 Attila D. Sándor
Attila D. Sándor Cristian Domșa4
Cristian Domșa4 Áron Péter
Áron Péter Sándor Hornok
Sándor Hornok- 1HUN-REN-UVMB Climate Change: New Blood-Sucking Parasites and Vector-Borne Pathogens Research Group, Budapest, Hungary
- 2Department of Parasitology and Zoology, University of Veterinary Medicine, Budapest, Hungary
- 3STAR-UBB Institute, Babes-Bolyai University, Cluj-Napoca, Romania
- 4Romanian Ornithological Society, Cluj Napoca, Romania
Bats in the Western Palearctic are host for diverse array of ectoparasites, including three ixodid ticks (Ixodes ariadnae, I. simplex, and I. vespertilionis), which are highly specialized to parasitize these mammals. In this study we collected and analyzed 3,965 host-tick records across 31 bat species from published literature, online sources, and unpublished field data. Individual bat-specialist ticks showed distinct host preferences, with cave-dwelling bats accounting for over 90% of all records. Ixodes vespertilionis was the most generalist of them, with a broad host range and distribution, while I. simplex was highly host-specific, primarily parasitizing a single host species, Miniopterus schreibersii. Ixodes ariadnae had a similar host spectrum as I. vespertilionis but more restricted geographical range, likely influenced by seasonal and life history factors. Our findings revealed substantial geographical overlap in tick distributions across Central and Eastern Europe. Free-living tick stages were predominantly found in caves, and males were observed more frequently than females. Non-bat specific, as well generalist ticks such as Ixodes ricinus and Rhipicephalus sanguineus s.l. were rare on bats, with larger bat species being the more common hosts. These ticks may host DNA of several bacterial, viral, and parasitic pathogens, suggesting an important role in pathogen transmission to bats and possibly other mammals. This study underscores the ecological significance of bat-specialist ticks and highlights the need for further research on their distribution, host interactions, and role in zoonotic disease transmission.
Background
Ixodid ticks (Acari: Ixodidae) are obligate parasites of vertebrates, widely distributed across all terrestrial biomes of Earth (1). They are an ancient group, showing long coevolution with vertebrates, initially being the parasites of feathered dinosaurs/birds (2), later evolving to infest all terrestrial vertebrate groups (3). Currently, there are over 700 valid species, with high diversity in the tropics (4). Most species are specialized to feed either on birds, mammals or reptiles, however, several species are generalists, capable of feeding on most available terrestrial vertebrates in their habitats. In contrast, some species exhibit strict host specificity, adapting to feed on a single or very few host species (5, 6). Most ixodid ticks use two or three different hosts throughout their life cycle, with each developmental stage taking a single blood meal (with the exception of males). They attach to the hosts skin, penetrate it using their hypostome and chelicerae, then extract blood from the host, through a process called engorgement. Fully engorged ticks detach from the host in specific areas, they molt into the next development stage (larva to nymph to adult) or lay eggs (females) and die. Throughout this process, ticks may transmit pathogens (viral, bacterial or protozoan) between hosts, playing a crucial role in the epidemiology of vector-borne diseases (7, 8). Ticks are likely the most important vectors of pathogens in the temperate regions and show constant adaptation to changing climatic and biotic conditions, thus being in the forefront of zoonotic disease emergence (9).
Bats are among the most widespread terrestrial mammals, with high mobility and species diversity and they are important ecosystem service providers, too (8). They also may serve as important reservoir hosts for a wide range of pathogens, including viruses, bacteria, and parasites, some of which have the potential to spill over into livestock or human populations and cause emerging infectious diseases (10).
Recent studies of bat associated ectoparasites showed that these may carry DNA of a diverse array of viral, bacterial or protozoan pathogens, some with proven zoonotic character (11), although most remain uncharacterized (12–15). Among these, DNA of several pathogenic bacteria was identified in bat specialist ticks in Europe, Africa but also in the New World (16). In addition, ixodid ticks of Palearctic bats were suggested to play a role in the cycles of several groups of protozoa (17) and viruses (18). Two of the three bat specialist ticks occurring in the Western Palearctic are known to attack humans as well (19, 20). Furthermore, research on bat ticks is important from a taxonomic point of view, as reflected by the descriptions of six new bat-specialist ticks from Europe and Asia during the last decade (21–25), while current assessments are neglected in the region (26). In conclusion, the knowledge of bat-tick relations may provide valuable insights into the mechanisms driving host–parasite interactions and the importance of bat and tick populations in the ecology and spatial evolution of pathogens they may harbor. Here we intend to construct a general spatial distribution of hard ticks hosted by bats in the Western Palearctic, using georeferenced occurrences (mostly published in literature, but also from databases and some unpublished, own records) of specialist and generalist ixodid ticks registered on bats (or in case of bat specialist ticks in bat roosts). In addition, we intend to characterize the role of both the host-, as well the tick ecology may play in building these relations, with a special focus on their role in vector-borne pathogen spread.
Methods
Database creation
Our methodology followed a three-step process. First, a keyword search was performed using terms as: ‘ticks’ or ‘Ixodidae’ + ‘bats’, + ‘Western Palearctic’, or ‘Ixodes ariadnae’/‘Ixodes simplex’ and ‘Ixodes vespertilionis’ + ‘Western Palearctic’ in the following literature databases: PUBMED, Web of Science and Google Scholar. In the next step, duplicates were eliminated, and abstracts were verified to contain relevant data. This process resulted in a database of suitable papers. Subsequently, copies of the original publications were obtained and the references cited in these works were traced. This process was repeated until no new references were found. In the third step we extracted each individual host-tick record from the references, noting the location, date, host and parasite species, development stage (for ticks) and pathogen (if) mentioned. To complete the collated records, we traced museum specimen collections and observation records using data repositories like Global Biodiversity Information Facility,1 Obervation.org and NBN Atlas,2 among others. Direct internet searches using the same keywords also provided hits, verified by photos of the tick species. Unpublished data from our field studies in Algeria, Bulgaria, Hungary, and Romania (2019–2023) were also included. These records were introduced into a database and individually georeferenced to create distribution maps.
Distribution maps
For the maps, we overlaid the range of each host species with the presence data for each tick species. Each host range was set with transparency, so the more ranges overlapped, the more intense the range color appeared—a proxy for multiple host species presence. For the primary bat host species, we used freely available shapefiles from the International Union for Conservation of Nature (IUCN) Red List (27). IUCN ranges were used previously primarily for conservation biology of bats (28) or other mammals (29), but also for establishing the relationships between bats and argasid soft ticks (30), as well for bats’ insect ectoparasites and vectored pathogens (31). In the next step, we intersected these ranges with the contour of the Western Palearctic, which was delimited according to previously published borders (30, 32, 33).
Host–parasite relationships
Using the database, we mapped each host–parasite relationship and classified hosts as primary or accidental. To determine primary or accidental hosts of any ixodid tick species, we applied an arbitrary rule: any bat species with more than 5.0% of the records for a particular tick species was considered a primary host. Hosts with fewer than 5.0% of cumulative records for a particular tick species were considered non-primary or accidental hosts, following a system previously proposed for bat-fly associations (34–36). Additional host-related information, such as roosting sites or reproductive stages, was also extracted from the primary publications where available. Hosts were assigned either into cave-dwelling, or crevice dwelling group, based on their roosting preferences in their active period (37).
Results
In total, 507 published references were included in the primary reference database (Supplementary Table S1), of which 317 contained records of bat ticks. Additionally, 27 records were extracted from online sources, supplemented by 207 unpublished host–parasite records from the authors. The bat host—tick reference database contains 3,965 individual entries (Supplementary Table S1), with the three bat-specialist ticks making up to 97.3% of the total (Table 1), while ticks with generalist host selection were recorded on bats in 110 instances (Supplementary Table S1). The complete database contains 3,855 entries of bat specialist ticks (8,997 individual ticks), collected from 3,162 individual bat hosts (5,680 ticks), together with a total of 730 instances of ticks collected from the environment (3,225 individuals of unengorged, free ticks, collected generally from underground roosts’ walls), while collection circumstances were unknown for 92 cases (n = 92 ticks, only tick species and geographic location were recorded). Altogether 31 bat species were recorded to host bat-specialist ticks, with most records noted for I. vespertilionis (Table 1). For 24 cases, records mentioned only generic ‘Chiroptera,’ while 10 cases were assigned to either Myotis spp., Pipistrellus spp., or Plecotus spp. Only 2 cases (0.005% of all records) involved bat ticks found on non-bat hosts—both on humans. Genetic analysis of previous blood meals identified nine cases of non-bat hosts across two tick species (all host species are listed in Table 2).
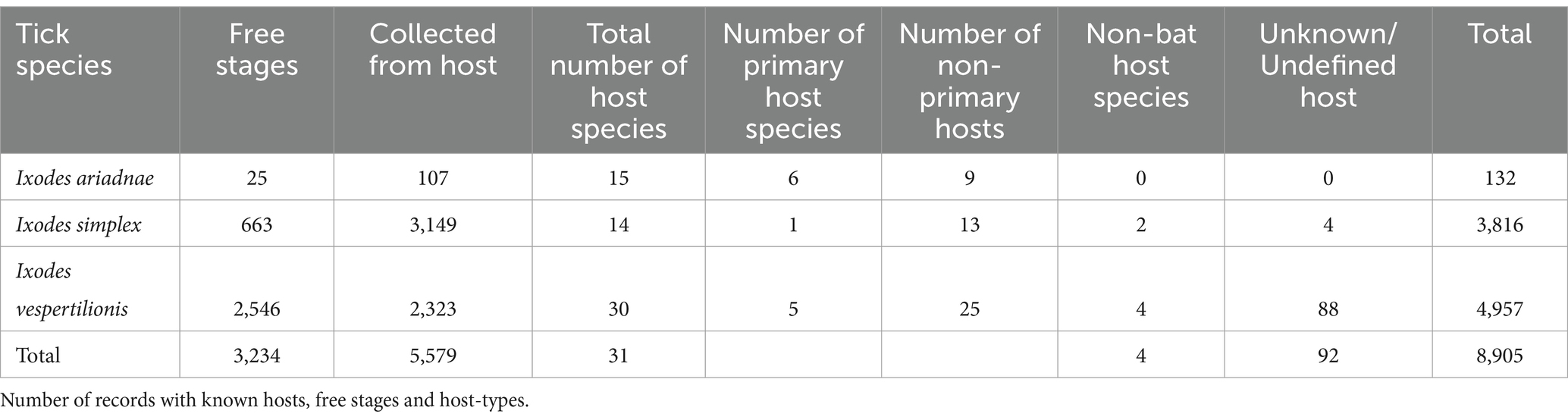
Table 1. Bat-specialist ticks recorded in the Western Palearctic.
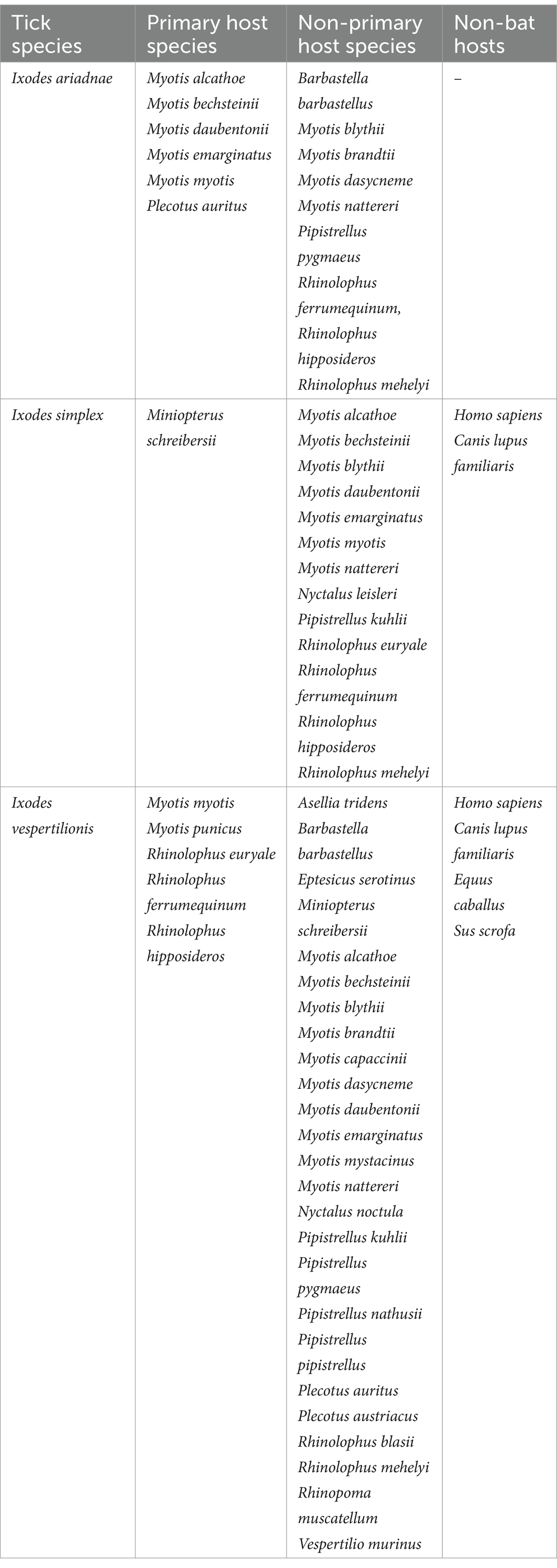
Table 2. Primary and non-primary bat host species of hard ticks (Ixodidae) in the Western Palearctic.
Ixodes vespertilionis had the most diverse host spectrum, with 30 different host species (5 primary and 25 non-primary hosts). Ixodes ariadnae had the most primary hosts (6), while I. simplex had a single primary host harboring 98.43% of all records. Most ticks were recorded on cave-dwelling bat species (93.6%), with a single species (I. ariadnae) occurring regularly on crevice-dwelling bat hosts (these include species which rely on rock-crevices, but also tree-hole roosting ones).
Host-collected ticks were mainly subadult stages (90.2%), together with 537 adult females (9.7%) and 10 males collected from hosts (only in case of I. vespertilionis were males found on bats). The different tick species showed distinct host selection, with small overlap in host palette, mainly among hosts of I. ariadnae and I. vespertilionis (Figure 1). Free-stages of bat-specialist ticks were represented mainly by adults and were dominated by I. vespertilionis (675 individual records of 2,547 ticks, 78.7% of all free ticks), with a highly biased sex ratio toward males (1–2.32). Records of free individuals for the other two species are rare or accidental (Table 1). Ixodes simplex showed highly aggregated off-host presence (a single visit to a site used as nursery colony by M. schreibersii in the previous year resulted in 554 ticks collected from a crevice with an area of ca. 0.02 sqm, involving all tick developmental stages and sexes), but altogether only 23 instances of free individuals are known.
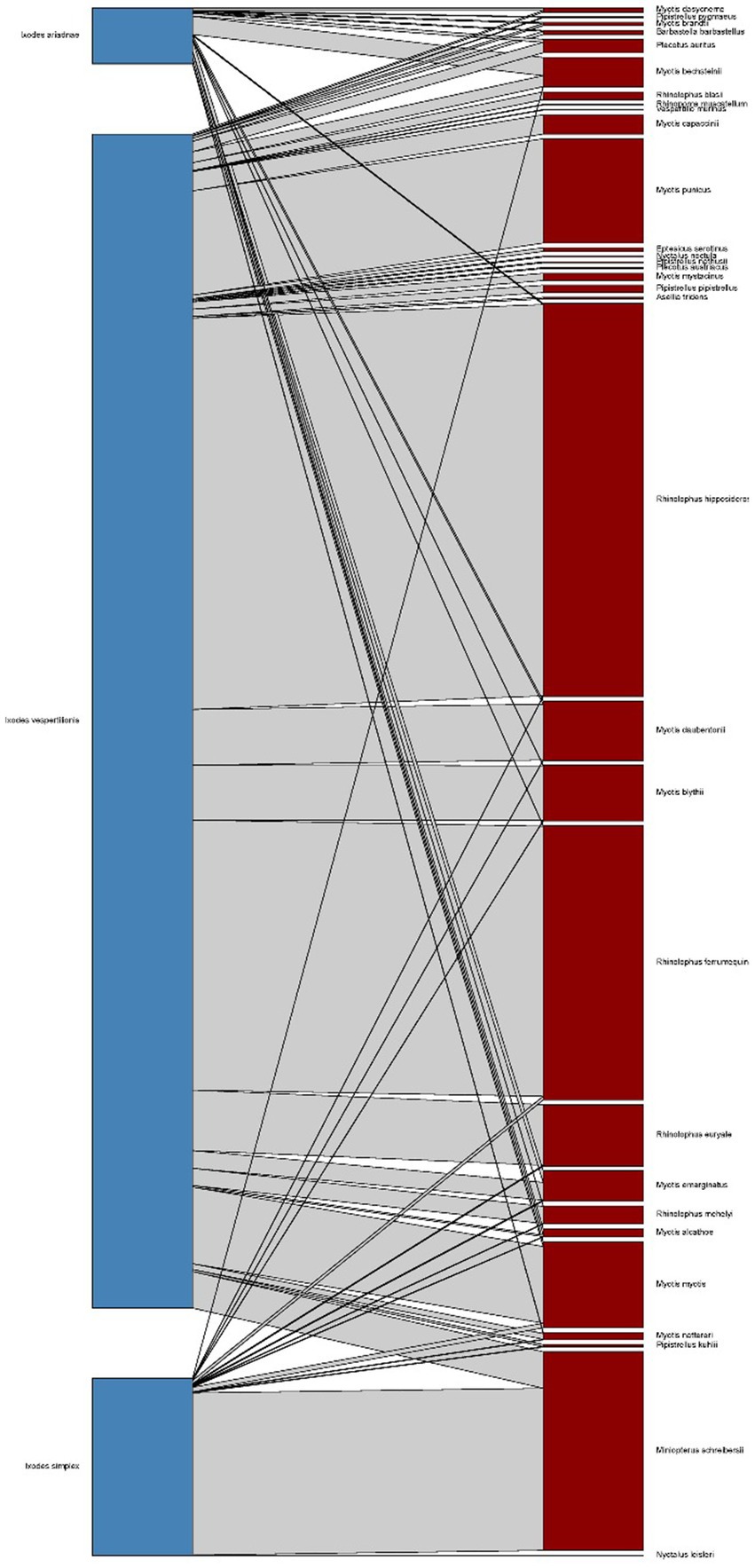
Figure 1. Bipartite representation of the parasite network of bats and their specific hard ticks using a quantitative interaction web based on individual host–parasite relations. Links between nodes represent the sum of individual bat tick occurrences for a given bat and tick species couple (blue bars – bat species, dark red bars – tick species, grey bars – host-parasite links).
Tick records showed wide geographic distribution, with range overlap in Central Europe and the Mediterranean for all three species (Figures 2–6). There was a considerable overlap between the distribution of the primary hosts and the range of I. simplex (Figure 4) and I. vespertilionis (Figure 5). Ixodes ariadnae shows the smallest range (Figure 3), followed by I. simplex (Figure 4) and I. vespertilionis (Figure 5). Two of the three species also occurred south of the Mediterranean Sea, in Africa, though all records of I. ariadnae lay in Central Europe and the Middle East (Anatolia). Most host-collected ixodid ticks came from bats caught close to underground roosts (90.2%), regardless of whether the hosts were cave-dwellers (91.3%, n = 2,367) or crevice dwellers (79.3%, n = 517). Records of hard ticks on crevice dwellers were made mainly in the autumn (71% of all records in August–October). A single tick species, I. ariadnae, showed strong seasonality, with 93.4% of records occurring from August to October. Adult females of I. simplex also showed clumped seasonal occurrence, with over 62% collected in spring (April–June), although only 22% of tick-infested hosts were recorded in spring. We found no marked seasonal differences in the distribution of I. vespertilionis collected from hosts, though slightly more records came from spring. Two bat-specialist tick species were found on humans (I. simplex and I. vespertilionis, each in a single instance). Both species were also collected from dogs, and I. vespertilionis was found on horses and wild boars.
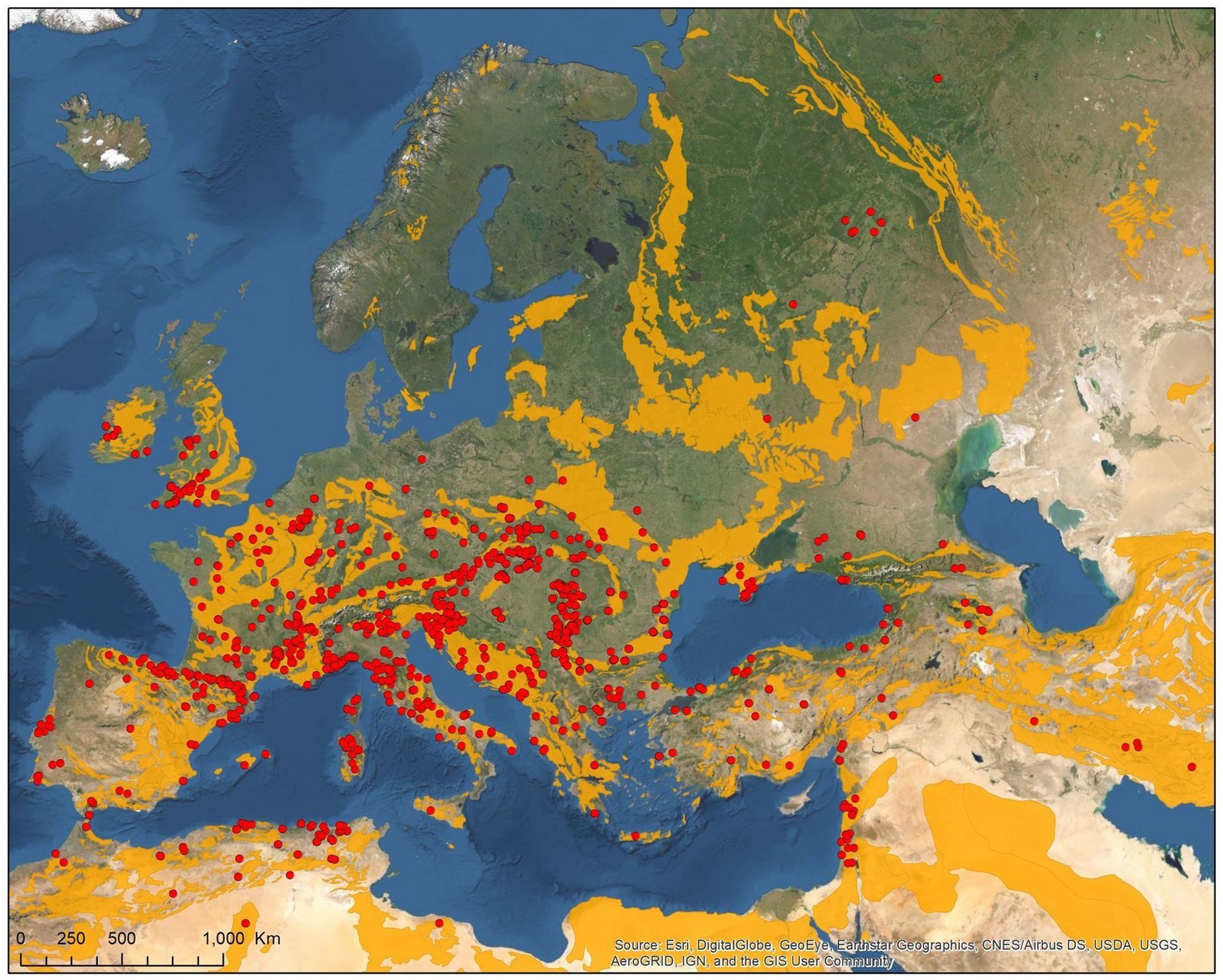
Figure 2. Geographic distribution of host-specialist bat ticks in the Western Palearctic (lime stone bedrock in yellow).
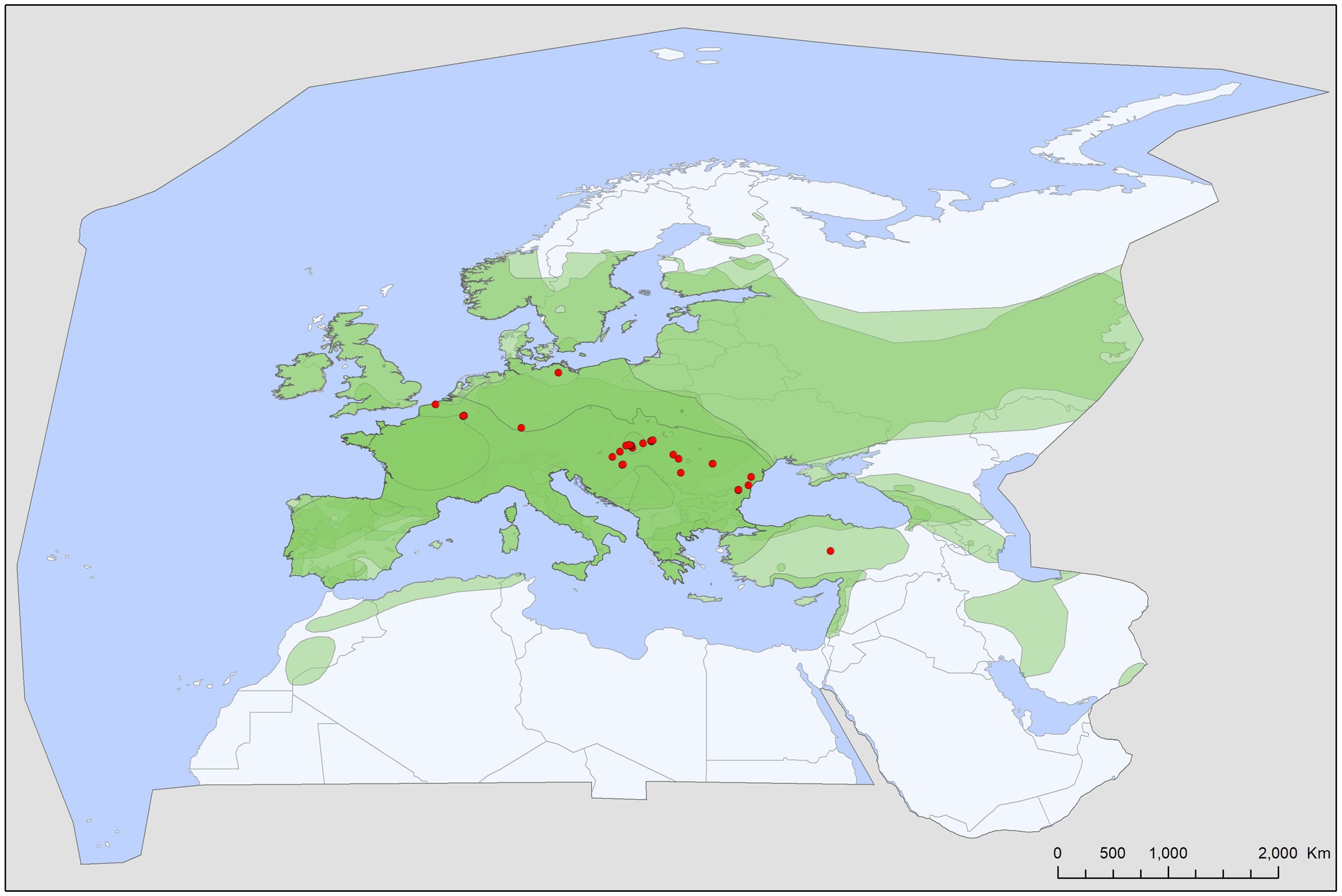
Figure 3. Geographic distribution of Ixodes ariadnae records in the Western Palearctic, overlaid to the geographic ranges for the six bat species studied as primary hosts (Myotis alcathoe, M. bechsteinii, M. daubentonii, M. emarginatus, M. myotis, Plecotus auritus) of this tick. Transparent layers were mapped on top of one another to highlight regions with dense range overlap.
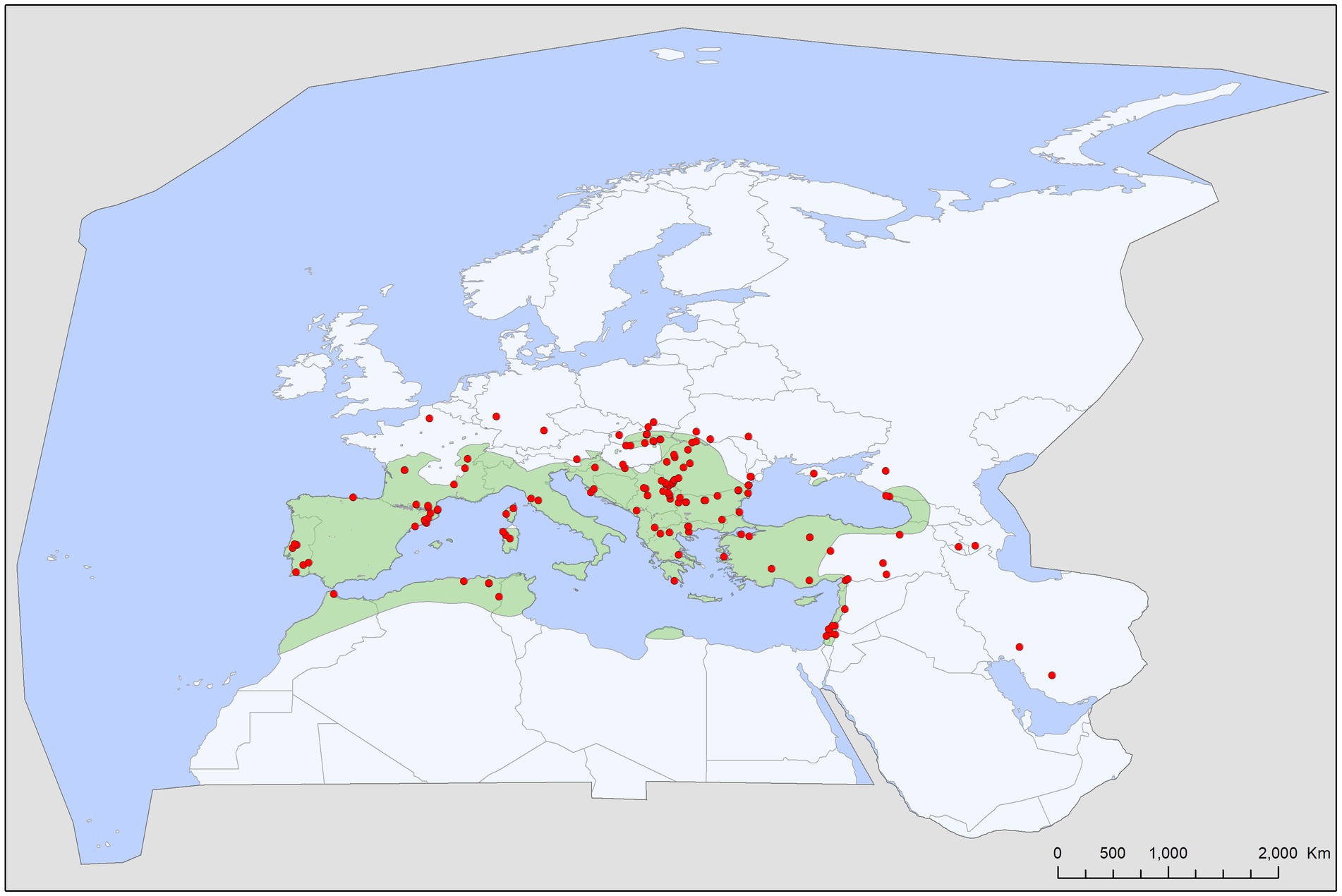
Figure 4. Geographic distribution of Ixodes simplex records in the Western Palearctic, overlaid to the geographic range for its primary host species, Miniopterus schreibersii.
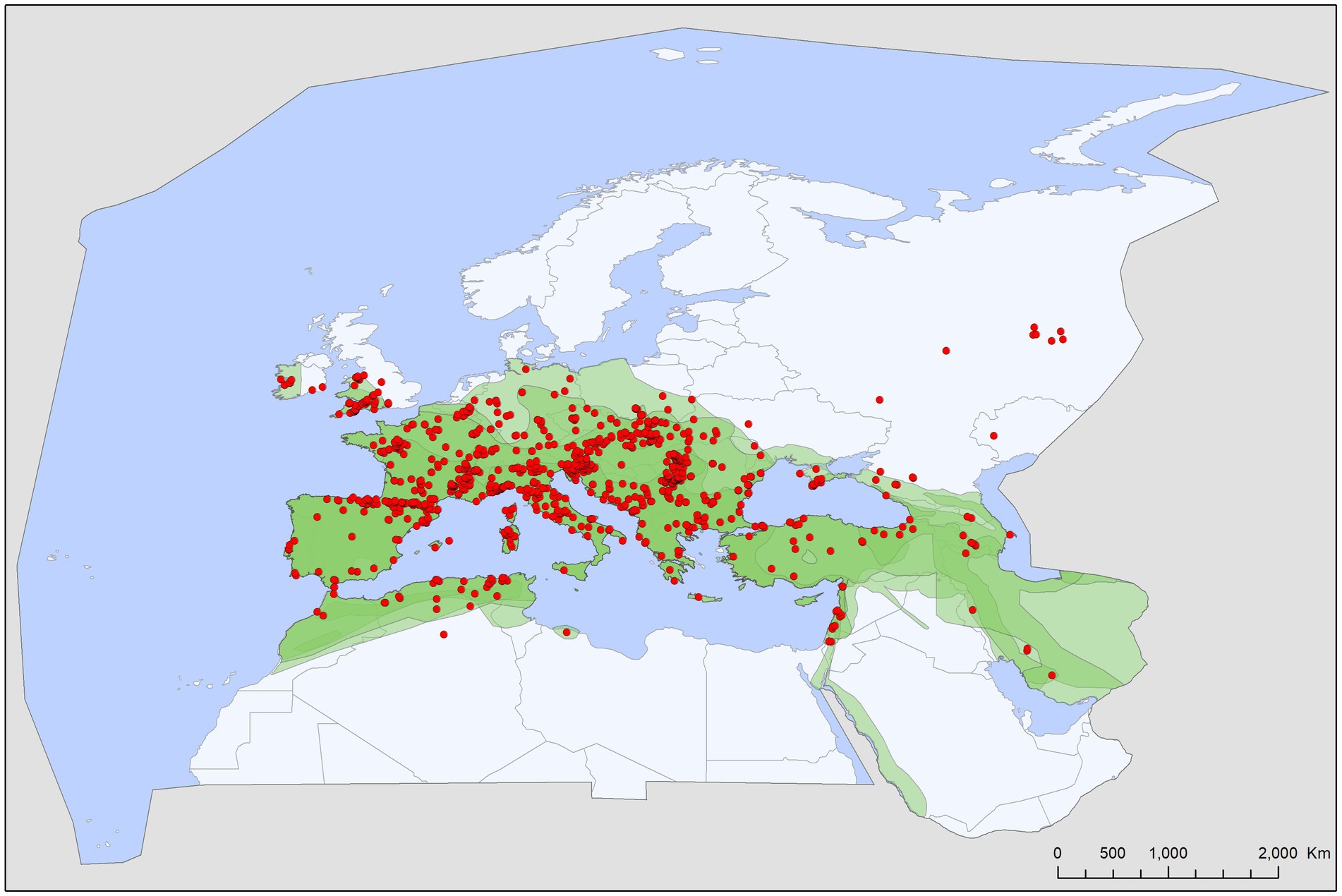
Figure 5. Geographic distribution of Ixodes vespertilionis records in the Western Palearctic, overlaid to the geographic ranges for the five bat species studied as primary hosts (Myotis myotis, M. punicus, Rhinolophus euryale, R. ferrumequinum, R. hipposideros) of this tick. Transparent layers were mapped on top of one another to highlight regions with dense range overlap. Some host species have additional range overlap in Africa and Central and South Asia.

Figure 6. Bipartite representation of the parasite network of generalist/non-specialist ticks and their bat hosts using a quantitative interaction web based on individual host–parasite relations. Links between nodes represent the sum of individual bat tick occurrences for a given bat and tick species couple (blue bars – bat species, dark red bars – tick species, grey bars – host-parasite links).
This survey recorded 110 host–parasite associations involving 27 bat species and other tick species (18 species, 149 individuals; Table 3). Most of these records involved generalist ticks, e.g., Ixodes ricinus (61 cases, 90 individuals, 60.4% of non-specialist tick records) and Rhipicephalus sanguineus s.l. (31 cases, 46 individuals, 30.8% of non-specialist tick records; Figures 6, 7). Even bird-specialists (I. arboricola, I. lividus) or rodent-specialists (I. redikorzevi, I. trinaguliceps) were occasionally recorded. The geographic range of generalist tick records showed a primarily southern distribution, with most being collected in the western part of the Mediterranean region (Figure 7). Wide-range, generalist ticks (I. ricinus and R. sanguineus s.l.) were mostly found on larger, heavier bat species (mean body weight for these hosts was 16.75 g vs. 15.16 g for the rest of generalist tick’s host). These ticks were evenly distributed all over the region (Figure 7, red dots), on both crevice-and cave-roosting species, with Pipistrellus pipistrellus hosting the most tick species (7 tick species), while most ticks were collected from M. myotis (16 cases) and M. schreibersii (11 records).
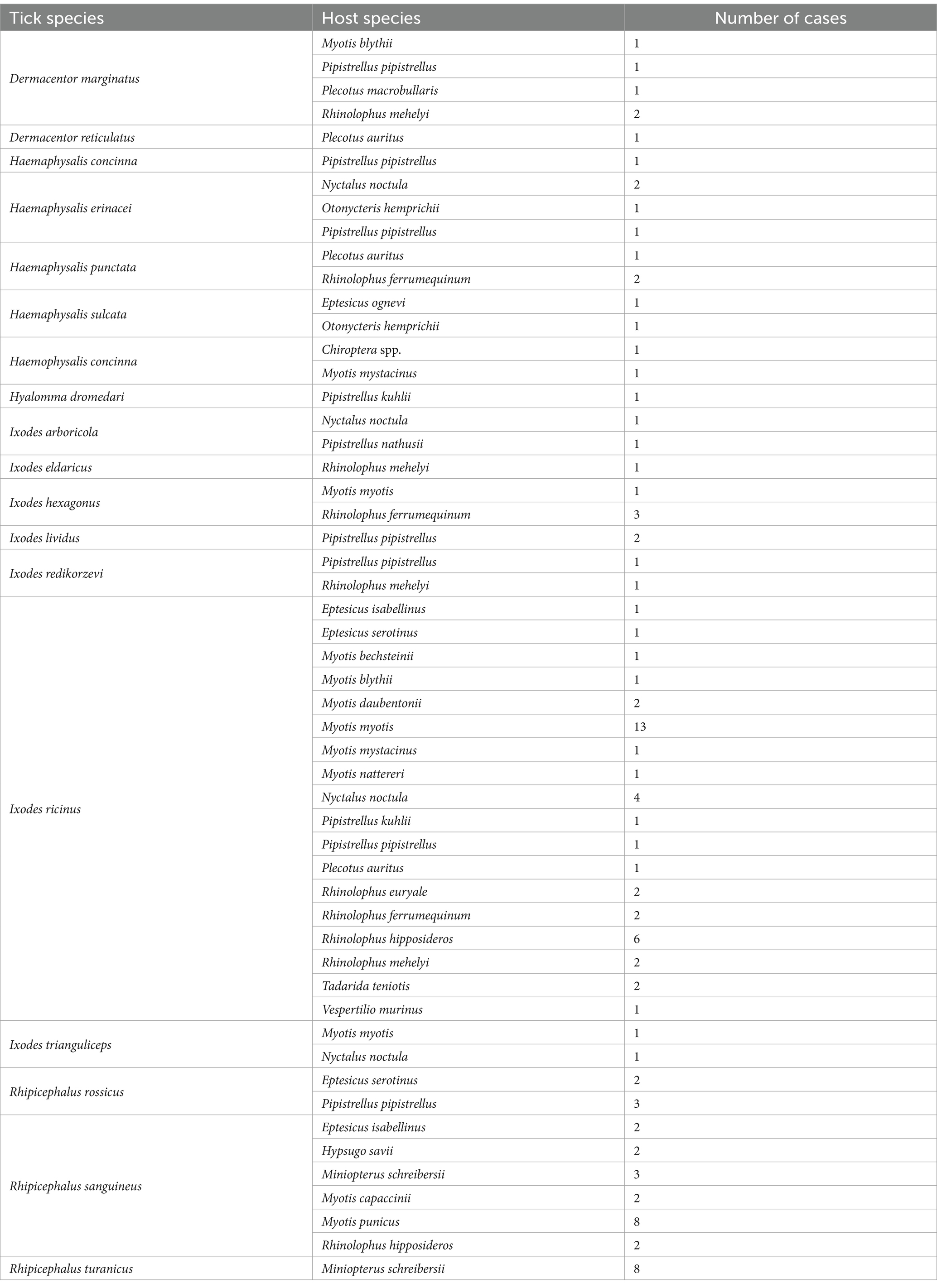
Table 3. List of other (generalist or bird specialist) tick species recorded on bats, with bat host species and number of occurrences.
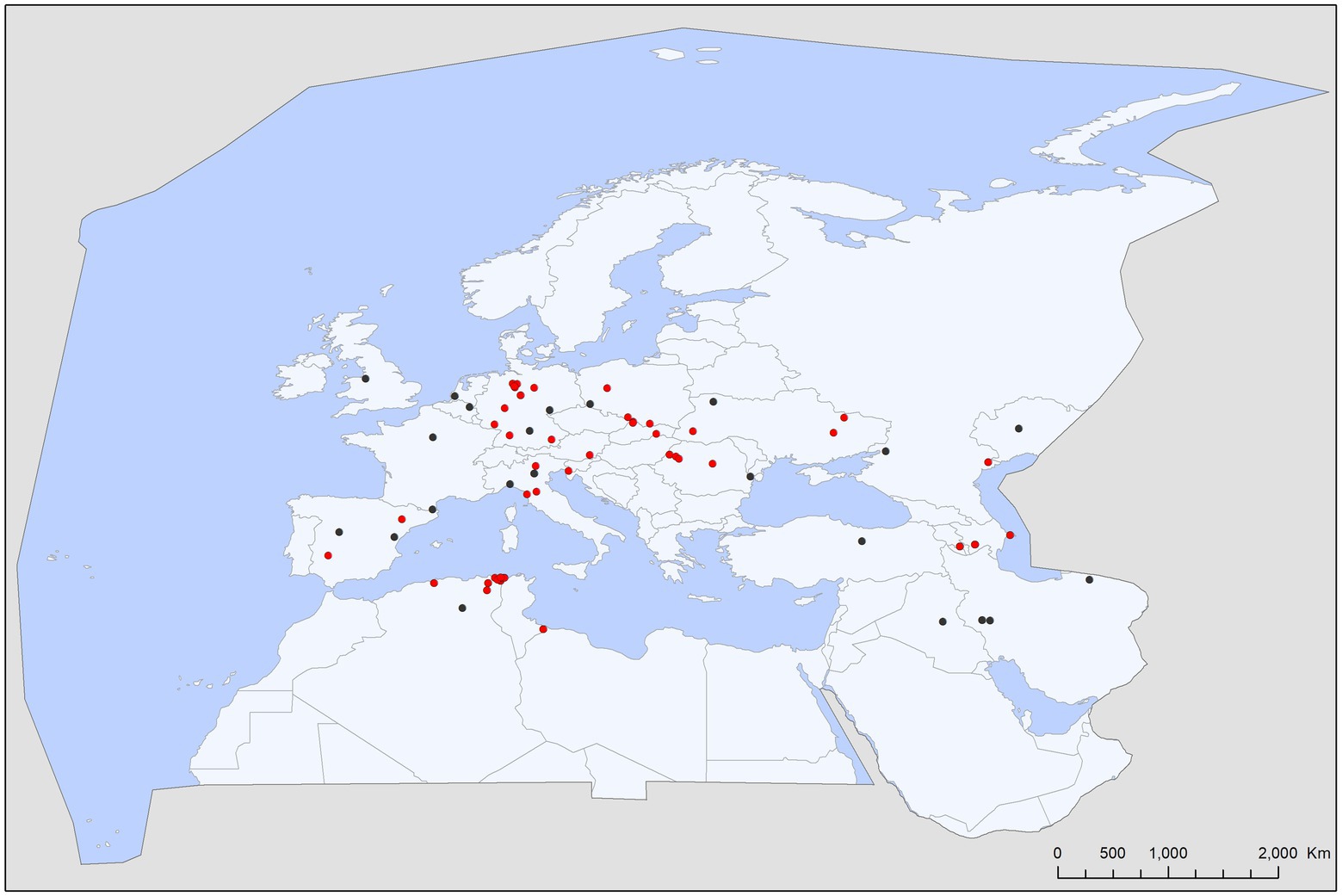
Figure 7. Map with the geographic distribution of other tick species (non-bat ticks) recorded on bats in the Western Palearctic.
Several viral, bacterial, and apicomplexan pathogens were identified in all three bat specialist tick species. DNA of at least eight bacteria, six piroplasmids, a haemosporidian and five viruses were identified in I. simplex, with similar number of bacteria, but less diverse apicomplexan and virus presence in I. vespertilionis. The least studied species (I. ariadnae, four studies), harbored DNA of two bacteria and a single piroplasmid (Table 4).
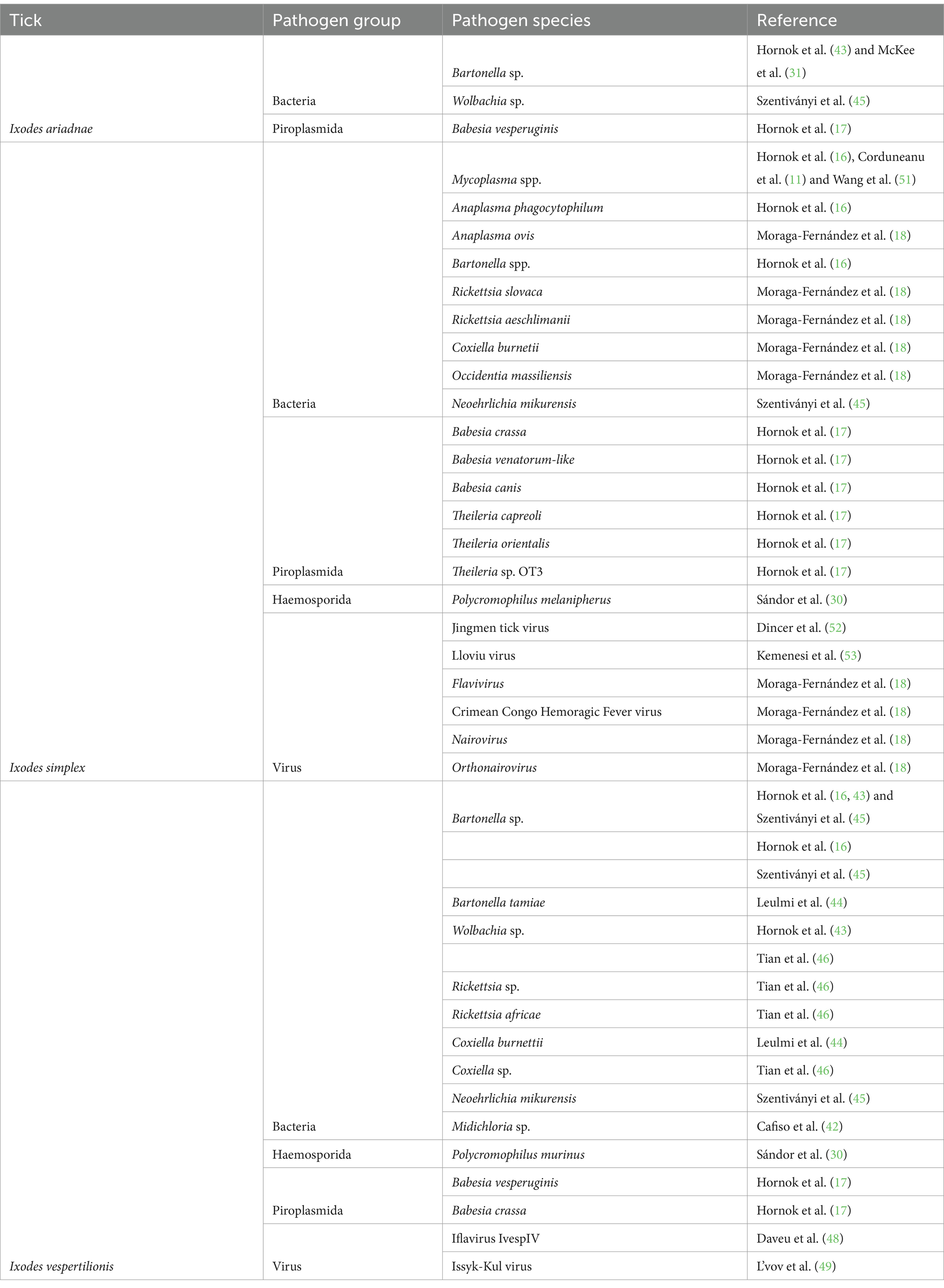
Table 4. DNA of pathogens detected in bat specialist ticks of the Western Palearctic bats.
Discussion
Our survey identified three ixodid ticks specialized on bats in the Western Palearctic, all of which belong to the genus Ixodes. These ticks were recorded from 31 bat species in the region (approximately 40% of all regularly occurring bats; Table 2), with hosts belonging to several bat families, including Hipposideridae, Miniopteridae, Molossidae, Rhinolophidae, Rhinopomatidae, and Vespertilionidae (37). The ticks primarily target cave-dwelling bat species (>90% of tick records with known hosts; see Table 2; Supplementary Table S1) but were also collected from crevice-dwellers during the swarming or hibernation periods when these bats regularly use underground habitats (37). Records of free stages for all three species were exclusively made underground, either inside active bat roosts in caves and mines (98.5%) or in large buildings and cellars with similar environmental conditions, often used by the same bat species. This co-occurrence is likely a result of shared evolutionary history or ecological limiting factors. Ixodes ariadnae, I. simplex and I. vespertilionis are close relatives, all three belong to the morphologically well documented Pholeoixodes subgenus, and their divergence supposedly happened only after a host shift of their common ancestor, likely originating from birds (38, 39). Ecological factors related to the hosts may also contribute to this sympatric occurrence, limiting tick-host interactions to specific spatial environments. All but one cave-dwelling bat species in the Western Palearctic are insectivorous (the fruit-eating Rousettus aegyptiacus is the exception, though no ixodid tick has been recorded from this bat). These bats spend most of their time in active flight away from roosts, spatially limiting the opportunity for ticks to access potential hosts to the interiors of the underground roosts.
Bat specialist ixodid ticks show wide distribution, two species occurring all over Europe, North Africa and the Middle East, however, the recently described I. ariadnae (21) was not yet found in Africa (Figure 3). The ranges of all three species overlap in Central and Eastern Europe and the Middle East, while only I. vespertilionis is found at northern latitudes, and I. ariadnae was not reported from most Mediterranean regions. There is significant overlap in the host spectrum of I. ariadnae and I. vespertilionis, with Myotis myotis serving as an important host for both species (Figure 1). The overlap with the hosts of I. simplex is less pronounced (Table 5) due to the strict host specificity of this species, which primarily parasitizes M. schreibersii (40, 41). Two tick species show distributions that extend well beyond the range of their primary bat host (Figures 4, 5), however, the range of I. ariadnae is far reduced in comparison to its primary hosts’ range, with documented records laying only in the central part of the overlapping range of its primary hosts (Figure 3). We suggest that this may be caused by several factors, like potential misidentification (for example the critical evaluation of samples collected and formerly identified as I. vespertilionis may complete this picture) and by reduced sampling effort in the main occurrence season (the species shows high seasonality in occurrence, limiting the chances of on-host capture, see also (40)).
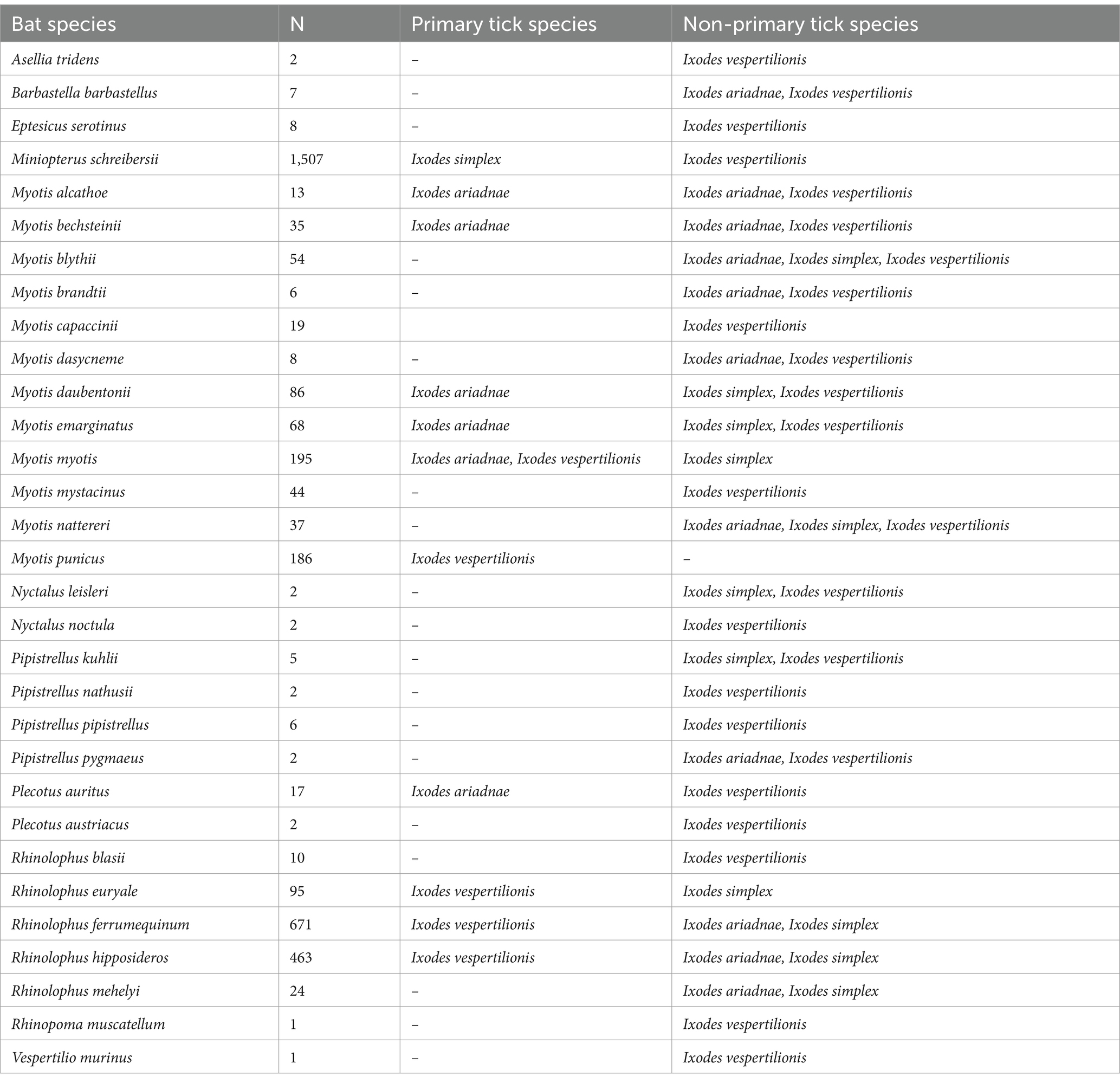
Table 5. List of bat species (Chiroptera) and their role as primary and non-primary bat-specialist hard tick (Ixodidae) hosts in the Western Palearctic (N—number of hosts with ticks).
The geographical distribution of I. ariadnae and I. vespertilionis only partially overlaps with the distribution of their main hosts (Figures 3, 5). This discrepancy is likely due to other limiting factors beyond host range, such as climatic conditions, which may differ at the southern and northern borders of their ranges. However, the presence of bat-specialist ticks is likely not directly limited by climate, as these ticks are primarily found off-host inside underground roosts with optimal climatic conditions. This pattern is clearly visible in Figure 2, where tick distribution is plotted against limestone bedrock, which hosts more than 91% of tick occurrences due to the presence of karst formations (caves).
Ixodes vespertilionis has the largest distribution range, extending from Britain in the west to the Urals in the east and covering North Africa and the Middle East (Figure 5). The easternmost limit likely extends beyond the borders of the Western Palearctic. However, recent assessments of I. vespertilionis specimens from the Eastern Palearctic and Oriental regions revealed several new species (23, 24). This species is primarily associated with horseshoe bats (Rhinolophus spp.) but is also a common parasite of the three large Myotis species (M. blythii, M. myotis, and M. punicus). It is also scarcely recorded on other vesper bats which frequent caves (Table 5 and Figure 1), fact which may help to interpret its occurrences far from the main hosts’ range (Figure 5). The species accounts for the bulk of unengorged tick records collected in caves, due to its habit of questing on cave walls (40). The highly biased sex ratio of free stages noted in this species may be explained by males not feeding and potentially living longer than females, which die after egg laying (hence more chances of encounter on roost walls). There is a slight seasonality in the occurrence of adult free stages, with more records noted during summer months, though this may be due to more frequent cave visits during this period rather than actual seasonality of the species. Several studies detected DNA of pathogenic bacteria (16, 42–46), piroplasms (17, 47) and viruses (48, 49) in I. vespertilionis individuals, both in host collected and free ticks (Table 4). While definitive proof of a vectorial role of this tick species for these pathogens is lacking, its wide distribution, diverse host range, and ubiquitous presence in most bat shelters suggest a significant potential role in pathogen transmission. Moreover, a recent study performing blood-meal analyses managed to detect DNA of non-bat provenience in several adult tick individuals, thus highlighting the chances for pathogen transfer between wide range of host species (31 known species of bat hosts) and other mammals (e.g., dogs, horse and wild boar, (45)), or humans (19).
Ixodes simplex is a nest-dwelling tick, highly gregarious by nature, staying hidden in crevices near its main host colonies (M. schreibersii) (40). It is strictly host-specific, being parasitic almost exclusively on M. schreibersii and rarely found on other bat species (<1.5% of occurrences collected from 13 different bat species, mainly cave-dwellers roosting in sympatry with M. schreibersii). The geographic distribution strongly overlaps with the main distribution of its host, showing a strong mutual relationship with this bat species. Northern outlier records were reported from areas where its host was present in the past (50), while records in the Middle East mostly represent observations on its sister species, the pale bent-winged bat (Miniopterus pallidus). It is common on its hosts, occurring in every roost regularly used by M. schreibersii, showing a constant presence and likely influencing the spatial organization of these bats (41). This tick shows high seasonality in its on-host occurrences, with the highest prevalence and intensity recorded in spring/early summer, sometimes causing detrimental effects on specific host individuals (20). While I. simplex is suspected to vector several bacterial (11, 16, 18, 45, 51), parasitic (17, 47), and viral pathogens (18, 52, 53), there is no unequivocal proof for these roles.
Ixodes ariadnae was recently described from Central European bats (21, 38) and remains a rare bat ectoparasite, with most records geographically limited to a narrow east–west belt between 44° and 51°N latitude, primarily in Europe. Compared to the distribution range of its primary hosts, I. ariadnae shows a highly reduced distribution area. We suggest that this range reflects the spatial extent of recent bat-tick studies rather than the actual distribution, which is expected to increase with future research efforts. This species displays strong seasonality, with 92.2% of host-collected ticks recorded during August–September, coinciding with the autumn swarming of bats (45). While there are fewer than 100 records of I. ariadnae, it has a relatively diverse host range, with 15 known bat hosts (Figure 1; Tables 1, 2). Most hosts (n = 10, 67%) are crevice-dwelling forest bats, which only use underground roosts during swarming or hibernation. Questing adults of I. ariadnae were mainly collected during winter months, though this is likely due to limited access to cave sections occupied by this species (S. Hornok, pers. comm.) rather than true seasonal activity peaks. Only a handful of studies have recorded pathogens in I. ariadnae (Table 4), detecting DNA from bacteria (16, 31, 45) and piroplasms (17).
All, but one Western Palearctic bat species are insectivorous and most species are hunting during flight, relying mainly on insects in flight. In consequence, ticks not using caves or other bat roosts rarely gain access to bat hosts. Thus, presence of generalist ticks on bats is a rare phenomenon, with <1.7% of all tick encounters related to bats represent other species than the three bat-specialist Ixodes. Truly generalist ticks (I. ricinus, R. sanguineus s.l.) made up the bulk of these records and these mainly targeted large-bodied species regularly hunting on the ground (M. blythii, M. myotis, and M. punicus). Other tick species are rarely recorded on bats and are mostly accidentals. Some of these ticks are bird-specialist nest-dwellers, e.g., I. arboricola (regular in tree crevices and bird nest boxes) or I. lividus (a tick species using nest-holes dug by sand martins, Riparia riparia (54)), species which may get access to bats roosting in these bird-nests. Other species are ticks associated to carnivora, which regularly occur in caves (Haemaphysalis erinacei and I. hexagonus (55)).
Bats are frequently parasitized by ticks, and these ticks can host pathogenic bacteria, parasites, or viruses. Certain bat species may act as bridging hosts, carrying not only bat-specialist ticks but also generalist ticks, thus they may have a particular importance from One Health perspective (56). Additionally, a recent study detected high levels of non-bat host DNA in free-living bat ticks, further highlighting the potential for bridging bat-related pathogens to other hosts.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was approved by Underground Heritage Commission (Romania) and the Bulgarian Ministry of Environment and Water (permit no. 718/24.08.2017 and 973/14.04.2023). Bat banding license numbers are 305/2015, 46/2016, 24/2017, 111/2018, 103/2019, 81/2021, and 122/2022. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
AS: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Writing – original draft, Writing – review & editing. CD: Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. ÁP: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Visualization, Writing – review & editing. SH: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. While working for this study, AS and SH were funded by the Office for Supported Research Groups, Hungarian Research Network (HUN-REN), Hungary (Project No. 1500107), and also supported by OTKA K-132794 of the National Research, Development and Innovation Office. ÁP was supported by the PD143382 NKFIH postdoctoral grant.
Acknowledgments
We thank for help provided by T. Szentiványi, M.L. Bendjeddou and M. Sevcik in gathering hard-to-find references.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1517704/full#supplementary-material
Footnotes
References
1. Randolph, SE. Ticks and tick-borne disease systems in space and from space. Adv Parasitol. (2000) 47:217–43. doi: 10.1016/S0065-308X(00)47010-7
2. de la Fuente, J. The fossil record and the origin of ticks (Acari: Parasitiformes: Ixodida). Exp Appl Acarol. (2003) 29:331–44. doi: 10.1023/A:1025824702816
3. Beati, L, and Klompen, H. Phylogeography of ticks (Acari: Ixodida). Annu Rev Entomol. (2019) 64:379–97. doi: 10.1146/annurev-ento-020117-043027
4. Guglielmone, AA, Robbins, RG, Apanaskevich, DA, Petney, TN, Estrada-Peña, A, and Horak, IG. The hard ticks of the world. Dordrecht: Springer (2014).
5. Nava, S, Guglielmone, AA, and Mangold, AJ. An overview of systematics and evolution of ticks. Front Biosci. (2009) 14:2857–77. doi: 10.2741/3418
6. Durden, LA. Taxonomy, host associations, life cycles and vectorial importance of ticks parasitizing small mammals In: S Morand, BR Krasnov, and R Poulin, editors. Micromammals and macroparasites. Tokyo: Springer Japan (2006). 91–102.
7. Sonenshine, DE, and Roe, RM. Biology of ticks, vol. 2. Oxford: Oxford University Press, USA (2014).
8. McCoy, KD, Léger, E, and Dietrich, M. Host specialization in ticks and transmission of tick-borne diseases: a review. Front Cell Infect Microbiol. (2013) 3:57. doi: 10.3389/fcimb.2013.00057
9. Parola, P, and Raoult, D. Ticks and tickborne bacterial diseases in humans: an emerging infectious threat. Clin Infect Dis. (2001) 32:897–928. doi: 10.1086/319347
10. Poel, WHMVD, Lina, PHC, and Kramps, JA. Public health awareness of emerging zoonotic viruses of bats: a European perspective. Vector-Borne Zoonotic Dis. (2006) 6:315–24. doi: 10.1089/vbz.2006.6.315
11. Corduneanu, A, Zając, Z, Kulisz, J, Wozniak, A, Foucault-Simonin, A, Moutailler, S, et al. Detection of bacterial and protozoan pathogens in individual bats and their ectoparasites using high-throughput microfluidic real-time PCR. Microbiol Spectr. (2023) 11:e0153123. doi: 10.1128/spectrum.01531-23
12. Tuttle, MD. Give bats a break: searches for new viruses in bats are unlikely to contribute substantially to human health, but they may threaten the future of bats. Issues Sci Technol. (2017) 33:41–51.
13. Zheng, X, Zhang, X, Huang, X, Yue, X, and Wang, Y. Biodiversity of Ectoparasites and molecular detection of Bartonella in Ectoparasites infesting Rhinolophus Affinis in Yunnan Province, China. Pak Vet J. (2024) 44:699–706.
14. Perumalsamy, N, Sharma, R, Subramanian, M, and Nagarajan, SA. Hard ticks as vectors: the emerging threat of tick-borne diseases in India. Pathogens. (2024) 13:556. doi: 10.3390/pathogens13070556
15. Yu, Z, Wang, H, Wang, T, Sun, W, Yang, X, and Liu, J. Tick-borne pathogens and the vector potential of ticks in China. Parasites Vectors. (2015) 8:24. doi: 10.1186/s13071-014-0628-x
16. Hornok, S, Szoke, K, Meli, ML, Sándor, AD, Görföl, T, Estók, P, et al. Molecular detection of vector-borne bacteria in bat ticks (Acari: Ixodidae, Argasidae) from eight countries of the old and new worlds. Parasites and Vectors. (2019) 12:50. doi: 10.1186/s13071-019-3303-4
17. Hornok, S, Szőke, K, Kováts, D, Estók, P, Görföl, T, Boldogh, SA, et al. DNA of piroplasms of ruminants and dogs in ixodid bat ticks. PLoS One. (2016) 11:e0167735. doi: 10.1371/journal.pone.0167735
18. Moraga-Fernández, A, Sánchez-Sánchez, M, Muñoz-Hernández, C, Pardavila, X, Sereno-Cadierno, J, Queirós, J, et al. Beware with the backpack! New hosts and pathogens identified for Ixodes simplex ticks collected from bats in the Iberian Peninsula. Res Vet Sci. (2024) 176:105316. doi: 10.1016/j.rvsc.2024.105316
19. Piksa, K, Nowak-Chmura, M, and Siuda, K. First case of human infestation by the tick Ixodes vespertilionis (Acari: Ixodidae). Int J Acarol. (2013) 39:1–2. doi: 10.1080/01647954.2012.737831
20. Péter, Á, Barti, L, Corduneanu, A, Hornok, S, Mihalca, AD, and Sándor, AD. First record of Ixodes simplex found on a human host, with a review of cases of human infestation by bat tick species occurring in Europe. Ticks Tick-Borne Dis. (2021) 12:101722. doi: 10.1016/j.ttbdis.2021.101722
21. Hornok, S, Kontschán, J, Kováts, D, Kovács, R, Angyal, D, Görföl, T, et al. Bat ticks revisited: Ixodes ariadnae sp. nov. and allopatric genotypes of I. vespertilionis in caves of Hungary. Parasit Vectors. (2014) 7:202. doi: 10.1186/1756-3305-7-202
22. Hornok, S, Kontschán, J, Estrada-Peña, A, de Mera, IGF, Tomanović, S, and de la Fuente, J. Contributions to the morphology and phylogeny of the newly discovered bat tick species, Ixodes ariadnae in comparison with I. vespertilionis and I simplex. Parasit Vectors. (2015) 8:47. doi: 10.1186/s13071-015-0665-0
23. Hornok, S, Görföl, T, Estók, P, Tu, VT, and Kontschán, J. Description of a new tick species, Ixodes collaris n. sp. (Acari: Ixodidae), from bats (Chiroptera: Hipposideridae, Rhinolophidae) in Vietnam. Parasit Vectors. (2016) 9:322. doi: 10.1186/s13071-016-1608-0
24. Takano, A, Yamauchi, T, Takahashi, M, Shimoda, H, Gotoh, Y, Mizuno, J, et al. Description of three new bat-associated species of hard ticks (Acari, Ixodidae) from Japan. ZooKeys. (2023) 1180:1–26. doi: 10.3897/zookeys.1180.108418
25. Hornok, S, Kontschán, J, Takano, A, Gotoh, Y, Hassanin, A, and Tu, VT. Description of Ixodes lanigeri sp nov., a new hard tick species (Acari, Ixodidae) collected from mouse-eared bats (Vespertilionidae, Myotis) in Vietnam. ZooKeys. (2024) 1215:107–25. doi: 10.3897/zookeys.1215.123624
26. Bendjeddou, ML, Khelfaoui, F, Abiadh, A, Mechouk, N, Mihalca, AD, and Sándor, AD. Bat Ectoparasites (Acari, Diptera, Hemiptera, Siphonaptera) in the grand Maghreb (Algeria, Libya, Mauritania, Morocco and Tunisia): a literature review and new data. Acta Parasit. (2024) 69:106–20. doi: 10.1007/s11686-023-00732-8
27. IUCN. The IUCN red list of threatened species. Version 2024–1. (2024). Available at: https://www.iucnredlist.org (Accessed October 21, 2023).
28. Frick, WF, Kingston, T, and Flanders, J. A review of the major threats and challenges to global bat conservation. Ann N Y Acad Sci. (2019) 1469:5–25. doi: 10.1111/nyas.14045
29. Sándor, AD. Underground life is still safest: comments on ‘danger underground and in the open – predation on blind mole rats (Rodentia, Spalacinae) revisited’. Mammal Rev. (2017) 47:230–5. doi: 10.1111/mam.12089
30. Sándor, AD, Mihalca, AD, Domşa, C, Péter, Á, and Hornok, S. Argasid ticks of Palearctic bats: distribution, host selection, and zoonotic importance. Front Vet Sci. (2021) 8:684737. doi: 10.3389/fvets.2021.684737
31. McKee, CD, Krawczyk, AI, Sándor, AD, Görföl, T, Földvári, M, Földvári, G, et al. Host phylogeny, geographic overlap, and roost sharing shape parasite communities in European bats. Front Ecol Evol. (2019) 7:69. doi: 10.3389/fevo.2019.00069
32. Ficetola, GF, Falaschi, M, Bonardi, A, Padoa-Schioppa, E, and Sindaco, R. Biogeographical structure and endemism pattern in reptiles of the Western Palearctic. Prog Phys Geogr Earth Environ. (2018) 42:220–36. doi: 10.1177/0309133318765084
33. Masseti, M, and Bruner, E. The primates of the western Palaearctic: a biogeographical, historical, and archaeozoological review. J Anthropol Sci. (2009) 87:33–91.
34. Patterson, BD, Dick, CW, and Dittmar, K. Nested distributions of bat flies (Diptera: Streblidae) on Neotropical bats: artifact and specificity in host-parasite studies. Ecography. (2009) 32:481–7. doi: 10.1111/j.1600-0587.2008.05727.x
35. Wenzel, RL. The streblid batflies of Venezuela (Diptera: Streblidae). Brigham Young Univ Sci Bull Biol Ser. (1976) 20:1–177. doi: 10.5962/bhl.part.5666
36. Sándor, AD, Corduneanu, A, Hornok, S, Mihalca, AD, and Péter, Á. Season and host-community composition inside roosts may affect host-specificity of bat flies. Sci Rep. (2024) 14:4127. doi: 10.1038/s41598-024-54143-4
37. Dietz, C, von Helversen, O, and Nill, D. Bats of Britain, Europe and Northwest Africa. London: A&C Black. (2009). 1–400 p.
38. Hornok, S, Estrada-Peña, A, Kontschán, J, Plantard, O, Kunz, B, Mihalca, AD, et al. High degree of mitochondrial gene heterogeneity in the bat tick species Ixodes vespertilionis, I ariadnae and I simplex from Eurasia. Parasit Vectors. (2015) 8:457. doi: 10.1186/s13071-015-1056-2
39. Hornok, S, Sándor, AD, Beck, R, Farkas, R, Beati, L, Kontschán, J, et al. Contributions to the phylogeny of Ixodes (Pholeoixodes) canisuga, I. (Ph.) kaiseri, I. (Ph.) hexagonus and a simple pictorial key for the identification of their females. Parasit Vectors. (2017) 10:545. doi: 10.1186/s13071-017-2424-x
40. Sándor, AD, Corduneanu, A, Péter, Á, Mihalca, AD, Barti, L, Csősz, I, et al. Bats and ticks: host selection and seasonality of bat-specialist ticks in eastern Europe. Parasit Vectors. (2019) 12:605. doi: 10.1186/s13071-019-3861-5
41. Sándor, AD, Péter, Á, Beke, B, Boldogh, SA, Bücs, SL, and Hornok, S. Ectoparasite-posed risk may affect the spatial organization of hibernating clusters of a social bat. Preprint. (2024). doi: 10.21203/rs.3.rs-4141372/v1
42. Cafiso, A, Bazzocchi, C, De Marco, L, Opara, MN, Sassera, D, and Plantard, O. Molecular screening for Midichloria in hard and soft ticks reveals variable prevalence levels and bacterial loads in different tick species. Ticks Tick-Borne Dis. (2016) 7:1186–92. doi: 10.1016/j.ttbdis.2016.07.017
43. Hornok, S, Kovács, R, Meli, ML, Gönczi, E, Hofmann-Lehmann, R, Kontschán, J, et al. First detection of bartonellae in a broad range of bat ectoparasites. Vet Microbiol. (2012) 159:541–3. doi: 10.1016/j.vetmic.2012.04.003
44. Leulmi, H, Aouadi, A, Bitam, I, Bessas, A, Benakhla, A, Raoult, D, et al. Detection of Bartonella tamiae, Coxiella burnetii and rickettsiae in arthropods and tissues from wild and domestic animals in northeastern Algeria. Parasit Vectors. (2016) 9:27. doi: 10.1186/s13071-016-1316-9
45. Szentiványi, T, Takács, N, Sándor, AD, Péter, Á, Boldogh, SA, Kováts, D, et al. Bat-associated ticks as a potential link for vector-borne pathogen transmission between bats and other animals. PLoS Negl Trop Dis. (2024) 18:e0012584. doi: 10.1371/journal.pntd.0012584
46. Tian, J, Hou, X, Ge, M, Xu, H, Yu, B, Liu, J, et al. The diversity and evolutionary relationships of ticks and tick-borne bacteria collected in China. Parasit Vectors. (2022) 15:352. doi: 10.1186/s13071-022-05485-3
47. Sándor, AD, Péter, Á, Corduneanu, A, Barti, L, Csősz, I, Kalmár, Z, et al. Wide distribution and diversity of malaria-related haemosporidian parasites (Polychromophilus spp.) in bats and their ectoparasites in eastern Europe. Microorganisms. (2021) 9:230. doi: 10.3390/microorganisms9020230
48. Daveu, R, Hervet, C, Sigrist, L, Sassera, D, Jex, A, Labadie, K, et al. Sequence diversity and evolution of a group of iflaviruses associated with ticks. Arch Virol. (2021) 166:1843–52. doi: 10.1007/s00705-021-05060-8
49. Lvov, DK, Kostiukov, MA, Daniiarov, OA, Tukhtaev, TM, and Sherikov, BK. Outbreak of arbovirus infection in the Tadzhik SSR due to the Issyk-Kul virus (Issyk-Kul fever). Vopr Virusol. (1984) 29:89–92.
50. Aulagnier, S, and Presetnik, P. Schreibers’ Bent-Winged Bat Miniopterus schreibersii (Kuhl, 1817) In: K Hackländer and FE Zachos, editors. Handbook of the mammals of Europe. Cham: Springer International Publishing (2020). 1–26.
51. Wang, R, Li, Z-M, Peng, Q-M, Gu, X-L, Zhou, C-M, Xiao, X, et al. High prevalence and genetic diversity of hemoplasmas in bats and bat ectoparasites from China. One Health. (2023) 16:100498. doi: 10.1016/j.onehlt.2023.100498
52. Dincer, E, Timurkan, MO, Yalcınkaya, D, Hekimoglu, O, Nayır, MB, Sertkaya, TZ, et al. Molecular detection of Tacheng tick Virus-1 (TcTV-1) and Jingmen tick virus in ticks collected from wildlife and livestock in Turkey: first indication of TcTV-1 beyond China. Vector-Borne Zoonotic Dis. (2023) 23:419–27. doi: 10.1089/vbz.2023.0029
53. Kemenesi, G, Tóth, GE, Mayora-Neto, M, Scott, S, Temperton, N, Wright, E, et al. Isolation of infectious Lloviu virus from Schreiber’s bats in Hungary. Nat Commun. (2022) 13:1706. doi: 10.1038/s41467-022-29298-1
54. Keve, G, Sándor, AD, and Hornok, S. Hard ticks (Acari: Ixodidae) associated with birds in Europe: review of literature data. Front Vet Sci. (2022) 9:928756. doi: 10.3389/fvets.2022.928756
55. Hornok, S, Wang, Y, Otranto, D, Keskin, A, Lia, RP, Kontschán, J, et al. Phylogenetic analysis of Haemaphysalis erinacei Pavesi, 1884 (Acari: Ixodidae) from China, Turkey, Italy and Romania. Parasit Vectors. (2016) 9:643. doi: 10.1186/s13071-016-1927-1
Keywords: Chiroptera, host-specificity, Ixodidae, vector-borne pathogens, zoonotic diseases
Citation: Sándor AD, Domșa C, Péter Á and Hornok S (2025) Ixodid ticks of Western Palearctic bats: ecology, host-parasite relationships, geographic distribution and zoonotic importance. Front. Vet. Sci. 12:1517704. doi: 10.3389/fvets.2025.1517704
Edited by:
Hussam Askar, Al Azhar University, EgyptReviewed by:
Marcos Rogério André, São Paulo State University, BrazilNehaz Muhammad, Hebei Normal University, China
Copyright © 2025 Sándor, Domșa, Péter and Hornok. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Attila D. Sándor, YWRzYW5kb3JAZ21haWwuY29t