 Ádám Kerek
Ádám Kerek Ábel Szabó1
Ábel Szabó1 Ákos Jerzsele
Ákos Jerzsele- 1Department of Pharmacology and Toxicology, University of Veterinary Medicine, Budapest, Hungary
- 2National Laboratory of Infectious Animal Diseases, Antimicrobial Resistance, Veterinary Public Health and Food Chain Safety, University of Veterinary Medicine, Budapest, Hungary
- 3Department of Animal Hygiene, Herd Health and Mobile Clinic, University of Veterinary Medicine, Budapest, Hungary
- 4Poultry-Care Kft., Újszász, Hungary
Introduction: One of the most pressing challenges of our time is the global spread of antimicrobial resistance (AMR). Regular surveillance studies are critical for advancing collaborative efforts between animal and public health sectors, aligning with the One Health concept.
Methods: In this study, we aimed to assess the antibiotic susceptibility of commensal Clostridium perfringens (C. perfringens) strains (n = 146) isolated from largescale poultry farms in Hungary, using minimum inhibitory concentration (MIC) determinations. Additionally, PCR was employed to investigate the presence of major and minor virulence factors.
Results: Our findings revealed a decrease in the efficacy of penicillin-based antibiotics, which are primary choices for treating C. perfringens-related infections, with resistance observed in 48.3% of isolates for penicillin and 20.7% for amoxicillin. Furthermore, virulence gene analysis identified 47 strains (32.2%) carrying the major beta toxin gene, one strain with the epsilon toxin gene (0.7%), 27 strains (18.5%) with the minor beta2 toxin gene, and four strains (2.7%) with the netB toxin gene.
Discussion: These results underscore the necessity of regular surveillance studies and highlight the significant role of commensal strains as reservoirs for sustaining resistance. Future research should include larger sample sizes to provide a more comprehensive understanding of resistance dynamics. Additionally, the genetic basis of resistance in multidrug-resistant strains should be elucidated using next-generation sequencing, enabling targeted interventions to address this growing concern.
1 Introduction
Bacteria can develop resistance to antimicrobial agents which previously were effective in treating infections caused by them. This phenomenon, known as antimicrobial resistance (AMR), represents a global challenge (1). Various microorganisms, such as fungi and saprophytic bacteria, have evolved mechanisms to produce antimicrobial compounds as part of their natural defense strategies (2). The production of antibiotic compounds provides an evolutionary advantage by inhibiting competitors in the struggle for resources (3). Additionally, antibiotics may function as signaling molecules in intercellular communication under natural conditions (4). While many antimicrobial agents are derived from natural compounds, modified to enhance their efficacy, others, such as sulfonamides and fluoroquinolones, are fully synthetic (2). The spread of AMR has emerged as one of the most pressing health concerns in both veterinary and human medicine (1). Consequently, extensive research efforts now focus on exploring alternatives to replace antibiotics partially or entirely. These alternatives include plant-based essential oils (5), plant extracts (6, 7), antimicrobial peptides (8), and propolis, which has demonstrated antimicrobial activity in multiple studies (9–11). Furthermore, medium-chain fatty acids and triglycerides also possess antimicrobial properties (12). The poultry industry, following the swine sector (13) ranks as the second-largest consumer of antibiotics in animal agriculture, underscoring the critical need for responsible and regulated use of antimicrobials within the sector (14). Effective biosecurity measures further play a vital role in reducing antibiotic use (15). To preserve the efficacy of antimicrobial agents, susceptibility testing, coupled with pharmacokinetic and pharmacodynamic studies, is essential before initiating therapeutic treatments (16).
Clostridium perfringens (C. perfringens) is a Gram-positive, anaerobic, spore-forming bacterium most commonly associated with necrotic enteritis in poultry, particularly broiler chickens, where it has significant economic consequences (17). Subclinical infections alone are estimated to cause losses of up to five cents per bird (18), while outbreaks globally result in a $2 billion annual loss to the broiler industry (18). A recently developed toxin-based classification system categorizes C. perfringens strains into seven types (A–G) based on their toxin production, which determines their pathogenicity (19). These types are associated with various diseases: for example, enterotoxin-producing strains (CPE) are classified as type F, while strains producing the critically important virulence factor netB toxin are grouped under type G, encompassing isolates responsible for necrotic enteritis (19). The presence of AMR and virulence factors can be interdependent, with antibiotic selection pressure exerting a positive or negative correlation with these traits, potentially influencing the pathogenicity of the microorganism (20–22). As an opportunistic pathogen, C. perfringens is part of the microbiota in the gastrointestinal tracts of healthy humans and animals but can also contribute to various diseases (23). It is a frequent cause of foodborne illnesses, typically linked to the consumption of fresh vegetables and fruits contaminated with C. perfringens-bearing soil or water, as well as improperly handled meats and cooked foods. The heat-resistant spores of the bacterium can germinate and proliferate rapidly in cooled foods. Additionally, seemingly healthy animals can transmit C. perfringens to humans through direct or indirect contact, potentially causing disease in new hosts (24).
In the treatment of anaerobic bacteria such as C. perfringens, penicillin remains a viable option (25). Amoxicillin, belonging to the class of time-dependent bactericidal antibiotics (26), is effective not only against Gram-negative infections but also in treating C. perfringens (27). Optimal dosing aims to ensure that the drug concentration exceeds the minimum inhibitory concentration (MIC) for 40–60% of the dosing interval, thereby achieving the necessary T > MIC threshold (28, 29). Most Clostridium species are non-pathogenic, with only a few identified as disease-causing. Their versatile metabolism allows them to degrade a wide variety of organic materials through multiple metabolic pathways, including proteins, peptides, amino acids, sugars, purines, pyrimidines, alcohols, and organic acids (30). This versatility provides them with an essential ecological role in biomass renewal (31). However, the historical overuse of antibiotics has contributed to the spread of AMR in Clostridium strains and recent studies have shown a decline in the sensitivity of C. perfringens isolates from food samples to tetracycline, erythromycin, and lincomycin (32).
Monitoring trends such as these is crucial for tracking AMR, which is crucial to combat and it thus underscores the importance of our study. We aimed to assess the antimicrobial susceptibility profiles of commensal C. perfringens strains isolated from large-scale poultry farms in Hungary. Additionally, we explored potential correlations between the presence of virulence factors, identified through PCR assays, and the resistance profiles of these strains.
2 Results
2.1 Regional distribution and origin of samples received
We examined a total of 146 C. perfringens isolates obtained from broiler chickens. The regional distribution of the isolates (Figure 1) was as follows: 8 isolates (5.5%) originated from the Dél-Alföld region, 23 isolates (15.8%) from the Dél-Dunántúl region, 24 isolates (16.4%) from the Észak-Alföld region, 17 isolates (11.6%) from the Észak-Magyarország region, 32 isolates (21.9%) from the Közép-Dunántúl region, 14 isolates (9.6%) from the Közép-Magyarország region, and 28 isolates (19.2%) from the Nyugat-Dunántúl region. These seven regions correspond to all NUTS-2 statistical regions of Hungary, providing nationwide geographic coverage in the isolate sampling.
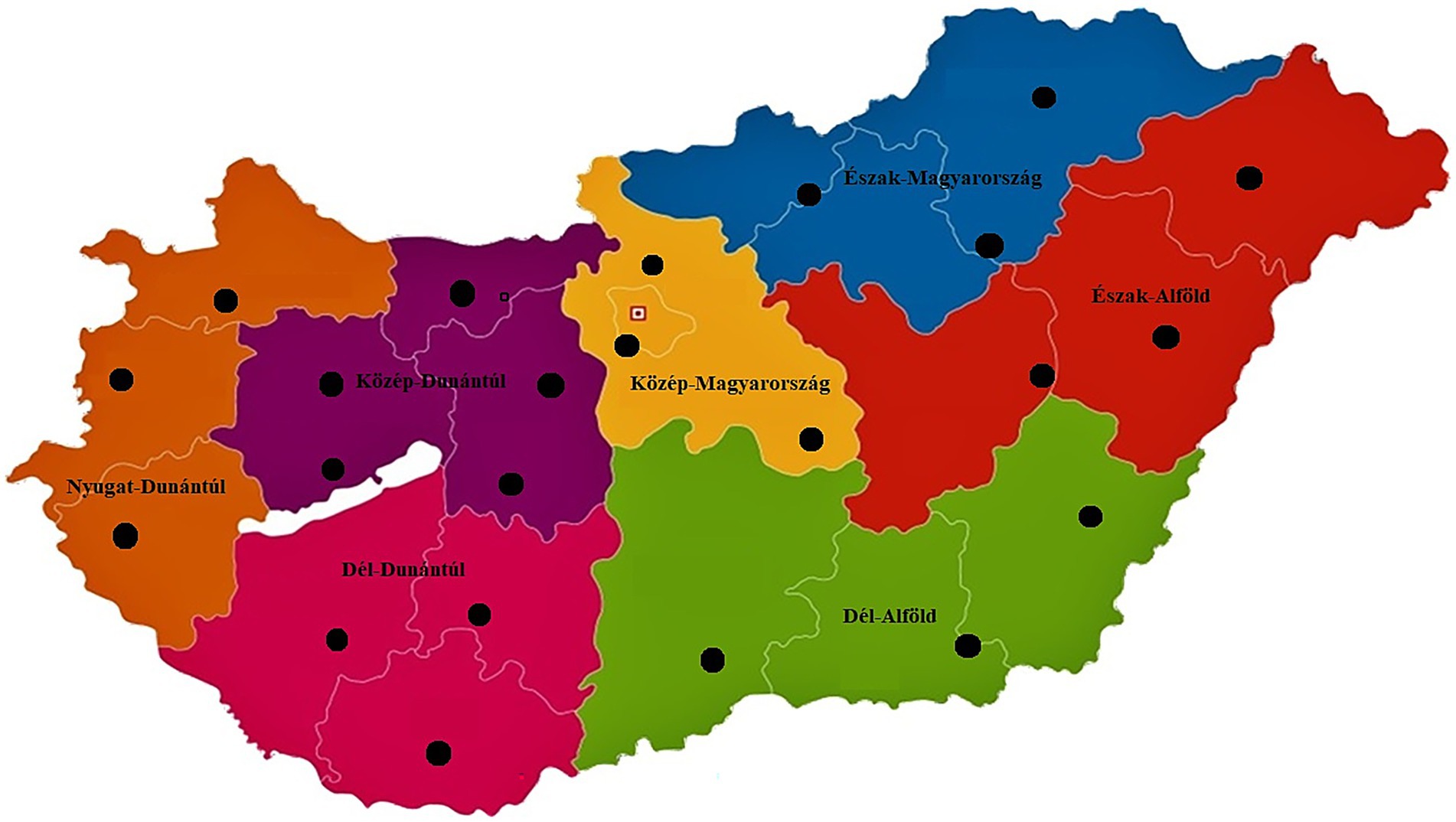
Figure 1. Regional origin of samples collected from chickens across the seven regions of Hungary. Black dots indicate the geographic locations (n = 23) of sample sources, demonstrating nationwide coverage.
Among the isolates, 39.7% were derived from broiler flocks, 35.6% from breeding flocks, and 24.7% from egg-laying flocks. Additionally, 39.7% of the isolates originated from juvenile flocks, while 60.3% were from adult flocks. Regarding flock size, 42.7% of the isolates were obtained from small flocks (5,001–50,000), 35.6% from medium-sized flocks (50,001–100,000), and 21.7% from large flocks (>100,001).
2.2 Correlation and clustering of resistance profiles
After determining the MIC values, we assessed the resistance rates for antibiotics with established clinical breakpoints. Subsequently, correlation analysis was performed to evaluate the relationships between the different active substances (Figure 2). We observed a strong positive correlation between lincomycin and clindamycin (0.96), tilozin and clindamycin (0.95), tilozin and lincomycin (0.92), as well as enrofloxacin and vancomycin (0.83).
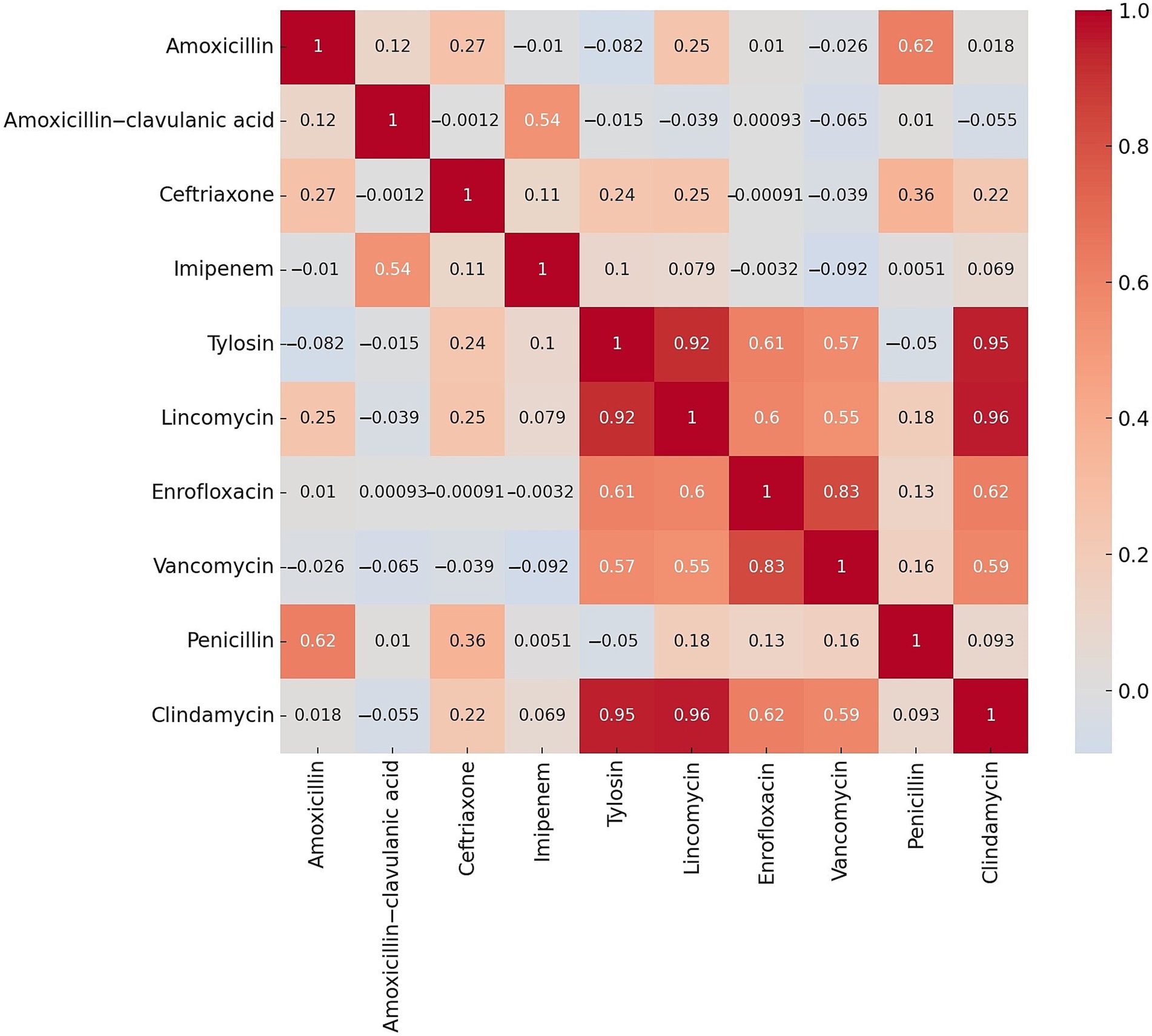
Figure 2. Correlation analysis of resistance rates between various active substances following the determination of resistance levels in Clostridium perfringens strains (n = 146) isolated from domestic chickens.
Hierarchical cluster analysis was conducted to uncover patterns and relationships among the examined strains based on their antimicrobial resistance profiles. Based on the clustering results, the 146 commensal C. perfringens strains were grouped into three main clusters, which reflect distinct resistance profiles. These hierarchical relationships are visualized in a dendrogram (Figure 3), which highlights structural similarities and potential outliers. The three clusters are further illustrated in the principal component analysis (PCA) plot (Figure 4), where they are color-coded for clarity.
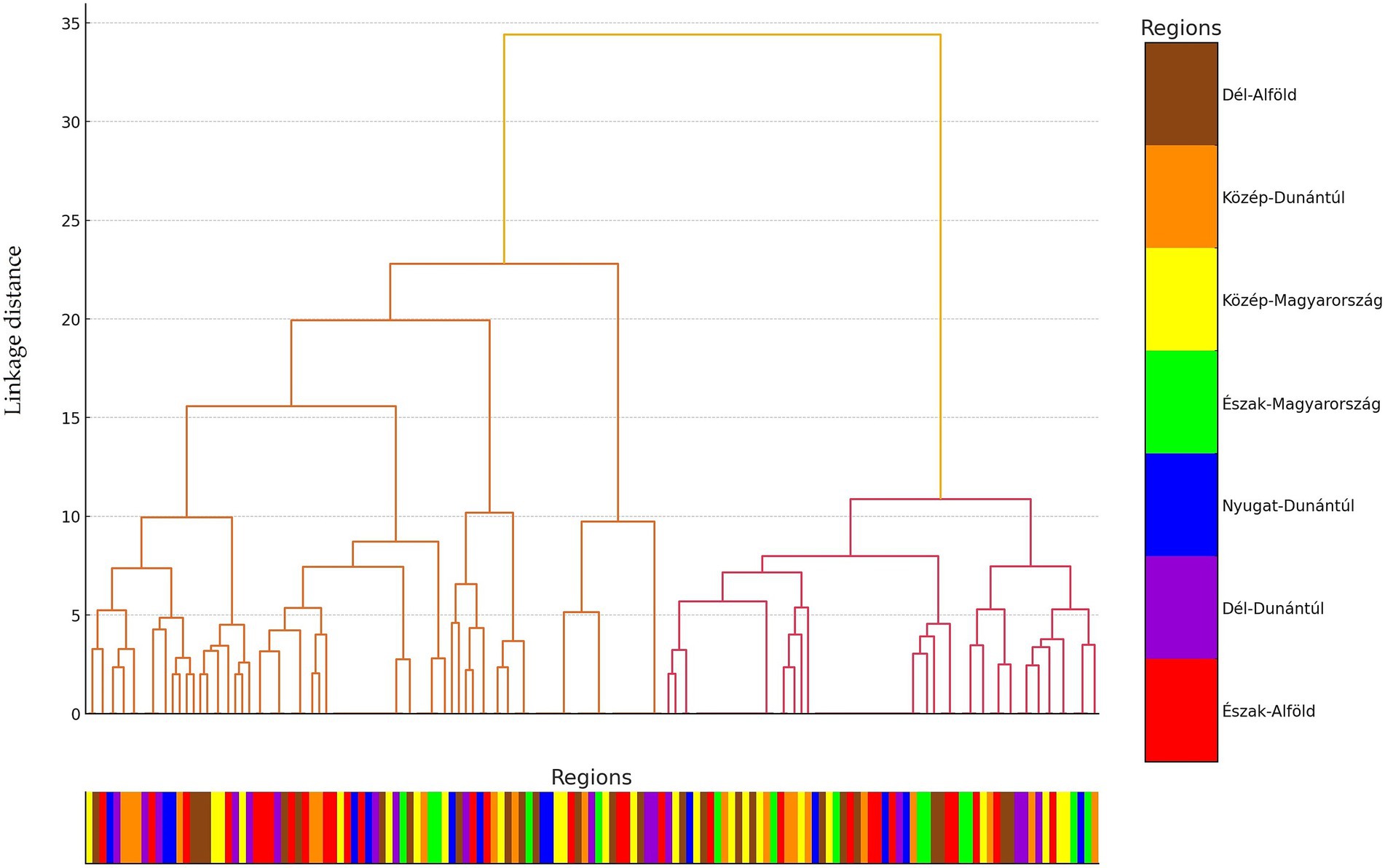
Figure 3. Hierarchical cluster analysis of commensal Clostridium perfringens strains (n = 146) isolated from chickens. The branches of the dendrogram represent clusters of isolates with similar resistance profiles, while the branch lengths indicate the degree of similarity or difference among the isolates.
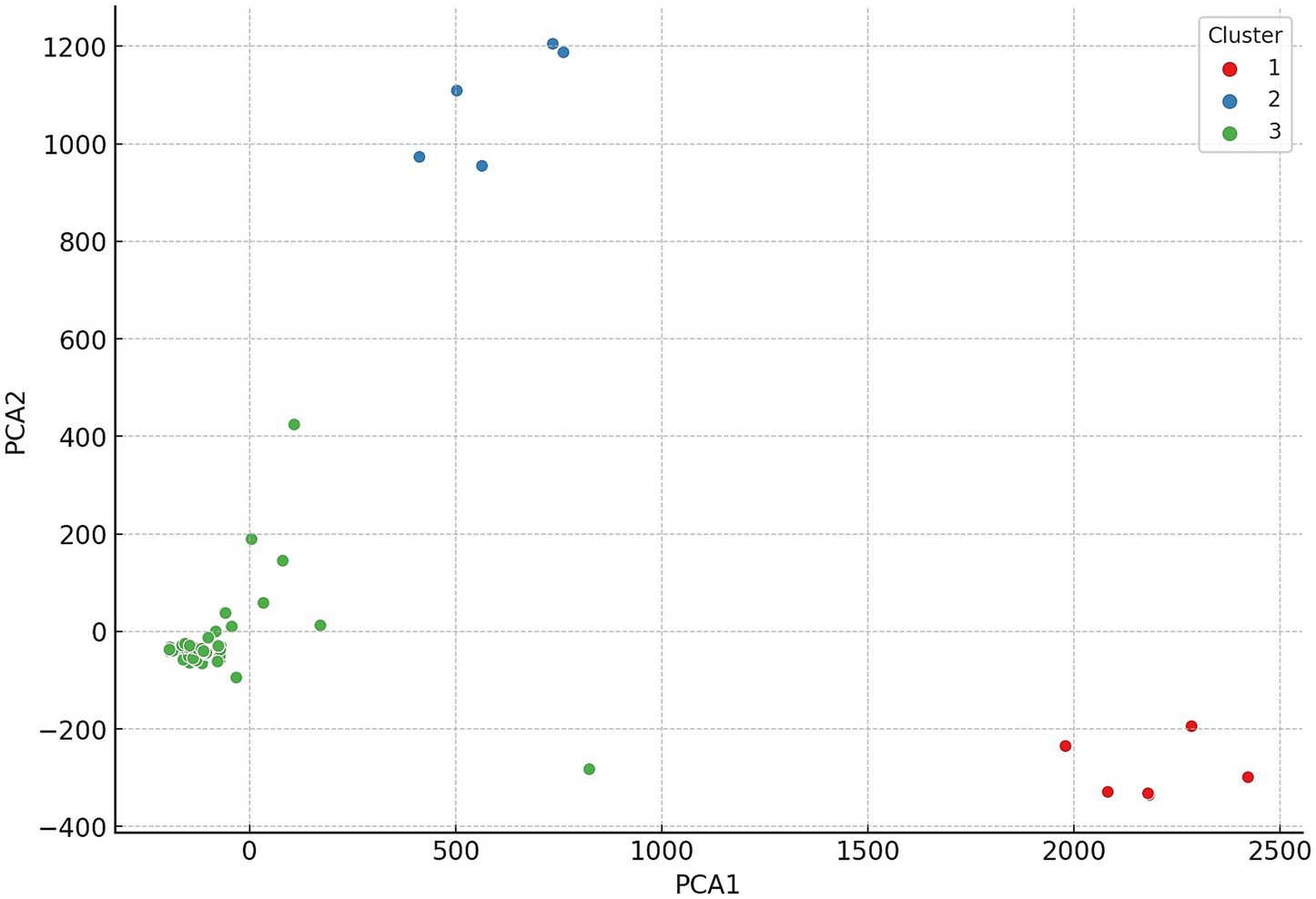
Figure 4. Principal component analysis (PCA) of commensal Clostridium perfringens strains (n = 146) isolated from chickens. The first principal component accounted for the largest variance, followed by the second principal component. Only a few strains were observed in the third cluster.
Subsequently, PCA was performed to reduce dimensionality and uncover major patterns in the antimicrobial resistance profiles (Figure 4). The first principal component accounted for the largest proportion of variance, while the second component, orthogonal to the first, explained the next highest amount. This analysis revealed clear clustering tendencies among the strains. The geographic origin of isolates strongly correlated with cluster membership: strains from the same region were predominantly grouped within the same cluster, a pattern supported by the horizontal color-coded axis. The cluster analysis revealed significant associations between certain antimicrobial resistance profiles and cluster membership. For instance, isolates resistant to lincosamides (lincomycin and clindamycin) were predominantly assigned to the second cluster. Additionally, the results suggest geographic patterns likely influenced by region-specific antibiotic usage practices.
2.3 Regional and usage-related patterns of resistance
Statistical analyses were conducted to evaluate resistance levels for each active substance based on utilization type (Table 1). Significant differences in resistance were observed between egg-laying and meat-producing flocks, as well as between breeding and meat-producing flocks, particularly for lincomycin, enrofloxacin, penicillin, and clindamycin. In all cases, broiler chickens exhibited significantly higher resistance levels compared to other utilization types.
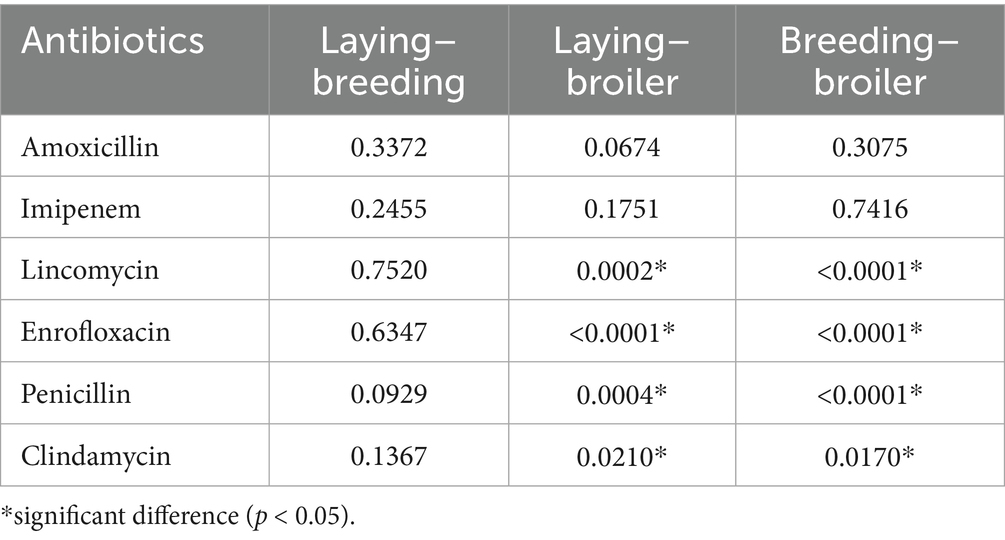
Table 1. Importance of each type of utilization in the resistance profile, by antibiotics.
The influence of age groups on resistance levels was also assessed for each active substance (Table 2). Significant differences were observed for lincomycin, enrofloxacin, penicillin, and clindamycin, with notably higher resistance levels identified in juvenile flocks compared to adult populations.
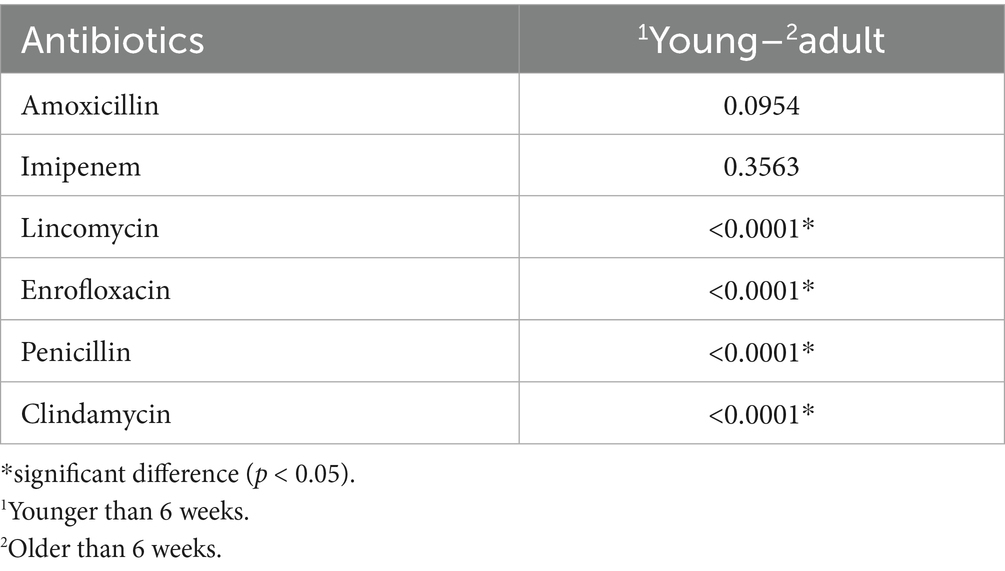
Table 2. The importance of age group on the degree of resistance to each antibiotic.
Additionally, farm size was evaluated as a potential factor influencing resistance levels across the tested active substances (Table 3). The most notable differences were observed between small and large-scale farms for imipenem, lincomycin, and enrofloxacin. In all cases, significantly higher resistance levels were identified on large-scale farms compared to smaller ones.
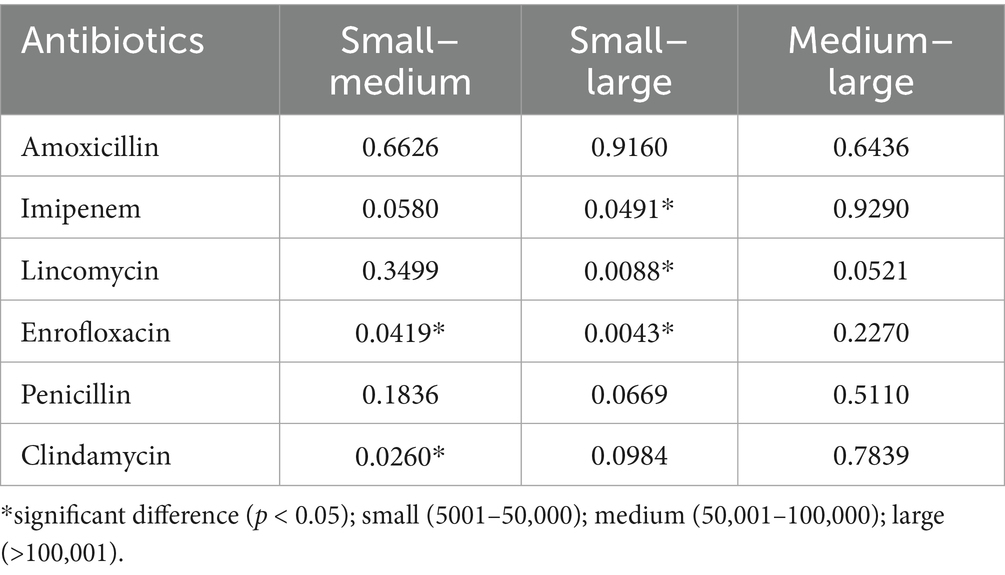
Table 3. The effect of colony size on the level of resistance to certain antibiotics.
2.4 MIC distributions and susceptibility classification
We generated frequency tables for the determined MIC values for each active substance which has clinical breakpoints, including the percentage distribution of these values (Table 4). Subsequently, we calculated the MIC50 and MIC90 values and included the epidemiological cut-off values (ECOFF) defined by European Committee on Antimicrobial Susceptibility Testing (EUCAST) for clindamycin. However, ECOFF values were not the other antibiotics we tested. Both the calculated MIC50 and MIC90 values for imipenem were below the clinical breakpoint. Additionally, the MIC50 value was below the clinical breakpoint for penicillin, clindamycin, and amoxicillin. The ECOFF value for clindamycin was also below the clinical breakpoint. A frequency table for active substances without clinical breakpoints is provided in Supplementary Table S1.
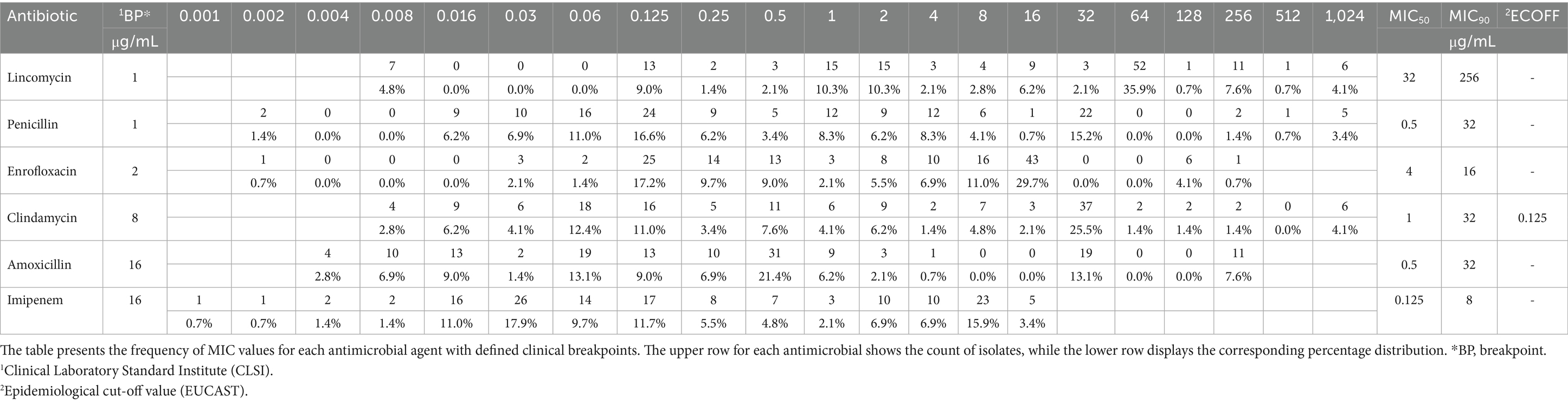
Table 4. Frequency table of minimum inhibitory concentration (MIC) values for antimicrobials with defined breakpoints in Clostridium perfringens isolates from domestic chickens (n = 145).
For antimicrobials with defined clinical breakpoints, the proportion of resistant and susceptible isolates was expressed in percentages (Figure 5). Resistance varied widely according to antibiotic, with isolates showing very low resistance to imipenem. Reduced susceptibility was observed for penicillin, with only 51.7% of isolates classified as sensitive. In contrast, 79.3% of isolates were sensitive to amoxicillin. Lincomycin was found to be the least effective, with just 17.2% of isolates displaying susceptibility.
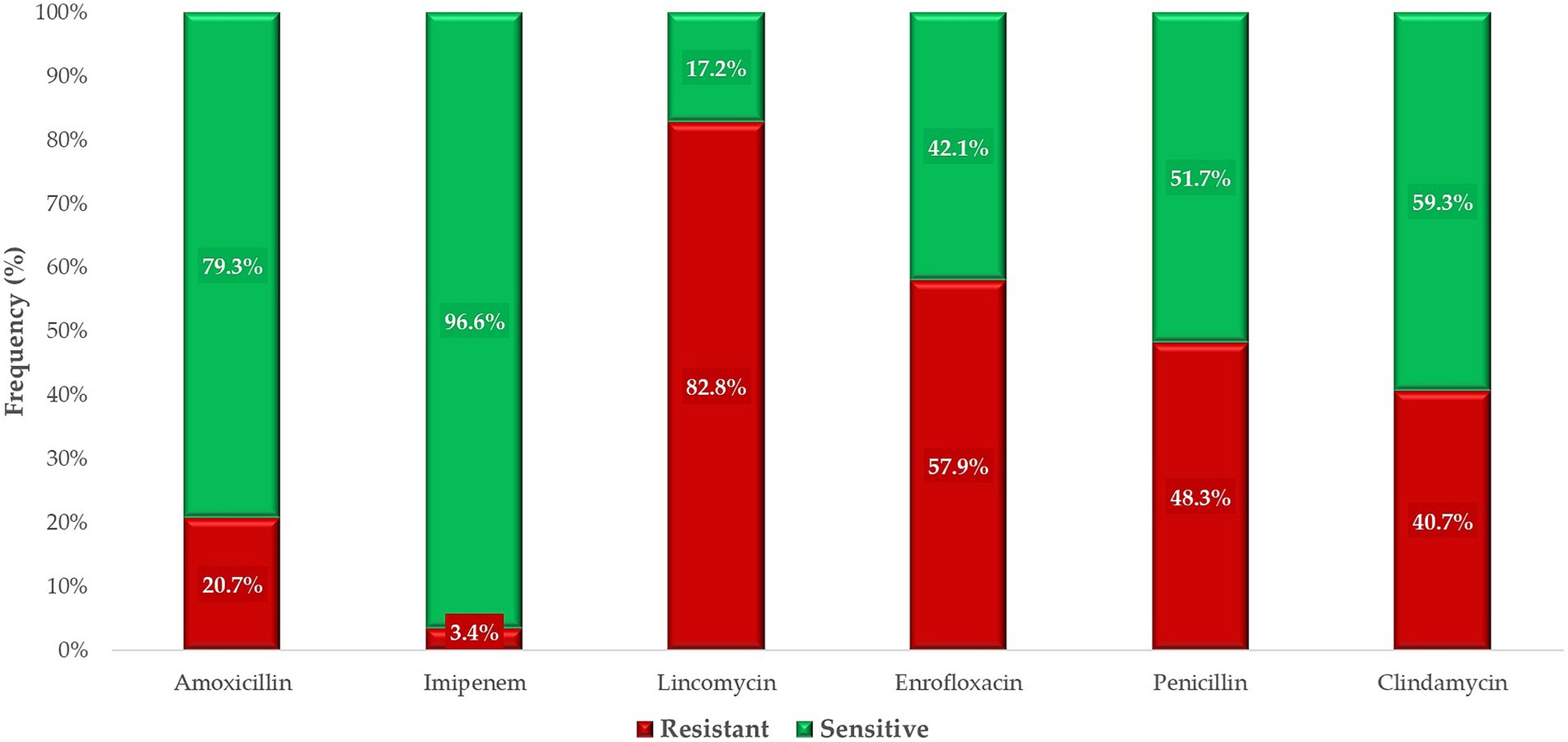
Figure 5. The susceptibility profile of commensal Clostridium perfringens strains (n = 146) isolated from domestic chickens to antibiotics with defined clinical breakpoints.
2.5 Detection and co-occurrence patterns of toxin genes
We analyzed the presence of major and minor toxin genes in each isolation using PCR. All examined isolates tested positive for the major alpha toxin gene, serving as a quality control measure, as this gene is inherently carried by all C. perfringens strains. The major beta toxin gene was detected in 47 isolates (32.2%), while the epsilon toxin gene was identified in a single isolate (0.7%). The minor beta2 toxin gene was present in 27 isolates (18.5%), and the netB toxin gene was found in four isolates (2.7%). None of the isolates carried the genes responsible for entero or iota toxin production.
When analyzing the co-occurrence of multiple toxin genes (excluding the universally present major alpha toxin gene), the most frequent combination—observed in 18 cases (12.3%)—was the major beta and minor beta2 toxin genes. This was followed by the concurrent presence of the major beta, minor beta2, and netB toxin genes in seven cases (4.8%), and the major beta, minor beta2, and epsilon genes in three cases (2.1%). Additionally, the combination of major beta and epsilon toxin genes and that of major beta and netB toxin genes were each observed in a single instance (0.7%).
2.6 Association of toxin genes with resistance
Statistical analysis revealed that the presence or absence of specific toxin genes influenced the degree of resistance to certain antibiotics. For enrofloxacin, a significant difference (p = 0.0049) was noted when the netB gene was present, with significantly lower resistance levels observed in isolates carrying the gene. Furthermore, when examining the impact of carrying multiple genes simultaneously, it was found that resistance to lincomycin was significantly higher (p = 0.0379) in isolates lacking multiple toxin genes compared to those carrying multiple genes.
3 Discussion
We examined a total of 146\u00B0C. perfringens isolates derived from broiler chickens. Following the assessment of resistance levels, a strong positive correlation was identified between several antibiotics, including lincomycin and clindamycin (0.96), tilozin and clindamycin (0.95), tilozin and lincomycin (0.92), and enrofloxacin and vancomycin (0.83). Cluster analysis revealed that isolates resistant to both lincomycin and clindamycin were predominantly grouped within the second cluster. These findings also suggest underlying geographic patterns influencing antibiotic resistance, potentially reflecting region-specific variations in antibiotic usage.
When examining utilization patterns, significant differences in resistance levels were observed across production types, with meat-producing animals exhibiting consistently higher resistance levels compared to breeding populations for most tested substances. Furthermore, age group analysis demonstrated significantly elevated resistance to multiple antibiotics in younger, finishing animals. This trend is likely attributable to the increased selection pressure exerted by the frequent use of antibiotics during rearing. By contrast, colony size had minimal impact on resistance variability.
In C. perfringens infections, first-line therapeutic agents generally retained their efficacy. However, resistance patterns incorporating toxin-producing genes in the isolates revealed significant resistance differences for enrofloxacin and lincomycin, particularly when multiple toxin genes were simultaneously present.
In this study, we found that resistance to amoxicillin among the isolates was only 20.7%. This antibiotic is one of the primary choices for treating infections. However, Wei et al. (2020) reported no resistance to amoxicillin among their isolates (33). A Belgian study found that all strains isolated from the gastrointestinal tracts of broiler chickens were sensitive to amoxicillin, with MIC values below 1 μg/mL (34). In Egypt, 7% of necrotic enteritis-causing isolates were resistant to amoxicillin (35). Similarly, in the United States in 2016, no resistance to amoxicillin-clavulanate was detected in in C. perfringens isolates recovered from broiler chicken fecal samples (36). Comparative studies on amoxicillin-clavulanate resistance in poultry are limited. This is partly due to the lack of a maximum residue limit (MRL) for poultry, and the fact that C. perfringens strains typically do not produce beta-lactamase, making the addition of clavulanate unnecessary. The investigation of amoxicillin-clavulanate is therefore more relevant for research purposes than practical veterinary applications.
The reduced sensitivity to amoxicillin observed in our study, compared to other international studies, may be attributable to differing antibiotic usage practices. Oral preparations of penicillins have been extensively used in poultry production for decades, exerting significant selective pressure on the gut microbiota and contributing to environmental contamination. The 48.3% resistance to narrow-spectrum penicillin observed in this study is particularly concerning. Silva et al. (2009) reported no penicillin-resistant strains in broiler chickens, with MIC values not exceeding 0.25 μg/mL (37). Regarding imipenem, 3.4% of the isolates in our study exhibited resistance, whereas Akhi et al. reported 38% resistance in human fecal isolates (38). In Vietnam, all chicken meat-derived isolates were sensitive to imipenem (39). Although imipenem is strictly reserved for human medicine as a last-resort, life-saving antibiotic, the emergence of resistance in veterinary settings is particularly concerning and undesirable. One limitation of imipenem analysis is its instability in aqueous solutions, which may impact results (40).
We observed a resistance rate of 57.9% to enrofloxacin, while Wei et al. reported a slightly lower rate of 43.8% (33). In contrast, Osman and Elhariri documented an even higher resistance rate of 82% in isolates from broiler chickens (35). These elevated resistance rates are not surprising, given the widespread use of enrofloxacin in the poultry industry and its demonstrated ineffectiveness against anaerobic bacteria, which typically show minimal susceptibility to this drug (41). Moreover, enrofloxacin is classified as a critically important antimicrobial for human health, listed among WHO’s highest-priority fluoroquinolones (42). Its use must therefore be significantly curtailed in veterinary applications to preserve its effectiveness for treating multidrug-resistant infections in humans.
The highest level of resistance was observed against lincomycin, with 82.8% of the isolates identified as resistant. In a study conducted in Egypt, all broiler chicken isolates were found to be resistant to lincomycin (35). Similarly, two Belgian studies reported resistance rates of 61.5 and 63.3%, respectively, among isolates. However, in Brazil, 89.1% of isolates from healthy broiler chickens were found to be sensitive to lincomycin (43). For clindamycin, the resistance rate was 40.7%. In Korea, between 2010 and 2011, 52.5% of isolates from domestic chickens were resistant to clindamycin, whereas in samples collected between 2012 and 2016, the resistance rate decreased to 29.7% (33). In Canada’s Ontario province, only 2% of isolates from domestic chickens collected in the spring of 2005 were resistant to clindamycin (44). The high levels of resistance observed for both clindamycin and lincomycin can be explained by cross-resistance, as both belong to the lincosamide class of antibiotics.
The observed strong positive correlations between lincomycin and clindamycin (0.96) suggest potential co-selection mechanisms or shared resistance determinants among these compounds. Such relationships could arise from the widespread use of these antibiotics in veterinary practice, where cross-resistance is driven by overlapping resistance genes or efflux pumps targeting structurally or functionally similar antimicrobial classes. Furthermore, the overuse of lincomycin has likely exerted selective pressure which has facilitated the survival and spread of resistant strains. Inadequate dosing or suboptimal treatment durations may also have contributed to the development of antibiotic resistance, as these practices fail to eradicate bacteria completely, allowing resistant strains to proliferate. Additional contributing factors include horizontal gene transfer, genetic adaptation, and varying patterns of antibiotic use.
In the case of lincomycin and clindamycin, the nearly complete correlation reflects their classification within the lincosamide group, which shares a similar mechanism of action targeting bacterial protein synthesis. This finding aligns with previous reports indicating that resistance to one often predicts resistance to the other, driven by the erm genes responsible for methylation of the 23S rRNA component of the 50S ribosomal subunit. Similarly, the correlations between tilozin and both lincomycin (0.92) and clindamycin (0.95) may reflect their use in livestock production.
The correlation between enrofloxacin and vancomycin (0.83), while lower, is noteworthy and warrants further exploration. Though these antibiotics target different cellular processes, resistance may result from indirect selective pressures or the co-localization of resistance determinants on integrons or other genetic platforms. For example, quinolone resistance is most often associated with mutations in the quinolone-resistance determining regions (QRDRs) of DNA gyrase or topoisomerase IV, and it may coexist with overexpression of efflux pumps such as norA or oepA (45). Additionally, these mechanisms can occur on the same mobile genetic elements that also carry vancomycin resistance determinants (e.g., vanA or vanB), thereby enabling co-transfer of multiple resistance traits (46).
The observed differences in utilization trends are likely influenced by variations in antibiotic selection practices associated with specific production stages and age groups. Notably, adult herds may undergo multiple treatment cycles with different classes of antibiotics over their lifetime, potentially leading to a broader spectrum of selective pressures. Additionally, higher antibiotic usage in larger colonies, driven by the increased need to manage disease outbreaks and ensure productivity, may further exacerbate selection pressure, contributing to the emergence and persistence of antimicrobial resistance.
Overall, the high levels of resistance are likely the result of a combination of these factors. Implementing proper antibiotic stewardship, conducting susceptibility testing, and continuously monitoring resistance levels while investigating their genetic basis are essential measures to curb the spread of resistance.
We examined the presence of major and minor toxin genes responsible for toxin production in all 146 commensal C. perfringens isolates. Statistical analysis revealed that the presence or absence of specific toxin genes influenced the degree of resistance to certain antibiotics. The major alpha toxin gene was identified in every strain, consistent with findings from Lyhs et al., who also reported its presence in all isolates. However, their results were limited to isolates from animals with necrotic enteritis or those that had been euthanized (47). We identified the minor netB toxin gene in 2.7% of isolates. In contrast, Lyhs et al. found this gene in 8% of clinical isolates but did not detect it in commensal strains (47). The beta2 toxin gene was detected in 18.5% of isolates, compared to only one isolate reported by Lyhs et al. (47). In contrast to the findings of Lyhs et al., who did not detect genes encoding the major beta, epsilon, or iota toxins in any of their isolates, our study identified the gene for beta toxin in 32.2% and for epsilon toxin in 0.7% of the examined isolates. Like Lyhs et al., we did not detect the major iota toxin gene in any strains (47). Furthermore, the minor entero toxin gene was not identified in any isolate. Mohiuddin et al. reported the presence of the netB toxin gene in two poultry isolates but did not detect genes coding for major beta, epsilon, iota, or minor entero toxins (48). In our study, statistical analysis of the presence or absence of toxin genes revealed significant findings. Notably, enrofloxacin resistance was significantly lower in our isolates harboring the netB gene (p = 0.0049), indicating a possible link between virulence and susceptibility profiles. In contrast, Slavic et al. reported that strains resistant to clindamycin and erythromycin were significantly less likely to carry the beta2-toxin gene; however, no significant relationships between antibiotic sensitivity and the presence of toxin genes was identified in isolates from other animal species, such as cattle, chickens, or turkeys (44). We also examined the effect of simultaneous presence of multiple toxin genes. The resistance to lincomycin was significantly higher (p = 0.0379) in strains lacking toxin genes than in those harboring multiple toxin genes. Wei et al. described significant differences between C. perfringens isolates from healthy chickens and those with clinical signs of necrotic enteritis. Specifically, isolates from diseased birds showed significantly reduced sensitivity to clindamycin compared to isolates from healthy birds (33).
Our findings underscore the critical importance of continuous monitoring of antimicrobial resistance patterns in both commensal and clinical C. perfringens strains. Future studies incorporating larger sample sizes, clinical isolates, and advanced molecular techniques, such as next-generation sequencing, could provide deeper insights into the relationships between resistance mechanisms and virulence factors. These efforts will be pivotal in developing effective, evidence-based antibiotic use strategies and preserving the efficacy of critically important antimicrobials for both veterinary and human health.
4 Materials and methods
4.1 The origin of the strains
The examined strains were collected between February 2022 and May 2023 during routine diagnostic investigations conducted by veterinarians serving large-scale livestock farms in collaboration with poultry health experts of the Department of Animal Hygiene, Herd Health and Mobile Clinic of University of Veterinary Medicine, Budapest. Samples, including 15 cloacal swabs per farm, were collected using Amies-type transport medium (Biolab Zrt., Budapest, Hungary). Samples were obtained from the intestinal contents or cloacal swabs of clinically healthy broiler chickens and submitted to the national reference laboratory. For the isolation of C. perfringens, samples were streaked onto CHROMagar™ C. perfringens selective agar (Chebio Fejlesztő Kft., Budapest, Hungary) and incubated at 37°C for 24–48 h under anaerobic conditions using the BD GasPak™ anaerobic system (VWR International Kft., Debrecen, Hungary). Colonies showing characteristic morphology were subcultured for purification. Species-level identification of the isolates was confirmed by MALDI-TOF mass spectrometry. The confirmed C. perfringens isolates were then provided to us for further analysis.
Pure cultures of the isolates were prepared on tryptic soy agar (Biolab Zrt., Budapest, Hungary) and stored at −80°C in a Microbank™ system (Pro-Lab Diagnostics, Richmond Hill, Canada).
Metadata accompanying the samples included information on the organ of origin (cloaca) and the specific location of collection. Based on the collection sites, the samples were categorized into Hungary’s seven administrative regions during recordkeeping.
4.2 Minimum inhibitory concentration determination
The phenotypic presence of AMR was determined by measuring MIC values following the guidelines of the Clinical Laboratory Standard Institute (CLSI) (49). Breakpoints were also defined according to CLSI guidelines (49) and compared with the ECOFF established by the EUCAST.
The isolates, stored at −80°C, were suspended in 3 mL of cation-adjusted Muller-Hinton broth (CAMHB) and incubated at 37°C for 18–24 h prior to testing. The MIC determinations were conducted using 96-well microtiter plates (VWR International, LLC., Debrecen, Hungary). All wells, except those in the first column, were filled with 90 μL of CAMHB. Stock solutions of the tested antibiotics (Merck KGaA, Darmstadt, Germany) were prepared following CLSI guidelines (49). Amoxicillin and amoxicillin-clavulanic acid were prepared in a 2:1 ratio using phosphate buffer (pH 7.2, 0.01 mol/L). Penicillin and imipenem were dissolved in phosphate buffer (pH 6, 0.1 mol/L), while ronidazole and metronidazole were prepared in dimethyl sulfoxide (DMSO). Distilled water was used for ceftriaxone, tilozin, tilmicosin, lincomycin, clindamycin, oxytetracycline, and vancomycin. Enrofloxacin was prepared in distilled water with 1 mol/L NaOH. From each 512 μg/mL working solution, 180 μL was added to the first column of the microtiter plate, followed by serial twofold dilutions across the plate. After the 10th column, 90 μL of excess solution was discarded, leaving 90 μL in each well. Bacterial suspensions adjusted to 0.5 McFarland standard using a nephelometer (ThermoFisher Scientific, Budapest, Hungary) were inoculated into the microtiter plates, starting from the 11th column and proceeding backward, with 10 μL per well (49). Evaluation was performed using the SensititreTM SWINTM automatic MIC reader (ThermoFisher Scientific, Budapest, Hungary) and VIZION system software version 3.4 (ThermoFisher Scientific, Budapest, Hungary, 2024). The quality control strain was C. perfringens (ATCC 13124).
4.3 PCR tests
Using PCR, we mapped the gene sets responsible for major and minor toxin production in each strain. DNA extraction was performed from bacterial suspensions using the Zymo Quick-DNA Fungal/Bacterial Miniprep Kit (Zymo Research, Murphy Ave., Irvine, CA, United States), following the manufacturer’s protocol. To disrupt the bacterial cells and release genomic DNA, a Qiagen TissueLyzer LT (Qiagen GmBH, Hilden, Germany) was used at 50 Hz for 5 min. The PCR assays utilized the Kylt Clostridium perfringens kit, specifically designed for real-time PCR detection of DNA (Kylt, Höltinghausen, Germany). This kit is optimized for detecting major (alpha, beta, epsilon, iota) and minor (beta2, entero, netB) toxin genes in bird-derived samples. All procedures adhered strictly to the manufacturer’s guidelines, with the extracted nucleic acids stored at −20°C until further use.
PCR assays were carried out using the CFX Opus Dx Real-Time PCR system (Bio-Rad Hungary Ltd., Budapest, Hungary). The data analysis was performed with the Bio-Rad CFX Maestro software version 5.3.022.1030 (Bio-Rad Hungary Ltd., Budapest, Hungary).
4.4 Statistical analysis
The data analysis was conducted using R version 4.1.0 (50). The normality of data distribution was tested using the Shapiro–Wilk test. For datasets that deviated from normal distribution, non-parametric statistical methods were employed. The Kruskal-Wallis test (51), was utilized to evaluate resistance levels across multiple sample groups. This method is particularly suitable for comparing medians when normality assumptions are not met. To further investigate group differences, post hoc pairwise comparisons were conducted using the Mann–Whitney U test (52), for non-parametric data and two-sample t-tests for normally distributed data. Bonferroni correction was applied to reduce the likelihood of Type I errors from multiple comparisons, although it is acknowledged that this approach may increase the risk of Type II errors (failing to detect true differences) (53).
Correlation analysis was performed to identify relationships among resistance profiles to various active substances. PCA (54) was employed to uncover underlying patterns in the data and to visualize similarities and differences across samples. PCA effectively reduces the dimensionality of complex datasets by transforming them into principal components, with each component representing a linear combination of the original variables. The first principal component captures the largest variance, with subsequent components accounting for progressively smaller variances. The orthogonality of these components ensures that they are uncorrelated, simplifying the interpretation of complex data relationships.
Hierarchical cluster analysis was also performed to identify clusters of isolates based on resistance profiles. The results were visualized using a dendrogram (55); a tree-like diagram illustrating the hierarchical relationships among clusters. This method groups observations into clusters that share similar characteristics, distinguishing them from other clusters. The dendrogram provides a clear visualization of the clustering process and the relative relationships between groups.
Correlation analysis further explored the strength and direction of relationships between variables. A positive correlation indicates that an increase in one variable corresponds to an increase in the other, while a negative correlation implies an inverse relationship. The correlation coefficient, ranging from −1 to +1, quantifies the strength of the relationship, with +1 representing a perfect positive correlation, −1 a perfect negative correlation, and 0 indicating no linear relationship.
5 Conclusion
In conclusion, our study highlights the significance of the global spread of AMR and underscores the importance of examining C. perfringens strains from both veterinary and public health perspectives. The resistance rates observed for penicillin and amoxicillin, alongside the detection of virulence factors, reaffirm the pivotal role of commensal-strains in maintaining resistance reservoirs. Our findings emphasize the necessity of regular monitoring, particularly in large-scale poultry operations, to quickly identify emerging resistance trends. Future studies with larger sample sizes and the use of next-generation sequencing to elucidate the genetic basis of resistance in multi-drug-resistant strains can provide deeper insights, ultimately contributing to the development of more effective treatment and prevention strategies.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The study was exempt from animal experimentation approval according to the Animal Welfare Committee of the University of Veterinary Medicine Budapest, as it involved only non-invasive diagnostic sampling (tracheal and cloacal swabs) from poultry during routine veterinary visits. All flock owners provided written informed consent for participation in the study, and all data were handled anonymously in full compliance with the General Data Protection Regulation (GDPR, EU 2016/679).
Author contributions
ÁK: Conceptualization, Methodology, Resources, Software, Writing – review & editing. ÁS: Writing – original draft, Data curation, Formal analysis, Project administration, Visualization. FB: Project administration, Writing – original draft. BC: Project administration, Writing – original draft. LK: Resources, Writing – original draft. ÁJ: Conceptualization, Funding acquisition, Investigation, Supervision, Validation, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by Project no. RRF-2.3.1-21-2022-00001 was implemented with the support provided by the Recovery and Resilience Facility (RRF), financed under the National Recovery Fund budget estimate, RRF-2.3.1–21 funding scheme.
Acknowledgments
We would like to thank Katalin Balogh and Tamásné Pénzes Imre for the preparation of the laboratory work.
Conflict of interest
LK was employed by Poultry-Care Kft.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1589747/full#supplementary-material
References
1. Prestinaci, F, Pezzotti, P, and Pantosti, A. Antimicrobial resistance: a global multifaceted phenomenon. Pathog Glob Health. (2015) 109:309–18. doi: 10.1179/2047773215Y.0000000030
2. Holmes, AH, Moore, LSP, Sundsfjord, A, Steinbakk, M, Regmi, S, Karkey, A, et al. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet. (2016) 387:176–87. doi: 10.1016/S0140-6736(15)00473-0
3. Martínez, JL. Antibiotics and antibiotic resistance genes in natural environments. Science. (2008) 321:365–7. doi: 10.1126/science.1159483
4. Davies, J. Are antibiotics naturally antibiotics? J Ind Microbiol Biotechnol. (2006) 33:496–9. doi: 10.1007/s10295-006-0112-5
5. Kovács, L, Nagy, D, Könyves, L, Jerzsele, Á, and Kerek, Á. Antimicrobial properties of essential oils – animal health aspects. Magy Allatorv Lapja. (2023) 145:497–510. doi: 10.56385/magyallorv.2023.08.497-510
6. Kerek, Á, Szabó, Á, Dobra, PF, Bárdos, K, Ózsvári, L, Fehérvári, P, et al. Determining the in vivo efficacy of plant-based and probiotic-based antibiotic alternatives against mixed infection with Salmonella enterica and Escherichia coli in domestic chickens. Vet Sci. (2023) 10:706. doi: 10.3390/vetsci10120706
7. Jerzsele, Á, Somogyi, Z, Szalai, M, and Kovács, D. Effects of fermented wheat germ extract on artificial Salmonella Typhimurium infection in broiler chickens. Magy Allatorv Lapja. (2020) 142:77–85.
8. Sebők, C, Márton, RA, Meckei, M, Neogrády, Z, and Mátis, G. Antimicrobial peptides as new tools to combat infectious diseases. Magy Allatorv Lapja. (2024) 146:181–91. doi: 10.56385/magyallorv.2024.03.181-191
9. Olasz, Á, Jerzsele, Á, Balta, L, Dobra, PF, and Kerek, Á. In vivo efficacy of different extracts of propolis in broiler salmonellosis. Magy Allatorv Lapja. (2023) 145:461–75. doi: 10.56385/magyallorv.2023.08.461-475
10. Kerek, Á, Csanády, P, and Jerzsele, Á. Antibacterial efficiency of propolis – part 1. Magy Allatorv Lapja. (2022) 144:285–98.
11. Kerek, Á, Csanády, P, Tuska-Szalay, B, Kovács, L, and Jerzsele, Á. In vitro efficacy of Hungarian propolis against bacteria, yeast, and trichomonas gallinae isolated from pigeons—a possible antibiotic alternative? Resources. (2023) 12:101. doi: 10.3390/resources12090101
12. Hetényi, N, Bersényi, A, and Hullár, I. Physiological effects of medium-chain fatty acids and triglycerides, and their potential use in poultry and swine nutrition: a literature review. Magy Allatorv Lapja. (2024) 146:651–9. doi: 10.56385/magyallorv.2024.11.651-659
13. Kovács, D, Palkovicsné Pézsa, N, Farkas, O, and Jerzsele, Á. Usage of antibiotic alternatives in pig farming: literature review. Magy Allatorv Lapja. (2021) 143:281–2.
14. Essősy, M, Fodor, I, Ihnáth, Z, Karancsi, Z, Kovács, D, Szalai, KV, et al. The possibilities of antibiotic-free broiler-hen fattening, with special reference to the use of pre- and probiotics. Magy Allatorv Lapja. (2020) 142:397–407.
15. Farkas, M, Könyves, L, Csorba, S, Farkas, Z, Józwiák, Á, Süth, M, et al. Biosecurity situation of large-scale poultry farms in Hungary according to the databases of National Food Chain Safety Office Centre for disease control and biosecurity audit system of poultry product Board of Hungary in the period of 2021–2022. Magy Allatorv Lapja. (2024) 146:723–42. doi: 10.56385/magyallorv.2024.12.723-742
16. Mag, P, Németh, K, Somogyi, Z, and Jerzsele, Á. Antibacterial therapy based on pharmacokinetic/ pharmacodynamic models in small animal medicine-1. Literature review. Magy Allatorv Lapja. (2023) 145:419–38. doi: 10.56385/magyallorv.2023.07.419-438
17. Timbermont, L, Haesebrouck, F, Ducatelle, R, and Van Immerseel, F. Necrotic enteritis in broilers: an updated review on the pathogenesis. Avian Pathol. (2011) 40:341–7. doi: 10.1080/03079457.2011.590967
18. Keyburn, AL, Boyce, JD, Vaz, P, Bannam, TL, Ford, ME, Parker, D, et al. NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens. PLoS Pathog. (2008) 4:e26. doi: 10.1371/journal.ppat.0040026
19. Rood, JI, Adams, V, Lacey, J, Lyras, D, McClane, BA, Melville, SB, et al. Expansion of the Clostridium perfringens toxin-based typing scheme. Anaerobe. (2018) 53:5–10. doi: 10.1016/j.anaerobe.2018.04.011
20. Beceiro, A, Tomás, M, and Bou, G. Antimicrobial resistance and virulence: a successful or deleterious association in the bacterial world? Clin Microbiol Rev. (2013) 26:185–230. doi: 10.1128/CMR.00059-12
21. Vilei, EM, Schlatter, Y, Perreten, V, Straub, R, Popoff, MR, Gibert, M, et al. Antibiotic-induced expression of a cryptic cpb2 gene in equine beta2-toxigenic Clostridium perfringens. Mol Microbiol. (2005) 57:1570–81. doi: 10.1111/j.1365-2958.2005.04789.x
22. Ahn, D, and Prince, A. Host-pathogen Interface: Progress in understanding the pathogenesis of infection due to multidrug-resistant Bacteria in the intensive care unit. J Infect Dis. (2017) 215:S1–8. doi: 10.1093/infdis/jiw405
23. Grenda, T, Jarosz, A, Sapała, M, Grenda, A, Patyra, E, and Kwiatek, K. Clostridium perfringens—opportunistic foodborne pathogen, its diversity and epidemiological significance. Pathogens. (2023) 12:768. doi: 10.3390/pathogens12060768
24. Hamza, D, Dorgham, SM, Elhariri, M, Elhelw, R, and Ismael, E. New insight of apparently healthy animals as a potential reservoir for Clostridium perfringens: A public health implication. J Vet Res. (2018) 62:457–62. doi: 10.2478/jvetres-2018-0073
25. Gadbois, P, Brennan, JJ, Bruce, HL, Wilson, JB, and Aramini, JJ. The role of penicillin G potassium in managing Clostridium perfringens in broiler chickens. Avian Dis. (2008) 52:407–11. doi: 10.1637/8114-091807-Reg
26. Lees, P, Concordet, D, Aliabadi, FS, and Toutain, P-L. Drug selection and optimization of dosage schedules to minimize antimicrobial resistance In: FM Aarestrup editor. Antimicrobial resistance in Bacteria of animal origin. Ames, Iowa, USA: John Wiley & Sons, Ltd (2005). 49–71.
27. Huang, T-M, Lin, TL, and Wu, CC. Antimicrobial susceptibility and resistance of chicken Escherichia coli, Salmonella spp., and Pasteurella multocida isolates. Avian Dis. (2009) 53:89–93. doi: 10.1637/8268-021608-Reg.1
28. Toutain, PL, Del Castillo, JRE, and Bousquet-Mélou, A. The pharmacokinetic-pharmacodynamic approach to a rational dosage regimen for antibiotics. Res Vet Sci. (2002) 73:105–14. doi: 10.1016/S0034-5288(02)00039-5
29. McKellar, QA, Sanchez Bruni, SF, and Jones, DG. Pharmacokinetic/pharmacodynamic relationships of antimicrobial drugs used in veterinary medicine. J Vet Pharmacol Ther. (2004) 27:503–14. doi: 10.1111/j.1365-2885.2004.00603.x
30. Dürre, P. Physiology and sporulation in Clostridium. Microbiol Spectr. (2014) 2:2012. doi: 10.1128/microbiolspec.tbs-0010-2012
31. Popoff, MR, and Bouvet, P. Genetic characteristics of toxigenic Clostridia and toxin gene evolution. Toxicon. (2013) 75:63–89. doi: 10.1016/j.toxicon.2013.05.003
32. Hassani, S, Pakbin, B, Brück, WM, Mahmoudi, R, and Mousavi, S. Prevalence, antibiotic resistance, toxin-typing and genotyping of Clostridium perfringens in raw beef meats obtained from Qazvin City, Iran. Antibiotics (Basel). (2022) 11:340. doi: 10.3390/antibiotics11030340
33. Wei, B, Cha, S-Y, Zhang, J-F, Shang, K, Park, H-C, Kang, J, et al. Antimicrobial susceptibility and association with toxin determinants in Clostridium perfringens isolates from chickens. Microorganisms. (2020) 8:1825. doi: 10.3390/microorganisms8111825
34. Martel, A, Devriese, L, and Cauwerts, K. Susceptibility of Clostridium perfringens strains from broiler chickens to antibiotics and anticoccidials. Avian Pathol. (2004) 33:3–7. doi: 10.1080/0307945031000163291
35. Osman, K, and Elhariri, M. Antibiotic resistance of Clostridium perfringens isolates from broiler chickens in Egypt. Rev Sci Tech. (2013) 32:841–50. doi: 10.20506/RST.32.2.2212
36. Mwangi, S, Timmons, J, Fitz-coy, S, and Parveen, S. Characterization of Clostridium perfringens recovered from broiler chicken affected by necrotic enteritis. Poult Sci. (2019) 98:128–35. doi: 10.3382/ps/pey332
37. Silva, ROS, Salvarani, FM, Assis, RA, Martins, NRS, Pires, PS, and Lobato, FCF. Antimicrobial susceptibility of Clostridium perfringens strains isolated from broiler chickens. Braz J Microbiol. (2009) 40:262–4. doi: 10.1590/S1517-838220090002000010
38. Akhi, MT, Asl, SB, Pirzadeh, T, Naghili, B, Yeganeh, F, Memar, Y, et al. Antibiotic sensitivity of Clostridium perfringens isolated from faeces in Tabriz, Iran. Jundishapur J Microbiol. (2015) 8:e20863. doi: 10.5812/jjm.20863v2
39. Duc, HM, Hoa, TTK, Ha, CTT, Van Hung, L, Van Thang, N, Minh Son, H, et al. Prevalence and antibiotic resistance profile of Clostridium perfringens isolated from pork and chicken meat in Vietnam. Pathogens. (2024) 13:400. doi: 10.3390/pathogens13050400
40. de Souza, BF, Capra Pezzi, L, Tsao, M, Franco de Oliveira, T, and Manoela Dias Macedo, S. Stability and degradation products of imipenem applying high-resolution mass spectrometry: an analytical study focused on solutions for infusion. Biomed Chromatogr. (2019) 33:e4471. doi: 10.1002/bmc.4471
41. Zych, S, Adaszyńska-Skwirzyńska, M, Szewczuk, MA, and Szczerbińska, D. Interaction between Enrofloxacin and three essential oils (cinnamon bark, clove bud and Lavender flower)—A study on multidrug-resistant Escherichia coli strains isolated from 1-day-old broiler chickens. Int J Mol Sci. (2024) 25:5220. doi: 10.3390/ijms25105220
42. Gehring, R, Mochel, JP, and Schmerold, I. Understanding the background and clinical significance of the WHO, WOAH, and EMA classifications of antimicrobials to mitigate antimicrobial resistance. Front Vet Sci. (2023) 10:1153048. doi: 10.3389/fvets.2023.1153048
43. Nhung, NT, Chansiripornchai, N, and Carrique-Mas, JJ. Antimicrobial resistance in bacterial poultry pathogens: A review. Front Vet Sci. (2017) 4:126. doi: 10.3389/fvets.2017.00126
44. Slavić, Đ, Boerlin, P, Fabri, M, Klotins, KC, Zoethout, JK, Weir, PE, et al. Antimicrobial susceptibility of Clostridium perfringens isolates of bovine, chicken, porcine, and Turkey origin from Ontario. Can J Vet Res. (2011) 75:89–97.
45. Hooper, DC, and Jacoby, GA. Mechanisms of drug resistance: quinolone resistance. Ann N Y Acad Sci. (2015) 1354:12–31. doi: 10.1111/nyas.12830
46. Ruiz, J. Unusual and unconsidered mechanisms of bacterial resilience and resistance to quinolones. Life. (2024) 14:383. doi: 10.3390/life14030383
47. Lyhs, U, Perko-Mäkelä, P, Kallio, H, Brockmann, A, Heinikainen, S, Tuuri, H, et al. Characterization of Clostridium perfringens isolates from healthy turkeys and from turkeys with necrotic enteritis. Poult Sci. (2013) 92:1750–7. doi: 10.3382/ps.2012-02903
48. Mohiuddin, M, Song, Z, Liao, S, Qi, N, Li, J, Lv, M, et al. Animal model studies, antibiotic resistance and toxin gene profile of NE reproducing Clostridium perfringens type A and type G strains isolated from commercial poultry farms in China. Microorganisms. (2023) 11:622. doi: 10.3390/microorganisms11030622
49. Clinical and Laboratory Standards Institute. CLSI. Methods for dilution antimicrobial susceptibility tests for Bacteria that grow aerobically. 11. th ed. Wayne, PA: Clinical and Laboratory Standards Institute (2018).
50. R Core Team. (2020). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available online at: http://www.r-project.org/index.html [Accessed September 4, 2022]
51. Kruskal, WH, and Wallis, WA. Use of ranks in one-criterion variance analysis. J Am Stat Assoc. (1952) 47:583–621.
52. Fay, MP, and Proschan, MA. Wilcoxon-Mann-Whitney or t-test? On assumptions for hypothesis tests and multiple interpretations of decision rules. Statistics Surveys. (2010) 4:1–39. doi: 10.1214/09-SS051
54. Jolliffe, IT, and Cadima, J. Principal component analysis: a review and recent developments. Philos Trans A Math Phys Eng Sci. (2016) 374:20150202. doi: 10.1098/rsta.2015.0202
Keywords: Clostridium perfringens, antimicrobial resistance, minimum inhibitory concentration (MIC), PCR, chickens, Hungary
Citation: Kerek Á, Szabó Á, Barnácz F, Csirmaz B, Kovács L and Jerzsele Á (2025) Antimicrobial susceptibility profiles of commensal Clostridium perfringens isolates from chickens in Hungarian poultry farms between 2022 and 2023. Front. Vet. Sci. 12:1589747. doi: 10.3389/fvets.2025.1589747
Edited by:
María-Aránzazu Martínez, Complutense University of Madrid, SpainReviewed by:
Jacob Kwada Kwaga, Ahmadu Bello University, NigeriaRogers Ruyu Azabo, Sokoine University of Agriculture, Tanzania
Copyright © 2025 Kerek, Szabó, Barnácz, Csirmaz, Kovács and Jerzsele. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ádám Kerek, a2VyZWsuYWRhbUB1bml2ZXQuaHU=