 Melikasadat Mollabashi1,2
Melikasadat Mollabashi1,2 Alonza Klopfer1,3
Alonza Klopfer1,3 Thainá Lunardon1,2
Thainá Lunardon1,2 Nikolia Darzenta1,2
Nikolia Darzenta1,2 Emily Davis1Matt Murray1
Emily Davis1Matt Murray1 Scarlett M. Sumner3*
Scarlett M. Sumner3* Maria C. Naskou1,2*
Maria C. Naskou1,2*- 1Scott-Ritchey Research Center, College of Veterinary Medicine, Auburn University, Auburn, AL, United States
- 2Department of Pathobiology, College of Veterinary Medicine, Auburn University, Auburn, AL, United States
- 3Department of Clinical Sciences, College of Veterinary Medicine, Auburn University, Auburn, AL, United States
Introduction: Platelet derived products have gained increasing attention as promising alternative biologicals for the treatment of canine wounds. Specifically, platelets play a crucial role during the inflammatory phase of wound healing due to the release of chemokines, proteins, cytokines, and growth factors. Additionally, platelets possess antimicrobial properties, which can be influenced by their manufacturing process, platelet and leukocyte concentration, activation method, and the presence of plasma and complement. The objective of this study was to assess how various preparation methods of platelet products affect their antimicrobial effect against bacteria commonly isolated from wounds.
Methods: In this study, blood was collected from eight purpose-bred dogs, and platelet-rich plasma was produced using two methods of centrifugation, one leukocyte-enriching and one leukocyte-reducing. Some samples were processed for plasma depletion and platelet lysate was subsequently generated through freeze–thaw cycles. Additionally, portions of platelet lysate samples underwent heat treatment for complement inactivation. All treatment groups were tested against four common bacteria found in canine skin wounds: Escherichia coli, Enterococcus faecalis, Staphylococcus pseudintermedius, and Staphylococcus aureus. The antimicrobial effect of various lysate formulations was evaluated using a bacteria-spiking (time-killing) assay.
Results: Platelet lysate significantly reduced the number of S. aureus and E. coli after 3 h compared to culture media. No significant differences were noted in the log reduction of bacteria between the centrifugation techniques. After depleting plasma, the log reduction of S. pseudintermedius was significantly less than before plasma depletion, whereas the opposite was seen for E. faecalis after 3 h. Complement-depleted plasma led to a significantly lower log reduction for E. faecalis after 24 h compared to platelet lysate.
Discussion: Therefore, the presence of plasma and complement proteins in platelet lysate appear to play a critical role in inhibiting the growth of certain bacterial strains, whereas the leukocyte concentration does not have a significant effect. Further research is needed to identify the ideal formulation and dose of canine platelet lysate as an antimicrobial and wound healing treatment.
Introduction
Wound healing is a complex multicellular dynamic event characterized by several processes such as inflammation, coagulation, epithelization, tissue regeneration, and granulation tissue formation (1). Platelets play a crucial role during the inflammatory phase of wound healing due to their alpha granules releasing most of the chemokines, proteins, cytokines, and growth factors responsible for activating and recruiting other cells involved in inflammatory responses (2). Platelet-rich plasma (PRP) is a platelet-derived product containing an increased concentration of platelets, widely known to benefit wound healing processes and inhibit common pathogens found in wounds (3–7). Platelet concentrates, such as PRP, can be subjected to several manufacturing techniques that include lysis of platelets to release their chemotactic and growth factors (8). The resulting product is called platelet lysate (PL) (8).
PL is an acellular product containing platelet-derived growth factors that can serve as an off-the-shelf alternative to PRP (9). As it is acellular, issues of immunogenicity are reduced, which is another attractive attribute of PL as a therapeutic (9). Additionally, PL can be pooled from several donors to decrease donor variability and can be stored for extended periods in the freezer (9–11). Another attractive aspect of PL regarding wound healing is its antimicrobial potential. Specifically, the antimicrobial properties originate from the release of antimicrobial peptides (AMPs) and an array of growth factors (12–15). The activity of AMPs has been shown to initiate pore formation within bacterial membranes, which can evoke cell death (16). New studies have demonstrated that canine platelet-derived products display antimicrobial properties in wounds infected with antibiotic-resistant Gram-negative (17) and Methicillin-resistant Staphylococcus aureus (MRSA) (18), while equine PL was recently found to inhibit the growth of Enterococcus faecalis and Staphylococcus aureus (8). Additionally, equine PL has shown both concentration-dependent and independent inhibition effects depending on the type of bacteria being treated (8). Therefore, PL has dual treatment potential, since it can be used for wound healing purposes, and as an alternative to traditional antibiotics for regulating wound-related bacterial infections, including those with antibiotic resistance.
PRP and PL can be manufactured in different ways, including various commercial systems, manual lab centrifugation methods, gravity filtration methods, and plateletpheresis (19). Final platelet concentrations of PRP vary greatly within the literature, even when using a single method of production (20, 21). Variability also exists regarding the leukocyte concentration (20, 21) and the method of platelet activation (22, 23). The variability of platelet products in the manufacturing processes and the inflammatory cytokine concentration, growth factors, and cellular composition can affect tissue metabolism in vitro and in vivo (7, 22–25). These factors can ultimately alter the efficacy of the final product.
Similarly, the antimicrobial potential of platelet products can be affected by the concentration of leukocytes, however, the results are controversial. Some studies showed that leukocytes were not essential for maintaining the antimicrobial properties of platelets (17, 26, 27). Specifically, leukocytes can produce pro-inflammatory protease and acid hydrolases that might induce an inflammatory response (28). On the contrary, some studies found that the presence of leukocytes in platelet products amplified their ability to inhibit bacterial growth (29, 30), because leukocytes produce myeloperoxidase that kills bacteria (31).
In additional to the platelet themselves, plasma can accelerate the wound healing process as well as provide antimicrobial properties (32). One specific component that likely contributes to its antimicrobial effect is the complement system. The complement system is comprised of unique plasma proteins that interact to opsonize pathogens while employing inflammatory responses that correspond to invading infections (33) and act as an extracellular and intracellular defense mechanism (34). Certain complement proteins, such as C3 and C5, can initiate cell differentiation, maintain immunological tolerance, and activate cell mediate responses (34). The literature demonstrates that specific plasma proteins, specifically C3a, contribute to the direct killing of Escherichia coli, Pseudomonas aeruginosa, and Enterococcus faecalis by binding and inducing breaks in bacterial membranes (35). Finally, Burnouf et al. found that complement activation pathways are essential for the antimicrobial function of human plasma and platelet-derived products against certain bacteria such as Escherichia coli, Staphylococcus aureus, Pseudomonas aeruginosa, and Klebsiella pneumoniae (36).
This study aimed to compare the antimicrobial activity of canine PL generated by different methods, including a leukocyte-reduced and a leukocyte-rich method, against bacteria commonly present in canine skin wounds. Subsequently, we aimed to identify whether the presence of plasma and complement are responsible for the antimicrobial action of PL. We hypothesized that canine PL would display antimicrobial effects against common bacteria found in canine skin wounds compared to standard bacteria growth media and that plasma and heat-sensitive components- namely the complement system- would improve its effectiveness.
Materials and methods
Canine blood donors
In this study, eight canine blood donors were recruited for whole blood donation. Dogs were purpose-bred beagles, four females and four males, with a mean age of 5.67 +/− 0.5 years and a mean weight of 14.83 +/− 2 kg. All donors were considered healthy based on history, preliminary blood work (complete blood cell count and chemistry panel), and physical examination. The study’s protocol (2022–5,085) was approved by the Auburn University Institutional Animal Care and Use Committee (IACUC).
Generation of platelet concentrates
Approximately 100 mL of blood was collected in ACD-A tubes from each canine donor. A portion of the blood was also collected into an EDTA tube for a complete blood count (CBC) using the Heska hematology analyzer (Heska Element HT5, Heska, Colorado, United States). Platelet rich plasma (PRP), leukocyte-rich platelet rich plasma (L-PRP), and platelet-poor plasma (PPP) were manufactured under a biosafety laminar flow cabinet (Thermo Scientific Biological Safety Cabinets) (37). Briefly, two different manual centrifugation methods were used to isolate platelets from whole blood. For PRP, whole blood was centrifuged at 1000 g for 5 min. The plasma layer was then collected and centrifuged at 1500 g for 15 min to pellet the platelets. For the L-PRP, whole blood was centrifuged at 180 g for 20 min. The plasma layer and buffy coat were then collected and centrifuged at 650 g for 15 min to pellet the platelets. For each method, the PPP was removed without disturbing the platelet pellet, leaving two milliliters remaining for pellet resuspension. The platelet pellet was resuspended, and a platelet count was performed. All platelet concentrations were adjusted to 0.8–1 × 106 platelets/μl using PPP. Samples were stored at −80°C.
Plasma depletion
Before storage at −80°C, a portion of the PRP, L-PRP, and PPP samples were plasma depleted by centrifugation (Sorvall X Pro Series Centrifuge, ThermoFisher Scientific, Osterode am Harz, Germany) at 3000 g for 30 min. Plasma was removed without disturbing the pellet and the pellet was resuspended with an equal volume of phosphate-buffered saline without calcium and magnesium (PBS −/−, VWR, United States) to produce leukocyte-reduced platelet pellet concentrate (PPC) and leukocyte-rich platelet pellet concentrate (LPPC). Samples were then stored at −80°C.
Generation of platelet lysates
PPP, PRP, LPRP, PPC, and LPPC formulations from eight dogs were thawed and pooled to generate three lots of each formulation, each containing lysate from three donors. All samples then underwent four additional freeze–thaw cycles using liquid nitrogen and a dry bath at 37°C (Precision GP 20, ThermoFisher Scientific, Newington, NH). Samples were placed in a microcentrifuge (Fresco 21 Centrifuge, ThermoFisher Scientific, Osterode am Harz, Germany) at 20,000 g for 20 min. The supernatant was collected as the final products: processed PPP (PPP), leukocyte-reduced platelet lysate (PL), leukocyte-rich platelet lysate (LPL), leukocyte-reduced platelet pellet lysate (PPL), and leukocyte-rich platelet pellet lysate (LPPL). Samples were stored at −80°C.
Heat treatment (complement inactivation)
Heat treatment was performed on a portion of the PL, LPL, PPL, and LPPL formulations via a high-heat dry bath at 56°C for 30 min, followed by an immediate ice bath for 5 min (37). Samples were placed in a microcentrifuge at 10,000 g for 15 min at 4°C. The supernatant was collected to provide the final products: heat-treated leukocyte-reduced platelet lysate (hPL), heat-treated leukocyte-rich platelet lysate (hLPL), heat-treated leukocyte-reduced platelet pellet lysate (hPPL), and heat-treated leukocyte-rich platelet pellet lysate (hLPPL). Samples were stored at −80°C.
Spiking assay (time-killing assay)
The spiking assay was performed with four clinically isolated bacteria obtained from ATCC: Escherichia coli (E. coli; ATCC #25922), Enterococcus faecalis (E. faecalis; ATCC #19433), Staphylococcus pseudintermedius (S. pseudintermedius; ATCC #49051), and Staphylococcus aureus (S. aureus; ATCC #12600). Experiments were performed under a biosafety hood using an aseptic technique. First, 1 mL of brain heart infusion broth (BHI; Becton Dickinson, Sparks MD) was used to rehydrate the freeze-dried bacteria. Resuspended bacteria were incubated at 37°C overnight with shaking at 200 rpm in a culture tube with a total of four milliliters of BHI broth. Following overnight incubation, the optical density (OD) was measured for each culture tube using duplicate aliquots in 96-well plates via Biotek Gen5 (Synergy HI Microplate Reader, Agilent Technologies Inc. Winooski, VT) at OD600. BHI broth was used as a positive control. Bacterial suspensions were adjusted to 0.3 OD, which corresponds to 108 CFU/mL (38) prior to the spiking assay.
Each treatment group (PL, LPL, PPL, LPPL, hPL, hLPL, hPPL, hLPPL) at 90% v/v was added to a sterile tube containing the appropriate volume (10%) of adjusted bacterial stock suspension (108 CFU/mL). BHI, PPP, and PBS were tested as positive and negative controls, accordingly. Treatments were incubated at 37°C under constant rotation (200 rpm) for 3 and 24 h. At each time point, cultures were serially diluted in PBS (−/−) (six 10-fold dilutions). Nutrient agar plates were inoculated with 200 μL aliquots from each dilution and incubated overnight at 37°C. Bacterial colonies were manually counted using a colony counter (Reichert Quebec Darkfield). Plates containing approximately 30–300 CFUs were selected for analysis from each experimental cohort.
Statistical analysis
To reduce individual variability, platelet lysates from three dogs were pooled per lot and a total of three lots were generated (11). A power analysis showed that a sample size of 3 would be sufficient to test the antimicrobial properties of lysates to achieve a statistical significance of p < 0.05 and power = 90%. Assessments between spiking assay results were based on the log reductions compared to bacteria cultures in BHI [Log reduction = Log (bacterial count of positive control/bacterial count of treatment)]. All data were imported into a statistical analysis program (Graphpad Prism; Graphpad Software Inc. San Diego, CA, United States). Normality was assessed by the visual examination of histograms of the residual, normal plots of residuals, and via utilizing the Shapiro-Wilks test. The equality of variances was calculated utilizing Levene’s test and plotting residuals against the fitted value. Hypotheses were tested by repeated measures or 2-way analysis of variance (ANOVA). Tukey’s test was used to adjust for multiple paired comparisons. All statistical analysis was done at p < 0.05 level of significance. Continuous data were summarized and reported as mean ± standard deviation. Samples were analyzed in duplicates.
Results
Hematologic values
The mean concentration of platelets, white blood cells (WBC), and hematocrit (HCT), from the whole blood and platelet products are shown in Table 1. The mean (± SD) platelet concentration for whole blood was 239.375 ± 80.627 × 103/μL, the WBC concentration was 7.89 ± 1.64 × 103/μL, and the HCT was 53.32% ± 6.79. Compared to whole blood, there was an 11.8-fold increase in platelet concentration for PRP (2,837 ± 1335.605 × 103/μL) and a 9.46-fold increase for L-PRP (2,266.6 ± 1494.311 × 103/μL). The WBC concentration of PRP decreased 4.55-fold compared to whole blood (1.728 ± 6.69 × 103/μL) while L-PRP had a 2-fold decrease in WBC concentration compared to whole blood (3.926 ± 2.842 × 103/μL). There was minimal blood hemodilution for both PRP and L-PRP. The concentration of platelets, WBC, and HCT were negligible in all lysate formulation groups (PL, LPL, PPL, and LPPL; data not shown).
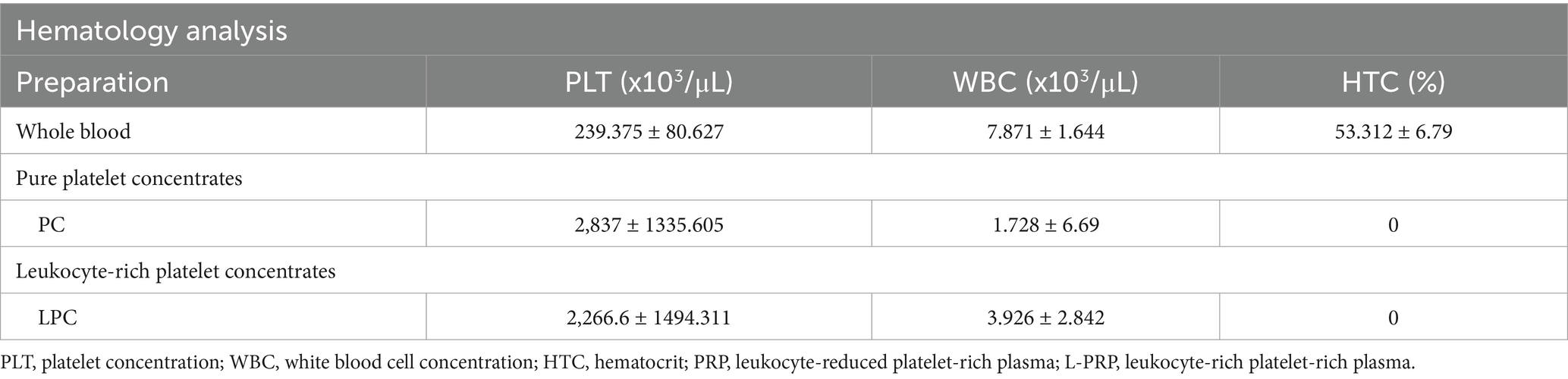
Table 1. Descriptive results of hematological data presented as mean ± standard deviation from whole blood, and platelet-derived concentrates.
Spiking assay
The effect of platelet concentration
The log reduction in growth of the E. coli, E. faecalis, S. pseudintermedius, and S. aureus after 3 and 24 h of PL or PPP treatment is demonstrated in Figure 1. PL caused a significant log reduction of S. aureus (1.724 ± 0.7120, p = 0027) and E. coli (4.238 ± 0.8227, p = 0.0486) after 3 h. However, this effect was not significant at 24 h. PPP resulted in a significant log reduction of E. faecalis after 3 (0.7937 ± 0.1841, p = 0.0220) and 24 h (1.215 ± 0.9646, p = 0.0149). A nonsignificant trend was noted for PL to achieve a greater log reduction than PPP for E. coli, S. pseudintermedius, and S. aureus after both 3 and 24 h. These findings suggest that platelets are essential for the antimicrobial action of PL against all bacteria except E. faecalis.
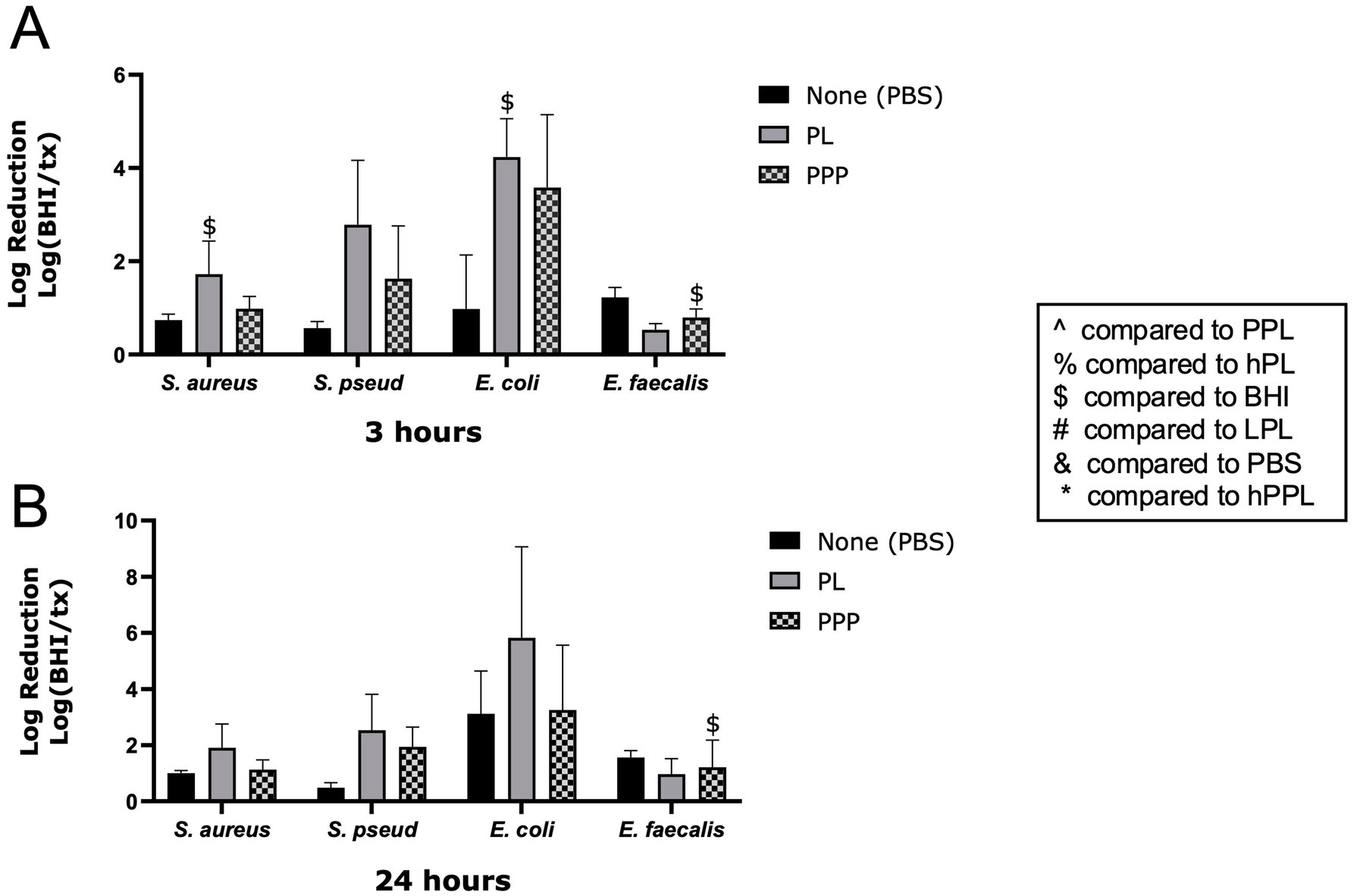
Figure 1. Log reduction of S. aureus, S. pseudintermedius, E. coli, and E. faecalis following the addition of PBS, PL, and PPP after 3 h (A) and 24 h (B). Data are presented as mean bacteria log reduction ± S. D. normalized to bacteria grown in BHI according to the following formula: Log (BHI/treatment). $ p < 0.05 compared to BHI; n = 3 lots of pooled platelet lysate. PBS, Phosphate Buffered Saline; PL, Platelet Lysate; PPP, Platelet-Poor Plasma; PPL, leukocyte-reduced platelet pellet lysate; hPL, Heat-treated Platelet Lysate; BHI, brain heart infusion broth; LPL, Leukocyte-Rich Platelet Lysate; hPPL, Heat-treated leukocyte-reduced platelet pellet lysate.
The effect of leukocyte concentration
The log reduction of bacteria treated with PL and LPL is shown in Figures 2. LPL caused a significant log reduction of S. aureus at 3 (1.576 ± 0.1900, p = 0.0237) and 24 h (1.932 ± 0.2428, p = 0.0258). At 3 h, LPL had a significantly greater log reduction of S. aureus than PBS (p = 0.0120). LPL (0.3728 ± 0.3514) caused a significant, weaker, log reduction of E. faecalis compared to PL (0.9720 ± 0.5482, p = 0.0045) and PBS (1.561 ± 0.2435, p = 0.0174) after 24 h. LPPP (0.8369 ± 0.3485, 1.171 ± 1.058) and PPP (0.7937 ± 0.1841, 1.215 ± 0.9646) had a significant log reduction of E. faecalis at 3 (p = 0.0140, p = 0.0220) and 24 (p = 0.0192, p = 0.0149) hours. PL treatment exhibited a nonsignificant trend of a greater log reduction of S. aureus and S. pseudintermedius than LPL after 3 h, and for S. pseudintermedius after 24 h. The opposite trend, with LPL achieving a greater log bacteria reduction than PL, was noted for E. coli after 3 h. Therefore, leukocytes do not appear to affect the ability of platelets to inhibit bacterial growth.
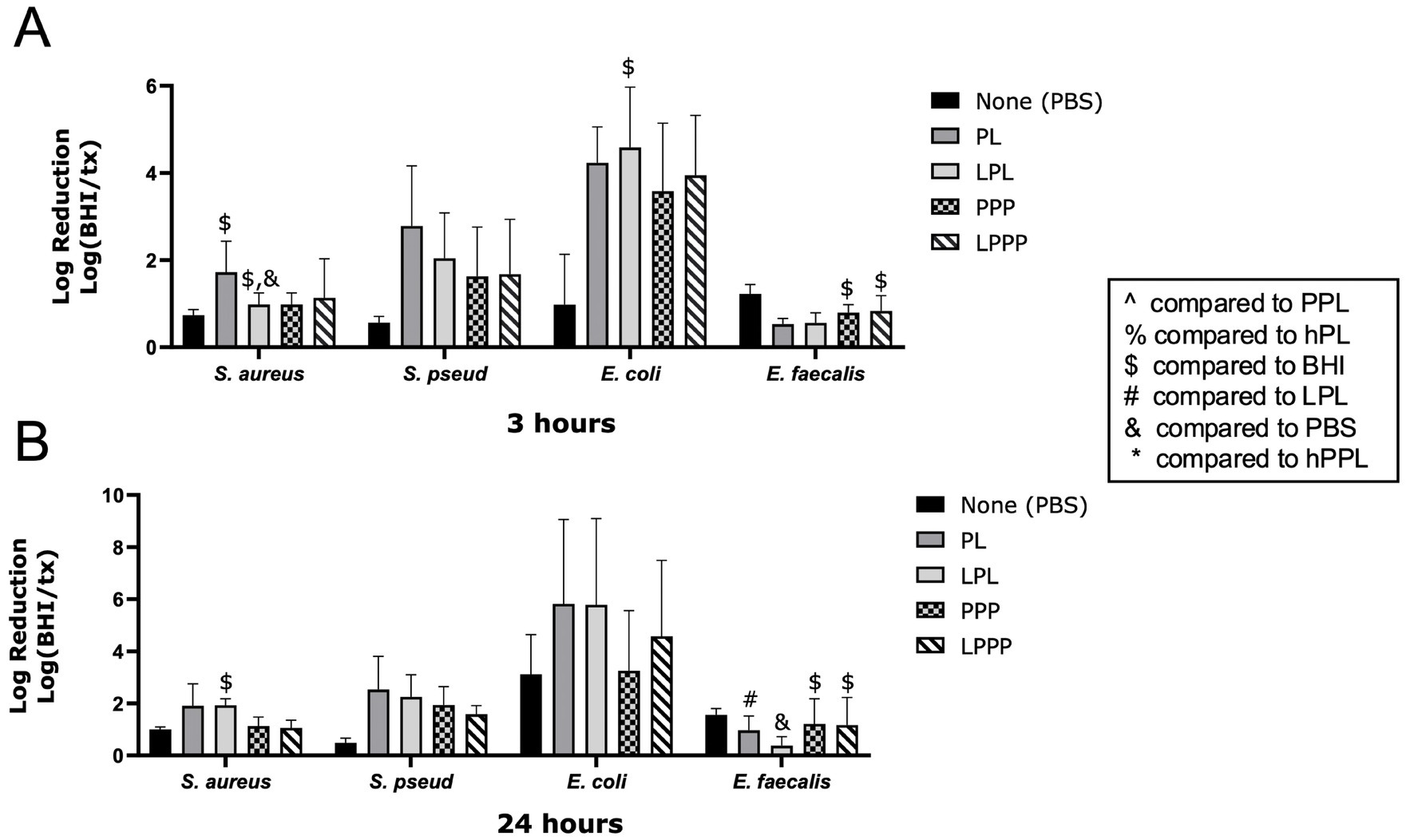
Figure 2. Log reduction of S. aureus, S. pseudintermedius, E. coli, and E. faecalis following the addition of PBS, PL, LPL, PPP, and LPPP after 3 h (A) and 24 h (B). Data are presented as mean bacteria log reduction ± S. D. normalized to bacteria grown in BHI according to the following formula: Log (BHI/treatment). $p < 0.05 compared to BHI; &p < 0.05 compared to PBS; #p < 0.05 compared to LPL; n = 3 lots of pooled platelet lysate generated from 8 donors. PBS, Phosphate Buffered Saline; PL, Platelet Lysate; LPL, Leukocyte-Rich Platelet Lysate; PPP, Platelet-Poor Plasma; LPPP, Leukocyte-Rich Platelet-Poor Plasma; PPL, leukocyte-reduced platelet pellet lysate; hPL, Heat-treated Platelet Lysate; BHI, brain heart infusion broth; hPPL, Heat-treated leukocyte-reduced platelet pellet lysate.
The effect of plasma content
The log reductions of bacteria in the presence of PL and PPL are illustrated in Figure 3. PPL caused a significant log reduction of E. faecalis at 3 (0.9623 ± 0.1493, p = 0.0396) and 24 h (1.057 ± 0.2413, p = 0.0372). At 3 h, the log reduction of E. faecalis was significantly greater with PBS than with PL (0.5296 ± 0.1331, p = 0.0260). In contrast, PL caused a greater log reduction of S. pseudintermedius at 3 (2.786 ± 1.377) and 24 h (2.538 ± 1,275) compared to PPL (1.166 ± 0.4440, p = 0.0478 and 0.8274 ± 0.4030, p = 0.0325). A nonsignificant trend toward greater log reduction with PL compared to PPL was noted with S. aureus and E. coli at 3 h and E. coli at 24 h (Figures 3A,B). These data demonstrate that plasma contributes to the antimicrobial properties of platelets.
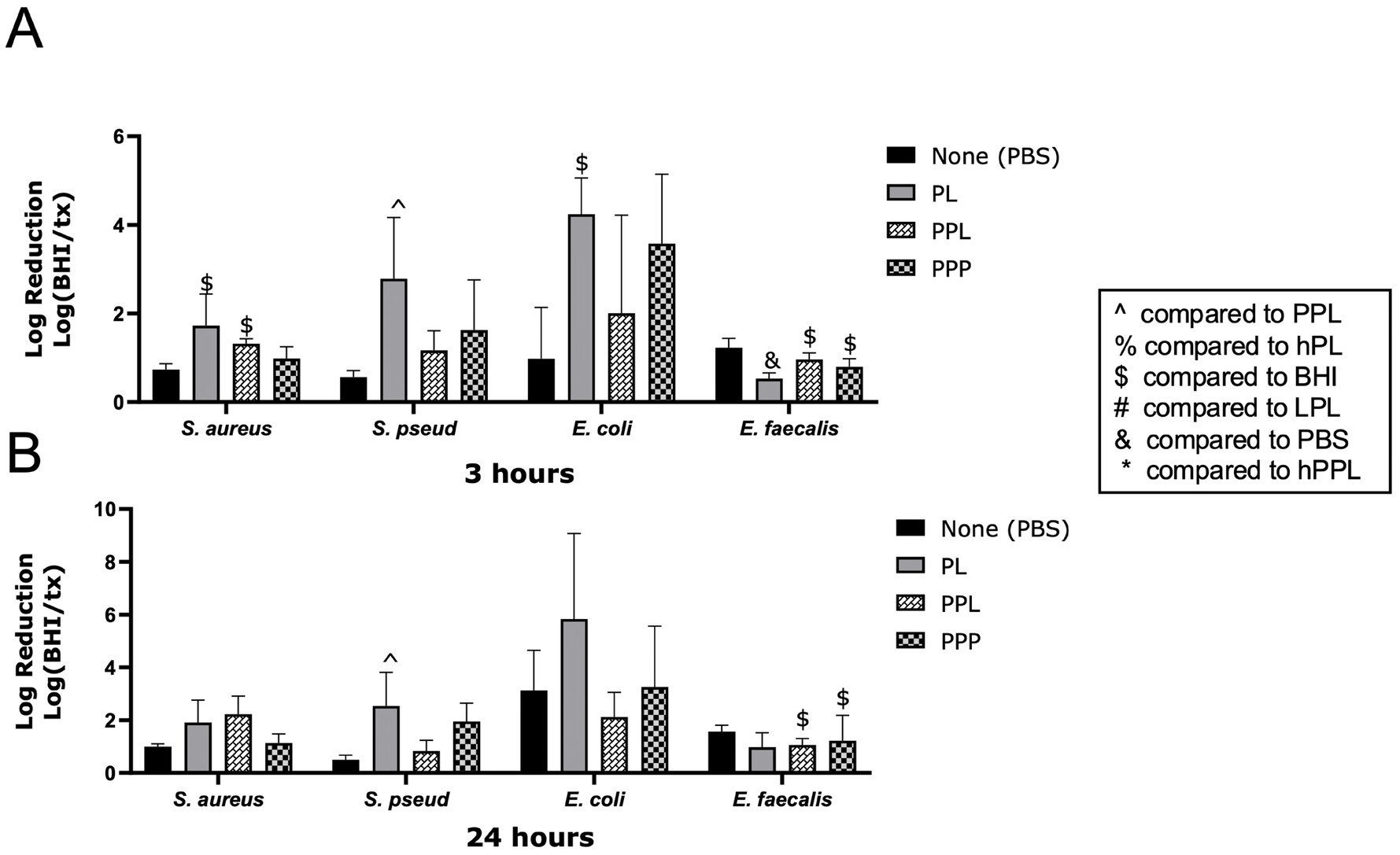
Figure 3. Log reduction of S. aureus, S. pseudintermedius, E. coli, and E. faecalis following the addition of PBS, PL, PPL, and PPP after 3 h (A) and 24 h (B). Data are presented as mean bacteria log reduction ± S. D. normalized to bacteria grown in BHI according to the following formula: Log (BHI/treatment). $p < 0.05 compared to BHI; ^p < 0.05 compared to PPL; n = 3 lots of pooled platelet lysate generated from 8 donors. PBS, Phosphate Buffered Saline; PL, Platelet Lysate; PPL, leukocyte-reduced platelet pellet lysate; PPP, Platelet-Poor Plasma; hPL, Heat-treated Platelet Lysate; BHI, brain heart infusion broth; LPL, Leukocyte-Rich Platelet Lysate; hPPL, Heat-treated leukocyte-reduced platelet pellet lysate.
The effect of complement inactivation
The log reduction of bacteria with PL and hPL to compare the effects of heat treatment is depicted in Figure 4. At 3 h, hPL had a significant log reduction of S. aureus (2.049 ± 0.2612, p = 0.0267), which was significantly greater than the effect of PBS (0.7370 ± 0.1307, p = 0.0320). PL led to a greater log reduction of E. faecalis (0.9720 ± 0.5482) compared to hPL (0.3381 ± 0.3916, p = 0.0026) after 24 h. No significant differences were noted between PL and hPL at either time point for E. coli or S. pseudintermedius (Figures 3A,B). This data suggests that heat-sensitive plasma proteins are necessary for the antimicrobial features of platelets.
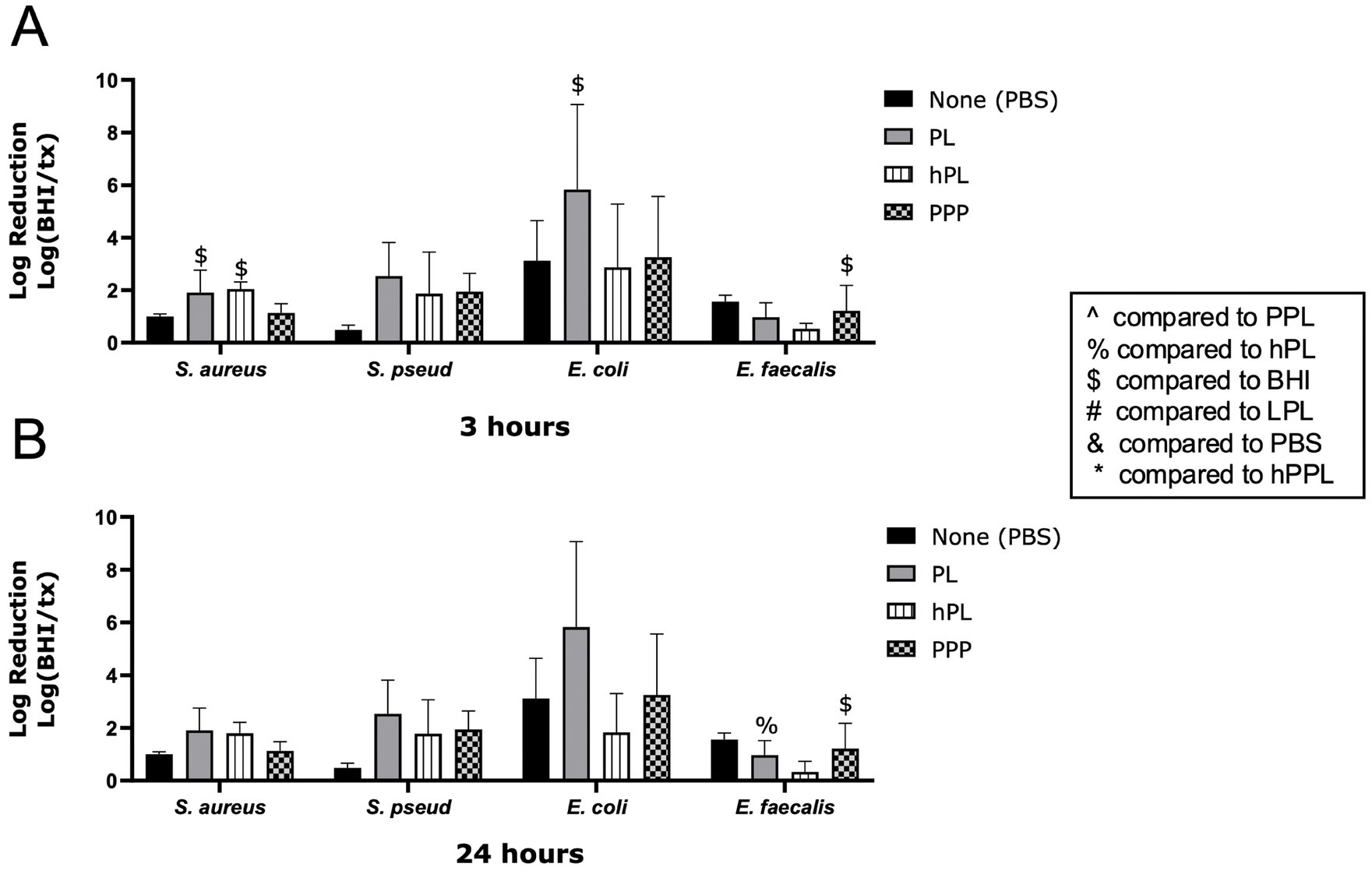
Figure 4. Log reduction of S. aureus, S. pseudintermedius, E. coli, and E. faecalis following the addition of PBS, PL, hPL, and PPP after 3 h (A) and 24 h (B). Data are presented as mean bacteria log reduction ± S. D. normalized to bacteria grown in BHI according to the following formula: Log (BHI/treatment). $p < 0.05 compared to BHI; %p < 0.05 compared to hPL; n = 3 lots of pooled platelet lysate generated from 8 donors. PBS, Phosphate Buffered Saline; PL, Platelet Lysate; hPL, Heat-treated Platelet Lysate; PPP, Platelet-Poor Plasma; PPL, leukocyte-reduced platelet pellet lysate; BHI, brain heart infusion broth; LPL, Leukocyte-Rich Platelet Lysate; hPPL, Heat-treated leukocyte-reduced platelet pellet lysate.
The effect of complement inactivation combined with plasma depletion
The log reduction of bacteria with PL and hPPL to compare the effects of plasma depletion combined with heat treatment is shown in Figure 5. Treatment with hPPL led to a significant log reduction of S. aureus at 3 h (1.819 ± 0.2719, p = 0.0368), which was also significantly greater reduction than PBS (0.7370 ± 0.1307, p = 0.0291), and at 24 h (; 1.001 ± 0.09561, p = 0.0049). Treatment with hPPL also led to a significant log reduction E. faecalis after 3 (0.9620 ± 0.04092, p = 0.0012) and 24 h (1.053 ± 0.3638, p = 0.0039). Conversely, the log reduction of E. coli after treatment with hPPL (1.645 ± 1.327) was significantly weaker compared to PPP (3.579 ± 1.566, p = 0.0282) after 3 h. PL was not significantly different from hPPL at 3 and 24 h, though trends were noted for PL to provide a greater log reduction of E. coli and S. pseudintermedius. Thus, the presence of plasma and its complement proteins is essential for maintaining the antimicrobial properties of platelets.
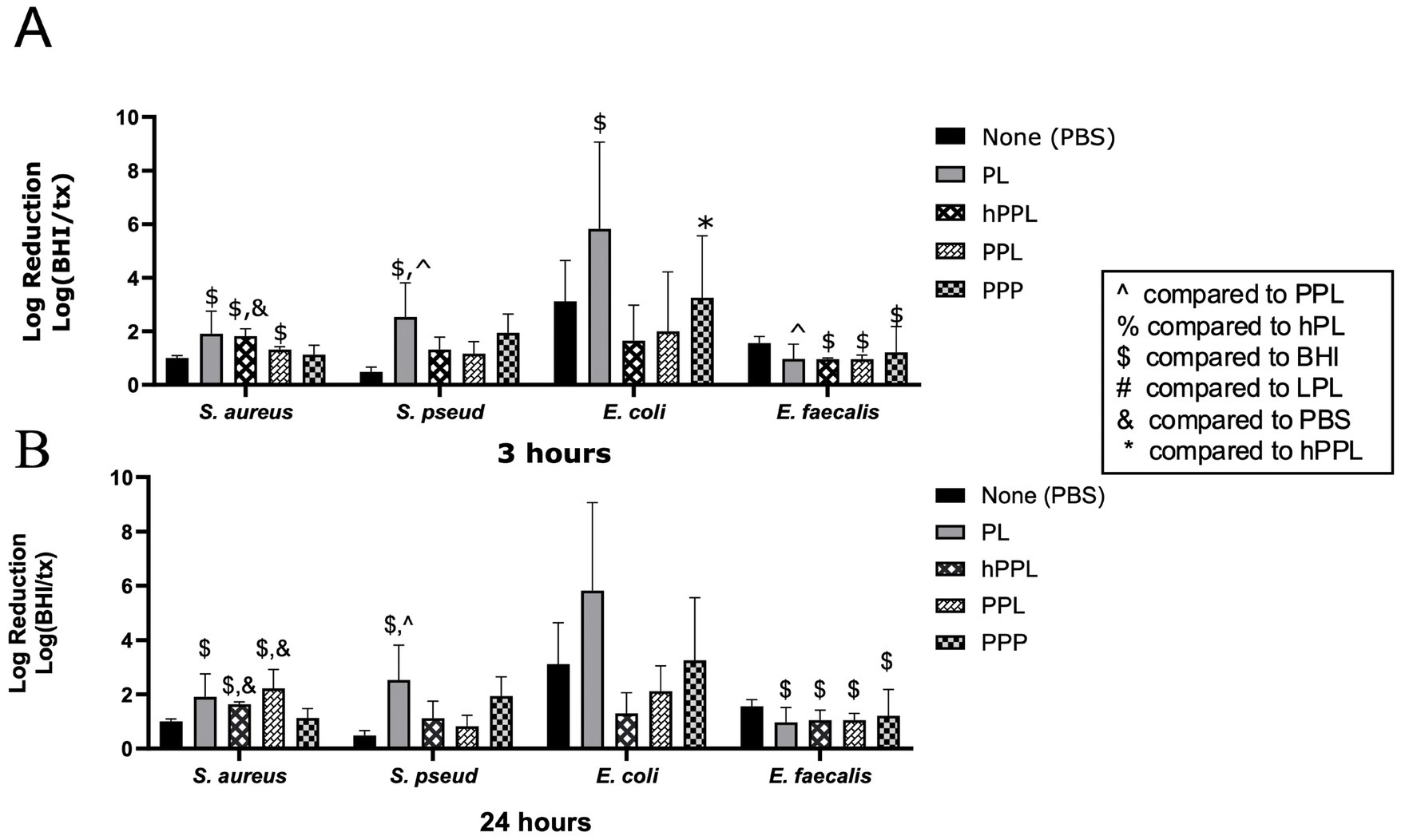
Figure 5. Log reduction of S. aureus, S. pseudintermedius, E. coli, and E. faecalis following the addition of PBS, PL, hPPL, PPL, and PPP after 3 h (A) and 24 h (B). Data are presented as mean bacteria log reduction ± S. D. normalized to bacteria grown in BHI according to the following formula: Log (BHI/treatment). $p < 0.05 compared to BHI; &p < 0.05 compared to PBS; ^p < 0.05 compared to PPL; *p < 0.05 compared to hPPL; n = 3 lots of pooled platelet lysate generated from 8 donors. PBS, Phosphate Buffered Saline; PL, Platelet Lysate; hPPL, Heat-treated leukocyte-reduced platelet pellet lysate; PPL, leukocyte-reduced platelet pellet lysate; PPP, Platelet-Poor Plasma; hPL, Heat-treated Platelet Lysate; BHI, brain heart infusion broth; LPL, Leukocyte-Rich Platelet Lysate.
The effect of nutrient depletion
In order to evaluate whether the inhibition of the bacteria growth following the addition of platelet derived products could be partially due to nutrient deprivation we set up a serial of experiments evaluating bacteria growth in the presence of PL, PPP and PBS. Specifically, Bacterial colony counts for S. aureus, S. pseudintermedius, E. coli and E. faecalis after 3 and 24 h of PL, PPP, and PBS treatment are demonstrated in Supplementary Figure 1. We found that, PL led to a nonsignificant inhibition of both S. aureus (Supplementary Figure 1A) and S. pseudintermedius (Supplementary Figure 1B) compared to baseline. However, PPP inhibited growth of S. pseudintermedius but supported the growth of S. aureus compared to baseline after 3 and 24 h. Moreover, we found that the addition of PL, PPP, and PBS significantly inhibited the growth of E. coli after 3 h compared to the baseline. Similarly, after 24 h, PL, PPP, and PBS significantly decreased E. coli growth. PBS significantly supported the growth of S. pseudintermedius at 24 h. The same nonsignificant trend was noted for S. aureus after the addition of PBS for two time points. Finally, PL significantly and PPP non significantly increased the growth of E. faecalis compared to baseline after 3 h. The same trends were observed for PL and PPP after 24 h. Even though PBS decreased its growth compered to baseline after 24 h. Given that bacteria growth was encountered following the addition of PBS, it is not reasonable to believe that inhibition of bacteria growth is due to the lack of nutrients that are essential for bacteria growth.
Discussion
In this study, highly concentrated PRP was achieved in both the leukocyte-reduced and leukocyte-rich formulations. The recommended platelet concentration for platelet concentrates is at least 300 × 103 platelets/ μL for a therapeutic effect to occur based on the recommendation by the American Red Cross (22, 39). In this study, the number of concentrated platelets was consistent and exceeded the guidelines of the American Red Cross and previous studies (39). Due to the high concentration of platelets and standardization, the final concentration of all platelet products was standardized to 800–1,000 × 103/μL prior to further processing. Additionally, leukocyte reduction in the leukocyte-reduced PRP was more successful compared to our previous work (37), which is in line with previous work by Shin et al. (22). However, leukocyte concentration was less successful in this study compared to previous studies using the leukocyte-rich method (17, 37). These variations are likely due to collection variations, donor variability, and processing time, particularly following the first centrifugation cycle.
When comparing PL and PPP, PL significantly reduced the growth of S. aureus and E. coli, and a trend was noted for PL to achieve a greater log reduction than PPP for E. coli, S. pseudintermedius, and S. aureus. These findings suggest that platelets are essential for the antimicrobial action of PL against E. coli, S. pseudintermedius, and S. aureus. Similar to PL, LPL demonstrated antimicrobial activity for S. aureus, E. coli, and S. pseudintermedius with a greater significance noted against S. aureus and E. coli at the 3-h timepoint. Significant antimicrobial activity against E. faecalis was not identified with treatment of PL or LPL. Our findings are consistent with the previous findings reported in the literature, noting platelet preparations inhibited S. aureus and E. coli (36), but not E. faecalis (40, 41). These results are also consistent with a previous study which proved leukocyte-reduced and leukocyte-rich platelet pellet concentrate have similar antimicrobial properties against five strains of bacteria (26). However, not all studies agree with this finding. Another study found the addition of neutrophils enhanced the antibacterial properties of platelet-rich plasma (30). Giusti et al. noticed that different leukocyte concentrations might have beneficial or detrimental impacts on the healing process, depending on the specific clinical circumstance (42). It is also worth noting that our results may be affected by the relatively low differential in leukocyte count between our formulations. Future studies should optimize the methodology used for leukocyte depletion during the manufacturing of platelet concentrates and guidelines may be needed for leukocyte concentrations depending on desired clinical applications.
Plasma has previously been noted to play a major role in the antibacterial properties of various orthobiologics (43). According to previous studies, the bactericidal properties of platelet products are mostly due to plasma factors (36). Other studies have also proved that the antimicrobial action of platelet products is related to the presence of plasma rather than platelets themselves (17, 36). Our results displayed that the depletion of plasma from PL (PPL) led to significantly less bacteria growth inhibition than PL after 3 and 24 h for S. pseudintermedius and E. coli. In contrast, PPL was more effective than PL against E. faecalis after 3 and 24 h and against S. aureus after 24 h. This latter finding is not in accordance with the previous literature, which concluded the antimicrobial properties of PRP against E. faecalis are due to the synergistic effects of plasma and platelet-derived factors (44). Additionally, in this study, plasma depletion inhibited the growth of S. aureus after 24 h. This might be due to the fact that S. aureus has a coagulase enzyme that converts fibrinogen to fibrin, which is used as an extracellular matrix to protect itself (45). By removing plasma, the above protection mechanisms are not available, which makes S. aureus more susceptible to therapeutic invasions. Thus, we believe that plasma is necessary for the antimicrobial properties of PL. Plasma contains protease, which seems critical for the cleavage of proteins in platelets. It is worth mentioning that the amount of plasma is a factor that needs to be considered carefully. In our study, platelet concentrates were resuspended in 100% plasma. However other studies found that variable plasma concentrations can affect the antimicrobial properties of the product (15, 46). Specifically, one study found that 10% of plasma is the most efficient concentration for maintaining an adequate amount of protease (15). Furthermore, another study demonstrated that 10% plasma is efficient and higher than 50% plasma is detrimental to the antimicrobial properties of platelets (46). Future studies should evaluate meticulously how various plasma concentrations can affect the antimicrobial features of platelet-derived products.
Based on previous studies, the heat-sensitive complement proteins within plasma are critical in inhibiting bacterial growth (27, 36). Our results further support this finding as heat-treated PL (hPL) did not provide the same bacterial growth reduction in any bacteria tested compared to PL. Accordingly, the presence of the protein complements seems crucial for maintaining the antimicrobial activity of PL. (47) Specifically, complement proteins use opsonin as labels on the bacteria to facilitate phagocytosis; in addition, they increase bacterial opsonization by antibodies since the antibodies kill the bacteria (33). Thus, our findings confirm the necessity of heat-sensitive complement proteins for maintaining the antimicrobial features of PL.
Combining plasma depletion and heat treatment was also evaluated. When plasma was depleted from PL, we found that plasma-depleted PL (PPL) was not efficient in inhibiting bacterial growth except for E. faecalis. Similarly, inactivating complements by utilizing heat had the same effect on PL (hPL). When we performed plasma depletion and heat treatment simultaneously, we confirmed a decreased potential of that formulation in inhibiting bacterial growth. Future studies on platelet products should consider the effects of plasma proteins and processing, including heat exposure, as these may change the product’s therapeutic potential.
To evaluate whether a nutritional deprivation is responsible for the increased bacteria reduction in the presence of platelet derived products we evaluated the growth of a specific concentration of bacteria in the presence of PL, PPP and PBS. We assumed that PBS would provide minimal notional support. According to our results, we confirmed that PL has a bactericidal effect against E. coli after both 3 and 24 h since it decreased the colony-forming units of a certain bacteria concentration more than 3 log 10 CFU/mL (28). Our finding is aligned with Burnouf et al. since a similar bactericidal activity of platelet preparations against E. coli was observed (36). In addition, both PL and LPL showed bacteriostatic activity against S. aureus, and S. pseud which is also consistent with previous studies (28). Thus, the antimicrobial effect of PL cannot be contributed to a nutrient deprivation effect.
Limitations of this study include a low differential between leukocyte concentrations in our “leukocyte-reduced” and “leukocyte-rich” preparation. This was likely affected by our inability to solely collect plasma from the top layer of the buffy coats, as contamination of leukocytes from accidentally aspirating the buffy coat is suspected. Protocol adjustments or use of different techniques or equipment may aid in obtaining the desired leukocyte concentration. Additional limitations include a small sample size, limited time points, and using commercially available bacterial strains collected from a human healthcare setting. In this study we decided to pool lysates generated from eight donors in an effort to decreased individual variability as has been previously reported (11).
The preparation process of platelet lysates included four freeze–thaw cycles in liquid nitrogen. It is possible that the number of freeze/thaw cycles could affect the release of factors from platelet granules and subsequently their biological activity. However, various studies have demonstrated that performing 3–5 freeze/thaw cycles achieves the optimal concentration of growth factors from platelet granules (48). Regardless, future studies should evaluate the effect of various numbers and conditions of freeze/thaw cycles for the optimal release of growth factors and antimicrobial properties. Finally, the generation process of lysates included the addition of acid-citrate-dextrose solution A (ACD-A) as an anticoagulant. Even though the addition of dextrose could have an impact on the composition of the growth factors, ACD-A is a common anticoagulant used in transfusion medicine for the preparation of platelet derived biologicals (49, 50). Specifically, a study evaluated the effect of various anticoagulants on the composition of growth factors and found that no differences were detected on PDGF concentration among the groups (49). However, a direct comparison has not been performed between ACD-A and acid-citrate to evaluate the effect of dextrose on factor composition.
Our results demonstrated that PL and LPL have antimicrobial properties independent of leukocyte concentration, specifically against S. aureus, E. coli, and S. pseudintermedius, but not E. faecalis. Plasma proteins and other plasma components seem to be critical for inhibiting the growth of some bacterial strains. In conclusion, platelet derived products have the potential to accelerate the wound healing process via the release of growth factors, anti-inflammatory and antimicrobial molecules. Future studies should investigate the most appropriate concentration of PL for targeted antimicrobial therapy and use in wound healing in dogs.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Institutional Animal Care and Use Committee (IACUC). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
MeM: Writing – original draft, Investigation, Formal analysis, Data curation, Methodology. AK: Data curation, Methodology, Writing – review & editing. TL: Writing – review & editing, Data curation. ND: Writing – review & editing, Data curation, Methodology. ED: Writing – review & editing, Methodology, Data curation, Supervision. MaM: Data curation, Writing – review & editing, Methodology. SS: Supervision, Data curation, Writing – review & editing, Writing – original draft, Methodology, Conceptualization, Investigation, Validation. MN: Visualization, Resources, Investigation, Data curation, Methodology, Conceptualization, Project administration, Validation, Supervision, Funding acquisition, Writing – original draft, Software, Writing – review & editing, Formal analysis.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Scott Fund and the Swaim Fund for Excellence in Wound Healing, Scott-Ritchey Research Center, College of Veterinary Medicine, Auburn University.
Acknowledgments
The authors would like to acknowledge Laura Huber for guidance in the interpretation of findings and assistance with statistical analysis.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1605649/full#supplementary-material
Supplementary Figure 1 | Mean number of bacterial colonies following treatment with PL, PPP, and PBS after 3 and 24 hours for S. aureus (A), S. pseudintermedius (B), E. coli (C), and E. faecalis (D). Data are presented as mean Colony Forming Unit per milliliter. The initial inoculum population was 108 CFUs/mL, represented by the dotted line (* = p < 0.05). n = 3 lots of pooled platelet lysate generated from 8 donors. PL, Platelet Lysate; PPP, Platelet-Poor Plasma; PBS, Phosphate Buffered Saline.
References
1. Pinto, JM, Pizani, NS, Kang, HC, and Silva, LA. Application of platelet-rich plasma in the treatment of chronic skin ulcer - case report. An Bras Dermatol. (2014) 89:638–40. doi: 10.1590/abd1806-4841.20143004
2. Barrientos, S, Stojadinovic, O, Golinko, MS, Brem, H, and Tomic-Canic, M. Growth factors and cytokines in wound healing. Wound Repair Regen. (2008) 16:585–601. doi: 10.1111/j.1524-475X.2008.00410.x
3. Alves, R, and Grimalt, R. A review of platelet-rich plasma: history, biology, mechanism of action, and classification. Skin Appendage Disord. (2018) 4:18–24. doi: 10.1159/000477353
4. Wroblewski, AP, Mejia, HA, and Wright, VJ. Application of platelet-rich plasma to enhance tissue repair. Oper Tech Orthop. (2010) 20:98–105. doi: 10.1053/j.oto.2009.10.006
5. Lynch, MD, and Bashir, S. Applications of platelet-rich plasma in dermatology: a critical appraisal of the literature. J Dermatolog Treat. (2016) 27:285–9. doi: 10.3109/09546634.2015.1094178
6. Smith, OJ, Wicaksana, A, Davidson, D, Spratt, D, and Mosahebi, A. An evaluation of the bacteriostatic effect of platelet-rich plasma. Int Wound J. (2021) 18:448–56. doi: 10.1111/iwj.13545
7. Kaushik, A, and Kumaran, MS. Platelet-rich plasma: the journey so far ! Indian Dermatol Online J. (2020) 11:685–92. doi: 10.4103/idoj.IDOJ_369_19
8. Gordon, J, Alvarez-Narvaez, S, and Peroni, JF. Antimicrobial effects of equine platelet lysate. Front Vet Sci. (2021) 8:703414. doi: 10.3389/fvets.2021.703414
9. da Fonseca, L, Santos, GS, Huber, SC, Setti, TM, Setti, T, and Lana, JF. Human platelet lysate - a potent (and overlooked) orthobiologic. J Clin Orthop Trauma. (2021) 21:101534. doi: 10.1016/j.jcot.2021.101534
10. Hagen, A, Holland, H, Brandt, VP, Doll, CU, Haussler, TC, Melzer, M, et al. Platelet lysate for mesenchymal stromal cell culture in the canine and equine species: analogous but not the same. Animals (Basel). (2022) 12:189. doi: 10.3390/ani12020189
11. Naskou, MC, Norton, NA, Copland, IB, Galipeau, J, and Peroni, JF. Innate immune responses of equine monocytes cultured in equine platelet lysate. Vet Immunol Immunopathol. (2018) 195:65–71. doi: 10.1016/j.vetimm.2017.11.005
12. Avellar, HK, Lutter, JD, Ganta, CK, Beard, W, Smith, JR, Jonnalagadda, N, et al. In vitro antimicrobial activity of equine platelet lysate and mesenchymal stromal cells against common clinical pathogens. Can J Vet Res. (2022) 86:59–64.
13. Tang, YQ, Yeaman, MR, and Selsted, ME. Antimicrobial peptides from human platelets. Infect Immun. (2002) 70:6524–33. doi: 10.1128/IAI.70.12.6524-6533.2002
14. Harman, RM, Yang, S, He, MK, and Van de Walle, GR. Antimicrobial peptides secreted by equine mesenchymal stromal cells inhibit the growth of bacteria commonly found in skin wounds. Stem Cell Res Ther. (2017) 8:157. doi: 10.1186/s13287-017-0610-6
15. Gilbertie, JM, Schaer, TP, Schubert, AG, Jacob, ME, Menegatti, S, Ashton Lavoie, R, et al. Platelet-rich plasma lysate displays antibiofilm properties and restores antimicrobial activity against synovial fluid biofilms in vitro. J Orthop Res. (2020) 38:1365–74. doi: 10.1002/jor.24584
16. Duong, L, Gross, SP, and Siryaporn, A. Developing antimicrobial synergy with AMPs. Front Med Technol. (2021) 3:640981. doi: 10.3389/fmedt.2021.640981
17. Attili, AR, Iacoucci, C, Serri, E, Cuteri, V, Cantalamessa, A, Linardi, M, et al. Antibacterial properties of canine platelet-rich plasma and other non-Transfusional Hemo-components: an in vitro study. Front Vet Sci. (2021) 8:746809. doi: 10.3389/fvets.2021.746809
18. Farghali, HA, AbdElKader, NA, AbuBakr, HO, Aljuaydi, SH, Khattab, MS, Elhelw, R, et al. Antimicrobial action of autologous platelet-rich plasma on MRSA-infected skin wounds in dogs. Sci Rep. (2019) 9:12722. doi: 10.1038/s41598-019-48657-5
19. Naskou, MC, Sumner, SM, Chocallo, A, Kemelmakher, H, Thoresen, M, Copland, I, et al. Platelet lysate as a novel serum-free media supplement for the culture of equine bone marrow-derived mesenchymal stem cells. Stem Cell Res Ther. (2018) 9:75. doi: 10.1186/s13287-018-0823-3
20. Franklin, SP, Birdwhistell, KE, Strelchik, A, Garner, BC, and Brainard, BM. Influence of cellular composition and exogenous activation on growth factor and cytokine concentrations in canine platelet-rich plasmas. Front Vet Sci. (2017) 4:40. doi: 10.3389/fvets.2017.00040
21. Franklin, SP, Garner, BC, and Cook, JL. Characteristics of canine platelet-rich plasma prepared with five commercially available systems. Am J Vet Res. (2015) 76:822–7. doi: 10.2460/ajvr.76.9.822
22. Shin, HS, Woo, HM, and Kang, BJ. Optimisation of a double-centrifugation method for preparation of canine platelet-rich plasma. BMC Vet Res. (2017) 13:198. doi: 10.1186/s12917-017-1123-3
23. Perazzi, A, Busetto, R, Martinello, T, Drigo, M, Pasotto, D, Cian, F, et al. Description of a double centrifugation tube method for concentrating canine platelets. BMC Vet Res. (2013) 9:146. doi: 10.1186/1746-6148-9-146
24. Carr, BJ, Canapp, SO Jr, Mason, DR, Cox, C, and Hess, T. Canine platelet-rich plasma systems: a prospective analysis. Front Vet Sci. (2015) 2:73. doi: 10.3389/fvets.2015.00073
25. Qian, Y, Han, Q, Chen, W, Song, J, Zhao, X, Ouyang, Y, et al. Platelet-rich plasma derived growth factors contribute to stem cell differentiation in musculoskeletal regeneration. Front Chem. (2017) 5:89. doi: 10.3389/fchem.2017.00089
26. Mariani, E, Canella, V, Berlingeri, A, Bielli, A, Cattini, L, Landini, MP, et al. Leukocyte presence does not increase microbicidal activity of platelet-rich plasma in vitro. BMC Microbiol. (2015) 15:149. doi: 10.1186/s12866-015-0482-9
27. Lopez, C, Carmona, JU, Giraldo, CE, and Alvarez, ME. Bacteriostatic effect of equine pure platelet-rich plasma and other blood products against methicillin-sensitive Staphylococcus aureus. An in vitro study. Vet Comp Orthop Traumatol. (2014) 27:372–8. doi: 10.3415/VCOT-14-04-0054
28. Anitua, E, Alonso, R, Girbau, C, Aguirre, JJ, Muruzabal, F, and Orive, G. Antibacterial effect of plasma rich in growth factors (PRGF(R)-Endoret(R)) against Staphylococcus aureus and Staphylococcus epidermidis strains. Clin Exp Dermatol. (2012) 37:652–7. doi: 10.1111/j.1365-2230.2011.04303.x
29. Bielecki, T, Dohan Ehrenfest, DM, Everts, PA, and Wiczkowski, A. The role of leukocytes from L-PRP/L-PRF in wound healing and immune defense: new perspectives. Curr Pharm Biotechnol. (2012) 13:1153–62. doi: 10.2174/138920112800624373
30. Prysak, MH, Lutz, CG, Zukofsky, TA, Katz, JM, Everts, PA, and Lutz, GE. Optimizing the safety of intradiscal platelet-rich plasma: an in vitro study with Cutibacterium acnes. Regen Med. (2019) 14:955–67. doi: 10.2217/rme-2019-0098
31. Lincoln, JA, Lefkowitz, DL, Cain, T, Castro, A, Mills, KC, Lefkowitz, SS, et al. Exogenous myeloperoxidase enhances bacterial phagocytosis and intracellular killing by macrophages. Infect Immun. (1995) 63:3042–7. doi: 10.1128/iai.63.8.3042-3047.1995
32. Powanda, MC, and Moyer, ED. Plasma proteins and wound healing. Surg Gynecol Obstet. (1981) 153:749–55.
33. Janeway, CA, and Travers, P. Immunobiology: the immune system in health and disease. Cambridge, MA: Current Biology (2001).
34. Reichhardt, MP, and Meri, S. Intracellular complement activation-an alarm raising mechanism? Semin Immunol. (2018) 38:54–62. doi: 10.1016/j.smim.2018.03.003
35. Nordahl, EA, Rydengard, V, Nyberg, P, Nitsche, DP, Morgelin, M, Malmsten, M, et al. Activation of the complement system generates antibacterial peptides. Proc Natl Acad Sci USA. (2004) 101:16879–84. doi: 10.1073/pnas.0406678101
36. Burnouf, T, Chou, ML, Wu, YW, Su, CY, and Lee, LW. Antimicrobial activity of platelet (PLT)-poor plasma, PLT-rich plasma, PLT gel, and solvent/detergent-treated PLT lysate biomaterials against wound bacteria. Transfusion. (2013) 53:138–46. doi: 10.1111/j.1537-2995.2012.03668.x
37. Lunardon, T, Sumner, SM, Mollabashi, M, Darzenta, N, Davis, E, and Naskou, MC. Growth factor and cytokine characterization of canine platelet lysate with variable leukocyte concentration, plasma content, and heat-sensitive proteins. Front Vet Sci. (2024) 11:1408080. doi: 10.3389/fvets.2024.1408080
38. Shariati, A, Moradabadi, A, Azimi, T, and Ghaznavi-Rad, E. Wound healing properties and antimicrobial activity of platelet-derived biomaterials. Sci Rep. (2020) 10:1032. doi: 10.1038/s41598-020-57559-w
39. Dohan Ehrenfest, DM, Rasmusson, L, and Albrektsson, T. Classification of platelet concentrates: from pure platelet-rich plasma (P-PRP) to leucocyte- and platelet-rich fibrin (L-PRF). Trends Biotechnol. (2009) 27:158–67. doi: 10.1016/j.tibtech.2008.11.009
40. Bielecki, TM, Gazdzik, TS, Arendt, J, Szczepanski, T, Krol, W, and Wielkoszynski, T. Antibacterial effect of autologous platelet gel enriched with growth factors and other active substances: an in vitro study. J Bone Joint Surg Br. (2007) 89:417–20. doi: 10.1302/0301-620X.89B3.18491
41. Mariani, E, Filardo, G, Canella, V, Berlingeri, A, Bielli, A, Cattini, L, et al. Platelet-rich plasma affects bacterial growth in vitro. Cytotherapy. (2014) 16:1294–304. doi: 10.1016/j.jcyt.2014.06.003
42. Giusti, I, Di Francesco, M, D'Ascenzo, S, Palumbo, P, Rughetti, A, Dell'Orso, L, et al. Leukocyte depletion does not affect the in vitro healing ability of platelet rich plasma. Exp Ther Med. (2018) 15:4029–38. doi: 10.3892/etm.2018.5887
43. Nicol, MJ, Brubaker, TR, Honish, BJ 2nd, Simmons, AN, Kazemi, A, Geissel, MA, et al. Antibacterial effects of low-temperature plasma generated by atmospheric-pressure plasma jet are mediated by reactive oxygen species. Sci Rep. (2020) 10:3066. doi: 10.1038/s41598-020-59652-6
44. Drago, L, Bortolin, M, Vassena, C, Romano, CL, Taschieri, S, and Del Fabbro, M. Plasma components and platelet activation are essential for the antimicrobial properties of autologous platelet-rich plasma: an in vitro study. PLoS One. (2014) 9:e107813. doi: 10.1371/journal.pone.0107813
45. Zapotoczna, M, O'Neill, E, and O'Gara, JP. Untangling the diverse and redundant mechanisms of Staphylococcus aureus biofilm formation. PLoS Pathog. (2016) 12:e1005671. doi: 10.1371/journal.ppat.1005671
46. Trier, DA, Gank, KD, Kupferwasser, D, Yount, NY, French, WJ, Michelson, AD, et al. Platelet antistaphylococcal responses occur through P2X1 and P2Y12 receptor-induced activation and kinocidin release. Infect Immun. (2008) 76:5706–13. doi: 10.1128/IAI.00935-08
47. Zimmer, J, Hobkirk, J, Mohamed, F, Browning, MJ, and Stover, CM. On the functional overlap between complement and anti-microbial peptides. Front Immunol. (2014) 5:689. doi: 10.3389/fimmu.2014.00689
48. Strandberg, G, Sellberg, F, Sommar, P, Ronaghi, M, Lubenow, N, Knutson, F, et al. Standardizing the freeze-thaw preparation of growth factors from platelet lysate. Transfusion. (2017) 57:1058–65. doi: 10.1111/trf.13998
49. Aizawa, H, Kawabata, H, Sato, A, Masuki, H, Watanabe, T, Tsujino, T, et al. A comparative study of the effects of anticoagulants on pure platelet-rich plasma quality and potency. Biomedicines. (2020) 8:42. doi: 10.3390/biomedicines8030042
Keywords: canine platelet lysate, leukocyte concentration, complement proteins, plasma, antimicrobial properties
Citation: Mollabashi M, Klopfer A, Lunardon T, Darzenta N, Davis E, Murray M, Sumner SM and Naskou MC (2025) Plasma and complement proteins are essential for the antimicrobial activity of canine platelet lysate. Front. Vet. Sci. 12:1605649. doi: 10.3389/fvets.2025.1605649
Edited by:
Luca Melotti, University of Padua, ItalyReviewed by:
Janina Burk, University of Veterinary Medicine Vienna, AustriaKevin Yaneselli, Universidad de la República, Uruguay
Copyright © 2025 Mollabashi, Klopfer, Lunardon, Darzenta, Davis, Murray, Sumner and Naskou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Scarlett M. Sumner, c21zMDE5M0BhdWJ1cm4uZWR1; Maria C. Naskou, bXpuMDA1MEBhdWJ1cm4uZWR1