shown), because NCF1 expression was down-regulated at day 3 compared with day 1. No interaction was significant between treatment and metritis (SELL P = 0.548, SOD1 P = 0.298, NCF1 P = 0.651, and TNFaR P = 0.343), or day and metritis for the evaluated genes.
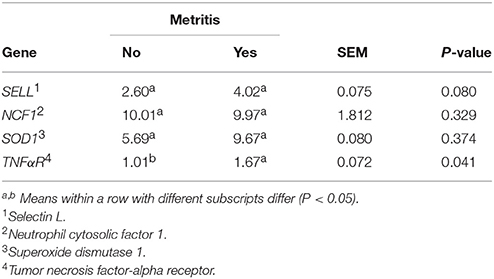
Table 3. Neutrophil gene expression as affected by presence or absence of metritis in dairy cows.
Non-esterified Fatty Acids (NEFA)
Blood NEFA concentration was associated with day (P < 0.01), and parity (P < 0.05), but not with treatment (P = 0.45). Interactions between treatment and parity (P < 0.01; Figure 2A), and parity and day (P < 0.01; Figure 2B) were significant; but no interaction between treatment and day (P = 0.48, Figure 2C) was observed. Blood NEFA concentration was higher in multiparous cows of the uterine group than in primiparous cows of both the uterine treatment and multiparous cows of CTRL group (Figure 2A). Blood NEFA concentration was 112.67 μM less before calving (d −10) than after calving (d +1, +3, +6) in primiparous cows, and was 239.57 μM higher after calving (d +1, +3, +6) than before calving (d −10) in multiparous cows. No differences were observed between heifers and cows before calving, but after calving, multiparous cows had at least 1.20-fold higher NEFA concentration in blood than primiparous cows. No differences in NEFA concentrations were found between metritic and healthy cows (data not shown). There were low or no correlations between neutrophil gene expression (SELL, NCF1, SOD1, and TNFaR) and blood NEFA concentrations. The r2 were 0.02, 0.04, 0.02, and 0.01 respectively with P- values of 0.10, 0.03, 019, and 0.32.
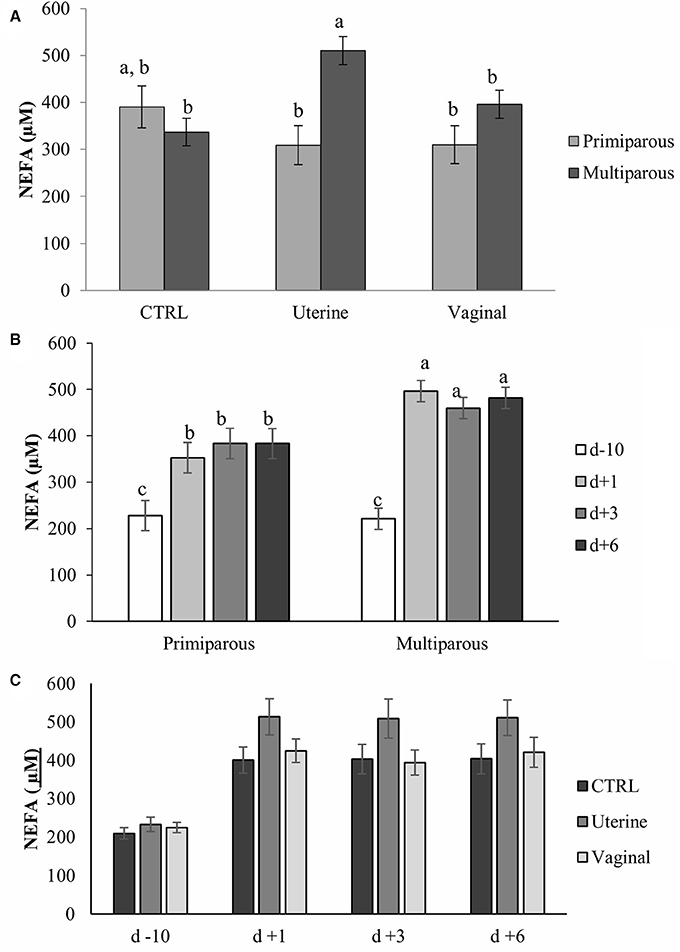
Figure 2. Blood non-esterified fatty acids (NEFA) concentrations by treatment and parity (A), by parity and day (B), and by treatment and parity (C) in dairy cows. Bars represent mean ± SE. Bars with different letters differ (P < 0.05). CTRL, Control cows (untreated); Uterine, one intra-uterine dose of lactic acid bacteria (LAB) 1 day after calving; Vaginal, two intravaginal doses of LAB per week during 3 week pre-calving; D-10, 10 days before calving; d1, day 1 after calving; d3, day 3 after calving; d6, day 6 after calving.
Pro-inflammatory Markers
The expression of genes coding for the pro-inflammatory cytokines C-X-C motif chemokine ligand 8 (CXCL8), interleukine 1 beta (IL1B), interleukin 6 (IL6), and tumor necrosis factor-alpha (TNFa) was evaluated as markers of inflammation in the endometrium. The gene expression of these cytokines was affected by time (P < 0.01), but not by treatment or metritis. No interactions were found between treatment and metritis, metritis and time, time and treatment. Nor was the triple interaction among treatment, metritis, and time.
As treatment did not affect the expression of pro-inflammatory profile, a second analysis was performed to evaluate differences in gene expression between 6 metritic and 6 healthy cows, only using cows in the CTRL group. The gene expression of CXCL8 was affected by day (P < 0.01), and there was an interaction between metritis and day (P < 0.05; Figure 3A). Healthy cows tended to express more CXCL8 at day 1 (P = 0.08, 2.30-fold) whereas metritic cows tended to express CXCL8 (P = 0.07) by 2.45-fold at day 3 compared with healthy cows. The gene expression of IL1B was affected by day (P < 0.01) and there was an interaction between day and metritis (P < 0.05; Figure 3B). Metritic cows tended (P = 0.053) to have a 3.05-fold greater expression of IL1B gene at day 3 compared with healthy cows. No differences were found at days 1 and 6. The expression of IL6 and TNFa was affected by day (P < 0.05), but no differences were observed between healthy and metritic cows (Figures 3C,D).
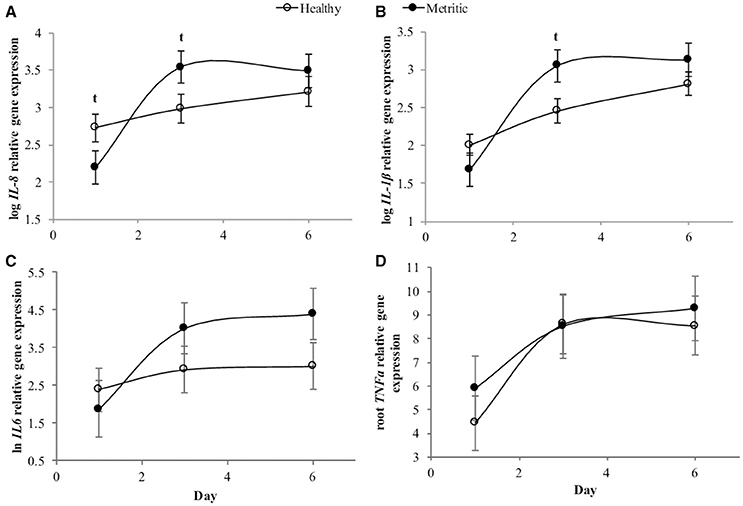
Figure 3. Cytokine gene expression from endometrial biopsies taken from healthy (open circles) and metritic dairy cows (solid circles) from the CTRL treatment. Expression of C-X-C motif chemokine ligand 8 (CXCL8) gene over time (A), interleukine 1 beta (IL1B, B), interleukin 6 (IL6, C), and tumor necrosis factor- alpha (TNFa, D). Points represent means ± SEM for different conditions. Points with t differ (P < 0.1) between healthy and metritic cows.
Discussion
The main objective of this study was to evaluate the capacity of a combination of LAB strains which previously showed to reduce E. coli infection in vitro and ex vivo (8), to modulate inflammation and reduce the prevalence of metritis after intra-vaginal or intra-uterine administration. Previous work by other authors showed that intra-vaginal administration of other LAB strains was able to reduce uterine infections in dairy cows (7). Most of the probiotics currently used in animals are LAB, especially in the case that they are used to try to modulate infection and/or inflammation. In our study, the intra-vaginal application of LAB twice per week during the 3 week before calving was able to reduce the prevalence of metritis compared with the CTRL cows. Recent studies had characterized the uterine microbiota from cows (25, 26) showing a different microbial community from metritic or healthy cows. On the first study (27) they found that Fusobacteria was the dominant group in metritic cows, while Gamaproteobacteria was more present in the uterus of healthy cows. The second study (28) also observed an increase abundance of Fusobacteria in metritic cows along with and increment of Bacteroidetes. Even then, E. coli is still being described as the main pathogen responsible for initiating uterine infection and inflammation, unbalancing the uterine microbiota and favoring the infection of other pathogens such as Bacteroides spp. or Fusobacterium necrophorum (28). Also, it has been previously shown that cows treated intra-vaginally with this combination of LAB tended to have lesser presence of E. coli in the vagina than not-treated cows (P = 0.09) (10). These findings could explain partially the results observed herein.
On the other hand, the intra-uterine application of the same dose of LAB did not show any difference (P = 0.872) in metritis prevalence compared to the CTRL group.
In this study, neutrophil gene expression was quantified as part of the inflammatory response. It is known that neutrophils, are a primary source of immune response in the uterus, with the neutrophil activity being associated with risk of metritis (12). Some authors (29) have stated that a desirable response should be a prompt, substantial flux of neutrophils into the uterus after calving. On the other hand, an excessive immune response could be counter-productive as it impairs uterine or ovarian function (15). In the current study metritic cows tended (P = 0.079; Table 2) to express more SELL (l-selectin), a protein that is expressed on the neutrophil surface and promotes neutrophil infiltration into the infected tissue, compared with healthy cows. Furthermore, metritic cows expressed more TNFaR [the TNF-α receptor that is involved in regulating neutrophil functions such as degranulation, release of oxygen metabolites, phagocytosis and adhesion to endothelium (30)] than healthy cows (P = 0.041; Table 3). In LAB-treated cows, the vaginal and uterine treatments down-regulated the expression of all genes studied as markers of neutrophil activity compared with CTRL cows (Table 2). Specifically, the expression of SELL and TNFaR was reduced along with the expression of NCF1 and SOD, both involved in the respiratory burst. In the vaginal treatment, the decrease of neutrophil activity could probably be related to a reduction of pathogenic bacteria in the vagina and a consequent reduction in the incidence of infection in the uterus. There were no differences between CTRL and uterine cows in the expression of CXCL8 nor with the other cytokines IL1B, IL6, and TNFa. This was an unexpected result because previous in vitro and ex vivo studies indicated that a direct treatment of the endometrium with LAB, modulated the levels of these cytokines (9). However, the uterine treatment was associated with a reduction in neutrophil gene expression; but probably the dose was not sufficient to modulate the expression of proinflammatory cytokines, or the metritis prevalence.
The differences between a pathological and physiological inflammation is not well understood and depends on the severity, timing and duration of inflammation (15). It is widely accepted that all early post-partum cows are in a pro-inflammatory state (3) and that damage to uterine tissue during calving leads to inflammation that likely contributes to the systemic condition. In this study, when the profile of pro-inflammatory cytokines was analyzed only in CTRL animals the results showed that metritic cows had impaired expression of CXCL8 at day 1 compared with healthy cows, but at day 3 this situation reversed, with CXCL8 and IL1B being over-expressed in metritic compared with healthy cows (Figure 3). It seems that cows with a less active immune system are more susceptible to bacterial infections [which is present in almost all dairy cows during the 2–3 week after calving (31)] ultimately resulting in metritis, as the immune systems reacts more slowly.
A previous study Hammon et al. (12) found that cows with puerperal metritis had greater plasma NEFA levels pre-partum than healthy cows. In the current study, no differences in blood NEFA concentration between healthy and metritic cows were observed (P = 0.183). The inconsistent results may be due to different timeline used to diagnose metritis. In the present study, metritis was diagnosed when temperature rose > 39.5°C along with the presence of vaginal purulent discharge at the 1st week postpartum, whereas others (12) classified cows as metritic if purulent discharge was observed between day 0 and 14 after parturition, regardless of presence of fever.
In the current study, multiparous cows receiving the uterine treatment had an increase of blood NEFA concentrations, which indicates a larger NEB in this group of cows compared with multiparous cows from the CTRL and vaginal groups. However, differences in NEB could not be attributed to changes in body condition or milk production since no differences in milk yield at 60 DIM (P = 0.55; data not shown) and BW between d −10 to d +21 related to calving were observed between treatments (P = 0.96; data not shown). Thus, as cows in the uterine treatment modified neutrophil gene expression (Table 2) that may indicate that LAB is affecting the energy metabolism either directly or indirectly consequence of a potentially greater energy consumption from the immune system (32, 33). It has been previously reported that high blood NEFA concentration is associated with impaired blood neutrophil functions (11, 12). Herein, we have not found any correlation between NEFA and the expression of some genes related to neutrophil activity, but we cannot discard other relationships with total neutrophil activity, not measured in the current study. It could be speculated that with an impaired neutrophil activity, the endometrium is recruiting more immune cells to sustain the involution processes, increasing the NEB of the animal, but this hypothesis could not be confirmed because total immune cell counts were not performed in this study neither activity assays.
Conclusions
The vaginal application of the combination of lactic acid bacteria used herein reduced metritis prevalence in dairy cows compared with untreated animals; whereas, the prevalence of metritis was the same between untreated cows and cows treated with LAB intra-uterine. The gene expression of pro-inflammatory cytokines was down-regulated in circulating neutrophils from intra-vaginal and intrauterine treatments with lactic-acid bacteria, but no change was observed in the post-partum endometrium. The combination of lactic acid bacteria administration prior to calving appears to be a promising strategy to prevent uterine infections.
Author Contributions
SG, AA, and RC: Design of the study; SG and MB: LAB preparation; SG, BS, JD-R, and MB: Collection of health data and blood; BS and JD-R: Collection of endometrial biopsies; SG: Laboratory analysis and statistical analysis; SG, AA, RC, and ÀB: Interpretation of the data and drafting of the manuscript. All authors read and approved the final manuscript.
Funding
This study was supported by a contribution from the Natural Sciences and Engineering Research Council (NSERC), the Dairy Industry Research and Education Committee (DIREC, British Columbia Dairy Association), and IRTA (Institut de Recerca i Tecnologies Agroalimentàries).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Nelson Dinn and all students that assisted with data collection at the University of British Columbia Dairy Education and Research Centre. This study has been included in the thesis of SG and it can be accessed online under the university policy (31).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2018.00135/full#supplementary-material
Abbreviations
LAB, lactic acid bacteria.
References
1. LeBlanc SJ. Postpartum uterine disease and dairy herd reproductive performance: a review. Vet J. (2008) 176:102–14. doi: 10.1016/j.tvjl.2007.12.019
2. Sheldon IM, Lewis GS, LeBlanc S, Gilbert RO. Defining postpartum uterine disease in cattle. Theriogenology (2006) 65:1516–30. doi: 10.1016/j.theriogenology.2005.08.021
3. Bradford BJ, Yuan K, Farney JK, Mamedova LK, Carpenter AJ. Invited review: inflammation during the transition to lactation: new adventures with an old flame. J Dairy Sci. (2015) 98:6631–50. doi: 10.3168/jds.2015-9683
4. Herath S, Fischer DP, Werling D, Williams EJ, Lilly ST, Dobson H, et al. Expression and function of toll-like receptor 4 in the endometrial cells of the uterus. Endocrinology (2006) 147:562–70. doi: 10.1210/en.2005-1113
5. Sheldon IM, Cronin JG, Healey GD, Gabler C, Heuwieser W, Streyl D, et al. Innate immunity and inflammation of the bovine female reproductive tract in health and disease. Reproduction (2014) 148:R41–51. doi: 10.1530/REP-14-0163
6. Deng Q, Odhiambo JF, Farooq U, Lam T, Dunn SM, Ametaj BN. Intravaginal lactic acid bacteria modulated local and systemic immune responses and lowered the incidence of uterine infections in periparturient dairy cows. PLoS ONE (2015) 10:e124167. doi: 10.1371/journal.pone.0124167
7. Deng Q, Odhiambo JF, Farooq U, Lam T, Dunn SM, Gänzle MG, et al. Intravaginally administered lactic acid bacteria expedited uterine involution and modulated hormonal profiles of transition dairy cows. J Dairy Sci. (2015) 98:6018–28. doi: 10.3168/jds.2014-8559
8. Genís S, Bach À, Fàbregas F, Arís A. Potential of lactic acid bacteria at regulating Escherichia coli infection and inflammation of bovine endometrium. Theriogenology (2016) 85:625–37. doi: 10.1016/j.theriogenology.2015.09.054
9. Genís S, Sánchez-Chardi A, Bach À, Fàbregas F, Arís A. A combination of lactic acid bacteria regulates Escherichia coli infection and inflammation of the bovine endometrium. J Dairy Sci. (2017) 100:479–92. doi: 10.3168/jds.2016-11671
10. Genís S, Bach À, Arís A. Effects of intravaginal lactic acid bacteria on bovine endometrium: implications in uterine health. Vet Microbiol. (2017) 204:174–9. doi: 10.1016/j.vetmic.2017.04.025
11. Ster C, Loiselle M-C, Lacasse P. Effect of postcalving serum nonesterified fatty acids concentration on the functionality of bovine immune cells. J Dairy Sci. (2012) 95:708–17. doi: 10.3168/jds.2011-4695
12. Hammon DS, Evjen IM, Dhiman TR, Goff JP, Walters JL. Neutrophil function and energy status in Holstein cows with uterine health disorders. Vet Immunol Immunopathol. (2006) 113:21–9. doi: 10.1016/j.vetimm.2006.03.022
13. Eger M, Hussen J, Drong C, Meyer U, von Soosten D, Frahm J, et al. Impacts of parturition and body condition score on glucose uptake capacity of bovine monocyte subsets. Vet Immunol Immunopathol. (2015) 166:33–42. doi: 10.1016/j.vetimm.2015.04.007
14. Hoeben D, Monfardini E, Opsomer G, Burvenich C, Dosogne H, De Kruif A, et al. Chemiluminescence of bovine polymorphonuclear leucocytes during the periparturient period and relation with metabolic markers and bovine pregnancy-associated glycoprotein. J Dairy Res. (2000) 67:249–59. doi: 10.1017/S0022029900004052
15. LeBlanc SJ. Interactions of metabolism, inflammation, and reproductive tract health in the postpartum period in dairy cattle. Reprod Domest Anim. (2012) 47(Suppl. 5):18–30. doi: 10.1111/j.1439-0531.2012.02109.x
16. Abbas AK, Lichtman AH. Cellular and Molecular Immunology, 5th Edn. Philadelphia, PA: Saunders (2005).
17. Hengst BA, Nemec LM, Rastani RR, Gressley TF, Larson R, Ziegler D, et al. Effect of conventional and intensified milk replacer feeding programs on performance, vaccination response, and neutrophil mRNA levels of Holstein calves. J Dairy Sci. (2012) 95:5182–93. doi: 10.3168/jds.2011-5261
18. McDougall S, Macaulay R, Compton C. Association between endometritis diagnosis using a novel intravaginal device and reproductive performance in dairy cattle. Anim Reprod Sci. (2007) 99:9–23. doi: 10.1016/j.anireprosci.2006.03.017
19. Carlson GP, Kaneko JJ. Isolation of leukocytes from bovine peripheral blood. Proc Soc Exp Biol Med. (1973) 142:853–6. doi: 10.3181/00379727-142-37131
20. Farinacci M, Colitti M, Sgorlon S, Stefanon B. A technique to screen plant extracts for anti-inflammatory activity on ovine neutrophils. Ital J Anim Sci. (2010) 6:548–50. doi: 10.4081/ijas.2007.1s.548
21. Domènech A, Parés S, Bach A, Arís A. Mammary serum amyloid A3 activates involution of the mammary gland in dairy cows. J Dairy Sci. (2014) 97:7595–605. doi: 10.3168/jds.2014-8403
22. Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. (2001) 29:e45. doi: 10.1093/nar/29.9.e45
23. Wathes DC, Cheng Z, Chowdhury W, Fenwick MA, Fitzpatrick R, Morris DG, et al. Negative energy balance alters global gene expression and immune responses in the uterus of postpartum dairy cows. Physiol Genomics (2009) 39:1–13. doi: 10.1152/physiolgenomics.00064.2009
24. Galvão KN, Felippe MJB, Brittin SB, Sper R, Fraga M, Galvão JS, et al. Evaluation of cytokine expression by blood monocytes of lactating Holstein cows with or without postpartum uterine disease. Theriogenology (2012) 77:356–72. doi: 10.1016/j.theriogenology.2011.08.008
25. Machado VS, Oikonomou G, Bicalho MLS, Knauer WA, Gilbert R, Bicalho RC. Investigation of postpartum dairy cows' uterine microbial diversity using metagenomic pyrosequencing of the 16S rRNA gene. Vet Microbiol (2012) 159:460–9. doi: 10.1016/j.vetmic.2012.04.033
26. Santos TMA, Bicalho RC. Diversity and succession of bacterial communities in the uterine fluid of postpartum metritic, endometritic and healthy dairy cows. PLoS ONE (2012) 7:e53048 doi: 10.1371/journal.pone.0053048
27. Santos TM, Gilbert RO, Bicalho RC. Metagenomic analysis of the uterine bacterial microbiota in healthy and metritic postpartum dairy cows. J Dairy Sci. (2011) 94:291–302. doi: 10.3168/jds.2010-3668
28. Bicalho MLS, Machado VS, Higgins CH, Lima FS, Bicalho RC. Genetic and functional analysis of the bovine uterine microbiota. Part I: metritis versus healthy cows. J Dairy Sci. (2017) 100:3850–62. doi: 10.3168/jds.2016-12058
29. Gilbert RO, Santos NR, Galvao KN, Brittin SB, Roman HB. The relationship between postpartum uterine bacterial infection (BI) and subclinical endometritis (SE). J Dairy Sci. (2007) 90:469.
30. Lopez S, Halbwachs-Mecarelli L, Ravaud P, Bessou G, Dougados M, Porteu F. Neutrophil expression of tumour necrosis factor receptors (TNF-R) and of activation markers (CD11b, CD43, CD63) in rheumatoid arthritis. Clin Exp Immunol. (1995) 101:25–32. doi: 10.1111/j.1365-2249.1995.tb02272.x
31. Sheldon IM. The postpartum uterus. Vet Clin North Am Food Anim Pract. (2004) 20:569–91. doi: 10.1016/j.cvfa.2004.06.008
32. Martin LB II, Scheuerlein A, Wikelski M. Immune activity elevates energy expenditure of house sparrows: a link between direct and indirect costs? Proc R Soc Lond B (2003) 270:153–8. doi: 10.1098/rspb.2002.2185
Keywords: intra-vaginal, intra-uterine, lactic acid bacteria, metritis, neutrophil
Citation: Genís S, Cerri RLA, Bach À, Silper BF, Baylão M, Denis-Robichaud J and Arís A (2018) Pre-calving Intravaginal Administration of Lactic Acid Bacteria Reduces Metritis Prevalence and Regulates Blood Neutrophil Gene Expression After Calving in Dairy Cattle. Front. Vet. Sci. 5:135. doi: 10.3389/fvets.2018.00135
Received: 09 April 2018; Accepted: 04 June 2018;
Published: 21 June 2018.
Edited by:
Subhash Verma, Chaudhary Sarwan Kumar Himachal Pradesh Krishi Vishvavidyalaya, IndiaReviewed by:
Pascal Rainard, Institut National de la Recherche Agronomique (INRA), FranceCarol Geralyn Chitko-McKown, U.S. Meat Animal Research Center (ARS-USDA), United States
Copyright © 2018 Genís, Cerri, Bach, Silper, Baylão, Denis-Robichaud and Arís. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anna Arís, YW5uYS5hcmlzQGlydGEuY2F0